the mechanical properties of the longitudinal … mechanical properties of the longitudinal muscle...
TRANSCRIPT

J. Exp. Biol. (1969), 50, 431-443 4 3 IWith 8 text-figuresPrinted in Great Britain
THE MECHANICAL PROPERTIES OF THE LONGITUDINALMUSCLE IN THE EARTHWORM
BY T. HIDAKA, H. KURIYAMA AND T. YAMAMOTO
Department of Physiology, Faculty of Medicine, Kyushu University, Fukuoka, Japan
{Received 10 July 1968)
INTRODUCTION
The longitudinal muscle of the earthworm is currently thought to consist of smooth-muscle fibres which contain intracellular supporting fibres arranged in a regularpattern.
Electron-microscopic examination, however, showed that the muscle fibres werecomparable with the striated muscle of vertebrates in that they contained inter-digitating arrays of thick and thin filaments. The membranes of the individual cellscontained desmosome or half-desmosome structures which were in contact with eachother. Just beneath the cell membrane large vesicles could be observed and they wereconnected with transverse tubules. The transverse tubules ran regularly amongst thecontractile proteins and connected one side of the membrane to another (Nishihara,1967).
The previous papers (Hidaka, Ito & Kuriyama, 1969; Hidaka, Ito, Kuriyama &Tashiro, 1969a, b) were concerned with the membrane properties, ionic effects andthe neuromuscular junctions in the longitudinal muscle of the earthworm, andtherefore this paper deals with a study of the mechanical properties of the longitudinalmuscle, carried out to investigate the specific features of this muscle in relation to itsmembrane properties.
EXPERIMENTAL METHODS
The membrane potentials, passive properties of the membrane and the spikesrecorded in these experiments were measured by the same methods as described inthe previous papers (Hidaka, Ito & Kuriyama, 1969). Tension development wasmeasured by the transducer produced by Nihon Kohden Ltd. The stimulatingelectrodes (platinum wire, 0-2 mm. in diameter) were arranged as a multigrid electrodealong a piece of tissue, 2 mm. wide, parallel with it for 10 mm. The stimulatingcurrent therefore passed transversely across the tissue with uniform intensity. Theionic composition of the Ringer's solution used in this experiment was the same asdescribed in the previous paper (Hidaka, Ito & Kuriyama, 1969).
RESULTS
Tension development related to membrane activity
The longitudinal muscle of earthworm was difficult to dissect completely free fromother tissues; the description of the results on the magnitude of the tension is thereforeonly qualitative.
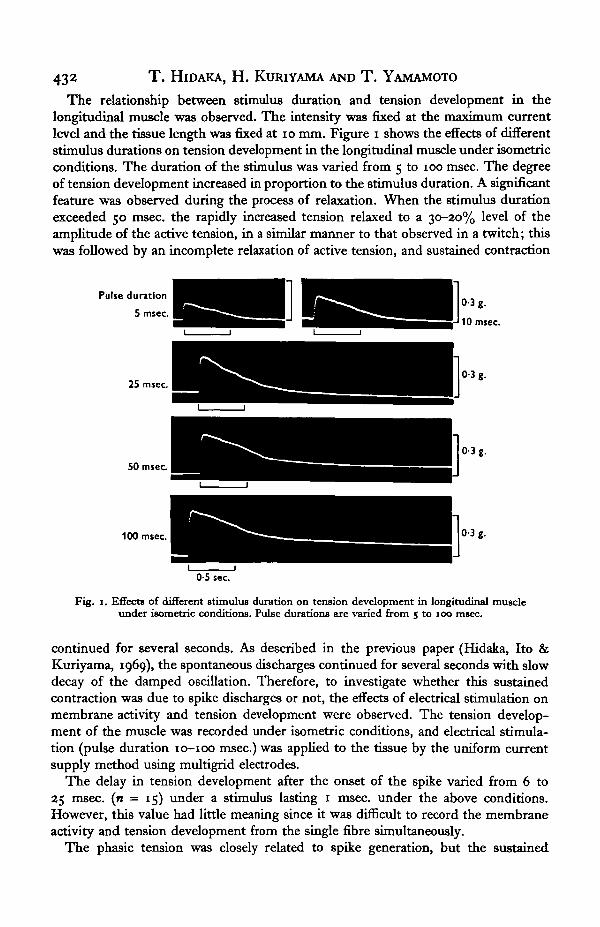
432 T . HlDAKA, H. KURIYAMA AND T . YAMAMOTO
The relationship between stimulus duration and tension development in thelongitudinal muscle was observed. The intensity was fixed at the maximum currentlevel and the tissue length was fixed at 10 mm. Figure i shows the effects of differentstimulus durations on tension development in the longitudinal muscle under isometricconditions. The duration of the stimulus was varied from 5 to 100 msec. The degreeof tension development increased in proportion to the stimulus duration. A significantfeature was observed during the process of relaxation. When the stimulus durationexceeded 50 msec, the rapidly increased tension relaxed to a 30-20% level of theamplitude of the active tension, in a similar manner to that observed in a twitch; thiswas followed by an incomplete relaxation of active tension, and sustained contraction
Pulse duration
5 msec.
25 msec
50 msec
100 msec 0-3 g.
0-5 sec
Fig. 1. Effects of different stimulus duration on tension development in longitudinal muscleunder isometric conditions. Pulse durations are varied from 5 to 100 msec.
continued for several seconds. As described in the previous paper (Hidaka, Ito &Kuriyama, 1969), the spontaneous discharges continued for several seconds with slowdecay of the damped oscillation. Therefore, to investigate whether this sustainedcontraction was due to spike discharges or not, the effects of electrical stimulation onmembrane activity and tension development were observed. The tension develop-ment of the muscle was recorded under isometric conditions, and electrical stimula-tion (pulse duration 10-100 msec.) was applied to the tissue by the uniform currentsupply method using multigrid electrodes.
The delay in tension development after the onset of the spike varied from 6 to25 msec, (n = 15) under a stimulus lasting 1 msec, under the above conditions.However, this value had little meaning since it was difficult to record the membraneactivity and tension development from the single fibre simultaneously.
The phasic tension was closely related to spike generation, but the sustained

Mechanical properties of earthworm longitudinal muscle 433
contraction was not so related. The sustained contraction never started at the peakof phasic tension in the physiological solution and the transition from phasic tensionto a sustained contraction could be observed clearly (Fig. 1 at 100 msec, pulse dura-tion). Summation of the twitch tension was observed on repetitive stimulation, with1 msec, pulses, for 5-7-5 sec.
The relationship between tension development and the membrane potential wasinvestigated. The membrane potential was varied by changing the external potassiumconcentration, [K]o. Figure 2 shows the relationship between tension development(contracture) and [K]o expressed on a logarithmic scale. The changes in membranepotential produced by various [K]o are also illustrated. When the tissue was exposed
1 0
Tens
ion
o Cn
-
-
- # 1—•- t 1
•*•••
••
t:#t•
: •
i i
2-7 10 20 30 50 80 m. (K)o
30 835-6-26
10
215-24-19
0-8
14-6 mV.16-2-13 mV.
03
36-2S.D. 40-6-31-8s.E 0-7
Fig. 2. Relationship between tension development and the changes in membrane potentialunder various external potassium concentrations, expressed on a logarithmic scale.
to a solution containing excess potassium (more than 30 mM), tension developedrapidly and was sustained at maximum amplitude for a few seconds, then graduallydecreased, but never returned to the normal resting state. The values plotted inFig. 2 were measured after exposing the tissue for 15 min. which was the timewhen stable amplitudes of the contracture were obtained.
The membrane potential in the normal solution (K+ = 27 mM) was - 36-2 mV.(s.D. = ± 2-6, n = 150) and when [K]o was increased up to 20 mM, the resting tensionincreased slightly, and the maximum upstroke of the amplitude of the contractureagainst the increased [K]o appeared at a potassium chloride concentration of 30 mM.

434 T. HroAKA, H. KURIYAMA AND T. YAMAMOTO
Further increase in [K]o increased the amplitude of the contracture but with widevariation. A [K]o in excess of 80 mM lowered the amplitude of the contracture. Theonset of a contracture appeared when the membrane was depolarized to — 20-5 mV.(S.D. = ± 2.8, n = 12). This critical membrane potential required to develop tensionwas very low compared with that observed in frog skeletal muscle (Hodgkin &Horowicz, i960) and in rabbit heart muscle (Liittgau, 1965). If the contracture wasgenerated by a similar mechanism to the twitch, a depolarization of approximately15 mV. might trigger phasic tension development.
Longitudinal muscle
I 400
1 -
1-5
g<4 -
OS
Resting tension
Total tension
Phasic tension
2-5Tissue length
Circular muscle
3 cm
Resting tension
Phasic tension
0-6 07 08Tissue length
0-9 cm.
Fig. 3. The relation between muscle length and tension development in both the longitudinalmuscle and the circular muscle, x , Resting tension; O, phasic tension; # , total tension.
Length-tension curve
The relation of muscle length to the tension developed was studied in both thelongitudinal muscle and the circular muscle of the earthworm, and the curves wereplotted in Fig. 3. The figure shows the phasic tension (active tension) elicited by

Mechanical properties of earthworm kmgitudinal muscle 435
electrical stimulation (1 msec.), the resting tension and the sum of the two (totaltension). It may be seen that the preparations developed phasic tension at far shorterlength than those at which resting tension developed (1*9 mm. in longitudinal muscleand o-6 mm. in circular muscle). The peak of the active tension was, however, notobserved at the point where the resting tension developed, and It was not sharply
ControlNa+ 140 0-5 g.
0-5 g.
0-5 g.
0-5 g.
1 sec
Fig. 4. Effects of various external sodium concentrations (substituted tris) on tension develop-ment; linear sustained lines indicate sustained contraction. Pulse duration, 10 msec.;frequency, O'2$ c./sec. Five successive stimuli -were applied .Tension wa» measured after 20 min.of exposure to the test solution.
defined. The active tension only declined when the preparation was severely stretched.In this respect the longitudinal and circular muscle of the earthworm differs fromskeletal muscle, and these muscles resembled the papillary muscle of the heart andthe taenia coli of the guinea-pig (Hill, 1951-2; Csapo & Goodall, 1954; Abbott &Mommaerts, 1959; Bulbring & Kuriyama, 1963). The smooth transition of the totaltension development is due to the fact that the different grades of stretch do notproduce sharply denned active tension.
a8 Exp. BioL 50, 3

436 T . HlDAKA, H . KURIYAMA AND T . YAMAMOTO
The effects of ions on the tension development
The effects of various ions (Na+, K+, Ca*4" and Cl~) on tension development wereinvestigated. The stimulus conditions were fixed as follows: d.c. pulses each lastingi msec, constant intensity, and a stimulus frequency of 0*4 c/sec. The effects of theabove ions were examined in three of four specimens. Five successive responses weresuperimposed in the same photograph.
Na+ (140 Na-free (Trls)
Control
K* 27 mM
6-8 mM
K* 13 5 mM
K* 27 mM
05 g.
0-5 g.
1 sec. 1 sec
Fig. s. Effects of various potassium concentrations in the presence of sodium and in sodium-freesolution. The solutions were maintained isotonic. The stimulus conditions to the tissue werethe same as for Fig. 4.
Figure 4 shows the effects of varying [Na]0 on tension development. Repetitivestimulation in both normal and sodium-deficient solution (substituted by tris) enhancedthe amplitude of the phasic tension development, to a constant extent at the crest.Reduction of [Na]0 in steps proportionally reduced the rates of rise and fall of the phasictension although the amplitude of the phasic tension was increased. Furthermore,sustained contractions were enhanced in sodium-deficient solution. This result alsoindicates that sustained contractions are not related to spike generation, since thespontaneous spike discharges and the spikes elicited by electrical stimulation were

Mechanical properties of earthzoorm longitudinal muscle 437
reduced in number in the train discharges in sodium-free solution (Hidaka, Ito &Kuriyama, 1969). The sustained contractions were, however, smaller in amplitude insodium-free solution than those recorded in a solution containing 14 mM sodium.
To investigate the effect of potassium ions solid KCl was added in the Ringersolution. Figure 5 shows the effects of [K]o in the presence of sodium and in sodium-free solution. In the presence of a normal sodium concentration (140 mM —Na+) theamplitude of the phasic tension developed was increased KCl concentration up to13*5 mM. Further increase of [K]o (> 30 mM) reduced or blocked the phasictension development, but developed a contracture, as described previously.The sustained tension, however, was not enhanced in either amplitude or
CaCI, 0-36 mM Control 1-8 mM
0-5 g.
1 sec.
4-5 mM 9 mM 18 mM
0 5g.
1 sec.Fig. 6. Effects of various calcium concentrations on tension development in longitudinal muscle.Calcium concentrations varied from 0-36 to 18 mM. The stimulus conditions were the sameas for Fig. 4.
duration in the excess-potassium solution. This inhibitory action on the sustainedcontractions by the excess potassium was also observed in sodium-free solution,i.e. even in sodium-free solution, excess potassium up to 13-5 mM enhanced phasictension development but reduced the amplitude of the sustained contractions.As a result, no summation of the phasic tension was observed and the sustainedcontraction ceased.
The phasic tension recorded in sodium-free solution was always higher than thatrecorded in solutions containing sodium since the membrane was more hyper-polarized in the sodium-free solution than in solutions containing sodium.
Figure 6 shows the effects of various calcium concentrations on tension develop-ment in longitudinal muscle. Excess calcium enhanced phasic tension development butreduced the amplitude of the sustained contractions. The reduction of the externalcalcium concentration of i-8 mM slightly enhanced the amplitude of the phasictension. However, a fifth of the normal calcium concentration reduced both thephasic tension and the sustained contractions. When the external chloride was replacedin stages by D-glutamate at concentrations from 148 to 5-6 mM the amplitude and
38-a

T . HEDAKA, H . KURIYAMA AND T . YAMAMOTO
duration of the phasic tension was not enhanced but those of the sustained contractionswere.
The effects of the various ions on tension development could be expected from thechanges in the membrane potential effected by the various ionic environments.Reduction of [Na]0 hyperpolarized the membrane and enhanced the amplitude of thephasic tension development, and increased [Ca]0 had effects similar to those observedin sodium-deficient solution. An inverse proportional relationship was observed inthe excess-potassium solutions until the membrane was depolarized to — 30 mV. Thevarious grades of chloride-deficient solutions (140-5-6 mM) did not change the mem-brane potential, and the amplitude of the active tension also remained nearly thesame.
Control Caffeine 0-25%
0-5 g.
0-3 g.
5 sec.
Fig. 7. Effects of caffeine (12 mM) on the tension development of longitudinal muscle,(a) Control; (b) after 15 min. of treatment with caffeine; (c) continued record of the tensiondevelopment by successive stimulation (5 msec., 0-25 c./sec.) after 20 min. of exposure.Stimulus conditions were the same as described for Fig. 4.
The effects of caffeine on membrane activity and tension development
Figure 7 shows the effects of caffeine (12 mM) on the tension development oflongitudinal muscle. The stimulus conditions were fixed at pulses of 5 msec, durationand stimulus frequency of 0-25 c/sec., and five successive stimuli were applied to thetissue.
In the control (a) phasic tension was not followed by a sustained contraction, but asecond stimulus enhanced the amplitude of the phasic tension with a similar cresttime.
After 15 min. exposure in caffeine-Ringer solution the phasic tension was enhancedin amplitude and its duration was prolonged, causing significant increase in the totaltension developed (i). After 20 min. exposure in caffeine—Ringer solution the relaxa-tion of the phasic tension was prolonged and differentiation between the delayedrelaxation and the sustained contraction became difficult (c).
During the potentiation which occurred in tension development, the membrane

Mechanical properties of earthworm longitudinal muscle 439
activity was recorded by the intracellular polarizing method. The mean values of theeffective resistance and the time constant of the membrane have been describedpreviously (Hidaka, Ito & Kuriyama, 1969).
The effect of caffeine (12 mM) on the membrane activity elicited by an intracellularpolarizing current was observed. The critical membrane potential required to elicita spike was 9 mV. in the normal solution, but a depolarization of more than 20 mV. didnot elicit any spike in the caffeine—Ringer solution.
2I
ControlCaffeine
/
x10"'A.)11 A
78 MO /: 36 Mfl / /
*/ 1m/ 1 —
/ / "0-25% caffeine J
/Control
10
20
30
40
50
60
70
80
mV.
Fig. 8. Current—voltage relationship in normal solution and in caffeine—Ringer solution (ia mM).
The current-voltage relationship was also observed in caffeine-Ringer solution.Figure 8 shows the current-voltage relationship curves in normal solution and incaffeine-Ringer solution, recorded from the same cell. In this cell the effectiveresistance of the membrane was initially 78 MD (s.D. = ±4*5, n = 11) but after20 min. exposure in caffeine-Ringer solution (12 mM) the membrane resistance haddropped to 35 MD (s.D. = ±3-1, n = 10).
When depolarizing currents were applied to the tissue, rectification by the mem-brane was observed in the current-voltage curve in caffeine-Ringer solution. Thetime-constant of the membrane, measured at 84% of the steady level of the electro-tonic potential produced by applying weak hyperpolarization, was 140-0 msec.(s.D. = ± 18, n = 8), and this value was reduced to 40-7 msec. (s.D. = ± 11, n = 8)in caffeine-Ringer solution. The calculated membrane capacitance was 1-7 x io^/ tF. /cm.2 in the normal solution and it was reduced to 0-9 x io"3 /iF./cm.2 in the solutioncontaining caffeine. This result indicates that caffeine reduces not only the membraneresistance but also the membrane capacitance.
The effect of 5-hydroxytryptamine on tension development was also observed,io~* g./ml. 5-hydroxytryptamine having no effect on the phasic tension or on the

44° T . HlDAKA, H. KURIYAMA AND T . YAMAMOTO
sustained contraction. When the concentration increased up to io"6 g./ml., the sus-tained tension ceased completely without any changes of the phasic tension. Theseresults confirmed the observation made on the 'catch' muscle of Mytihu (ABRM) byTwarog (19670,^), i.e. 5-hydroxytryptamine blocked generation of the catch tensiononly, while the phasic tension remained as it was before the treatment.
DISCUSSION
Electron-microscopic examination of the longitudinal muscle of the earthworm hasshown that the muscle fibres are comparable with the striated muscle of vertebratesin that they contain inter-digitating arrays of thick and thin filaments. The peripheralthick filaments are surrounded by thin filaments but the central thick filaments arenot surrounded by thin filaments. Two kinds of tubular structures were observed inthe muscle cell: open tubules and packed tubules. Both types of tubule are distributedregularly and the open tubules might correspond to the so-called transverse tubuleswhile the others might correspond to the J-rod, which is thought to be analogous to theZ-line in vertebrate striated muscle. The bridge structure distributed between theactin and the myosin in vertebrate skeletal muscle has not yet been observed in thelongitudinal muscle of the earthworm (Ikemoto, 1963; Nishihara, 1967).
At the present time it is difficult to discuss in detail the mechanical properties of thelongitudinal muscle in relation to its structure. Sustained contraction and activetension (twitch or phasic tension) in the earthworm were distinguished according tothe type of stimulus, the tissue being bathed in various ionic environments. Thetwitch tension had roughly similar properties to those observed in striated muscle andcardiac muscle. The positive staircase effect on the twitch tension produced by repeti-tive stimulation at low frequency was observed in this tissue, as in cardiac muscle(cf. the review of Sandow, 1965). In sodium-deficient solution the amplitude of thetwitch tension in skeletal muscle was enhanced, in spite of reductions in the spikeamplitude (Nastuk & Hodgkin, 1950). In the longitudinal muscle of the earthwormthe amplitude of the active tension was enhanced by reduction in (NaJo, but the spikeamplitude remained the same. Excess potassium depolarized the membrane andreduced the spike amplitude, but enhanced the amplitude of the active tension up toa (K)o of 20 mM, as observed in skeletal muscle. Excess calcium enhanced the amplitudeof the active tension and deficiency of calcium reduced it; these effects were welldefined in both cardiac and skeletal muscle (Luttgau & Niedergerke, 1958; Davis,1963; Sandow, 1965; Luttgau, 1965). When the external Cl~ was replaced by NO8~,the twitch tension of frog skeletal muscle was enhanced in amplitude without anychange in spike amplitude (Kahn & Sandow, 1950). This effect was confirmed byHodgkin & Horowicz (i960), although the amplitude of the twitch tension in thelongitudinal muscle of earthworm was only slightly increased in the presence ofnitrate (unpublished observation), and the substitution of glutamate for chloride didnot enhance the amplitude of twitch tension .development, unlike the situation in frogskeletal muscle (Mashima & Matsumura, 1962).
However, changing the various ionic concentrations affected the sustained con-tractions differently from the twitch tension, i.e. excess calcium showed no remarkableeffect, excess potassium stopped the generation of sustained contractions, and

Mechanical properties of earthworm longitudinal muscle 441
reduction in sodium enhanced the amplitude and prolonged the duration of thesustained contractions.
Von Uexkull (1907, 1912, 1929; referred to by Johnson, 1962) showed that theadductor muscles of the bivalve molluscus could remain contracted for considerableperiods of time without an intact innervation and apparently with little utilization ofmetabolic energy. Similar observations were also made on another muscle, namelythe anterior byssus retractor muscle of Mytihis eduUs (Winton, 1937). The name'catch mechanism* was proposed for this mechanism. A significant feature in this' catch' muscle was observed to be the presence of paramyosin as contractile protein.The mechanical properties of 'catch' muscle have been extensively studied by Jewell(1959), Lowy & Hanson (1962), Johnson & Twarog (i960) and Twarog (1967a, b, c).The sustained contraction recorded from longitudinal muscle in our experimentscannot be directly compared with the ' catch' phenomena, since the structural speci-ficity of the contractile protein has not been investigated. Furthermore, Bozler (1962)observed that the relaxation of smooth muscle became progressively slower during aseries of responses, and also became more economical in the maintenance of tension,as shown by measurements of heat production, and he thought that the tonic statewas not sharply distinguishable from the phasic state, but he agreed that in 'catch'muscle paramyosin might play a part in sustaining the tonus. The sustained contrac-tions in earthworm muscle did not follow spike discharges, as observed in 'catch'muscle, although the contractions lasted less than 1 min. under normal conditions.Treatment with caffeine or acetylcholine, however, prolonged relaxation for more thanseveral minutes. In contrast with the above chemicals, 5-hydroxytryptainine completelyblocked the generation of the sustained tension without any influence on the phasictension as observed in the 'catch' muscle by Twarog (1967a, b).
The effects of caffeine on tension development have been studied by Gasser (1930)and Saslow (1937); however, since the extensive investigation of Axelsson & Thesleff(i958),manyrelevant experiments have been carried out. At low concentrations caffeinelowered the mechanical threshold, prolonged the mechanical effective period, reducedthe time required to reach maximum tension and prolonged the active state. Theseactions of caffeine are thought to be produced by potentiation of twitch-tensiondevelopment. At high concentrations caffeine induced contracture of the muscle,thought to be due to activation of the initial step needed to trigger a contracture, butnot via the spike-activation link (Sandow, 1965). Both the potentiation and the con-tractures produced by caffeine are thought to be related to the release of calcium ionsfrom the sarcoplasmic reticulum. In longitudinal muscle, caffeine, in fact, loweredthe membrane potential, reduced the membrane resistance and capacitance andblocked spike generation. Yet electrical stimulation produced active tension with aprolonged relaxation time, and without spike generation. These facts also indicatethat caffeine produces contractures, not through the spike-generation link, but by directaction on the activation step of the mechanical changes, probably involving the sarco-plasmic reticulum. It has been postulated that spike generation in longitudinal muscleis due to the inward movement of calcium ions (Hidaka, Ito & Kuriyama, 1969).If caffeine also releases the bound calcium as free ions in the cell, as observed inskeletal muscle (Bianchi, 1961,1962), the block of spike generation might be explainedby a shift in the calcium equilibrium potential to zero or even to a negative value.

442 T . HlDAKA, H. KURIYAMA AND T . YAMAMOTO
Furthermore, caffeine reduced the membrane capacitance of the longitudinalmuscle to a half of the value before treatment. This result also might suggest that themolecular structure not only of the cell membrane but also of the sarcoplasmicreticulum were rearranged by caffeine: the capacitance of skeletal muscle and cardiacmuscle were thought to be composed of the capacitances of both the sarcoplasmicmembrane and the tubular system (Falk & Fatt, 1964; Adrian & Peachey, 1965;Fozzard, 1966). The large vesicles distributed just beneath the sarcoplasmic membranecompose a diad structure with the sarcoplasmic membrane and thus might easily beaffected by external treatment with caffeine.
SUMMARY
1. A study of the mechanical properties of longitudinal muscle in relation to themembrane properties was carried out under isometric conditions.
2. When the stimulus duration exceeded 50 msec., active tension development wasfollowed by sustained contraction. The sustained contractions were not-related tospike generation.
3. The critical potassium concentration to produce the contracture depolarized themembrane from —36 to —20 mV.
4. Reduced sodium concentration and increased calcium and increased potassium(up to 27 HIM) concentrations enhanced the amplitude of the active tension.
5. Reduced sodium concentration enhanced the amplitude and duration of thesustained tension, but increased potassium and calcium concentrations reduced them.
6. Caffeine (12 HIM) induced contractures of the muscle, and reduced the membraneresistance and capacitance.
7. Spikes were not elicited by intracellular stimulation.- 8. 5-Hydroxytryptamine (io~* g-/ml.) blocked the generation of the sustainedcontraction but no effect was observed on the phasic tension.
REFERENCES
ABBOTT, B. C. & MOMMAERTS, W. F. (1959). A study of iontropic mechanisms in the papillary musclepreparation. J. gen. Pkytiol. 42, 533-51.
ADMAN, R. H. & PEACHEY, L. D. (1965). The membrane capacity of frog twitch and slow muscle fibres.J. Pkysiol., Land. 181, 324-36.
AxELSSON, J. & THESLEFF, S. (1958). Activation of the contractile mechanism in striated muscle. Acta.pkytiol. tcand. 44, 55-66.
BIANCHI, C. P. (1961). The effect of caffeine on radio calcium movement in frog sartorius. J. gen Physiol.44. 845-58.
BIANCHI, C. P. (1962). Kinetics of radio caffeine uptake and release in frog sartorius. J. Pharmacol. 138,4I-7-
BOZLER, E. (196a). Initiation of contraction in smooth muscle. Pkysiol. Rev. 43, 179-86.BtJisMNO, E. & KURIYAMA, H. (1963). The effectof adrenaline on the smooth muscle of guinea-pig taenia
coli in relation to the degree of stretch. J. Pkysiol., Lond. 169, 198-212.CSAPO, A. & GOODAIX, M. C. (1954). Excitability length-tension relation and kinetics of uterine contrac-
tion in relation to hormonal status. J. Pkysiol., Lond. 136, 384-95.DAVIS, R. E. (1963). A molecular theory of muscle contractions. Calcium-dependent contraction with
hydrogen bond formation plus ATP-dependent extensions of part of the myosin-actin bridges.Nature, Lond. 199, 1068—074.
FALK, G. & FATT, P. (1964). Linear electrical properties of striated muscle fibres observed with intra-cellular electrodes. Proc. R. Soc. B 160, 69-123.
FOZZARD, H. A. (1966). Membrane capacity of the cardiac purkinje fibre. J. Pkysiol., Lond. 18a, 255-67.

Mechanical properties of earthworm longitudinal muscle 443GASSER, H. (1930). Contractures of skeletal muscle. Pkytiol. Rev. 10, 35-109.HIDAKA, T., ITO, Y. & KURIYAMA, H. (1969). Membrane properties of the somatic muscle (obliquely
striated muscle) of earthworm. J. exp. Biol. 50, 387-403.HIDAKA, T., ITO, Y., KURIYAMA, H. & TASHIRO, N. (1909a). Effects of various ions on the resting and
active membrane on the somatic muscle of earthworm. J. exp. Biol. 50, 405-415.HIDAKA, T., ITO, Y., KURIYAMA, H. & TASHIRO, N. (1969ft). Neuromuscular transmission in the
longitudinal layer of somatic muscle in the earthworm. J. exp. Biol., 50, 417-430.HTI.T., A. V. (1951—2). A discussion on the thermodynamics of elasticity in biological tissues under the
leadership of A. V. Hill. Proc. R. Soc. B 139, 464-526.HODGKM, A. L. & HOROWICZ, P. (i960). Potassium contractures in single muscle fibres. J. Pkytiol.,
Lond. 153, 386-403.IKKMOTO, N. (1963). Further studies in electron microscopic structures of the obliquely-striated muscle
of the earthworm, Eisema factida. Biol. J. Okayama Univ. 9, 81-126.JEWELL, B. R. (1959). Nature of the phasic and tonic responses of the anterior byssus retractor muscle of
Mytilus. J. Pkytiol., Lond. 149, 154-77.JOHNSON, W. H. (1962). Ton?ic mechanisms in smooth muscle. Pkytiol. Rev. 43, 113-59.JOHNSON, W. H. & TWAROG, B. M. (i960). The basis for prolonged contractions in molluscan muscle.
J. gen. Pkytiol. 43, 941-60.KAHN, A. J. & SANDOW, A. (1950). The potentiation of muscular contraction by the nitrate ion. Science,
N. Y. 113, 647-9.LOWY, J. & Hanson, J. (1962). Ultrastructure of invertebrate smooth muscles. Pkytiol. Rev. 43, 34-42.L/OTTGAU, H. C. (1965). The effect of metabolic inhibitors on the fatigue of action potential in single
muscle fibres. J. Pkytiol., Lond. 178, 45-67.LtfTTGAU, H. C. & NffiDERGERKE, R. (1958). The antagonism between Ca and Na ions on the frog's heart.
J. Pkytiol, Lond. 143, 486-505.MASHIMA, H. & MATSUMURA, M. (1962). Roles of external ions in the excitation-contraction coupling
of frog skeletal muscle. Jap. J. Pkytiol. 13, 639-53.NASTUK, W. L. & HODGKIN, A. L. (1950). The electrical activity of single muscle fibres. J. Cell. Comp.
Pkytiol. 35, 39-74.NISHIHARA, H. (1967). The fine structure of the earthworm body wall muscle. Acta. Anat. Nippon 43,
38-9.SANDOW, A. (1965). Excitation-contraction coupling in skeletal muscle. Pharmac. Rev. 17, 365-320.SASLOW, G. (1937). Oxygen consumption and respiratory quotient of caffeinized frog muscles. J. Cell.
Comp. Pkytiol. 10, 385-94.TWAROG, B. M. (1967a). The regulation of catch in molluscan muscle. J. gen. Pkytiol. 50, 157-69.TWAROG, B. M. (1967 A). Factors influencing contraction and catch in Mytilus smooth muscle. J. Pkytiol.,
Lond. 193, 847-56.TWAROG, B. M. (1967c). Excitation of Mytilut smooth muscle. J. Pkytiol., Lond. 193, 857—68.WrNTON, F. R. (1937). Changes in viscosity of an unstriated muscle (Mytilut edulii) during and after
stimulation with alternating, interrupted and uninterrupted direct currents. J. Pkytiol., Lond. 88,492-5"-