the journal of bio~ical chemistry 269, no. 6, of … · · 2001-06-26carrier demonstrates that...
TRANSCRIPT

THE JOURNAL OF B I O ~ I C A L CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269, No. 6, Issue of February 11, pp. 4668-4674, 1994 Printed in U.S.A.
The Axonal y-Aminobutyric Acid Transporter GAT4 Is Sorted to the Apical Membranes of Polarized Epithelial Cells*
(Received for publication, July 27, 1993, and in revised form, October 21, 1993)
Grazia PietriniSI, Young J. Suhn, Lambert Edelmannn, Gary Rudnickl, and Michael J. CaplanSII From the Departments of $Cellular and Molecular Physiology and Wharmacology, Yale University School of Medicine, New Haven, Connecticut 06510 and the IConsiglio Nazionale delle Ricerche, Center for Cytopharmacology, and the Department of Pharmacology, University of Milan, Milan 20129, Italy
Recent studies suggest that epithelial cells and neu- rons employ similar mechanisms to target proteins to the distinct subdomains of their polarized cell surface membranes. We have examined the sorting behavior of the neuronal y-aminobutyric acid (GABA) transporter GAT-1 expressed by transfection in the polarized epithe- lial Madin-Darby canine kidney (MDCK) cell line. We find that the GABA transporters endogenously ex- pressed by polarized hippocampal neurons in culture are restricted to axonal plasma membranes. In trans- fected MDCK cells, the GABA transporter is found to be localized primarily to the apical cell surface when ex- amined by immunocytochemistry, cell surface biotinyla- tion, and transport assay. MDCK cells exposed to hyper- osmotic stress express a close relative of GAT-1, the betaine transporter (BGT-1). We find that BGT-1 ex- pressed by transfection in MDCK cells accumulates pre- dominantly at the basolateral cell surface. These obser- vations suggest that the sorting information required for axonal targeting may be similar to that which medi- ates apical localization in epithelia. Furthermore, it would appear that despite their high degree of homol- ogy, the BGT-1 and GAT-1 transporters manifest sorting signals which specify their targeting to distinct cell sur- face domains.
The plasma membranes of neurons and epithelial cells are divided into distinct domains characterized by markedly differ- ent protein compositions (1-3). The epithelial plasmalemma is composed of an apical region separated by tight junctions from a basolateral surface which rests upon a basement membrane. Neuronal plasma membranes are subdivided into an axolemma and a somatodendritic membrane (4), which become function- ally distinct at the level of the axon hillock. In the epithelial case, plasmalemma1 polarity is a prerequisite for vectorial sol- ute and fluid transport. The domains of the epithelial plasma membrane must possess distinct classes of transport proteins to mediate fluxes against concentration gradients. Neuronal polarity is necessary to ensure rapid and unidirectional infor- mation flow. For both cell types, the maintenance of normal
* This work was supported by a Brown-Coxe fellowship (to G. P.), a fellowship from the American Heart Association, Connecticut affiliate (to Y. J. S.), National Institute on Drug Abuse Grant 07259 (to G. R.), a fellowship from the David and Lucille Packard Foundation (to M. J. C.), and National Institutes of Health Grant GM-42136 (to M. J. C.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "adver- tisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Molecular Physiology, Yale University School of Medicine, 333 Cedar /I To whom correspondence should be addressed: Dept. of Cellular and
St., New Haven, CT 06510. Tel.: 203-785-7316; Fax: 203-785-4951.
physiologic function requires that newly synthesized mem- brane proteins be differentially sorted to their appropriate sites of functional residence.
Recent evidence suggests that similar mechanisms may function to generate and maintain this polarity in both sys- tems. Viral glycoproteins that are sorted to the epithelial apical and basolateral surfaces have been shown to accumulate in the axonal and dendritic domains, respectively, of infected neurons (5). Furthermore, glycophospholipid-linked proteins are re- stricted to the apical surfaces of epithelial cells and to the axolemmas of neurons (6) . Thus, the signals and mechanisms responsible for apical sorting in epithelia may result in axonal targeting in neurons. To test this model further we have exam- ined the sorting behavior of an axonal protein expressed by transfection in polarized epithelial cells.
The Na,C1-dependent GABA' carrier functions to terminate GABA-ergic transmission by mediating the transport of GABA from the synaptic cleft into the cytoplasm of the nerve terminal (7,8). The recent identification of the cDNAencoding the GABA carrier demonstrates that this protein is part of a large gene family that includes the systems responsible for serotonin, nor- epinephrine, and dopamine transport (9). Each of these trans- porters is composed of a single polypeptide characterized by 12 putative transmembrane helices. Further characterization of this gene family reveals that at least four distinct isoforms of GABA transporter can be discerned (10,111. Designated GAT-1, GAT-2, GAT-3, and GAT-B, these transporters manifest differ- ent tissue localizations and pharmacologic sensitivities (12). The pharmacologic properties of the GAT-1 isoform suggest that it is expressed predominantly in neurons and not in glia (13). Electron microscopic immunolocalization studies per- formed on rat brain suggest that the GABA transporter is con- centrated in the plasma membranes of axons and axon termi- nals of GABA-ergic neurons (14). I t would appear, therefore, that neurons in situ are capable of targeting this protein to axonal membrane domains.
We wondered whether the sorting information that functions to ensure the GABA transporter's axonal localization in neu- rons would also serve to mediate apical targeting in epithelial cells. To test this possibility, we have generated stably trans- fected MDCK cell lines which express the rat GAT-1 trans- porter. We find that GAT-1 does, in fact, accumulate preferen- tially at the apical surface. These results suggest that axonal and apical sorting mechanisms may, in fact, share common features. This observation is especially interesting in light of the recent demonstration that the betaine transporter (BGT-1) endogenously expressed at the basolateral surfaces of MDCK cells shares -50% sequence identity with the GAT-1 system (15).
The abbreviations used are: GABA, y-aminobutyric acid; MDCK, Madin-Darby canine kidney; M A P , microtubule-associated protein.
4668

GABA Dansporter Is Sorted to Epithelial Apical Membranes 4669
MATERIALS AND METHODS Antibodies and Immunocytochemistry-Hippocampal neurons (the
kind gift of G. Banker, University of Virginia) were isolated from day 19 embryonic rats and cultured on glass coverslips according to the meth- ods of Goslin and Banker (16). Neurons were fured in ice-cold methanol and processed for immunofluorescence as described previously (17). The "-2 antibody was purchased from Boehringer Mannheim and used at a dilution of 1:lOO.
Antibody R24, directed against the GABA transporter, was provided by R. Jahn (Yale University) and used at a dilution of 1:50 for all immunofluorescence studies. This antibody was generated against a synthetic peptide whose sequence corresponds to amino acids 571-586 of the rat brain GABA transporter. Coupling the peptide to keyhole limpet hemocyanin and immunization in rabbits was carried out ac- cording to Lerner et al. (18).
Transfected MDCK cells were grown to confluence on transparent filters (Cyclopore, Becton Dickson, Lincoln Park, NJ), fixed in metha- nol, and processed for immunofluorescence as described in Pietrini et al. (17). The Na,K-ATPase a-subunit was localized using monoclonal anti- body 6H at a dilution of 1:lO. Production and characterization of this antibody are described elsewhere (17).
Immunofluorescence was observed with a Zeiss Axiophot epifluores- cence photomicroscope and photographed with Kodak T"AX 100 ASA film (Eastman Kodak Co.). Confocal microscopy was carried out on a Zeiss laser scanning confocal microscope, and X Z sections were gener- ated as described (19).
Subcloning and lkansfection-The cDNA encoding the GABA trans- porter, provided by B. Kanner, was subcloned into the ClaI and XbaI sites of the mammalian expression vector pCB6 (20) (gift of C. Brewer and M. Roth, University of Texas, Southwestern). MDCK cells were passaged and subjected to CaP0,-mediated transfection as described (21). Transfected cell lines were selected by growth in the antibiotic G418 (0.9 mg/ml) (Life Technologies, Inc.), and expression of the GABA transporter was assayed initially by GABA uptake assay (see below).
The cDNA encoding the betaine transporter subcloned into the pSPORTl vector was kindly provided by J . S. Handler (Johns Hopkins University). The insert was excised through digestion with MluI and ClaI, whose sites are present in the Sal1 adapter at the insert's 5'-end and in the insert's 3'-untranslated sequence, respectively. The excised insert was subcloned into pCB6 at the corresponding sites.
Cell Surface Biottnylation-Cells were plated at confluent density on 0.45-pm filter inserts (Transwell, Costar Co., Cambridge, MA). Seven days after seeding, GAT-1-transfected MDCK cells were biotinylated through exposure to NHS-ss-biotin present in the medium bathing the apical or basolateral surfaces. The biotinylation protocol employed was that of Sargiacomo et al. (22), with the exception that the reaction was carried out at pH 9.0 in 10 m~ triethanolamine, 2 mM CaCl,, 125 mM NaCI. We have demonstrated previously that this modification in- creases the efficiency of NHS-ss-biotin incorporation (23). Following biotinylation, filters were excised from the cups with a razor blade, and the attached monolayers were lysed in 1% Triton X-100 in 150 mM NaCl, 5 mM EDTA, 50 m~ Tris, pH 7.5. Biotinylated proteins were recovered from cell lysates by incubation with avidin-agarose beads (Pierce) as described (22). Bound proteins were eluted from the beads in Laemmli sample buffer (241, separated by SDS-polyacrylamide gel electrophore- sis and transferred to nitrocellulose. Transfers were probed with anti- body R24 at a dilution of 1:200 as described (19). Specific staining was visualized by the Enhanced Chemiluminescence technique (Amersham Corp.). For quantitation of the biotinylation experiments, autoradio- graphs were scanned with an LKB Ultrascan XL laser densitometer. Multiple samples were analyzed, corresponding to different quantities of total protein. In this manner, the linearity of the signal with respect to sample load was verified.
GABA Uptake Assay-Transfected and untransfected MDCK cells were grown to confluence for >5 days on 0.45-pm pore size Transwell filter inserts. GABAuptake was performed at room temperature accord- ing to a modification of the method of Yamauchi et al. (25). T h e filters were washed twice with uptake buffer (100 mM NaCl, 2 mM KCI, 1 m~ CaCl,, 1 m~ MgCI,, 10 m~ HEPES, pH 7.5), and each well was incubated from the apical or basolateral side with 0.2 ml of uptake buffer containing PHIGABA (DuPont NEN) at the final concentrations indicated in the figure legends. Following a 10-min incubation the uptake assay was ter- minated by aspirating the medium, and the filters were successively dipped into three beakers, each of which contained 100 ml of ice-cold uptake buffer. The filters were excised from the cups and placed into scintillation vials, where the attached epithelial cells were solubilized in 0.2 ml of 1% SDS. Protein was determined with the BCA protein assay
kit (Pierce) as described previously (25). The cell lysates were counted in 5 ml of Atomlight scintillation fluid (DuPont NEN).
RESULTS
Axonal Localization of the GABA Dansporter Endogenously Expressed in Polarized Hippocampal Neurons in Culture- Prior to examining the distribution of GABA transporter ex- pressed by transfection in epithelial cells, we wished to deter- mine whether this protein is restricted to the axolemma of polarized hippocampal neurons in culture. An antibody di- rected against a synthetic peptide derived from the carboxyl- terminal sequence of the GABA transporter (R24) was provided by R. Jahn. Indirect immunofluorescence employing the R24 anti-GABA transporter antibody was carried out on 7- and 14-day-old cultures of rat hippocampal neurons (gift of G. Banker). The identity of dendritic processes was established by double labeling with an antibody directed against the dendritic marker MAP-2 (26) (Fig. 1, a and c ) . The distribution of the GABA transporter is shown in Fig. 1, b and d . In Fig. 1, a and b, two groups of 7-day neurons, consisting of four and three cells on the the right bottom and left sides, respectively, are recognizable by their MAP-2 staining. A long, thin and branched process emanates from one of the neurons on the left side of Fig. l b (empty arrowhead). This process is brightly stained with the GABA transporter antibody (arrows) and negative for the dendritic marker (compare arrows in b with a ) . The GABA transporter-positive process follows a dendrite along its entire length and seems to ramify twice in descending branches (asterisks) which surround the group of cells on the bottom right of the figure. The process extends beyond the tip of the colinear dendrite and terminates shortly after the second ramification (double arrows). The process is recognizable as an axon both by its morphology (i.e. it is thinner and longer than neighboring processes) and by its lack of MAP-2 staining. In dense neuronal cultures it is difficult to find dendrites that are entirely free of axons running along their surfaces. There are, however, processes recognizable as dendrites by both their mor- phology and MAP-2 reactivity which are clearly negative when stained with the GABAtransporter antibody R24 (filled arrow- heads, compare a with b) . Although some of the dendrites are stained with both MAP-2 and R24, the patterns of staining with the two antibodies are quite distinct. MAP-2 immunostaining. appears to fill the cytoplasm, as would be expected for a cyto- plasmic antigen. In contrast, the GAT-1 staining pattern is fibrillar and lacy, consistent with its origin in individual axons running along and criss-crossing over the surfaces of dendrites. In older neurons (Fig. 1, c and d ) the complex network of axons and axon bundles that originate at GABA transporter-positive neurons runs along most of the dendritic surfaces of the neuron depicted and are clearly detected by the GABA transporter antibody. It should be noted that only a small fraction (-10%) of the cultured hippocampal neurons exhibited positive stain- ing for GABA transporter. This observation is consistent with the fact that only a minority population of hippocampal neu- rons is GABA-ergic (27). No staining of the occasional glial cells present in the primary cultures of the hippocampal neurons was ever noticed, consistent with the exclusively neuronal lo- calization of the GAT-1 transporter (data not shown). Thus, in culture as well as in situ the GABA transporter is restricted in its distribution to the axonal plasma membrane (14).
Apical Expression of the GABA Dansporter in Stably Duns- fected MDCK Cells-We next determined whether the sorting signals that specify this axonal localization would function to mediate apical targeting in polarized epithelial cells. The :DNA encoding the rat brain GABA transporter (kind gift of B. Kan- ner) was subcloned into the pCB6 mammalian expression vec- tor and used to generate stably transfected MDCK cells, as

4670 GABA Dansporter Is Sorted to Epithelial Apical Membranes
7 (panels a and h ) and 14 (panels c and d ) days were double labeled with a monoclonal antibody directed against MAF"2 (panels a and c ) and the FIG. 1. Immunolocalization of the GABA transporter in polarized hippocampal neurons in culture. Hippocampal neurons cultured for
GAFJA transporter-specific polyclonal antibody R24 (panels b and d ). Long and thin processes recognizable as axons by their morphology and negativity for MAP-2 are positively stained with R24 (arrows). The double arrows indicate an axon running along the surface of a dendrite and beyond the dendrite termination. MAP-2-positive processes (dendrites) can be found which are devoid of GABA transporter staining (arrowheads). Empty arrowheads indicate the cell body of a neuron from which a GAFJA transporter-positive axon departs. Asterisks follow an axon's ramifica- tions. Axon bundles are denoted by ax. Bar: panels a and b, 65 pm; panels c and d , 25 pm).
described under "Materials and Methods." MDCK cells are de- rived from the renal distal tubule and retain their parent tis- sue's polarized phenotype in culture (1-3). Cell lines stably expressing the GABA transporter were initially identified in GABA uptake assays, and their identities were confirmed by Western blotting. The GABA transporter antibody used in these studies does not cross-react with any endogenous pro- teins in untransfected MDCK cells, as demonstrated both by immunofluorescence and Western blot analysis. When antibody R24 was used to probe Western blots of total cellular homog- enates, a broad band of the expected molecular mass (-85 kDa) was detected only in material derived from the transfected cells (data not shown). Similarly, no immunofluorescent labeling could be detected when R24 was used to stain untransfected cells (not shown). Thus, the antibodies employed in these stud- ies are specific for the GABA transporter and detect this pro- tein expressed exogenously in epithelial cells.
The subcellular dietribution of thc CARA trnnsporter in transfected MDCK cells was established by immunologic and functional assays. Double label immunofluorescence was car- ried out using tho antibody directed against the GABA trans- porter as well as an antibody specific for the a-subunit of the Na,K-ATPase (Fig. 2). Labeled cells were examined both enface
(Fig. 2, bottom) and in X Z cross-sections generated by confocal microscopy (Fig. 2, top). As can be seen in Fig. 2u, the GABA transporter is restricted to the apical surfaces of transfected MDCK cells (up) . In contrast, the Na,K-ATPase retains its normal basolateral localization (28) (Fig. 2b), demonstrating that the transfected cells remain appropriately polarized. The same predominantly apical localization has been revealed in experiments employing R21, an antipeptide polyclonal anti- body directed against the amino terminus of GAT-1 (data not shown).
.The apical localization of the GABA transporter was further supported hy the cell surface biotinylation experiments and by the flux studies presented in Figs. 3 and 4, respectively. The steady-state hiotinylation experiment presented in Fig. 3 dem- onstrates that the GABA transporter is available to cell surface biotinylation predominantly from the apical side. A broad band of the molecular mass expected of the mature fully glycosylated transporter ( ~ 85 kDa) was readily detected in Western blots of material recovered from nvidin bend precipitntions when the NHS biotin reagent was added to the apical surface (panel A ). Little if any 85-kDa protein was detected when the biotinyla- tion was performed at the basolateral surface (panel B ) . The same pattern was observed when the amino-terminal R21 an-
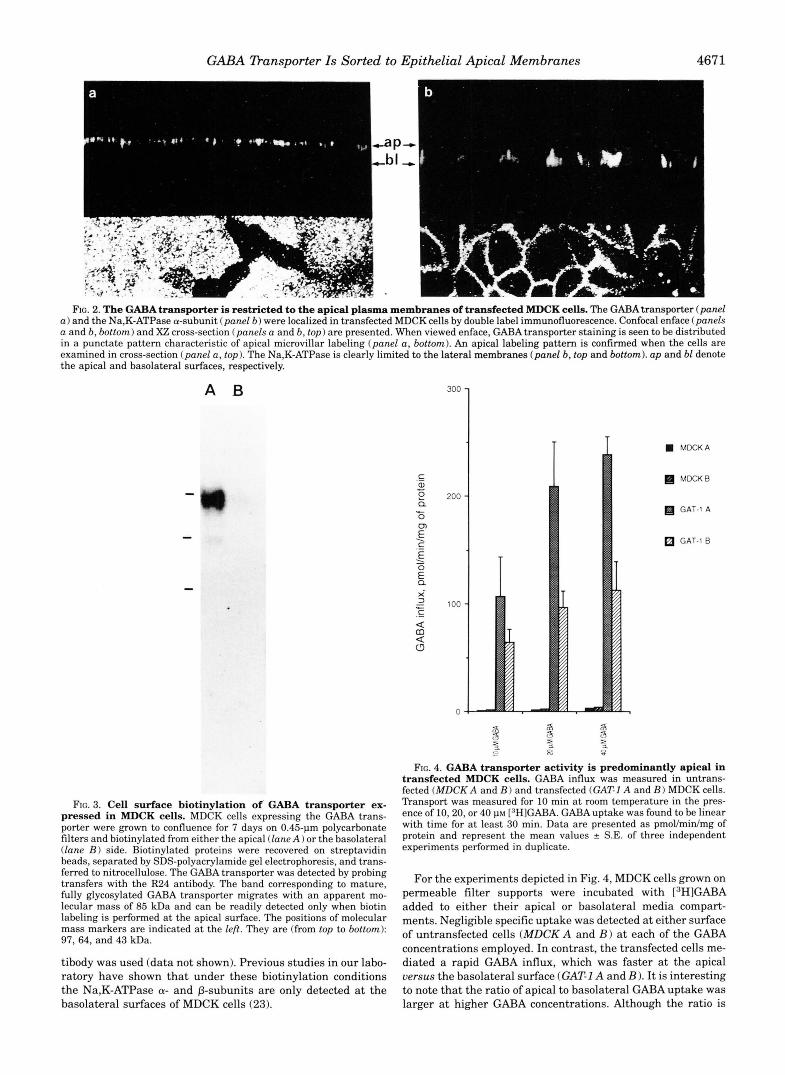
GABA Dansporter Is Sorted to Epithelial Apical Membranes 4671
FIG. 2. The GABA transporter is restricted to the apical plasma membranes of transfected MDCK cells. The GABA transporter (panel a ) and the Na,K-ATPase a-subunit (panel 6 ) were localized in transfected MDCK cells by double label immunofluorescence. Confocal enface (panels a and 6 , bottom) and X Z cross-section (panels a and b, t o p ) are presented. When viewed enface, GABA transporter staining is seen to be distributed in a punctate pattern characteristic of apical microvillar labeling (panel a, bottom). An apical labeling pattern is confirmed when the cells are examined in cross-section (panel a, top). The Na,K-ATPase is clearly limited to the lateral membranes (panel b, top and bottom). up and bl denote the apical and basolateral surfaces, respectively.
A B 300
200
100
0
T T W MDCKA ~
MDCKB
GAT-1 A
GAT-1 B
FIG. 3. Cell surface biotinylation of. GABA transporter ex- pressed in MDCK cells. MDCK cells expressing the GABA trans- porter were grown to confluence for 7 days on 0.45-pm polycarbonate filters and biotinylated from either the apical (lane A ) or the basolateral (lane B) side. Biotinylated proteins were recovered on streptavidin beads, separated by SDS-polyacrylamide gel electrophoresis, and trans- ferred to nitrocellulose. The GABA transporter was detected by probing transfers with the R24 antibody. The band corresponding to mature, fully glycosylated GAl3A transporter migrates with an apparent mo- lecular mass of 85 kDa and can be readily detected only when biotin labeling is performed at the apical surface. The positions of molecular
97,64, and 43 kDa. mass markers are indicated at the left. They are (from top to bottom):
tibody was used (data not shown). Previous studies in our labo- ratory have shown that under these biotinylation conditions the Na,K-ATPase a- and P-subunits are only detected at the basolateral surfaces of MDCK cells (23).
FIG. 4. GABA transporter activity is predominantly apical in transfected MDCK cells. GABA influx was measured in untrans- fected (MDCK A and B) and transfected (GAT-I A and B 1 MDCK cells. Transport was measured for 10 min a t room temperature in the pres- ence of 10.20, or 40 p~ [3HIGABA. GABA uptake was found to be linear with time for at least 30 min. Data are presented as pmol/min/mg of protein and represent the mean values f S.E. of three independent experiments performed in duplicate.
For the experiments depicted in Fig. 4, MDCK cells grown on permeable filter supports were incubated with 13H1GABA added to either their apical or basolateral media compart- ments. Negligible specific uptake was detected at either surface of untransfected cells (MDCKA and B) at each of the GABA concentrations employed. In contrast, the transfected cells me- diated a rapid GABA influx, which was faster at the apical versus the basolateral surface (GAT1 A and B ) . It is interesting to note that the ratio of apical to basolateral GABA uptake was larger at higher GABA concentrations. Although the ratio is

4672 GABA Dansporter Is Sorted to Epithelial Apical Membranes
- (P
0 c
c - 0
100
80
60
40
20
0
P 19 P
19 4 4
4 m
3
r
FIG. 5. Quantitation of the polarized distribution of the GABA transporter. The GABA transporter surface distributions determined functionally (GABA uptake A and E ) or through the biotinylation assay (Biotinylation A and B ) are presented above. GABA influx measure- ments from the apical ( A ) or the basolateral ( E ) surfaces were camed out in the presence of 40 VM GABA for 10 min at room temperature. Values are presented as the percent of total cell surface transporter activity and represent means e S.E. of three independent experiments performed in duplicate. Quantitation of the accessibility of GABA trans- porter to biotinylation from the apical or the basolateral surfaces was performed as described under “Materials and Methods.” Linearity of the biotinylation assay was determined by examining several dilutions of samples from labeled cells. Bars indicate S.E.
- 1.6 when the GABA concentration is 10 PM (which is close to the transporter’s expected K,,, of 4-5 1.1~ (29)), the ratio is >2 at 40 p~ GABA. It remains to be determined whether this dis- crepancy is the product of technical factors relating to the ge- ometry of the assay or if it in fact represents actual differences in the properties of transporters localized to the two surface domains.
A comparison of measurements of functional and biochemical polarity is presented in Fig. 5. The apical to basolateral polarity ratio revealed by cell surface biotinylation experiments was determined by densitometric scanning of autoradiographs of Western blots, as described under “Materials and Methods.” Although steady-state cell surface biotinylation suggests a po- larity ratio of -6.6, GABA uptake measurements indicate a ratio of -2.4, In spite of the method to method variation, how- ever, these data demonstrate that the GABA transporter be- haves as a predominantly apical protein when expressed by transfection in polarized MDCK cells. Finally, it should be noted that this phenomenon is not unique to MDCK cells. We have found that LLC-PK1 cells (derived from the pig kidney
100
80
60
40
20
0
I
eral in stably transfected MDCK cells. GABAuptake was measured FIG. 6. Betaine transporter activity is predominantly basolat-
in untransfected (MDCK-A and B ) and transfected (BGT-IA and E ) MDCK cells. Values represent three different experiments performed in duplicate for 30 min at room temperature with a GABA concentration of 100 PM. The assay is linear with time for at least 40 min. The S.E. is indicated by the bars.
cortex (30)) also target the GABA transporter to their apical plasma membranes (data not shown).
The Betaine Tkansporter Behaves as a Basolateral Protein in Tkansfected MDCK Cells-The Na,Cl-dependent betaine trans- porter is endogenously expressed at the basolateral surfaces of MDCK cells that have been exposed to hyperosmotic stress (25). Recent cloning studies demonstrate that this protein be- longs to the GABA transporter family and is 50% identical to GAT-1 at the amino acid level (15). It has also been shown that the betaine transporter can utilize GABA as a substrate for transport (15). In fact, the betaine transporter’s affinity for GABA (K,,, = -100 1.1~) is higher than that for betaine ( K , = -500 p ~ ) . We have used GABA uptake assays to assess the distribution of betaine transporter expressed by transfection in MDCK cells that have been grown under conditions of normal osmolarity. As can be seen in Fig. 6, untransfected cells express low levels of endogenous GABA transport activity (see also Fig. 4). In contrast, cells transfected with a cDNA encoding the betaine transporter (kind giR of J. Handler) express readily detectable GABA uptake, -80% of which is present at the ba- solateral surface. It would appear, therefore, that when ex- pressed by transfection as well as under conditions of hyperos- molarity, the betaine transporter behaves as a basolateral membrane protein.

GABA Dansporter Is Sorted to Epithelial Apical Membranes 4673
DISCUSSION
Previous comparisons of neuronal and epithelial membrane protein sorting have examined the distributions in cultured hippocampal neurons of proteins whose epithelial sorting be- havior had been established earlier (5, 6). The studies pre- sented here have extended this approach by examining the sorting properties of a neuronal protein, the GAT-1 isoform of the Na,Cl-dependent GABA transporter family, exogenously ex- pressed in epithelial cells. We first determined the localization of the neuronal transporter in mature, fully polarized hippo- campal neurons. We find that expression of the transporter is restricted to the axonal surfaces of the cultured neurons. This result is consistent with previous observations on the distribu- tion of GABA transporter in intact rat hippocampus (14).
To compare the sorting behavior of the GABA transporter expressed in neurons and epithelial cells, we stably transfected an epithelial cell line (MDCK) with the cDNA encoding GAT-1. The subcellular distribution of the GABA transporter was es- tablished by both immunochemical and functional methods. Our results demonstrate that MDCK cells express GAT-1 in a polarized manner and that the protein is concentrated pre- dominantly at the apical plasma membrane. There is a discrep- ancy between the apical to basolateral polarity ratios deter- mined by functional techniques, on the one hand, and biochemical methods on the other. When measured by GABA uptake assay, the distribution of the transporter appeared to be only about half as polarized as would be inferred from the results of the immunochemical experiments. Whether this phe- nomenon reflects actual differences in the kinetic behavior of the transporters inserted in the two surface domains or is sim- ply the product of assay conditions remains to be determined. In either event, our observations demonstrate that MDCK cells accumulate the GABA transporter at their apical surfaces, con- sistent with the hypothesis that the mechanisms involved in axonal targeting in neurons are functionally related to those that mediate apical sorting in epithelia.
In the context of this apparent rule equating the axonal and apical sorting pathways, it is interesting to note that the Na,K-ATPase comprises a puzzling exception. We have found that cultured hippocampal neurons express two isoforms of the Na,K-ATPase a-subunit (a1 and a3) (17). Both of these proteins are restricted to the basolateral surfaces of polarized epithelial cells. Surprisingly, they are also present in the axo- nal as well as in the dendritic domains of the neuronal plasma membrane. This apparent inconsistency might be explained by the involvement of cytoskeletal elements in determining or stabilizing membrane protein distributions. The Na,K-ATPase has been shown to interact directly with ankyrin and, conse- quently, to be associated with the fodrin-based subcortical cy- toskeleton (32). Although ankyrin and fodrin are restricted to the basolateral surfaces of polarized MDCK cells (33), iso- forms of these proteins have been found in both the axom and dendrites of neurons (34, 35). Studies on the targeting of the Na,K-ATPase suggest that in a thyroid epithelial cell line (FRT) (36) as well as in at least one clone of MDCK cells (23, 28) the sodium pump is delivered vectorially to the basolateral surface. Experiments performed on a different MDCK sub- clone, however, indicate that in this cell line Na,K-ATPase is randomly delivered to both cell surface domains (37). In this system, the sodium pump’s steady-state polarized distribution appears to be the product of selective stabilization at the baso- lateral plasmalemma produced through interactions with the cytoskeleton (38). If Na,K-ATPase sorting in neurons is simi- larly dependent upon selective stabilization, the presence of cytoskeletal elements in both types of neuronal processes may explain the sodium pump’s nonpolarized distribution. I t is
possible, therefore, that the relationship between neuronal and epithelial sorting may extend to a number of distinct classes of membrane proteins.
Finally, it is worth noting that another member of the GABA transporter’s gene family is endogenously expressed by MDCK cells. The epithelial cells of the renal distal tubule are bathed in an interstitial fluid which can become extremely hyperosmolar. To adapt to this environment, renal epithelial cells import as well as manufacture osmotically active compounds which allow the cells to hold on to cytosolic water (39). Betaine, whose structure is related to that of GABA, serves as one of these osmolytes (40). Hyperosmotic stress induces distal tubule epi- thelial cells to synthesize a Na,Cl-dependent betaine carrier which is -50% identical at the amino acid level to the GABA transporter (15, 40). Recent studies reveal that MDCK cells grown under hyperosmotic conditions express this protein and target it to their basolateral surfaces (25). Our data demon- strate that the betaine transporter is also basolateral when expressed by transfection in cells grown under normal osmotic conditions. Thus, despite their high degree of homology, the GABA and betaine transport systems are differentially sorted in epithelial cells. We are in the process of generating chimeras between these two transport systems. Analysis of the sorting behavior of the resulting constructs should allow us to identify the residues that contribute to these molecules’ sorting signals and to determine if these signals are, in fact, identical in epi- thelia and in neurons.
Acknowledgments-We are grateful to Drs. P. de Camilli, R. Jahn, G. Banker, B. Kanner, J. Handler, and J. Ahn for helpful discussions and for providing reagents. We also thank Dr. N. Borgese (in whose labora- tory a portion of this work was completed) for hospitality and helpful suggestions.
REFERENCES
2. 1.
3.
4.
6. 5.
7. 8. 9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
20. 19.
21.
22.
23. 24. 25.
26. 27.
Rodriguez-Boulan, E., and Nelson, W. J. (1989) Science 246,718-725
Caplan, M. J., and Matlin, K. S. (1989) in Functionul Epithelial Cells in Simons, K., and Fuller, S. D. (1985) Annu. Reu. Cell Biol. 1, 295-340
Culture (Matlin, K. S., and Valentich, J. D., eds) pp. 71-127, Alan R. Liss. New York
Black, M. M., and Baas, P. W. (1989) 2hvd.s Neurosci. 12,211-214 Dotti, C. G., and Simons, K. (1990) Cell 62,63-72 Dotti, C. G., Parton, R. G., and Simons, K. (1991) Nature 349, 15E-161 Iversen, L. L. (1971) Br J. Pharmacol. 41, 571-591 Radian, R., and Kanner, B. I. (1983) Biochemistry 22, 12361241 Guastella, J., Nelson, N., Nelson, H., Czyzyk, L., Kenyan, S., Miedel, M.,
Davidson, N., Lester, H., and Kanner, B. (1990) Science 249,1303-1306 Borden, L. A,, Smith, K. E., Hartig, P. R., Branchek, T. A., and Weinshank, R.
L. (1992) J. Biol. Chem. 267,2109&21104 Clark, J. A., Deutch, A. Y., Gallipoli, P. Z., and Amara, S. G. (1992) Neuron 9,
337448 Liu, Q. R., Lopez-Corcuera, B., Mandiyan, S., Nelson, H., and Nelson, N. (1993)
J. Biol. Chem. 288,21OCM?112 Mabjeesh, N. J., Frese, M., Rauen, T., Jeserich, G., and Kanner, B. I. (1992)
FEES Lett. 299.99-102 Radian, R., Ottersen, 0. P., Storm-Mathisen, J., Castel, M., and Kanner, B.
(1990) J. Neurosci. 10, 1319-1330 Yamauchi, A., Uchida, S., Kwon, H. M., Preston, A. S., Robey, R. B.,
Garcia-Perez, A,, Burg, M. B., and Handler, J. S. (1992) J. Biol. Chem. 267, 649-652
Goslin, K., and Banker, G. (1991) in Culturing Nerue Cells (Banker, G., and
Pietrini, G., Matteoli, M., Banker, G., and Caplan, M. J. (1992) Proc. Natl. Goslin, K., eds) pp. 251-281, MIT Press, Cambridge, MA
Lerner, R. A,, Green, H., Alexander, F., Liu, T., Sutcliffe, G., and Shinnick, T. Acad. Sci. U. S. A. 89, 8414-8418
Gottardi, C. J., and Caplan, M. J. (1993) J. Cell B i d . 121, 283-293 M. (1981) Proc. Natl. Acad. Sci. U. S. A. 78, 34033407
Brewer, C. B., and Roth, M. G. (1991) J. Cell Biol. 114, 413421 Puddington, L., Woodgett, C., and Rose, J. K. (1987) Proc. Natl. Acad. Sei.
Sargiacomo, M., Lisanti, M., Graeve, L., Le Bivic, A,, and Rodriguez-Boulan, E.
Gottardi, C. J., and Caplan, M. J. (1993) Science 260, 552-554 Laemmli, U. K. (1970) Nature 227,680-685 Yamauchi, A,, Kwon, H. M., Uchida, S., Preston, A,, and Handler, J. S. (1991)
Caceres, A,, Banker, G. A., and Binder, L. (1986) J. Neurosci. 6, 714-722 Mugnaini, E., and Oertel, W. H. (1985) in Handbook of Chemical Neuro-
anatomy (Bjorklund, A., and Hokfelt, T., eds) Vol. IV, Part I, Elsevier Science Publishing Co., Amsterdam
U. S. A. 84,2756-2760
(1989) J. Membr Biol. 107, 277-286
Am. J. Physiol. 261, F197-F202

4674 GABA nansporter Is Sorted to Epithelial Apical Membranes 28. Caplan, M. J., Anderson, H. C., Palade, G. E., and Jamieson, J. D. (1986) Cell
29. Keynan, S., Suh, Y. J., Kanner, B. I., and Rudnick, G. (1992) Biochemistry 51,
30. F'faller, W., Gstraunthaler, G., and Loidl, P. (1990) J. Cell. Physiol. 142, 247-
31. Deleted in prwf 32. Nelson, W. J., and Veshnock, P. J. (1987) Nature 528,533-536 33. Morrow, J. S. , Cianci, C. D., Ardito, T., Mann, A. S. , and Kashgarian, M. (1989)
34. Kordeli, E., and Bennett, V. (1991) J. Cell Biol. 114, 1243-1259
46,623-631
1974-1979
254
J. Cell Biol. 108,455-465
35. Kunimoto, M., Otto, E., and Bennett, V. (1991) J. Cell Bwl. 115,
36. Zunolo, C., and Rodriguez-Boulan, E. (1993) Science 280,55&552 37. Siemers, K. A,, Wilson, R., Mays, R. W., Ryan, T. A,, Wollner, D. A,, and Nelson,
38. Hammerton, R. W., Krzemiski, K A., Mays, R. W., Ryan, T. A,, Wollner, D. A,,
39. Bagnasco, S. , Balaban, R., Fales, H. M., Yang, Y., and Burg, M. (1986) J. Biol.
40. Nakanishi, T., Turner, R. J . , and Burg, M. B. (1990) Am. J. Physiol. 258.
1319-1331
W. J. (1993) Science 260,552456
and Nelson, W. J. (1991) Science 254,847450
Chem. 261,5872-5877
F1061-F1067