the of biochemistry and university seattle, · erythrocyte metabolism ... via the...
TRANSCRIPT

ERYTHROCYTE METABOLISM
IV. ISOLATION AND PROPERTIES OF METHEMOGLOBIN REDUCTASE*
BY F. M. HUENNEKENS, RUTH WADE CAFFREY, R. E. BASFORD,t AND BEVERLY W. GABRIO
WITH THE TECHNICAL ASSISTANCE OF L. LIU
(From the Departments of Biochemistry and Medicine, University of Washington, Seattle, Washington)
(Received for publication, November 26, 1956)
In mature, non-nucleated erythrocytes the breakdown of glucose to pen- tose, via the “phosphogluconate oxidation” pathway, includes an oxida- tive step, whereby glucose 6-phosphate is converted to 6-phosphogluconate in the presence of TPN’ and the appropriate dehydrogenase (2). It has been shown previously by other investigators (2-4), and amplified in Paper III of this series (5), that TPNH, formed in the primary dehydrogenation, is linked to oxygen through a TPNH oxidase. The isolated enzyme re- quires an added electron carrier such as methylene blue for its activity.
A similar situation obtains in the reduction of methemoglobin by the erythrocyte. The low (about 1 per cent (6)) steady state level of methe- moglobin in the cell indicates that the slow, continuous chemical oxidation of hemoglobin by molecular oxygen is nearly compensated for by the en- zymatic reduction of methemoglobin. Kiese (7) has partially purified the enzyme, methemoglobin reduct.ase, and has shown that TPNH is the sub- strate for the enzyme and that methylene blue serves as a cofactor. Evi- dence has been presented also for the existence of a similar, but separate, system requiring DPNH. In accordance with varying sources of evidence Kiese has suggested that the TPNH oxidase and the TPNH methemoglo- bin reductase are the same enzyme.
The present communication describes the isolation, in highly purified form, and the properties of the methemoglobin reductase from human erythrocytes. Observations are presented concerning TPNH and DPNH
* This investigation was supported by grants from the National Institutes of Health, United States Public Health Service, Department of the Army, Office of the Surgeon General, Eli Lilly and Company, and Initiative 171, State of Washington. A preliminary account of this work was presented at the Forty-fifth meeting of the American Society of Biological Chemists at Atlantic City, April, 1954 (1).
t Present address, Institute for Enzyme Research, University of Wisconsin. * The following abbreviations are used: DPN and TPN, di- and triphosphopyridine
nucleotides; DPNH and TPNH, reduced DPN and TPN; Hb, reduced hemoglobin; HbOz, oxyhemoglobin; MHb, methemoglobin; MeB, methylene blue.
261
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from

262 METHEMOGLOBIN REDUCTASE
as substrates, and oxygen, methemoglobin, and cytochrome c as terminal electron acceptors.
EXPERIMENTAL
Materials-Crystalline hemoglobin was prepared from human blood2 by the method of Drabkin (8), and methemoglobin by the oxidation of hemo- globin with NaNOz or Hz02 (9). Isocitric dehydrogenase was isolated from pig heart (10). dl-Isocitric acid la&one was obtained from the Cali- fornia Foundation for Biochemical Research, crystalline alcohol dehydro- genase was a product of the Mann Research Laboratories, Inc., TPNH, DPNH, and cytochrome c were obtained from the Sigma Chemical Com- pany, 1, lo-o-phenanthroline from the G. Fredrick Smith Chemical Com- pany, and iron-specific Versene from the Bersworth Chemical Company. Other materials were obtained as described in Paper III (5).
Methods-Protein concentrations were determined by the biuret method (11) with bovine serum albumin as the standard. Pyridine hemochromo- gen determinations were carried out as described by Basford et al. (12) with a molar extinction coefficient, E, equal to 34.7 X lo6 sq. cm. per mole (13) at 557 rnp for the hemochromogen of iron protoporphyrin. Total iron was estimated by the method of Sandell (14).
Spectrophotometric measurements were made in 1 cm. cuvettes in the Beckman spectrophotometer, model DU. Absorption spectra were ob- tained with a Cary recording spectrophotometer, model 11M.
Assay of the enzyme as a TPNH oxidase (i.e. with oxygen as the ter- minal electron acceptor) was carried out as follows: The experimental cuvette contained 0.15 pmole of TPNH, 100 pmoles of phosphate buffer,) pH 7.5, 0.2 ml. of enzyme, and water to make 3.0 ml. The blank cuvette was identical except for the omission of TPNH. After a stable, initial log lo/l value at 340 ml had been obtained, 0.01 ml. of 0.1 per cent methylene blue (0.027 pmole) was added to each cuvette at zero time and the reac- tion was followed by the decrease in light absorption over a 10 minute pe- riod. The reaction velocity was corrected for the slow, chemical oxidation of TPNH by the dye (Alo=0.022) in a blank cuvette from which the en- zyme was omitted.
The reaction velocity is linear with time and can be expressed as a Alo value, i.e. the change in log I,/1 at 340 mp over the 10 minute period, un- der conditions of enzyme concentration whereby Alo is less than 0.060. 1
2 We are indebted to Dr. Richard Czajkowski and Mr. Soren Jule of the King County Blood Bank, Seattle, for collecting and making available the blood for this study.
3 At the optimal pH for the enzyme (pH 7.5), there was no diminution of activity when tris(hydroxymethyl)aminomethane or Verona1 replaced phosphate as the buffer.
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from

HUENNEKENS, CAFFREY, BASFORD, AND GABRIO 263
unit of enzyme is defined as that amount which causes A,, = 0.010 under these conditions. Specific activity is defined as units of enzyme per mg. of protein. DPNH oxidase activity is det.ermined by the above spectro- photometric system with an equivalent amount of DPNH replacing the TPNH.
In order to study the enzyme as a methemoglobin reductase or cyto- chrome c reductase, the spectrophotometric assay system was modified to include 0.12 pmole of MHb or 0.10 to 0.20 pmole of cytochrome c. The methemoglobin reductase reaction was followed by the disappearance of the MHb band at 630 rnp, or the appearance of the HbOz band at 575 mp. The reaction with cytochrome c was followed by the appearance of the re- duced cytochrome c band at 550 rnp.
RESULTS AND DISCUSSION
Purijication of Enzyme
Step 1. Removal of Hemoglobin from Hemolysates-Fresh, human eryth- rocytes (125 ml. of packed cells) were washed three times by suspension and centrifugationh in cold, 0.9 per cent NaCl solution,6 diluted to the orig- inal volume (about 300 ml.) with saline, and hemolyzed by repeated freez- ing and thawing. As described in Paper III (5), the hemolysate was treated at low temperature with ethanol and chloroform to remove hemo- globin and was dialyzed.
Step 2. Fractionation at pH 5.4-The dialyzed enzyme from Step 1 (865 ml.) was lyophilized to dryness in order to remove traces of organic sol- vents and redissolved in 80 ml. of water. The dilute solution was allowed to stand overnight at 5”, and any precipitate which formed was removed by centrifugation. The solution was then adjusted to pH 5.4 with 10 per cent acetic acid, the precipitate removed by centrifugation, and the super- natant solution readjusted to pH 7.0 with 1 N sodium hydroxide. The amount of inert protein removed by the aging step varied somewhat with different preparations; however, the data given in Table I are representa- tive of a number of individual preparations.
Step 3. Ammonium Sulfate Fractionation-To the solution from Step 2 (80 ml.), immersed in an ice bath, were added slowly with stirring 80 ml. of saturated ammonium sulfate solution neutralized to pH 7.0 with concen- trated ammonium hydroxide. The precipitate (0 to 50 per cent fraction)
4 All centrifugations were carried out at 4’ for 15 minutes at 2300 X g in the Inter- national refrigerated centrifuge, model PR-1.
6 It has been found that cells washed thoroughly with 0.9 per cent NaCl before hemolysis yield enzyme preparations at the end of Step 1 which have a 5- to IO-fold higher specific activity compared to cells washed with Krebs-Ringer-phosphate solution.
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from
264 METHEMOGLOBIN REDUCTASE
was removed by centrifugation and discarded. Successive addition of 40, 67, and 135 ml. of ammonium sulfate to the 0 to 50 per cent supernatant fraction yielded the 50 to 60, 60 to 70, and 70 to 80 per cent precipitabes, which were removed by centrifugation, dissolved in 40 ml. of water, and dialyzed overnight against water. After dialysis, each of the above three fractions was centrifuged to remove any precipitate and assayed for activ- ity. The highest specific activity and most of the total activity were found in the 60 to 70 per cent fraction, which was used for all the experi- ments reported in this paper. The 50 to 60 or 70 to 80 per cent fractions could be refractionated with ammonium sulfate to obtain additional quan- tities of the enzyme.
For a number of preparations the specific activity after ammonium sul- fate fractionation ranged between 68 and 72. No further purification
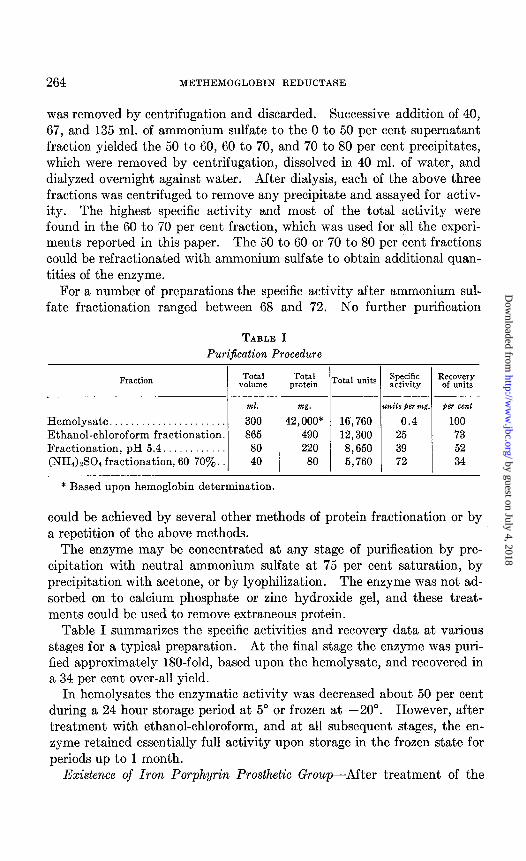
TABLE I Purification Procedure
Hemolysate...................... 300 42,000* 16,760 0.4 100 Ethanol-chloroform fractionation. 865 490 12,300 25 73 Fractionation, pH 5.4.. 80 220 8,650 39 52 (NHa) &!04 fractionation, 60-70s. 40 80 5,760 72 34
* Based upon hemoglobin determination.
could be achieved by several other methods of protein fractionation or by a repetition of the above methods.
The enzyme may be concentrated at any stage of purification by pre- cipitation with neutral ammonium sulfate at 75 per cent saturation, by precipitation with acetone, or by lyophilization. The enzyme was not ad- sorbed on to calcium phosphate or zinc hydroxide gel, and these treat- ments could be used to remove extraneous protein.
Table I summarizes the specific activities and recovery data at various stages for a typical preparation. At the final stage the enzyme was puri- fied approximately 180-fold, based upon the hemolysate, and recovered in a 34 per cent over-all yield.
In hemolysates the enzymatic activity was decreased about 50 per cent during a 24 hour storage period at 5” or frozen at -20”. However, after treatment with ethanol-chloroform, and at all subsequent st.ages, the en- zyme retained essentially full activity upon storage in the frozen state for periods up to 1 month.
Existence of Iron Porphyrin Prosthetic GroupAfter treatment of the
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from
HUENNEKENS, CAFFREY, BASFORD, AND GABRIO 265
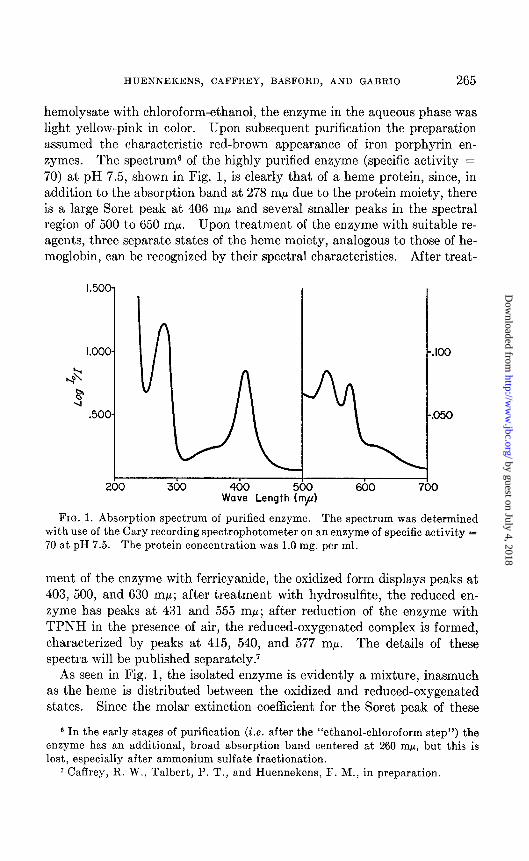
hemolysate with chloroform-ethanol, the enzyme in the aqueous phase was light yellow-pink in color. Upon subsequent purification the preparation assumed the characteristic red-brown appearance of iron porphyrin en- zymes. The spectrum6 of t,he highly purified enzyme (specific activity = 70) at pH 7.5, shown in Fig. 1, is clearly that of a heme protein, since, in addition to the absorption band at 278 rnp due to the protein moiety, there is a large Soret peak at 406 rnp and several smaller peaks in the spectral region of 500 to 650 rnp. Upon treatment of the enzyme with suitable re- agents, three separate states of the heme moiety, analogous to those of he- moglobin, can be recognized by their spectral characteristics. After treat-
FIG. 1. Absorption spectrum of purified enzyme. The spectrum was determined with use of the Cary recording spectrophotometer on an enzyme of specific activity = 70 at pH 7.5. The protein concentration was 1.0 mg. per ml.
ment of the enzyme with ferricyanide, the oxidized form displays peaks at 403, 500, and 630 rnp; after treatment with hydrosulfite, the reduced en- zyme has peaks at 431 and 555 rnp; after reduction of the enzyme with TPNH in the presence of air, the reduced-oxygenated complex is formed, characterized by peaks ab 415, 540, and 577 rnp. The details of these spectra will be published separately.7
As seen in Fig. 1, the isolated enzyme is evidently a mixture, inasmuch as the heme is distributed between the oxidized and reduced-oxygenated states. Since the molar extinction coefficient for the Soret peak of these
6 In the early stages of purification (i.e. after the “ethanol-chloroform step”) the enzyme has an additional, broad absorption band centered at 260 rnp, but this is lost, especially after ammonium sulfate fractionation.
7 Caffrey, R. W., Talbert, P. T., and Huennekens, F. M., in preparation.
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from

266 METHEMOGLOBIN REDUCTASE
two states is almost identical,8 the ratio of the protein peak at 278 rnp to the composite Soret peak, located between 405 and 408 rnp, may be used as a measure of the purification of the enzyme. The ratio is 1.45 for the pu- rified enzyme.
That the iron porphyrin is associated with the enzyme itself and is not due to traces of contaminating hemoglobin is substantiated by several ob- servations. First, the absorption maxima of the various states of the en- zyme are slightly, but significantly, different from those of hemoglobin. Second, repeated treatment of the enzyme with ethanol-chloroform under conditions identical with those of Step 1 in the purification procedure, whereby hemoglobin is denatured, does not lead to any further decrease in color or to further denaturation. Third, the ratio of enzymatic activity to the light absorption due to the porphyrin moiety is constant after the first stage during purification. Fourth, the enzyme is eluted from ion exchange columns under conditions whereby the multiple forms of hemoglobin (16) are still adsorbed.7 Fifth, crystalline hemoglobin itself displays no activ- ity as a methemoglobin reductase. Finally, under anaerobic conditions and with substrate amounts of enzyme, the iron porphyrin moiety can be reduced quantitatively by substrate amounts of reduced methylene blue or by substrate amounts of TPNH in the presence of catalytic amounts of methylene blue. These results will be reported in a subsequent communi- cation.7
It is proposed, then, that the iron porphyrin group is a functional part of the enzyme, and that it does not represent traces of contaminating he- moglobin, although the iron porphyrin moiety of the two proteins may be identical.
Molecular Weight Determination from Total Iron and Hemochromogen Analyses-Two separate aliquots of the purified enzyme were converted to the pyridine hemochromogen. The absorption maxima were at 418, 522, and 553 ml. From the optical density values at 553 rnp, the heme con- tent of the enzyme was calculated to be 5.47 and 5.51 X 10e6 mmole per mg. of protein. Assuming 1 heme per mole, this would correspond to a minimal molecular weight of 182,000. Total iron determinations, with cytochrome c as an internal control for the analytical method, gave an av- erage value of 5.31 X 1OW mmole per mg. of protein, or a molecular weight of 188,000. The actual molecular weight of the enzyme may be lowered slightly by a factor representing the absolute purity of the enzyme.
Spectrophotometric Demonstration of Activity-At all stages of purification
8 A solution of the enzyme (specific activity = 72) containing 1.63 mg. per ml. of total protein had an optical density of 1.050 at 406 mp. Assuming a molecular weight of 1.85 X 104, this would correspond to E = 119 X lo6 sq. cm. per mole for the Soret peak. The r values for the Soret peaks of HbOt and MHb are each approximately 130 X lo6 sq. cm. per mole (15).
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from
HUENNEKENS, CAFFREY, BASFORD, .4ND GABRIO 267
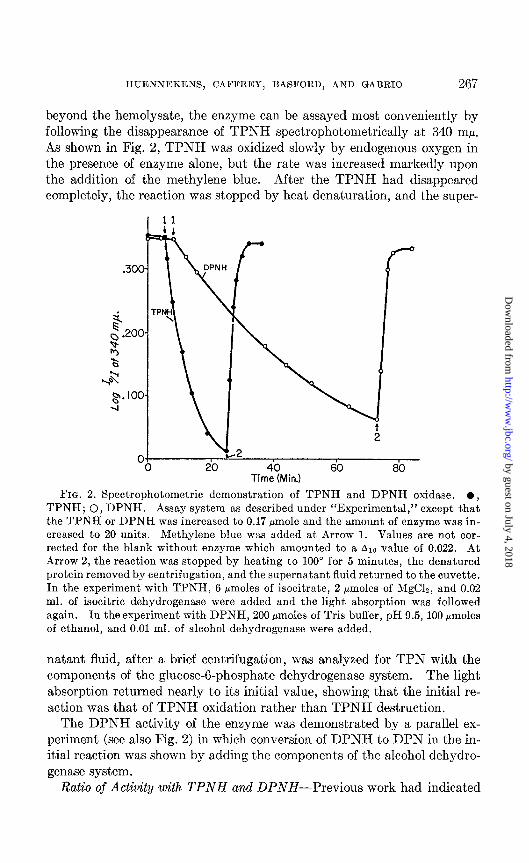
beyond the hemolysate, the enzyme can be assayed most conveniently by following the disappearance of TPNH spectrophotometrically at 340 rnp. As shown in Fig. 2, TPNH was oxidized slowly by endogenous oxygen in the presence of enzyme alone, but the rate was increased markedly upon the addition of the methylene blue. After the TPNH had disappeared completely, the reaction was stopped by heat denaturation, and the super-
2
TirrZ%4in.) 60 80
FIG. 2. Spectrophotometric demonstration of TPNH and DPNH oxidase. l , TPNH; 0, DPNH. Assay system as described under “Experimental,” except that the TPNH or DPNH was increased to 0.17 pmole and the amount of enzyme was in- creased to 20 units. Methylene blue was added at Arrow 1. Values are not cor- rected for the blank without enzyme which amounted to a Alo value of 0.022. At Arrow 2, the reaction was stopped by heating to 100” for 5 minutes, the denatured protein removed by centrifugation, and the supernatant fluid returned to the cuvette. In the experiment with TPNH, 6 pmoles of isocitrate, 2 pmoles of MgClz, and 0.02 ml. of isocitric dehydrogenase were added and the light absorption was followed again. In the experiment with DPNH, 200 Hmoles of Tris buffer, pH 9.5, 100 pmoles of ethanol, and 0.01 ml. of alcohol dehydrogenase were added.
natant fluid, after a brief centrifugation, was analyzed for TPN with the components of the glucose-g-phosphate dehydrogenase system. The light absorption returned nearly to its initial value, showing that the initial re- action was that of TPNH oxidation rather than TPNH destruction.
The DPNH activity of the enzyme was demonstrated by a parallel ex- periment (see also Fig. 2) in which conversion of DPNH to DPN in the in- itial reaction was shown by adding the components of the alcohol dehydro- genase system.
Ratio of Activity with TPNH and DPNH-Previous work ha.d indicated
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from

268 METHEMOGLOBIN REDUCTASE
t.he existence of two separate enzymes for the reduction of MHb, one spe- cific for TPNH and t,he other for DPNH. For example, Kiese (7) demon- strated in hemolysates that glucose and lactate were additive in their abil- ity to reduce MHb and that, when each of these substrates was employed, a different K, value was obtained for the cofactor, methylene blue.
With the enzyme at the highest stage of purification, the ratio of TPNH to DPNH activity was 5.4 when the assays were performed under parallel conditions. This ratio is not appreciably different from that at Step 1, where t,he value was approximately 3.5. In the latter case, the ratio is less reliable, owing to the relatively large amount of protein required for the assay. These data suggest that the present enzyme, as isolated, is spe- cific for TPNH, but that it has some activity with DPNH. There may exist, in addition, a DPN-specific reductase which is destroyed during the above isolation procedure.
Inhibitors-The enzyme is inhibited (a) approximately 25 per cent by Ca++, Ba++, Mn++, sulfate, and pyrophosphate, each at, a final concentra- tion of 10F2 M; (b) 40 per cent by Zn++ or atebrin at 10e3 M; (c) 20 per cent by p-chloromercuribenzoate at lo+ M; and (d) 50 per cent by Hg++ at 1O-5 M. The enzyme is not inhibited appreciably by riboflavin, riboflavin phos- phate, flavin adenine dinucleotide, 2,4-dinitrophenol, gramicidin, 8-hy- droxyquinoline, isonicotinic hydrazide, arsenite, arsenate, iodoacetate, io- dosobenzoate, or Versenate, each at a final concentration of 10F2 M. The pattern of inhibition for this enzyme is similar to that observed previously for the DPNH oxidase isolated from pig heart (17).
Terminal Electron Acceptors--In Paper III (5) and in the present inves- tigation it has been shown that oxygen serves as a terminal electron accep- tor in the presence of catalytic amounts of methylene blue. Under an- aerobic conditions, i.e. in evacuated cuvettes or in Thunberg tubes, methylene blue or 2,6-dichlorophenol-indophenol may be used as an acceptor.
Kiese has presented evidence that either methemoglobin or oxygen can be used as terminal electron acceptors for the enzyme and that they are competitive with each other. Using the purified enzyme, we have con- firmed his findings and have found that cytochrome cg may be used also as a terminal acceptor. A summary of comparative rates of reduction for the three acceptors, oxygen, MHb, and cytochrome c, is presented in Table II.
For either cytochrome c or MHb, the stoichiometry is as follows:
TPNH + 2 cytochrome c+++ -+ TPN+ + 2 cytochrome c++ + H+ (1)
9 There is no crossed specificity in this reaction, since methemoglobin will not replace cytochrome c in the DPNH cytochrome c reductase from pig heart (18) or the TPNH cytochrome c reductase from yeast (19). Altman (20) has found, however, that a preparation from brewers’ yeast contains MHb reductase activity.
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from

HUENNEKENS, CAFFREY, BASFORD, AND GABRIO 269
The reduction of these acceptors was followed by means of the optical den- sity changes at 550 rnp for cytochrome c and 630 rnp or 575 rnp for MHb. The initial rates, linear over a 0 to 5 minute interval, were 39.6 X lo-’ mmoles per ml. per minute for cytochrome c and 3.6 X lo-’ for MHb. Ac- cording to Equation 1, these values would correspond to rates of TPNH oxidation of 19.8 X lo-’ and 1.8 X lo-‘, respectively. Since the actual rates of TPNH oxidation, as measured by changes in optical density at
TABLE II
Terminal Electron Acceptors
Acceptor
Rate of TPNH disappearance, mmoles per ml. per min. X 10’
Observed Due to porphyrin Due to 0,
Cytochrome c.............................. 22.5 19.8 2.7 Oxygen.................................... 12.5 12.5 Methemoglobin. _..__..__..._..__........ 10.0 1.8 8.2
The standard assay system was used as described under “Experimental,” except that 0.3 pmole of TPNH was used. Successive spectra at given time intervals were run on the Cary recording spectrophotometer. For oxygen, the reaction was fol- lowed at 340 rnp, with AC of 6.22 X lo6 sq. cm. per mole being used for the difference between TPNH and TPN. For cytochrome c, the reaction was followed at 550 rnh by use of AE = 18.5 X lo6 (21) for the difference between oxidized and reduced cyto- chrome c, and at 340 rnp for TPNH (AC = 0 for oxidized and reduced cytochrome c at 340 mp). For MHb, the reaction was followed at 630 m/L (AE = 4.0 X lo6 (15))) at 575 rnp (A6 = 14.0 X lo6 (15)), and at 340 mp (AE = 7.0 X lo6 for MHb and HbOz at 340 mp). The rate of reduction of MHb calculated from the optical density changes at 630 rnp was 3.0 X 10-r, and 3.6 X 10-7, the changes at 575 rnp being utilized. The latter value was used, since the Alle at 575 rnp is larger. All the rates were taken over an initial period (0 to 5 minutes) and were corrected for blanks without enzgme.
340 rnp, were 22.5 X lo-’ and 10.0 X lo-‘, it is apparent that the differ- ences, 2.7 X lo-’ and 8.2 X lo-‘, represent the oxidation of TPNH due to oxygen, acting in competition with the porphyrins.
When oxygen is used alone as the acceptor, the rate of TPNH oxidation is 12.5 X lo-’ mmole per ml. per minute. From this value the rate of oxy- gen reduction would be either 6.3 X lo-’ or 12.5 X lo-‘, depending upon whether Equation 2 or 3 is operative.
TPNH + H+ + $0, -+ TPN+ + Hz0 (2)
TPNH + H+ + 02 --t TPN+ + Hz02 (3)
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from

270 METHEMOGLOBIN REDUCTASE
At present, it has not been possible to determine conclusively whether H20 or Hz02 is the product of oxygen reduction in this system.‘O
It is apparent that cytochrome c is a preferred acceptor to oxygen, for the initial concentration of the heme compound was approximately 3 X 10e5 M, as compared to the oxygen concentration of approximately 2.4 X l(k4 M (22).
The favored utilization of cytochrome c by the enzyme is somewhat sur- prising, inasmuch as the mature erythrocyte contains no endogenous cyto- chrome c. It is possible that, in the nucleated or immature stage, the erythrocyte uses the enzymes as a cytochrome reductase, but that, upon maturation and the concomitant reversion to a more anaerobic type of me- tabolism, the enzyme functions only as a methemoglobin reductase.
Electron Transport Scheme-From the evidence presented above, it seems permissible to formulate the electron transport scheme of the enzyme as follows:
TPNH 5 MeB -% heme -% Oz, MHb, or cytochrome c (4)
The enzyme is thus considered to have two prosthetic groups: (1) an un- known carrier, which is detached during purification and must be substi- tuted for by methylene blue or other autoxidizable dyes; and (2) a tightly bound iron porphyrin moiety. Under anaerobic conditions it can be shown7 that TPNH reacts first with the dye, t,hat the reduced dye inter- acts readily with the iron porphyrin, and that the iron porphyrin, in turn, reduces molecular oxygen. Under aerobic conditions it is possible that the reduced dye would interact directly with oxygen, thus by-passing the por- phyrin component. However, we have been unable to demonstrate the formation of hydrogen peroxide under these conditions.l”
Role of Methylene Blue and Other Carriers-As shown in this investiga- tion and previously (5), the enzyme, as isolated from human erythrocytes, has an absolute requirement for methylene blue, or other autoxidizable dyes, as an electron carrier. Toxoflavin, a pigment elaborated by Pseudo- monas cocovenen (Bacterium bongkrek), also stimulates the enzyme (23). In the search for the physiological electron carrier, a great variety of known coenzymes and metal ions were tried, including riboflavin, flavin mono- nucleotide, flavin adenine dinucleotide,” reduced glutathione, ascorbic acid,
10 When excess catalase and ethanol were added to the manometric system, there was observed no doubling of the oxygen uptake, as noted previously with the DPNH oxidase from pig heart (17). This fact would suggest that oxygen is being reduced to water rather than to hydrogen peroxide in this system.
11 The purified enzyme contains no flavin, as evidenced by the absorption spectrum (see Fig. 1) or by paper chromatographic examination (24) of the supernatant fluid of heat- and trichloroacetic acid-denatured preparations.
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from

HUENNEKENS, CAFFREY, BASFORD, AND GABRIO 271
and ergothioneine. All were without effect. Recently, we have found that the methemoglobin reductase, isolated from beef erythrocytes, does not re- quire methylene blue for activity. When the beef blood enzyme is boiled, a material is obtained which activates the human reductase.7 The puri- fication and properties of this natural cofactor are currently under investi- gation.
It is of interest to recall that the disease, congenital methemoglobinemia, which is characterized by steady state levels of MHb in the erythrocytes as high as 10 to 30 per cent, has been treated successfully by periodic in- fusion of massive amounts of methylene blue (25).
The authors are indebted to Dr. Preston Talbert for his contribution to this problem and to Dr. Clement Finch for many helpful discussions on the clinical aspects of methemoglobinemia.
SUMMARY
1. Reduced triphosphopyridine nucleotide oxidase, or methemoglobin re- ductase, has been isolated from hemolysates of human, mature erythrocytes by means of ethanol-chloroform treatment, precipitation of inert proteins at pH 5.4, and ammonium sulfate fractionation.
2. The enzyme is a hemoprotein with a molecular weight of approxi- mately 185,000, based upon pyridine hemochromogen and total iron de- terminations.
3. The enzyme oxidizes reduced triphosphopyridine nucleotide, and to a lesser extent reduced diphosphopyridine nucleotide, with oxygen, methe- moglobin, or cytochrome c as the terminal electron acceptor. Methylene blue is required as an electron carrier.
BIBLIOGRAPHY
1. Huennekens, F. M., and Gabrio, B. W., Federation Proc., 13, 232 (1954). 2. Warburg, O., Kubowitz, F., and Christian, W., Biochem. Z., 227, 245 (1930). 3. Harrop, G. A., and Barron, E. S. G., J. Ezp. Med., 48, 207 (1928). 4. Michaelis, L., and Salomon, K., J. Gen. Physiol., 13, 683 (1930). 5. Huennekens, F. M., Liu, L., Myers, H. A. P., and Gabrio, B. W., J. Biol. Chem.,
227, 253 (1957). 6. Bodansky, O., Pharmacol. Rev., 3, 144 (1951). 7. Kiese, M., Biochem. Z., 316, 264 (1944). 8. Drabkin, D. L., Arch. Biochem., 21, 224 (1949). 9. Austin, J. H., and Drabkin, D. L., J. Biol. Chem., 112, 67 (1935-36).
10. Grafflin, A. L., and Ochoa, S., Biochim. et biophys. acta, 4, 205 (1950). 11. Weichselbaum, T., Am. J. Clin. Path., 7, 40 (1946). 12. Basford, R. E., Tisdale, H. D., Glenn, J. L., and Green, D. E., Biochim. et biophys.
acta, in press. 13. Paul, K.-G., Theorell, H., and Akeson, A., Acta &em. Scud., ‘7, 1284 (1953).
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from

272 METHEMOGLOBIN REDUCTASE
14. Sandell, E. B., Calorimetric determination of traces of metals, New York (1944). 15. Horecker, B. L., in Lemberg, R., and Legge, J. W., Hematin compounds and bile
pigments, New York, 748 (1949). 16. Morrison, M., and Cook, J. L., Science, 132, 920 (1955). 17. Huennekens, F. M., Basford, R. E., and Gabrio, B. W., J. Biol. Chem., 213, 951
(1955). 18. Mahler, H. R., Sarkar, N. K., Vernon, L. P., and Alberty, R. A., J. Biol. Chem.,
199, 585 (1952). 19. Haas, E., Horecker, B. L., and Hogness, T. R., J. BioZ. Chem., 136, 747 (1940). 20. Altman, K. I., Biochim. el biophys. acta, 16, 155 (1954). 21. Margoliash, E., Biochem. J., 66, 535 (1954). 22. Chance, B., and Williams, G. R., J. Biol. Chem., 21’7, 383 (1955). 23. Stern, K. G., Biochem. J., 29, 500 (1935). 24. Kilgour, G. L., Felton, S. P., and Huennekens, F. M., J. Am. Chem. Sot., 79, 2254
(1957). 25. Wintrobe, M. M., Clinical hematology, Philadelphia, 147 (1951).
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from

technical assistance of L. LiuBasford, Beverly W. Gabrio and With the
F. M. Huennekens, Ruth Wade Caffrey, R. E.METHEMOGLOBIN REDUCTASEISOLATION AND PROPERTIES OF
ERYTHROCYTE METABOLISM: IV.
1957, 227:261-272.J. Biol. Chem.
http://www.jbc.org/content/227/1/261.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/227/1/261.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on July 4, 2018http://w
ww
.jbc.org/D
ownloaded from