the journal of biological chemistry printed in … · the von hippel-lindau tumor suppressor...
TRANSCRIPT

The von Hippel-Lindau Tumor Suppressor Stabilizes Novel PlantHomeodomain Protein Jade-1*
Received for publication, May 22, 2002, and in revised form, July 29, 2002Published, JBC Papers in Press, August 6, 2002, DOI 10.1074/jbc.M205040200
Mina I. Zhou‡, Hongmei Wang‡, Jonathan J. Ross§, Igor Kuzmin¶, Chengen Xu‡,and Herbert T. Cohen‡§�
From the Departments of ‡Medicine and §Pathology, Sections of Nephrology and Hematology/Oncology, Boston UniversitySchool of Medicine and Boston Medical Center, Boston, Massachusetts 02118 and ¶NCI, National Institutes of Health,Frederick Cancer Research Center, Frederick, Maryland 21702
The von Hippel-Lindau disease gene (VHL) is the caus-ative gene for most adult renal cancers. However, themechanism by which VHL protein functions as a renaltumor suppressor remains largely unknown. To identifylow occupancy VHL protein partners with potential rel-evance to renal cancer, we screened a human kidneylibrary against human VHL p30 using a yeast two-hybridapproach. Jade-1 (gene for Apoptosis and Differentia-tion in Epithelia) encodes a previously uncharacterized64-kDa protein that interacts strongly with VHL proteinand is most highly expressed in kidney. Jade-1 proteinis short-lived and contains a candidate destabilizing(PEST) motif and plant homeodomains that are not re-quired for the VHL interaction. Jade-1 is abundant inproximal tubule cells, which are clear-cell renal cancerprecursors, and expression increases with differentia-tion. Jade-1 is expressed in cytoplasm and the nucleusdiffusely and in speckles, where it partly colocalizeswith VHL. VHL reintroduction into renal cancer cellsincreases endogenous Jade-1 protein abundance up to10-fold. Furthermore, VHL increases Jade-1 proteinhalf-life up to 3-fold. Thus, direct protein stabilization isidentified as a new VHL function. Moreover, Jade-1 pro-tein represents a novel candidate regulatory factor inVHL-mediated renal tumor suppression.
VHL gene defects are responsible for both von Hippel-Lindaudisease (1) and most sporadic clear-cell renal cancers (2–5).VHL is therefore the most commonly affected renal cancergene. Clear-cell renal cancer is also the most malignant VHL1
disease lesion, which suggests VHL protein exerts strongesttumor suppressor activity in renal proximal tubules, which are
the precursor cells of this common malignancy.VHL disease manifestations, which include retinal angio-
mas; central nervous system hemangioblastomas; renal, pan-creatic, and epididymal cysts; and pheochromocytomas, pan-creatic neuroendocrine tumors, and clear-cell renal cancers,suggest VHL protein has multiple functions (6). VHL binds andpromotes ubiquitination of hypoxia-inducible transcription fac-tors HIF-1� and HIF-2� (7), protein kinase C (PKC) lambda (8),heterogeneous nuclear ribonucleoprotein A2 (9), and VHL-interacting deubiquitinating enzyme-1 (VDU1) (10). VHL in-hibits transcription elongation (11–14), mRNA stability (9,15–17), Sp1-related promoter activity (18, 19), and PKC activ-ity (8, 20, 21). VHL also increases abundance of the directlyinteracting protein fibronectin and promotes its incorporationinto extracellular matrix (22). VHL induces morphogenesis,cellular differentiation, and contact inhibition of renal cancercells or proximal tubule cells (23–27). Like the retinoblastomatumor suppressor, VHL inhibits apoptosis, particularly in re-sponse to cell stresses, such as serum depletion (28), glucosedepletion, endoplasmic reticulum (ER) stress (29), or UV irra-diation (30). VHL functional heterogeneity is further supportedby its residence in cytoplasm (31), the nucleus (32), mitochon-dria (33), ER (34), and perhaps Golgi (22).
VHL functions most important for renal tumor suppressionremain unclear. Based on the well-recognized association ofspecific VHL mutations with partial VHL disease phenotypes,a likely hypothesis is that VHL missense mutations may dis-rupt some, but not necessarily all, VHL functional pathways.For example, VHL mutations that prevent HIF ubiquitinationand therefore promote HIF and vascular endothelial growthfactor overexpression are precisely those that correlate withhemangioblastoma development (35–38). Although HIF over-expression contributes importantly to renal cancer pathogene-sis (39, 40), no VHL biochemical function or protein interactionhas been found that correlates with renal cancer risk, leavingunresolved the full role of VHL in renal tumor suppression (41).
To identify molecules potentially important in the pathogen-esis of clear-cell renal cancer, we screened an adult humankidney library with human VHL p30 using a yeast two-hybridapproach. We have named the gene encoding a novel, strongVHL-interactor as Jade-1 (gene for Apoptosis and Differentia-tion in Epithelia). Jade-1 protein is short-lived and most highlyexpressed in kidney. It contains a candidate PEST degradationdomain and plant homeodomain (PHD) motifs, which are notrequired for the VHL interaction. Jade-1 protein is directlystabilized by VHL protein, which is a new VHL function. Pre-liminary results suggest Jade-1 is also growth suppressive.Jade-1 may therefore participate in VHL-mediated renal tumorsuppression.
* This work was supported by the National Institutes of HealthGrants T32-DK07053 and F32-CA79133 (to M. Z.) and R01-CA79830(to H. T. C.). The costs of publication of this article were defrayed inpart by the payment of page charges. This article must therefore behereby marked “advertisement” in accordance with 18 U.S.C. Section1734 solely to indicate this fact.
The nucleotide sequence(s) reported in this paper has been submittedto the GenBankTM/EBI Data Bank with accession number(s) AF520952.
� To whom correspondence should be addressed: Evans BiomedicalResearch Center, X-535, Boston University Medical Center, 650 AlbanySt., Boston, MA 02118. Tel.: 617-638-7322; Fax: 617-638-7326; E-mail:[email protected].
1 The abbreviations used are: VHL, von Hippel-Lindau; X-gal, 5-bromo-4-chloro-3-indolyl-�-D-galactopyranoside; PKC, protein kinaseC; VDU1, VHL-interacting deubiquitinating enzyme-1; ER, endoplas-mic reticulum; HIF, hypoxia-inducible transcription factor; PHD, planthomeodomain; CMV, cytomegalovirus; HA, hemagglutinin; MPT,mouse proximal tubule; nt, nucleotide(s); aa, amino acid(s); del, dele-tion; dd, double PHD deletion; GST, glutathione S-transferase; FITC,fluorescein isothiocyanate; cy3, cyanine-3; C4HC3, Cys4-His-Cys3.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 42, Issue of October 18, pp. 39887–39898, 2002Printed in U.S.A.
This paper is available on line at http://www.jbc.org 39887
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from

EXPERIMENTAL PROCEDURES
Constructs—Plasmids pFLAG-cytomegalovirus (CMV)-2 VHL andderivatives have been described previously (18, 19). pRc-hemagglutinin(HA)-VHL (42), FLAG-p53 (43), and FLAG-PKC � were generouslyprovided by Drs. W. Kaelin (Dana-Farber Cancer Institute), U. Moll(Stony Brook), and A. Toker (Boston Biomedical Research Institute),respectively. VHL nucleotide (nt) sequence encoding amino acids (aa)2–213 and 55–143 was PCR-cloned into pGilda (CLONTECH) usingEcoRI and SalI sites. A mouse VHL cDNA clone was generously pro-vided by Dr. M. Lerman (NCI, National Institutes of Health, Frederick,MD) and subcloned similarly into pGilda. The Jade-1 5�-untranslatedregion and coding sequence were cut from pB42AD with NotI and XhoIand subcloned into NotI and SalI cut pFLAG-CMV2. The Jade-1 com-plete coding sequence (aa 1–509) and truncations (deletion 1 (del1), aa202–509; deletion 2 (del2), aa 372–509; and double PHD deletion (dd),aa 1–201, 254–311, 372–509) (see Fig. 1, A and B) were PCR-amplifiedand inserted using NotI and XbaI into pcDNA3.1 (Invitrogen) as anuntagged expression vector and into pCR3.1 uni (Invitrogen) that hadbeen modified to contain an HA tag. Deletion of the Jade-1 PHD regionswas performed by recombinant PCR. Additional details about plasmidconstructs can be obtained from the authors.
Yeast Two-hybrid Analysis—An adult human kidney cDNA library inpB42AD (CLONTECH) was screened against human VHL aa 2–213 inthe LexA-expressing, inducible yeast expression vector pGilda (CLON-TECH), according to the manufacturer’s instructions (19). pB42ADlibrary clones initially positive by growth on deficient medium and byX-gal staining were rescued and individually screened against bothhuman and mouse VHL in pGilda in yeast before sequencing. Interac-tion strength in yeast was categorized by the amount of time post-cotransformation required for yeast colonies to appear blue on X-galplates. Clones positive at 24 h were designated as the strongest (3�)interactors. Those positive at 48 or 72 h were designated as 2� or 1�interactors, respectively.
Cell Lines and Transfection—293T17 human embryonic kidney cellsand HT1080 fibrosarcoma cells were generously provided by Drs. Z. Luo(Boston University School of Medicine) and R. Widom (Boston Univer-sity School of Medicine), respectively. 786-O renal cancer cells andHeLa cells were obtained from American Type Culture Collection(Manassas, VA). These lines were grown in Dulbecco’s modified Eagle’smedium supplemented with 10% fetal bovine serum, glutamine, andpenicillin-streptomycin (Invitrogen). HA-VHL 786-O (42) and A498 re-nal cancer lines (44) were generously provided by Drs. W. Kaelin (Dana-Farber Cancer Institute) and W. Krek (Friedrich Miescher Institut,Basel), respectively. 786-O (18, 19) and UMRC6 (45) stably-transfectedrenal cancer lines were described previously. Stable renal cancer lineswere maintained as above in 0.2–0.4 mg/ml G418 (Invitrogen). SV40 Tantigen-transformed mouse proximal tubule (MPT) cells, generouslyprovided by Dr. M. Loghman-Adham (University of Utah) (46), weregrown in the above medium supplemented with 5 units/ml interferon �at 33 °C. MPT cells were differentiated without interferon at 37 °C for10 days. Primary culture mouse proximal tubule cells were generouslyprovided by Dr. W. Lieberthal (Boston University School of Medicine)(47). Cells were transfected using LipofectAMINE 2000 (Invitrogen)according to the manufacturer’s instructions or by calcium phosphateprecipitation (48).
Antibodies—To generate Jade-1 antisera, 2 New Zealand White rab-bits were injected with the same carboxyl-terminal 20 residue Jade-1peptide amino terminally linked to keyhole limpet hemocyanin. Anti-serum 1 was affinity-purified using the same peptide coupled to Sepha-rose (Alpha Diagnostics International, San Antonio, TX). Human VHLantiserum was generously provided by Dr. R. Burk (Albert EinsteinCollege of Medicine, Bronx, NY) (34). Human VHL monoclonal antibodyand FLAG M5 monoclonal antibody were from Pharmingen and Sigma,respectively. Sp1, glutathione S-transferase (GST), and HA antiserawere from Santa Cruz Biotechnology, as were fluorescein isothiocya-nate (FITC)- and cyanine-3 (cy3)-tagged anti-rabbit and anti-mousesecondary antibodies. Horseradish peroxidase-linked anti-rabbit andanti-mouse IgGs were from Bio-Rad.
Immunoprecipitation and Western Blotting—Cultured cells were ly-sed in lysis buffer (Tris 50 mM, pH 7.6, NaCl 150 mM, EDTA 30 mM,Triton X-100 0.5%,) with Complete protease inhibitor (Roche MolecularBiochemicals), precleared with protein A-agarose bead (Santa CruzBiotechnology), and mixed with antibody at excess (18, 19). Complexeswere pelleted with protein A-agarose, washed with lysis buffer, elutedwith sample buffer, and analyzed by SDS-PAGE. Immunoblotting wasperformed as previously described using the antibodies above. Formultitissue Western analysis, tissues were minced and homogenized
with a Teflon pestle in lysis or KETN buffers (100 mM KCl, 1 mM EDTA,10 mM Tris, pH 7.5, 0.1% Nonidet P-40) containing protease inhibitor.Human tissue lysates were obtained from Genotech (St. Louis, MO).Relative intensities of positive bands were assessed by densitometryusing Image 1.62 (National Institutes of Health). Backgrounds weresubtracted to assign densitometry values.
Northern Analysis—Total RNA was prepared using RNAzol (Tel-Test) according to the manufacturer’s instructions. A multitissueNorthern blot was probed as recommended (CLONTECH) using a 1-kb5� Jade-1 coding sequence fragment. Northern analysis was otherwiseperformed as described earlier (19).
Immunocytochemistry—Cells were grown on Chamber slides (Lab-Tek) and fixed with 1:1 methanol and acetone for 2 min at roomtemperature. Immunodetection was performed with affinity-purifiedanti-Jade-1 serum or VHL monoclonal antibody followed by FITC- orcy3-conjugated anti-rabbit or anti-mouse antibodies. Images were ob-tained by fluorescence microscopy (Nikon Optiphot) and digital imaging(RT Color camera, Diagnostic Instruments).
Metabolic Labeling—Cells were starved in cysteine (Cys)- and me-thionine (Met)-free medium for 1–2 h, then fed deficient medium con-taining 100 �Ci/ml 35S-Met and 35S-Cys (EasyTag express,PerkinElmer Life Sciences), followed by medium containing 100-foldexcess unlabeled Met and Cys as chase for times indicated. Labelingtimes were 15 min or 1 h for Jade-1 in transiently transfected cells or2.5 h for endogenous Jade-1 in renal cancer cells. Immunoprecipitationswere carried out as above, and proteins were separated by SDS-PAGE.Correct identification of labeled endogenous Jade-1 was aided by con-trol lanes of labeled untagged Jade-1 from transfected 293T17 cellsimmunoprecipitated with Jade-1 antiserum, with and without compet-itor peptide. Dried gels were subjected to autoradiography, and bandswere quantitated by densitometry using Image 1.62. Detection of la-beled endogenous Jade-1 required 3 weeks of autoradiography. Proteinhalf-life was determined by log densitometry plotting. Alternatively,protein half-life was estimated based on densitometry results at the 0-and 1-h chase times, using linear regression to identify the time pointat which the initial signal strength would be halved. Errors are re-ported as �1 S.D.
RESULTS
The Jade-1 Gene and Protein—The human Jade-1 gene wasidentified as a particularly strong VHL-interacting clone in ayeast two-hybrid screen of human VHL p30. VHL is a lowabundance intracellular protein that resides in multiple sub-cellular compartments (22, 31–34). Thus, the yeast screen wasperformed to identify potentially rare or short-lived VHL pro-tein interactions. An adult human kidney library was chosenbecause half of the mass of an adult human kidney is composedof proximal tubule cells. One million library clones in pB42ADyeast expression vector were screened against human VHL aa2–213 in LexA-expressing, inducible yeast expression vectorpGilda. Positive yeast colonies were confirmed by restreakingunder double-selection conditions. Library clones rescued fromconfirmed positive colonies were tested individually for inter-action with human VHL aa 2–213 and 55–143 as well asfull-length mouse VHL in pGilda. VHL aa 55–143 were chosenas a minimum substrate-binding VHL beta domain. Jade-1 wasone of 14 strongest interactors (3�) in yeast of 40 differentgenes identified, because VHL-Jade-1-cotransformed coloniesappeared blue at 24 h of incubation. Jade-1 also interactedconvincingly with human VHL aa 55–143 (3�) as well asmouse VHL (2�) in yeast. VDU1 (10) was also recovered in thescreen, supporting the validity of this approach for identifyingbona fide VHL interactors. Additional confirmed positive inter-actors in yeast include four other novel genes, 17 known genesencoding a wide range of proteins, and 14 known genes encod-ing chaperones or chaperone-like proteins, which are commonfalse-positives. As reported in another screen, VHL-bindingprotein-1 (3�) and filamin (2�) were also recovered (49),whereas other known VHL-interacting proteins were not.
GenBankTM BLAST searches indicate that the Jade-1 li-brary clone contains the complete coding sequence of a novel,single gene (Fig. 1A). The 3570-nt clone was found in-frame
VHL Stabilizes Novel PHD Protein Jade-139888
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from
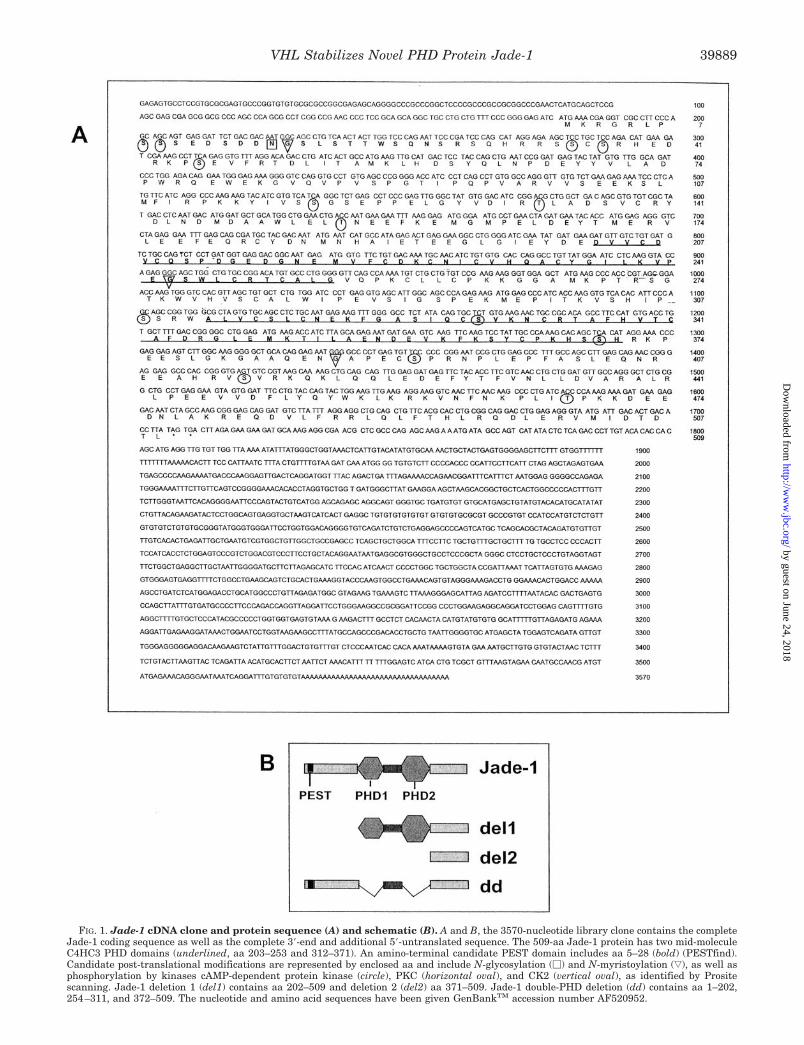
FIG. 1. Jade-1 cDNA clone and protein sequence (A) and schematic (B). A and B, the 3570-nucleotide library clone contains the completeJade-1 coding sequence as well as the complete 3�-end and additional 5�-untranslated sequence. The 509-aa Jade-1 protein has two mid-moleculeC4HC3 PHD domains (underlined, aa 203–253 and 312–371). An amino-terminal candidate PEST domain includes aa 5–28 (bold) (PESTfind).Candidate post-translational modifications are represented by enclosed aa and include N-glycosylation (�) and N-myristoylation (ƒ), as well asphosphorylation by kinases cAMP-dependent protein kinase (circle), PKC (horizontal oval), and CK2 (vertical oval), as identified by Prositescanning. Jade-1 deletion 1 (del1) contains aa 202–509 and deletion 2 (del2) aa 371–509. Jade-1 double-PHD deletion (dd) contains aa 1–202,254–311, and 372–509. The nucleotide and amino acid sequences have been given GenBankTM accession number AF520952.
VHL Stabilizes Novel PHD Protein Jade-1 39889
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from

with the B42 activation domain and has 178-nt 5� untrans-lated, 1527-nt coding, and 1831-nt 3� untranslated sequence, aswell as a polyadenylation signal and poly(A)� tail. The codingsequence is followed by several stop codons (Fig. 1A).
The deduced Jade-1 509 aa sequence (Fig. 1A) has a pre-dicted 58.4-kDa mass and 5.25 isoelectric pH. It has two con-sensus mid-molecule PHDs (50), also known as leukemia-asso-ciated protein (51) or trithorax consensus domains (52), whichare 50- to 70-aa C4HC3 zinc-binding motifs (Fig. 1, A and B).Alternatively, the second PHD may represent an extendedPHD (53) and include aa 257–371. Jade-1 residues 5–28 com-prise a strong candidate PEST domain (PESTfind score of �11)(Fig. 1, A and B), which is a charged, unstructured region thatpromotes susceptibility to degradation (54). Jade-1 has no signalor transmembrane sequences. Candidate sites for N-glycosyla-tion, myristoylation, and serine or threonine phosphorylation areshown, based on Prosite analysis, although the patterns foundare short and not highly specific. No closely homologous proteinshave been characterized. However, a transcript called E9 wasidentified in a differential screen of genes induced with apoptosisin a breast cancer line and is predicted to encode a closely relatedPEST- and PHD-containing protein (55).
To determine whether transfected Jade-1 encodes a proteinof the anticipated size, the protein coding sequence was sub-cloned into CMV promoter-driven tagged and untagged expres-sion vectors. A 64-kDa expressed protein is consistently seen inWestern blots of Jade-1 transfected but not untransfected orcontrol transfected cells (see Figs. 2A, 3A, and 3E), indicatingproduction of Jade-1 protein that is just larger than the pre-dicted 58.4-kDa molecular mass. Jade-1 may therefore be post-translationally modified.
To confirm the identify of endogenous Jade-1 protein, immu-noprecipitability and electrophoretic mobility of vector-ex-pressed and endogenous human Jade-1 proteins were com-pared. Antisera were generated against a 20-aa peptidecorresponding to the Jade-1 carboxyl terminus in two rabbits.By enzyme-linked immunosorbent assay, both antisera detectthe immunizing peptide to 1:100,000 dilution. Both Jade-1immune sera immunoprecipitate from 2 mg of cell lysates asimilar prominent 64-kDa band, whereas neither preimmuneserum does so, as assessed by SDS-PAGE and Jade-1 Westernanalysis (Fig. 2A). 293T17 cells were also transiently trans-fected with an untagged human Jade-1 expression vector. Anband identical in appearance was immunoprecipitable fromonly 250 �g of transfected cell lysate (Fig. 2A, far right lane),consistent with vector-expressed Jade-1. Similarly, tagged,transfected Jade-1 was identical in immunoprecipitability andappearance, although it was of higher molecular weight (datanot shown). Endogenous Jade-1 (Fig. 2B) and transfectedJade-1 immunoprecipitation (data not shown) could also becompletely blocked with the immunizing peptide. Thus, bothendogenous and transfected Jade-1 protein can be immunode-tected and immunoprecipitated in a highly specific manner.Moreover, these observations support the notion that the en-dogenous and transfected Jade-1 aa sequences are identical.Jade-1 antiserum 1 was used for subsequent experiments.
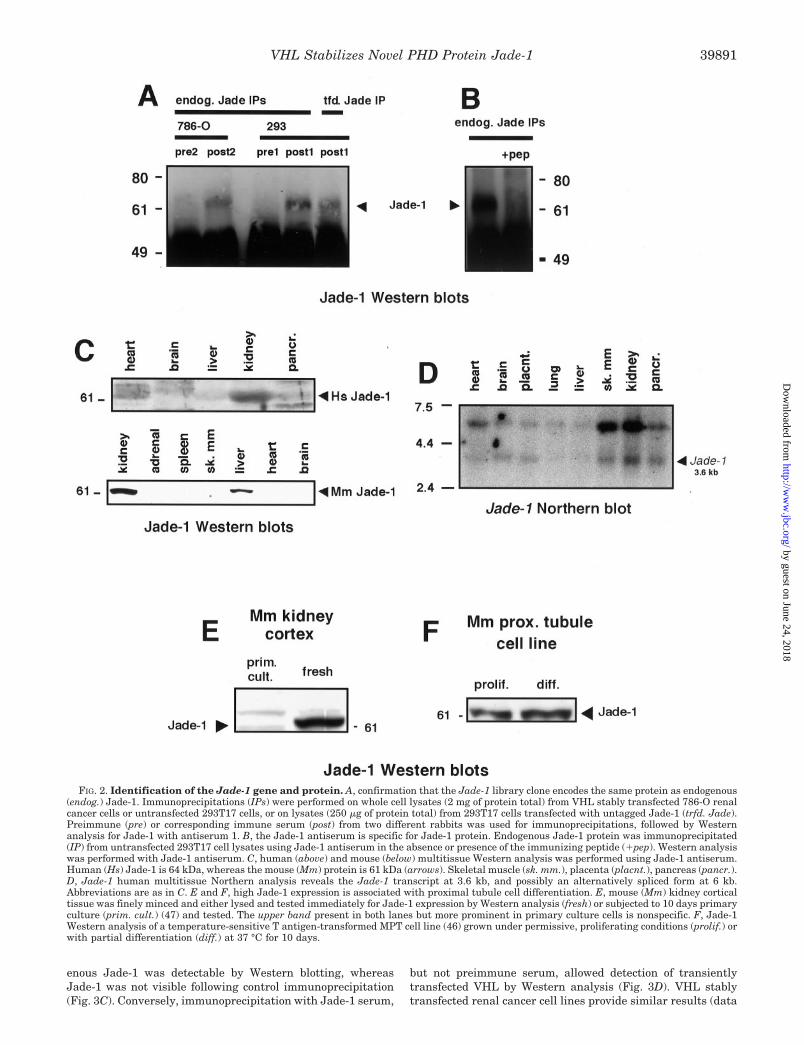
To examine Jade-1 protein distribution, Western analysiswas performed on several tissues (Fig. 2C). The human andmouse Jade-1 proteins, 64 and 61 kDa, respectively, were by farmost highly expressed in the kidney. Lower Jade-1 expressionwas also seen in human pancreas, liver, and heart and inmouse liver. Jade-1 was not readily detectable in human brain.In human cell lines, Jade-1 has been observed in HeLa, 293,and in multiple renal cancer cell lines, although at low levels(data not shown). Thus, although some difference in tissue
distribution was found between human and mouse, the kidneywas the major site of Jade-1 protein expression.
Jade-1 message and expression pattern were characterizedwith a human multitissue Northern blot (Fig. 2D). A 3.6-kbJade-1 transcript, identical to the expected size, was mostprominent in kidney. This message was also highly expressedin pancreas and skeletal muscle, but was found in all tissuestested with longer exposure (data not shown). The absence ofJade-1 protein in tissues, such as brain and skeletal muscle,where message was clearly expressed suggests that the pri-mary level of Jade-1 control may not be at the RNA level.Another major 6-kb transcript was also seen with a distributionsimilar to Jade-1 (Fig. 2D). Thus, as observed for Jade-1 pro-tein, Jade-1 message was most highly expressed in kidney, anda major band corresponds to the library clone.
To confirm that Jade-1 is expressed in renal proximal tubulecells, Western blots of kidney cortex and cultured mouse prox-imal tubule cells were performed. Kidney cortex is roughly 80%proximal tubule cells by mass. Jade-1 was prominently ex-pressed in renal cortex (Fig. 2E, right lane) and had lowerexpression in renal medulla (data not shown). However, whenmouse proximal tubule cells proliferated in primary culture,Jade-1 expression was greatly reduced, and the protein exhib-ited a slight reduction in molecular mass (Fig. 2E, left lane).These cells are at least 95% mouse renal proximal tubule cells(47). Jade-1 expression was also high in a temperature-sensi-tive SV40 T antigen transformed mouse proximal tubule (MPT)line and increases with differentiation (Fig. 2F). Together theseobservations indicate that Jade-1 is expressed in renal cancerprecursors and that differentiation or perhaps quiescence in-creases Jade-1 expression.
Jade-1 and VHL Proteins Interact—To verify the strongVHL-Jade-1 interaction observed in yeast, binding was testedin transiently transfected mammalian cells. Coding sequencefor Jade-1 and the other positive library clones was insertedinto mammalian expression vector pFLAG-CMV2. HA-taggedVHL was cotransfected into 293T17 cells with FLAG-Jade-1 orother FLAG-tagged library clones, several of which are in-cluded for comparison. In these experiments, immunoprecipi-tation of HA-VHL coimmunoprecipitated much FLAG-Jade-1,less FLAG-clone 10, little FLAG-clone 4, and no FLAG-clone 5,as shown by anti-FLAG Western blotting (Fig. 3A, HA-IPs).Jade-1, clone 10, and clone 4 expression levels in whole celllysates were comparable (Fig. 3A, lysates), which suggests theVHL interaction with Jade-1 is stronger than with clones 10 or4. As shown, more than 30% of Jade-1 coimmunoprecipitatedwith VHL, based on comparison of input and detected amountsof protein. FLAG-clone 5 expression levels in whole cell lysateswere undetectable in this experiment. Conversely, FLAG im-munoprecipitation of the Jade-1 library clone and to a lesserextent library clone 10 robustly coimmunoprecipitated HA-VHL (Fig. 3B). In contrast, no interaction was observed withclones 4 or 5. Thus, Jade-1 exhibited a strong bidirectionalinteraction with VHL, whereas clone 10 had a modest bidirec-tional interaction. Clone 4 had a weak unidirectional interac-tion. A strong VHL-Jade-1 interaction was also observed inHeLa and HT1080 cells (data not shown). The Jade-1 interac-tion with VHL in mammalian cells was stronger than that ofany other library clone tested. VHL therefore binds Jade-1avidly and selectively. Because Jade-1 did not bind p53 (datanot shown), its interaction with VHL is also selective.
To detect an interaction between VHL and endogenousJade-1 protein, immunoprecipitations were performed in VHLtransiently transfected 293T17 cells and in VHL stably trans-fected renal cancer cells. Following transient transfection ofVHL and immunoprecipitation using a VHL antiserum, endog-
VHL Stabilizes Novel PHD Protein Jade-139890
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from
enous Jade-1 was detectable by Western blotting, whereasJade-1 was not visible following control immunoprecipitation(Fig. 3C). Conversely, immunoprecipitation with Jade-1 serum,
but not preimmune serum, allowed detection of transientlytransfected VHL by Western analysis (Fig. 3D). VHL stablytransfected renal cancer cell lines provide similar results (data
FIG. 2. Identification of the Jade-1 gene and protein. A, confirmation that the Jade-1 library clone encodes the same protein as endogenous(endog.) Jade-1. Immunoprecipitations (IPs) were performed on whole cell lysates (2 mg of protein total) from VHL stably transfected 786-O renalcancer cells or untransfected 293T17 cells, or on lysates (250 �g of protein total) from 293T17 cells transfected with untagged Jade-1 (trfd. Jade).Preimmune (pre) or corresponding immune serum (post) from two different rabbits was used for immunoprecipitations, followed by Westernanalysis for Jade-1 with antiserum 1. B, the Jade-1 antiserum is specific for Jade-1 protein. Endogenous Jade-1 protein was immunoprecipitated(IP) from untransfected 293T17 cell lysates using Jade-1 antiserum in the absence or presence of the immunizing peptide (�pep). Western analysiswas performed with Jade-1 antiserum. C, human (above) and mouse (below) multitissue Western analysis was performed using Jade-1 antiserum.Human (Hs) Jade-1 is 64 kDa, whereas the mouse (Mm) protein is 61 kDa (arrows). Skeletal muscle (sk. mm.), placenta (placnt.), pancreas (pancr.).D, Jade-1 human multitissue Northern analysis reveals the Jade-1 transcript at 3.6 kb, and possibly an alternatively spliced form at 6 kb.Abbreviations are as in C. E and F, high Jade-1 expression is associated with proximal tubule cell differentiation. E, mouse (Mm) kidney corticaltissue was finely minced and either lysed and tested immediately for Jade-1 expression by Western analysis (fresh) or subjected to 10 days primaryculture (prim. cult.) (47) and tested. The upper band present in both lanes but more prominent in primary culture cells is nonspecific. F, Jade-1Western analysis of a temperature-sensitive T antigen-transformed MPT cell line (46) grown under permissive, proliferating conditions (prolif.) orwith partial differentiation (diff.) at 37 °C for 10 days.
VHL Stabilizes Novel PHD Protein Jade-1 39891
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from

FIG. 3. Jade-1 and VHL proteins interact. A and B, cotransfected Jade-1 and VHL interact strongly. A, left panel: FLAG (FL)-tagged Jade-1and other library clones were cotransfected with hemagglutinin (HA)-tagged VHL in 293T17 cells. HA antibody immunoprecipitations from 1.5 mgof cotransfected cell lysates were followed by FLAG antibody Western blotting. Right panel: FLAG Western blot of transfected 293T17 whole celllysates (60 �g per lane), the same as those used in immunoprecipitations in A and B. Other FLAG-tagged library clones, FL-4, FL-5, and FL-10,or empty vector (FL-vec), are shown for comparison. B, cotransfections were performed as in A but were followed by FLAG immunoprecipitationand VHL Western blotting. C and D, endogenous Jade-1 binds transfected VHL. C, 293T17 cells were transiently transfected with VHL, and wholecell lysates were immunoprecipitated using a VHL antiserum or control (ctrl) GST antiserum and assayed for the presence of Jade-1 by Westernblotting. D, lysates from VHL-transfected 293T17 cells were immunoprecipitated using preimmune serum (pre) or Jade-1 immune serum (post).VHL was detected using a VHL monoclonal antibody. E–G, the Jade-1 PHD regions are not required for interaction with VHL. E, expression levelsof cotransfected Jade-1 (J) (upper panels) or VHL protein (V) (lower panels) as measured by Western blotting of the same whole cell lysates usedfor immunoprecipitations in F and G. In 293T17 cells, VHL or empty pFLAG-CMV2 (ev) was cotransfected with FLAG- or HA-tagged Jade-1 ortruncations (see Fig. 1B for construct schematics), or with empty pCR3.1 uni HA (ev). Lower panels are independent immunodetections of the sameblots probed in the upper panels. F, Jade-1 immunoprecipitations of cell lysates from E were followed by VHL immunoblotting. G, VHLimmunoprecipitations of the same cell lysates were followed by Jade-1 immunodetection.
VHL Stabilizes Novel PHD Protein Jade-139892
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from

not shown). These observations indicate that endogenousJade-1 binds VHL.
To determine the Jade-1 regions responsible for the VHLinteraction, HA-tagged Jade-1 truncations were generatedlacking the amino terminus (and candidate PEST domain)(del1), the amino terminus and PHD regions (del2), or bothPHDs alone as a double internal deletion (dd), as diagrammedin Fig. 1B. In whole cell lysates, transient expression of full-length Jade-1 and the dd and del2 proteins was robust,whereas del1 expression was lower (Fig. 3E, upper panels).VHL expression was comparable in these cotransfections (Fig.3E, lower panels). Jade-1 immunoprecipitation of these samecell lysates permitted coimmunoprecipitation of VHL with full-length Jade-1, dd, and del1, but not with del2 (Fig. 3F). Like-wise, VHL immunoprecipitation allowed coimmunoprecipita-tion of full-length Jade-1, dd, and del1, but not del2 (Fig. 3G).The VHL interaction with del1 did appear reduced, even takinginto account lower del1 expression. As expected, immunopre-cipitation in the absence of either partner did not show theinteraction (Fig. 3, F and G, end lanes). Thus, these experi-ments demonstrate that the Jade-1 carboxyl terminus andPHD regions themselves are not absolutely required and thatthe amino terminus and the inter-PHD region in particularmay be most important for interaction with VHL. These find-ings also support the specificity of the VHL-Jade-1 interaction.
Localization of Jade-1 and VHL—To determine subcellularcompartments where Jade-1 might reside, cells were tran-siently transfected with HA-tagged Jade-1. In 293T17 cells,transiently transfected HA-Jade-1 gives a strong diffuse andspeckled cytoplasmic signal (Fig. 4A). Occasionally, intenseperinuclear Jade-1 fluorescence was seen, as shown. Cotrans-fected FLAG-VHL exhibited a nearly identical immunofluores-cence pattern (Fig. 4B). Moreover, the Jade-1 and VHL proteinswere almost completely colocalized (Fig. 4C). The HA anti-serum and FITC-linked anti-rabbit secondary antibodies gaveno detectable fluorescence with untransfected or vector trans-fected cells (data not shown), which indicates signal is highlyspecific for HA-tagged Jade-1 protein. VHL was detected witha monoclonal antibody and cy3-conjugated anti-mouse second-ary antibody, a combination that also showed negligible back-ground or bleedthrough fluorescence to the other fluorophorewavelength (data not shown). In MPT cells, transiently trans-fected HA-Jade-1 was seen in prominent nuclear speckles, inaddition to the diffuse and speckled cytoplasmic pattern (Fig.4D). Nuclear speckle localization has also been described forother PHD proteins (56, 57).
Endogenous Jade-1 was localized using Jade-1 antiserum 1that had been affinity-purified against the immunizing pep-tide. Several lines of evidence suggest this antibody is specificfor human Jade-1 in immunofluorescence studies despite itsrecognition of several bands by Western blotting. First, theantiserum specificity for Jade-1 in immunoprecipitations washigh, because no cross-reacting bands detectable by Westernblot were immunoprecipitable (see Fig. 2, A and B). Second,although antiserum 1 readily detected 61-kDa mouse Jade-1 byWestern blot as well as nonspecific bands, it could not immu-noprecipitate any of these proteins (data not shown). Moreover,the mouse Jade-1 immunofluorescence signal was negligible atthe antibody dilution used for human cells. These observationssuggest strongly that the antiserum 1 Jade-1 immunofluores-cence signal corresponds only to the highest affinity target,human Jade-1, and not any lower affinity, non-immunoprecipi-table protein, such as mouse Jade-1.
Endogenous Jade-1 appeared in prominent nuclear specklesin 786-O (Fig. 4E) and A498 renal cancer cells (Fig. 4H). Incytoplasm, diffuse, filamentous, and speckled endogenous
Jade-1 fluorescence was seen as well. Stably transfected VHLcolocalized with endogenous Jade-1 in a subset of Jade-1-posi-tive nuclear, perinuclear, and cytoplasmic speckles (Fig. 4,F–I). Endogenous Jade-1 and stably transfected VHL also co-localized diffusely in the nucleus of this cell. Jade-1- and VHL-positive cytoplasmic speckles were present in roughly equalabundance, however, only about 10% of each protein colocalizedwith the other (Fig. 4I). Some Jade-1- and VHL-positive cyto-plasmic speckles appeared in ring-like groups, as indicated bythe circles. Thus, VHL and Jade-1 colocalized in several com-partments, although the colocalizing protein fractions weresmall, which suggests that the protein interactions may bedynamic.
VHL Stabilizes Jade-1 Protein Expression—To establish abiological relationship between VHL and Jade-1 protein,Jade-1 protein expression was analyzed in VHL-deficient renalcancer cell lines and compared with VHL-expressing stablederivatives by Western analysis and immunoprecipitation. In786-O, UMRC6, and A498 renal cancer cells, endogenousJade-1 protein expression is 3- to 10-fold higher in VHL stablytransfected derivatives than in parental or empty vector lines(Fig. 5A, upper panels). Protein loading for each sample pair iscomparable based on Ponceau S membrane staining (Fig. 5A,lower panel). The UMRC6 cells have lower Jade-1 expressionthan 786-O and A498 cells; consequently, UMRC6 Jade-1 sig-nal detection here required longer exposure. These results havebeen confirmed by Jade-1 immunoprecipitation (data notshown). Despite expressing even less wild-type VHL proteinthan many non-cancer renal lines, including 293T17 cells (datanot shown), UMRC6 VHL cells still exhibited 3-fold increasedJade-1 expression. This observation supports the notion thatelevated Jade-1 protein levels are not merely the result of VHLoverexpression. Furthermore, increased Jade-1 expression wasreadily seen in other VHL-transfected 786-O stable lines incomparison with additional 786-O empty vector lines and wasevident regardless of the degree of cell confluence (data notshown). These results establish a consistent biological relation-ship whereby the presence of VHL increases Jade-1 expression.
To initially assess how VHL might increase Jade-1 expres-sion, Jade-1 was transiently transfected into 293T17 cells withand without VHL. One day following transfection the amountof transfected Jade-1 protein was largely unaffected by VHLcotransfection (Fig. 5B, far left panel). In contrast, VHL co-transfection substantially increased Jade-1 abundance 3 dayspost-transfection (Fig. 5B, right panels). In HeLa and HT1080cancer cell lines in particular VHL dramatically increasedJade-1 protein. As controls, cotransfected empty vector or betagalactosidase did not increase Jade-1. In addition, VHL co-transfection did not increase the expression of p53 or VHL-binding proteins PKC � and the C2H2 zinc-finger transcriptionfactor Sp1 (Fig. 5C), supporting the notion that VHL specifi-cally increases Jade-1 abundance. VHL-dependent increases intransfected Jade-1 suggest that the VHL effect on Jade-1 is notat the transcriptional or mRNA level, because Jade-1 genecontrol elements were not present and the regulatory elementsin the different expression vectors were similar. Because in-creased Jade-1 expression was most notable by late post-trans-fection, we examined whether VHL affects Jade-1 proteinstability.
To explore the mechanism whereby VHL increases Jade-1abundance, pulse-chase metabolic labeling experiments wereperformed in renal cancer cell lines. 786-O cells stably trans-fected with wild-type VHL or an empty expression vector werepulsed with radiolabeled 35S-Met and 35S-Cys. Labeled Jade-1was immunoprecipitated from 786-O cell lysates and subjectedto SDS-PAGE, followed by autoradiography and densitometry.
VHL Stabilizes Novel PHD Protein Jade-1 39893
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from

FIG. 4. Jade-1 resides in cytoplasmic and nuclear speckles and partly colocalizes with VHL. A–C, Jade-1 and VHL colocalize intransiently transfected 293T17 cells. 293T17 cells were transiently transfected with HA-Jade-1 and FLAG-VHL. A, anti-HA polyclonal antibodyand FITC-labeled anti-rabbit secondary antibody were used to localize transfected Jade-1. B, anti-VHL monoclonal antibody and cy3-labeledanti-mouse secondary antibodies were used to detect VHL. C, merged view of A and B. D, transfected Jade-1 is found in nuclear speckles in mouseproximal tubule cells. MPT cells were transiently transfected with HA-Jade-1. Anti-HA polyclonal antibody and FITC-conjugated anti-rabbit
VHL Stabilizes Novel PHD Protein Jade-139894
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from

Endogenous Jade-1 protein half-life as estimated by linearregression increases from 40 � 6 to 81 � 21 (S.D.) minutes withreintroduction of wild-type VHL in 786-O cells, based on re-sults from three experiments. A single, representative experi-ment is shown in Fig. 6A, in which the Jade-1 protein half-lifeis 41 min without VHL and 62 min with VHL. Similar resultswere obtained with A498 stable lines. The amount of labeled
Jade-1 at the end of the 2.5-h labeling period was higher inVHL-expressing cells than VHL-deficient cells, most likely dueto increased degradation of Jade-1 during labeling withoutVHL. This hypothesis is supported by the fact that the Jade-1half-life was shorter than the labeling period itself, particularlyin VHL-null cells. Furthermore, a similar discrepancy inJade-1 abundance with and without VHL was seen at the0-chase time point in transient transfections with 1-h labeling(Fig. 6B, right panel) but not with 15-min labeling (Fig. 6B, leftpanel). Because relatively small fractions of the proteins inter-act (Fig. 4, F–I), VHL most likely stabilizes Jade-1 through a“hit-and-run” mechanism.
To model the in vivo protein stabilization mechanism, pulse-chase metabolic labeling experiments were performed in293T17 cells transiently transfected with Jade-1, with andwithout wild-type VHL, using 1-h labeling periods. Wild-typeVHL cotransfection increased immunoprecipitable, radiola-beled Jade-1 at the 0-, 2-, and 4-h chase time points (Fig. 6B,right panel). Labeled, coimmunoprecipitated VHL appeared inabundance at the bottom of the blot. Using 0- and 1-h chasetime points from four experiments and estimating by linearregression, VHL increased cotransfected Jade-1 half-life from39 � 3 to 106 � 14 (S.D.) min, similar to the VHL effect onendogenous Jade-1. A multipoint decay log plot provided sim-ilar transfected Jade-1 half-life results (Fig. 6C). The plottedline slopes differ by more than 3-fold with and without VHL(slope �0.326 versus �1.03, respectively), indicating VHL sub-stantially increases the Jade-1 protein half-life. Shortening thelabeling period to 15 min equalized immunoprecipitablelabeled Jade-1 with or without VHL at chase time 0 (Fig. 6B,left panel), supporting the notion that VHL does not increaseJade-1 protein synthesis. In fact, Jade-1 production appearedslightly decreased with cotransfected VHL, a reproducible find-ing consistent with VHL-mediated repression of a cotrans-fected promoter (18, 19). Thus, these results support the en-dogenous Jade-1 data that VHL increases Jade-1 abundanceprimarily by prolonging Jade-1 protein half-life.
DISCUSSION
Jade-1 is a novel protein that interacts physically and func-tionally with the VHL tumor suppressor. No closely homolo-gous protein has been characterized previously. Jade-1 is short-lived and contains a candidate PEST degradation domain andPHD motifs as major structural features. Jade-1 is most highlyexpressed in kidney and resides in cytoplasm and the nucleus,both diffusely and in prominent speckles. Jade-1 expression ishigh in differentiated proximal tubule cells, which are renalcancer precursors, and expression falls with proliferation. Thestrong VHL-Jade-1 interaction does not depend on intact PHDfingers, but may require the inter-PHD region and the PEST-containing amino terminus. Most importantly, VHL increasesboth transfected and endogenous Jade-1 levels by directly sta-bilizing the protein, which represents a new VHL function.Preliminary results not presented here also suggest thatJade-1 is growth suppressive and may promote apoptosis.Thus, Jade-1 protein is a candidate regulatory molecule thatmay participate in VHL-mediated renal tumor suppression.
Jade-1 is a novel protein-coding gene. Initially, the Jade-1 nt
secondary antibodies were used to localize Jade-1 in this single cell, shown in a background of untransfected cells. E, endogenous Jade-1 is foundin nuclear speckles in renal cancer cells. Untransfected 786-O renal cancer cells were probed with affinity-purified anti-Jade-1 serum andFITC-labeled anti-rabbit secondary antibody. A single 786-O cell of typical appearance is shown. As in D, nuclear speckles are in the same visualplane as the nucleus. F–I, colocalization of endogenous Jade-1 with stably transfected VHL. Wild-type VHL stably transfected A498 cells wereprobed for both VHL and Jade-1. F, VHL was detected using anti-VHL monoclonal and cy3-labeled anti-mouse secondary antibodies. G, a mergedview of the cell from F and H is shown. H, Jade-1 was detected using affinity-purified anti-Jade-1 serum and an FITC-labeled anti-rabbit secondaryantibody. I, colocalization of endogenous Jade-1 and stably transfected VHL to cytoplasmic speckles. Higher magnification view of perinuclearregion from G, with a 90° rotation counterclockwise. Nucleus is to the right. Circled regions highlight groups of speckles with Jade-1-VHLcolocalization.
FIG. 5. VHL increases Jade-1 protein abundance. A, VHL in-creases Jade-1 protein expression in three of three renal cancer celllines. Upper panels: Jade-1 Western analysis of whole cell lysates fromrenal cancer lines stably transfected with either empty vector (786-Oand A498) (�) or wild-type VHL p30 (V). UMRC6 control cells (�) arethe parental line. Paired samples from a single blot are boxed sepa-rately, because UMRC6 cells require longer exposure to demonstrateJade-1. Lower panel: Ponceau S stain of same membrane probed above.B, VHL increases Jade-1 expression 3 days, but not 1 day, post-cotrans-fection. Cultured cell lines as indicated were transiently cotransfectedwith a Jade-1 expression vector and a second expression vector, eitherempty (�), or containing wild-type VHL (V) or �-galactosidase (Bgal). 1or 3 days following transfection Jade-1 protein levels were determinedby Western analysis. C, the VHL-mediated increase in expression isspecific for Jade-1. FLAG-tagged p53 (FL-p53), FLAG-tagged proteinkinase C � (FL-PKCz), or transcription factor Sp1 were cotransfectedwith either vector alone (�) or wild-type VHL (V). FLAG or Sp1 West-ern blotting was performed on cell lysates harvested 3 days aftertransfection.
VHL Stabilizes Novel PHD Protein Jade-1 39895
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from

and aa sequences were not found in GenBankTM. Subse-quently, cDNA clones were deposited with predicted proteincoding sequences identical to Jade-1, including “hypotheticalprotein” FLJ22479, which is represented by protein accessionnumbers NP_079176 (New Energy Development Organization(Japan) human cDNA sequencing project, Japan) andXP_033946 (NCBI annotation project). Protein FLJ14714 isidentical to FLJ22479 and has been assigned accession numberAK027620 (New Energy Development Organization (Japan)project, Japan). Another cDNA encodes a putative “unnamedprotein product” with accession number BAB55239 (New En-ergy Development Organization (Japan) project, Japan). Noprotein expression or functional information accompaniesthese GenBankTM reports. Characterization of the humanJade-1 cDNA clone identified revealed the presence of a com-
plete 3�-end. Expressed sequence tag sequences were reviewedto determine the Jade-1 initiation codon, and the MKF site waschosen because no clone with substantially more 5� sequencewas found. Correct identification of the initiation codon is sup-ported by the identical sizes of untagged, transfected Jade-1protein and the endogenous, immunoprecipitable Jade-1 pro-tein. Other Jade-1 transcripts exist as well, as suggested by theprominent 6-kb message on Northern analysis and a 95-kDaJade-1 immunoreactive band on Western blotting (data notshown). Although such bands may reflect homologs or alterna-tive transcripts, longer Jade-1 coding sequence clones havebeen deposited in public databases. For example, an 846-aaJade-1 protein would be predicted based on combined overlap-ping Celera Genomics cDNA clone CT8385 and cloneKIAA1807 (accession number AB058710) (Kazusa DNA Re-
FIG. 6. VHL stabilizes Jade-1 protein. A, VHL stabilizes endogenous Jade-1 protein. Vector or VHL stably transfected 786-O renal cancer cellswere labeled for 2.5 h with 35S-Cys and 35S-Met and chased with excess unlabeled Cys and Met for times shown. Autoradiography (below) andcorresponding densitometry in pixels with background subtracted (above) is shown for the specific radiolabeled, immunoprecipitated Jade-1 proteinband (arrow). The Jade-1 band was confirmed both by size and as the only band competed away by the immunizing peptide. Linear regression ofthis representative result approximates a Jade-1 protein half-life of 41 min without VHL and 62 min with VHL. B, VHL stabilizes transfectedJade-1 protein. 293T17 cells were transiently transfected with Jade-1 and either empty vector (�) or wild-type VHL (�V), metabolically labeledand chased as in A for the times shown. Labeled Jade-1 was immunoprecipitated and subjected to SDS-PAGE and autoradiography. C, VHLstabilizes transfected Jade-1 protein. Logarithmic densitometry plot of Jade-1 protein degradation with and without VHL from a more detailedpulse-chase experiment than B, including duplicate time points. 293T17 cell transient transfections and Jade-1 immunoprecipitations wereperformed as in B.
VHL Stabilizes Novel PHD Protein Jade-139896
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from

search Institute) (58), although their non-coding sequences areincomplete. The predicted 846-aa Jade-1 protein would have amass near 95 kDa and would contain the two PHD regions butwould lack the candidate PEST sequence or any additionalrecognizable domains. This larger protein also contains inter-nally 13 of the 20 aa in the immunizing peptide, which stillmight be sufficient for immunoreactivity. Although alternativesplicing may change interaction with and regulation by VHL orother functions, 509-aa Jade-1 may be the first member of agroup of proteins stabilized by the VHL tumor suppressor.
Several observations suggest Jade-1 is growth suppressive ormight participate in apoptosis. Stable Jade-1 overexpression in786-O renal cancer cells so far has not been possible. We typi-cally achieve 30–40% success rates for stable expression of anytransgene and 20% success rates with VHL, which is growthsuppressive in these cells. However, 0 of 25 Jade-1-transfected,drug-resistant stable 786-O colonies exhibited increased Jade-1expression. In addition, transient transfection of Jade-1 into293T17 cells increases apoptosis by 50%, as measured both byHoechst fluorescence and direct visualization and by taggedAnnexin V binding and fluorescence-activated cell scanning.2
This latter observation may explain the difficulty generating aJade-1 stable cell line. In addition, the closely homologous E9gene is increased 9-fold with induction of apoptosis in breastcancer cells (55). The predicted E9 polypeptide has a candidatePEST domain and two PHD regions, which further suggestssuch proteins may participate in apoptosis. As shown here,Jade-1 expression also correlates with differentiation of mouseproximal tubule cells and VHL status. The role of Jade-1 inrenal cancer is unclear, but the differentiation or quiescencerelationship is intriguing. Jade-1 expression may fall in anyproliferative renal epithelial cell lesion, such as renal cysts,even in the absence of VHL gene defects. The prominent Jade-1message in normal kidney and pancreas is also interesting,because both tissues develop cysts and cancers in VHL disease.Jade-1 may therefore be an important negative regulator ofepithelial cell growth.
These data associate VHL with a PHD protein, which has notbeen described previously. The C4HC3 PHD finger binds twozinc ions and is structurally similar to the C3HC4 RING zinc-binding domain. The PHD is a critically important motif, be-cause it is lost or mutated in several diseases, including theacute leukemias, reviewed in Ref. 59, X-linked alpha thalasse-mia with mental retardation (60), and others. Many PHD pro-teins are transcription factors or cofactors and have chromatinmodifying roles (50, 61–64). Thus, by increasing Jade-1 pro-tein, VHL might alter chromatin, which has not been previ-ously suggested and might help explain its promotion of a renalepithelial differentiation program (26). A subset of PHD pro-teins is involved in apoptosis, such as requiem (65), ing1 (66),and perhaps E9 (55). Given the well-recognized roles of VHL intranscriptional control of gene expression (7, 18) and protectionagainst stress-induced apoptosis (28–30, 67), PHD proteinJade-1 could be an important participant in these VHLpathways.
Direct protein stabilization by VHL is also a new finding.Although VHL promotes the ubiquitination and destruction ofseveral interacting proteins (7–10), no protein partner thus faris stabilized by VHL like Jade-1. Jade-1 is also stabilized in ahighly specific manner, because this effect is not seen withVHL-interacting proteins Sp1 or PKC � nor with p53. VHLmight increase Jade-1 stability in several ways. Because theVHL-Jade-1 interaction appears transient, a “hit-and-run”mechanism appears most likely, which may involve a Jade-1
post-translational modification rather than VHL-dependentsteric hindrance of Jade-1 degradation. Moreover, the Jade-1PEST domain is a strong candidate degradation-susceptibilityregion, or degron. VHL might therefore alter PEST domainphosphorylation, for example, which is well known to affectprotein stability. Although Jade-1 protein appears tissue-re-stricted, Jade-1 message is more widely expressed, suggestingthat control of Jade-1 expression may occur predominantly atthe protein level. Several proteins that do not interact withVHL are increased with VHL reintroduction, such as cell cycleinhibitors p27 (28) and p21, and Bcl2 (30, 67). As expected ofproteins increased by a tumor suppressor, they can be growthsuppressive (68), which by inference suggests Jade-1 mightalso inhibit growth. Jade-1 stabilization also supports the no-tion that VHL may play a global role in determining proteinfate. VHL-mediated control of the deubiquitinating enzymeVDU1 (10) supports this assertion, as does the widening role ofVHL in defense against cell stress (28–30, 67).
We pursued a yeast two-hybrid approach to identify lowoccupancy VHL protein partners that might have importancein renal cancer. This work identifies Jade-1, a novel PEST- andPHD-containing protein, as a strong VHL interactor that mayhelp control gene expression and differentiation in renal cancerprecursor cells. Moreover, these studies identify direct proteinstabilization as a new VHL function. Further elucidation of therole of Jade-1 in renal tumor suppression and renal epithelialcell growth and development may therefore be of considerableinterest.
Acknowledgments—We sincerely thank Dr. W. Lieberthal for helpfuldiscussions; Drs. D. Cohen, D. Salant, and D. Seldin for critical readingof the manuscript; Drs. R. Burk, W. Kaelin, W. Krek, M. Lerman,W. Lieberthal, J. Lisztwan, M. Loghman-Adham, Z. Luo, U. Moll,D. Salant, A. Toker, and R. Widom for generously providing reagents;Dr. H. Mahmud for generating pGilda constructs; and S. Waitzman fortechnical assistance.
REFERENCES
1. Latif, F., Tory, K., Gnarra, J., Yao, M., Duh, F. M., Orcutt, M. L., Stackhouse,T., Kuzmin, I., Modi, W., Geil, L., Schmidt, L., Zhou, F., Li, H., Wei, M. H.,Chen, F., Glenn, G., Choyke, P., Walther, M. M., Wend, Y., Duan, R.-S. R.,Dean, M., Glavac, D., Richard, R. M., Crossey, P. A., Ferguson-Smith, M. A.,Paslier, D. L., Chumakov, L., Cohen, D., Chinault, A. C., Maher, E. R.,Linehan, W. M., Zbar, B., and Lerman, M. I. (1993) Science 260, 1317–1320
2. Crossey, P. A., Richards, F. M., Foster, K., Green, J. S., Prowse, A., Latif, F.,Lerman, M. I., Zbar, B., Affara, N. A., Ferguson-Smith, M. A., and Maher,E. R. (1994) Hum. Mol. Genet. 3, 1303–1308
3. Foster, K., Prowse, A., van den Berg, A., Fleming, S., Hulsbeek, M. M.,Crossey, P. A., Richards, F. M., Cairns, P., Affara, N. A., Ferguson-Smith,M. A., Buys, C. H., and Maher, E. R. (1994) Hum. Mol. Genet. 3, 2169–2173
4. Gnarra, J. R., Tory, K., Weng, Y., Schmidt, L., Wei, M. H., Li, H., Latif, F., Liu,S., Chen, F., Duh, F. M., Lubensky, I., Duan, D. R., Florence, C., Pozzatti,R., Walther, M. M., Bander, N. H., Grossman, H. B., Brauch, H., Pomer, S.,Brooks, J. D., Isaacs, W. B., Lerman, M. I., Zbar, B., and Linehan, W. M.(1994) Nat. Genet. 7, 85–90
5. Chen, F., Kishida, T., Yao, M., Hustad, T., Glavac, D., Dean, M., Gnarra, J. R.,Orcutt, M. L., Duh, F. M., Glenn, G., Green, J., Hsia, Y. E., Lamiell, J., Li,H., Wei, M., Schmidt, L., Tory, K., Kuzmin, I., Stackhouse, T., Latif, F.,Linehan, W. M., Lerman, M., and Zbar, B. (1995) Hum. Mutat. 5, 66–75
6. Cohen, H. T. (1999) Curr. Opin. Nephrol. Hypertens. 8, 325–3317. Maxwell, P. H., Wiesener, M. S., Chang, G. W., Clifford, S. C., Vaux, E. C.,
Cockman, M. E., Wykoff, C. C., Pugh, C. W., Maher, E. R., and Ratcliffe,P. J. (1999) Nature 399, 271–275
8. Okuda, H., Saitoh, K., Hirai Si, S., Iwai, K., Takaki, Y., Baba, M., Minato, N.,Ohno, S., and Shuin, T. (2001) J. Biol. Chem. 276, 43611–43617
9. Pioli, P. A., and Rigby, W. F. (2001) J. Biol. Chem. 276, 40346–4035210. Li, Z., Na, X., Wang, D., Schoen, S. R., Messing, E. M., and Wu, G. (2002)
J. Biol. Chem. 277, 4656–466211. Aso, T., Lane, W. S., Conaway, J. W., and Conaway, R. C. (1995) Science 269,
1439–144312. Duan, D. R., Pause, A., Burgess, W. H., Aso, T., Chen, D. Y., Garrett, K. P.,
Conaway, R. C., Conaway, J. W., Linehan, W. M., and Klausner, R. D.(1995) Science 269, 1402–1406
13. Kibel, A., Iliopoulos, O., DeCaprio, J. A., and Kaelin, W. G., Jr. (1995) Science269, 1444–1446
14. Kroll, S. L., Paulding, W. R., Schnell, P. O., Barton, M. C., Conaway, J. W.,Conaway, R. C., and Czyzyk-Krzeska, M. F. (1999) J. Biol. Chem. 274,30109–30114
15. Iliopoulos, O., Levy, A. P., Jiang, C., Kaelin, W. G., Jr., and Goldberg, M. A.(1996) Proc. Natl. Acad. Sci. U. S. A. 93, 10595–10599
16. Gnarra, J. R., Zhou, S., Merrill, M. J., Wagner, J. R., Krumm, A.,2 J. J. Ross, M. I. Zhou, H. Wang, and H. T. Cohen, unpublished data.
VHL Stabilizes Novel PHD Protein Jade-1 39897
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from

Papavassiliou, E., Oldfield, E. H., Klausner, R. D., and Linehan, W. M.(1996) Proc. Natl. Acad. Sci. U. S. A. 93, 10589–10594
17. Levy, A. P., Levy, N. S., and Goldberg, M. A. (1996) J. Biol. Chem. 271,25492–25497
18. Mukhopadhyay, D., Knebelmann, B., Cohen, H. T., Ananth, S., and Sukhatme,V. P. (1997) Mol. Cell. Biol. 17, 5629–5639
19. Cohen, H. T., Zhou, M., Welsh, A. M., Zarghamee, S., Scholz, H.,Mukhopadhyay, D., Kishida, T., Zbar, B., Knebelmann, B., and Sukhatme,V. P. (1999) Biochem. Biophys. Res. Commun. 266, 43–50
20. Pal, S., Claffey, K. P., Dvorak, H. F., and Mukhopadhyay, D. (1997) J. Biol.Chem. 272, 27509–27512
21. Pal, S., Claffey, K. P., Cohen, H. T., and Mukhopadhyay, D. (1998) J. Biol.Chem. 273, 26277–26280
22. Ohh, M., Yauch, R. L., Lonergan, K. M., Whaley, J. M., Stemmer-Rachamimov,A. O., Louis, D. N., Gavin, B. J., Kley, N., Kaelin, W. G., Jr., and Iliopoulos,O. (1998) Mol Cell 1, 959–968
23. Lieubeau-Teillet, B., Rak, J., Jothy, S., Iliopoulos, O., Kaelin, W., and Kerbel,R. S. (1998) Cancer Res. 58, 4957–4962
24. Koochekpour, S., Jeffers, M., Wang, P. H., Gong, C., Taylor, G. A., Roessler,L. M., Stearman, R., Vasselli, J. R., Stetler-Stevenson, W. G., Kaelin, W. G.,Jr., Linehan, W. M., Klausner, R. D., Gnarra, J. R., and Vande Woude, G. F.(1999) Mol. Cell. Biol. 19, 5902–5912
25. Baba, M., Hirai, S., Kawakami, S., Kishida, T., Sakai, N., Kaneko, S., Yao, M.,Shuin, T., Kubota, Y., Hosaka, M., and Ohno, S. (2001) Oncogene 20,2727–2736
26. Davidowitz, E. J., Schoenfeld, A. R., and Burk, R. D. (2001) Mol. Cell. Biol. 21,865–874
27. Kamada, M., Suzuki, K., Kato, Y., Okuda, H., and Shuin, T. (2001) Cancer Res.61, 4184–4189
28. Pause, A., Lee, S., Lonergan, K. M., and Klausner, R. D. (1998) Proc. Natl.Acad. Sci. U. S. A. 95, 993–998
29. Gorospe, M., Egan, J. M., Zbar, B., Lerman, M., Geil, L., Kuzmin, I., andHolbrook, N. J. (1999) Mol. Cell. Biol. 19, 1289–1300
30. Schoenfeld, A. R., Parris, T., Eisenberger, A., Davidowitz, E. J., De Leon, M.,Talasazan, F., Devarajan, P., and Burk, R. D. (2000) Oncogene 19,5851–5857
31. Los, M., Jansen, G. H., Kaelin, W. G., Lips, C. J., Blijham, G. H., and Voest,E. E. (1996) Lab. Invest. 75, 231–238
32. Lee, S., Chen, D. Y., Humphrey, J. S., Gnarra, J. R., Linehan, W. M., andKlausner, R. D. (1996) Proc. Natl. Acad. Sci. U. S. A. 93, 1770–1775
33. Shiao, Y. H., Resau, J. H., Nagashima, K., Anderson, L. M., and Ramakrishna,G. (2000) Cancer Res. 60, 2816–2819
34. Schoenfeld, A. R., Davidowitz, E. J., and Burk, R. D. (2001) Int. J. Cancer 91,457–467
35. Clifford, S. C., Cockman, M. E., Smallwood, A. C., Mole, D. R., Woodward,E. R., Maxwell, P. H., Ratcliffe, P. J., and Maher, E. R. (2001) Hum. Mol.Genet. 10, 1029–1038
36. Hoffman, M. A., Ohh, M., Yang, H., Klco, J. M., Ivan, M., and Kaelin, W. G., Jr.(2001) Hum. Mol. Genet. 10, 1019–1027
37. Cockman, M. E., Masson, N., Mole, D. R., Jaakkola, P., Chang, G. W., Clifford,S. C., Maher, E. R., Pugh, C. W., Ratcliffe, P. J., and Maxwell, P. H. (2000)J. Biol. Chem. 275, 25733–25741
38. Ohh, M., Park, C. W., Ivan, M., Hoffman, M. A., Kim, T. Y., Huang, L. E.,Pavletich, N., Chau, V., and Kaelin, W. G. (2000) Nat. Cell Biol. 2, 423–427
39. Kondo, K., Klco, J., Nakamura, E., Lechpammer, M., and Kaelin, W. G., Jr.(2002) Cancer Cell 1, 237–246
40. Maranchie, J. K., Vasselli, J. R., Riss, J., Bonifacino, J. S., Linehan, W. M., andKlausner, R. D. (2002) Cancer Cell 1, 247–255
41. Ivan, M., and Kaelin, W. G., Jr. (2001) Curr. Opin. Genet. Dev. 11, 27–3442. Iliopoulos, O., Kibel, A., Gray, S., and Kaelin, W. G., Jr. (1995) Nat. Med. 1,
822–82643. Zaika, A., Marchenko, N., and Moll, U. M. (1999) J. Biol. Chem. 274,
27474–2748044. Lisztwan, J., Imbert, G., Wirbelauer, C., Gstaiger, M., and Krek, W. (1999)
Genes Dev. 13, 1822–183345. Ivanov, S. V., Kuzmin, I., Wei, M. H., Pack, S., Geil, L., Johnson, B. E.,
Stanbridge, E. J., and Lerman, M. I. (1998) Proc. Natl. Acad. Sci. U. S. A.95, 12596–12601
46. Loghman-Adham, M., Rohrwasser, A., Helin, C., Zhang, S., Terreros, D.,Inoue, I., and Lalouel, J. M. (1997) Kidney Int. 52, 229–239
47. Lieberthal, W., Triaca, V., Koh, J. S., Pagano, P. J., and Levine, J. S. (1998)Am. J. Physiol. 275, F691–F702
48. Cohen, H. T., Bossone, S. A., Zhu, G., McDonald, G. A., and Sukhatme, V. P.(1997) J. Biol. Chem. 272, 2901–2913
49. Tsuchiya, H., Iseda, T., and Hino, O. (1996) Cancer Res. 56, 2881–288550. Aasland, R., Gibson, T. J., and Stewart, A. F. (1995) Trends Biochem. Sci. 20,
56–5951. Saha, V., Chaplin, T., Gregorini, A., Ayton, P., and Young, B. D. (1995) Proc.
Natl. Acad. Sci. U. S. A. 92, 9737–974152. Koken, M. H., Saib, A., and de The, H. (1995) C R Acad. Sci. III 318, 733–73953. Linder, B., Newman, R., Jones, L. K., Debernardi, S., Young, B. D., Freemont,
P., Verrijzer, C. P., and Saha, V. (2000) J. Mol. Biol. 299, 369–37854. Rechsteiner, M., and Rogers, S. W. (1996) Trends Biochem. Sci 21, 267–27155. Szelei, J., Soto, A. M., Geck, P., Desronvil, M., Prechtl, N. V., Weill, B. C., and
Sonnenschein, C. (2000) J. Steroid Biochem. Mol. Biol. 72, 89–10256. Bjorses, P., Pelto-Huikko, M., Kaukonen, J., Aaltonen, J., Peltonen, L., and
Ulmanen, I. (1999) Hum. Mol. Genet. 8, 259–26657. Nakamura, T., Blechman, J., Tada, S., Rozovskaia, T., Itoyama, T., Bullrich,
F., Mazo, A., Croce, C. M., Geiger, B., and Canaani, E. (2000) Proc. Natl.Acad. Sci. U. S. A. 97, 7284–7289
58. Nagase, T., Nakayama, M., Nakajima, D., Kikuno, R., and Ohara, O. (2001)DNA Res. 8, 85–95
59. Hayashi, Y. (2000) Semin. Hematol. 37, 368–38060. Gibbons, R. J., Bachoo, S., Picketts, D. J., Aftimos, S., Asenbauer, B.,
Bergoffen, J., Berry, S. A., Dahl, N., Fryer, A., Keppler, K., Kurosawa, K.,Levin, M. L., Masuno, M., Neri, G., Pierpont, M. E., Slaney, S. F., and Higgs,D. R. (1997) Nat. Genet. 17, 146–148
61. Schindler, U., Beckmann, H., and Cashmore, A. R. (1993) Plant J. 4, 137–15062. Bjorses, P., Halonen, M., Palvimo, J. J., Kolmer, M., Aaltonen, J., Ellonen, P.,
Perheentupa, J., Ulmanen, I., and Peltonen, L. (2000) Am. J. Hum. Genet.66, 378–392
63. Pitkanen, J., Doucas, V., Sternsdorf, T., Nakajima, T., Aratani, S., Jensen, K.,Will, H., Vahamurto, P., Ollila, J., Vihinen, M., Scott, H. S., Antonarakis,S. E., Kudoh, J., Shimizu, N., Krohn, K., and Peterson, P. (2000) J. Biol.Chem. 275, 16802–16809
64. Schultz, D. C., Friedman, J. R., and Rauscher, F. J., 3rd. (2001) Genes Dev. 15,428–443
65. Gabig, T. G., Mantel, P. L., Rosli, R., and Crean, C. D. (1994) J. Biol. Chem.269, 29515–29519
66. Helbing, C. C., Veillette, C., Riabowol, K., Johnston, R. N., and Garkavtsev, I.(1997) Cancer Res. 57, 1255–1258
67. Devarajan, P., De Leon, M., Talasazan, F., Schoenfeld, A. R., Davidowitz, E. J.,and Burk, R. D. (2001) J. Biol. Chem. 276, 15823–15831
68. Veis, D. J., Sorenson, C. M., Shutter, J. R., and Korsmeyer, S. J. (1993) Cell 75,229–240
VHL Stabilizes Novel PHD Protein Jade-139898
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from

T. CohenMina I. Zhou, Hongmei Wang, Jonathan J. Ross, Igor Kuzmin, Chengen Xu and Herbert
Protein Jade-1The von Hippel-Lindau Tumor Suppressor Stabilizes Novel Plant Homeodomain
doi: 10.1074/jbc.M205040200 originally published online August 6, 20022002, 277:39887-39898.J. Biol. Chem.
10.1074/jbc.M205040200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/277/42/39887.full.html#ref-list-1
This article cites 68 references, 35 of which can be accessed free at
by guest on June 24, 2018http://w
ww
.jbc.org/D
ownloaded from