the lactobin a and amylovorin l471 encoding genes are identical, and their distribution seems to be...
TRANSCRIPT

www.elsevier.com/locate/ijfoodmicro
International Journal of Food Microbiology 90 (2004) 93–106
The lactobin A and amylovorin L471 encoding genes are identical,
and their distribution seems to be restricted to the species
Lactobacillus amylovorus that is of interest for cereal fermentations
L. De Vuysta,*, L. Avontsa, P. Neysensa, B. Hosteb, M. Vancanneytb,J. Swingsb, R. Callewaerta
aResearch Group of Industrial Microbiology, Fermentation Technology and Downstream Processing,
Department of Applied Biological Sciences, Vrije Universiteit Brussel, Pleinlaan 2, B-1050 Brussels, BelgiumbLaboratory of Microbiology, Department of Biochemistry, Physiology and Microbiology, BCCM/LMG Bacteria Collection,
Universiteit Gent, K.L. Ledeganckstraat 35, B-9000 Gent, Belgium
Received 28 October 2002; received in revised form 22 April 2003; accepted 30 May 2003
Abstract
Lactobin A and amylovorin L471 are two bacteriocins produced by the phenotypically different strains Lactobacillus
amylovorus LMG P-13139 and L. amylovorus DCE 471, respectively. A 110-bp PCR fragment of the structural gene of lactobin
Awas obtained from total genomic DNA of L. amylovorus LMG P-13139, which was used as a probe to isolate a 3.6-kb HindIII
chromosomal fragment for sequencing. PCR amplification revealed that both the structural genes of both the bacteriocins
lactobin A and amylovorin L471 were identical. These bacteriocins will be further referred to as amylovorin L. Amylovorin L can
be defined as a small, strongly hydrophobic, antibacterial peptide consisting of 50 amino acids. It is synthesized as a precursor
peptide of 65 amino acids processed at a characteristic double-glycine proteolytic cleavage site. Amylovorin L hence belongs to
the class II bacteriocins. It has a narrow inhibitory spectrum, being most active towards Lactobacillus delbrueckii subsp.
bulgaricus LMG 6901T. Among 38 strains of the Lactobacillus acidophilus DNA homology group, another 6 L. amylovorus
strains were also inhibitory towards the L. delbrueckii subsp. bulgaricus LMG 6901T strain. The lactobin A or amylovorin L471
structural genes could be detected in the genomes of three of these L. amylovorus strains, but only after extensive PCR
amplification, indicating that the inhibitory substances were slightly different. The bacteriocins were characterized as small
(approximately 4800 Da), heat-stable peptides that were active in a wide pH range (2.2–8.0). Finally, preliminary experiments
indicated that the production of amylovorin L by L. amylovorusDCE 471 took place during a natural rye fermentation, indicating
its potential importance in the development of a functional (probiotic) starter culture for cereal fermentations.
D 2003 Elsevier B.V. All rights reserved.
Keywords: Amylovorin L471; Lactobin A; Lactobacillus amylovorus; Lactobacillus acidophilus; Bacteriocin distribution
0168-1605/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
doi:10.1016/S0168-1605(03)00298-8
Abbreviations: LBN, lactobin; AMY, amylovorin.
* Corresponding author. Tel.: +32-2-629-32-45; fax: +32-2-
629-27-20.
E-mail address: [email protected] (L. De Vuyst).
1. Introduction
Lactic acid bacteria (LAB) are well known for
their antagonistic activity against a variety of micro-
organisms. Bacteriocin production is one of the

L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–10694
properties responsible for the antibacterial activity
against closely related species, but some Gram-pos-
itive food spoilers and pathogens are often inhibited
as well (De Vuyst and Vandamme, 1994; Klaenham-
mer, 1988). Bacteriocins are low-molecular-mass
peptides or high-molecular-mass proteins that are
mostly hydrophobic and thermostable or hydrophilic
and heat sensitive, respectively (De Vuyst and Van-
damme, 1994; Klaenhammer, 1993). Bacteriocin pro-
duction has been found among almost all genera of
LAB (De Vuyst and Vandamme, 1994; Wood and
Holzapfel, 1995).
The structural genes of a bacteriocin produced by a
strain belonging to a certain species are often wide-
spread among strains within this species. This has
been revealed through PCR amplification for instance
for the genes of nisin A/Z in Lactococcus lactis (De
Vos et al., 1993; Martınez et al., 1995; Meghrous et
al., 1999; Rodrıguez et al., 1995a; Ward et al., 1994),
lactocin S in Lactobacillus spp. (Rodrıguez et al.,
1995b), enterocin AS-48 in Enterococcus spp. (Joos-
ten et al., 1997; Rodrıguez et al., 1998), and pediocin
AcH/PA-1 in Pediococcus acidilactici (Rodrıguez et
al., 1997). Furthermore, it frequently happens that the
same bacteriocin is isolated from cultures of strains
from different species. Usually, this becomes clear
when the amino acid or gene sequence have been
elucidated. Numerous examples are available. For
instance, the bacteriocins sakacin A from Lactobacil-
lus sakei Lb706 (Holck et al., 1992), curvacin A from
Lactobacillus curvatus LTH 1174 (Tichaczek et al.,
1992), and sakacin K from L. sakei CTC 494
(Remiger et al., 1996) are identical. Further, among
different strains, variants of a bacteriocin are often
found, which differ only in one or a few amino acids.
The most common example is nisin A and nisin Z
produced by L. lactis subsp. lactis, which only differ
in one amino acid (Mulders et al., 1991). Further-
more, acidocin 1229 from Lactobacillus acidophilus
JCM 1229 (Tahara and Kanatani, 1996), gassericin
B3 from Lactobacillus gasseri JCM 2124 (Tahara et
al., 1997), and acidocin J1132 from L. acidophilus
JCM 1132 (Tahara et al., 1996) also only differ in
one or two amino acids. Therefore, bacteriocins seem
to be genus specific. Until now, only one exception
has been reported: pediocin AcH/PA-1 is naturally
produced by both P. acidilactici and Lactobacillus
plantarum strains (Ennahar et al., 1996).
Lactobacillus amylovorus, an obligate homofer-
mentative strain and a member of the L. acidophilus
DNA homology group, plays an important role in
several cereal fermentation processes such as type II
sourdoughs based on rye (Muller et al., 2001; Vogel
et al., 1999), and many traditional African and Asian
fermented food products, e.g. Sudanese Kisra pro-
duction from sorghum (Hamad et al., 1992), and
Indian idli made from rice and black gram dough
(Chavan and Kadam, 1989). Furthermore, strains of
L. amylovorus deserve special attention as they have
the capability to adhere to human intestinal cells
(Muller et al., 1998). The combination of adhesion
and formation of antagonistic compounds, in partic-
ular, bacteriocins, are especially tempting for the
development of probiotic strains from cereal origin
(Vogel et al., 1999).
In earlier reports, we described the isolation of
two bacteriocinogenic peptides produced by L. amy-
lovorus LMG P-13139 (Contreras et al., 1997) and
L. amylovorus DCE 471, two strains that differ
phenotypically as well as in their total cell protein
pattern (Callewaert et al., 1999). Isolation and puri-
fication of the antibacterial peptides revealed two
bacteriocins with estimated molecular masses of 4.5
and 6.0 kDa (Contreras et al., 1997), and 4.8 and 5.8
kDa (De Vuyst et al., 1996a), respectively. In both
cases, the smallest peptide was sequenced and des-
ignated as lactobin A and amylovorin L471, respec-
tively (Callewaert et al., 1999; Contreras et al.,
1997). The N-terminal part of 35 amino acids
determined so far for amylovorin L471 is identical
to the N-terminal amino acid sequence of lactobin A
(Callewaert et al., 1999). However, amylovorin L471
differs in molecular mass from lactobin A in 2 Da,
indicating the presence of a disulphide bond in
amylovorin L471 (Callewaert et al., 1999). Despite
this strong similarity, differences in inhibitory spec-
trum are observed in that amylovorin L471 inhib-
its more target bacteria than lactobin A (Callewaert
et al., 1999).
The aim of this paper was to assess the distri-
bution of the amylovorin structural gene among
strains of the L. acidophilus DNA homology group.
First, the structural genes of the bacteriocins lacto-
bin A and amylovorin L471 were sequenced. Next,
the distribution of the amylovorin L471 structural
gene was surveyed through PCR amplification in 38

L. De Vuyst et al. / International Journal of
other Lactobacillus strains of the L. acidophilus
DNA homology group. Further, the bacteriocins
from three other L. amylovorus strains were isolated
and characterized. Finally, based on its compe-
titiveness in rye fermentation, the potential impor-
tance of bacteriocin production by L. amylovorus is
underlined.
2. Material and methods
2.1. Bacterial strains and media
L. amylovorus LMG P-13139 (Contreras et al.,
1997) and L. amylovorus DCE 471 (De Vuyst et al.,
1996a) were used as the lactobin A and the amylo-
vorin L471 producer, respectively. Lactobacillus del-
brueckii subsp. bulgaricus LMG 6901T was used as
the bacteriocin-sensitive indicator organism (De
Vuyst et al., 1996a). The strain Enterococcus fae-
cium CTC 492 was used as a negative control in the
PCR reactions (Aymerich et al., 1996). The latter
strain was kindly provided by Dr. Marta Hugas
(IRTA, Monells, Spain). The strains L. plantarum
LMG 6907T and L. sakei subsp. carnosus LMG
17302T were used as a control in the rep-PCR anal-
ysis. Other LAB strains used are listed in Table 1.
Escherichia coli Sure2 was provided by Stratagene
(La Jolla, CA).
All strains were stored at � 80 jC in their ap-
propriate cultivation medium plus glycerol (final
concentration of 25.0%, v/v): MRS (Oxoid, Basing-
stoke, UK) for lactobacilli and E. faecium, and Luria
Bertani (LB) medium (Sambrook et al., 1989) for E.
coli. Before experimental use, the cultures were
propagated twice at 37 jC for 12 h, along with
shaking at 250 rpm in the case of E. coli, to obtain
fresh cultures; the transfer inoculum was 1.0% (v/v).
Lactobacillus iners was cultivated microaerophili-
cally (controlled atmosphere of 10% oxygen, 10%
carbon dioxide, and 80% nitrogen) in Columbia
medium (Oxoid) supplemented with 5% (v/v) horse
blood. MRS medium adjusted to pH 6.5 was used for
bacteriocin production. Bottom and overlay agar
media were prepared by addition of 15 and 7 g of
granulated agar (Oxoid), respectively, to 1 l of me-
dium. The media were sterilized by heating at 121
jC for 20 min.
2.2. DNA isolation from L. amylovorus LMG P-
13139, PCR-mediated isolation of the structural gene
of lactobin A, and cloning and sequencing of the PCR
fragments
Total genomic DNA from L. amylovorus LMG P-
13139 was prepared by the method described by Boot
et al. (1993). Two 35-mer degenerate lactobin A-
specific primers for PCR were deduced from the N-
and C-terminal amino acid sequence of lactobin A,
respectively (Contreras et al., 1997). The sense primer
(5VAAT/C G C I TAT/C A/T C I G C I G C I T/C T G
G G I T G 3V) correlated with the lactobin A DNA
sequence coding for the amino acids at positions 5–13,
whereas the antisense primer (5VC G TA I C C I G C I
A G I G C G C A I A C I G C 3V) stretched between theamino acids at positions 47 and 39. PCR reactions
were performed according to the standard procedures
in a total volume of 50 Al in the presence of 0.2 pmol
Al� 1 of sense primer, antisense primer, and dNTP mix.
Briefly, 100 ng of template DNA and 1 U Taq DNA
polymerase (Amersham Biosciences, Uppsala, Swe-
den) were added to each reaction. Before cycling,
template DNA was denatured at 95 jC for 3 min. In
each cycle, template DNA was denatured again at 95
jC for 30 s, annealing took place at 50 jC. Polymer-
ization was performed at 72 jC for 2 min. PCRs
consisted of 35 cycles and were followed by an extra
polymerization step at 72 jC for 5 min. Samples were
kept at 4 jC after the reactions were completed. Two
different PCR fragments of approximately 90 and 110
bp were generated, cloned in pBSK+ (2.9-kb vector;
Stratagene), and sequenced in both directions using T3
and T7 primers.
2.3. Cloning and sequencing of the structural gene of
lactobin A
Chromosomal DNA of L. amylovorus LMG P-
13139 was digested with HindIII according to the
manufacturer’s instructions (Boehringer Mannheim,
Mannheim, Germany). The 110-bp PCR fragment
was used as a homologue probe in digoxigenin-
labeled, nonradioactive, and random a-32P-dATP-la-
beled Southern hybridizations, carried out according
to the manufacturers’ instructions (Boehringer Man-
nheim; Amersham Biosciences). A selected DNA
fragment of 3.6 kb was isolated from the complete
Food Microbiology 90 (2004) 93–106 95

Table 1
Strains used for detection of the structural genes of lactobin A and
amylovorin L471
Strain Origin PCR I PCR II
Lactobacillus acidophilus
LMG 11430
human � �
L. acidophilus LMG 11467 human � �L. acidophilus LMG 11469 rat intestine � �L. acidophilus LMG 11472 unknown � �Lactobacillus amylovorus
DCE 471
corn steep liquor + +
L. amylovorus LMG P-13139 corn steep liquor + +
L. amylovorus LMG 9434 pig’s small
intestine
+/� +/�
L. amylovorus LMG 9496T cattle waste–corn
fermentation
� �
L. amylovorus LMG 13049 cattle waste–corn
fermentation
+/� +/�
L. amylovorus LMG 13135 cattle waste–corn
silage
+/� +/�
L. amylovorus LMG 18179 swine intestine � �L. amylovorus LMG 18192 human faeces � �L. amylovorus LMG 18197 pig faeces � �L. amylovorus LMG 18198 calf faeces � �Lactobacillus crispatus
LMG 9479Teye � �
L. crispatus LMG 12004 human urine � �L. crispatus LMG 18191 chicken faeces � �L. crispatus LMG 18199 human faeces � �Lactobacillus gallinarum
LMG 14751
chicken faeces � �
L. gallinarum LMG 14753 chicken faeces � �L. gallinarum LMG 14754 chicken faeces � �L. gallinarum LMG 14755 chicken faeces � �Lactobacillus gasseri
LMG 11413
human saliva � �
L. gasseri LMG 13134 vaginal tract � �L. gasseri LMG 18176 human intestine � �L. gasseri LMG 18203 human faeces � �Lactobacillus helveticus
LMG 13522
unknown � �
L. helveticus LMG 18182 dairy products � �L. helveticus LMG 18183 unknown � �L. helveticus LMG 18225 unknown � �Lactobacillus iners
LMG 18913
human vaginal
discharge
� �
L. iners LMG 18914 human urine � �L. iners LMG 18915 unknown � �L. iners LMG 18916 unknown � �Lactobacillus johnsonii
LMG 9436Thuman blood � �
L. johnsonii LMG 18175 human intestine � �L. johnsonii LMG 18204 mouse faeces � �L. johnsonii LMG 18206 pig faeces � �Enterococcus faecium
CTC 492
fermented
sausage
� �
L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–10696
HindIII digest of chromosomal DNA separated by
agarose gel electrophoresis, cloned in the unique
HindIII site of the pBSK+ cloning vector, and trans-
formed into competent E. coli Sure2 cells, using
general protocols (Sambrook et al., 1989). Nucleotide
sequence analysis was performed by sequencing dou-
ble-stranded DNA in two orientations by the dideoxy-
terminator chain method of Sanger et al. (1977) using
the nonradioactive Taq Dye Deoxy Terminator Cycle
sequencing kit (Applied Biosystems, Foster City, CA)
and a Model 377 A Sequencer (Applied Biosystems).
2.4. DNA isolation from, and primers and PCR
conditions, for detection of the lactobin A structural
gene
Total genomic DNA was isolated by use of the
Puregene kit (Gentra Systems, Minneapolis, MN).
DNA was checked on 2% (m/v) agarose gels and
quantified by the Dynaquant kit (Amersham Bioscien-
ces) with calf thymus DNA as standard (DQ 202,
Sigma, St. Louis, MO).
Three primers were chosen from the DNA se-
quence of the coding part of the lactobin A structural
gene (Fig. 1). The first primer (5V TGG ACT AAT
GCATAC AGC GC 3V), named (LBN) lactobin sense
primer, corresponded to the N-terminal end of lacto-
bin A (Contreras et al., 1997). The second primer (5VCCA ATTACA GCA CCC CATAC 3V), named LBN
antisense primer, corresponded to the C-terminal part
of lactobin A (Contreras et al., 1997). The third primer
(5VCTT TAC GAA CAT AAC CCG CC 3V), named
amylovorin (AMY) antisense primer, corresponded to
the C-terminal part of the known N-terminal amylo-
vorin L471 amino acid sequence, which is identical to
the N-terminal lactobin A amino acid sequence (Call-
ewaert et al., 1999).
A Perkin Elmer GeneAmp PCR System 9600
(Applied Biosystems) was used. The PCR reaction
Notes to Table 1:
CTC, Centre de Tecnologia de la Carn (Institut de Recerca i Tec-
nologia Agroalimentaries, Monells, Spain); DCE, Department of
Chemical Engineering (Vrije Universiteit Brussel, Brussels, Bel-
gium); LMG, Laboratorium Microbiologie Gent Culture Collection
(Ghent University, Gent, Belgium).
PCR I: LBN sense and LBN antisense primer set; PCR II: LBN
sense and AMY antisense primer set: +, amplification; � , no am-
plification; +/� , very weak amplification.

Fig. 1. Nucleotide sequence of the lactobin A structural gene of L. amylovorus P-13139 and primer selection for the detection of the lactobin A
and amylovorin L471 structural genes. The amino acid sequence of lactobin A including the leader is displayed in the one-letter code. The
double-glycine (� 1/� 2), leader-processing site is underlined. The 35 amino acids identical to amylovorin L471 are displayed in bold
(Callewaert et al., 1999). The nucleotide sequences corresponding to the primers are in italic and underlined. They are named the LBN sense,
AMY antisense, and LBN antisense primer, respectively (from left to right). The possible ribosome-binding site is in bold and underlined. The
terminator codon is indicated by an asterisk. Only part of the sequenced 3.6-kb HindIII fragment is shown.
L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–106 97
was done in a volume of 50 Al of Gold PCR buffer
(Applied Biosystems) containing 2.5 mM of MgCl2,
0.2 mM of dNTPs, 0.6 AM of each primer, 1.25 U of
AmpliTaq Gold DNA polymerase (Applied Biosys-
tems), and 5 ng of template DNA (unless stated
otherwise). The amplification was started by a DNA
denaturation and an enzyme activation step at 95 jCfor 10 min. The amplification consisted of 35 cycles of
denaturation and enzyme activation at 95 jC during 30
s, and annealing and extension at 67 jC during 30 s
(unless stated otherwise). Final annealing and exten-
sion occurred at 67 jC for 12 min. At the end, the
temperature was decreased to 4 jC to stop the reaction.
First, the PCR conditions for detection of the
lactobin A structural gene were optimized. PCR ampli-
fications were carried out with 1, 5, 10, and 100 ng of
template DNA at an annealing and elongation temper-
ature of 63, 66, 67, 68, and 69 jC. The lactobin A
producer, L. amylovorus LMG P-13139, and the amy-
lovorin L471 producer, L. amylovorus DCE 471, were
used as positive controls; the enterocin A producer, E.
faecium CTC 492, was used as negative control. Total
genomic DNA was used as template.
Amplified DNA fragments were visualized on a
4.5% (m/v) LSI MP agarose gel (Boehringer Man-
nheim) after staining with ethidium bromide. A
øX174/HinfI DNA marker (726 to 24 bp; Stratagene)
was used to determine the length of the amplified
fragments. The gel was run at 150 V for 150 min.
Based on the nucleotide sequence of the lactobin A
structural gene (Fig. 1), a 144-bp DNA fragment and a
96-bp DNA fragment was expected to be amplified
with the LBN sense and LBN antisense primer set
(PCR I), and the LBN sense and AMY antisense
primer set (PCR II), respectively.
Cleaning of the PCR products to be sequenced was
done by use of the QIA quick spin PCR purification
kit (Qiagen, Hilden, Germany). The sequencing reac-
tion was carried out with 4 Al of Big Dye (Applied
Biosystems), 3 Al of PCR product, and 3 Al of primer
(LBN sense and LBN antisense or AMY antisense) of
20 ng Al� 1. The sequence of double-stranded DNA in
two orientations was determined on an ABI PRISM
377 DNA Sequencer (Applied Biosystems).
To study the distribution of the lactobin A structural
gene among strains of the L. acidophilus DNA ho-
mology group, two PCR experiments were performed,
one with the LBN sense and antisense primers (PCR I)
and another with the LBN sense and AMY antisense
primers (PCR II). Total DNA from L. amylovorus
LMG P-13139 and E. faecium CTC 492 were used
as positive and negative controls, respectively.
2.5. Bacteriocin isolations and assay of bacteriocin
activity and inhibitory spectrum
Lactobin A, amylovorin L471, and the bacteriocins
from L. amylovorus LMG 9434, L. amylovorus LMG
9496T, L. amylovorus LMG 13049, and L. amylovorus
LMG 13135 were produced and isolated as described
previously (Callewaert et al., 1999; De Vuyst et al.,
1996a). The material thus obtained was considered as
a purified bacteriocin preparation and could be stored
at � 80 jC (Callewaert et al., 1999).
Bacteriocin activity was measured by an agar spot
assay as described previously (De Vuyst et al., 1996a).
Briefly, twofold serial dilutions of cell-free culture
supernatant containing bacteriocin were spotted (10
Al) onto fresh lawns of the sensitive indicator organ-
ism L. delbrueckii subsp. bulgaricus LMG 6901T.

L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–10698
These lawns were prepared by propagating fresh
cultures to an optical density at 600 nm (OD600)
between 0.40 and 0.45, and adding 100 Al of the cell
suspension to 3.5 ml of overlay agar. Overlaid agar
plates were incubated at 37 jC for at least 12 h. The
bactericidal activity was defined as the reciprocal of
the highest dilution demonstrating complete inhibition
of the indicator lawn and was expressed in activity
units (AU) per milliliter of culture medium. To assay
the spectrum of activity of a bacteriocin towards other
lactobacilli, the antimicrobial activity of a purified
bacteriocin preparation was examined against the
target bacteria (Table 2) in at least two separate tests.
2.6. Characterization of the bacteriocins
For a characterization of the bacteriocins of L.
amylovorus LMG 9434 (solution of 50 AU ml� 1),
L. amylovorus LMG 13049 (3200 AU.ml� 1), and L.
amylovorus LMG 13135 (1200 AU.ml� 1), compared
to L. amylovorus DCE 471 (6400 AU ml� 1) and L.
amylovorus LMG P-13139 (1600 AU ml� 1), purified
bacteriocin was treated with several proteases in the
Table 2
Inhibitory spectra of the purified bacteriocin preparations from L.
amylovorus strains DCE 471, LMG P-13139, LMG 13135, LMG
13049, LMG 9496T, and LMG 9434
Bacteriocin
preparation
Indicator strain
DCE
471
LMG
P-13139
LMG
13049
LMG
13135
LMG
9496TLMG
9434
L. delbrueckii subsp.
bulgaricus LMG
6901T
+ + + + + +
L. amylovorus LMG
9434
+ + + + + �
L. amylovorus LMG
9496T+ + + � � �
L. amylovorus LMG
13135
+ + � � � �
L. amylovorus LMG
13049
+ + � � � �
L. amylovorus LMG
P-13139
+ � � � � �
L. amylovorus DCE
471
� � � � � �
The activities against L. delbrueckii subsp. bulgaricus LMG 6901T
of all preparations were 6400 AU ml� 1, except for the purified
bacteriocin preparation from L. amylovorus LMG 9434 that was
200 AU ml� 1.
+, inhibition; � , no inhibition.
appropriate buffer and final concentration indicated:
proteinase K, 1 mg ml� 1, 50 mM phosphate buffer, pH
7.0, 37 jC; trypsin, 2 mg ml� 1, 50 mM phosphate
buffer, pH 7.0, 37 jC; a-chymotrypsin, 5 mg ml� 1, 50
mM phosphate buffer, pH 7.5, 37 jC; and pepsin, 1 mg
ml� 1, 200 mM citrate buffer, pH 2.2, 37 jC. All
enzymes were purchased from VWR International
(Darmstadt, Germany). The residual activity of the
bacteriocin solutions was tested towards L. delbrueckii
subsp. bulgaricus LMG 6901T as described above. The
appropriate buffers and enzyme/buffer solutions were
used as controls. The activity was further tested after
various heat treatments (15, 30, and 60 min at 60 and
100 jC, and autoclaving during 15 min at 121 jC);nonheat-treated samples were used as a control. The
influence of pH on activity was tested in the following
buffers: 200 mM citrate buffer for pH 2.2, 3.0, 4.0, 4.5,
5.0, 5.5, and 6.0, and 100 mM sodium phosphate
buffer for pH 6.5, 7.0, and 8.0. The activity was
measured after 1 h and after 24 h of incubation at 25
jC. Buffer solutions were used as a negative control.
To estimate the molecular mass of the bacteriocins,
tricine–SDS–PAGE was carried out as described
previously (De Vuyst et al., 1996a).
2.7. Rye fermentation with L. amylovorus DCE 471
To test its suitability for rye fermentation, a freshly
prepared inoculum of L. amylovorus DCE 471 was
added to a mixture of rye (2.5 kg) and water (7.5 l) in
a Biostat C fermentor (B. Braun Biotech International,
Melsungen, Germany). The water was sterilized in the
fermentor at 121 jC for 20 min; after cooling to 37
jC, the rye was added to the fermentor, immediately
followed by inoculation. The fermentation was carried
out at a controlled temperature of 37 jC. The pH was
monitored on line. The stirring speed was 600 rpm; it
was decreased to 400 rpm after 12 h as the sourdough
viscosity decreased during the fermentation. Samples
were aseptically removed from the fermentation ves-
sel after 0, 3, 6, 12, 15, and 24 h, and analyzed for
total titratable acidity, total bacteria and yeast counts,
and soluble bacteriocin activity. Total titratable acidity
(TTA) was determined as the amount (ml) of 0.1 M
NaOH to titrate an aliquot of 10 g of a sample blended
with 100 ml of distilled water to a final pH of 8.5.
Total cell counts (CFU ml�1) of fresh samples were
determined on plates of Sourdough Simulation Medi-

L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–106 99
um (SSM) agar adjusted to pH 5.4 (Messens et al.,
2002). Soluble bacteriocin activity of culture super-
natants obtained after microcentrifugation (13,000
rpm, 20 min) was tested by bioassay towards L.
delbrueckii subsp. bulgaricus LMG 6901T as indicator
organism as described above. Finally, to verify strain
domination of the added culture of L. amylovorus
DCE 471, four bacterial colonies from an agar plate
with approximately 10–100 colonies were randomly
picked up. The authenticity of these colonies was
identified by rep-PCR according to the method of
Gevers et al. (2001). The ability to produce bacterio-
cin was tested by overlaying the agar plates with soft
agar containing the sensitive indicator strain L. del-
brueckii subsp. bulgaricus LMG 6901T and observing
the plates for inhibition zones around the colonies.
Fig. 2. Visualization of the amplified PCR fragments. PCR am-
plification reactions were done on a blank sample (lanes 2 and 6)
and on total DNA of the following strains: E. faecium CTC 492
(lanes 3 and 7), L. amylovorus LMG P-13139 (lanes 4 and 8), and L.
amylovorus DCE 471 (lanes 5 and 9). Lanes 1 and 10 contain DNA
molecular mass markers (726 to 24 bp, top to bottom). In lanes 6 to
9, the LBN sense and LBN antisense primer set was used for the
amplification of a 144-bp fragment of the lactobin A structural gene.
In lanes 2–5, the LBN sense and AMY antisense primer set were
used for the amplification of a 96-bp fragment of the lactobin A
structural gene corresponding to the known N-terminal amino acid
sequence of amylovorin L471.
3. Results
3.1. PCR-mediated isolation of the structural gene of
lactobin A, cloning and sequencing of the PCR
fragments, and cloning and sequencing of the
structural gene of lactobin A
The 110-bp PCR fragment encoded a part of the
amino acid sequence of lactobin A, produced by L.
amylovorus LMG P-13139, and was located between
amino acids 5 and 47 (Fig. 1). The smaller 90-bp PCR
fragment concerned a 5Vtruncated form of the 110-bp
fragment due to nonspecific downstream hybridiza-
tion of the sense primer.
L. amylovorus LMG P-13139 contained only one
plasmid of approximately 30 kb that could be cured
from the strain by three consecutive overnight incu-
bations in the presence of 200 Ag of novobiocin per
ml without loss of bacteriocin production (results not
shown). The plasmid did not hybridize with chromo-
somal DNA of L. amylovorus LMG P-13139 in
Southern blot experiments, excluding the possibility
of chromosomal incorporation (results not shown).
Partial sequencing of the 3.6-kb HindIII chromo-
somal DNA fragment revealed the complete nucleotide
sequence of the structural gene of lactobin A, lbnA. A
relevant part of the sequencing results is represented in
Fig. 1. The structural gene encodes a prebacteriocin of
50 amino acids with a 15 amino acid leader peptide and
a G(� 2)G(� 1) proteolytic processing site, character-
istic of class II bacteriocins. Upstream of lbnA, a
possible ribosome-binding site is located.
3.2. PCR conditions for detection of the lactobin A
structural gene, sequencing of the lactobin A and
amylovorin L471 PCR amplicons, and detection of the
lactobin A and the amylovorin L471 structural genes
among other strains of the L. acidophilus DNA
homology group
An annealing and extension temperature of 67 jCand an amount of 5 ng of template DNAwere optimal
for the detection of the 144-bp and 96-bp DNA
fragments. At an annealing and elongation tempera-
ture of 63 jC, similar amplifications were obtained
with 1, 5, and 10 ng of template DNA. Increasing the
amount of template DNA to 100 ng decreased the
amplification. Increasing the annealing and elongation
temperature from 63 to 66, 67, 68, and 69 jC also
resulted in a decreasing amplification.
Both amplicons were detected in total DNA of the
lactobin A producer as well as in total DNA of the
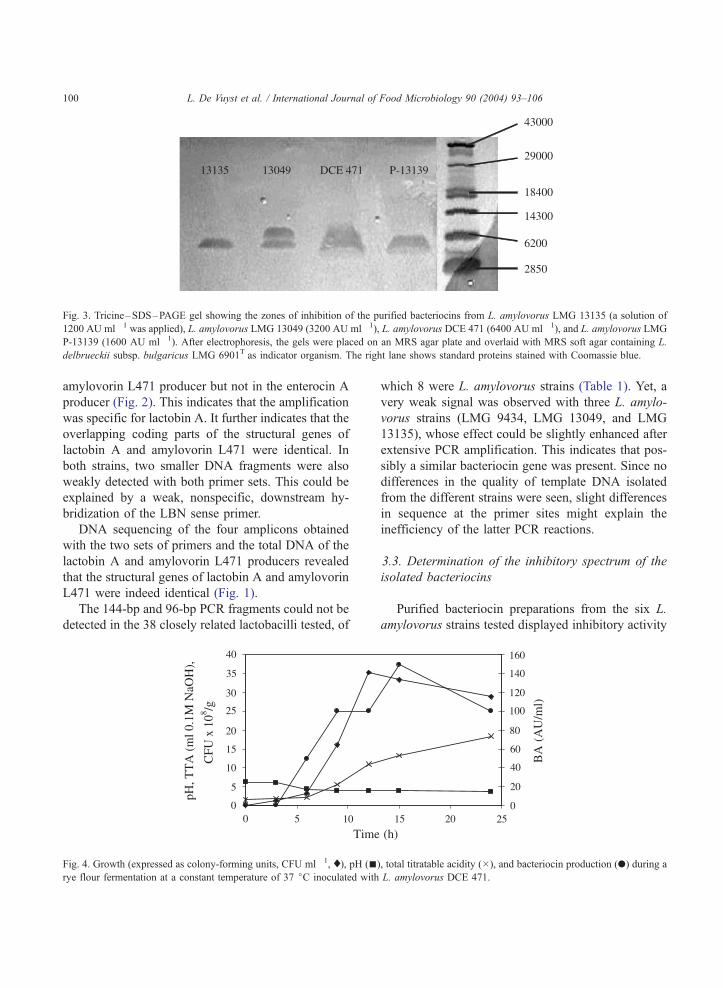
Fig. 3. Tricine–SDS–PAGE gel showing the zones of inhibition of the purified bacteriocins from L. amylovorus LMG 13135 (a solution of
1200 AU ml� 1 was applied), L. amylovorus LMG 13049 (3200 AU ml� 1), L. amylovorus DCE 471 (6400 AU ml� 1), and L. amylovorus LMG
P-13139 (1600 AU ml� 1). After electrophoresis, the gels were placed on an MRS agar plate and overlaid with MRS soft agar containing L.
delbrueckii subsp. bulgaricus LMG 6901T as indicator organism. The right lane shows standard proteins stained with Coomassie blue.
L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–106100
amylovorin L471 producer but not in the enterocin A
producer (Fig. 2). This indicates that the amplification
was specific for lactobin A. It further indicates that the
overlapping coding parts of the structural genes of
lactobin A and amylovorin L471 were identical. In
both strains, two smaller DNA fragments were also
weakly detected with both primer sets. This could be
explained by a weak, nonspecific, downstream hy-
bridization of the LBN sense primer.
DNA sequencing of the four amplicons obtained
with the two sets of primers and the total DNA of the
lactobin A and amylovorin L471 producers revealed
that the structural genes of lactobin A and amylovorin
L471 were indeed identical (Fig. 1).
The 144-bp and 96-bp PCR fragments could not be
detected in the 38 closely related lactobacilli tested, of
Fig. 4. Growth (expressed as colony-forming units, CFU ml� 1, x), pH (nrye flour fermentation at a constant temperature of 37 jC inoculated with
which 8 were L. amylovorus strains (Table 1). Yet, a
very weak signal was observed with three L. amylo-
vorus strains (LMG 9434, LMG 13049, and LMG
13135), whose effect could be slightly enhanced after
extensive PCR amplification. This indicates that pos-
sibly a similar bacteriocin gene was present. Since no
differences in the quality of template DNA isolated
from the different strains were seen, slight differences
in sequence at the primer sites might explain the
inefficiency of the latter PCR reactions.
3.3. Determination of the inhibitory spectrum of the
isolated bacteriocins
Purified bacteriocin preparations from the six L.
amylovorus strains tested displayed inhibitory activity
), total titratable acidity (�), and bacteriocin production (.) during a
L. amylovorus DCE 471.

L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–106 101
against L. delbrueckii subsp. bulgaricus LMG 6901T
(Table 2). L. amylovorus DCE 471 showed an activity
against all L. amylovorus strains tested. The other L.
amylovorus strains could be ranked in order of increas-
ing ability to inhibit other L. amylovorus strains. For
instance, L. amylovorus LMG 13049 inhibited two out
of six of the L. amylovorus strains tested, while L.
amylovorus LMG 9434 did not inhibit any of the L.
amylovorus strains tested. The same order also corre-
Fig. 5. Rep-PCR fingerprints using the (GTG)5 primer of randomly picked
12 h (lanes 13–16), 15 h (lanes 17–20), and 24 h (lanes 21–24) of ferment
fermentor (lane 25), L. plantarum LMG 6907T (lane 26) and L. sakei sub
sponded with a decreasing sensitivity against each
other. The most sensitive strain was L. amylovorus
LMG 9434, followed by L. amylovorus LMG 9496T, L.
amylovorus LMG 13135, L. amylovorus LMG 13049,
L. amylovorus LMG P-13139, and L. amylovorusDCE
471. Bacteriocin preparations from L. amylovorusDCE
471 and L. amylovorus LMG P-13139 displayed sim-
ilar inhibiting activities against other L. amylovorus
strains (Table 2). It was confirmed that the DCE 471
up colonies after 3 h (lanes 1–4), 6 h (lanes 5–8), 9 h (lanes 9–12),
ation; the preculture of L. amylovorus DCE 471 used to inoculate the
sp. carnosus LMG 17302T (lane 27). M: molecular mass marker.

Fig. 6. Bacteriocin production of colonies of L. amylovorus DCE
471 after 3 h (dilution, 10� 6, plate 1), 6 h (dilution, 10� 6, plate 2),
9 h (dilution, 10� 7, plate 3), 12 h (dilution, 10� 7, plate 4), and 15 h
(dilution, 10� 7, plate 5) of fermentation of rye flour that was
inoculated with a culture of L. amylovorus DCE 471 at a constant
temperature of 37 jC. The plates were overlayed with the indicator
strain L. delbrueckii subsp. bulgaricus LMG 6901T.
L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–106102
bacteriocin preparation inhibited L. amylovorus LMG
P-13139 as well; in contrast, the LMG P-13139 prep-
aration did not inhibit L. amylovorus DCE 471 (Call-
ewaert et al., 1999).
3.4. Characterization of the bacteriocins
The inhibitory activity of the purified bacteriocin
preparations of L. amylovorus LMG 9434, L. amylo-
vorus LMG 13049, L. amylovorus LMG 13135, and
of L. amylovorus DCE 471 and L. amylovorus LMG
P-13139 was lost after incubation with the proteolytic
enzymes proteinase K, trypsin, a-chymotrypsin, and
pepsin. This confirmed that an active protein moiety
was responsible for the inhibition of the indicator
strain. Further, the compounds retained full activity
after various heat treatments, including 60 min at 60
jC and 15 min at 100 jC for all L. amylovorus strains
tested. They were only partially inactivated after 60
min at 100 jC in the case of L. amylovorus LMG P-
13139 and L. amylovorus DCE 471 (a decrease with a
factor two) or after autoclaving during 15 min at 121
jC (a decrease with a factor four to eight). All
bacteriocin preparations were active over a wide range
of pH (2.2–8.0). All these data contribute to the
bacteriocinogenic nature of the antibacterial substan-
ces. Finally, tricine–SDS–PAGE showed that all L.
amylovorus strains produced a bacteriocin that dis-
played a molecular mass of approximately 4800 Da,
comparable to that of amylovorin L471 (Fig. 3). In the
case of L. amylovorus LMG 13049, two antibacterial
peptides were detected with a molecular mass of 4800
and 5800 Da, respectively. The purified bacteriocin of
L. amylovorus LMG 9434 was also run on this gel but
did not give an inhibition zone because of its too low
activity (200 AU ml� 1).
3.5. Rye fermentation with L. amylovorus DCE 471
Fig. 4 displays the growth and bacteriocin produc-
tion of L. amylovorus DCE 471 during a rye flour
fermentation at a constant temperature of 37 jC. Theinitial pH value of the dough mixture was 6.3
corresponding with a TTA of 1.4. Samples taken
immediately after inoculation showed a bacterial count
of 2.5� 105 CFU g� 1. Exponential growth of L.
amylovorus DCE 471 started after about 3 h and lasted
till 12 h of fermentation to reach a bacterial count of
3.5� 109 CFU g� 1. This coincided with a fast acidi-
fication of the medium as indicated by the observed pH
drop and increasing TTAvalue. An acidification rate of
0.13 h� 1 could be calculated. The growth was paral-
leled with amylovorin L471 production, underlining its
growth-associated kinetics and potential contribution
in competitiveness of the strain (De Vuyst et al., 1996b;
Messens et al., 2002). Maximum bacteriocin titers (150
AU ml� 1) were observed at the end of the exponential
growth phase, followed by a decrease most probably
due to adsorption to cells, flour particles, etc. (De Vuyst
et al., 1996b). The bacterial count dropped to 2.9� 108
CFU g� 1 after 24 h of fermentation. Then, a maximum
TTA of 18.4 was reached, which coincided with a final
pH value of 3.5. The dominance and competitiveness

L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–106 103
of the added L. amylovorus DCE 471 strain has been
shown in two manners. First, identical rep-PCR finger-
prints were found for the preculture and the 24 samples
of four bacterial colonies randomly picked up after 3, 6,
9, 12, 15, and 24 h of rye fermentation (Fig. 5a,b).
Second, all colonies were shown to produce amylo-
vorin L471 throughout the fermentation as revealed by
their inhibition zones towards L. delbrueckii subsp.
bulgaricus LMG 6901T in an overlay assay (Fig. 6).
4. Discussion
The phenotypically different strains L. amylovorus
LMG P-13139 and L. amylovorus DCE 471 have
been identified as producers of a bacteriocin, called
lactobin A and amylovorin L471, respectively, with a
small difference in molecular mass and an identical N-
terminal amino acid sequence (Callewaert et al., 1999;
Contreras et al., 1997). Analysis of the amplified PCR
fragments from the structural bacteriocin genes iso-
lated from total DNA of both strains revealed that
lactobin A and amylovorin L471 are identical bacter-
iocins. They will be further referred to as amylovorin
L. The bacteriocin amylovorin L can be defined as a
small, strongly hydrophobic peptide, consisting of 50
amino acids, that is synthesized as a precursor peptide
of 65 amino acids processed at a characteristic double-
glycine proteolytic cleavage site, and hence belongs to
the class II bacteriocins (Nes et al., 1996). It has a
narrow inhibitory spectrum, being most active to-
wards L. delbrueckii subsp. bulgaricus LMG 6901T
among the strains tested (De Vuyst et al., 1996a;
Callewaert et al., 1999). The production of amylo-
vorin L seems to be associated with only some L.
amylovorus strains since the structural genes were not
detected in other strains of the L. acidophilus DNA
homology group. Nevertheless, almost all L. amylo-
vorus strains (LMG 13135, LMG 13049, LMG 9496T,
and LMG 9434) used in this study displayed antago-
nistic activity against L. delbrueckii subsp. bulgaricus
LMG 6901T. Their antibacterial peptides could be
concentrated and purified by ammonium sulphate
precipitation and chloroform/methanol extraction as
was done for amylovorin L471, lactobin A, and other
small, hydrophobic bacteriocins (De Vuyst et al.,
1996a; Zamfir et al., 1999; Foulquie Moreno et al.,
2003). The sensitivity of all these antibacterial pep-
tides towards several proteases further indicates their
bacteriocin nature. Hence, our data strongly indicate
that all L. amylovorus strains examined produced a
small bacteriocin that is characterized by a narrow
inhibitory spectrum. The latter property seems to be
common for bacteriocins from L. acidophilus (Bare-
foot and Klaenhammer, 1983).
Bacteriocin production and resistance among L.
amylovorus strains seems to be very common. L.
amylovorus DCE 471 showed a bactericidal activity
against all L. amylovorus strains tested. It was resis-
tant against the bactericidal compounds produced by
the other L. amylovorus strains. In contrast, L. amy-
lovorus LMG 9434, the most sensitive strain, dis-
played bactericidal activity only against L. delbrueckii
subsp. bulgaricus LMG 6901T. The other L. amylo-
vorus strains studied varied in bactericidal activity and
resistance. These data could suggest the production of
multiple bacteriocins and their corresponding immu-
nity proteins by each strain. Bacteriocin immunity
proteins specifically protect the bacteriocin producer
against the production of its own bacteriocin (Quadri
et al., 1995). In addition, bacteriocin production may
render bacteria more resistant against other bacterio-
cins (Eijsink et al., 1998). This observation was
suggested to be responsible for the specific action of
bacteriocins within closely related bacteria. It may
play an important role in the competitiveness of the
strains in a particular ecosystem. Moreover, the adapt-
ability of the strain in a food matrix is a very
important criterion in the selection procedure of a
suitable starter strain (Leroy et al., 2002).
All L. amylovorus strains used were isolated from a
similar corn source, except for L. amylovorus LMG
9434 that was isolated from the small intestines of a
pig (Table 1). L. amylovorus LMG 9496T, L. amylo-
vorus LMG 13135, and L. amylovorus LMG 13049
originated from a cattle waste–corn fermentation. L.
amylovorus DCE 471 (De Vuyst et al., 1996a) and L.
amylovorus LMG P-13139 (Contreras et al., 1997)
were isolated from corn steep liquor. Their cereal
origin may indicate the usefulness of L. amylovorus
strains in such an environment. Since an in situ
bacteriocin activity from L. amylovorus DCE 471
was detected during a rye fermentation inoculated
with this culture, and apparently being dominated by
the strain, the bacteriocin production may be respon-
sible for an improved competitiveness of this strain in

L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–106104
a cereal environment. This opens the way for the use
of bacteriocin-producing L. amylovorus strains in both
rye and sorghum fermentations (Messens et al., 2002).
An ecosystem of increasing importance is the
gastrointestinal tract, in particular, with respect to
the use of probiotic bacteria. It is well known that
probiotic LAB contribute to the stabilization of the gut
microflora (Salminen et al., 1996), thereby avoiding
the colonization of pathogenic bacteria through for
instance the production of antimicrobials like organic
acids (Ibrahim and Bezkorovainy, 1993), and preven-
tion of adherence to the epithelial cells through
competitive exclusion (Bernet et al., 1993; Chauviere
et al., 1992) or inhibition of adhesion (Coconnier et
al., 1993). Also, bacteriocins produced by LAB may
play an important role in this complex ecosystem. For
instance, from the 96 strains of the L. acidophilus
group isolated from human intestines, 62 displayed
bacteriocinogenic activity, indicating that bacteriocin
production is widespread among these bacteria
(Kawai et al., 1997). Indeed, almost all members of
the L. acidophilus group produce bacteriocins: L.
acidophilus (e.g. acidocin A, Kanatani et al., 1995),
L. amylovorus (e.g. amylovorin L471, De Vuyst et al.,
1996a), Lactobacillus crispatus (e.g. crispacin A,
Tahara and Kanatani, 1997), L. gasseri (e.g. gassericin
KT7, Zhu et al., 2000), and Lactobacillus johnsonii
(e.g. lactacin F, Fremaux et al., 1993). Interestingly,
the L. acidophilus group encompasses important pro-
biotic strains (Klein et al., 1998). However, up to now,
only bacteriocins from cultures of LAB strains isolat-
ed from faeces have been purified (Kawai et al., 1998,
2000, 2001). Remarkably, several of these bacterio-
cins as well as other bacteriocins from the L. acid-
ophilus DNA homology group share the same primary
sequence such as gassericin T (Kawai et al., 2000) and
lactacin X (Fremaux et al., 1993), lactobin A (Con-
treras et al., 1997) and amylovorin L471 (Callewaert
et al., 1999), and gassericin A (Kawai et al., 1998) and
acidocin B (Leer et al., 1995). They may therefore
play an important role in this complex ecosystem.
As a conclusion, it is likely that bacteriocin forma-
tion by L. amylovorus is interesting for the develop-
ment of probiotic LAB strains of cereal origin, cereals
becoming of growing importance in the development
of functional foods (Charalampopoulos et al., 2002).
It may contribute to a controlled development of the
cereal microflora during cereal fermentation, simulta-
neously involving the addition of a starter culture with
potential health-promoting properties.
Acknowledgements
The research presented was financially supported by
the Research Council of the Vrije Universiteit Brussel
(VUB) and the Fund for Scientific Research—
Flanders. Supports from Dr. Hilde Revets from the
Research Group of Cellular Immunology (VUB), from
Dr. Bruno Pot and Karen Lefebvre from the Laboratory
of Microbiology of Ghent University (RUG), and from
Dr. Danielle Janssens from the BCCM/LMG Bacteria
Collection (RUG) are gratefully acknowledged.
References
Aymerich, T., Holo, H., Havarstein, L.S., Hugas, M., Garriga, M.,
Nes, I.F., 1996. Biochemical and genetic characterization of
enterocin A from Enterococcus faecium, a new antilisterial bac-
teriocin in the family of pediocins. Applied and Environmental
Microbiology 62, 1676–1682.
Barefoot, S.F., Klaenhammer, T.R., 1983. Detection and activity of
lactacin B, a bacteriocin produced by Lactobacillus acidophilus.
Applied and Environmental Microbiology 45, 1808–1815.
Bernet, M.F., Brassart, D., Neeser, J.-R., Servin, A.L., 1993. Adher-
ing bifidobacterial strains inhibit interactions of enteropathogens
with cultured human intestinal epithelial cells. Applied and En-
vironmental Microbiology 59, 4121–4128.
Boot, H.J., Kolen, C.P.A.M., Van Noort, S.M., Pouwels, P.H., 1993.
S-layer protein of Lactobacillus acidophilus ATCC 4356: pu-
rification, expression in Escherichia coli, and nucleotide se-
quence of the corresponding gene. Journal of Bacteriology 175,
6089–6096.
Callewaert, R., Holo, H., Devreese, B., Van Beeumen, J., Nes, I.,
De Vuyst, L., 1999. Characterization and production of amylo-
vorin L471, a bacteriocin purified from Lactobacillus amylovo-
rus DCE 471 by a novel three-step method. Microbiology 145,
2559–2568.
Charalampopoulos, D., Wang, R., Pandiella, S.S., Webb, C., 2002.
Application of cereals and cereal components in functional
foods: a review. International Journal of Food Microbiology
79, 131–141.
Chauviere, G., Coconnier, M.-H., Kerneis, S., Darfeuille-Michaud,
A., Joly, B., Servin, A.L., 1992. Competitive exclusion of diar-
rheagenic Escherichia coli (ETEC) from enterocyte-like Caco-2
cells in culture. FEMS Microbiology Letters 91, 213–218.
Chavan, J.K., Kadam, S.S., 1989. Nutritional improvement of ce-
reals by fermentation. Critical Reviews in Food Science and
Nutrition 28, 349–400.
Coconnier, M.-H., Bernet, M.F., Kerneis, S., Chauviere, G., Fourn-
iat, J., Servin, A.L., 1993. Inhibition of adhesion of enteroinva-

L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–106 105
sive pathogens to human intestinal Caco-2 cells by Lactobacil-
lus acidophilus LB strain decreases bacterial invasion. FEMS
Microbiology Letters 110, 299–306.
Contreras, B.G.L., De Vuyst, L., Devreese, B., Busanyova, K.,
Raymaeckers, J., Bosman, F., Sablon, E., Vandamme, E.J.,
1997. Isolation, purification, and amino acid sequence of lacto-
bin A, one of the two bacteriocins produced by Lactobacillus
amylovorus LMG P-13139. Applied and Environmental Micro-
biology 63, 13–20.
De Vos, W.M., Mulders, J.W.M., Siezen, R.J., Hugenholtz, J.,
Kuipers, O.P., 1993. Properties of nisin Z and distribution of
its gene, nisZ, in Lactococcus lactis. Applied and Environmental
Microbiology 59, 213–218.
De Vuyst, L., Vandamme, E.J., 1994. Bacteriocins of Lactic Acid
Bacteria: Microbiology, Genetics and Applications. Blackie
Academic & Professional, London.
De Vuyst, L., Callewaert, R., Pot, B., 1996a. Characterization of the
antagonistic activity of Lactobacillus amylovorus DCE 471 and
large scale isolation of its bacteriocin amylovorin L471. System-
atic and Applied Microbiology 19, 9–20.
De Vuyst, L., Callewaert, R., Crabbe, K., 1996b. Primary metabolite
kinetics of bacteriocin biosynthesis by Lactobacillus amylovorus
and evidence for stimulation of bacteriocin production under
unfavourable growth conditions. Microbiology 142, 817–827.
Eijsink, V.G.H., Skeie, M., Middelhoven, P.H., Brurberg, M.B.,
Nes, I.F., 1998. Comparative studies of class IIa bacteriocins
of lactic acid bacteria. Applied and Environmental Microbiol-
ogy 64, 3275–3281.
Ennahar, S., Aoude-Werner, D., Sorokine, O., van Dorsselaer, A.,
Bringel, F., Hubert, J.C., Hasselmann, C., 1996. Production of
pediocin AcH by Lactobacillus plantarum WHE 92 isolated
from cheese. Applied and Environmental Microbiology 62,
4381–4387.
Foulquie Moreno, M.R., Devreese, B., Van Beeumen, J., De Vuyst,
L., 2003. Isolation and biochemical characterisation of enter-
ocins produced by enterococci from different sources. Journal
of Applied Microbiology 94, 214–229.
Fremaux, C., Ahn, C., Klaenhammer, T.R., 1993. Molecular anal-
ysis of the lactacin F operon. Applied and Environmental Micro-
biology 59, 3906–3915.
Gevers, D., Huys, G., Swings, J., 2001. Applicability of rep-PCR
fingerprinting for identification of Lactobacillus species. FEMS
Microbiology Letters 205, 31–36.
Hamad, S.H., Bocker, G., Vogel, R.F., Hammes, W.P., 1992. Micro-
biological and chemical analysis of fermented sorghum dough
for Kisra production. Applied Microbiology and Biotechnology
37, 728–731.
Holck, A., Axelsson, L., Birkeland, S.-T., Aukrust, T., Blom, H.,
1992. Purification and amino acid sequence of sakacin A, a
bacteriocin from Lactobacillus sake Lb706. Journal of General
Microbiology 138, 2715–2720.
Ibrahim, S.A., Bezkorovainy, A., 1993. Inhibition of Escherichia
coli by bifidobacteria. Journal of Food Protection 56, 713–715.
Joosten, H.M.L.J., Rodrıguez, E., Nunez, M., 1997. PCR detection
of sequences similar to the AS-48 structural gene in bacteriocin-
producing enterococci. Letters in Applied Microbiology 24,
40–42.
Kanatani, K., Oshimura, M., Sano, K., 1995. Isolation and charac-
terization of acidocin A and cloning of the bacteriocin gene
from Lactobacillus acidophilus. Applied and Environmental
Microbiology 61, 1061–1067.
Kawai, Y., Saito, T., Uemura, J., Itoh, T., 1997. Rapid detection
method for bacteriocin and distribution of bacteriocin-producing
strains in Lactobacillus acidophilus group lactic acid bacteria
isolated from human feces. Bioscience, Biotechnology, and Bi-
ochemistry 61, 179–182.
Kawai, Y., Saito, T., Kitazawa, H., Itoh, T., 1998. Gassericin A; an
uncommon cyclic bacteriocin produced by Lactobacillus gas-
seri LA39 linked at N- and C-terminal ends. Bioscience, Bio-
technology, and Biochemistry 62, 2438–2440.
Kawai, Y., Saitoh, B., Takahashi, O., Kitazawa, H., Saito, T., Na-
kajima, H., Itoh, T., 2000. Primary amino acid and DNA se-
quences of gassericin T, a lactacin F-family bacteriocin
produced by Lactobacillus gasseri SBT 2055. Bioscience, Bio-
technology, and Biochemistry 64, 2201–2208.
Kawai, Y., Ishii, Y., Uemura, K., Kitazawa, H., Saito, T., Itoh, T.,
2001. Lactobacillus reuteri LA6 and Lactobacillus gasseri
LA39 isolated from faeces of the same human infant produce
identical cyclic bacteriocin. Food Microbiology 18, 407–415.
Klaenhammer, T.R., 1988. Bacteriocins of lactic acid bacteria. Bio-
chimie 70, 337–349.
Klaenhammer, T.R., 1993. Genetics of bacteriocins produced by
lactic acid bacteria. FEMS Microbiology Reviews 12, 39–86.
Klein, C., Pack, A., Bonaparte, C., Reuter, G., 1998. Taxonomy and
physiology of probiotic lactic acid bacteria. International Journal
of Food Microbiology 41, 103–125.
Leer, R.J., van der Vossen, J.M., van Giezen, M., van Noort, J.M.,
Pauwels, P.H., 1995. Genetic analysis of acidocin B, a novel
bacteriocin produced by Lactobacillus acidophilus. Microbiol-
ogy 141, 1629–1635.
Leroy, F., Verluyten, J., Messens, W., De Vuyst, L., 2002. Model-
ling contributes to the understanding of the different behaviour
of bacteriocin-producing strains in a meat environment. Interna-
tional Dairy Journal 12, 247–253.
Martınez, B., Suarez, J.E., Rodrıguez, A., 1995. Antimicrobials
produced by wild lactococcal strains isolated from homemade
cheeses. Journal of Food Protection 58, 1118–1123.
Meghrous, J., Fliss, I., Bouksaim, M., Lacroix, C., 1999. Digox-
igenin-labeled probe for rapid identification of nisogenic Lacto-
coccus lactis strains. FEMS Microbiology Letters 171, 43–48.
Messens, W., Neysens, P., Vansieleghem, W., Vanderhoeven, J., De
Vuyst, L., 2002. Modelling growth and bacteriocin production
by Lactobacillus amylovorus DCE 471 in response to temper-
ature and pH values used for sourdough fermentations. Applied
and Environmental Microbiology 68, 1431–1435.
Mulders, J.W.M., Boerrigter, I.J., Rollema, H.S., Siezen, R.J.,
de Vos, W.M., 1991. Identification and characterization of the
lantibiotic nisin Z, a natural nisin variant. European Journal of
Biochemistry 201, 581–584.
Muller, M.R.A., Rouvet, M., Brassart, D., Bocker, G., Ehrmann,
M.A., Vogel, R.F., 1998. Adhesion of Lactobacillus strains from
cereal fermentations to human intestinal cells. International
Dairy Journal 8, 584.
Muller, M.R.A., Wolfram, G., Stolz, P., Ehrmann, M.A., Vogel, R.F.,

L. De Vuyst et al. / International Journal of Food Microbiology 90 (2004) 93–106106
2001. Monitoring the growth of Lactobacillus species during a
rye flour fermentation. Food Microbiology 18, 217–227.
Nes, I.F., Diep, D.B., Havarstein, L.S., Brurberg, M.B., Eijsink, V.,
Holo, H., 1996. Biosynthesis of bacteriocins in lactic acid bac-
teria. Antonie van Leeuwenhoek 70, 113–128.
Quadri, L.E.N., Sailer, M., Terebiznik, M.R., Roy, K.L., Vederas,
J.C., Stiles, M.E., 1995. Characterization of the protein confer-
ring immunity to the antimicrobial peptide carnobacteriocin B2
and expression of carnobacteriocins B2 and BM1. Journal of
Bacteriology 177, 1144–1151.
Remiger, A., Ehrmann, M.A., Vogel, R.F., 1996. Identification of
bacteriocin-encoding genes in lactobacilli by polymerase
chain reaction (PCR). Systematic and Applied Microbiology
19, 28–34.
Rodrıguez, J.M., Cintas, L.M., Casaus, P., Horn, N., Dodd, H.M.,
Hernandez, P.E., Gasson, M.J., 1995a. Isolation of nisin-produc-
ing Lactococcus lactis strains from dry fermented sausages.
Journal of Applied Bacteriology 78, 109–115.
Rodrıguez, J.M., Cintas, L.M., Casaus, P., Suarez, A., Hernandez,
P.E., 1995b. PCR detection of the lactocin S structural gene in
bacteriocin-producing lactobacilli from meat. Applied and En-
vironmental Microbiology 61, 2802–2805.
Rodrıguez, J.M., Cintas, L.M., Casaus, P., Martınez, M.I., Suarez,
A., Hernandez, P.E., 1997. Detection of pediocin PA-1-produc-
ing pediococci by rapid molecular biology techniques. Food
Microbiology 14, 363–371.
Rodrıguez, E., Martınez, M.I., Medina, M., Hernandez, P.E., Ro-
drıguez, J.M., 1998. Detection of enterocin AS-48-producing
dairy enterococci by dot-blot and colony hybridization. Journal
of Dairy Research 65, 143–148.
Salminen, S., Isolauri, E., Salminen, E., 1996. Clinical uses of pro-
biotics for stabilizing the gut mucosal barrier: successful strains
and future challenges. Antonie van Leeuwenhoek 70, 347–358.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning:
A Laboratory Manual, 2nd ed. Cold Spring Harbor Laboratory
Press, New York, NY.
Sanger, F., Nicklen, S., Couson, A.R., 1977. DNA sequencing with
chain-terminating inhibitors. Proceedings of the National Acad-
emy of Science of the United States of America 74, 5463–5467.
Tahara, T., Kanatani, K., 1996. Isolation, partial characterization
and mode of action of acidocin J1229, a bacteriocin produced
by Lactobacillus acidophilus JCM 1229. Journal of Applied
Bacteriology 81, 669–677.
Tahara, T., Kanatani, K., 1997. Isolation and partial characterization
of crispacin A, a cell-associated bacteriocin produced by Lacto-
bacillus crispatus JCM 2009. FEMS Microbiology Letters 147,
287–290.
Tahara, T., Oshimura, M., Umezawa, C., Kanatani, K., 1996. Iso-
lation, partial characterization, and mode of action of acidocin
J1132, a two-component bacteriocin produced by Lactobacillus
acidophilus JCM 1132. Applied and Environmental Microbiol-
ogy 62, 892–897.
Tahara, T., Yoshioka, S., Utsumi, R., Kanatani, K., 1997. Isolation
and partial characterization of bacteriocins produced by Lacto-
bacillus gasseri JCM-2124. FEMS Microbiology Letters 148,
97–100.
Tichaczek, P.S., Nissen-Meyer, J., Nes, I.F., Vogel, R.F., Hammes,
W.P., 1992. Characterization of the bacteriocins curvacin A from
Lactobacillus curvatus LTH1174 and sakacin P from Lactoba-
cillus sake LTH673. Systematic and Applied Microbiology 15,
460–468.
Vogel, R.F., Knorr, R., Muller, M.R.A., Steudel, U., Ganzle, M.G.,
Ehrmann, M.A., 1999. Non-dairy lactic fermentations: the ce-
real world. Antonie van Leeuwenhoek 76, 403–411.
Ward, L.J.H., Brown, J.C.S., Graham, P.D., 1994. Application of
the ligase chain reaction to the detection of nisin A and nisin Z
genes in Lactococcus lactis subsp. lactis. FEMS Microbiology
Letters 117, 29–34.
Wood, B.J.B., Holzapfel, W.H., 1995. The Genera of Lactic Acid
Bacteria. Blackie Academic & Professional, London, UK.
Zamfir, M., Callewaert, R., Cornea, C.P., Savu, L., Vatafu, I.,
De Vuyst, L., 1999. Purification and characterization of a bac-
teriocin produced by Lactobacillus acidophilus IBB 801. Jour-
nal of Applied Microbiology 87, 923–931.
Zhu, W.M., Liu, W., Wu, D.Q., 2000. Isolation and characterization
of a new bacteriocin from Lactobacillus gasseri KT7. Journal of
Applied Microbiology 88, 877–886.