the of chemistry no. july 1, in molecular cloning, primary … · 2001-06-28 · the journal of...
TRANSCRIPT

THE JOURNAL OF B I O ~ I C A L CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269, No. 26, Issue of July 1, pp. 17713-17722, 1994 Printed in U.S.A.
Molecular Cloning, Primary Structure, and Characterization of Two Members of the Mammalian Electroneutral Sodium-(Potassium)- Chloride Cotransporter Family Expressed in Kidney*
(Received for publication, January 15, 1994, and in revised form, April 19, 1994)
Gerard0 GambaS, Akihiko MiyanoshitaSn, Michael Lombardi, Jonathan Lytton, Wen-Sen Lee, Matthias A. Hediger, and Steven C. Hebertll From the Haruard Center for Study of Kidney Disease, Haruard Medical School and the Laboratory of Molecular Physiology and Biophysics, Renal Diuision, Department of Medicine, Brigham and Women’s Hospital, Boston, Massachusetts 02115
Electrically silent Na+-(K+)-Cl- transporter systems are present in a wide variety of cells and serve diverse physiological functions. In chloride secretory and ab- sorbing epithelia, these cotransporters provide the chlo- ride entry mechanism crucial for transcellular chloride transport. We have isolated cDNh encoding the two ma- jor electroneutral sodium-chloride transporters present in the mammalian kidney, the bumetanide-sensitive Na+- K+-Cl- symporter and thiazide-sensitive Na+-Cl- cotrans- porter, and have characterized their functional activity in Xenopus laevis oocytes. Despite their differing sensi- tivities to bumetanide and thiazides and their different requirements for potassium, these -115-kDa proteins share significant sequence similarity (-60%) and exhibit a topology featuring 12 potential membrane-spanning helices flanked by long non-hydrophobic domains at the NH, and COOH termini. Northern blot analysis and in situ hybridization indicate that these transporters are expressed predominantly in kidney with an intrarenal distribution consistent with their recognized functional localization. These proteins establish a new family of Na+-(K+)-Cl- cotransporters.
In many epithelial and non-epithelial cells, sodium and chlo- ride movement are interdependent, providing a efficient means of directly coupling the energetically favorable entry of sodium into cells with that of chloride. All of the sodium-chloride- coupled transport (Na+-Cl- cotransport) systems identified so far are electrically silent and can be grouped according to whether K+ accompanies Na+-Cl- cotransport and according to
* This work was supported by grants from the American Heart Asso- ciation Massachusetts Affiliate and the Mexican Council of Science and Technology (CONACYT) (to G. G.), by a grant from the American Heart Association Massachusetts Affiliate (to A. M.), by a grant from the Juvenile Diabetes Foundation International (to W.-S. L.), and by grants from the National Institutes of Health (to S. C. H., M. H., and J. L.). A preliminary report of some of these studies has been presented (Gamba et al., 1993a). The costs of publication of this article were defrayed in
hereby marked “advertisement” in accordance with 18 U.S.C. Section part by the payment of page charges. This article must therefore be
1734 solely to indicate this fact.
to the GenBankmIEMBL Data Bank with accession number(s) U10096 The nucleotide sequence(s) reported in this paper has been submitted
and U10097. $ The contributions of the first two authors should be considered as
equal. 5 Present address: Dept. of Nephrology, Instituto Nacional de la Nu-
trici6n Salvador ZubirBn, Tlalpan 14000, Mexico City, Mexico. ll Present address: Dept. of Pediatrics, Hokkaido University School of
Medicine, Sapporo 060, Japan. 11 To whom correspondence should be addressed: Dept. of Medicine,
Brigham and Women’s Hospital, 76 Francis St., Boston, MA 02115.
their sensitivities to specific classes of inhibitors (i .e. “high ceiling” (or “loop”)-type diuretics of the sulfamoyl-benzoic acid class such as bumetanide or benzothiadiazine (or thiazide)- type agents like hydrochlorothiazide) (as detailed in the fol- lowing reviews (Geck and Heinz, 1986; O’Grady e t al., 1987; Palfrey and O’Donnell, 1992; Saier and Boyden, 1984; Stokes, 1989)).
Geck and co-workers (Geck et al., 1980) provided the first clear evidence that the movement of Na’ and C1- in Ehrlich ascites tumor cells was tightly coupled to that of K+ with a stoichiometry of lNa+-lK+-2Cl-. Since that initial discovery, functionally similar electroneutral cotransporters, exhibiting both a requirement for potassium and sensitivity to bumet- anide, have been found in many different cell types including epithelial cells in the thick ascending limb of the loop of Henle in the mammalian kidney (Geck and Heinz, 1986; O’Grady et al., 1987; Palfrey and O’Donnell, 1992; Saier and Boyden, 1984). Potassium is not universally required, however, for all sodium-chloride-coupled transport systems. A number of stud- ies have suggested that a bumetanide-sensitive Na+-Cl- co- transporter may also be expressed in some cells under certain conditions (Alvo et al., 1985; Hoffman and Simonsen, 1989; Sun et al., 1991). Moreover, a second type of electroneutral and K+-independent Na+-Cl- cotransporter has been functionally identified in the distal tubule of the mammalian kidney (Beau- mont et al., 1989; Costanzo, 1985; Ellison et al., 19871, the urinary bladder of the teleost Pseudopleuronectes americanus (Stokes, 1984), and several non-renal tissues (Calder et al., 1992; Cremaschi e t a l . , 1992; Drewnowska and Baumgarten, 1991). This latter Na+-Cl- cotransporter is insensitive to bu- metanide but highly sensitive to the thiazides.
The major differences in potassium requirements and inhibi- tor (diuretic) sensitivities among these Na+-(K+)-Cl- cotrans- port systems, the diversity in molecular weights of burnetanide/ furosemide-binding (Feit et al., 1988; Jessen et al., 1989; Jorgensen et al., 1984) and [3H]4-benzoyl-5-sulfamoyl-3-(3-the- ny1oxy)benzoic acid-photolabeled proteins (Haas et al . , 1991; Haas and Forbush, 1987a, 198713; Lytle and Forbush, 1992b), as well as the substantial variations in both ion kinetic prop- erties and inhibitor sensitivities of these cotransporters among several cells or tissues (Geck and Heinz, 1986; O’Grady et al., 1987; Palfrey and O’Donnell, 1992; Saier and Boyden, 1984), strongly suggested that different isoforms of the cotransporter may exist or that the cotransporter may be composed of mul- tiple subunits (see Haas (1989)). Although several strategies have been employed in attempts to identify the molecular fea- tures of the proteins that mediate electroneutral Na+-(K+)-Cl- cotransport (Burnham et al., 1990; Feit et al., 1988; Jessen et al., 1989; Lytle and Forbush, 1992a), the nucleotide and amino acid sequence of only one of these cotransporter proteins has
17713

17714 l b o Electroneutrul Nu'-(K+)-Cl- Cotrunsporters in Kidney
been reported to date. We recently isolated by expression clon- ing a 3.7-kb' cDNA, flTSC (Gamba et al., 1993b) encoding the thiazide-sensitive lNa+-lCl- cotransporter present in the win- ter flounder (l? arnericanus) urinary bladder (Stokes, 1984).
Using an approach based on homology to flTSC, we now report the isolation, sequence, and functional expression of cDNAs encoding the thiazide-sensitive Na+-Cl- and the bumet- anide-sensitive Na'-K+-Cl- cotransporters from rat kidney. These molecules, together with the Na+-Cl- cotransporter from the flounder urinary bladder, which exhibit a significant ho- mology consistent with a common ancestry, define a new family of electroneutral Na+-(K+)-Cl- cotransporters. The kidney-spe- cific distribution of transcripts encoding these cotransporters suggests that other, probably related, genes encode non-renal Na+-(K+)-Cl- cotransporters.
EXPERIMENTAL. PROCEDURES Extraction and Selection of Cellular RNA-Total RNA was extracted
from several tissues of adult male Sprague-Dawley rats using the guanidine/isothiocyanate method. Poly(A)+ RNA from renal superficial cortex, cortex, inner stripe of the outer medulla, inner medulla, and papilla was selected with oligo(dT)-cellulose affinity chromatography (type 7, Pharmacia Biotech).
Construction and Screening of cDNA Libraries-To isolate rTSC1, a rat renal cortex non-directional cDNA library was constructed using the plasmid pcDNAII (Invitrogen), and screened under low stringency con- ditions (hybridized in 40% formamide, 4 x SSC, 4 x Denhardt's solution, 0.5% sodium dodecyl sulfate (SDS), 200 pg/ml salmon sperm DNA, for 12 h at 37 "C and washed in 1 x SSCP, 0.1% SDS at 42 "C) using a randomly primed 32P-labeled DNA probe derived from the BamHI frag- ment of flTSC (nucleotides 1194-2543; Gamba et al. (199313)). Several partial cDNA clones were isolated. Then, a directional, size-selected (3.0-9.4-kb) rat renal superficial cortex cDNA library was constructed using the plasmid pSPORTl (Superscript, Life Technologies, Inc.) and was screened under high stringency conditions (hybridized in 50% form- amide, 5 x SSCP, 2% SDS, 0.1% N-lauroylsarcosine, 5% blocking rea- gent (Boehringer Mannheim) at 65 "C, and washed at 65 "C in 0.1 x SSCP), using a digoxigenin-UTP riboprobe constructed from the 5' end of the previously isolated clones, until a full-length clone of 4,394 bp was isolated. To isolate rBSC1, a directional cDNA library was constructed into pSPORTl (Superscript) using poly(AY RNA prepared from rat renal inner stripe of outer medulla. The library was size-selected to 3-7 kb, electroporated into Escherichia coli (DHlOB cells, Life Technologies, Inc.), and screened under low stringency conditions (as above) using the randomly primed 32P-labeled DNA probe derived from the BamHI- BamHI fragment of flTSC. Two overlapping clones were eventually isolated and used to construct a full-length cDNA clone of 4,612 bp.
cDNA Sequencing and Computer Analysis-Several restriction frag- ments of rTSCl and rBSCl were subcloned into pBluescript (Strat- agene) or pSPORTl (Life Technologies, Inc.). Both strands of the sub- cloned cDNA inserts were sequenced by the dideoxy chain termination method using a Sequenase version 2.0 DNA sequencing kit (U. S. Bio- chemical Corp.). Internal regions of subcloned fragments were se- quenced using 17-mer primers. Sequencing reactions were run on 5% polyacrylamide gels. Analysis of nucleotide and amino acid sequence was performed using the GENEWORKS software package (Intelli- Genetics, version 2.2.1).
Oocyte Procurement and cRNA Injection-Clusters of oocytes were surgically harvested from adult female Xenopus laevis (Nasco) anesthe- tized with 0.17% tricaine and incubated in Ca2+-free Ringer (82.5 mM NaC1, 2 mM KCl, 1 mM MgCl,, 5 mM HEPES, pH 7.4) for 1 h (Gamba et al., 1993b). Then oocytes were manually separated and incubated in modified Barth's Ringer (ND96) (in m: 96 NaCl, 2 KCI, 1.8 CaCl,, 1 MgCI,, and 5 HEPES/Tris, pH 7.4) containing collagenase (Boehringer Mannheim type B, 2 mg/ml) for 90 min. After collagenase treatment, the remaining follicular layer was removed using fine forceps. Stage V-VI oocytes were incubated overnight at 17 "C in ND96. Surviving oocytes (>95%) were then injected with 50 nl of H,O or with 50 nl of cRNA-
NotI-linearized rBSCl or rTSCla or rTSClb cDNA, using T7 RNA containing solution (0.1-0.5 pg/pl). cRNAwas transcribed in uitro, from
The abbreviations used are: kb, kilobase pair(s); bp, base pair(s); UTR, untranslated region; PCR, polymerase chain reaction; DIDS, 4,4'-
Henle; NMDGCl, methyl-o-glucamine chloride. diisothiocyanostilbene-2,2'-disulfonic acid; TAL, thick ascending limb of
polymerase (Stratagene) in the presence of Cap analog (7-methyl-G~- ppG, Pharmacia). Before injection, cRNA samples were denatured by heating at 65 "C for 2 min. Injected oocytes were incubated at 17 "C for 4 days in ND96 containing gentamicin (5 mg/dl) and sodium pyruvate (2.5 mM). To inhibit electroneutral Na+-(K+)-Cl- cotransport and reduce cell C1- activity (thereby enhancing the driving force for tracer 86Rb+ or "Na+ uptake), oocytes were incubated for the last 24 h in a Cl--free ND96 medium (containing, in mM: 96 Na' isethionate, 2 K+ gluconate, 1.8 Ca" gluconate, 1 Mg2' gluconate, 5 Hepes/Tris, 5 mg/dl gentamicin, and 2.5 Na' pyruvate, pH 7.4).
Functional Expression-Functional expression of rTSC was assessed by measuring "Na+ uptake in oocytes injected with H,O or with rTSCla (or rTSClb) cRNA using the following protocol (Gamba et al., 1993b); a 30-min preincubation in C1"free medium (CI- was replaced by gluco- nate) that also contained 1 mM ouabain, 100 p~ amiloride and 100 p~ bumetanide was followed by a 60-min uptake period in isotonic uptake medium (in mM: 58 NaCI, 38 methyh-glucamine chloride (NMDGCI), 2 KCI, 1.8 CaCI,, 1.0 MgCI,, 5 H e p e f i s , pH 7.4), in the presence of the same drugs and 20 pCi/ml ,,Na+ (DuPont NEN).
Functional expression of rBSCl was assessed by measuring 86Rb+ uptake in groups of oocytes injected with H,O or with rBSCl cRNA using the following protocol. Oocytes were incubated for 30 min in a hypotonic Na+K+-free solution (in mM: 75 NMDGCl, 1.8 CaCI,, 1 MgCI,, 5 Hepesms , pH 7.4) to inhibit the oocyte endogenous Na-K-Cl cotrans- porter (see "Results"). This was followed by a 15-min incubation in the same solution containing 1 mM ouabain (to inhibit oocyte endogenous Na+-K+-ATPase), and finally a 60-min uptake period in a hypotonic (in mM: 58 NaCI, 2 KC1, 10 BaC1, 1.8 CaCl,, 1 MgCI,, 5 Hepes/Tris, pH 7.4) or isotonic (same solution + 38 mM NMDGCI) uptake medium, contain- ing 1 mM ouabain and 20 pCi/ml 86Rb+ (DuPont NEN). To study the C1- and Na' dependence of 86Rb+ uptake, GI- was replaced by gluconate and Na' by NMDG in the uptake medium.
All uptakes were performed at 30 "C and were linear over the first 60 min. At the end of the uptake period, oocytes were washed three times in ice-cold uptake solution without isotope and then centrifuged through oil (1:l dioctylphthalate and silicone; Aldrich) to remove extra- cellular fluid. After the oocytes were dissolved by overnight incubation in 10% sodium dodecyl sulfate (Sigma), tracer activity was determined by p-scintillation counting.
In Vitro Danslation-0.5 pg of each cDNA (rBSC1 and rTSCla) was translated in vitro using nuclease-treated rabbit reticulocyte lysate for 60 min at 30 "C in the absence or presence of canine pancreatic micro- somal membranes (Promega). In the absence of membranes, 0.5% Tri- ton X-100 was included in the reaction mixture. Endoglycosidase H (Boehringer Mannheim) treatment was performed in 2 pl of translate in the presence of 0.1 M sodium acetate and 1% Triton X-100. Protein products were resolved by Laemmli SDS-polyacrylamide gel electro- phoresis (6-8% acrylamide).
Northern Blot Analysis-For Northern blot analyses, 20 pg of total RNA from tissues or 2.5 pg of poly(A)' RNA from renal cortex, outer medulla, inner medulla, or papilla was resolved by electrophoresis (5% formaldehyde, 1.0% agarose gel) and transferred to a nylon membrane (Stratagene). Quality of RNA was assessed by ethidium bromide stain- ing. The conditions used for high stringency were: hybridization at 65 "C in 5 x SSCP, 50% formamide, 2% SDS, 0.1% N-lauroylsarcosine, 5% blocking reagent (Boehringer Mannheim), followed by washing in 0.1 x SSCP, 0.1% SDS at 65 "C. The conditions used for low stringency were: hybridization a t 60 "C in 5 x SSCP, 45% formamide, 2% SDS, 0.1% N-lauroylsarcosine, 5% blocking reagent (Boehringer Mannheim), fol- lowed by washing in 0.5 x SSCP, 0.1% SDS at 50 "C. The probes used were digoxigenin-UTP-labeled riboprobe (Boehringer Mannheim) con- structed by in vitro transcription using SP6 RNA polymerase from a BamHI-BamHI fragment of flTSC or from an EcoRI-digested rBSC 1 or rTSCla. Hybridization was detected by chemiluminescence (Boeh- ringer Mannheim).
In Situ Hybridization: Autoradiograms-The intrarenal localization of rBSCl and rTSCl transcripts was determined in the rat kidney by in situ hybridization using methods similar to those described previously (Kanai et al., 1992). 35S-UTP-labeled, sense and antisense, riboprobes were synthesized with T7 and SP6 RNA polymerase from full-length rBSCl and rTSCla cDNAs. RNA probes were degraded by partial hy- drolysis for 80 min at 60 "C, forming products of -100 base pairs. Paraformaldehyde-fixed sections of rat kidney were prepared, hybrid- ized to probe, and washed at high stringency. The hybridized tissue slices were exposed to the film for -24 h.

Two Electroneutral Na+-(K+)-Cl- Cotransporters in Kidney 17715
Mouse Rat
IM OM C IM OM C kb 7.5 '
4.4 '
2.4
FIG. 1. Identification of transcripts in rat and mouse kidneys homologous to the thiazide-sensitive Na+-Cl- cotransporter cloned from flounder urinary bladder. Low stringency Northern blot of poly(A)'RNA isolated from rat and mouse kidney inner medulla ( I M ) , inner stripe of outer medulla (OM), and cortex ( C ) using a probe generated from the flTSC cotransporter. Figure shows 2.5 pg of poly(AY RNMane; the autoradiograph was developed after 6 h of exposure at room temperature. RNA molecular weight markers are indicated.
RESULTS
Molecular Cloning We employed a homology-based strategy to isolate cDNAs
encoding Na+-(K+)-Cl- cotransporters form rat kidney. Fig. 1 shows a Northern blot of poly(A)+ RNA isolated from the cortex, inner stripe of outer medulla, and inner medulla of rat and mouse kidney performed under low stringency conditions using a riboprobe constructed from a BamHI fragment of flTSC (nucleotides 1194-2543 encoding amino acids 363-813) cDNA (Gamba et al., 199313). Consistent with localization of the thia- zide-sensitive Na+-Cl- cotransporter in the cortical distal con- voluted segment of the nephron (Beaumont et al., 1989; Costanzo, 1985; Ellison et al., 1987; Stokes, 1989; Velazquez and Wright, 19861, three transcripts of -3.3, -4.5, and -8.0 kb were observed in rat and mouse renal cortex. Two additional transcripts (-4.6-kb transcript in both rat and mouse, and a -3.3-kb transcript in mouse, but not rat) were detected in the inner stripe of outer medulla, a kidney region not known to express the thiazide-sensitive Na+-Cl- cotransporter (Kikeri et al., 1990). Alarge percentage of the renal outer medulla cellular mass, however, is composed of thick ascending limb of Henle segments, which exhibit a high functional activity of the bu- metanide-sensitive Na+-K+-Cl- cotransporter (Burg et d., 1973; Greger, 1985; Hebert and Andreoli, 1984), suggesting the pos- sibility that the -4.6-kb transcript common to both mouse and rat might encode this latter cotransporter.
To isolate rat renal cortical cDNAs homologous to flTSC, we first screened a non-directional cDNA library under low strin- gency conditions using a randomly primed 32P-labeled DNA probe derived from the coding region of flTSC (Gamba et al., 1993b). Several partial-length clones were isolated, which were lacking poly(A) tails and ATG initiation codons. Two full-length clones of -4.4 kb (rTSCla) and -3.3 kb (rTSClb) were even- tually isolated by screening a second, oligo(dT)-primed and size-selected (3-9.4 kb) cDNAlibrary (pSPORT1, Life Technolo- gies, Inc.) from rat "superficial" renal cortex at high stringency with a digoxigenin-UTP riboprobe derived from the 5' end of one of the partial clones. Sequencing of the 4,394-bp rTSCla cDNA yielded a 3,018-bp open reading frame, with a transla- tion initiation codon assigned to the first in-frame ATG con- tained within a Kozak consensus sequence (Kozak, 1991), C A T A m , starting at nucleotide 9. The 1343-bp 3"untrans- lated region (UTR) of rTSCla contained the polyadenylation sequence (Manley, 1988) AATAAA, located 19 bases upstream from the poly(A) tail. The nucleotide sequence of the 3,315-bp rTSClb cDNA was identical to rTSCla except for a truncated 3"UTR of 231 bp.
A 5.0 r
+ z m
0
P I 0 P0lyTZ
B 10 .
al A? m P 3 c
+ 5 '
2
0 -
,:, , I, + - + + - + " + " +
[K+l0 + - - + + + + + Burnet + + - + + + + +
zzNa+ uptake (expressed as nmolloocyteh) in oocytes injected with wa- FIG. 2. Functional expression of rTSC in X. laevis oocytes. A,
ter (H,O, open burs) or with rTSCla cRNA (cRNA, solid burs). Expres-
of extracellular C1- ([Cl-]J or in the presence of 100 PM concentration of sion was assessed under isotonic conditions, in the presence or absence
the thiazide, polythiazide (PoZyTZ). :2 indicatesp c 0.05 from the control + [Cl-I,, - polythiazide group. B, effect of extracellular K+ ([K+l0) and transport inhibitor agents on "Na' uptake in oocytes injected with rTSCla cRNA (12.5 ngloocyte). Extracellular K+ ([K+], = 5 mM) and 100 PM bumetanide (Burnet) were present in all groups, except where noted (-1. The uptakes shown by the striped burs were also performed in the presence of one of the following transport inhibitors (100 PM): the thia- zides metolazone (MTZ), hydrochlorothiazide (HCTZ), chlorothiazide (CTZ), or polythiazide (PolyTZ), or the stilbene DIDS. Each bur repre- sents mean -c S.E. of 20 oocytes from a single toad. * indicates p c 0.05 from the control + [K'],, + bumetanide group.
We also screened a directional, size-selected cDNA library from rat renal inner stripe of outer medulla under low strin- gency conditions using the same flTSC BamHI fragment to isolate renal outer medullary cDNAs homologous to flTSC. Two different groups of clones with overlapping cDNA inserts were isolated and used to construct a full-length -4.6-kb cDNA, rBSC1. We used the reverse transcriptase-based polymerase chain reaction (PCR) to demonstrate that the spliced rBSCl cDNA represented a legitimate reverse transcript from outer medulla. Two pairs of specific primers, corresponding to se- quences on opposite sides of the rBSCl cDNA splice site and with one primer of each pair derived from sequence upstream of the 5' end of the clone forming the 3' end of rBSC1, were used to amplify poly(A)' RNA isolated from rat inner stripe of outer medulla. The PCR product sizes and restriction digest frag- ment patterns from outer medullary mRNA were identical to those observed from the rBSCl cDNA. Sequencing of the 4,612-bp rBSCl clone yielded a 3,308-bp open reading frame with the first in-frame ATG, within a Kozak consensus se- quence ( M G m T ) (Kozak, 19911, 215 bp downstream from the 5' end. The 1045-bp 3'-UTR contained the polyadenylation sequence (Manley, 1988) AATAAA, 14 bases upstream from the poly(A) tail. Comparison of the nucleotide sequences of rBSCl and rTSCla revealed 51% identity in the open reading frames and lower levels of identity in the 5'- and 3"UTRs.

17716 flu0 Electroneutral Na+-(K+)-Cl- Cotransporters in Kidney
A ,o I HYPOTONIC ISOTONIC
Burnet
0 7.5
2 5.0 m a += 2.5 n
I
K 0
Burnet
IClI.
[Na'l.
- + - + - + - +
- + "
+ + + - + + - +
- + - -
+ + + - + + - +
K O rBSC rTSC
FIG. 3. Functional expression of rBSCl in Xenopus Zaeuis
oocyteh) in oocytes injected with water (open bars) or with 12.5 ng of oocytes. A, effect of osmolality on "Rb+ uptakes (expressed as nmol/
rBSCl cRNA (solid bars). 86Rb+ uptakes were measured under hypo- tonic (58 mM NaCI) or isotonic conditions (58 m~ NaCl + 38 mM n- methyl-D-ghcamine, NMDGCI), and in the presence or absence of 100 p~ bumetanide (Burnet). Each bar represents mean S.E. of 20 oocytes from a single toad. B, "Rb+ uptake in oocytes injected with H,O (open bars) or with 12.5 ng of rBSCl cRNA (solid bars). Uptakes were meas- ured using the hypotonic uptake medium in the absence or presence of 100 p~ bumetanide (Burnet) or in hypotonic uptake medium without extracellular Na' (fNa+lo; replaced by NMG') or extracellular C1- UCl-lo; replaced by isethionate) as indicated. C , effect of inhibitors on the s6Rb+ uptake in oocytes injected with water (H,O), or with 12.5 ng of rBSCl or rTSCla cRNA. Uptakes were performed in the absence (open bars) or presence of 100 VM concentration of bumetanide (solid bars), metola- zone (striped bars), or DIDS (hatched bars). DIDS effects were not examined in rTSCla cRNA-injected oocytes. For all experiments, each bar represents mean f S.E. of 20 oocytes.
rTSCla, rTSClb, and rBSC1 Encode Na+-Cl- Cotransporters with Different Functional and
Pharmacological Properties rTSCla Encodes a Thiazide-sensitive Na'-Cl- Cotrans-
porter-As we have shown previously (Gamba et al., 1993b), H,O-injected X. laevis oocytes exhibit a very low rate of zzNa+ uptake that is not affected by C1- removal or addition of the thiazide diuretic, polythiazide (Fig. 2 A ) ; 22Na+ uptakes (nmoV oocyteh) were 0.15 2 0.005 in the presence of C1-, 0.16 2 0.006 in the absence of C1-, and 0.14 2 0.004 in the presence of C1- and 100 p polythiazide. In contrast, oocytes injected with rTSCla cRNA (- 12 ng/oocyte) exhibited significantly higher "Na+ up- takes (4.35 2 0.19 versus 0.15 2 0.05 nmoVoocyteh) that could be nearly completely inhibited by extracellular C1- removal (0.58 0.03 nmoVoocyteh), or by the addition of 100 1.1~ poly- thiazide to the C1"containing uptake medium (0.17 2 0.01 nmol/oocyteh). Consistent results were obtained in two sepa- rate experiments using different toads: 6.07 2 0.43 nmol/ oocyteh (cRNA-injected control; n = 35 oocytes), 0.58 2 0.03 nmol/oocyteh (zero C1-; n = 20), 0.13 2 0.14 nmoVoocyteh (100 p~ thiazide; n = 33). Injection of f fSClb cRNAinto oocytes also gave rise to a large zzNa+ uptake that was inhibited by thia- zides: 13.15 2 0.62 nmolhloocyte (control) versus 0.85 f 0.34 (100 p metolazone). I t is unlikely that the high level of z2Na+
rTSC M-AELPVTEMPGDA- ---LCSGRFT--- IS TLLGGDEPP---PAA CDNSOPSH----LTH 45 rBSC MSVNIPSNSVPSGAS RFOVHVINEGHGSGA AMSDSTDPPHYEETS FGDEAONRLKISFRP 6 0
f lTSC M- - - - - - -ELPGDG- - - -VHLASYGP-RAP OLAVNGKAP"-AGG LDGSGYYQ---CYRD 4 2
rBSC GNOECYENFLOTGET AKTDTTFHAYDSHTN TYYLOTFGHNTMDAV PKIEYYRNTGSVSGP 1 2 0 r T S C G S - - - - T L Y L R - - - - - - - - - - - - - - - - - - - - - - - - T F G Y N T I D V Y PAYEHYANRPLPGEP 7 7 f lTSC GS- - - -SVASRGSD- - - . - - - - - - - - - - - - ----APTGYDTVDAP PHYDFYANTEVWGRR 7 8
rBSC KVNRPSLOEIHEOLA KNVA--VAPGSADRV AN--GDGMPGDEOAE NKEEOVTGVVKFGWV 1 7 6
f l T S C RRVRPSLYOLS--VD PEODDFKPPMYEETA GERMGGGDSSEEEEE EHKEPPPEPPRFGWV 1 3 6 rTSC RKVRPTLADLHSFLK QEGSHLHALAFDGRP GHELTDGLVEDETGA NSEKSPGEPVRFGWV 1 3 7
rBSC KGVLVRCMLNIHGVM LFIRLSWIVGEAGIG L G V I I I G L S V V V T T L TGISMSAICTNGVVR 2 3 6 rTSC KGVMIRCMLNINGVI LYLRLPWITAOAGIV LTWLIILLSVMVTSI TGLSISAISTNGKVK 197 f l T S C OGVMIRCMLNIWGVI LYLRLPWITAOAGIG L T W V I I L L S S F I T G I TGLSTSAIATNGKVK 1 9 6
L
- MI I " P I rBSC GGGAYYLISRSLGPE FGGSIGLIFRFANAV RVAiiVVGFAETVVD LLKESDSMMVDPTND 2 9 6 rTSC SGGTYFLISRSLGPE LGGSIGLIFAFANAV GVAMHTVGFAETVRD LLOENGTPIVDPIND 257 f l T S C GGGTYFLISRSLGPE LGGSIGLIFAFANAV AVAMHTVGFAETVTD
LMRENGVVMVDPP 256 r B S C I R I I G S I T V V I L L G I SVAGMEWEAKAOVIL L V I L L I G I A N F F I G T VIPSNNEKKSRGFFN 3 5 6 I M3 I
r T S C I R I I G V V T V T V L L A I SLAGMEWESKAOVLF FLVIMVSFANYLVGT LIPASKDKASKGFYS 3 1 7 f l T S C I R I V G V I T V T C L L G I SMAGMEWESKAOVLF FLVINVSFVNYIVGT IIPASPDKOAKGFFS 316
rBSC YOASIFAENFGPSFT EGEG-FFSVFAIFFP AATGILAGANISGDL EDPODAIPRGTMLAI 4 1 5 rTSC YHGDIFVONLVPDWR GIDGSFFGMFSIFFP SATGILAGANISGDL KDPAVAIPKGTLMAI 377 f l T S C YKAEIFAANFVPGWR GKEGSFFGMFSIFFP SATGILAGANISGOL KDPTVAIPRGTLMAI 376
r 8 S C FITTVAYIGVAICVR ACVVRDATGSMNDTV --VSGMNCNGSAACG LGYDFSRC-OHEPCO 4 7 2 r T S C FWTTISYLAISATIG SCVVRDASGDVNDTI "TPGPGLCEGLACV HAGTSRNAPOOHSCR 4 3 5 f l T S C F W T T I S Y L I I S A T I G ACVVRDASGELNDTL SYSSSSENCSGLACO YRUDFSECIKNNTCK 4 3 6
"6- L
YGLMNNFOVMSMVSG FGPLITAGIFSATLS YGLINYYOTMSMVSA FAPLITAGIFGATLS HGIMNYYQSMSLVSA FAPLISAGIFGATLS
rBSC
f 1 TSC rTSC
rBSC GYGKNNEPLRGYFLT rTSC GYGKNKEPVRGYLLA f l T S C GYGKNAEPLRAYLLT
rBSC
f 1 TSC rTSC
rBSC rTSC f 1 TSC
WRPAYGIYNMWVSLF I
WRPSFRYYSKWAALF WRPSFRFYSKWLSLL
ALSYVSALDNALELT "11
AGSYNLALSYSVGLN ASSYSMALNOCVGLN
" 8 . F V I A H A F I L I A E L N V
YVIAVCFVLIAELNT Y A I A V A F I I I A E L N T
"9- GAILCCAVMFVINWW GAVISVVIMFLLTWW GAVCCVVIMFLLTWW
1- L ~
TVEDHVKNFRPOCIV
OVEDHVKNYRPOCLV EVEDHIKNYRPDCLV
SALASLVSAPKVFOA LCKDNIFKGLOFFAK SALACLVSAAKVFOC LCEDOLYPLIGFFGK SALACLVSAPKVFOC LCKDOLYPLIGFFGK I __
I A P I I S N F F L A S Y A L INFSCFHASYAKSPG I A P I I S N F F L C S Y A L INFSCFHASITNSPG I A P I I S N F F L C S Y A L INFSCFHASVTNSPG
A A V I T Y V I E L F L Y I Y VTYKKPDVNWGSSTO AALIAIGVVLFLLLY VIYKKPEVNWGSSVO AALIAFGYVFFLLGY TLYKKPAVNWGSSVQ
LTGGPMTRPALLDIT HAFTKNSGLCICCEV LTGPPNFRPALVDFV STFTQNLSLMICGHV LTGPPCCRPALVDLV GCLTKRLSLMMCGHV
L M 1 0 -
"12 I
5 3 2 4 9 5 4 9 6
5 9 2 5 5 5
5 5 6
6 5 2 6 1 5 6 1 6
7 1 2 6 7 5 6 7 6
rBSC FVGPRKLCVKEMNSG MAKKOAWLMKNKIKA FYAAVAADCFRDGVR SLLOASGLGRHKPNT 7 7 2 r T S C LIAPGKORVPELRLI ASGHTKWLNKRKiKA FYSDVIAEDLRSGVO ILMOASGLGRMKPNI 7 3 5 f l T S C VTA-GPSPVSE-R-- ---HVTWLNORKVRS FYRGVVAADLRSGVN MLLDGAGLGRIKPNV 7 2 9
rBSC LVIGYKKNWRKAPLS ELENYVGIIHDAFDF EIGVVIVRISOGFDI SPVLOVODELEKLEO 8 3 2 rTSC LVVGFKKNWQSAHPA TLEDYIGILHDAFDF NYGVCIMRMREGLNV SEALPTHTAPEALVO 7 9 5 f l T S C LLMGFKKDWGCDSPO AAHHYIGILHDAFOL HYGVCVLAVKEGLDA SHPPOCHVNP----- 784
rBSC ERLALEAAIKONDCE EGKGGIRGLFKKAGK LNITK--PAPKKDSN ISTI-OSMHVGEFNO 8 8 9
f l T S C - - - G F D G G P E S - - - - - - - - - - - - - - - - - - - IN-TVCAPACVOTSV TSSV--SMDPDPOP- 818 8 0 5 rTSC EE ............. ............... .--.-..--..Q.. .STIFOS........
rESC KLVEASAOFKKKOGK GTIDVWWLFDDGGLT LLIPYILTLRKKWKO CKLRIYVGGKINRIE 949 r T S C - - - - - - - - - - - E O G K KTIDIYWLFDDGGLT LLIPYLLHRKKRWGK CKIRVFVGGOINRMD 854 f l T S C - - - - -SSVFOKKOGK KTIDIYWLSDDGGLT LLLPYLLTRRKRWAG CYVRVFVGGDTDKKE 8 7 3
rBSC EEKISMASLLSKFRI K F A D I H I I G D I N I K P NKESWKVFEEMIEPY RLHESHKDLTTAE" 1 0 0 7 r T S C EERKAIISLLSKFRL GFHEVHVLPOINOKP OVEHTKRFEDMIAPF RLNDGFKDEATVA" 9 1 2 f l T S C EOKEEVLALIKKFRL GFHDVEVLPDIHOPP OPGNVDHFEDSVNRF RLETNPKODSDSGPO 9 3 3
rBSC KLKRESPWKITDAEL EAVKEKSYROVRLNE LLQEHSRAANLIVLS LPVARKGSISDLLYM 1 0 6 7
f l T S C OOOOEEPWMITEODL ERNRAKSLROIRLNE VLOVHSREAALIVIT MPVGRRGVCPSTLFL 9 9 3 rTSC EMRROYPWKISDEEI NKNRIKSLRQVRLNE ILLDYSRDAALI ILT LPIGRKGKCPSSLYM 970
rBSC AWLEILTKNL-PPVL LVRGNHKNVLTFYS
f l T S C A'iiLDVLSRDLRPPVL LVRGNOENVLTFYCO rTSC AWLETLSOOLSPPVL LIRGNOENVLTFYCQ
1 0 9 5 1 0 0 2 1023
Na+-(K+)-Cl- cotransporter family members. The deduced amino FIG. 4. Deduced amino acid sequences of the electroneutral
acid sequences of the rat renal outer medulla rBSCl (rBSC, top lane), the rat renal cortex rTSCla (rTSC, middle lane), and the flounder urinary bladder TSC (fZTSC, bottom lane; Gamba et al., (199313)) are
lines. Gaps have been added to obtain the best alignment. Amino acid shown. Amino acid residues are numbered on the right of individual
highlighted. Twelve stretches of amino acid sequence (Ml-Ml2) pre- residues that are common for at least two of the cotransporters are
dicted to encode membrane-spanning helices are indicated by brackets.
uptake induced in rTSCla or rTSClb cRNA-injected oocytes was due to either activation or expression of Na+:H+ exchang- ers, epithelial sodium channels, or Na+-K+-ATPase since amilo- ride (100 PM) and ouabain (1 mM) were present in all uptake media.
We also assessed the potassium dependence and transport inhibitor sensitivity of "Na+ uptake in X. laevis oocytes ex- pressing f lSCla . The rate of "Na' uptake in LlSCla cRNA- injected oocytes was unaffected by removal of extracellular K+ or by the addition of 100 p~ bumetanide in the presence or absence of extracellular K+ (Fig. 2 B ) (nmoVoocyteh: 9.25 2 0.72 (- [K'],, 100 PM bumetanide) uersus 9.94 2 0.63 (- [K+] , ) , p = not significant; 8.36 2 0.55 (+[K+],, 100 PM bumetanide) versus 9.25 2 0.72 (- [K'],, +lo0 1.1~ bumetanide), p = not significant). Furthermore, rTSCla cRNA-injected oocytes did not express any significant bumetanide- or thiazide-sensitive "Rb+ uptake (Fig. 3C). When taken together, these results show that

%o Electroneutral Na'-(K+)-Cl- Cotransporters in Kidney 17717
FIG. 5. The electroneutral Na+-(K+)- C1- cotransporter proteins belong to a new family of proteins. Hydrophobic- ity profiles of the deduced amino acid se- quences of flTSC (A), rTSCla ( B ) , and rBSCl ( C ) according to the Kyte-Doolittle algorithm (Kyte and Doolittle, 1982) us-
N-linked glycosylation sites are indicated ing a window of 11 amino acids. Potential
(A). D and E, models for rTSCla and BSCl Na+-(K)+-Cl- cotransporters with 12
tential CAMP-dependent protein kinase- putative membrane-spanning helices. Po-
dependent ( +) and protein kinase C-de- pendent (m) phosphorylation sites are shown. The N-linked glycosylation sites between transmembrane spans 7 and 8 are indicated by the branched structures.
A flTSC 1023 amino acid residues
3-
0-
-3- I
200 400 600 800 1000
B rTSC 1002 amino acid residues
3-
0-
A -34 I I I 1 I I I I I I
200 400 600 800 1000
C rBSC 1095 amino acid residues
3- I 1 5 3 4 Ls 6 7 8 9 . 1 1 . 1 2 I
0-
-3 I I I I I I I I I I A A
I
200 400 600 800 1000
rTSCla and, likewise, rTSClb cDNAs encode for a thiazide- sensitive Na+-Cl- transporter that does not require potassium and is insensitive to high ceiling diuretics.
"Na+ uptake in rTSCla cRNA-injected oocytes could be in- hibited to varying degrees by several different benzothiadia- zine derivatives (or thiazides; see Fig. 2). The relative inhibi- tion of C1--dependent "Na+ uptake by these thiazides (100 ) 1 ~ ) was polythiazide = metolazone > hydrochlorothiazide > chlo- rothiazide. This diuretic profile is similar to that previously shown for inhibition C1--dependent Na' absorption oocytes in- jected with cRNA transcribed from flTSC (Gamba et al., 1993b), for inhibition of C1--dependent Na' absorption (measured as the short circuit current) in the flounder urinary bladder (Li et al., 19901, and for thiazide competition for the high-affinity [3H]metolazone binding site on kidney cortical membranes (Beaumont et al., 1988). As a group the thiazides appeared to be more potent inhibitors of the mammalian rTSC cotransporter than the flounder flTSC cotransporter (Fig. 2 B ; Gamba et al. (1993b)), e.g. CTZ inhibited "Na+ uptake by 83% in oocytes expressing rTSC uersus 52% for flTSC. Interestingly, "Na+ up- take observed in rTSCla cRNA-injected oocytes was also inhib- ited by 100 p DIDS, an inhibitor of anion exchange processes like Cl-:HCO; exchange (Cabantchik and Rothstein, 1972; Kopito and Lodish, 1985) (see Fig. 2B). In this regard, stilbene
disulfonates have also been shown to reduce irreversibly the binding of [3Hlmetolazone to rat renal cortical membranes (ICsO
380 ) 1 ~ ) at a site(s) different than the metolazone-anion bind- ing site (Luo and Fanestil, 1991; Tran et al., 1990).
rBSCl Encodes for a Burnetanide-sensitiue Na+-K+-CI- cotransporter-X. laeuis oocytes express an endogenous Na+- K+-Cl- cotransporter activity (Shetlar et al., 1990) that medi- ates cell volume regulatory responses to changes in extracellu- lar osmolality. Our preliminary experiments, as well as those reported by Shetlar and Kinne (1990) and by Suvitayavat et al. (1994), indicated that the bumetanide-sensitive Na+-K+-Cl- co- transporter inX. laeuis oocytes could be activated by hypertonic cell shrinkage and inhibited by hypotonic cell swelling. In ad- dition, we had previously shown that hypotonicity has little effect on apical Na+-K+-Cl- cotransporter activity in the mam- malian medullary thick ascending limb of Henle (Hebert and Sun, 1988). Accordingly, we assessed the effects of extracellular osmolality on bumetanide-sensitive 86Rb+ uptake in H,O-in- jected or rBSCl cRNA-injected oocytes. Fig. 3A shows "Rb+ uptakes performed using a hypotonic or isotonic medium in oocytes obtained from a single toad. It is evident that bumet- anide-sensitive "jRb+ uptake in H,O-injected oocytes is almost abolished under hypotonic conditions (in nmol/oocyte/h: 0.09 5
0.005 (hypotonic uptake medium) uersus 0.88 2 0.08 (isotonic

17718 lho Electroneutral Na+-(K+)-Cl- Cotransporters in Kidney
uptake medium)). In contrast, total ssRb+ uptake in rBSCl cRNA-injected oocytes in the hypotonic medium was -40-fold higher than in H,O-injected oocytes (0.12 2 0.09 (H,O) versus 4.52 2 0.23 (rBSC) nmoYoocyteh, p < 0.01) and was virtually completely inhibited by 100 p~ bumetanide (0.17 2 0.002 nmoV oocyteih). Consistent results were obtained in three separate experiments using different toads: (in nmoVoocyteih) 3.89 2 0.12 (cRNA-injected control; n = 65 oocytes) and 0.24 2 0.01 nmol/oocyte/h (cRNA-injected oocytes with 100 PM bumetanide, n = 63); 0.15 2 0.01 nmoYoocyteih (H,O-injected control; n = 52 oocytes), and 0.04 -c 0.01 nmoYoocyteh (H,O-injected oocytes with 100 JIM bumetanide; n = 46 oocytes). In addition, bumet- anide-sensitive 86Rb+ uptake in rBSCl cRNA-injected oocytes was only -2-fold higher when performed in isotonic uptake media (8.9 2 0.6 (isotonic) versus 4.3 2 0.23 (hypotonic) nmoV oocyteih) compared with an -9-fold increase in tracer uptake observed in H,O-injected oocytes for a similar change in osmo- lality. These results demonstrate that the hypotonic protocol can be used to distinguish the rBSCl cotransporter activity from that endogenous to the oocyte.
We assessed the specific ionic requirements and inhibitor drug sensitivities of 86Rb+ uptake in rBSCl cRNA-injected X. laevis oocytes using the hypotonic protocol. In these experi- ments (Fig. 3B), oocytes injected with rBSCl cRNA exhibited an -130-fold higher bumetanide-sensitive 86Rb+ uptake than water-injected oocytes (6.68 2 0.41 versus 0.05 2 0.003 nmoY oocyteih). In addition, the increased tracer uptake in rBSCl cRNA-injected oocytes could be nearly completely inhibited by the addition of 100 PM bumetanide to the uptake medium (0.27 5 0.003 nmoYoocytek) or by extracellular Na+ or C1- deletion (0.08 2 0.04 and 0.02 2 0.01 nmolloocyteih, respectively). 10 and 1 p~ bumetanide reduced ,,Na+ uptake by 89% and 44% of that observed with a 100 p~ concentration of this diuretic (n = 14 oocytes), suggesting a K,, for bumetanide of about 1 p ~ . In contrast, s6Rb+ uptake in rBSCl cRNA-injected oocytes was not inhibited by 100 PM concentrations of either metolazone or DIDS (see Fig. 3C). Finally, the enhanced *'Rb+ uptake induced in oocytes by rBSCl cRNA could not be due to either activation or expression of potassium channels or Na+-K+-ATPase, since 10 mM barium and 1 mM ouabain were present in the uptake medium. Thus, the physiological and pharmacological proper- ties of 86Rb+ uptake in rBSCl cRNA-injected oocytes show that rBSCl encodes for a sodium-potassium-chloride cotransporter that is sensitive to bumetanide but unaffected by thiazide type diuretics, i.e. rBSCl encodes a bumetanide-sensitive Na+-K+- C1- cotransporter.
Deduced Primary Structure of the Electroneutral Cotransporters
The Thiazide-sensitive Na+-C1- Cotransporter Protein-The 3,006-bp coding segment of the rTSCla cDNA predicts a pro- tein of 1002 amino acid residues (Fig. 4) with a calculated molecular mass of 110 kDa. Hydrophobicity analysis of the rTSC protein predicts three major domains (Fig. 5 A ) : a hydro- philic NH,-terminal region of 133 amino acids, which lacks a characteristic eukaryotic signal segment suggesting an intra- cellular location; a predominantly hydrophobic region contain- ing 12 putative membrane-spanning helices (Ml-Ml2; Figs. 4 and 5); and a long COOH-terminal hydrophilic region of 304 amino acids. Two of the three potential N-linked glycosylation sites (Asn-404, Asn-356, and Asn-543) are located within the putative membrane-spanning segments S6 and S10, respec- tively. The other N-linked glycosylation site (Asn-404) is lo- cated on the putative extracellular loop between potential membrane-spanning segments M7 and M8 and is conserved in flTSC (Gamba et al., 199313). The glycosylated -115-kDa pro- tein product, obtained by in vitro translation of rTSCla cRNA
A H*O rTSC cRNA n t I
Membranes - - - + + Endo H - - + - +
1 2 3 4 5 kDa
110 % --n- 80 %
B H2O rBSC cRNA - 1 1
Membranes - + - - + + Endo H - - - + - +
1 2 3 4 5 6 kDa
110,
80 %
FIG. 6. In vitro translation products of the electroneutral Na+- (K+)-CI- cotransporters. A, autoradiography of a 6% SDS-PAGE showing the in vitro translation products of rTSCla: lune 1, water control (no RNA added); lunes 2-5, with rTSCla cRNA. B , autoradiog- raphy of a 6% SDS-PAGE showing the in oitro translation products of rBSC1: lunes I and 2, water control (no RNA added) with or without membranes; lunes 34, with rBSCl cRNA. Memb, microsomal mem- branes; endo, endoglycosidase H.
in the presence of canine pancreatic microsomes, is in agree- ment with the possible single glycosylation site (Fig. 6A). The rTSCla cotransporter protein contains five potential protein kinase C phosphorylation sites (Kemp and Pearson, 1990) in predicted cytosolic regions: Ser-16, Ser-124, Ser-559, Thr-889, and Ser-934 (Figs. 4 and 5 0 ) . No potential CAMP-dependent protein kinase phosphorylation sites (Kemp and Pearson, 1990) were found in the rTSCla protein sequence.
The Bumetanide-sensitive Na+-K+-Cl- Cotransporter Pro- tein-The 3,285-bp coding segment of the rBSCl cDNApredicts a cotransporter protein of 1095 amino acid residues (M, = 120) with general structural features similar to those of rTSC: an NH,-terminal hydrophilic region of 174 amino acids, followed by a hydrophobic domain with 12 putative membrane-spanning helices, and a long hydrophilic COOH terminus of 454 amino acids. The rBSCl protein sequence contains six potential N- linked glycosylation sites; two are located in the hydrophilic loop between membrane-spanning segments M7 and M8 (Fig. 5C) with the Asn-442 site being conserved in rTSC and flTSC (Figs. 4 and 5B) (Gamba et al., 1993b); of the four other poten- tial N-linked glycosylation sites, two are positioned within po- tential membrane-spanning segments (Asn-396 and Asn-79) and two (Asn-864 and Asn-875) are located in the putative cytosolic COOH terminus. A -120-kDa product was obtained by in vitro translation of rBSCl cRNA, however, a significant increase in protein size could not be clearly resolved in the presence of canine pancreatic microsomes, possibly owing to the rather broad and indistinct bands seen on SDS-PAGE (Fig. 6B). Thus, we were not able to confirm that rBSCl can be glycosylated in vitro. rBSCl contains seven potential protein kinase C phosphorylation sites (Kemp and Pearson, 1990) (Figs. 4 and 5E): two in the NH, terminus (Ser-57 and Thr-75) and five in the COOH terminus (Thr-639, Thr-927, Ser-983,

'Ituo Ekctroneutral Na+-(K+)-Cl- Cotransporters in Kidney 17719
FIG. 7. Tissue distribution of elec- troneutral Na+-(K+)-Cl- cotransport- ers. Northern blot hybridization of ribo- probes from rTSCla cDNA (A) or from rBSCl ( B ) to total RNA (20 pgflane) from the indicated tissues at high stringency. Autoradiograms were exposed for 18 h. The spot in lane Testis of B was not repro- ducible.
A
kb 7.5 > 4.4>
1.4)
kb 7.5 > 4.4 >
1.4 >
Ser-999, and Ser-1029). In addition, three potential CAMP-de- pendent protein kinase phosphorylation sites are present in the putative cytosolic COOH terminus of rBSC1: Ser-874, Ser-1013, and Ser-1060. Interestingly, a 10-amino acid segment recently identified as containing a CAMP-dependent protein kinase phosphorylation site in the shark rectal gland Na-K-Cl cotransporter, Thr-189 in the peptide 'e5FGHNT'AgIDAVP'94 (Lytle and Forbush, 1992a), is strikingly conserved in the NH,- terminal domain of rBSCl(90% identity; g7FGHNTMDAVP'06). These potential phosphorylation sites in rBSCl are of consid- erable interest since Na+-K+-Cl- cotransporter activity can be regulated by agents that modulate CAMP-dependent protein kinase and protein kinase C in kidney (Di Stefan0 et al., 1989, 1990; Elalouf et al., 1984; Hebert et al., 1981; Homma et al., 1990; Molony et al., 1987) and other non-renal cells (Palfrey and O'Donnell, 1992; Pewitt et al., 1990).
Tissue-specific Expression Non-renal Tissue Distribution-The tissue distributions of
rBSCl and rTSCl mRNA transcripts were investigated by Northern blot analyses (Fig. 7) of total RNA extracted from each of several rat tissues, using full-length digoxigenin-UTP riboprobes constructed from rBSCl or rTSCla cDNA. Although rTSCl and rBSCl transcripts were abundant in kidney, all non-renal tissues tested were negative at high stringency.
Zntrarenal Distribution-The expression of rTSCl and rBSCl was assessed in poly(A)' RNA isolated from selected regions of the rat kidney, including superficial cortex, cortex, inner stripe of outer medulla, inner medulla, and papilla. Three transcripts were detected in superficial cortex and whole cortex mRNA, using a riboprobe constructed from rTSCla (Fig. 8B): major transcripts of -4.4 and -3.3 kb, corresponding to the
sizes of rTSCla and rTSClb cDNAs, and a minor band a t -10 kb. In situ hybridization of rat kidney sections (Fig. 9, A and B ) , using a 35S-UTP labeled probe constructed from rTSCla, re- vealed that TSC mRNA is expressed predominantly in the outer cortex in short tubule segments. This distribution is vir- tually identical to that observed for high aftinity [3H]metola- zone binding to rat kidney sections (Beaumont et al., 1989) and is consistent with the major expression of rTSC mRNA in the distal convoluted tubule, the nephron segment known to ex- press native thiazide-sensitive Na+-Cl- cotransporter activity (Beaumont et al., 1989; Costanzo, 1985; Ellison et al., 1987; Stokes, 1989; Velazquez and Wright, 1986).
Northern blot of mRNA isolated from rat kidney using a riboprobe constructed from rBSCl (Fig. 8A ), revealed a major transcript of -4.6 kb in superficial cortex, cortex, outer me- dulla, and inner medulla. Although not shown in Fig. 8, a -4.6-kb transcript was also present in papillary mRNA on films exposed for several days. In addition, two minor tran- scripts of -8 and -10 kb were observed in outer medulla. In situ hybridization of rat kidney slices using 35S-UTP labeled probes constructed from rBSCl showed a similar overall pat- tern of distribution except for the lack of rBSCl hybridization in inner medulla or papillary tip (Fig. 9, C and D). The reason for this absence of rBSCl hybridization in the inner medulla/ papillary region was not determined, but may relate to mRNA abundance or to differences in sensitivities of the assays or to the presence of an homologous, but not identical, transcript in the inner medullary region. In this regard, other investigators have clearly demonstrated bumetanide-sensitive Na+-K+-Cl- cotransporter activity in the rat inner medullary collecting duct (Grupp et al., 1989) and rabbit papillary surface epithelium (Sands et al., 1986). Nevertheless, the predominant localization

17720
A
kb 9.5 > 7.5 * 4.4 >
2.4 %
kb
9.5 > 7.5 * 4.4 >
%o Electroneutral Na+-(K+)-Cl- Cotransporters in Kidney
sc c
rrrd
IM P
sc c OM IM P
2.4 * FIG. 8. Expression of electroneutral Na+-(K+)-Cl- cotransport-
ers in kidney regions. Northern blot hybridization of a riboprobe rBSCl cDNA (A) or rTSCla cDNA(B) to poly(A)+ RNA(2.5 pg from each region) isolated from rat renal superficial cortex (SC), cortex (C), inner stripe of outer medulla (OM), inner medulla (ZM), and papilla (PT). Autoradiographs were exposed for 1 h. RNA molecular weight markers (Life Technologies, Inc.) are indicated a t left.
of rBSCl hybridization to the outer medulla and cortical med- ullary rays, is consistent with the major expression of rBSCl mRNA in the cortical and medullary thick ascending limbs of Henle.
DISCUSSION We report the cloning and deduced primary structure of the
two major mammalian renal electroneutral Na+-(K+)-Cl- co- transporters. Functional expression inX. laevis oocytes (Figs. 2 and 31, coupled with the different intrarenal distributions of specific cotransporter mRNA (Figs. 8 and 91, indicate that rBSCl cDNA encodes the bumetanide-sensitive Na+-K+-Cl- co- transporter expressed on apical membranes of the thick ascending limb of Henle (TAL) (Greger, 1981; Greger and Schlatter, 1983; Hebert and Andreoli, 1984) and rTSCla (and rTSClb) cDNA encodes the thiazide-sensitive Na+-Cl- cotrans- porter, which is functionally expressed on apical membranes of the renal distal convoluted tubule (Costanzo, 1985; Ellison et al., 1987, 1993; Stokes, 1989).
We have previously shown that vasopressin action (via cyclic AMP) is required for expression of the K+-dependence of the apical Na+-(K+)-CI- cotransporter in the mouse medullary TAL (Sun et al., 1991). However, we observed functional expression of K+-dependent Na+-Cl- cotransport in oocytes injected with cRNA transcribed from the rat kidney rBSCl clone without stimulating oocyte cyclic AMP production (e.g. with forskolin). Although oocytes have a measurable level of cyclic AMP even in the absence of forskolin stimulation (data not shown) and this may have been sufficient to account for expression of K+-de- pendent, bumetanide-sensitive Na+-Cl- cotransport in rBSC1- cRNA-injected oocytes, an alternative possibility is species dif-
ferences in the cotransporter. Good (1988) has shown that rat medullary TAL segments express furosemide-sensitive NH; transport in the absence of vasopressin stimulation. Since NH; and K+ appear to compete for a single site on the cotrans- porter (Good, 1988; Kinne et al., 1986), the results from Good’s study argue rather strongly that the cotransporter in the rat TAL functions in the Na+-K+-Cl- cotransport mode, even in the absence of vasopressin. Whether the K+-independent, bumet- anide-sensitive Na+-Cl- cotransport activity previously ob- served in mouse medullary TAL segments (Sun et al., 1991) represents a different functional state of the rBSCl Na+-K+-Cl- cotransporter or is mediated by a different, but related, protein is currently under investigation. I t seems clear, however, that rTSCl does not encode for this activity, given the sensitivity of rTSCl to thiazides, but not bumetanide, and the superficial cortical distribution of rTSCl mRNA.
rBSC1, rTSC, and flTSC Belong to the Same Family of Proteins-While the bumetanide-sensitive Na+-K+-Cl- and thia- zide-sensitive Na+-Cl- cotransporters share the property of transporting sodium and chloride in an electroneutral fashion, they differ in Na+:Cl- stoichiometry (1Na’:lCI- for TSCs (Gamba et al., 1993b; Velazquez et al., 1984) and lNa+:lK+:2Cl- for BSCs (Brown and Murer, 1985; Geck et al., 1980; Greger, 1981; Greger and Schlatter, 1983)), the ability to transport K+(Rb+), and the sensitivity to inhibitors (diuretic agents and stilbene disulfonates). However, the two mammalian renal NaCl cotransporters reported here and the flounder urinary bladder thiazide-sensitive Na+-Cl- cotransporter (Gamba et al., 1993b) cloned previously exhibit remarkably similar hydropho- bicity profiles (Fig. 5, A-C) and proposed topologies (Fig. 5, D and E ) , and they share significant overall amino acid sequence homology (40% identity, 48% similarity; Fig. 4). Among the three cotransporters, the highest degree of similarity is be- tween rTSCla and flTSC (62% identity, 72.6% similarity); how- ever, even between rBSCl and the TSCs, the similarity is -60% (for rBSC1-rTSCla, 48% identity and 61.7% similarity; for rBSC1-flTSC, 46% identity and 59.7% similarity). The re- gions corresponding to the membrane-spanning segments are more conserved, whereas the putative extracellular loops, the NH,- and COOH-terminal loops, are less well conserved. The highest degree of similarity is in the membrane-spanning seg- ments S1, S3, S6, S8, and S10 (-85% identity). The lowest degree of similarity is located in the NH,-terminal loop (-5% identity).
A search of protein sequence data bases (GenBank, EMBL, and SWISS-PROT) revealed no significant overall sequence similarities between rTSCla (rTSClb) or rBSCl and other re- ported proteins. Specifically, there was no significant homology with Na+-coupled cotransporters of glucose (Hediger et al., 1987), y-aminobutyric acid (Amara and Kuhar, 1993), or gluta- mate (Kanai et al., 1993) or with NaPO, cotransporters (Werner et al., 1991), Na+-H+ exchangers (Collins et al., 1992; Orlowski et al., 1992; Sardet et al., 1989), or Cl-:HCO, exchangers (Alper et al., 1988; Kopito and Lodish, 1985). Thus rBSC1, rTSCla (and rTSClb), and flTSC define a new family of membrane transporters, the electroneutral Na+-(K+)-Cl- cotransporter family. Although the full amino acid sequence of the basolateral Na+-K+-Cl- cotransporter from shark rectal gland is not cur- rently published, the conservation of the NH,-terminal phos- phopeptide (Lytle and Forbush, 1992a) in rBSCl (see “Results”) suggests that the rectal gland cotransporter belongs to this family.
While determination of the molecular size of the native co- transporter protein(.$ will require production of specific anti- BSC antibodies, the -120-kDa rBSCl core protein (Fig. 6B) is similar in size to the largest of the [3H]bumetanide-binding or
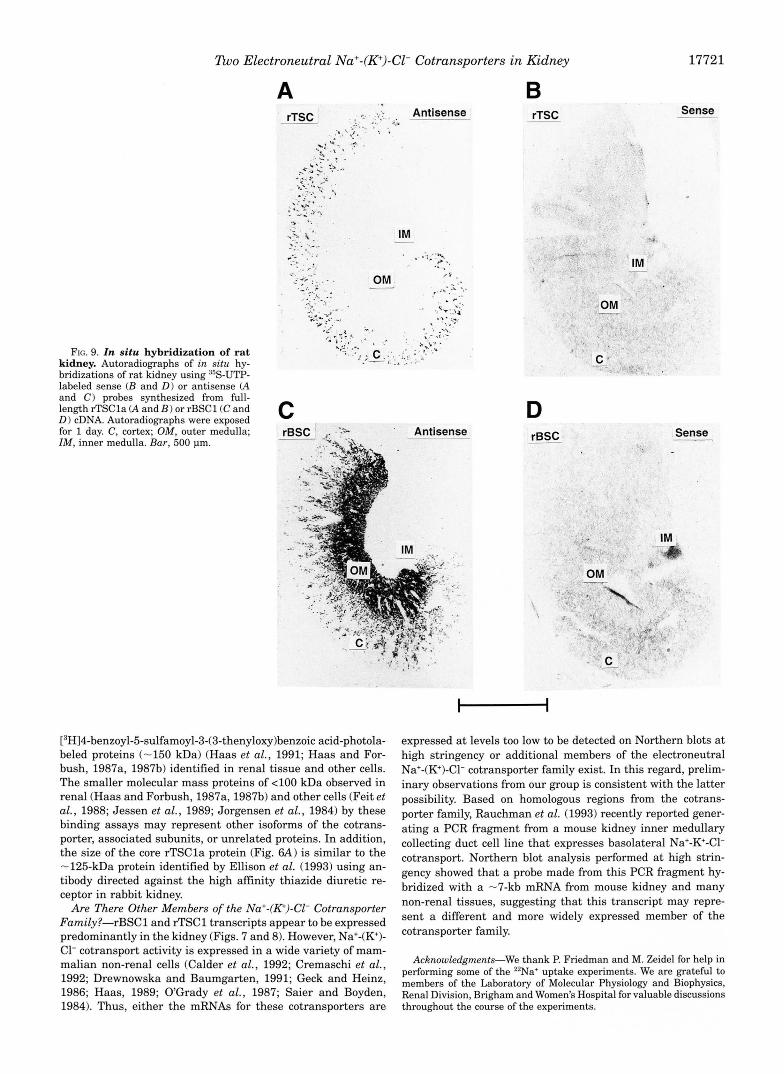
l tvo Electroneutral Na+-(K+)-Cl- Cotransporters in Kidney 17721
FIG. 9. In situ hybridization of rat kidney. Autoradiographs of in situ hy- bridizations of rat kidney using 35S-UTP- labeled sense (B and D) or antisense (A and C ) probes synthesized from full- length rTSCla (A and B ) or rBSCl (C and D ) cDNA. Autoradiographs were exposed for 1 day. C , cortex; OM, outer medulla; IM, inner medulla. Bur, 500 pm.
A B
C rBSC _" -5 ',. ' Antisense ~-
[3H14-benzoyl-5-sulfamoyl-3-(3-thenyloxy)benzoic acid-photola- beled proteins (-150 kDa) (Haas et al., 1991; Haas and For- bush, 1987a, 1987b) identified in renal tissue and other cells. The smaller molecular mass proteins of d o 0 kDa observed in renal (Haas and Forbush, 1987a, 1987b) and other cells (Feit et al., 1988; Jessen et al., 1989; Jorgensen et al., 1984) by these binding assays may represent other isoforms of the cotrans- porter, associated subunits, or unrelated proteins. In addition, the size of the core rTSCla protein (Fig. 6 A ) is similar to the -125-kDa protein identified by Ellison et al. (1993) using an- tibody directed against the high affinity thiazide diuretic re- ceptor in rabbit kidney.
Are There Other Members of the Na+-(K+)-Cl- Cotransporter Family?-rBSC1 and rTSCl transcripts appear to be expressed predominantly in the kidney (Figs. 7 and 8). However, Na+-(K+)- C1- cotransport activity is expressed in a wide variety of mam- malian non-renal cells (Calder et al., 1992; Cremaschi et al., 1992; Drewnowska and Baumgarten, 1991; Geck and Heinz, 1986; Haas, 1989; O'Grady et al., 1987; Saier and Boyden, 1984). Thus, either the mRNAs for these cotransporters are
. - r .
D rBSC- - Sense ""
H
expressed at levels too low to be detected on Northern blots at high stringency or additional members of the electroneutral Na+-(K+)-Cl- cotransporter family exist. In this regard, prelim- inary observations from our group is consistent with the latter possibility. Based on homologous regions from the cotrans- porter family, Rauchman et al. (1993) recently reported gener- ating a PCR fragment from a mouse kidney inner medullary collecting duct cell line that expresses basolateral Na+-K+-Cl- cotransport. Northern blot analysis performed at high strin- gency showed that a probe made from this PCR fragment hy- bridized with a -7-kb mRNA from mouse kidney and many non-renal tissues, suggesting that this transcript may repre- sent a different and more widely expressed member of the cotransporter family.
Acknowledgments-We thank P. Friedman and M. Zeidel for help in performing some of the 22Na+ uptake experiments. We are grateful to members of the Laboratory of Molecular Physiology and Biophysics, Renal Division, Brigham and Women's Hospital for valuable discussions throughout the course of the experiments.

17722 lluo Electroneutral Na+-(K+)-Cl- Cotransporters in Kidney Note Added in Proof-The sequence of the shark rectal gland Na-
K-CI cotransporter has recently been reported (Xu et al., 1994) and clearly belongs to this cotransporter family.
REFERENCES
Alper, S. L., Kopito, R. R., Libresco, S. M., and Lodish, H. F. (1988) J. Biol. Chem.
Alvo, M., Calamia, J., and Eveloff, J . L. (1985) Am. J. Physiol. 249, F36F39 Amara, S. G., and Kuhar, M. J. (1993) Annu. Reu. Neurosci. 16,73-93 Beaumont, IC, Vaughn, D. A., and Fanestil, D. D. (1988) Proc. Natl. Acad. Sci.
Beaumont, K., Vaughn, D. A,, and Healy, D. P. (1989) J. Pharmacol. Exp. Ther. 250,
Brown, C. D. A,, and Murer, H. (1985) J. Membr. Biol. 87, 131-139 Burg, M. B., Stoner, L. C., Cardinal, J., and Green, N. (1973)Am. J. Physiol. 225,
Bumham, C. E., Kidd, J., and Palfrey, H. C. (1990)Am. J. Physiol. 259, F383-F388 Cabantchik, 2. I., and Rothstein, A. (1972) J. Membr. Biol. 10, 311-330 Calder, J . A., Schachter, M., and Sever, P. S. (1992) Eur. J. Pharmacol. 220,19-26 Collins, J. F., Honda, T., Knobel, S., Bulus, N. M., Conary, J., DuBois, R., and
Costanzo, L. S. (1985) Am. J. Physiol. 248, F527-F535 Cremaschi, D., Porta, C., Botta, G., and Meyer, G. (1992) J. Membr. Biol. 129,
Di Stefano, A,, Wittner, M., Nitschke, R., Braitsch, R., Greger, R., Bailly, C., Amiel, 221-235
640446 C., Elalouf, J. M., Roinel, N., and De Rouffignac, C. (1989) Pfliigers Arch. 414,
Di Stefano, A,, Wittner, M., Nitschke, R., Braitsch, R., Greger, R., Bailly, C., Amiel, C., Roinel, N., and De Roufflgnac, C. (1990) Pfliigers Arch. 417, 161-167
Drewnowska, K., and Baumgarten, C. M. (1991)Am. J. Physiol. 260, C122-C131 Elalouf, J. M., Roinel, N., and De Roufignac, C. (1984) Am. J. Physiol. 246,
Ellison, D. H., Velazquez, H., and Wright, F. S. (1987)Am. J. Physiol. 253, F546- F213-F220
Ellison, D. H., Biemesderder, D., Momsey, J., Lauring, J., and Desir, G. V. (1993) F554
Feit, P. W., Hoffmann, E. K., Schiodt, M., Kristensen, P., Jessen, F., and Dunham, Am. J. Physiol. 264, F141-F148
Gamba, G., Miyanoshita, A,, Lombardi, M., Lytton, J., Lee, W.-S., Hediger, M. A,, P. B. (1988) J. Membr. Biol. 103, 135-147
Gamba, G., Saltzberg, S. N., Lombardi, M., Miyanoshita,A., Lytton, J., Hediger, M. and Hebert, S. C. (1993a) J. Am. SOC. Nephrol. 4, 866 (abstr.)
A,. Brenner. B. M.. and Hebert, S. C. (1993b) Proc. Natl. Acad. Sci. U. S. A. 90,
263, 17092-17099
U. S. A. 85, 2311-2314
414-419
119-124
Ghishan, F. K. (1993) Proc. Natl. Acad. Sci. U. S. A. 90,39383942
Geck, P., and Heinz, E. (1986) J. Membr. Biol. 91, 97-105 Geck, P., Pietrzyk, C., Burckhardt, B.-C., Pfeiffer, B., andHeinz, E. (198O)Biochim.
2749-2753
Good, D. W. (1988) Am. J. Physiol. 255, F78-F87 Greger, R. (1981) Scand. J. Audiol. Suppl. 14, 1-15 Greger, R. (1985) Physiol. Reu. 65, 760-797 Greger, R., and Schlatter, E. (1983) KZin. Wochenschr. 61, 1019-1027 Grupp, C., Pavenstadt-Grupp, I., Grunewald, R. W., Bevan, C., Stokes, J. B., 111,
Haas, M. (1989) Annu. Reu. Physiol. 51 ,443457 Haas, M., and Forbush, B., I11 (1987a) Am. J. Physiol. 253, C243-C250
Haas, M., Dunham, P. B., and Forbush, B., 111 (1991) Am. J. Physiol. 260, C791- Haas, M., and Forbush, B., 111 (1987b) Kidney Znt. 32, S134-S140
Hebert, S. C., and Andreoli, T. E. (1984)Am. J. Physiol. 246, F745-F756 Hebert, S. C., and Sun, A. (1988) Am. J. Physiol. 255, F962-F969 Hebert, S. C., Culpepper, R. M., and Andreoli, T. E. (1981) Am. J. Physiol. 241,
Biophys. Acta 600,432447
and Kinne, R. K. H. (1989) Kidney Int. 36, 201-209
C804
F412-F431
Hediger, M. A., Coady, M. J., Ikeda, T. S., and Wright, E. M. (1987) Nature 330,
Hoffmann, E. K., and Simonsen, L. 0. (1989) Physiol. Reu. 69,315-382 Homma, T., Bums, K. D., and Hams, R. C. (199O)J. Biol. Chem. 265,17613-17620 Jessen, F., Cherksey, B. D., Zeuthen, T., and Hoffmann, E. K. (1989) J. Membr. Biol.
Jorgensen, P. L., Petersen, J., and Rees, W. D. (1984) Biochim. Biophys. Acta 775,
Kanai, Y., Stelzner, M. G., Lee, W.-S., Wells, R. G., Brown, D., and Hediger, M. A.
Kanai, Y., Smith, C. P., and Hediger, M. A. (1993) Dends Neurosci. 16, 365-370
Kikeri, D., Azar, S., Sun, A,, Zeidel, M. L., and Hebert, S. C. (1990)Am. J. Physiol. Kemp, B. E., and Pearson, R. B. (1990) Dends Biochem. Sci 15,342-346
Kinne, R., Kinne-Saffran, E., Schutz, H., and Scholermann, B. (1986) J. Membr.
Kopito, R. R., and Lodish, H. F. (1985) Nature 316, 234-238 Kozak, M. (1991) J. Biol. Chem. 266, 19867-19870 Kyte, J., and Doolittle, R. F. (1982) J. Mol. Biol. 157, 105-132 Li, J. H., Zuzack, J. S., and Kau, S. T. (1990) in Diuretics III; Chemistry, Pharma-
cology, and Clinical Applications (Puschett, J. B., and Greenberg, A,, eds) pp. 107-110, Elsevier Science Publishing Co., New York
379381
108, 139-151
105-110
(1992) Am. J. Physiol. 263, F1087-F1093
258, F445-F456
Biol. 94,279-284
Luo, H., and Fanestil, D. D. (1991) Biochem. Pharmacol. 41, 1745-1750
Lytle, C.,Xu, J.-C., Biemesderfer, D., Haas, M., andForbush, B., I11 (1992b)J. Biol. Lytle, C., and Forbush, B., I11 (1992a) J. Biol. Chem. 267,25438-25443
Manley, J. L. (1988) Biochim. Biophys. Acta 950, 1-12 Molony, D. A,, Reeves, W. B., Hebert, S. C., and Andreoli, T. E. (1987) Am. J.
O'Grady, S. M., Palfrey, H. C., and Field, M. (1987)Am. J. Physiol. 253, C177-C192 Orlowski, J., Kandasamy, R. A., and Shull, G. E. (1992) J. Biol. Chem. 267,9331-
Palfrey, H. C., and O'Donnell, M. E. (1992) Cell Physiol. Biochem. 2, 293307 Pewitt, E. B., Hegde, R. S., Haas, M., and Palfrey, H. C. (1990) J. Biol. Chem. 265,
Rauchman, M., Delpire, E., Hebert, S. C., and Gullans, S. R. (1993) J. Am. SOC.
Saier, M. H., and Boyden, D. A. (1984) Mol. Cell. Biochem. 59, 1 1 3 2 Sands, J. M., Knepper, M. A., and Spring, K. R. (1986) Am. J. Physiol. 251,
Sardet, C., Franchi, A,, and Pouyssegur, J. (1989) Cell 66,271-280 Shetlar, R. E., and Kinne, R. K. H. (1990) F U E B J . 4, A563 (abstr.) Shetlar, R. E., Scholermann, B., Morrison, A. I., and Kinne, R. K. H. (1990) Bio-
Stokes, J. B. (1984) J. Clin. Invest. 74, 7-16
Sun, A,, Grossman, E. B., Lombardi, M., and Hebert, S. C. (1991) J. Membr. Biol. Stokes, J. B. (1989) Kidney Int. 36, 427433
Suvitayavat, W., Palfrey, H. C., Haas, M., Dunham, P. B., Kalmar, E, and Rao, M.
Tran, J. M., Farrell, M. A., and Fanestil, D. D. (1990)Am. J. Physiol. 258, F908-
Velazquez, H., and Wright, F. S. (1986) Am. J. Physiol. 250, F1013-F1023 Velazquez, H., Good, D. W., and Wright, F. S. (1984)Am. J. Physiol. 247, F904-
Werner, A., Moore, M. L., Mantei, N., Biber, J., Semenza, G., and Murer, H. (1991)
Xu, J.-C., Lytle, C., Zhu, T. T., Payne, J. A,, Benz, E., Jr., and Forbush, B., 111 (1994)
Chem. 267,25428-25437
Physiol. 252, F177-F187
9339
20747-20756
Nephrol. 4, 877
F475-F484
chim. Biophys. Acta 1023, 184-190
120,83-94
C. (1994)Am. J. Physiol. 266, C28PC292
F915
F911
Proc. Natl. Acad. Sci. U. S. A. 88, 9608-9612
Proc. Natl. Acad. Sei. U. S. A. 91, 2201-2205