the journal of biological chemistry vol. 257. … · veolar macrophages produce a nondialyzable...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 257. No. 14, Issue of July 25, pp. 8098-8105. 1982 Printed in U.S.A.
Modulation of Collagen Production Following Bleomycin-induced Pulmonary Fibrosis in Hamsters PRESENCE OF A FACTOR IN LUNG THAT INCREASES FIBROBLAST PROSTAGLANDIN E2 AND CAMP AND SUPPRESSES FIBROBLAST PROLIFERATION AND COLLAGEN PRODUCTION*
(Received for publication, September 4, 1981, and in revised form, January 22, 1982)
Joan G. Clark$, Katherine M. Kostal, and Beverly A. Marino From the Pulmonarv Division. Department of Medicine, Washington University School of Medicine,
I _
St. Louis, Missouri 631 10
To elucidate mechanisms involved in the regulation of lung collagen content we studied hamsters with bleo- mycin-induced pulmonary fibrosis. Lung collagen in this model is increased as the result of greatly increased lung collagen synthesis rates. However, collagen syn- thesis rates are subsequently restored to normal. Ham- ster lung explants from both normal and bleomycin- exposed hamsters were cultured, and the effects of explant conditioned medium (CM) on lung fibroblast (IMR-90) proliferation and collagen production in ui tm were determined. Lung explant CM increased fibroblast prostaglandin (PG)E2 production and intracellular CAMP, and decreased both fibroblast proliferation and collagen production in a dose-dependent manner. Greater activity was observed with lung explant CM from bleomycin-exposed lungs. Incubation of fibro- blasts with indomethacin prior to addition of CM blocked CM-mediated changes in PGEa and CAMP and inhibited changes in fibroblast proliferation and colla- gen production. Exogenous PGEz or dibutyryl CAMP also suppressed fibroblast proliferation and collagen production. The suppressive activity in lung-condi- tioned medium is nondialyzable, has an apparent mo- lecular weight of 15,000-20,000 by gel filtration, and is heat-stable. It is not species-restricted since CM from hamster lung affected human and hamster lung fibro- blasts similarly. Activity is present preformed in lung and bronchoalveolar lavage fluid, although bronchoal- veolar macrophages produce a nondialyzable factor in culture which suppresses fibroblast proliferation. The suppressive activity identified in fibrotic lung may rep- resent a means for limiting collagen accumulation fol- lowing lung injury.
Diffuse interstitial pulmonary fibrosis results from various injuries to the lung, although the cause is frequently unknown. Regardless of etiology, the disease is characterized physiolog- ically by diminished lung volumes and compromised gas ex-
* This work was supported by United States Public Health Service Grants HL16118 and HL07317 from the National Institutes of Health. A preliminary report of these studies was presented at the National Meeting of the American Federation for Clinical Research, San Fran- cisco, CA, April, 1981 ((1981) CZin. Res. 29, 444). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby m.arked “aduer- tisernent” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom requests for reprints should be addressed at: Pulmonary Division, Department of Medicine, Washington University School of Medicine, 660 South Euclid Ave., St. Louis, MO 63110.
change, and histologically by increased numbers of inflam- matory cells and collagen. Pulmonary fibrosis may progress rapidly, remain stable for variable times, or even resolve (1-4). Biochemical correlates have been difficult to obtain because noninvasive methods are not available to study biochemical events in the lung. However, postmortem studies have dem- onstrated increased total lung collagen 10-20 days after the onset of acute, severe respiratory failure (5), suggesting in- creased lung collagen synthesis. Despite biopsy evidence of fibrosis, some patients with this disorder survive and recover pulmonary function, indicating that the fibrotic process can be modified (4). In idiopathic pulmonary fibrosis, collagen concentration was not increased in lung biopsy specimens (6), but total lung collagen could not be evaluated. Collagen synthesis rates in lung tissue explants from these patients were usually normal (6). These observations suggest that collagen synthesis rates may change during the course of pulmonary fibrosis, but mechanisms responsible for these changes are unknown.
Bleomycin-induced pulmonary fibrosis in hamsters has his- tological and physiological similarities to interstitial pulmo- nary fibrosis in humans (7-9). Lung collagen content increases to twice control values 3 weeks after intratracheal bleomycin (9-11). This increased collagen results from increased synthe- sis rates occurring 1 to 2 weeks after bleomycin injection (11). However, synthesis rates soon decline (ll), becoming identical with control values at 10 weeks.’ The biochemical mechanisms by which collagen synthesis is restored to normal could oper- ate at two levels. First, the number of collagen-producing cells, primarily fibroblasts, could change. Second, the production of collagen per cell could be modified. Both fibroblast prolifera- tion and collagen synthesis rates can be altered in vitro by culture conditions (12, 13), drugs (14), hormones (151, and biological products from other cell types (16-19). The sup- pressive effect of dibutyryl cAMP on cell growth (20) and collagen production (21) is of special interest since changes in intracellular cAMP effected by changes in prostaglandin pro- duction could be a common pathway for modulation of fibro- blast proliferation and collagen synthesis.
In this study, we have attempted to determine if the decline in collagen synthesis following bleomycin administration is related to changes in fibroblast proliferation and/or collagen production and whether these effects are mediated by pros- taglandin or CAMP. Hamster lung explants obtained after bleomycin injection were cultured, and the effects of this lung- conditioned medium upon fibroblast proliferation and colla- gen production were determined. We found a potent factor in
’ J. G. Clark, K. M. Kostal, and B. A. Marina, unpublished obser- vation.
8098

Modulation of Collagen Production in Pulmonary Fibrosis 8099
lung that suppresses both fibroblast proliferation and collagen production. This dual suppression is mediated, in part, by alterations of fibroblast endogenous PGEz' production and intracellular CAMP levels. These findings represent a poten- tially important mechanism for the regulation of collagen synthesis in lung.
EXPERIMENTAL PROCEDURES
Bleomycin Treatment-Ten-week-old male Syrian hamsters re- ceived a single intratracheal dose of 1 unit of bleomycin (Blenoxane; Bristol Laboratories, Syracuse, NY) in 0.1 ml of sterile physiologic saline. Age-matched, uninjected hamsters served as controls.
Conditioned Medium from Lung Cultures-At intervals after bleo- mycin administration, animals were anesthetized with thiamylal so- dium. The lungs were excised, dissected free of large bronchi and blood vessels, and minced into sections (1 to 2 mm) with scalpels. The entire lung mince was suspended in 10 ml of DMEM containing 10% fetal calf serum, 200 units/ml of penicillin, 200 pg/ml of strepto- mycin (basal medium), and 50 pg/ml of ascorbic acid in 25-ml Erlen- meyer flasks. The cultures were equilibrated with 95% 0 2 , 5% COZ and incubated at 37 "C. After 1 h the medium was replaced with 10 ml of fresh medium, and the lung mince was incubated an additional 6 h. The CM was removed and stored at -20 "C. In some experiments the final incubation time was varied to study the time course of activity appearance in the medium. To determine if protein synthesis was required for the appearance of suppressive activity, some explants were frozen at -20 "C prior to incubation. Thawed tissue from these experiments was incubated for 6 h in basal medium. The incorporation of nondialyzable ['Hlproline into the tissue (11) was used to assess protein synthetic activity.
Conditioned Medium from Bronchoalveolar Macrophage Cul- tures-EAMs were obtained by bronchoalveolar lavage of anesthe- tized hamsters with an 18-gauge intratracheal catheter using 10 ml of 0.9 g/dl of NaCl containing 3 mM Na2EDTA. The lavage fluid was centrifuged at 250 X g for 15 min at 25 "C, and the cell pellet was suspended in DMEM and recentrifuged. Cells (>90% BAM) were suspended in DMEM at 5 X IO6 cells/ml, and 5 X IO5 cells were plated in 1.0 ml of DMEM in 1.7-cm diameter multiple well culture plates (Multiwell Tissue Culture Plate, Falcon Plastic, Oxnard, CA). After incubation for 1 h at 37 "C in 5% Con, 95% air atmosphere, cell layers were rinsed once with DMEM to remove nonadherent cells, and adherent cells were cultured in basal medium. CM was removed after 1,2, 4, 6, or 24 h and stored at -20 "C. To determine if BAM release of suppressive activity was constant, BAMs were cultured for 24 h, and the medium was replaced with fresh medium which was collected 6 h later.
Lavage Fluid-Hamster lungs were lavaged and cells were re- moved by centrifugation as described above. Cell-free lavage fluid was dialyzed for 24 h against three changes of DMEM (100x volume) at 4 "C.
Fibroblast Cultures-Human fetal lung fibroblasts (IMR-90, Insti- tute for Medical Research, Camden, NJ) were plated at 3 X lo4 cells/ 1.7-cm diameter well in 1 ml of basal medium and incubated in a 5% COZ, 95% air atmosphere at 37 "C. Cells were in the tenth to fifteenth passage (1:4 split ratio).
Hamster lung fibroblasts were cultivated from 10-week-old male Syrian hamsters. Lungs were removed aseptically and finely minced with two scalpels. The tissue was allowed to adhere to plastic flasks (T-25, Falcon Plastics) for 6 h, and the cultures were incubated in DMEM containing 20% fetal calf serum, penicillin, and streptomycin, as described above. After 1 week, fibroblast outgrowth was visible, and the medium was changed twice weekly until the cells were confluent. The cells were then removed by trypsin and subcultivated in T-75 or T-25 flasks. After reaching visual confluence, cells were removed by trypsin and subcultivated at a 1:3 ratio in 24-well plates. By the second passage, only fibroblastic cells were visible. Hamster lung fibroblasts obtained by this method required 2 to 3 weeks to reach visual confluence, while IMR-90 fibroblasts plated at the same density were confluent in less than 1 week. Because of their slow growth, hamster lung fibroblasts were plated at high density and used in the second to third passage. Unless otherwise specified, we used IMR-90 cells, referred to as fibroblasts.
When fibroblasts were nearly confluent (usually 5 days after pas-
2The abbreviations used are: PG, prostaglandin; DMEM, Dul- becco's modified Eagle's medium; CM, conditioned medium; BAM, bronchoalveolar macrophage.
sage), varying amounts of CM from cultured lung explants or BAM were added with fresh basal medium to a total volume of 500 pllwell. In some experiments, CM was dialyzed (M, cutoff = 12,000, Spectra- por, Los Angeles, CA) against three changes of DMEM (100X volume) for 36 h at 4 "C before addition to the fibroblast cultures. CM was filter-sterilized (0.2-pm pore size) prior to addition to the fibroblast cultures. Fibroblasts were then incubated in the presence of CM for 24 h, and proliferation and collagen synthesis were assayed as below.
Fibroblast Proliferation Assay-After incubation of the fibro- blasts with CM, 1 pCi of ['HIdThd (25 Ci/mol, Amersham Corp.) in 100 pl of basal medium was added to each well, and the incubation was continued for an additional 4 h. The labeling medium was removed, the cell layer was rinsed with cold (4 "C) Hanks' balanced salt solution, and cold trichloroacetic acid (1 ml, 5% w/v) was added to the cell layer. After 30 min at 4 "C, trichloroacetic acid was removed and replaced with 300 p1 of 0.2 M Na2C03 in 0.1 M NaOH. One hundred pl of 50% trichloroacetic acid were added, and ["HldThd in the solution was then determined in triplicate by liquid scintillation spectroscopy. In some experiments cell layer protein was measured as an index of cell number (22). Cell counts of trypsinized cell layers were performed with a hemocytometer. Cell viability was determined by trypan blue exclusion.
Fibroblast Collagen Production Assay-After incubation of the fibroblasts with CM, 10 pCi of ['Hlproline (130 Ci/mol, Amersham Corp.) in 100 pl of basal medium containing ascorbic acid and p- aminopropionitrile to yield final concentrations of 50 pg/ml and 25 pg/ml was added to each well, and the incubation was continued for an additional 6 h. The medium was removed and combined with a 500-pI Hanks' balanced salt solution wash of the cell layer. The cell layer was scraped into 1 ml of 0.5 M acetic acid. After the addition of protease inhibitors (IO mM N-ethylmaleimide, 20 mM disodium EDTA, and 0.3 mM phenylmethylsulfonyl fluoride in final concentra- tion), the medium and cell layer were dialyzed separately for 24 h against running tap water, hydrolyzed in 6 N HCl at 110 "C for 18 h, evaporated, and dissolved in 0.2 M sodium citrate buffer, pH 2.2. ["HI Hydroxyproline and ['Hlproline were separated chromatographically, as previously described (14). Since ["Hlhydroxyproline in medium was proportional to total ['H]hydroxyproline (see below), measure- ments were made on medium alone unless otherwise indicated.
In some experiments, medium 'H-proteins were digested with bacterial collagenase (type VI, Sigma), purified, and examined for noncollagen protease activity as described by Peterkovsky and Die- gelmann (23). Proline specific activity in fibroblast cell layers was measured by a modification of the method of Troll and Lindsley (24). In these experiments, the fibroblasts were cultured in P-100 culture dishes (Falcon Plastics) with 7 ml of medium.
Fibroblast PGE2 Production-PGE, in fibroblast culture medium was measured after exposure to CM. Medium was removed, frozen at
TABLE I Suppression of lung fibroblast proliferation and collagen
production by lung explant CM obtained a t intervals after intratracheal bleomycin
Fibroblasts were cultured with CM (5% concentration) from normal hamster lung explants or bleomycin lung explants obtained at varying intervals after bleomycin instillation. After 24-h incubation with CM, ["HIdThd incorporation or [3H]hydroxyproline production was meas- ured. Values are mean f S.D., n = 3 or 4 cultures. All values are significantly different from cultures containing no CM (p < 0.01). Per cent suppression indicated in parentheses is (basal medium value - experimental value) X 100/basal medium value.
Time after bleomycin ["HIdThd incorpora- ['HIHydroxypro- ~ ~ ~...
tlon line production
days
Basal medium (no CM) 3.33 f 0.08 118 f 5 0 (uninjected) 4 5 6 8
1.63 & 0.21 (51) 56 f 8 (52)
2.36 f 0.12" (29) 64 f 8 (46) 1.67 f 0.24 (50) 1.24 f 0.18* (62) 43 f 3' (64)
15 0.98 f 0.04" (70) 47 f 3" (60)
2.64 f 0.18" (21) 59 f 7 (50)
12 1.22 f 0.12* (64) 46 f 3* (61)
plant CM (p < 0.01).
plant CM (p < 0.05).
a Significantly different from uninjected, normal hamster lung ex-
* Significantly different from uninjected, normal hamster lung ex-

8100 Modulation of Collagen Production in Pulmonary Fibrosis
-70 "C, and assayed within 1 week after obtaining the samples. Radioimmunoassay was performed as described by Dray et al. (25), using antibody specific for PGEZ (Institute Pasteur Production, Manes-la-Coquette, France), ['H]PGE2 probe (170 Ci/mM; New Eng- land Nuclear), and authentic PGE2 standard (The Upjohn Go.). Assays of each culture were performed in triplicate.
Fibroblast CAMP Levels-After incubation of fibroblasts with CM, medium was removed, the cell layer rinsed once with Hanks' balanced salt solution, and ice-cold 5% (w/v) trichloroacetic acid (1.0 mi) added. After 30 min at 4 "C, the trichloroacetic acid supernatant was removed and extracted three times with 5 ml of anhydrous ether. The aqueous phase was lyophilized, and CAMP content was measured in duplicate by radioimmunoassay (Becton-Dickinson, Orangeburgh, NY).
Physical Properties of CM-Heat stability of suppressive activity in CM was assessed by incubating CM at 56 or 80 "C for 1 h prior to incubation with cultured fibroblasts.
CM was fractionated on a column (40 X 2.5 cm) of Sephadex G75 (Pharmacia Fine Chemicals, Piscataway, NJ) calibrated with molec- ular weight standards and equilibrated with 0.5 M acetic acid. Di- alyzed CM or basal medium (3 ml) was lyophilized and resuspended in water at one-third the original volume. Concentrated CM or basal
0
FIG. 1. Effect of CM on [3H]dThd incorporation by lung fi- broblasts. Fibroblasts (IMR-90) were incubated for 24 h with varying concentrations of CM from normal lung or bleomycin-treated lung (11 days after bleomycin). ['HIdThd (1 pCi) was added to each culture, the cultures were incubated an additonal4 h, and ['HIdThd incorporated into the cell layer was measured. CM from normal lung (0) suppressed fibroblast ["HIdThd incorporation in a dose-depend- ent manner. CM from bleomycin-treated lung (0) in equivalent concentration suppressed ['HIdThd incorporation to a greater degree. Values are mean f. S.D., n = 3.
TABLE I1 Effect of CM on cell number und cell layer protein in fibroblast
cultures Subconfluent fibroblasts were cultured in basal medium (-CM) or
in 10% CM (+CM) from bleomycin-exposed lung explants obtained 15 days after bleomycin instillation. At the indicated times, cells were counted or cell Iayer protein measured. Values are mean 2 S.D., n = 3 cultures.
Duration of culture
Cell number Cell layer protein
-CM +CM -CM +CM h Io"/culture pg/cuZture 0 0.93 & 0.14 197 -C 25
24 48 4.59 k 0.60 3.33 k 0.15" 371 & 38 285 & 31"
1.66 & 0.36 1.50 f 0.36 236 k 22 195 f 60
" Significantly different from cultures in basal medium after the same duration of culture ( p < 0.05).
CM CONCENTRATION (Yo)
FIG. 2. Effect of CM on [3H]hydroxyproline production by lung fibroblasts. Fibroblasts (IMR-90) were incubated for 24 h with varying concentrations of CM from normal lung or bleomycin-treated lung (11 days after bleomycin). ['HIProline (10 pCi) was added to each culture, the cultures were incubated an additional 6 h, and nondialyzable ["Hlhydroxyproline in the medium was measured. CM from normal lung (0) suppressed fibroblast ['H]hydroxyproline pro- duction in a dose-dependent manner. CM from bleomycin-treated lung (8) in equivalent concentration suppressed ['HH]hydroxyproline production to a greater degree. Values are mean f S.D., n = 3.
TABLE I11 Effect of CM on proline specific activity, prolyl hydroxylation, and
collagen and noncollagenproteinproduction in lung fibroblast cultures
Fibroblasts were cultured in basal medium or CM (10% concentra- tion) from hamster lung explants obtianed 11 days after bleomycin. After 24 h (except as noted), ["Hlproline was added for 6 h. Parame- ters of collagen production were measured as described under "Experimental Procedures." Values are mean f S.D., n = 3 cultures.
CM treatment
[3H]Proline specific activity (cpm X 10-~&g proline)
Prolyl hydroxylation (96)' 30-h exposure to CM 44-h exposure to CM
[3H]Hydroxyproline (cpm X
Collagen productione r3H]Proline (cpm X 10-4/cul-
Noncollagen protein produc-
Per cent coliaeen aroduction*
lo-'))/culture"
ture)'
tiorP
None Bleomvcin lung
2.70 f 0.37
39.5 & 2.1 38.0 f 2.3 4.30 f 0.14
4.03 f 0.13 6.26 & 0.37
23.2 & 1.4
6.9 k 0.53
2.03 +- 0.23"
39.6 2 1.4 26.7 +- 0.7' 2.18 f 0.48"
2.71 f. 0.50' 2.53 f 0.76'
12.5 f 3.7'
8.7 j; 0.71" ~~ ~
" Significantly different from untreated cultures ( p < 0.05). ' ['H]Hydroxyproline in collagenase-sensitive protein X 100 + total
' Significantly different from untreated cultures ( p < 0.01). " Nondialyzable C3H]hydroxyproline in cells and medium.
radioactivity in collagenase-sensitive protein.
['HlHydroxyproline + % hydroxylation + proline specific activity;
Nondialyzable ["Hlproline in cells and medium. C3H]Proline + proline specific activity; units are arbitrary. (Nondialyzable [3H]hydroxyproline + nondialyzable ["Hlproline)
units are arbitrary.
x loo.
Modulation of Collagen Production in Pulmonary Fibrosis 8101
medium was chromatographed, and pooled fractions were lyophilized and resuspended in 6 ml of basal medium. Aliquots (500 pl) of the pooled fractions were then added to fibroblast cultures.
RESULTS
Effect of Lung CM on Fibroblast Proliferation and Colla- gen Synthesis-CM from both normal hamster lung cultures and bleomycin-induced fibrotic lung suppressed both r3H] dThd incorporation and [3H]hydroxyproline production by cultured IMR-90 fibroblasts (Table I). CM from lung obtained 4 or 5 days after bleomycin instillation suppressed fibroblast r3H]dThd incorporation and [3H]hydroxyproline significantly less than normal lung CM. However, CM from lung obtained 8 days or more after bleomycin injection suppressed r3H]dThd incorporation and ['H]hydroxyproline synthesis to a greater degree than normal lung CM. To explore the mechanisms of the suppression of fibroblast proliferation and collagen syn-
TABLE IV Effect of CM on hamster lung fibroblast proliferation and collagen
production Hamster lung fibroblasts were cultured for 24 h with basal medium
or CM from normal hamster lung explants or bleomycin-exposed lung explants obtained 15 days after bleomycin instillation. ['HIdThd incorporation or [3H]hydroxyproline production was measured as described under "Experimental Procedures." Values are mean f S.D., n = 3 cultures.
CM treatment ['HIdThd incorpora- ['H]Hydroxyproline tion Drodurtion
None 6.37 f 0.94 Normal lung CM
0.57 f 0.04
1 .O% 2.51 f 0.16" 0.46 f 0.03' 10.0% 1.12 f 0.20" 0.57 C 0.10
1.08 1.88 f 0.39" 0.44 f 0.05' 0.44 f 0.25" 0.42 f 0.05'
Bleomycin lung CM
10.0% " Significantly different from cultures in basal medium ( p < 0.01). * Significantly different from cultures in basal medium ( p < 0.05).
~ ~
(24h) (30h) 3.3 6.5
thesis by CM from lung cultures, we studied in detail the effects of lung CM obtained from normal hamsters and ham- sters 11 to 15 days after bleomycin injection, a time when suppressive activity was increased compared to controls.
Suppression of Fibroblast ProEiferation by Lung CM-The suppression of fibroblast proliferation by CM was dose-de- pendent (Fig. I), although increases in CM concentration above 25% resulted in relatively little additional suppression. CM obtained from bleomycin-treated lung consistently re- sulted in greater suppression of fibroblast proliferation than CM.from normal lung. Moreover, very small amounts of CM significantly suppressed fibroblast proliferation.
Fibroblast viability (>go%) was not different from control cultures after 24- to 48-h exposure to CM at concentrations as high as 50%. Smaller increases in cell number and cell layer protein were observed after 48 h of incubation with CM than in untreated cultures (Table 11). The suppressive effects of CM on proliferation were reversible as fibroblasts regained
TABLE V Effect of CM on fibroblast PGEz production and intracellular
CAMP concentration Fibroblasts were incubated for 24 h with basal medium or varying
concentrations of CM from normal lung explants or bleomycin-ex- posed lung explants obtained 11 days after bleomycin. Intracellular cAMP and PGEz in medium were measured by radioimmunoassay. Values are mean f S.D., n = 3 cultures.
CM concentration PGE2 CAMP 4% pg/culture pmol/culture
None 21 r 1 1.82 f 0.44 Normal lung CM
60 f 13" 2.16 f 0.01 184 f 10" 2.41 f O.lOh
10.0 7400 f 830" 5.30 -t 0.31"
0.1 278 f 26" 1.0
2.59 -t 0.06'
10.0 4.05 -t 0.50"
0.1 1.0
Bleomycin lung CM
2800 & 298" 8200 f 743" 5.60 2 0.71"
" Significantly different from cultures in basal medium (p < 0.01). ' Significantly different from cultures in basal medium ( p < 0.05).
20.9 (48h)
0.20) I r I
I- o:
1 I I
12 24 48
DURATION OF EXPOSURE TO CM (hours)
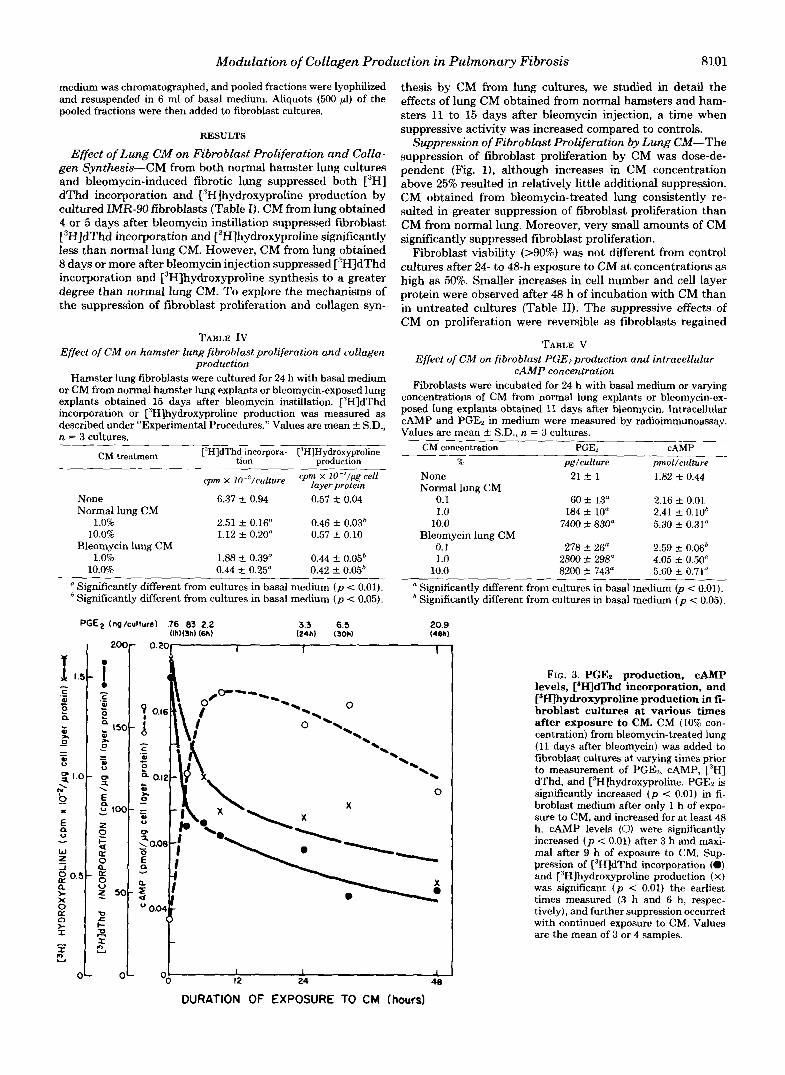
FIG. 3. PGEz production, cAMP levels, C3H]dThd incorporation, and [3H]hydroxyproline production in fi- broblast cultures at various times after exposure to CM. CM (10% con- centration) from bleomycin-treated lung (11 days after bleomycin) was added to fibroblast cultures at varying times prior to measurement of PGE,, CAMP, ["HI dThd, and [3H]hydroxyproline. PGEz is significantly increased ( p < 0.01) in fi- broblast medium after only 1 h of expo- sure to CM, and increased for at least 48 h. cAMP levels (0) were significantly increased ( p < 0.01) after 3 h and maxi- mal after 9 h of exposure to CM. Sup- pression of ["HIdThd incorporation (0) and [3H]hydroxyproline production (X) was significant ( p < 0.01) the earliest times measured (3 h and 6 h, respec- tively), and further suppression occurred with continued exposure to CM. Values are the mean of 3 or 4 samples.

8102 Modulation of Collagen Production in Pulmonary Fibrosis
their ability to proliferate after removal of CM. For example, proliferation of fibroblasts continuously exposed to CM for 24 h exhibited 82% suppression of r3H]dThd incorporation, while proliferation of cells exposed to CM for 8 h, rinsed, and cultured in fresh basal medium for an additional 16 h was suppressed only 51%.
Suppression of Fibroblast Collagen Production by Lung CM-CM caused a dose-dependent decrease in nondialyzable
TABLE VI Effect of exogenous PGEz and dibutyryl CAMP on lung fibroblasts
Lung fibroblasts (IMR-90) were incubated for 24 h with varying amounts of PGE:! or dibutyryl cAMP in 500 pl of basal medium. ["HI dThd incorporation, [3H]hydroxyproline production, or cAMP was measured as described under "Experimental Procedures." Values are mean f S.D.. n = 3 cultures.
Treatment [iH]dThd in- [ 'HIHydrox~- corporation proline pro- cAMP
duction
PGEz (ng/ml) 0 0.5 5.0
50.0 500.0
0 1 5
Dibutyryl cAMP (mM)
cpm X lo-.'/ culture
16.7 f 0.29 7.80 f 0.20" 3.40 f 0.15" 2.12 f 0.04" 1.88 f 0.04"
13.3 f 0.07 4.90 f 0.23" 0.73 f 0.05"
cpm X lo+/ pg cell layer pmol/culture
protein
1.12 f 0.18 2.14 1- 0.15 2.50 1- 0.34 3.02 -C 0.736
0.30 & 0.03' 2.68 C 0.17b 0.27 f 0.02* 3.27 +- 0.57*
1.19 & 0.26 0.92 f 0.05 0.44 k 0.09"
~~
Significantly different from untreated cultures ( p < 0.01). Significantly different from untreated cultures ( p < 0.05).
c
I
0 -
0 -
0 -
I I
I
I I
I
I
NONE DETECTED C
[3H]hydroxyproline in fibroblast medium (Fig. 2). The de- crease was greater in cultures exposed to bleomycin lung CM than in cultures exposed to normal lung CM. The threshold for detection of normal lung CM activity was 1076, and for bleomycin lung CM, 1%.
[3H]Hydroxyproline in medium was proportional to total [3H]hydroxyproline in the cultures under both control and experimental conditions. Medium from control cultures with no CM contained 74.7 f 4.9% of the [3H]hydroxyproline; normal lung CM-exposed (10% concentraion cultures, 73.7 & 12.8%; and bleomycin lung CM-exposed cultures, 74.0 k 6.3%. We expressed ["H]hydroxyproline/pg of cell layer protein to reflect cell number.
Table 111 presents the data required for a precise calculation of collagen production (26, 27). CM decreased [3H]proline specific activity, reflecting an increased free proline pool fol- lowing CM. Prolyl hydroxylation of newly synthesized colla- gen was not affected after 30-h incubation with CM, but decreased significantly after 44 h. Collagen production and noncollagen protein production were both reduced by approx- imately 50%.
Effect of CM on Hamster Lung Fibroblast Proliferation and Collagen Production-Hamster lung fibroblast response to CM exposure was similar to that of human lung fibroblasts. Both [3H]dThd incorporation and nondialyzable medium ["HI hydroxyproline were suppressed by CM from normal lung and bleomycin-exposed lung (Table IV).
Effect of CM on Fibroblast Endogenous PGE2 Production and Intracellular cAMI"PGE2 levels in medium after 24-h incubation increased strikingly with increasing concentration of CM (Table V). Prostaglandin production was significantly
cAMP
L
20
I "
-IN00 +INDO 2
WOLIFERATION
T
" -IN00 +IN00
COLLAGEN SYNTHESIS
T
" -INDO +INDO
FIG. 4. Effect of indomethacin on CM activity. Fibroblasts exposure resulted in increased PGEL and cAMP and decreased ["HI were incubated 2 h with indomethacin (INDO, 5 pg/ml). Basal me- dThd incorporation and ["Hlhydroxyproline production. When the dium (hatched bars) or CM at 10% concentration (normal lung CM, fibroblasts were pretreated with indomethacin, the CM-mediated open bars; bleomycin lung CM, solid bars) was added for an addi- increase in PGE, and cAMP was blocked, and suppression of ["HI tional 24 h. PGE2 production, CAMP, ['HldThd incorporation, and dThd incorporation and ['H]hydroxyproline production was in- ['H]hydroxyproline production were measured as described under hibited. The comparisons are between cultures with the same CM "Experimental Procedures." In the absence of indomethacin, CM treatment, with and without indomethacin ( * p < 0.05; **p < 0.01).
Modulation of Collagen Production in Pulmonary Fibrosis 8103
greater in fibroblast cultures exposed to bleomycin lung CM than in cultures exposed to normal lung CM at concentrations of 0.1 and 1.0%. Preliminary studies showed that conditioned medium and basal medium contained PGE2. Although this amount was small (about 20 pg/ml in basal medium and 100 pg/ml in 10% CM from bleomycin lung) compared to the amounts observed after incubation with fibroblasts, the data were corrected for this exogenous PGEz.
Fibroblasts incubated with CM for 24 h also had significant increases in cAMP compared to controls (no CM) (Table V). Concentrations of bleomycin lung CM as low as 0.1% stimu- lated CAMP. CM from normal lung stimulated cAMP at 1.0% concentration.
Changes in all four parameters studied are related to dura- tion of fibroblast exposure to CM in Fig. 3. PGE, was present in fibroblast culture medium after 1 h and increased during the entire 48-h incubation. The level of PGEz present in the fibroblast medium after only 1 h is similar to an amount of exogenous PGE2 that significantly decreased ["HIdThd incor- poration by fibroblasts (Table VI). Intracellular cAMP was significantly increased after 3 h and maximal a t 9 h; thereafter, the level of CAMP decreased, but remained elevated compared to control cultures (t = 0 h). The suppression of ["HIdThd incorporation and ["H]hydroxyproline production was signif- icant at the earliest times measured, i.e. 3 h and 6 h, respec- tively. Additional suppression was noted with continued in- cubation with CM.
Pretreatment of the cultured fibroblasts for 2 h with indo- methacin (5.0 pg/ml) blocked CM-mediated increases in PGE, production and intracellular cAMP levels and partially pre- vented the CM-mediated suppression of ['HIdThd incorpo- ration and ['H]hydroxyproline synthesis (Fig. 4). Indometha- cin concentrations ranging between 0.1 and 10.0 m g / d had the same quantitative effect on ['HIdThd incorporation and ['Hlhydroxyproline production. The effect of CM concentra- tions of 1.0% was also only partially inhibited by indomethacin (data not shown). Indomethacin treatment alone resulted in changes in PGEz, fibroblast CAMP, ["HIdThd incorporation, and ["Hlhydroxyproline production. However, the per cent
BLUE DEXTRIN ALBUMIN
I 1 LrrOZYUE
I PHENOL REO
ELUTION VOLUME (ml)
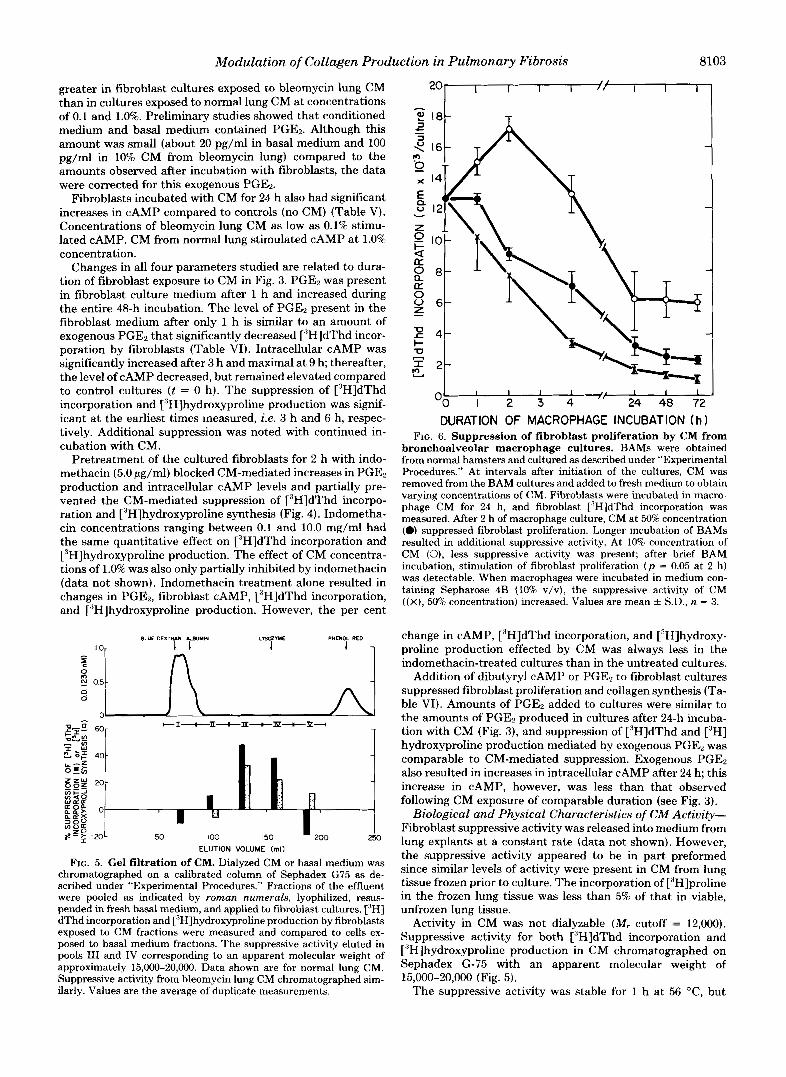
FIG. 5. Gel filtration of CM. Dialyzed CM or basal medium was chromatographed on a calibrated column of Sephadex G75 as de- scribed under "Experimental Procedures." Fractions of the effluent were pooled as indicated by roman numerals, lyophilized, resus- pended in fresh basal medium, and applied to fibroblast cultures. ['HI dThd incorporation and ['H]hydroxyproline production by fibroblasts exposed to CM fractions were measured and compared to cells ex- posed to basal medium fractions. The suppressive activity eluted in pools I11 and IV corresponding to an apparent molecular weight of approximately 15,000-20,000. Data shown are for normal lung CM. Suppressive activity from bleomycin lung CM chromatographed sim- ilarly. Values are the average of duplicate measurements.
DURATION OF MACROPHAGE INCUBATION (h 1 FIG. 6. Suppression of fibroblast proliferation by CM from
bronchoalveolar macrophage cultures. BAMs were obtained from normal hamsters and cultured as described under "Experimental Procedures." At intervals after initiation of the cultures, CM was removed from the BAM cultures and added to fresh medium to obtain varying concentrations of CM. Fibroblasts were incubated in macro- phage CM for 24 h, and fibroblast ["HIdThd incorporation was measured. After 2 h of macrophage culture, CM at 50% concentration (0) suppressed fibroblast proliferation. Longer incubation of BAMs resulted in additional suppressive activity. At 10% concentration of CM (0), less suppressive activity was present; after brief BAM incubation, stimulation of fibroblast proliferation ( p = 0.05 at 2 h) was detectable. When macrophages were incubated in medium con- taining Sepharose 4B (10% v/v), the suppressive activity of CM ( ( X ) , 50% concentration) increased. Values are mean k S.D., n = 3.
change in CAMP, ["HIdThd incorporation, and ['Hlhydroxy- proline production effected by CM was always less in the indomethacin-treated cultures than in the untreated cultures.
Addition of dibutyryl cAMP or PGE2 to fibroblast cultures suppressed fibroblast proliferation and collagen synthesis (Ta- ble VI). Amounts of PGEz added to cultures were similar to the amounts of PGEz produced in cultures after 24-h incuba- tion with CM (Fig. 3), and suppression of ['HIdThd and ["HI hydroxyproline production mediated by exogenous PGEB was comparable to CM-mediated suppression. Exogenous PGE2 also resulted in increases in intracellular cAMP after 24 h; this increase in CAMP, however, was less than that observed following CM exposure of comparable duration (see Fig. 3).
Biological and Physical Characteristics of C" Activity- Fibroblast suppressive activity was released into medium from lung explants at a constant rate (data not shown). However, the suppressive activity appeared to be in part preformed since similar levels of activity were present in CM from lung tissue frozen prior to culture. The incorporation of ['Hlproline in the frozen lung tissue was less than 5% of that in viable, unfrozen lung tissue.
Activity in CM was not dialyzable (M, cutoff = 12,000). Suppressive activity for both ['HIdThd incorporation and ['H]hydroxyproline production in CM chromatographed on Sephadex G-75 with an apparent molecular weight of 15,000-20,000 (Fig. 5).
The suppressive activity was stable for 1 h at 56 "C, but

8104 Modulation of Collagen Production in Pulmonary Fibrosis
reduced 50% by incubation at 80 “C for 1 h. Repeated freezing at -20 “C and thawing a t 25 “C (three times) did not alter activity in CM.
Suppressive Activity in Bronchopulmonary Lavage Fluid and BAM Cultures-Fibroblast proliferation was suppressed 35% following incubation with dialyzed cell-free normal lung lavage fluid diluted 1:l with DMEM containing 20% fetal calf serum, while bleomycin-exposed lung lavage similarly diluted resulted in 53% suppression. Macrophages obtained by bron- choalveolar lavage of normal hamsters also produced in cul- ture a nondialyzable inhibitor of fibroblast proliferation. Sup- pressive activity in the medium increased with incubation time and was also dependent on the concentration of CM (Fig. 6). However, we also noted that fibroblast proliferation was stimulated (p = 0.05) by low concentrations of CM (10%) obtained after 2 h of BAM culture. The release of suppressive activity by BAM in culture for 6 h was identical, whether studied immediately after adherence or after being held in culture for 24 h. Incubation of the macrophages with Sepha- rose 4B resulted in increased suppressive activity.
DISCUSSION
A factor that suppresses fibroblast proliferation and colla- gen production is present in conditioned medium from both normal and bleomycin-exposed lung explants. The difference in level of activity in normal and bleomycin-treated lung is likely attributable to concentration, and the potency of CM suggests amplification following interaction of CM with fibro- blasts. Moreover, suppressive activities for both fibroblast proliferation and collagen production have the same biochem- ical characteristics, indicating that a single factor in CM affects a common mechanism in fibroblast proliferation and collagen synthesis.
CM-mediated suppression of proliferation and collagen pro- duction is associated with increased endogenous PGEz pro- duction and intracellular cAMP in proportion to the concen- tration of CM. Several lines of evidence in this study support the conclusion that CM stimulates PGEz synthesis, and PGEz in turn increases CAMP, perhaps by PGEz-mediated stimula- tion of adenylate cyclase (28), with resultant suppression of both fibroblast proliferation and collagen production. 1) Ex- ogenous PGEz or dibutyryl cAMP decreases fibroblast prolif- eration and collagen production, and exogenous PGEz in- creases fibroblast CAMP, 2) within 6 h of CM exposure, endogenous PGEz is increased to levels that increase intracel- lular cAMP and decrease cell proliferation and collagen pro- duction; 3) indomethacin, an inhibitor of prostaglandin syn- thesis, blocks CM-mediated changes in PGE2 and cAMP and partially prevents CM-mediated suppression of fibroblast pro- liferation and collagen production.
While increases in PGEz and cAMP level appear to be important mechanisms, indomethacin, which completely blocks prostaglandin production, incompletely blocks CM-me- diated suppression of fibroblast proliferation and collagen synthesis. It is possible that the small amounts of PG in CM contribute to the suppressive effect, or that CM contains other components with different suppressive mechanisms. Alterna- tively, a single component in CM may suppress by more than one mechanism.
Unlike the brief increase in fibroblast CAMP which follows incubation with exogenous PG (29), cAMP is increased for at least 24 h after CM exposure, This is not unexpected, since PGEz levels are constantly increasing after CM exposure. Some refractoriness to PGEz does occur, since cAMP levels decline after 24 h of CM exposure. However, suppression of fibroblast proliferation and collagen production is sustained even as cAMP levels are decreasing, suggesting that other
mechanisms perhaps not directly involving cAMP also regu- late fibroblast function.
Lung CM decreases fibroblast collagen production, and after extended exposure to CM, newly synthesized collagen is underhydroxylated, perhaps by CM-mediated changes in other proteins such as prolyl hydroxylase. Since underhydrox- ylation increases intracellular degradation of procollagen (30), collagen production could be reduced by this mechanism. Also, collagen produced by fibroblasts incubated with dibu- tyryl cAMP or PGE undergoes increased intracellular degra- dation (29). Such post-translational modulation of collagen production, however, does not exclude control at transcrip- tional or translational levels. In our experiments, both collagen and noncollagen protein synthesis was decreased by CM, but since proline is not equally distributed in proteins, [3H]proline incorporation into collagenase-resistant polypeptides may not be a sensitive measure of specific noncollagen protein. The mechanism and biological specificity of CM-mediated changes in protein synthesis require further study.
Fibroblast suppressive activity is, in part, preformed in lung since it is present in cell-free lavage fluid and extractable from nonviable lung tissue. The alveolar macrophage may be a source of suppressive activity in lung as increased suppressive activity in CM after bleomycin coincides with a mononuclear infiltrate in bleomycin-treated lung (7, 9) and increased yield of BAM in lavage fluid.’ Moreover, suppressive activity for fibroblast proliferation and collagen synthesis has previously been demonstrated in conditioned medium from activated peripheral blood mononuclear cells (16, 17), the presumed precursor of BAM. In this study, cultured hamster BAMs clearly release a nondialyzable product that suppresses fibro- blast proliferation. Studies are currently underway in our laboratory to further characterize suppressive activity from BAM and to define the biochemical mechanisms responsible for its effects.
Under some experimental circumstances, macrophage prod- ucts increase fibroblast proliferation (31, 32), and it is likely that BAM secrete several products that influence fibroblasts differently. In these studies stimulatory activity was detected at low concentrations of CM from freshly adherent BAM, suggesting the presence of a factor that can stimulate fibro- blasts to proliferate. Several explanations may account for the predominance of suppressive activity in our experiments. We utilized culture conditions which are near-optimal for fibro- blast proliferation (12) and collagen synthesis (13) SO that inhibitory activity could be assayed with maximal sensitivity. The culture conditions might be especially relevant to the injured, “leaky” lung in which fibroblasts are exposed to serum products (32), but are likely to underestimate stimula- tory effects. Alternatively, stimulatory substances may be released in a different pattern than suppressive substances SO
that eventually suppression predominates. Also, fibroblast receptors or “receptiveness” to stimulators may be more lim- ited, allowing inhibitors to predominate.
Various stimuli accelerate production of collagen in the lung, but mechanisms which control the total amount of lung collagen are unknown. One mode of collagen synthetic control in the lung is suggested by our observation that conditioned medium from hamster lung explants suppresses fibroblast proliferation and collagen synthesis in vitro. This finding in normal lung explants suggests that fibroblasts in normal lung might be continuously under suppression and maintained in a quiescent state which could account for the very low level of collagen turnover observed in normal lung (33). Fibroblast suppressive activity decreased in the first days after bleomycin instillation and then increased after the fibrogenic process was established (2.e. 8 days after bleomycin), suggesting that levels

Modulation of Collagen Production in Pulmonary Fibrosis 8105
of suppressive activity are also important in the modulation of fibroblast proliferation and collagen production in lung fibrosis.
Although the in vivo significance of these observations remains uncertain, similar changes in levels of fibroblast sup- pressive activity after a fibrogenic event have also been dem- onstrated in other models of fibrosis, including nephritis (34) and granulation tissue formation (35). In addition, Dayer et al. (36) have demonstrated that peripheral blood mononuclear cells produce a factor which suppresses proliferation of cul- tured rheumatoid synovial cells by stimulating increases in PGE2 and CAMP. Collagenase production is also increased by this factor which co-purifies with interleukin 1 (37). A product of mononuclear cells has also been shown by Korn et al. (16) to suppress fibroblast proliferation and concomitantly to in- crease endogenous fibroblast PGE production. Partial inhibi- tion of growth suppression occurred with indomethacin pre- treatment of these fibroblasts. In a similar system, Jimenez et al. (17) showed that mononuclear cell culture supernatants selectively suppressed collagen synthesis. The results of these experiments and our demonstration of similar activity in bleomycin-treated lung suggest that active suppression of fibroblasts may occur in various circumstances and represent an important means for regulation of connective tissue for- mation.
Acknowledgments-We thank Drs. John A. McDonald, Robert P. Mecham, WiUiam Stenson, and Aubrey Morrison for helpful discus- sions and suggestions.
REFERENCES I. Karlinsky, J. B., and Goldstein, R. H. (1980) J. Lab. Clin. Med.
2. Crystal, R. G., Fulmer, J . D., Roberts, W. C., Moss, M. L., Line, B. R., and Reynolds, H. Y. (1976) Ann. Intern. Med. 85,769-788
3. Crystal, R. G., Fulmer, J. D., Baum, B. J., Bernardo, J., Bradley, K. H., Bruel, S. D., Elson, N. A., Fells, G. A,, Ferrans, V. J., Gadek, J. E., Hunninghake, G. W., Kawanami, O., Kalman, J. A., Line, B. R., McDonald, J. A., McLees, B. D., Roberts, W. C., Rosenberg, D. M., Tolstoshev, P., VonGal, E., and Wein- berger, S. E. (1978) Lung 155, 199-224
4. Lamy, M., Fallat, R. J., Koeniger, E., Dietrich, H.-P., Ratliff, J. L., Eberhart, R. C., Tucker, H. J., and Hill, J . D. (1976) Am. Rev. Respir. Dis. 1, 267-284
5. Zapol, W. M., Trelstad, R. L., Coffey, J . W., Tsai, I., and Salvador, R. A. (1979) Am. Rev. Respir. Dis. 119, 547-554
6. Fulmer, J . D., Bienkowski, R. S., Cowan, M. J., Breul, S. D., Bradley, K. M., Ferrans, V. J., Roberts, W. C., and Crystal, R. G. (1980) Am. Rev. Respir. Dis. 122,289-301
7. Snider, G. L., Hayes, J . A., and Korthy, A. L. (1978) Am. Rev. Respir. Dis. 117, 1099-1108
8. Snider, G. L., Celli, B. R., Goldstein, H. H., O’Brien, J. J., and
96,939-942
Lucey, E. C. (1978) Am. Rev. Respir. Dis. 117,289-297
Respir. Dis. 117, 299-305
(1979) Am. Rev. Respir. Dis. 120,67-73
B. C. (1980) J. Lab. Clin. Med. 96,943-953
1976
9. Starcher, B. C., Kuhn, C., and Overton, J . E. (1978) Am. Rev.
10. Goldstein, R. H., Lucey, E. C., Franzblau, C., and Snider, G. L.
11. Clark, J . G., Overton, J . E., Marino, B. A., Uitto, J., and Starcher,
12. Rutherford, R. B., and Ross, R. (1976) J. Cell Biol. 69, 196-203,
13. Booth, B. A,, Polak, K. L., and Uitto, J. (1980) Biochim. Biophys.
14. Clark, J. G., Starcher, B. C., and Uitto, J. (1980) Biochim. Bio-
15. Pastan, I. H., Johnson, G. S., and Anderson, W. B. (1975) Annu.
16. Korn, J. H., Halushka, P. V., and LeRoy, E. C. (1980) J . Clin.
17. Jimenez, S. A,, McArthur, W., and Rosenbloom, J. (1979) J. Exp.
18. Johnson, R. L., and Ziff, M. (1976) J. CEin. Invest. 58,240-252 19. Wahl, S. M., Wahl, L. M., and McCarthy, J . B. (1978) J. Immunol.
20. Johnson, G. S., Friedman, R. M., and Pastan, I. (1971) Proc. Natl.
21. Baum, B. J., Moss, J., Breul, S. D., and Crystal, R. G. (1978) J.
22. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J .
23. Peterkovsky, B., and Diegelmann, R. (1971) Biochemistry 10,
24. Troll, W., and Lindsley, J . (1955) J. Biol. Chem. 215,655-660 25. Dray, F., Charbonnel, B., and Maclouf, J . (1975) Eur. J . CEin.
Invest. 5, 311-318 26. Bradley, K. H., McConnell, S. D., and Crystal, R. G. (1974) J .
Biol. Chem. 249, 2674-2683 27. Breul, S. D., Bradley, K. H., Hance, A. J., Schaffer, M. P., Berg,
R. A,, and Crystal, R. G. (1980) J. Biol. Chem. 255,5250-5260 28. Samuelsson, B., Goldyne, M., Granstrom, S., Hamburg, M., Ham-
marstrom, S., and Malmsten, C . (1978) Annu. Rev. Biochem.
29. Baum, B. J., Moss, J., Breul, S. D., Berg, R. A., and Crystal, R. G.
30. Berg, R. A,, Schwartz, M. L., and Crystal, R. G. (1980) Proc. Natl.
31. Martin, B. M., Gimbrone, M. A., Jr., Unanue, E. R., and Cotran,
32. Liebovich, S. J., and Ross, R. (1976) Am. J. Pathol. 84, 501-514 33. Pierce, J . A., Resnick, H., and Henry, P. H. (1967) J. Lab. Clin.
34. Nelson, E. G., Jimenez, S. A,, and Phillips, S. M. (1980) J.
35. Kulonen, E., and Potila, M. (1980) Exp. Pathol. (Jena) 18,
36. Dayer, J.-M., Goldring, S. R., Robinson, D. R., and Krane, S. M.
37. Mizel, S. B., Dayer, J.-M., Krane, S. M., and Mergenhagen, S. E.
Acta 607, 145-160
phys. Acta 631,359-370
Rev. Biochem. 44,491-522
Invest. 65, 543-554
Med. 150, 1421-1431
121, 942-946
Acad. Sci. U. S. A . 68,425-429
Biol. Chem. 253, 3391-3394
(1951) J. Biol. Chem. 193,265-275
988-994
477,997-1029
(1980) J. Biol. Chem. 255, 2843-2847
Acad. Sci. U. S. A . 77,4746-4750
R. S. (1981) J. Immunol. 126, 1510-1515
Med. 69,485-493
Immunol. 125, 1708-1714
456-461
(1979) Biochim. Biophys. Acta 586,87-105
(1981) Proc. Natl. Acad. Sei. U . S. A. 78, 2474-2477