the vestibular system modulates masseter · pdf filetaxic frame. a thermistor probe ... and...
TRANSCRIPT

! .. I
Journal of Vestibular Research, Vol. 3, pp. 163-171, 1993 Printed in the USA. All rights reserved.
0957-4271/93 $6.00 + .00 Copyright © 1993 Pergamon Press Ltd.
THE VESTIBULAR SYSTEM MODULATES MASSETER MUSCLE ACTIVITY
E. Tolu and M. Pugliatti
Institute of Human Physiology, University of Sassari, Viale S. Pietro 43/b, 07100 Sassari, Italy Reprint address: Eusebio Tolu, Institute of Human Physiology, University of Sassari,
Viale S. Pietro 43/b, 07100 Sassari, Italy
o Abstract - The aim of this study was to investigate whether, and in what way, the vestibular input may influence the activity of the masseter muscles. The variations in the spontaneous electrical activity and the evoked responses in the masseter motor units to natural or electrical activation of the vestibular afferents were recorded in anesthetized guinea pigs. The effects of a unilateral lesion of the labyrinth on the firing rate of the masseter motor units were also studied. Results show that: 1) vestibular input elicited an excitatory tonic control on masseter muscle activity; 2) a faster labyrinthine control is driven to the contralateral than the homolateral masseter muscles; 3) vestibular macular input does exert an asymmetrical control on masseteric muscles of both sides, in relation to the head displacement in space. The latencies of responses recorded from the masseter motor units suggest that polysynaptic pathways are involved in connecting the vestibular system to the trigeminal complex. The possible anatomical substrates for this vestibulomasseteric reflex are discussed.
o Keywords - vestibular system; masseter muscles; vestibulomasseteric reflex.
I utroduc:ion
I: is wei k n Ov.T. ina: .. iH. veSli:JUlal SYSL::m
play~ an im;1onan! role ir con:rolling the activity of the motoneurons inne:-vating the extrinsic eye muscles (1-4) and in modulating the activity of the alpha-motoneurons and gamma-motoneurons projecting to the neck (5,6), trunk, and limb extensor muscles (7-9). These activities are aimed at maintaining posture and equilibrium and at governing loco-
motion. In addition, recent experiments have demonstrated that macular and ampullar inputs participate in controlling the activity of the intrinsic and extrinsic tongue muscles (lOB). These vestibulo-hypoglossal connections are important in controlling the tongue position in the mouth during head displacement and jaw movements in the course of the masticatory cycles. However, it is not yet clear whether impulses coming from the vestibular system can also influence the activity of trigeminal masticatory motoneurons. There is clinical evidence that patients suffering from a vestibular syndrome also show functional impairment of the jaw-closing muscles (14). Furthermore, in man, a modulation of the masseter muscle's monosynaptic reflex during labyrinthine stimulation has been described (15).
Because present knowledge of these specific nervous connections is still incomplete, the aim of this study was to investigate whether, and in what way, the vestibular input influences the activity of the masseter muscle, chosen as the main j2.\\-ciosing m:;scie .
T wo set~ of experiments were perfo:-meci if. 35 guinea pigs. The general experimental procedure for all animal groups was as follows: under anesthesia with ketamine (60 mg/kg i.m.), supplemented as required throughout the experiments, the animals were tracheotomized and placed in a prone position with the
RECEIVED 4 August 1992; REVISED MANUSCRIPT RECEIVED 21 October 1992; ACCEPTED 23 October 1992.
163

164
head and mandibular bone fixed to a stereotaxic frame. A thermistor probe was placed in the rectum and the body temperature was kept at 39 ± 0.5°C. Heart rate and pupil diameter were continuously controlled for the animal's perception of pain. The left epitympanic recessus was opened to exposed the ampullae of the horizontal and vertical canals, the iJoste:-ior .:anal being surgically unapj:roac!1abie. "!" 'mgs[e:1 5 11m ong ~!ec[rodes. insulated ex;: ~P{ ~ G:· :nl;! :lP SotO- ~ CG( j :C.) i:.:: -;!:>iSi..!.-:_=
'.vere nsertec :nro :he :nLlsserer :nusc!es :'c r reccrding :he ~!ect ricaj activity ;)[ 3ing!e :nctor ;,rnits. The spontaneous dectricallctivity and the evoked responses of single motor units to natural or electrical activation of the vestibular system were recorded and stored on magnetic tape (Racal Recorders Ltd.) for later analysis. At the end of the experiments the animals were killed with a barbiturate overdose. The motor unit spikes, converted by a window discriminator to standard pulses, were fed from the magnetic tape to the Techfen Computer-scope (ISC-16 program) to construct the pulse rate and the poststimulus time histograms (PSTHs) of the responses elicited by 100 consecutive electrical stimuli of the labyrinth. The spontaneous spikes of the masseter motor units and the raw data of single stimulus responses were also displayed on the memory oscilloscope computer and fed to a plotter for charting the analogical signals.
First Set (19 Experiments)
Five out of 15 experiments were designed to study the effects of a unilateral lesion of the labyrinth on the firing rate of the single motor units recorded in the contralateral masseter muscle. Recordings were performed in basal conditions and after destruction of the labyrinth by drilling the internal ear (16,17). A control of 4 animals was studied in the same way: the surgery and the bone drilling to expose the middle ear was quite extensive, while the only experimental difference was the drilling of the internal ear. This was short, and could not have caused additional algetic stimulus capable of independently modifying the masseter muscle activity in fully anesthetized
E. TOlu and ~/1 , : ugliatti
animals . In a further group of animals (5 experiments) the firing rate of the masseter motor units was analyzed following caloric stimulation of the contralateral labyrinth. This classic procedure produces endolymphatic currents that mechanically stimulate the ampullar receptors, thus allowing the unilateral stimulation of the I/estibular receptors. Caloric stimulaLion '.vas ~erformed ,'or : 0 :; oy directly l rri ~
sat ing ~he ~xposed vestiiJ ular <.:anais with Wee :} i y ~!·-:r4 · ·;va~c :· ::..;.- . : ~ . _:;lrl ic:.l izu." J.l t~ ~[ ion
'NilS ;JU!U .10[ :0 inv·.::;ive a ny 0[her neili by " ' ~ 'l''' '' rI'C in ' ''e i r ;: crati r cr 1"I r o,"'dur ;" .-y·['as se-...; L .. U"" ....... ... _.J ... ... J.l ~ .. 1 0 .1.t~ :' "".... .. .... . . ,
teric activity was recorded by means of a mngsten electrode, previously implanted into the contralateral muscle.
In another group of animals (5 experiments), the masseteric muscle activity evoked by the specific stimulation of the vestibular macular receptors was analyzed. The animal, fixed in the stereotaxic apparatus, was locked on a tilting frame consisting of a special platform which could be slowly revolved on its longitudinal axis by a crank connected to a worm screw. In this way, the whole animal could be tilted without changing the position of the head in relation to the body, thus excluding neck reflexes. Lateral tilting up to 20° was performed at a steady angular speed (less than 0.05°/s) with smooth starting and stopping in order to avoid stimulation of the ampullar receptors. The firing rate of single masseter motor units was analyzed in basal conditions and following ipsilateral and contralateral static tilting.
Second Set (16 Experiments)
These experiments were aimed at clarifying the vestibulomasseteric connections by using electrical stimulation of vestibular afferents. In the animals, with the left exposed epitympanic recessus, 2 silver wires (25 /Lm diam., insulated except for the tips) were inserted into the ampullae of the horizontal and vertical canals, respectively. A third silver wire was placed in the surrounding bone as reference electrode. The wires were then fixed in place with dental cement and connected to a model S11 Grass stimulator. Electrical stimulus of
I·

L
Vestibular Input to Masseter Motoneurons
the vestibular receptors was performed using the following parameters: 0.1 ms of duration, 1 Hz frequency, intensity just above the threshold for inducing ocular jerks. In control experiments, this stimulus adjustment proved to be twice the threshold for the N J
response in the ipsilateral vestibular nuclei (18,19). The tympanic exposure was protected with paraffm. The response of the motor units were recorded by means of tungsten microelectrode previously implanted into the masseter muscle of both sides. As further evidence of specific labyrinthine stimulation, a monophasic potential can be evoked in the masseteric nerve following electrical stimulation of the contralateral vestibular nerve in 4 animals. However, because isolating the vestibular nerve involved destroying the labyrinthine receptors, in the experimental animals the electrical stimuli were applied directly on intact vestibular receptors, as reported in the paper.
Results
Effects of a Vestibular Lesion on Masseter Muscle Activity
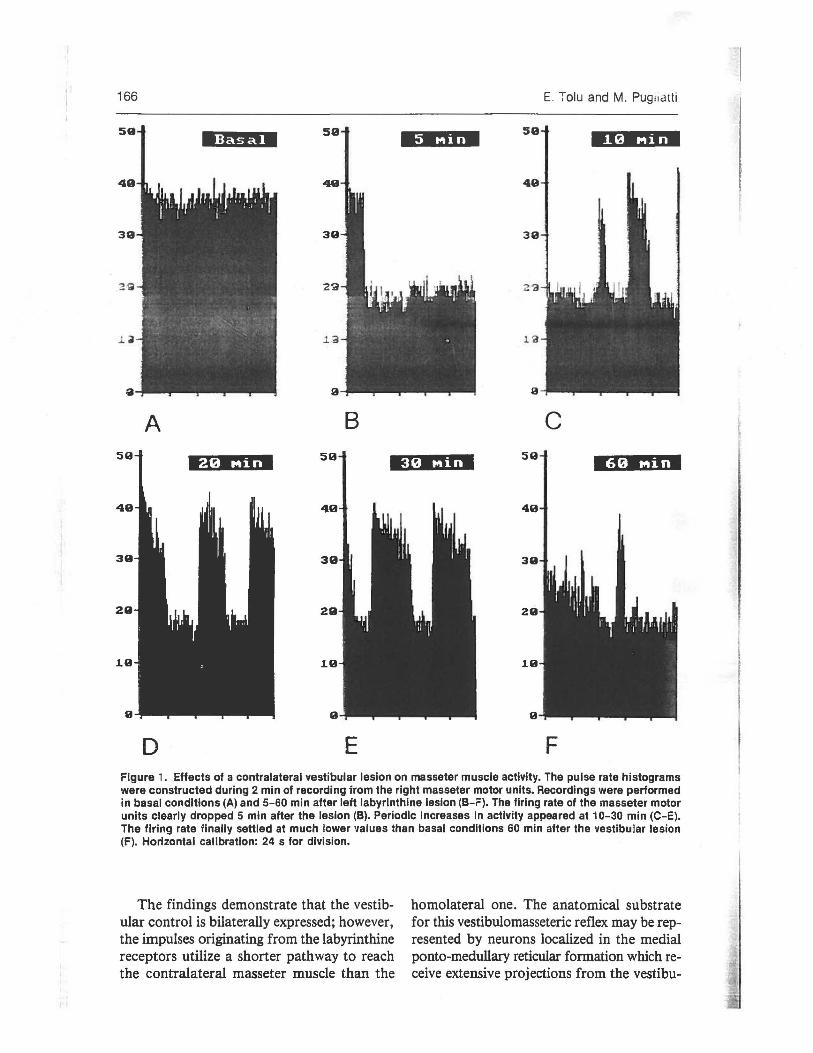
Figure 1 shows the spontaneous pulse rate of a masseteric motor unit recorded in basal conditions (A) and following a unilateral lesion of the contralateral vestibular receptors (B-F). Five minutes after the lesion (B), there was a marked drop in the pulse rate, followed by bursts of activity lasting from 7 to 35 s, mainly 10 and 30 min after hemilabyrinthectomy (C-E). The pulse rate finally settled at much lowe:- values thar; basal conditions (F). These results show that the vestibular input elicited an excitatory tonic control OD the COD-
Effects oj Naturai Vestibular Stimulation on j'viasseter . 1l1uscle Activity
Figure 2 shows the spontaneous firing rate of masseter motor units recorded in basal conditions (A) and following caloric stimulation of the contralateral labyrinth (B-D). The firing rate of the motor units increased as soon as the stimulus was applied (B); after its re-
165
moval (C), the firing rate changed back to basal conditions (0). These observations confirm a vestibular excitatory influence on the contralateral masseteric muscle fibers. Figure 3 shows the spontaneous firing rate of masseter motor units recorded with the animal in horizontal position (B) and during contra- or ipsilateral static tilting. The firing rate increased after contralateral (A) and decreased after ipsilateral (C) tilting. Prevalentlyopposite effects were observed following caloric stimulation with cold water. Three repeated stimulation cycles were performed for selected units, and in each case the response pattern was maintained. It is evident that vestibular macular input does exert an asymmetrical control on masseteric muscle, in relation to the head displacement in space.
Effects of Electrical Stimulation of the Vestibular Afferents on the Masseter Muscle Activity
Figure 4 shows the patterns of response of masseter motor units of both sides evoked after the electrical stimulation was applied in the left anterior and lateral vestibular canals. A and B show the contralateral and C and D the homolateral recordings; the insets show examples of masseter motor unit responses to a vestibular stimulus. The histograms were constructed following 100 consecutive stimuli applied to anterior (A and C) and lateral (B and D) ampUllae. Excitatory responses were observed most frequently. The latency values were found to be lowe:- in cont;-aiate;-a: recordings: 9.26::: :.04 ms (mean ± SDj foliowing anterior ampullar stimulation and 9.02 ±
.25 ~5 ("!fcnr·~l!"lp iar.e-a am!1uIi2.- s:imuictior .. comparee witt latencies 0; 1 t." :::: :.4S ane J9.58 ± 2.:: ms. respectivel~. if' hom~llate;-airecordings. These finding~ show that a faster labyrinthine control is driven to the contralateral masseter muscle.
Discussion
These experiments provide clear evidence that the vestibular system does modulate the activity of the masseteric motoneurons.
,I
166 E. iolu and M. PUglldlti
58 M:4*,- 58 •• :n.M 5 Mlo:_:.'._
A B c 50 50 50
·a~·:';,'
D E F Figure 1. Effects of a contralateral vestibular lesion on masseter muscle activity. The pulse rate histograms were constructed during 2 min of recording Trom the right masseter motor units. Recordings were performed in basal conditions (A) and 5-60 min after left labyrinthine lesion (B-F). The firing rate of the masseter motor units clearly dropped 5 min after the lesion (B). Periodic increases in activity appeared at 10-30 min (C-E). The firing rate iinally settled at much lower values than basal conditions 60 min after the vestibular lesion (F). Horizontal calibration: 24 s for division.
The findings demonstrate that the vestibular control is bilaterally expressed; however, the impulses originating from the labyrinthine receptors utilize a shorter pathway to reach the contralateral masseter muscle than the
homolateral one. The anatomical substrate for this vestibulomasseteric reflex may be represented by neurons localized in the medial ponto-medullary reticular formation which receive extensive projections from the vestibu-

i
i' I
Vestibular Input to Masseter Motoneurons 167
A
B
t
D
Figure 2. Effects of caloric stimulation of the labyrinth on the masseter motor units. Spontaneous electrical activity of masseter motor units recorded in basal conditions (A) and during caloric stimulation of the contralateral labyrinth IS-C), The arrows indicate the beginnin£ an~ ent' of the stimulation. The firing rate of the motor units in::reaseC::, as soon'as the stimulu~ was apphec (8;' after its remova: 1: \ the firin£ rate changed ba:!: iC besa' conditions (D;, Ve~ti:a ' calibratior. : 10(, FI:', norlzon;a ' calibrati on: " s,
iar nuclea:' _ompie), (20), Responses in th e medial reticuiar neuron:, were observed following natural activation of the labyrinthine receptors (21,22) and after electrical stimulation of the vestibular nerves (23) or vestibular nuclei (24), The reticular formation contains a mixed population including neurons with axons ascending to midbrain or projecting to trigeminal nuclear complexes including the V motor nuclei (25-27). Hence, the vestibular
a:f ~··ents coule D~ojec~ direc:; ~ oniO the t~jgemina! mowr nucleus 0;- ontO the adjacent premotor neurons involved in transmitting the various centrally and peripherally evoked modulatory influences to V motoneurons (28,29). As far as the vestibulomasseteric pathway is concerned, polysynaptic connections must be assumed. The latencies of responses recorded from the masseter motor units (9.26 ms for anterior and 9.02 ms for lateral ampullae
:;"1 !t I
/.

168 E. Tolu ',,",j H. Pugliatti
A
HI"
7S
B
7:5
(Scale = 9.792 S /Div)
c .1_ I
J sa .
zs
(Scale = ~.7~2 S /Div)
Figure 3. Effects of static tilting on the masseter motor units. Each pulse rate histogram was constructed during 60 s of recording from the masseter motor units. The Insets show the'raw data of the electrical activity of the motor units. Recordings were performed with the animal in horizontal position (8) and during contraor Ipsilateral static tilting (A,e). The firing rate increased after contralateral (A) and decreased after Ipsilateral tilting (C). Insets: vertical calibration 100 "V, horizontal calibration 1.8 s.

CJ) (0
A B 25
~ P l ~
A
~ p l ~
III!JIJ ID I d , II B I IIII , , t t
C D
··t dli 28-1 I .,1
p l
25
~. P l
I 1.5
1.8
5
8' ' 'l'a ''''';TJlU~~Jml~lrn jl_II.llLU~'' II I, II II I , t t (Soale 18.89 n$/Dlv)
Figure 4. Effects of electrir.fll !"tirn'lIFlti"n 01 thl'! anterior and lateral canals on masseter motor unit activity. (A,B) Contralateral masseter muscle recordings; (C,D) Ipsilateral masseter muscle rpcr:>nUnQ<; . Thp insets show spike potentials evoked In the masseter muscles following a single electrical stimulus applied to the vestibular ampullae (0.1 ms of r1\/IFllip". 1 117 IrPfI'.,ency, intensity twice the threshold for evoking the N1 response In the Ipsilateral vestibular nuclei). A: anterior; L: lateral; P: posterior canals. Vprtir'" ('alihratinn: 1000 p.V; horizontal calibration: 20 ms. The histogram responses were constructed following 100 consecutive stimuli. The arrows mark thp <;lirn .. I,,<; ",lil"r. I<;.

170
stimulations) observed following contralateral vestibular stimulation are consistent with polysynaptic pathways such as: primary vestibular neurons, reticular formation, premo tor interneurons, and trigeminal motoneurons. It is likely that secondary vestibular neurons are involved in the vestibulo-masseteric reflex. The duration ,)f ;: he respcnse and :he ~ong
la[e!1des :joservd (o llowing :psi lateral vesribuiar;[!mulaticn ( ~ 6..+ :us ;"o r J.nten or .lna : Q.58 ::-IS ~'or .U:':::l! ·lffi iJ1.!iine :iI:m ... .! .Ofll suggest ,:lat .)[~er ::;oiysynapt:c ;JaL hway'l lre involved ~ n :onnec[ing :he 'Jes[i iJ ular system w the trigeminal .:ompiex. i t :s :mown : ha[ labyrinthine stimulation evokes vestibulocollie reflex, which acts through the vestibulospinal axons on C1-Cs motoneurons controlling the neck muscles to stabilize the head position in space (30-33). There is evidence of the importance of parallel pathways, presumably reticulospinal tracts, in producing the reflex (33,34). Anatomical and electrophysiological studies demonstrated a direct projection of spinal afferents arising from cells localized in the cervical dorsal horn, to the sensory spinal trigeminal complex in the rat (35,36). In addition, trigeminal spinal neurons have been shown to project to the spinal cord (37,38). The complex spatial relationship and the ex-
E. ,vlU and M. PUglidttl
tensive overlap between the spinotrigeminal and trigeminospinal pathways could represent another anatomical substrate for reciprocal functional loops involving the spinal trigeminal complex and the cervical spinal cord. Because the cervical spinal cord is implicated in the processing of vestibular information from :he maculae and ampuilae :-eceptors it could aiso Je invoived ~n the vestibulo-[rigeminai :-eiat ionsnip shown in :he ;Jresem ;!:~periments.
: '_El~~!C;l~i ! :i :~lc /c':Jli C~~G- ,_r: ~emii1ai re!J.~lCnsn i p ;1<15 [Wo impenam roies:
: . ,he maimenance of muscle :ohe: :nasse[eric :notorunits are subject to excitatory tonic control from the vestibular system in basal conditions, in the same way as in other physiological extensor muscles. These effects appeared more evident in the contralateral muscle although the control was bilateral.
2. Its involvement in the masseter function: responses obtained by tilting showed that there is vestibular asymmetrical control on both the homolateral and contralateral muscles, so as to adapt their muscle tone to the head displacement in order to keep the mandibular axis in the right position.
Acknowledgments- This research was supported
by Grants from the M.U.R.S.T. and C.N.R ..
REFERENCES
1. Raphan T, Cohen B. Velocity storage and the ocular response to multidimensional vestibular stimuli. In: Berthoz A, Melvill-Jones G, eds. Adaptive mechanisms in gaze control. Amsterdam: Elsevier Science Publishers BV; 1985: 123-43.
2. Robinson DA. The coordinate of neurons in the vestibular-ocular reflex. In: Berthoz A, Melvill-Jones G, eds. Adaptive mechanisms in gaze control. Amsterdam: Elsevier Science Publishers BV; 1985:297-311.
3. Precht w. The functional synaptology of brainstem oculomotor pathways . In: Baker R, Berthoz A, eds. Control of gaze by brain stem neurons. Amsterdam, New York: Elsevier/North-Holland Biomedical Press; 1977: 131-41.
4. Cohen B. The vestibulo-ocular reflex arc. In: Kornhuber HH, ed. Handbook of sensory physiology, vol. 6, part 1. New York: Springer-Verlag; 1974:478-516.
5. Wilson VJ, Peterson BW. Vestibular and reticular projection to the neck. In: Peterson BW, Richmond FJ, eds. Control of head movement. New York: Oxford University Press; 1988:129-40.
6. Wilson VJ, Yoshida M. Bilateral connections between
labyrinths and neck motoneurons. Brain Res. 1969; 13:603-7.
7. Gernandt BE. Vestibulo-spinal mechanisms. In: Kornhuber HH, ed. Handbook of sensory physiology, vol. 6, part 1. New York: Springer-Verlag; 1974: 541-61.
8. Abzug C, Maeda M, Peterson BW, Wilson VJ. Cervical branching of lumbar vestibulospinal axons. J Physiol (Lond). 1974;243:449-522.
9. Pompeiano O. Vestibulospinal relations: Vestibular influences on gamma motoneurons and primary afferents . In: Brodal A, Pompeiano 0, eds. Basic aspect of central vestibular mechanisms. Progress in Brain Res, vol. 37. New York: Elsevier; 1972;197-232.
10. Mamell 0, Tolu E. Labyrinthine volleys to the tongue motor nucleus. IRCS Med Sci. 1985; 13:785-6.
11. Mamell 0, To1u E. Vestibular ampullar modulation of hypoglossal neurons. Physiol Behav. 1986;37: 773-5.
12. Mamell 0, Tolu E. Hypoglossal responses to macular stimulation in the rabbit. Physiol Behav. 1987; 39:273-5.
13. Mamell 0, Tolu E, Metis F, Carla MA. Labyrinthine

Vestibular Input to Masseter Motoneurons
projection to the hypoglossal nucleus. Brain Res Bull. 1988; 20:83-8.
14. Hopf HC. Vertigo and masseter paresis. A new local brainstem syndrome probably of vascular origin. J Neurol. 1987;235:42-5.
15. Hickenbottom RS, Bishop B, Moriarty MT. Effects of whole-body rotation on masseteric motoneuron excitability. Exp Neurol. 1985;89:442-53.
16. Mameli 0, Tolu E. Involvement of the inferior olivary neurons in vestibular compensation. Brain Res Bull. 1986;16:739-43.
17. Tolu E, Mameli 0, Azzena MT, Azzena GB. Dynamic responses of vestibular cells during spinal decompensation. Physiol Behav. 1980;25:637-40.
18. Azzena GB, Mameli 0, Tolu E. Vestibular nuclei of hemilabyrinthectomized guinea pigs during decompensation. Arch Ital BioI. 1976; 114:389-98.
19. Precht W, Shimazu H, Markham CH. A mechanism of central compensation of vestibular function following hemilabyrinthectomy. J Neurophysiol. 1966; 29:996-10 10.
20. LadpJi R, Brodal A. Experimental studies of the commissural and reticular formation projections from the vestibular nuclei in the cat. Brain Res. 1968;8:65-96.
21. Duensing F, Schaefer KP. Die Aktivitiit einzelner Neurone der Formatio reticularis des Nicht geffesseiten Kaninchens bie Kopfwendungen und vestibuliiren Reizen. Arch Psychiatr Ztg Ges Neurol. 1975;201: 97-122.
22. Spyer KM, Ghelarducci B, Pompeiano O. Responses of brain stem reticular neurons to tilting. Brain Res. 1973;56:321-6.
23. Peterson BW, Filion M, Felpel LP, Abzug C. Response of medial reticular neurons to stimulation of the vestibular nerve. Exp Brain Res. 1975;22:335-50.
24. Peterson BW, Abzug C. Properties of projections from vestibular nuclei to medial reticular formation in the cat. J Physiol. 1975;38:1421-35.
25. Chandler SH, Goldberg LJ. Effects of ponto-medullary reticular formation on the neuronal networks responsible for rhythmical jaw movements in the guinea pig. J Neurophysiol. 1988;59:819-32.
26. Gurahian SM, Chandler SH, Goldberg LJ. Intracellular analysis of trigeminal motoneuron rhythmical activity during stimulation of pontomedullary retic-
171
ular formation in anesthetized guinea pig. J Neurophysiol. 1989;62:1225-36.
27. Nakamura Y, Takatori M, Nozaki S, Kikuchi M. Monosynaptic reciprocal control of trigeminal motoneurons from the medial bulbar reticular formation. Brain Res. 1975;89:144-8.
28. Sessle BJ. Modulation of alpha and gamma trigeminal motoneurons by various peripheral stimuli. Exp Neurol. 1977;54:323-39.
29. Westberg KG, Olsson KA. Integration in trigeminal premotor interneurones in the cat. 1. Functional characteristics of neurones in the subnucleus of the oral nucleus of the spinal trigeminal tract. Exp Brain Res. 1991;84:102-14.
30. Baker JF, Perlmutter SI, Peterson B. Spatial and temporal properties of the vestibulocollic reflex in decerebrate cats. J Neurophysiol. 1985;54:735-56.
31. Berthoz A, Anderson JH. Frequency analysis of vestibular influence on extensor motoneurons: II Relationship between neck and forelimb extensor motoneurons. Brain Res. 1971;376-80.
32. Dutia MB, Price RF. Interaction between the vestibulocollic reflex and the cervicocollic stretch reflex in the decerebrate cat. J Physiol (Lond). 1987;387: 19-30.
33. Miller AD, Roossin PS, Schor RH. Roll tilt reflexes after vestibulospinal tract lesions. Exp Brain Res. 1982;48: 107-12.
34. Wilson VJ, Peterson BW, Fukushima K, Hirai N, Uchino Y. Analysis of vestibulocollic reflexes by sinusoidal polarization of vestibular afferent fibers. J Neurophysiol. 1979;42:331-46.
35. Torvik A. Afferent connections to the sensory trigeminal nuclei, the nucleus of the solitary tract and adjacent structures. An experimental study in the rat. J Comp Neurol. 1956;106:51-142.
36. Phelan KD, Falls WM. The spinotrigeminal pathway and its spatial relationship to the origin of trigeminospinal projections in the rat. Neuroscience. 1991; 40:477-96.
37. Ruggiero DA, Ross CA, Reis OJ. Projections from the spinal trigeminal nucleus to the entire length of the spinal cord in the rat. Brain Res. 1981;225:225-33.
38. Burton H, Loewy AD. Projections of the spinal cord from medullary somatosensory relay nuclei. J Camp Neural. 1977;173:773-92.