the map kinase kpp2 regulates mating and pathogenic development in ustilago maydis
TRANSCRIPT

The MAP kinase Kpp2 regulates mating andpathogenic development in Ustilago maydis
Philip Mu ller, Christian Aichinger,Michael Feldbru¨gge and Regine Kahmann *
Institut fur Genetik und Mikrobiologie, Maria-Ward-Str. 1a,D-80638 Munchen, Germany.
Summary
In the phytopathogenic fungus Ustilago maydis ,fusion of compatible haploid cells is a prerequisitefor infection. This process is genetically controlledby the biallelic a locus, encoding pheromone precur-sors and receptors. These are presumed to becoupled to a heterotrimeric G protein and a MAPkinase cascade, leading to activation of the HMGdomain transcription factor Prf1. Here, we havedemonstrated that putative MAP kinase sites in Prf1are required for its activity during mating. In addition,we have identified a gene, kpp2 , which encodes aputative MAP kinase related to Pmk1 of Magnaporthegrisea and Fus3p of Saccharomyces cerevisiae. kpp2deletion mutants are attenuated in several steps ofdevelopment: cell fusion, induction of pheromone-responsive genes and pathogenicity. Epistasis analy-sis shows that kpp2 does not affect pheromone geneexpression through the cAMP signalling cascade.Pathogenicity of kpp2 mutants can be partiallyrestored by overexpressing the b genes, indicating aregulation of Prf1 by Kpp2. These data support thehypothesis that the MAP kinase Kpp2 transmits thepheromone signal.
Introduction
In the large group of smut fungi, pathogenic developmentis initiated after two compatible haploid cells fuse and forma dikaryotic filament. In the most extensively studied mem-ber of this group, the pathogen of corn Ustilago maydis,cell fusion and subsequent pathogenic development arecontrolled by two mating type loci termed a and b (seeBanuett, 1995). The a locus encodes the components ofa cell recognition system, i.e. a pheromone precursorand a seven transmembrane receptor (Bolker et al.,
1992). Haploid cells are cigar shaped and grow in ayeast-like form by budding. Binding of pheromone to itscognate receptor induces the formation of conjugationtubes, which are directed towards each other and fuse attheir tips (Snetselaar et al., 1996). From these fused mat-ing pairs, growth of a dikaryotic filament is initiated.Although this morphological transition can be visualizedon medium plates containing activated charcoal, sustainedgrowth of the dikaryon is observed only after penetrationof plant tissue (Banuett and Herskowitz, 1989; Snetselaarand Mims, 1993). Characteristic for disease developmentis the initial bleaching of leaves and accumulation of antho-cyanins. Subsequently, plant tumours are induced inwhich the fungus proliferates and eventually producesdiploid spores (Christensen, 1963). The entire series ofevents occurring after cell fusion is regulated through theproducts of the multiallelic b locus (Banuett and Hersko-witz, 1989; Bolker et al., 1995a). The b locus codes fortwo homeodomain proteins, bE and bW, which form het-erodimers when derived from different alleles (Gillissenet al., 1992; Kamper et al., 1995). This active heterodi-meric complex then promotes the switch to pathogenicdevelopment.
The HMG domain transcription factor, Prf1, has beenidentified as central in connecting the pheromone-signal-ling pathway with b gene expression. Prf1 binds phero-mone response elements located either 58 or 38 to allpheromone-inducible genes like the mfa pheromone, thepra receptor or the b genes. In addition, Prf1 autoregu-lates its own synthesis (Hartmann et al., 1996). As a result,transcription of all of these genes is strongly increasedafter pheromone stimulation (Urban et al., 1996). Thepheromone signal transduction pathway leading to theactivation of Prf1 has not been worked out in detail.
Recently, the cAMP signalling pathway has also beenshown to affect mating, besides playing an importantrole in regulating cell morphology and pathogenicity (Kron-stad, 1997; Kruger et al., 1998). Several components ofthe cAMP pathway have been identified, including gpa3(encoding a G protein a subunit), uac1 (encoding adeny-late cyclase), ubc1 (encoding the regulatory subunit ofprotein kinase A, PKA) and adr1 (encoding the catalyticsubunit of PKA; Gold et al., 1994; 1997; Durrenberger etal., 1998; Kruger et al., 1998). A reduction in intracellularcAMP level leads to filamentous growth that is indepen-dent of the b locus, whereas an increase in intracellular
Molecular Microbiology (1999) 34(5), 1007–1017
Q 1999 Blackwell Science Ltd
Received 15 June, 1999; revised 7 September, 1999; accepted 10September, 1999. *For correspondence. E-mail [email protected]; Tel. (þ49) 89 2180 6150; Fax (þ49) 89 1785633.

cAMP induces a phenotype termed multiple budding (Goldet al., 1994; 1997). Besides these effects on morphology,cAMP also affects pathogenic development. In particular,it has been demonstrated that gpa3, uac1 and ubc1 deletionmutants are non-pathogenic (Gold et al., 1997; Regenfelderet al., 1997; Durrenberger et al., 1998;). Furthermore, prf1gene expression is regulated by internal cAMP levels, bothtranscriptionally as well as post-transcriptionally, and thisregulation is passed on to the pheromone genes (Krugeret al., 1998; Hartmann et al., 1999). However, the activa-tion of Prf1 via the cAMP pathway does not allow cellfusion in the absence of pheromone stimulation (Krugeret al., 1998). Thus, there must be a second pathway lead-ing to the activation of Prf1.
The hypothesis that the additional route of activation isconferred by a MAP kinase module is supported by the fol-lowing: (i) Prf1 activity is regulated by pheromones on thepost-transcriptional level (Hartmann et al., 1999), (ii) theprotein contains six sequence motifs fitting the consensusL/PXS/ TP of MAP kinase sites (Clark-Lewis et al., 1991),and (iii) Prf1 harbours a putative MAP kinase docking siteof the FXFP type (Jacobs et al., 1999).
To gain more insight into the pheromone signalling path-way leading to the activation of Prf1 we have addressedthe role of the putative MAP kinase sites in Prf1 geneti-cally, and have isolated and characterized a gene encod-ing a MAP kinase implicated in transducing the pheromonesignal.
Results
Putative MAP kinase recognition sites in Prf1 areimportant for its biological activity
In order to investigate whether Prf1 is subject to phosphor-ylation by a MAP kinase, we changed six serine or threo-nine residues at position 3 of the consensus sequenceL/PXS/ TP to alanine (Clark-Lewis et al., 1991). In addition,a presumed MAP kinase docking site TPNFAFDP (Jacobset al., 1999) spanning amino acid positions 586–592, wassubstituted by APNAAAAP. At the same time, the N- andC-termini of Prf1 were modified by a hexa-histidine tagand triple-myc epitope respectively. The resulting alleleis termed prf1-M1 (Fig. 1A). As a control, the alleleprf1-E was constructed, which differs from the wild-typeallele only in the terminal epitopes (Fig. 1A). Thesemutant alleles were introduced into the prf1 locus instrains FB1 (a1b1) and FB2 (a2b2) using homologousrecombination, and the replacement of the wild-type allelewas verified using Southern analysis (data not shown). Toanalyse mating competence, compatible strains wereinoculated on charcoal-containing potato dextrose plates.The formation of dikaryotic hyphae results in white-lookingcolonies, which have a fuzzy appearance. Compatible
wild-type pairs FB1 and FB2, as well as FB1prf1-E andFB2prf1-E, react strongly (Fig. 1B), illustrating that theepitope-tagged allele of prf1 is fully functional. In contrast,FB1, as well as FB2, strains expressing prf1-M1 wereattenuated in the formation of dikaryotic hyphae in crosseswith compatible wild-type strains. When both mating part-ners harboured the prf1-M1 allele, dikaryotic hyphaecould not be detected, a phenotype also seen in crossesof two Dprf1 strains (Fig. 1B). However, strains containingthe prf1-M1 allele behaved differently from Dprf1 strainsin two respects. prf1-M1 strains were able to fuse withwild-type strains and could stimulate filamentous growthin the pheromone tester strain CL13 (a1bE1bW2),whereas Dprf1 mutants were completely sterile andwere unable to produce pheromone (Fig. 1B; Hartmann
Q 1999 Blackwell Science Ltd, Molecular Microbiology, 34, 1007–1017
Fig. 1. Reduced mating competence of prf1-M1 strains.A. Schematic representation of different prf1 alleles: His,N-terminal hexa-histidine tag; Myc, C-terminal triple-myc tag;HMG, HMG box containing DNA-binding domain; black rectangle,MAP kinase docking site of the FXFP type; open rectangle,docking site altered by four amino acid substitutions.B. Strains indicated on the left and on the top were spotted aloneor in combination on charcoal-containing potato dextrose platesand incubated at 248C for 48 h. Dikaryotic filaments form colonieswith a characteristic white, fuzzy appearance.
1008 P. Muller, C. Aichinger, M Feldbrugge and R. Kahmann

et al., 1996). Thus, the elimination of the putative MAPkinase sites did not destroy the activity of prf1 completely,but affected its function during mating. This provides evi-dence that a MAP kinase is involved in the transmissionof the pheromone signal to Prf1.
Identification of a MAP kinase gene
Using the degenerate primers MAP3 and MAP5, designedaccording to known fungal MAPK genes FUS3, SPK1 andpmk1 (Elion et al., 1990; Toda et al., 1991; Xu and Hamer,1996), and genomic DNA from U. maydis strain SG200D
fuz7 as a template, we were able to amplify a single frag-ment of 490 bp, which, upon sequencing, displayed sig-nificant similarity to known MAPK genes (Fig. 2). Usingthis fragment as a probe, a genomic clone was isolatedfrom a lEMBL3 library (Schulz et al., 1990), and after sub-cloning hybridizing fragments, a plasmid clone, pKpp2H,containing a 4.5 kb HindIII fragment was derived. Sub-sequently, several incomplete cDNA clones were isolatedfrom a cDNA library in lgt10 (Schauwecker et al., 1995).Sequence analysis revealed an open reading frame
(ORF) for a polypeptide of 345 amino acids that was notinterrupted by introns (Fig. 2). Comparisons with database entries demonstrated striking similarities to otherfungal MAP kinases, such as Pmk1 of Magnaporthegrisea (75% identity), CEK1 of Candida albicans (67%identity) or Fus3p from Saccharomyces cerevisiae(58% identity, Fig. 2). We designate the correspondingU. maydis gene kpp2.
Phenotype of Dkpp2-1 strains
To analyse the function of the kpp2 gene in U. maydis,mutants were generated in which the region encompass-ing the first 161 amino acids of the kpp2 ORF weredeleted. To this end, plasmid pDkpp2-1 (see Experimentalprocedures) was introduced, using transformation, intohaploid strains FB1 (a1b1) or FB2 (a2b2), as well as intothe solopathogenic haploid strains CL13 (a1bE1bW2) orSG200 (a1::mfa2bE1bW2). Nourseothricin-resistant trans-formants, in which the resident kpp2 gene was replacedby the Dkpp2-1 allele, were identified using Southern analy-sis (not shown). Replacement mutants could be generated
Q 1999 Blackwell Science Ltd, Molecular Microbiology, 34, 1007–1017
Fig. 2. Alignment showing homology of Kpp2 to other MAP kinases from fungi and yeasts. Identical amino acids are shaded. The primerpositions are indicated by horizontal arrows, asterisks denote the invariant TXY motif of MAP kinases, which has been shown to bephosphorylated upon stimulation through its MAPK kinase (Errede et al., 1993; Gartner et al., 1992). The alignment was performed usingCLUSTAL W1.7 (Thompson et al., 1994). GenBank accession numbers for the proteins displayed are as follows: Kpp2 from U. maydis(AF193614), Pmk1 from Magnaporthe grisea (U70134), CEK1 from Candida albicans (P28869), SPK1 from Schizosaccharomyces pombe(P27638) and Fus3p and Kss1p from S. cerevisiae (P16892) and (P14681) respectively.
MAP kinase Kpp2 in U. maydis 1009
with similar efficiency (about 20%) in all strains, andthese mutants had no obvious phenotypic alterationsupon growth in PD medium, indicating that kpp2 is anon-essential gene.
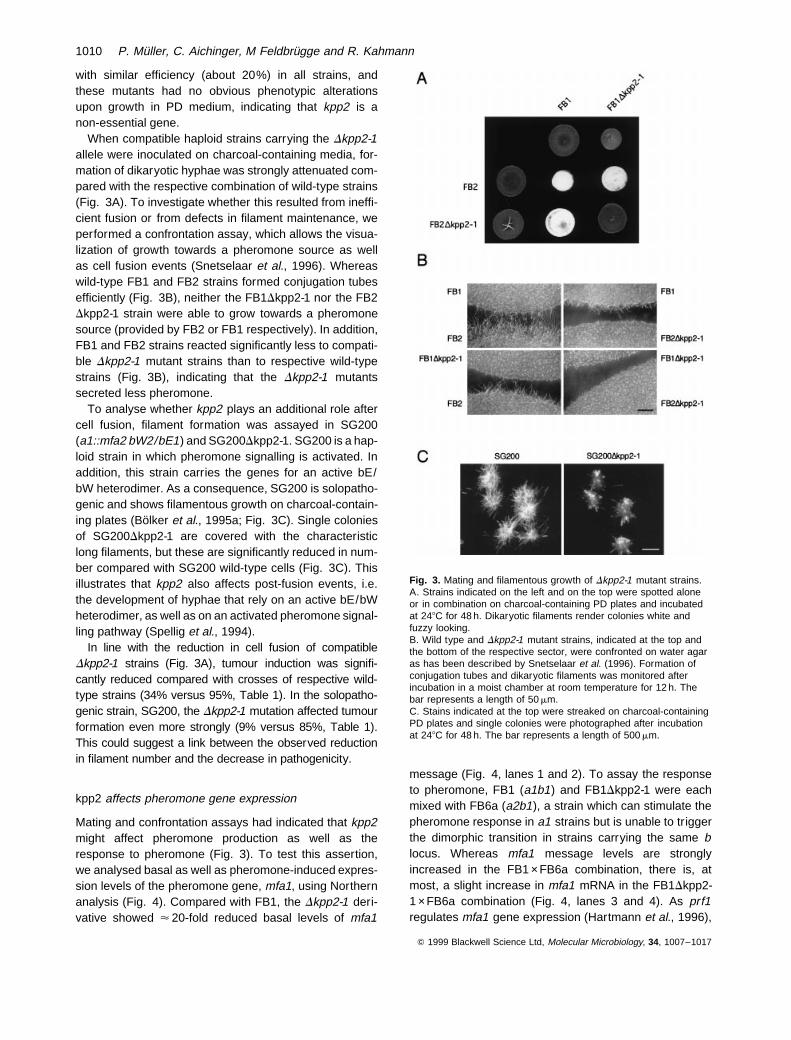
When compatible haploid strains carrying the Dkpp2-1allele were inoculated on charcoal-containing media, for-mation of dikaryotic hyphae was strongly attenuated com-pared with the respective combination of wild-type strains(Fig. 3A). To investigate whether this resulted from ineffi-cient fusion or from defects in filament maintenance, weperformed a confrontation assay, which allows the visua-lization of growth towards a pheromone source as wellas cell fusion events (Snetselaar et al., 1996). Whereaswild-type FB1 and FB2 strains formed conjugation tubesefficiently (Fig. 3B), neither the FB1Dkpp2-1 nor the FB2Dkpp2-1 strain were able to grow towards a pheromonesource (provided by FB2 or FB1 respectively). In addition,FB1 and FB2 strains reacted significantly less to compati-ble Dkpp2-1 mutant strains than to respective wild-typestrains (Fig. 3B), indicating that the Dkpp2-1 mutantssecreted less pheromone.
To analyse whether kpp2 plays an additional role aftercell fusion, filament formation was assayed in SG200(a1::mfa2 bW2/bE1) and SG200Dkpp2-1. SG200 is a hap-loid strain in which pheromone signalling is activated. Inaddition, this strain carries the genes for an active bE/bW heterodimer. As a consequence, SG200 is solopatho-genic and shows filamentous growth on charcoal-contain-ing plates (Bolker et al., 1995a; Fig. 3C). Single coloniesof SG200Dkpp2-1 are covered with the characteristiclong filaments, but these are significantly reduced in num-ber compared with SG200 wild-type cells (Fig. 3C). Thisillustrates that kpp2 also affects post-fusion events, i.e.the development of hyphae that rely on an active bE/bWheterodimer, as well as on an activated pheromone signal-ling pathway (Spellig et al., 1994).
In line with the reduction in cell fusion of compatibleDkpp2-1 strains (Fig. 3A), tumour induction was signifi-cantly reduced compared with crosses of respective wild-type strains (34% versus 95%, Table 1). In the solopatho-genic strain, SG200, the Dkpp2-1 mutation affected tumourformation even more strongly (9% versus 85%, Table 1).This could suggest a link between the observed reductionin filament number and the decrease in pathogenicity.
kpp2 affects pheromone gene expression
Mating and confrontation assays had indicated that kpp2might affect pheromone production as well as theresponse to pheromone (Fig. 3). To test this assertion,we analysed basal as well as pheromone-induced expres-sion levels of the pheromone gene, mfa1, using Northernanalysis (Fig. 4). Compared with FB1, the Dkpp2-1 deri-vative showed < 20-fold reduced basal levels of mfa1
message (Fig. 4, lanes 1 and 2). To assay the responseto pheromone, FB1 (a1b1) and FB1Dkpp2-1 were eachmixed with FB6a (a2b1), a strain which can stimulate thepheromone response in a1 strains but is unable to triggerthe dimorphic transition in strains carrying the same blocus. Whereas mfa1 message levels are stronglyincreased in the FB1 ×FB6a combination, there is, atmost, a slight increase in mfa1 mRNA in the FB1Dkpp2-1 ×FB6a combination (Fig. 4, lanes 3 and 4). As prf1regulates mfa1 gene expression (Hartmann et al., 1996),
Q 1999 Blackwell Science Ltd, Molecular Microbiology, 34, 1007–1017
Fig. 3. Mating and filamentous growth of Dkpp2-1 mutant strains.A. Strains indicated on the left and on the top were spotted aloneor in combination on charcoal-containing PD plates and incubatedat 248C for 48 h. Dikaryotic filaments render colonies white andfuzzy looking.B. Wild type and Dkpp2-1 mutant strains, indicated at the top andthe bottom of the respective sector, were confronted on water agaras has been described by Snetselaar et al. (1996). Formation ofconjugation tubes and dikaryotic filaments was monitored afterincubation in a moist chamber at room temperature for 12 h. Thebar represents a length of 50 mm.C. Stains indicated at the top were streaked on charcoal-containingPD plates and single colonies were photographed after incubationat 248C for 48 h. The bar represents a length of 500 mm.
1010 P. Muller, C. Aichinger, M Feldbrugge and R. Kahmann

we have also analysed the effect of the Dkpp2-1 mutationon expression of the prf1 gene. Similar to mfa1 geneexpression, we observed a 15-fold reduction in prf1mRNA levels in Dkpp2-1 strains. After pheromone induc-tion, we detected an induction of prf1 message (Fig. 4,lane 4). It is most likely that this increase in prf1 mRNAoccurs in the pheromone donor strain, FB6a, which canbe stimulated by the residual amounts of pheromone pro-duced by the Dkpp2-1 mutant strain. The absence ofpheromone gene induction (which can be specificallyassayed in the Dkpp2-1 strain without interference fromthe pheromone donor) suggests that the MAP kinaseKpp2 participates in pheromone signal transduction.
kpp2 and cAMP signalling
Pheromone gene transcription is known to be influencedby pheromone signalling as well as by the cAMP pathway
(Kruger et al., 1998; Hartmann et al., 1999). Therefore, wedecided to investigate the reaction of Dkpp2-1 strains toexternally added cAMP. In the presence of 6 and 15 mMcAMP, the FB1Dkpp2 strain behaved like the respectivewild-type strain, and displayed the characteristic multiplebudding phenotype at the higher concentration (Gold etal., 1994; Kruger et al., 1998; not shown). This result indi-cates that transmission of the external cAMP signal isunaffected by the Dkpp2-1 mutation.
To address the epistatic relationship between cAMP sig-nalling and kpp2, we generated a FB1Dkpp2-1Dubc1 dou-ble mutant strain (see Experimental procedures). Dubc1strains display a multiple budding phenotype (Gold et al.,1994), and the FB1Dkpp2-1Dubc1 double mutant strainwas unaltered with respect to this trait (not shown). Com-pared with FB1, basal prf1 and mfa1 expression levelswere dramatically increased in the FB1Dubc1 strain thathas been described before (Hartmann et al., 1999).Although transcripts of these two genes were barelydetectable in Dkpp2-1 strains, we observed a significantinduction of both transcripts in the double mutant, althoughthe levels were significantly below what is seen in theDubc1 single mutant strain (Fig. 5). We conclude thatthe MAP kinase Kpp2 is not a component of the cAMP cas-cade. The differences in mfa1 and prf1 transcript levels inthe Dubc1 and Dubc1Dkpp2-1 double mutant strains canbe explained by assuming that the cAMP cascade andthe pheromone signalling cascade act in concert on Prf1.
Constitutive expression of prf1 does not restorepathogenicity in Dkpp2-1 strains
With respect to pathogenic development, prf1 plays a cru-cial role in activating transcription of the b genes. In parti-cular, it has been shown that in the solopathogenic haploidstrain, CL13 (a1bE1bW2), the deletion of prf1 causes lossof pathogenicity. This defect can be overcome by expres-sing bE1 and bW2 from constitutive promoters (Hartmannet al., 1996). In view of this data, the reduction in tumourformation observed in Dkpp2-1 strains could simply reflectinsufficient prf1 gene expression. Although tumour forma-tion in SG200Dkpp2-1 was reduced to 9%, we observedeven fewer tumours develop in CL13Dkpp2-1 infections(CL13 differs from SG200 only in lacking the activatedpheromone pathway; Bolker et al., 1995a). In addition,these tumours were clearly distinguishable from thoseinduced by wild-type strains with respect to their sizeand their location (Table 1). We have consequently triedto restore pathogenicity in CL13Dkpp2-1 by expressingeither the prf1 gene (prf1con) or the bE1 and bW2 genesconstitutively (bcon). The CL13prf1conDkpp2-1 strain showedhigh levels of prf1 gene transcript (not shown), however,pathogenicity was not rescued (Table 1). On the otherhand, transformants expressing the bcon allele were
Q 1999 Blackwell Science Ltd, Molecular Microbiology, 34, 1007–1017
Table 1. Tumour formation induced by Dkpp2-1 strains.
Inoculum Infected plants Tumourformation (%)
FB1 (a1b1) ×FB2 (a2b2) 114 95FB1Dkpp2-1 ×FB2Dkpp2-1 106 34SG200 (a1::mfa2 bE1 bW2) 99 85SG200Dkpp2-1 96 9a
CL13 (a1bE1bW2) 20 70CL13Dkpp2-1 107 1.8a
CL13 prf1con 20 80CL13 prf1con Dkpp2-1 23 4a
HA103 (a1bcon) 40 95HA103Dkpp2-1 40 52
a. Tumour diameter did not exceed 1 mm and tumour developmentwas observed only on leaves
Fig. 4. Pheromone-induced gene expression in Dkpp2-1 strains.Strains and strain combinations, indicated at the top, were grownon charcoal-containing CM plates for 48 h at 248C. RNA wasisolated and subjected to Northern analysis, after loading 15 mgtotal RNA per lane. The same filter was hybridized in successionwith probes for mfa1, prf1 and cbx as a loading control.
MAP kinase Kpp2 in U. maydis 1011

significantly improved in tumour formation with respectto the CL13Dkpp2-1 progenitor strain (from 4% to 52%,Table 1). These results illustrate that reduced tumour for-mation in solopathogenic haploid Dkpp2-1 strains can bepartially overcome by increasing the amounts of bE1/bW2 heterodimer. For the wild-type situation, these resultsimply that kpp2 is needed for the pheromone response aswell as for pathogenic development.
Discussion
In this communication we describe the isolation and char-acterization of the first MAP kinase gene of the basidiomy-cete U. maydis. kpp2 is needed for recognition betweenmating partners and has an additional function duringpathogenic development.
In several systems, MAP kinases are directly responsi-ble for phosphorylation of target transcription factors,which then activate specific genes (Treisman, 1996).The best-studied example with respect to the pheromoneresponse pathway is S. cerevisiae. In particular, all com-ponents of the MAP kinase module and the transcriptionfactor responsible for pheromone-dependent geneexpression have been characterized in detail (Herskowitz,1995). The respective MAP kinase, Fus3p, has beenshown to phosphorylate the Ste12p homeodomain tran-scription factor in vitro (Elion et al., 1993). However, it isstill a matter of debate whether Fus3p-mediated phosphor-ylation of Ste12p accounts for the pheromone-dependentgene activation in vivo (Song et al., 1991; Hung et al.,1997; Pi et al., 1997).
In U. maydis, the HMG box transcription factor, Prf1, isresponsible for pheromone-induced gene expression(Hartmann et al., 1996). The activity of prf1 is regulatedby pheromone on the transcriptional as well as the post-transcriptional level (Hartmann et al., 1999). In thisstudy, we describe the analysis of a prf1 allele, containingmutations in six putative MAP kinase sites as well as in apresumed MAP kinase docking site. Strains harbouringthis allele still produce pheromone but exhibit a reducedmating competence, indicating that mutant Prf1 proteinhas retained its ability to drive pheromone gene expres-sion. However, the full mating response after pheromonestimulation is lost, arguing that protein phosphorylationby a MAP kinase may regulate the activity of Prf1.
A putative candidate for this MAP kinase is Kpp2. Fromits deduced amino acid sequence, Kpp2 is most closelyrelated to other fungal MAP kinases like Pmk1 of M. gri-sea, CEK1 of C. albicans, SPK1 of Schizosaccharomycespombe and Kss1p and Fus3p of S. cerevisiae. Common toall these MAP kinases is the characteristic TEY motif in theactivation loop of subdomain VIII (Hunter and Plowman,1997). SPK1 and Fus3p are components of the phero-mone signalling pathway in their respective hosts (Hersko-witz, 1995). The pmk1 gene is needed for appressoriumformation in M. grisea (Xu and Hamer, 1996), and canpartially complement the mating defect of a S. cerevisiaefus3–kss1 double mutant (Xu and Hamer, 1996). TheKSS1 gene in S. cerevisiae, as well as its functional homo-logue CEK1 in C. albicans, regulates morphogenesis(Whiteway et al., 1992; Madhani and Fink, 1998; Brownand Gow, 1999).
The hypothesis that kpp2 is the MAP kinase transmittingthe pheromone signal is supported by characteristic phe-notypes of kpp2 deletion strains. Dkpp2-1 mutants werereduced in cell fusion but were not completely sterile, aphenotype that resembles the situation in strains harbour-ing the prf1-M1 allele. The residual fusion competence ofkpp2 mutant strains might also reflect the recruitment ofanother, as yet unknown, MAP kinase to the signallingmodule. Such a scenario has been uncovered in S. cere-visiae (Madhani et al., 1997). The most prominent pheno-type of kpp2 mutants is that they produce less pheromoneand cannot react to pheromone stimulation. Consequently,such strains are unable to initiate the primary morphologi-cal response to pheromone, which is the formation of con-jugation tubes. A comparable defect in conjugation tubeformation has also been observed in mutants lacking theMAPK kinase Fuz7 (Banuett and Herskowitz, 1994). fuz7mutants have a severe mating defect, cannot maintain fila-ments, induce tumours less efficiently and produce telios-pores that are reduced in germination. Based on thismultitude of phenotypes, fuz7 was initially placed in thepheromone cascade as well as in another pathway respond-ing to a putative plant signal (Banuett and Herskowitz,
Q 1999 Blackwell Science Ltd, Molecular Microbiology, 34, 1007–1017
Fig. 5. Epistatic relationship of kpp2 and the cAMP signallingpathway. Strains indicated at the top were grown on charcoal-containing CM plates for 48 h at 248C. RNA was isolated andsubjected to Northern analysis, after loading 15 mg total RNA perlane. The same filter was hybridized in succession with probes formfa1, prf1 and cbx as a loading control.
1012 P. Muller, C. Aichinger, M Feldbrugge and R. Kahmann

1994). However, in contrast to kpp2 mutants, pheromone-induced gene expression in fuz7 mutants was similar tothat in the wild type (Regenfelder et al., 1997). Therefore,it appears unlikely that the MAPK kinase, Fuz7, acts in themodule containing Kpp2 (Fig. 6). However, we cannotexclude the possibility that Kpp2 and Fuz7 interact duringsubsequent stages of development.
Because cAMP signalling in U. maydis is intimatelylinked with the pheromone cascade (Kruger et al., 1998;Hartmann et al., 1999), the mating defects seen in kpp2mutant strains could be indirect and result from perturba-tions in cAMP signalling. In the following, we shall argueagainst this hypothesis. Lowering the internal cAMPlevel usually results in filamentation (Gold et al., 1994),which we do not observe in Dkpp2-1 strains. In addition,Dkpp2-1 strains respond to high external cAMP levelswith multiple budding, and thus show the same responseas wild-type strains (Gold et al., 1994), indicating thatkpp2 is not an effector for this morphological transition.Furthermore, the Dubc1-Dkpp2-1 double mutant straindisplays elevated levels of mfa1 transcript comparedwith the Dkpp2-1 strain, excluding the possibility thatkpp2 provides a link between cAMP signalling and prf1.Taken together, these independent observations suggestthat kpp2 is not an integral component of the known cAMPcascade (Fig. 6).
An unexpected phenotype of kpp2 mutant strains was asignificant reduction in the ability to induce plant tumours,even under conditions in which cell fusion was notrequired. In such haploid solopathogenic strains, like
CL13, it has been demonstrated that an activated phero-mone pathway is not a prerequisite for pathogenicity(Regenfelder et al., 1997). However, one component ofthe pheromone signalling cascade, prf1, also serves anessential function during pathogenesis because it is theactivator of b gene transcription. In the absence of phero-mone, Prf1 appears to be activated through differentsignals and guarantees sufficient b gene expression (Hart-mann et al., 1996). Interestingly, in CL13Dkpp2-1, constitu-tive expression of prf1 did not positively affect pathogenicdevelopment. Constitutive transcription is likely to increasethe amount of Prf1 protein, but this protein would not beactivated by Kpp2 (Fig. 6). The overexpression of bE1/bW2, on the other hand, led to a significant increase intumour formation. In this situation, the need for activatedPrf1 protein is bypassed and pathogenicity is increased.For the wild-type situation, this probably reflects theMAP kinase Kpp2 transmitting other signals, which alsoresult in the activation of the b genes through Prf1, inthe absence of pheromone stimulation. The notable differ-ences in pathogenicity observed between SG200Dkpp2-1and CL13Dkpp2-1, two strains that differ only with respectto an activated pheromone signalling cascade, mightreflect a higher residual activity of the pheromone pathwayin the SG200 background. In addition, we assume thatKpp2 has several targets, besides Prf1, which need tobe phosphorylated for full pathogenic development(Fig. 6). This may explain why pathogenicity is not res-cued completely by overexpression of the active bE/bWheterodimer in a Dkpp2-1 strain. As it is known that aregulated cAMP signalling pathway is needed for patho-genicity (Gold et al., 1997; Durrenberger et al., 1998),one or more of these targets may coincide with targetsfor Adr1.
In the rice pathogen M. grisea, cAMP signalling hasalready been linked to very discrete steps of developmentduring infection. For example, an increase in intracellularcAMP levels promotes early steps of appressorium devel-opment (Mitchell and Dean, 1995; Xu and Hamer, 1996;Xu et al., 1997). Furthermore, pmk1 mutants of M. grisea,which lack a MAP kinase with strong similarity to Kpp2(Fig. 2), are defective in both appressorium formationand invasive growth, although they have retained theirability to respond to extracellular cAMP (Xu and Hamer,1996). Pmk1 was suggested to either act downstream ofa cAMP-dependent signal or function in parallel duringappressorium formation (Xu and Hamer, 1996). Thus, acomplex interplay between cAMP and MAP kinase signal-ling also emerges in this system as a prerequisite for a suc-cessful infection. It will be an interesting and challengingtask to identify separate, as well as shared targets, forboth MAP kinases and cAMP signalling, and to determinewhether these may be conserved between Ascomyceteand Basidiomycete fungi.
Q 1999 Blackwell Science Ltd, Molecular Microbiology, 34, 1007–1017
Fig. 6. Proposed signalling processes during mating andpathogenic development in U. maydis. Different signalling inputs(top) are transmitted by three different cascades, eliciting variouscellular processes (bottom). Broken arrows indicate missingcomponents or putative signalling events. See text for detaileddiscussion.
MAP kinase Kpp2 in U. maydis 1013

Experimental procedures
Isolation of the kpp2 gene
Degenerate primers MAP3 (GCITAYGGIRTNGTNTG) andMAP5 (AYYTCIGGIGCICKRTAVYA) were used for theamplification of U. maydis DNA from strain SG200Dfuz7 (J.Kruger, unpublished). Reactions contained 10 mM Tris-HCl,pH 8.3, 3 mM MgCl2, 50 mM KCl, 20 pmol primers and 2 UTaq polymerase (Boehringer Mannheim). Amplification wasachieved in 35 cycles of 1 min at 958C, 1 min at 528C and1 min at 728C. For sequencing, polymerase chain reaction(PCR) products of 490 bp were cloned, after blunting withPfu polymerase (Stratagene) and phosphorylation with poly-nucleotide kinase (Boehringer Mannheim), into the HincIIsite of pTZ19R. The amplified kpp2 fragment was used toscreen a genomic lEMBL3 library (Schulz et al., 1990).From a hybridizing clone, kpp2 was subcloned as a 4.5 kbHindIII fragment in pTZ19R, and the resulting plasmid wasdesignated pkpp2H.
Plasmids and plasmid constructions
Plasmid pTZ19R (Pharmacia) was used for cloning, subclon-ing and sequencing. pNEBHyg(¹) and pNEBNat(þ) arederived from pNEB193 (New England Biolabs) and containa hygromycin and a nourseothricin resistance cassetterespectively (A. Brachmann and R. Kahmann, unpublished).In the nourseothricin resistance cassette, the nat1 ORF ofStreptomyces noursei is expressed from the gap promoterfrom U. maydis (Smith and Leong, 1990; Krugel et al.,1993; G. Bakkeren, unpublished). The same resistance cas-settes are present in pSLNat(þ) and pSLHyg(þ), which differfrom pNEBNat(þ) and pNEBHyg(¹) in their multiple cloningsites (A. Brachmann and R. Kahmann, unpublished). pNE-BUH is an autonomously replicating plasmid for U. maydis,containing the UARS and a cassette conferring hygromycinresistance (G. Weinzierl and R. Kahmann, unpublished).
pRFcon is a plasmid carrying the prf1 gene under the con-trol of the constitutive tef1 promoter, 58 and 38 flanking sequ-ences for homologous recombination and a carboxinresistance cassette inserted upstream of the tef1 promoter(Hartmann et al., 1999).
In the deletion construct pDprf1 a 2487 bp Sal I–SphI frag-ment, extending from position þ 66 to þ 2553 bp of the prf1gene, was replaced by a 1618 bp fragment with XhoI andSphI ends, containing the nourseothricin resistance cassettederived from pSLNat(þ). pprf1-E was constructed in severalsteps. First, NcoI and XhoI sites were generated, using stan-dard PCR techniques, at the start and stop codon of the prf1gene respectively. Next, the NcoI site was used to add ahexa-histidine tag derived from plasmid pET-15b (Novagen)and the XhoI site served to insert three copies of the mycepitope tag GEQKLISEEDLN from plasmid pUC19tm (Hart-mann, 1997). Finally, a 2927 bp ClaI–Nsi I fragment frompSLHyg(þ), containing a hygromycin resistance cassette,was inserted in the 38 untranslated region of the prf1 gene.The prf1-M1 allele was derived from prf1-E by introducingpoint mutations, using the megaprimer PCR technique(Sarkar and Sommer, 1990). The following oligonucleotidemegaprimers were used to substitute the indicated aminoacid(s) by alanine: S267, CCGGGCCCGCTTTGCACGTGG
AGCTGACGGCTTGCC; S375, GTACCGGTCATGCACCCTTGCGCGCCAACGCTAGCCT; S448, GCTCGTACGACGACCAGTCACTACCTGCGCCGCAATCCTTTG; T473, GCTCGAGCTTGGCGCCGAAGGCCATGC; S498, CTCGATGCTCATCGCGCCCATGACAGCGTC; S812, CGGCAAACCGCTTCCTGCGCCGTTGCAGCTCGTC; T586, F589, F590 andD591, TCCGCCCTCCAGAGCCGGTGCAGCGGCCGCATTGGGAGCTTCATCATAGTCAGC.
The deletion construct pDkpp2-1 was derived from pkpp2Hby replacing a 620 bp MluI–Bgl II fragment, encompassingthe region between ¹138 to þ 482 of the kpp2 ORF, withthe nourseothricin resistance cassette isolated as a Bgl II–Asc I fragment from pNEBNat(þ). The plasmid pubc1-1 wasderived from pubc1 (Hartmann et al., 1999) by deleting the1.5 kb BssHII fragment, encompassing the ORF of ubc1from amino acid 20 to 517, and replacing it with the hygromy-cin resistance cassette isolated as a 2.9 kb AscI fragmentfrom pNEBHyg(¹). For the construction of pNEBUHkpp2, a2.5 kb Pst I–HindIII fragment of pkpp2H was cloned into pNE-BUH cut with HindIII and Sbf I.
Strains and strain constructions
For cloning purposes, the E. coli K12 derivative DH5a wasused.
Haploid Ustilago maydis strains FB1 (a1b1), FB2 (a2b2)and FB6a (a2b1), have been described previously (Banuettand Herskowitz, 1989). The haploid solopathogenic strainsCL13 (a1 bE1bW2) and the derivative SG200 (a1::mfa2bE1bW2) differ with respect to the mfa2 gene, which wasintroduced in the resident a1 locus of CL13 and which pro-vides for autocrine pheromone stimulation resulting in fila-mentous growth (Bolker et al., 1995a; 1995b). Strain HA103(a1 bcon) is a CL13 derivative in which transcription of thebE1 and bW2 genes is driven by constitutive promoters.The strain was constructed by homologous recombinationbetween pbcon and the resident b locus (Hartmann et al.,1996; Hartmann, 1997). FB1Dubc1 (a1b2 ubc1::nat ) is aFB1 derivative, containing an internal 1.5 kb deletion withinthe ubc1 gene (corresponding to amino acids 20–517),which was substituted by a nourseothricin resistance cassette(Hartmann et al., 1999).
The strains FB1Dprf1 and FB2Dprf1 were generated bytransformation of the respective wild-type strains with plasmidpDprf1 that had been linearized with DraI. Nourseothricinresistant transformants were subjected to Southern analysisin order to verify homologous recombination. prf1-E and prf1-M1 strains were derived from FB1Dprf1 and FB2Dprf1 strains,using plasmids pprf1-E and pprf1-M1. Hygromycin-resistanttransformants were first screened for loss of nourseothricinresistance and then subjected to Southern analysis.
All Dkpp2-1 deletion strains were constructed by transfor-mation of the respective progenitor strains with pDkpp2-1 lin-earized with BamHI. Homologous recombination wasverified using Southern analysis, after cleaving the DNAwith Sal I. The 2.8 kb HindIII–Sal I fragment of pkpp2H wasused as a probe. To verify that it was only kpp2 that wasresponsible for the observed, FB1Dkpp2-1 and FB2Dkpp2-1 were transformed with pNEBUHkpp2, an autonomouslyreplicating plasmid containing the kpp2 gene, including 1 kb58 as well as 0.5 kb 38 sequences. Introduction of this plasmid
Q 1999 Blackwell Science Ltd, Molecular Microbiology, 34, 1007–1017
1014 P. Muller, C. Aichinger, M Feldbrugge and R. Kahmann

completely restored the mating defect of respective strains(not shown).
CL13prf1con and CL13prf1conDkpp2-1 strains were con-structed by transformation of the respective progenitor strainswith the plasmid pRFcon cut with DraI. Homologous recombi-nation was detected using Southern analysis after cleavingthe DNA with Pst I using a genomic 3.2 kb HindIII–BamHIfragment, encompassing parts of the prf1 gene, as a probe.
To generate the double mutant FB1Dkpp2-1-Dubc1, theFB1Dkpp2-1 strain was transformed with plasmid pubc1-1 lin-earized with BamHI. DNA from individual transformants wascleaved with EcoRI and homologous recombination wasdetected using Southern analysis, employing the 3.6 kbEcoRI fragment of pubc1 as a probe.
Growth conditions for U. maydis
Strains were grown at 288C in YEPS (modified after Tsukadaet al., 1988), potato dextrose (PD) or CM medium (Holliday,1974). cAMP (Sigma) was added directly to the medium,which was filter sterilized before use. Hygromycin B was pur-chased from Boehringer Mannheim, nourseothricin from theHans-Knoll-Institute and carboxin from Riedel de Haen. Allchemicals used were of analytical grade and were obtainedfrom Sigma or Merck.
The mating reaction was observed by co-spotting strains oncharcoal-containing potato dextrose plates, after incubation at248C for 48 h. Confrontation assays were performed as hasbeen described (Snetselaar et al., 1996), except that thestrains were resuspended in H2O to an OD600 of 5.0 prior tospotting. The formation of mating structures was observedafter 12 h of incubation at room temperature.
Pathogenicity assay
Plant infections of the variety Early Golden Bantam (OldsSeeds) were performed essentially as has been describedpreviously (Gillissen et al., 1992), with the following modifica-tions. Strains were grown overnight in YEPS liquid medium toan OD600 of 0.8, harvested and resuspended in water to anOD600 of 2.4 prior to infection. For each strain tested, atleast two independent pathogenicity assays were performed.Initial disease symptoms were observed one week postinoculation and tumour formation was scored after 4 weeks.Tumours induced by wild-type strains ranged in size from0.5–5 cm in diameter, and developed at stems and leaves.Significantly smaller tumours (<1 mm in diameter) wereobserved in solopathogenic strains carrying the Dkpp2-1allele (see Table 1).
DNA and RNA procedures
Standard molecular techniques followed Sambrook et al.(1989). Transformation of Ustilago maydis was performedas published by Schulz et al. (1990). U. maydis DNA was iso-lated as described by Hoffmann and Winston (1987). RNAwas isolated from strains grown on charcoal-containing CMplates as has been described previously (Schauwecker etal., 1995). To assay the effects of cAMP on gene expressionand morphology, strains were grown in PD liquid medium and
RNA was prepared as described by Kruger et al. (1998). Thefollowing probes were used for Northern analysis: a 675 bpEcoRV fragment spanning the mfa1 gene isolated frompUMa1 (Urban et al., 1996), a 1.5 kb Sal I–Bgl II fragmentcontaining prf1 derived from pHA89 (Hartmann et al., 1996)and a 2.2 kb EcoRI–EcoRV fragment of pCBX122 (Koen etal., 1991) that detects the iron–sulphur succinate dehydro-genase message (cbx) as loading control. Radioactive label-ling of the DNA was performed with the megaprime DNAlabelling kit (Amersham). A Storm phosphorimager (Molecu-lar Dynamics) was used for signal detection. The intensity ofthe signals was calculated and compared using IMAGEQUANT
software (Molecular Dynamics).The sequence of kpp2 was derived from pkpp2H, using an
ABI 373 automated sequencer. Sequence analysis was per-formed with standard bioinformatic tools.
Acknowledgements
We thank Dr S. Gold for exchanging data prior to publicationand Dr G. Bakkeren (University of British Columbia) for theNAT1 resistance cassette. The invaluable technical assist-ance of K.-H. Braun in performing plant infections is gratefullyacknowledged. We thank J. Kruger and A. Brachmann fortheir critical reading of the manuscript and Dr C. Koch forscientific advice. This work was supported by the Leibniz pro-gramme of the DFG, SFB369 and the EU project CEREPAT.
References
Banuett, F. (1995). Genetics of Ustilago maydis, a fungalpathogen that induces tumors in maize. Annu Rev Genet29: 179–208.
Banuett, F., and Herskowitz, I. (1989). Different a alleles ofUstilago maydis are necessary for maintenance of fila-mentous growth but not for meiosis. Proc Natl Acad SciUSA 86: 5878–5882.
Banuett, F., and Herskowitz, I. (1994). Identification of fuz7, aUstilago maydis MEK /MAPKK homolog required for a-locus-dependent and -independent steps in the fungal lifecycle. Genes Dev 8: 1367–1378.
Bolker, M., Urban, M., and Kahmann, R. (1992). The a mat-ing type locus of U. maydis specifies cell signalling compo-nents. Cell 68: 441–450.
Bolker, M., Genin, S., Lehmler, C., and Kahmann, R.(1995a). Genetic regulation of mating and dimorphism inUstilago maydis. Can J Bot 73: 320–325.
Bolker, M., Bohnert, H.U., Braun, K.H., Gorl, J., and Kah-mann, R. (1995b). Tagging pathogenicity genes in Ustilagomaydis by restriction enzyme-mediated integration(REMI). Mol Gen Genet 248: 547–552.
Brown, A.J.P., and Gow, N.A.R. (1999). Regulatory networkscontrolling Candida albicans morphogenesis. TrendsMicrobiol 7: 333–338.
Christensen, J.J. (1963). Corn smut caused by Ustilago may-dis. Am Phytopathol Soc Monogr no. 2.
Clark-Lewis, I., Sanghera, J.S., and Pelech, S.L. (1991). Defi-nition of a consensus sequence for peptide substraterecognition by p44mpk, the meiosis-activated myelinbasic protein kinase. J Biol Chem 266: 15180–15184.
Q 1999 Blackwell Science Ltd, Molecular Microbiology, 34, 1007–1017
MAP kinase Kpp2 in U. maydis 1015

Durrenberger, F., Wong, K., and Kronstad, J.W. (1998).Identification of a cAMP-dependent protein kinase cataly-tic subunit required for virulence and morphogenesis inUstilago maydis. Proc Natl Acad Sci USA 95: 5684–5689.
Elion, E.A., Grisafi, P.L., and Fink, G.R. (1990). FUS3encodes a cdc2þ /CDC28-related kinase required for thetransition from mitosis into conjugation. Cell 60: 649–664.
Elion, E.A., Satterberg, B., and Kranz, J.E. (1993). FUS3phosphorylates multiple components of the mating signaltransduction cascade: evidence for STE12 and FAR1.Mol Biol Cell 4: 495–510.
Errede, B., Gartner, Z., Zhou, Z., Nasmyth, K., and Ammerer,G. (1993). MAP kinase related FUS3 from S. cerevisiae isactivated by STE7 in vitro. Nature 362: 261–264.
Gartner, A., Nasmyth, K., and Ammerer, G. (1992). Signaltransduction in Saccharomyces cerevisiae requires tyro-sine and threonine phosphorylation of FUS3 and KSS1.Genes Dev 6: 1280–1292.
Gillissen, B., Bergemann, J., Sandmann, C., Schroeer, B.,Bolker, M., and Kahmann, R. (1992). A two-componentregulatory system for self/non-self recognition in Ustilagomaydis. Cell 68: 647–657.
Gold, S.E., Duncan, G., Barrett, K., and Kronstad, J. (1994).cAMP regulates morphogenesis in the fungal pathogenUstilago maydis. Genes Dev 8: 2805–2816.
Gold, S.E., Brogdon, S.M., Mayorga, M.E., and Kronstad,J.W. (1997). The Ustilago maydis regulatory subunit ofcAMP dependent protein kinase is required for gall forma-tion in maize. Plant Cell 9: 1585–1594.
Hartmann, A. (1997). Die Pheromonantwort in U MaydisKontrolle von Zellfusion und Pathogenitat. PhD thesis.Ludwig-Maximilians-Universitat Munchen.
Hartmann, A., Kahmann, R., and Bolker, M. (1996). The phero-mone response factor coordinates filamentous growth andpathogenicity in Ustilago maydis. EMBO J 15: 1632–1641.
Hartmann, A., Kruger, J., Lottspeich, F., and Kahmann, R.(1999). Environmental signals controlling sexual develop-ment of the corn smut fungus Ustilago maydis throughthe transcriptional regulator Prf1. Plant Cell 11: 1293–1305.
Herskowitz, I. (1995). MAP kinase pathways in yeast: formating and more. Cell 80: 187–197.
Hoffmann, C., and Winston, F. (1987). A ten-minute DNApreparation from yeast efficiently releases autonomousplasmids for transformation of Escherichia coli. Gene 57:267–272.
Holliday, R. (1974). Ustilago maydis. In Handbook of Genet-ics, Vol 1. King, R.C. (ed.). New York, NY: Plenum Press,pp. 575–595.
Hung, W., Olson, K.A., Breitkreutz, A., and Sadowski, I.(1997). Characterization of the basal and pheromone-stimulated phosphorylation states of Ste12p. Eur J Bio-chem 245: 241–251.
Hunter, T., and Plowman, G.D. (1997). The protein kinases ofbudding yeast: six score and more. Trends Genet 22: 18–22.
Jacobs, D., Glossip, D., Xing, H., Muslin, A.J., and Kornfeld,K. (1999). Multiple docking sites on substrate proteins forma modular system that mediates recognition by ERK MAPkinase. Genes Dev 13: 163–175.
Kamper, J., Reichmann, M., Romeis, T., Bolker, M., and Kah-mann, R. (1995). Multiallelic recognition: nonself-depen-dent dimerization of the bE and bW homeodomainproteins in Ustilago maydis. Cell 81: 73–83.
Koen, J.P.R., White, G.A., and Hargreaves, J.A. (1991). Iso-lation, characterization and sequence of a gene conferringresistance to the systemic fungicide carboxin from themaize smut pathogen Ustilago maydis. Curr Genet 19:475–481.
Kronstad, J.W. (1997). Virulence and cAMP in smuts, blastsand blights. Trends Plant Sci 2: 194–199.
Krugel, H., Fiedler, G., Smith, C., and Baumberg, S. (1993).Sequence and transcriptional analysis of nourseothricinacetyltransferase-encoding gene nat1 from Streptomycesnoursei. Gene 127: 127–131.
Kruger, J., Loubradou, G., Regenfelder, E., Hartmann, A.,and Kahmann, R. (1998). Crosstalk between cAMP andpheromone signalling pathways in Ustilago maydis. MolGen Genet 260: 193–198.
Madhani, H.D., and Fink, G.R. (1998). The riddle of MAPkinase signalling specificity. Trends Genet 14: 151–155.
Madhani, H.D., Styles, C.A., and Fink, G.R. (1997). MAPkinases with distinct inhibitory functions impart signallingspecificity during yeast differentiation. Cell 91: 673–684.
Mitchell, T.K., and Dean, R.A. (1995). The cAMP-dependentprotein kinase catalytic subunit is required for appressor-ium formation and pathogenesis by the rice blast pathogenMagnaporthe grisea. Plant Cell 7: 1869–1878.
Pi, H., Chien, C.T., and Fields, S. (1997). Transcriptionalactivation upon pheromone stimulation mediated by asmall domain of Saccharomyces cerevisiae Ste12p. MolCell Biol 17: 6410–6418.
Regenfelder, E., Spellig, T., Hartmann, A., Lauenstein, S.,Bolker, M., and Kahmann, R. (1997). G proteins in Ustilagomaydis: transmission of multiple signals. EMBO J 16:1934–1942.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). Molecu-lar Cloning: a Laboratory Manual. Cold Spring Harbor, NY:Cold Spring Harbor Laboratory Press.
Sarkar, G., and Sommer, S.S. (1990) The ‘megaprimer’method of site-directed mutagenesis. Biotechniques 8:404–407.
Schauwecker, F., Wanner, G., and Kahmann, R. (1995).Filament-specific expression of a cellulase gene in thedimorphic fungus Ustilago maydis. Biol Chem Hoppe-Seyler 376: 617–625.
Schulz, B., Banuett, F., Dahl, M., Schlesinger, R., Schafer,W., Martin, T., et al. (1990). The b alleles of U. maydis,whose combinations program pathogenic development,code for polypeptides containing a homeodomain-relatedmotif. Cell 60: 295–306.
Smith, T.L., and Leong, S.A. (1990). Isolation of a Ustilagomaydis glyceraldeyde-3-phosphate dehydrogenase-encoding gene. Gene 93: 111–117.
Snetselaar, K.M., and Mims, C.W. (1993). Light and electronmicroscopy of Ustilago maydis hyphae in maize. MycolRes 98: 347–355.
Snetselaar, K.M., Bolker, M., and Kahmann, R. (1996). Usti-lago maydis mating hyphae orient their growth towardpheromone sources. Fungal Genet Biol 20: 299–312.
Song, O., Dolan, J.W., Yuan, Y.O., and Fields, S. (1991)
Q 1999 Blackwell Science Ltd, Molecular Microbiology, 34, 1007–1017
1016 P. Muller, C. Aichinger, M Feldbrugge and R. Kahmann

Pheromone-dependent phosphorylation of the yeastSTE12 protein correlates with transcriptional activation.Genes Dev 5: 741–750.
Spellig, T., Bolker, M., Lottspeich, F., Frank, R.W., and Kah-mann, R. (1994). Pheromones trigger filamentous growthin Ustilago maydis. EMBO J 13: 1620–1627.
Thompson, J.D., Higgins, D.G., and Gibson, T.J. (1994).CLUSTAL W: improving the sensitivity of progressive multiplesequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. NucleicAcids Res 22: 4673–4680.
Toda, T., Shimanuki, M., and Yanagida, M. (1991). Fissionyeast genes that confer resistance to staurosporineencode a AP-1-like transcription factor and a protein kinaserelated to mammalian ERK1/MAP2 and budding yeastFUS3 and KSS1 kinases. Genes Dev 5: 60–73.
Treisman, R. (1996). Regulation of transcription by MAPkinase cascades. Curr Op Cell Biol 8: 205–215.
Tsukada, T., Carleton, S., Fotheringham, S., and Holloman,
W.K. (1988). Isolation and characterisation of an autono-mously replicating sequence from Ustilago maydis. MolCell Biol 8: 3703–3709.
Urban, M., Kahmann, R., and Bolker, M. (1996). Identificationof the pheromone response element in Ustilago maydis.Mol Gen Genet 251: 31–37.
Whiteway, M., Dignard, D., and Thomas, D.Y. (1992). Domi-nant negative selection of heterologous genes: isolation ofCandida albicans genes that interfere with S. cerevisiaemating factor-induced cell cycle arrest. Proc Natl AcadSci USA 89: 9410–9414.
Xu, J.-R., and Hamer, J.E. (1996). MAP kinase and cAMPsignalling regulate infection structure formation and patho-genic growth in the rice blast fungus Magnaporthe grisea.Genes Dev 10: 2696–2706.
Xu, J.-R., Urban, M., Sweigard, J.A., and Hamer, J.E. (1997).The CPKA gene of Magnaporthe grisea is essential forappressorial penetration. Mol Plant Microbe Interact 10:187–194.
Q 1999 Blackwell Science Ltd, Molecular Microbiology, 34, 1007–1017
MAP kinase Kpp2 in U. maydis 1017