the j b c vol. 278, no. 30, issue of july 25, pp. 27502 ... · the obligate predatory bdellovibrio...
TRANSCRIPT

The Obligate Predatory Bdellovibrio bacteriovorus Possesses aNeutral Lipid A Containing �-D-Mannoses That ReplacePhosphate ResiduesSIMILARITIES AND DIFFERENCES BETWEEN THE LIPID As AND THE LIPOPOLYSACCHARIDES OF THEWILD TYPE STRAIN B. BACTERIOVORUS HD100 AND ITS HOST-INDEPENDENT DERIVATIVE HI100*
Received for publication, March 24, 2003, and in revised form, May 7, 2003Published, JBC Papers in Press, May 12, 2003, DOI 10.1074/jbc.M303012200
Dominik Schwudke‡§, Michael Linscheid‡¶, Eckhard Strauch§, Bernd Appel§, Ulrich Zahringer�,Hermann Moll�, Mareike Muller�, Lothar Brecker�, Sabine Gronow�, and Buko Lindner�
From the ‡Department of Chemistry, Humboldt Universitat zu Berlin, D-12489 Berlin, Germany,the §Project Group Biological Safety, Robert Koch Institute Berlin, D-13353 Berlin, Germany,and the �Research Center Borstel, Center for Medicine and Biosciences, D-23845 Borstel, Germany
Bdellovibrio bacteriovorus are predatory bacteria thatpenetrate Gram-negative bacteria and grow intrap-eriplasmically at the expense of the prey. It was suggestedthat B. bacteriovorus partially degrade and reutilize li-popolysaccharide (LPS) of the host, thus synthesizing anouter membrane containing structural elements of theprey. According to this hypothesis a host-independentmutant should possess a chemically different LPS. There-fore, the lipopolysaccharides of B. bacteriovorus HD100and its host-independent derivative B. bacteriovorusHI100 were isolated and characterized by SDS-polyacryl-amide gel electrophoresis, immunoblotting, and massspectrometry. LPS of both strains were identified assmooth-form LPS with different repeating units. The lipidAs were isolated after mild acid hydrolysis and theirstructures were determined by chemical analysis, bymass spectrometric methods, and by NMR spectroscopy.Both lipid As were characterized by an unusual chemicalstructure, consisting of a �-(136)-linked 2,3-diamino-2,3-dideoxy-D-glucopyranose disaccharide carrying six fattyacids that were all hydroxylated. Instead of phosphategroups substituting position O-1 of the reducing and O-4�of the nonreducing end �-D-mannopyranose residueswere found in these lipid As. Thus, they represent the firstlipid As completely missing negatively charged groups. Areduced endotoxic activity as determined by cytokine in-duction from human macrophages was shown for thisnovel structure. Only minor differences with respect tofatty acids were detected between the lipid As of the host-dependent wild type strain HD100 and for its host-independent derivative HI100. From the results of thedetailed analysis it can be concluded that the wild typestrain HD100 synthesizes an innate LPS.
Bdellovibrionaceae were discovered in 1962 by their lyticactivity against Gram-negative bacteria in experiments de-
signed for the isolation of bacteriophages in soil samples (1).They are small, motile Gram-negative bacteria and possess apredatory lifestyle that includes an obligate growth and repli-cation phase taking place in the periplasm of the prey. Bdell-ovibrionaceae are widely spread in the environment, e.g. soil,marine sediments, rhizosphere of plants, sewage, etc. (1–4). Inthe latter environment it was found that Bdellovibrionaceaeare involved in the reduction of bacterial counts thus support-ing the self-purification of domestic waste waters (4, 5). Fur-thermore, Bdellovibrionaceae were found in the intestinal tractof mammals and might be important for the reduction of path-ogenic bacteria in this environment (6, 7).
Despite the unique predatory lifestyle and common morpho-logical features, Bdellovibrionaceae show a great phylogeneticdiversity based on 16 S rRNA analyses. Bdellovibrionaceae aredivided into the species Bdellovibrio bacteriovorus, Bacterio-vorax stolpii, Bacteriovorax starrii, and some strains yet to beassigned (3, 7, 8). The strain B. bacteriovorus HD1001 investi-gated in this work is a reference strain, which was isolatedfrom soil (1).
B. bacteriovorus possesses a life cycle consisting of an attackphase followed by attachment and invasion of the periplas-matic space of the prey bacterium. After penetration into theperiplasmatic space of the prey the peptidoglycan layer of thehost is partially degraded in a short period of time leading toswelling of the prey bacteria and the formation of sphericalbdelloplasts. The former outer membrane of the prey forms abarrier against the surrounding environment and thus retainsthe available nutrients in a confined space (9, 10). Inside theprey elongation and multiplication take place and finally theprey bacteria are lysed (11, 12). B. bacteriovorus wild typestrains solely grow on living bacteria. However, in a multistepselection procedure including streptomycin tolerance asmarker host-independent (HI) mutants can be isolated thatgrow slowly on rich media but show a number of aberrantmorphological features (13, 14).
* The work was supported by Deutsche ForschungsgemeinschaftGrants LI 448/1-1 (to B. L.), SFB 470-A1 (to S. G.), SFB 470-B4 (toU. Z.), and LI 309/22-1 (to M. L.) and the Fonds der ChemischenIndustrie (FCI). The costs of publication of this article were defrayed inpart by the payment of page charges. This article must therefore behereby marked “advertisement” in accordance with 18 U.S.C. Section1734 solely to indicate this fact.
¶ To whom correspondence should be addressed: Professor of AnalyticalChemistry, Humboldt Universitat zu Berlin, Dept. of Chemistry, Brook-Taylor-Str. 2, 12489 Berlin, Germany. Tel.: 49-0-30-2093-7575; Fax:49-0-30-2093-6985; E-mail: [email protected].
1 The abbreviations used are: HD, host-dependent; HI, host-independ-ent; COSY, correlation spectroscopy; DQF, double quantum filter; ESI-FT, electrospray ionization Fourier transformation; FT-IR, Fouriertransform infrared spectroscopy; HMBC, heteronuclear multiple bondcoherence spectroscopy; hMNC, human mononuclear cells; HMQC, het-eronuclear multiple quantum coherence spectroscopy; LPS, lipopolysac-charide; mAb, monoclonal antibody; MALDI, matrix-assisted laser de-sorption ionization; MS, mass spectrometry; ROESY, rotating frameOverhauser effect spectroscopy; TNF-�, tumor necrosis factor-�;TOCSY, total correlation spectroscopy.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 278, No. 30, Issue of July 25, pp. 27502–27512, 2003© 2003 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org27502
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

In former studies enzymatic activity of host-dependent (HD)B. bacteriovorus against the cell wall of Gram-negative bacteriaincluding the LPS of the outer membrane was detected (15, 16).However, in addition to the degradation of macromolecularcompounds of the prey several studies also indicated thatB. bacteriovorus reutilizes outer membrane proteins, lipid A,and fatty acids of the prey bacteria by integration into itsmembrane system (15–19). To understand the cell wall degrad-ing mechanisms detailed information about its own membranesystem is needed, as the predatory lifestyle requires that itsown cell wall is protected against degradation. Nelson andRittenberg (18) detected two different lipid A species in LPSpreparations from host-dependent B. bacteriovorus 109J bythin layer chromatography (TLC). One lipid A showed moresimilarity to that of the prey bacteria whereas the other sharedcommon features with the lipid A from a host-independentB. bacteriovorus strain. Chemical analysis revealed a nonade-cenoic acid and (OH)-13:0 as characteristic fatty acids, andfurthermore, glucosamine was determined as constituent of thelipid A backbone. However, these results did not include acomplete structural description of lipid A. The structural de-termination of LPS and lipid As derived form wild typeB. bacteriovorus (HD100) and its derivative (HI100) is a pre-requisite to elucidate the molecular interaction between pred-ator and prey bacteria. The structures of the lipid As of thehost-dependent strain B. bacterivorus HD100 and its host-independent mutant HI100 were determined in detail and cyto-kine release were measured for assessment of biological activity.
EXPERIMENTAL PROCEDURES
Bacterial Strains and Culture Condition—The host-dependentB. bacteriovorus HD100 (DSM 50701) was grown on Escherichia coliK12 (DSM 423) as described earlier (7). The host-independent mutantB. bacteriovorus HI100 (DSM 12732) was grown in PYE medium (ATCCmedium 526) at 30 °C for 3 to 5 days (7).
LPS Isolation—The bacterial pellets were washed twice with 1%phenol, ethanol, and acetone and than dried at room temperature. Forthe host-dependent strain the yield was 4 g (pooled from a 22-literculture) and for the host-independent strain 3.8 g (pooled from a 7-literculture). After enzymatic degradation of nucleic acids and incubationwith proteinase K the phenol/chloroform/light petroleum, 2:5:8 (v/v/v),method was used for LPS extraction (20).
Water was added to the extract of the host-dependent strain HD100until a fraction H1 precipitated. The precipitate was collected by cen-trifugation. The remaining supernatant was concentrated until thephenol crystallized on ice water. A second LPS fraction, H2, was ob-tained by adding ethanol to the remaining solution on ice water and theprecipitate was collected by centrifugation. The LPS of the host-inde-pendent strain HI100 could only be precipitated with ethanol. All threeLPS fractions were washed with acetone three times and dried. Theyield was 206 mg for H1 and 110 mg for H2. From the host-independentstrain HI100 190 mg of LPS was isolated. For further purification thecrude LPS fractions were resuspended in twice distilled water andproteinase K was added to a final concentration of 100 �g ml�1 andincubated at 37 °C overnight. These suspensions were dialyzed againsttwice distilled water and the LPS solutions of about 12 mg ml�1 werecentrifuged at 100,000 � g. The sediments were lyophilized giving ayield of 161.5 mg of LPS of H1, 79.6 mg of LPS of H2, and 111.4 mg ofLPS of B. bacteriovorus HI100.
Isolation of Lipid A—Free B. bacteriovorus lipid As were obtained byhydrolysis of LPSs (49.6 mg of HD100, 52.7 mg of HI100) with 1% aceticacid for 90 min at 100 °C. Both lipid As were centrifuged and fraction-ated on a silica gel column (Silica Gel F60, Merck) with chloroform,chloroform/methanol, 9:1 (v/v), chloroform/methanol, 8:2 (v/v), and chlo-roform/methanol, 1:1 (v/v), each. The preparations were analyzed byTLC (Silica Gel 60 F254 plate, Merck; CHCl3/CH3OH/H2O, 100:75:15(v/v/v)) and both lipid As were found in the respective last fractions. Thelipid A yield of strain HD100 was 7.3 and 8.1 mg of strain HI100.
Fatty Acid Analysis—Fatty acids were released and converted totheir methyl esters with HCl in methanol (for determination of totalfatty acid: 2 M HCl, 24 h, 120 °C; for ester-linked fatty acids: 0.5 M HCl,30 min, 85 °C). Fatty acid residues were extracted with chloroform andincubated with diazomethane to obtain the methylated products. After
treatment with bis(trimethylsilyl)trifluoroacetamide the trimethylsil-ylethers of the fatty acids were analyzed by GC-MS. The absoluteconfiguration of the hydroxy fatty acids was determined by GC-MS ofthe 1-phenylethylamide derivatives (21). GC-MS was performed on aHewlett Packard mass spectrometer 5989A equipped with a fused silicacapillary column (HP-5MS: 30 m, inner diameter: 0.25 mm, film thick-ness 0.25 �m). Helium served as carrier gas and the GC temperaturewas initially 150 °C for 3 min, then raised to 320 °C at 5 °C min�1.Electron impact was carried out at 70 eV and chemical ionization massspectra (chemical ionization-MS) were recorded with ammonia as reac-tant gas (0.1 kilopascal).
Sugar Analysis—Qualitative examination of the sugar portion ofLPS was performed by methylation (0.5 M HCl/CH3OH, 45 min, 85 °C,and 2 M HCl/CH3OH, 16 h, 85 °C) followed by peracetylation (22). Theabsolute configuration of the mannose and 2,3-diamino-2,3-dideoxyglu-cose of the lipid A from B. bacteriovorus HD100 was determined withthe acetylated (R)-2-butylglycosides (23). The parameters of the GC-MSanalysis were identical as described for fatty acid analysis.
Matrix-assisted Laser Desorption Ionization-Time of Flight MassSpectrometry (MALDI-TOF MS)—MALDI-TOF MS of LPS and lipid Awere performed with a Bruker-Reflex III (Bruker-Franzen Analytik,Bremen, Germany) in linear (LIN-) and/or reflector (REF-) TOF config-uration at an acceleration voltage of 20 kV and delayed ion extraction.Samples were dispersed in H2O (LPS), or chloroform/methanol, 1:1 (v/v)(lipid A), at a concentration of 10 �g �l�1 and mixed on the target withequal volumes of matrix solution. The best results were obtained usinga saturated solution of recrystallized 2,5-dihydroxybenzoic acid (gentis-ic acid, DHB, Aldrich) in 0.1% aqueous trifluoroacetic acid/acetonitrile,1:1, as matrix solution. The mass spectra show the average of at least 50single laser shots. Mass scale calibration was performed externally withsimilar compounds of known chemical structure.
Electrospray Ionization Fourier Transform-Ion Cyclotron ResonanceMass Spectrometry (ESI FT-ICR MS)—ESI FT-ICR MS was performedin the negative and positive ion mode using an APEX II Instrument(Bruker Daltonics, Billerica, MA) equipped with a 7-tesla magnet andan Apollo ion source. Samples were dissolved at a concentration ofabout 10 ng �l�1 in a 50:50:0.001 (v/v/v) mixture of 2-propanol, water,and triethylamine and sprayed at a flow rate of 2 �l min�1. Capillaryvoltage was set to 3.8 kV, and drying gas temperature to 150 °C.Capillary skimmer dissociation was induced by increasing the capillaryexit voltage from �100 to �350 V. Infrared multiphoton dissociation ofisolated parent ions was performed with a 35 W, 10.6-�m CO2 laser(Synrad, Mukilteo, WA). The unfocused laser beam was directedthrough the center of the ICR cell for 40 ms and the fragment ions weredetected after a delay of 0.5 ms.
Nuclear Magnetic Resonance (NMR) Spectroscopy—For NMR spec-troscopy all exchangeable lipid A protons were replaced with deuteriumby dissolving 5.0 mg of lipid A in 1.0 ml of chloroform-d/methanol-d4, 1:1(v/v). After evaporating it in a slight N2 stream, lipid As were dissolvedin 0.5 ml of chloroform-d/methanol-d4 7:3 (v/v) and transferred into5-mm high precision NMR sample tubes (Promochem, Wesel,Germany). Tetramethylsilane was used as internal standard for 1H(0.00 ppm) and 13C (0.00 ppm) spectra.
Proton and all two-dimensional spectra were recorded at 600.1 MHzon a 14-tesla AVANCE DRX-600 (Bruker, Rheinstetten, Germany) and13C spectra were measured on a 8.4-tesla AVANCE DPX-360 (Bruker)at 90.6 MHz. All spectra were measured at 310 K and performed withBruker XWINNMR 2.6 software. One-dimensional measurements wererecorded with a 90° pulse angle, an acquisition of 16,384 data points,and a relaxation delay of 1.0 s. After zero filling to 32,768 data pointsthey were Fourier transformed to spectra with a range of 7200 Hz (1H)or 22,000 Hz (13C). To determine DQF-COSY, TOCSY (100-ms mixingtime), ROESY (250-ms mixing time), HMQC (coupled and decoupled),HMQC-TOCSY, and HMBC, between 256 and 512 experiments in theF1-dimension were recorded, each with 2048 data points in the F2-dimension. Sinusoidal multiplication and Fourier transformation led totwo-dimensional spectra with a range of 4800 Hz in the proton dimen-sion as well as 22,000 Hz and 32,000 Hz in the carbon dimension forHMQC and HMBC, respectively.
Immunological Characterization of LPS—Purified LPS was sepa-rated by SDS-PAGE gels and either stained with alkaline silver nitrate(24) or detected with monoclonal antibodies (mAb) after electrotransferonto polyvinylidene difluoride membranes by tank-blotting (Bio-RadMini Trans-Blot cell). Prior to use, polyvinylidene difluoride mem-branes were wetted in methanol and rinsed carefully in distilled water(at least 10 min), where they were kept until further use. Blotting wascarried out at 4 °C for 16 h at 10 mA, all following steps were performedat room temperature. After transfer, membranes were placed in dis-
Structure of B. bacteriovorus Lipid A 27503
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

tilled water for 30 min, washed six times for 5 min each in blot buffer(50 mM Tris-HCl, 0.2 M NaCl, pH 7.4), blocked 1 h in blot buffersupplemented with 10% nonfat dry milk, and incubated for 1 h withmAb A6, directed against a bisphosphorylated lipid A backbone (25),A20, recognizing a terminal Kdo residue (26), or mAb WN1, reactingwith the core region of E. coli LPS (27). Antibody A6 was used as the cellculture supernatant (RPMI, supplemented with 10% fetal calf serum),A20 (2 �g ml�1) and WN1 (5 �g ml�1) were diluted in the same medium.Blots were washed six times (5 min each in blot buffer) to remove theprimary antibody, followed by incubation for 1 h with alkaline phos-phatase-conjugated goat anti-mouse immunoglobulin G (H�L, Di-anova, diluted 1:2000 in blot buffer supplemented with 10% nonfat drymilk), washed as before, and developed with 5-bromo-4-chloro-3-indo-lyl-phosphate and p-nitro blue tetrazolium chloride as substrates ac-cording to the supplier’s instructions.
Release of Cytokines from Human Mononuclear Cells—Human mono-nuclear cells (hMNC) were isolated from healthy donors. Heparinizedblood (20 IU ml�1) was processed directly by mixing with an equalvolume Hanks’ balanced salt solution and centrifugation on Ficoll den-sity gradient for 40 min (21 °C, 500 � g). The interphase layer ofmononuclear cells was collected and washed twice in serum free Hanks’solution and once in serum-free RPMI 1640 containing 2 mM L-gluta-mine, 100 units ml�1 penicillin, and 100 �g ml�1 streptomycin. Cellswere resuspended in serum-free medium, and the cell number wasadjusted to 5 � 106 ml�1.
For the stimulation experiment 200 �l of hMNC were transferred toeach well of 96-well culture plates and LPS and lipid A were added togive final concentrations of 100 ng ml�1, 10 ng ml�1, and 1 ng ml�1. Thesamples were incubated for 4 h at 37 °C and 5% CO2. Supernatantswere collected after centrifugation of the culture plates for 10 min at400 � g and stored at �20 °C until further use.
The release of TNF-� in the cell supernatants was performed in asandwich enzyme-linked immunosorbent assay as described elsewhere(28). Briefly, microtiter plates (Greiner, Solingen, Germany) werecoated with a mAb against TNF-� (Intex AG, Switzerland). Cell culturesupernatants and standard (recombinant TNF-�, rTNF-�, Intex) wereadded as appropriately diluted test samples and serial dilutions ofrTNF-�. The horseradish peroxidase-conjugated rabbit anti-rTNF-� an-tibody was added and the plates were incubated while shaking for 16 to24 h at 4 °C. After six washings with distilled water the color reactionwas initiated by addition of tetramethylbenzidine/H2O2 in alcoholicsolution and stopped with the same amount of 1 M H2SO4. The resultingyellow oxidation product was determined at a wavelength of 450 nm onan enzyme-linked immunosorbent assay reader (Rainbow, Tecan,Crailsheim, Germany). Interleukin-6 enzyme-linked immunosorbentassay was performed according to the manufacturer’s recommendation(Intex AG, Switzerland).
Determination of (� 7 �) Gel to Liquid-Crystalline Phase Transi-tion—The phase behavior was characterized by Fourier transform in-frared (FT-IR) spectroscopy on a IFS55 (Bruker, Karlsruhe, Germany)using 10 mM lipid A and LPS suspensions isolated from HI100 and fromE. coli K12 according to methods as described in the literature (29). Thepeak position of the symmetric stretching vibration of the methylenegroups was taken as a measure of the acyl chain order, lying below 2850cm�1 in the highly ordered gel phase (low fluidity) and above 2852 cm�1
in the less ordered liquid-crystalline phase (high fluidity).
RESULTS
LPS Isolation and SDS-PAGE—Extraction of dried bacterialcells using a modified phenol/chloroform/light petroleummethod gave LPS preparations, which were analyzed by SDS-PAGE (Fig. 1). The sample H2 and HI100 LPS showed char-acteristic patterns for S-form LPS (lanes 1, 4, and 5). Thesample H1 and E. coli K12 LPS preparations showed profilestypical for R-form LPS (lanes 2, 3, and 6–8). As can be deducedfrom the obvious similarity between H1 of HD100 (lanes 2 and3) and the LPS of E. coli K12 (lanes 6 and 7) the water-precipitated LPS fraction H1 contains mainly the R-form LPSof E. coli K12. In contrast, the ethanol-precipitated fraction H2of HD100 (lanes 4 and 5) as well as the LPS of HI100 (lane 1)showed ladders of repeating units typical for S-form LPS. How-ever, the distances of the bands are larger for the host-inde-pendent strain HI100 indicating a size difference of the repeat-ing units of both strains. Small amounts of the LPS of B.bacteriovorus HD100 were also observed in H1 (lanes 2 and 3).
Immunological Characterization of LPS of B. bacteriovorus—For further characterization of the different LPS preparationsthree monoclonal antibodies (A20, A6, and WN1) were used forWestern blots. The mAb A20 recognizes a terminal Kdo resi-due, mAb A6 reacts specifically with bisphosphorylated lipid Abackbone, and mAb WN1 identifies the core region of E. coliK12 (Fig. 2). With mAb A20 (panel B) with water and ethanol-precipitated fractions H1 and H2 from strain HD100 (lane 6and 1, respectively), LPS from strain HI100 (lane 3), and LPSfrom E. coli K12 LPS (lane 5) showed reactivity. This showsthat B. bacteriovorus possess Kdo residues, which are accessi-ble to mAb A20. However, it does not allow a differentiation ofB. bacteriovorus LPS and that of E. coli K12. To detect residualhost LPS in preparations of B. bacteriovorus strain HD100mAb WN1 was used (Fig. 2, panel A). Strong reactivity wasvisible with the control LPS of K12 (lane 5) as well as with H1of strain HD100 (lane 6). However, the fraction H2 of HD100contained considerably less of the host LPS (lane 1). In con-trast, purified LPS of the host-independent strain HI100 didnot show any reaction with mAb WN1 (lane 3), indicatingstructural differences to the core region of E. coli. A structuraldifference of the lipid A backbones could be demonstrated withmAb A6 (panel C). In addition to E. coli K12 LPS (lane 5) andboth B. bacteriovorus HD100 preparations (lanes 1 and 6), thepurified lipid A of B. bacteriovorus HD100 showed a positivereaction with this mAb (lane 2). Neither the LPS of strainHI100 nor the isolated lipid A was recognized by mAb A6 (lanes3 and 4). Taken together, these results indicate that B. bacte-riovorus LPS possesses Kdo, that its core region and the lipid Abackbone are different from that of E. coli, and that residualLPS from the host E. coli K12 is found in the HD100 LPSpreparation. The free lipid A of B. bacteriovorus HI100 did notreact with any available antibody (lane 4).
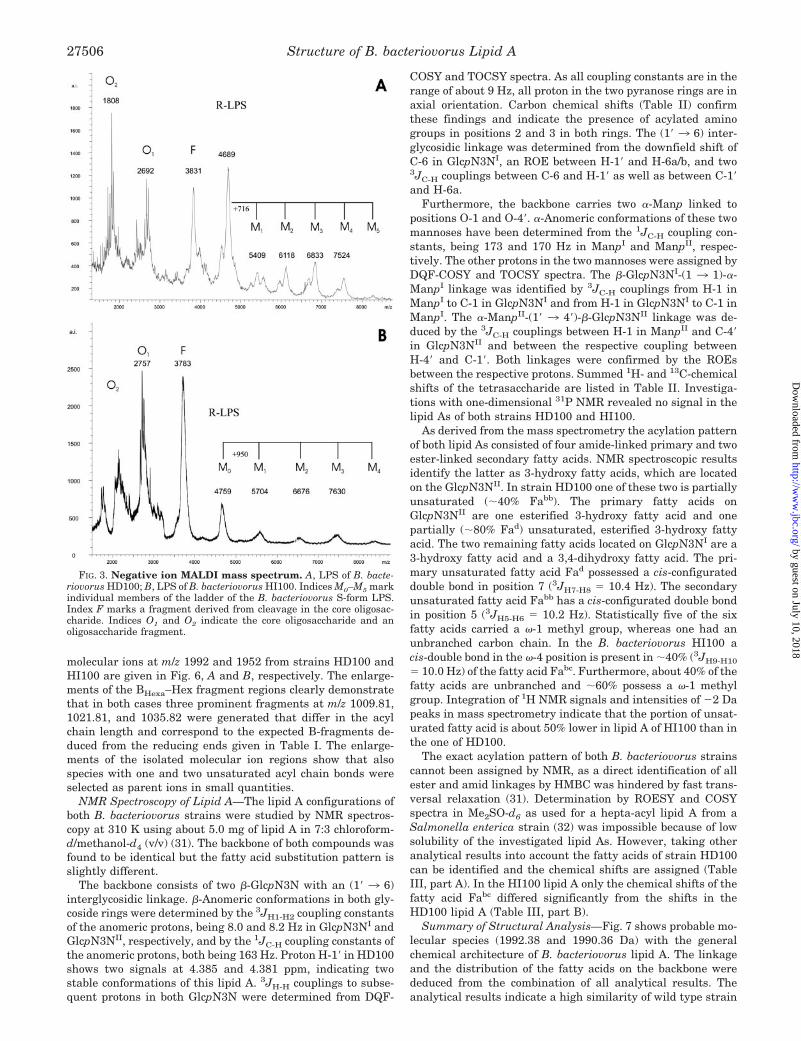
Mass Spectrometry of LPS of B. bacteriovorus—The negativeion MALDI LIN-TOF mass spectra of the complete LPS ofB. bacteriovorus HD100 and B. bacteriovorus HI100 (Fig. 3, Aand B) show typical patterns of S-form LPS with a series ofmolecular ion peaks, representing LPS species with differentnumbers of repeating units (M0, M1, M2, M3, . . . ) and laser-induced in-source fragment ions originating from the cleavageof the labile linkage between lipid A and core oligosaccharides.Fragments representing the core oligosaccharide (O1, O2) wereidentified by accompanying fragments (–44 Da) originatingfrom decarboxylation of KdoI. These results are in good agree-ment with the data from experiments using �-Kdo-mAb A20(Fig. 2B). Although the spectra are not well resolved because ofthe heterogeneity and adduct ion formation, they reveal impor-tant data concerning the differences of LPS HD100 and HI100.The R-form LPS (M0) can be assigned to the peaks at m/z 4759
FIG. 1. Alkaline silver-stained SDS-PAGE of the LPS B. bacte-riovorus HI100, B. bacteriovorus HD100, and E. coli K12. 1, 0.5 �gof LPS of HI100; 2, 0.5 �g of LPS for fraction H1; 3, 1 �g of LPS forfraction H1; 4, 0.5 �g of LPS of HD100 (H2); 5, 1 �g of LPS of HD100(H2); 6, 0.5 �g of LPS of E. coli K12; 7, 1 �g of LPS of E. coli K12; 8, 1�g from the ethanol-precipitated fraction of E. coli K12.
Structure of B. bacteriovorus Lipid A27504
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

(HI100) and m/z 4689 (HD100). The same mass differenceswere observed for the core oligosaccharides (O1). The repeatingunit of HI100 has an average mass of 950 Da whereas HD100has an average mass of 716 Da. Laser-induced cleavage in thecore oligosaccharide lead to fragments at m/z 3831 and m/z3783 for HD100 and HI100, respectively. Furthermore, thehost-dependent strain HD100 shows up to three peaks in theLPS population attributing to heterogeneity in the core oligo-saccharide. High resolution ESI FT-ICR MS of the R-form LPS(M0) gave a complex pattern of peaks differing by 14 Da com-bined with species at �2 and �4 Da indicating that the fattyacid composition must be very heterogeneous with respect tochain length and the degree of saturation (data not shown).
Chemical Analysis of B. bacteriovorus LPS (GC-MS)—Kdo,Glc, GlcN, and Hep were detected as part of the oligosaccharideportion of the complete LPS of B. bacteriovorus by GC-MS aftermethylation. The sugar portion of the lipid A was identifiedas 2,3-diamino-2,3-dideoxy-D-glucopyranose (GlcpN3N) and D-mannopyranose (Manp).
Fatty acid analysis revealed only hydroxylated fatty acids forboth strains. The most abundant fatty acid component ofB. bacteriovorus HD100 was (R)-3-hydroxy-11-methyl-dodeca-noic acid (iso-C13:0(3-OH)). In small amounts unbranched3-hydroxy fatty acids were detected. In the host-independentstrain HI100 the major components could be identified as thebranched and unbranched 13:0(3-OH) in nearly equal amounts.For both strains longer and shorter homologues were detectedand the absolute configurations of all 3-hydroxy fatty acidswere determined as R-form. Furthermore, in both strains ad-ditional dihydroxy fatty acids (14:0(3,4-OH) and 15:0(3,4-OH))as well as unsaturated 3-hydroxy acids were present.
Because of rearrangement, reactions and the formation ofbyproducts during derivatization of the unsaturated and dihy-droxy fatty acid molar ratios of the identified fatty acids couldnot accurately be determined. Thus a detailed study of thefragmentation of the lipid A was performed using mass spec-trometry and NMR spectroscopy.
Mass Spectrometry of B. bacteriovorus Lipid A—Free lipid Asamples were mass analyzed by MALDI TOF-MS. In agree-
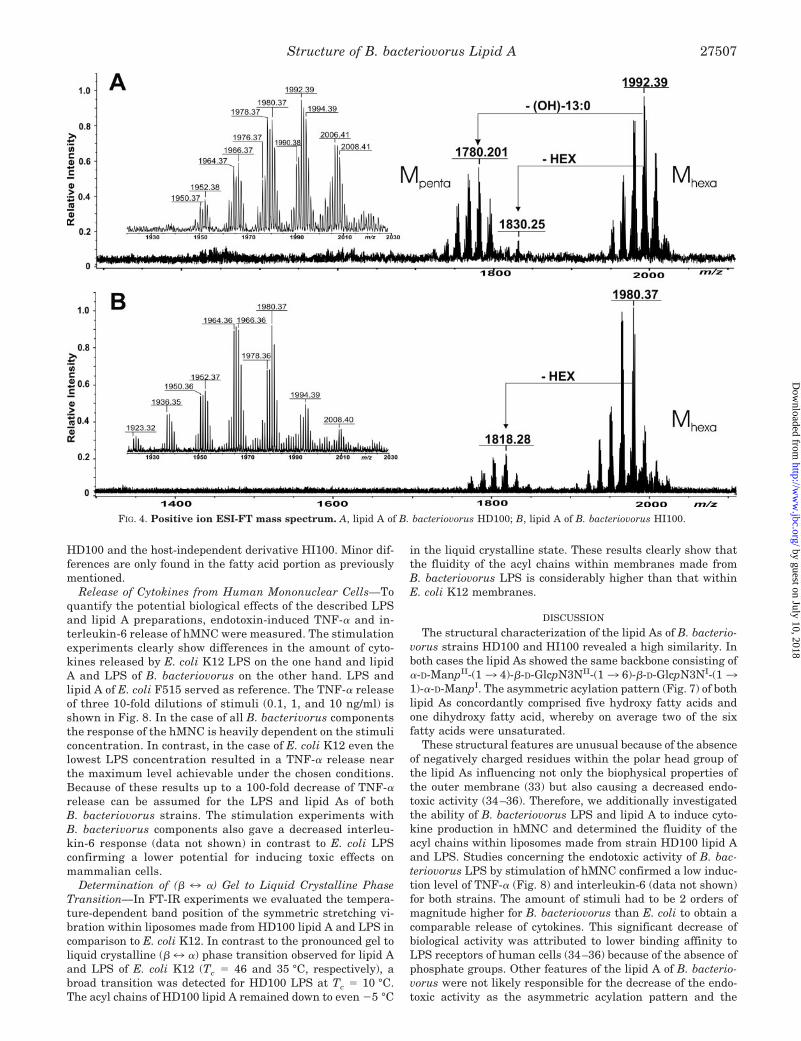
ment with the results of the laser-induced in-source fragmen-tation experiments of the complete LPS the same complexpatterns of lipid A ion peaks were detected in the positive ionmode (around m/z 2000; data not shown). Using high resolutionESI-MS the positive ion mass spectrum of HD100 lipid A (Fig.4A) exhibits two abundant groups [M � H]� ions around m/z1992.39 and m/z 1780.20 as the most intense signals. The firstgroup was assigned to lipid A consisting of a 2,3-diamino-2,3-dideoxyglucose (GlcpN3N) disaccharide carrying two hexosesand six fatty acids (MHexa). The second group of molecular ionsis missing one (OH)-13:0 fatty acid (–212 Da: MPenta). Thus thepeak at m/z 1992.39 was in excellent agreement with the cal-culated mass (m/z 1992,385) of a lipid A carrying one di(OH)-15:0, one (OH)-14:0, and four (OH)-13:0 fatty acids, one ofwhich possesses a double bond. A third group of peaks aroundm/z 1830.25 missing one hexose (–162 Da) can be seen withonly minor intensity. For HI100 lipid A (Fig. 4B) the group ofmolecular ions around m/z 1980.34 (MHexa) and m/z 1818.28corresponds to hexa-acylated lipid A carrying two or one hexoseresidues, respectively. The inlets in Fig. 4, A and B, representenlargements of the respective molecular ion region each show-ing more than 14 different molecular peaks expressing theheterogeneity in the fatty acid composition. These findingswere in good agreement with the data of the chemical compo-nent analysis. A comparison of the lipid As of both B. bacterio-vorus strains revealed that the average of the fatty acids of wildtype strain HD100 possess longer chains length (�14, �2 � 14Da) and a higher portion of double bonds (–2 Da) than HI100.
Lipid As from both strains were O-deacylated by hydrazinetreatment and then mass analyzed. For both lipid As thegroups of molecular species were reduced mainly by �424.6 Da(spectra not shown) demonstrating that two fatty acid (mainly(OH)-13:0) are ester-linked thus confirming that the backboneconsists only of a GlcpN3N-disaccharide.
Capillary skimmer dissociation generated fragment ions ofdiagnostic importance (Fig. 5, A and B). By cleavage the glyco-sidic linkage between the two GlcpN3Ns fragment ions (BHexa,BPenta) were formed representing the non-reducing GlcpN3NII
moiety (B-fragments according to the nomenclature of Domonand Costello (30)) followed by the subsequent loss of hexoseand, to a lesser extent, also by the cleavage of one (OH)-13:0fatty acid (–230 Da, peaks at m/z 800). From the masses of theB-fragments and known fatty acid composition of the GC-MS itis evident that the non-reducing GlcpN3NII of the hexa-acy-lated lipid A carries four residues, the reducing GlcpN3NI onlytwo fatty acid residues. Furthermore, a comparison of the B-fragments (BHexa-Hex) of the two lipid As missing the hexose(enlargements shown in Fig. 5, C and D) demonstrate that bothsamples comprise identical fragment ions, however, with dif-ferent intensities. An unambiguous determination of the fourfatty acid residues linked to GlcpN3NII is not possible. There-fore, the molecular masses of all possible GlcpN3NI, carryingtwo fatty acid residues besides hexose, were calculated fromthe mass differences between all measured molecular ions (Fig.4A) and the B-fragments observed in Fig. 5C. These differenceswere compared with the masses calculated of all combinationsof fatty acids detected by the component analysis (see Table I).Most possible combinations consist of one dihydroxy fatty acidand one hydroxy fatty acid. Based on this calculation andresults of O-deacylation by hydrazine treatment it can be con-cluded that four hydroxylated fatty acids are linked toGlcpN3NII. Furthermore, up to two double bounds are detect-able in the B-fragments (Fig. 5, C and D).
To prove which fatty acid combinations are realized, MS/MSexperiments were performed. As an example, the infrared mul-tiphoton dissociation-MS/MS spectra of the hexa-acyl lipid A
FIG. 2. Immunological characterization of LPS. Reactivity ofmonoclonal antibodies with LPS of B. bacteriovorus HD100 H2 (lane 1),HD100 H1 (lane 6), HI100 (lane 3), and E. coli K12 (lane 5) as well aswith isolated lipid A from B. bacteriovorus HD100 (lane 2) and HI100(lane 4). Samples (2.5 �g/lane) were separated by SDS-PAGE, blottedonto polyvinylidene difluoride membranes, and developed with mono-clonal antibodies: A, mAb WN-1 reacting with the core region of E. coliLPS; B, mAb A20 �-Kdo reacting with a terminal Kdo-residue; C, mAbA6 reacting with a bisphosphorylated glucosamine disaccharidebackbone.
Structure of B. bacteriovorus Lipid A 27505
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from
molecular ions at m/z 1992 and 1952 from strains HD100 andHI100 are given in Fig. 6, A and B, respectively. The enlarge-ments of the BHexa–Hex fragment regions clearly demonstratethat in both cases three prominent fragments at m/z 1009.81,1021.81, and 1035.82 were generated that differ in the acylchain length and correspond to the expected B-fragments de-duced from the reducing ends given in Table I. The enlarge-ments of the isolated molecular ion regions show that alsospecies with one and two unsaturated acyl chain bonds wereselected as parent ions in small quantities.
NMR Spectroscopy of Lipid A—The lipid A configurations ofboth B. bacteriovorus strains were studied by NMR spectros-copy at 310 K using about 5.0 mg of lipid A in 7:3 chloroform-d/methanol-d4 (v/v) (31). The backbone of both compounds wasfound to be identical but the fatty acid substitution pattern isslightly different.
The backbone consists of two �-GlcpN3N with an (1� 3 6)interglycosidic linkage. �-Anomeric conformations in both gly-coside rings were determined by the 3JH1-H2 coupling constantsof the anomeric protons, being 8.0 and 8.2 Hz in GlcpN3NI andGlcpN3NII, respectively, and by the 1JC-H coupling constants ofthe anomeric protons, both being 163 Hz. Proton H-1� in HD100shows two signals at 4.385 and 4.381 ppm, indicating twostable conformations of this lipid A. 3JH-H couplings to subse-quent protons in both GlcpN3N were determined from DQF-
COSY and TOCSY spectra. As all coupling constants are in therange of about 9 Hz, all proton in the two pyranose rings are inaxial orientation. Carbon chemical shifts (Table II) confirmthese findings and indicate the presence of acylated aminogroups in positions 2 and 3 in both rings. The (1� 3 6) inter-glycosidic linkage was determined from the downfield shift ofC-6 in GlcpN3NI, an ROE between H-1� and H-6a/b, and two3JC-H couplings between C-6 and H-1� as well as between C-1�and H-6a.
Furthermore, the backbone carries two �-Manp linked topositions O-1 and O-4�. �-Anomeric conformations of these twomannoses have been determined from the 1JC-H coupling con-stants, being 173 and 170 Hz in ManpI and ManpII, respec-tively. The other protons in the two mannoses were assigned byDQF-COSY and TOCSY spectra. The �-GlcpN3NI-(1 3 1)-�-ManpI linkage was identified by 3JC-H couplings from H-1 inManpI to C-1 in GlcpN3NI and from H-1 in GlcpN3NI to C-1 inManpI. The �-ManpII-(1� 3 4�)-�-GlcpN3NII linkage was de-duced by the 3JC-H couplings between H-1 in ManpII and C-4�in GlcpN3NII and between the respective coupling betweenH-4� and C-1�. Both linkages were confirmed by the ROEsbetween the respective protons. Summed 1H- and 13C-chemicalshifts of the tetrasaccharide are listed in Table II. Investiga-tions with one-dimensional 31P NMR revealed no signal in thelipid As of both strains HD100 and HI100.
As derived from the mass spectrometry the acylation patternof both lipid As consisted of four amide-linked primary and twoester-linked secondary fatty acids. NMR spectroscopic resultsidentify the latter as 3-hydroxy fatty acids, which are locatedon the GlcpN3NII. In strain HD100 one of these two is partiallyunsaturated (�40% Fabb). The primary fatty acids onGlcpN3NII are one esterified 3-hydroxy fatty acid and onepartially (�80% Fad) unsaturated, esterified 3-hydroxy fattyacid. The two remaining fatty acids located on GlcpN3NI are a3-hydroxy fatty acid and a 3,4-dihydroxy fatty acid. The pri-mary unsaturated fatty acid Fad possessed a cis-configurateddouble bond in position 7 (3JH7-H8 � 10.4 Hz). The secondaryunsaturated fatty acid Fabb has a cis-configurated double bondin position 5 (3JH5-H6 � 10.2 Hz). Statistically five of the sixfatty acids carried a �-1 methyl group, whereas one had anunbranched carbon chain. In the B. bacteriovorus HI100 acis-double bond in the �-4 position is present in �40% (3JH9-H10
� 10.0 Hz) of the fatty acid Fabc. Furthermore, about 40% of thefatty acids are unbranched and �60% possess a �-1 methylgroup. Integration of 1H NMR signals and intensities of �2 Dapeaks in mass spectrometry indicate that the portion of unsat-urated fatty acid is about 50% lower in lipid A of HI100 than inthe one of HD100.
The exact acylation pattern of both B. bacteriovorus strainscannot been assigned by NMR, as a direct identification of allester and amid linkages by HMBC was hindered by fast trans-versal relaxation (31). Determination by ROESY and COSYspectra in Me2SO-d6 as used for a hepta-acyl lipid A from aSalmonella enterica strain (32) was impossible because of lowsolubility of the investigated lipid As. However, taking otheranalytical results into account the fatty acids of strain HD100can be identified and the chemical shifts are assigned (TableIII, part A). In the HI100 lipid A only the chemical shifts of thefatty acid Fabc differed significantly from the shifts in theHD100 lipid A (Table III, part B).
Summary of Structural Analysis—Fig. 7 shows probable mo-lecular species (1992.38 and 1990.36 Da) with the generalchemical architecture of B. bacteriovorus lipid A. The linkageand the distribution of the fatty acids on the backbone werededuced from the combination of all analytical results. Theanalytical results indicate a high similarity of wild type strain
FIG. 3. Negative ion MALDI mass spectrum. A, LPS of B. bacte-riovorus HD100; B, LPS of B. bacteriovorus HI100. Indices M0–M5 markindividual members of the ladder of the B. bacteriovorus S-form LPS.Index F marks a fragment derived from cleavage in the core oligosac-charide. Indices O1 and O2 indicate the core oligosaccharide and anoligosaccharide fragment.
Structure of B. bacteriovorus Lipid A27506
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from
HD100 and the host-independent derivative HI100. Minor dif-ferences are only found in the fatty acid portion as previouslymentioned.
Release of Cytokines from Human Mononuclear Cells—Toquantify the potential biological effects of the described LPSand lipid A preparations, endotoxin-induced TNF-� and in-terleukin-6 release of hMNC were measured. The stimulationexperiments clearly show differences in the amount of cyto-kines released by E. coli K12 LPS on the one hand and lipidA and LPS of B. bacteriovorus on the other hand. LPS andlipid A of E. coli F515 served as reference. The TNF-� releaseof three 10-fold dilutions of stimuli (0.1, 1, and 10 ng/ml) isshown in Fig. 8. In the case of all B. bacterivorus componentsthe response of the hMNC is heavily dependent on the stimuliconcentration. In contrast, in the case of E. coli K12 even thelowest LPS concentration resulted in a TNF-� release nearthe maximum level achievable under the chosen conditions.Because of these results up to a 100-fold decrease of TNF-�release can be assumed for the LPS and lipid As of bothB. bacteriovorus strains. The stimulation experiments withB. bacterivorus components also gave a decreased interleu-kin-6 response (data not shown) in contrast to E. coli LPSconfirming a lower potential for inducing toxic effects onmammalian cells.
Determination of (� 7 �) Gel to Liquid Crystalline PhaseTransition—In FT-IR experiments we evaluated the tempera-ture-dependent band position of the symmetric stretching vi-bration within liposomes made from HD100 lipid A and LPS incomparison to E. coli K12. In contrast to the pronounced gel toliquid crystalline (� 7 �) phase transition observed for lipid Aand LPS of E. coli K12 (Tc � 46 and 35 °C, respectively), abroad transition was detected for HD100 LPS at Tc � 10 °C.The acyl chains of HD100 lipid A remained down to even �5 °C
in the liquid crystalline state. These results clearly show thatthe fluidity of the acyl chains within membranes made fromB. bacteriovorus LPS is considerably higher than that withinE. coli K12 membranes.
DISCUSSION
The structural characterization of the lipid As of B. bacterio-vorus strains HD100 and HI100 revealed a high similarity. Inboth cases the lipid As showed the same backbone consisting of�-D-ManpII-(1 3 4)-�-D-GlcpN3NII-(1 3 6)-�-D-GlcpN3NI-(1 31)-�-D-ManpI. The asymmetric acylation pattern (Fig. 7) of bothlipid As concordantly comprised five hydroxy fatty acids andone dihydroxy fatty acid, whereby on average two of the sixfatty acids were unsaturated.
These structural features are unusual because of the absenceof negatively charged residues within the polar head group ofthe lipid As influencing not only the biophysical properties ofthe outer membrane (33) but also causing a decreased endo-toxic activity (34–36). Therefore, we additionally investigatedthe ability of B. bacteriovorus LPS and lipid A to induce cyto-kine production in hMNC and determined the fluidity of theacyl chains within liposomes made from strain HD100 lipid Aand LPS. Studies concerning the endotoxic activity of B. bac-teriovorus LPS by stimulation of hMNC confirmed a low induc-tion level of TNF-� (Fig. 8) and interleukin-6 (data not shown)for both strains. The amount of stimuli had to be 2 orders ofmagnitude higher for B. bacteriovorus than E. coli to obtain acomparable release of cytokines. This significant decrease ofbiological activity was attributed to lower binding affinity toLPS receptors of human cells (34–36) because of the absence ofphosphate groups. Other features of the lipid A of B. bacterio-vorus were not likely responsible for the decrease of the endo-toxic activity as the asymmetric acylation pattern and the
FIG. 4. Positive ion ESI-FT mass spectrum. A, lipid A of B. bacteriovorus HD100; B, lipid A of B. bacteriovorus HI100.
Structure of B. bacteriovorus Lipid A 27507
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

chain length of the fatty acids were comparable with the potentcytokine inducer E. coli hexa-acyl lipid A. As expected, theinduction level of TNF-� for the complete S-form LPS ofB. bacteriovorus HD100 and HI100 was higher compared withpure lipid A preparations (37). With respect to the unchargedbackbone of strain HD100 lipid A the fluidity of acyl chainswithin liposomes was affected. The results revealed that theacyl chains within HD100 LPS are in a significantly less or-dered state most probably because of the missing negativecharges inhibiting cation bridging within the outer membrane
and because of the presence of at least two unsaturated acylchains per lipid A molecule. This increased fluidity may lead toan increased permeability of HD100 outer membrane (38)meaning an advantage for consumption of degraded prey com-ponents during the intraperiplasmatic growth phase at a tem-perature range typical for soil and aquatic habitats.
It should be emphasized that our experiments cannot answerthe question if B. bacteriovorus integrates complete lipid A orLPS molecules of the prey bacteria into its cell wall (39). Westructurally determined the LPS of HD100 after completing its
FIG. 5. Positive ion ESI-capillary skimmer dissociation FT mass spectrum. A, lipid A of B. bacteriovorus HD100; B, lipid A of B.bacteriovorus HI100. C and D, enlargement of the BHexa-Hex fragment region of strains HD100 and HI100. Bhexa, B-fragments originating fromlipid A species MHexa; Bpenta, B-fragments originating from lipid A species MPenta.
TABLE ICalculation of fatty acid composition of the reducing end of B. bacteriovorus HD100 lipid A based upon
chemical analysis and mass spectrometry
Structure of B. bacteriovorus Lipid A27508
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

predatory life cycle on E. coli K12. The special structure ofB. bacteriovorus lipid A probably excluded incorporation pro-cesses of components of the outer membranes of the prey dur-ing the life cycle. This was confirmed by chemical separation ofB. bacteriovorus LPS from the E. coli K12 LPS of the prey withour isolation protocol. This finding supports electron micro-scopic observations (11) showing no membrane fusion processduring the invasion of B. bacteriovorus with the outer mem-brane of the prey. It was proposed that B. bacteriovorus enzy-matically hydrolyzes LPS of prey bacteria. However, the pres-ence of high amounts of unmodified LPS of preys after completedegradation of the prey bacteria by B. bacteriovorus suggeststhat this does not occur to a great extent and may be restrictedto the invasion process (12, 15). Furthermore, we microscopi-cally observed complete cell hulls of former prey bacteria sug-gesting that the outer membrane of the prey bacteria ratherprovides a protected environment (10) in which B. bacterio-vorus hydrolyzes and consumes the macromolecular compo-nents of the prey. In contrast to former investigations (18) twoLPS fractions clearly distinguishable from each other were
found in HD100 grown on E. coli K12. On the one hand a LPSfraction was synthesized by the prey bacteria and on the otherhand a B. bacteriovorus synthesized LPS. However, detailedstructural analysis of the B. bacteriovorus LPS revealed a newtype of substitution pattern of the lipid A backbone and anuncommon acylation pattern. Thus, the earlier described com-ponent analysis of B. bacteriovorus lipid A by Nelson et al. (18)could not be confirmed. We assume that an insufficient isola-tion procedure led to a contamination of the preparations withphospholipids, as we also noticed traces of phosphatidylethano-lamine in the lipid A preparation of B. bacteriovorus HI100.
Dissimilarities between the two B. bacteriovorus lipid Aswere different portions of iso-fatty acids and unsaturated fattyacids. In strain HD100 statistically five of the six fatty acidswere branched, whereas strain HI100 only possessed about 3.6branched fatty acids. Furthermore, a double bond in the sec-ondary fatty acid Fab (Table III, Fig. 7), which was present inabout 40% of both lipid As was at a different position. Whereasthe double bond was located between C-9 and C-10 in strainHI100, its position was between C-5 and C-6 in strain HD100.
FIG. 6. Positive ion infrared multiphoton dissociation MS/MS mass spectrum. A, lipid A molecule species m/z 1992.42 of B. bacteriovorusHD100; B, lipid A molecule species m/z 1952.42 of B. bacteriovorus HI100.
TABLE IIProton and carbon chemical shifts of the lipid A backbones in B. bacteriovorus strains HD100 and HI100
H-1 H-2 H-3 H-4 H-5 H-6a/b C-1 C-2 C-3 C-4 C-5 C-6
�-D-Manp�-(13 4.86 3.61 3.54 3.51 3.52 3.61/3.80 101.1 70.3 70.4 66.7 73.7 61.34)-�-D-GlcpN3N�-(13 4.385/4.381 3.73 3.97 3.65 3.32 3.75/3.75 101.6 52.9 53.7 74.2 75.9 60.16)-�-D-GlcpN3B�-(13 4.58 3.71 3.86 3.32 3.45 3.73/3.93 100.5 53.1 54.8 67.8 76.3 68.11)-�-D-Manp� 4.92 3.74 3.63 3.51 3.76 3.61/3.80 100.6 69.6 70.3 66.7 73.4 61.3
Structure of B. bacteriovorus Lipid A 27509
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

In the latter case the double bond caused a different lipid Aconformation compared with the accompanied saturated spe-cies, indicated by two 1H NMR signals of H-1� in a 6:4 ratio.Obviously, the double bond close to the backbone caused adifferent conformation from the saturated species and shiftedthe 1H NMR signals of H-1� from 4.386 to 4.381 ppm. Theprerequisite for such conformational differences was a flexibleinterglycosidic linkage in the backbone (31), which was con-firmed by NOEs from H-1� to H-6a/b and H-5. In strain HI100that possessed the double bond in a different position, thesedissimilar conformations could not be observed. Furthermore,
the amount of unsaturated fatty acids was about 50% lower instrain HI100 than in strain HD100. With regard to these minordifferences it can be assumed that wild type strain HD100synthesizes an innate lipid A without generating hybrid formsby recycling components of the preys lipid A. The high portionof iso-fatty acids in strain HD100 further indicates that theyare not consumed from the preys LPS because only unbranchedfatty acids are known for E. coli K12 (40).
How can the differences between the structure of the LPS ofhost-dependent strain HD100 and the host-independent strainHI100 be interpreted? To isolate host-independent strains of
FIG. 7. Structure of lipid A from B. bacteriovorus. A, structure of the backbone with two (1�36)-linked �-D-GlcpN3N in the central regionand two �-D-Manp linked in positions 1 and 4�. B, structure and distribution of the fatty acids for strain HD100. A 3-hydroxy-11-methyldedocanoicacid and a 3-hydroxy-11-methyldedodec-7-enoic acid (80% unsaturated) are amide linked to position 2� and 3� in �-D-GlcpN3NII. The two hydroxyfunctions of these primary acids were esterified with a 3-hydroxy-11-methyldodecanoic acid and a 3-hydroxy-11-methyldodec-5-enoic acid (40%unsaturated). Positions 2 and 3 in �-D-GlcpN3NI were substituted with a 3,4-dihydroxy-13-methyltetradecanoic acid and a 3-hydroxy-tetradecanoicacid. Chain length and branching of the fatty acids representing their predominant distribution were derived from the GC-MS analysis, infraredmultiphoton dissociation-MS/MS, and NMR. The exact position of each fatty acid could not be determined from the analytical results.
Structure of B. bacteriovorus Lipid A27510
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

B. bacteriovorus the selection of streptomycin-resistant mu-tants is necessary. Additionally, several more passages arenecessary before the mutants are able to grow on enrichedmedia (13). During this procedure the mutants probably ac-quire several mutations that enable them to grow without ahost. Host-independent mutants show morphologically distinc-tive features like spheroplast forming, long spiral-shaped cells,and pigmentation (14). Although a genetic locus (hit) was iden-tified that is involved in the conversion from the predatory tothe host-independent lifestyle, all authors are in agreementthat this phenotype cannot be explained by a single mutation inthe hit locus (13, 14, 42). Our hypothesis is that the differencesin the oligosaccharide portion of the HI100 LPS and HD100LPS are caused by the selection procedure (13). It is likely thatstreptomycin resistance of HI strains is caused by alterations ofthe ribosomal protein S12 that interferes with protein synthe-sis leading to pleiotropic changes (43, 44). As a consequence theproteins for the biosynthesis of the outer membrane might beaffected causing morphological aberrations in host-independ-
ent mutants. However, the main structural features of theHI100 lipid A are conserved in comparison to wild type strainHD100.
The unique predatory lifestyle of B. bacteriovorus requiresthe evolutionary development of special structures. The lipid Athat was found in B. bacteriovorus and is described here for thefirst time may represent such a unique structure. Analyses ofother Gram-negative bacteria living in highly specialized envi-ronments have indicated that an unusual lipid A structure isimportant for the lifestyle and may serve as an evolutionarymarker. It was shown that in Aquifex pyrophilus, living in hotsprings, the conserved phosphate substituents of the lipid A arereplaced by galacturonic acid residues (45). Chlamydia tracho-matis, an obligate intracellular pathogen, comprises a highlyheterogeneous LPS in regard to fatty acid composition (41).This may be a parallel to the intracellular growth of B. bacte-riovorus inside the prey bacteria. The correlation between anunusual lipid A structure and a specialized environment cannow be extended to B. bacteriovous.
TABLE IIIProton and carbon chemical shifts of the lipid A fatty acids in B. bacteriovorus (A) and Fabc in B. bacteriovorus strain HI100 (B)
H-1C-1
H-2a/bC-2
H-3C-3
H-4C-4
H-5C-5
H-6C-6
H-7C-7
H-8C-8
H-9C-9
H-10C-10
H-11C-11
H-12C-12
H-13C-13
H-14C-14
H-15C-15
A (HD100)Fatty acid a (Faa) 2.34/2.34 3.92 1.42 1.19 NDa ND ND ND ND 1.43 0.79 0.79
171.8 41.5 68.1 36.7 28.9 ND ND ND ND ND 38.5 13.0 13.0Fatty acid ba (Faba) 2.34/2.34 3.92 1.42 1.19 ND ND ND ND ND 1.43 0.79 0.79
171.8 41.5 68.1 36.7 28.9 ND ND ND ND ND 38.5 13.0 13.0Fatty acid bb (Fab) 2.34/2.34 3.99 2.19 5.33 5.45 1.96 1.28 ND ND 1.43 0.79 0.79
171.8 41.5 68.1 34.2 123.5 132.5 24.9 28.3 ND ND 38.5 13.0 13.0Fatty acid c (Fac) 2.44/2.46 5.15 1.51 1.22 1.10 ND ND ND ND 1.43 0.79 0.79
171.5 40.5 70.7 33.5 28.7 26.5 ND ND ND ND 38.5 13.0 13.0Fatty acid d (Fad) 2.37/2.43 5.08 1.52 1.20 1.94 5.30 5.23 1.97 1.27 1.43 0.79 0.79
171.1 40.2 71.0 33.4 28.8 26.5 130.0 128.3 26.5 29.1 38.5 13.0 13.0Fatty acid e (Fae) 2.17/2.29 3.85 1.38 1.20 ND ND ND ND ND ND ND 1.18 0.83
173.9 43.0 68.2 36.4 28.7 ND ND ND ND ND ND ND 38.5 13.3Fatty acid f (Faf) 2.26/2.26 3.75 3.28 1.39 1.22 ND ND ND ND ND 1.18 1.43 0.79 0.79
173.3 39.9 70.3 73.4 32.7 29.4 ND ND ND ND ND 27.3 38.5 13.0 13.0B (HI100)
Fatty acid bc (Fabc) 2.34/2.34 3.95 1.40 1.21 ND ND 1.96 5.32 5.32 1.43 0.81 0.81171.8 41.3 68.4 36.2 29.1 ND ND 25.1 128.6 128.6 38.5 13.1 13.1
a ND, not determined due to signal overlap.
FIG. 8. Release of TNF-� from human mononuclear cells by stimulation with lipid A and LPS of B. bacteriovorus HD100, HI100, andE. coli K12.
Structure of B. bacteriovorus Lipid A 27511
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

Acknowledgments—We gratefully acknowledge H. P. Cordes (Re-search Center Borstel) for taking several NMR measurements. Wethank L. Brade (Research Center Borstel, Borstel, Germany) for pro-viding the monoclonal antibodies used in this study and I. von Cube(Research Center Borstel, Borstel, Germany) for expert technical as-sistance. Furthermore, we thank G. von Busse (Research Center Bors-tel, Borstel, Germany) for performing FT-IR spectroscopy.
REFERENCES
1. Stolp, H., and Starr, M. P. (1963) Antonie Leeuvewenhoek 29, 217–2482. Ravenschlag, K., Sahm, K., Pernthaler, J., and Amann, R. (1999) Appl. Envi-
ron. Microbiol. 65, 3982–39893. Jurkevitch, E., Minz, D., Ramati, B., and Barel, G. (2000) Appl. Environ.
Microbiol. 66, 2365–23714. Richardson, I. R. (1990) J. Appl. Bacteriol. 69, 134–1405. Lambina, W. A., Ledowa, L. A., and Tschurkina, L. G. (1987) Mikrobiologiia
56, 860–8646. Ibragimov, F. (1980) Zh. Mikrobiol. Epidemiol. Immunobiol. 97–997. Schwudke, D., Strauch, E., Krueger, M., and Appel, B. (2001) Syst. Appl.
Microbiol. 24, 385–3948. Baer, M. L., Ravel, J., Chun, J., Hill, R. T., and Williams, H. N. (2000) Int. J.
Syst. Evol. Microbiol. 50, 219–2249. Cover, W. H., and Rittenberg, S. C. (1984) J. Bacteriol. 157, 391–397
10. Cover, W. H., Martinez, R. J., and Rittenberg, S. C. (1984) J. Bacteriol. 157,385–390
11. Abram, D., Castro e Melo, J., and Chou, D. (1974) J. Bacteriol. 118, 663–68012. Tudor, J. J., McCann, M. P., and Acrich, I. A. (1990) J. Bacteriol. 172,
2421–242613. Seidler, R. J., and Starr, M. P. (1969) J. Bacteriol. 100, 769–78514. Barel, G., and Jurkevitch, E. (2001) Arch. Microbiol. 176, 211–21615. Thomashow, M. F., and Rittenberg, S. C. (1978) J. Bacteriol. 135, 998–100716. Rosson, R. A., and Rittenberg, S. C. (1979) J. Bacteriol. 140, 620–63317. Talley, B. G., McDade, R. L., Jr., and Diedrich, D. L. (1987) J. Bacteriol. 169,
694–69818. Nelson, D. R., and Rittenberg, S. C. (1981) J. Bacteriol. 147, 860–86819. Kuenen, J. G., and Rittenberg, S. C. (1975) J. Bacteriol. 121, 1145–115720. Galanos, C., Luderitz, O., and Westphal, O. (1969) Eur. J. Biochem. 9, 245–24921. Wollenweber, H. W., and Rietschel, E. T. (1990) J. Microbiol. Methods 11,
195–21122. Bryn, K., and Jantzen, E. (1982) J. Chromatogr. 240, 405–41323. Gerwig, G. J., Kammerling, J. P., and Vliegenthart, J. F. (1979) Carbohydr.
Res. 77, 10–17
24. Tsai, C. M., and Frasch, C. E. (1982) Anal. Biochem. 119, 115–11925. Kuhn, H. M., Brade, L., Appelmelk, B. J., Kusumoto, S., Rietschel, E. T., and
Brade, H. (1992) Infect. Immun. 60, 2201–221026. Brade, L., Kosma, P., Appelmelk, B. J., Paulsen, H., and Brade, H. (1987)
Infect. Immun. 55, 462–46627. Di Padova, F. E., Brade, H., Barclay, G. R., Poxton, I. R., Liehl, E., Schuetze,
E., Kocher, H. P., Ramsay, G., Schreier, M. H., McClelland, D. B. L., andRietschel, E. T. (1993) Infect. Immun. 61, 3863–3872
28. Gallati, H. (1982) J. Clin. Chem. Clin. Biochem. 20, 907–91429. Brandenburg, K., Kusumoto, S., and Seydel, U. (1997) Biochim. Biophys. Acta
1329, 183–20130. Domon, B., and Costello, C. E. (1988) Glycoconj. J. 5, 397–40931. Brecker, L. (2003) Chem. Phys. Lipids, in press32. Janusch, H., Brecker, L., Lindner, B., Heine, H., Alexander, C., Gronow, S.,
Ulmer, A. J., Rietschel, E. T., and Zahringer, U. (2002) J. Endotoxin Res. 8,343–356
33. Schromm, A. B., Brandenburg, K., Loppnow, H., Zahringer, U., Rietschel,E. T., Carroll, S. F., Koch, M. H., Kusumoto, S., and Seydel, U. (1998)J. Immunol. 161, 5464–5471
34. Alexander, C., and Rietschel, E. T. (2001) J. Endotoxin Res. 7, 167–20235. Rietschel, E. T., Kirikae, T., Schade, F. U., Ulmer, A. J., Holst, O., Brade, H.,
Schmidt, G., Mamat, U., Grimmecke, H-D., Kusumoto, S., and Zahringer,U. (1993) Immunobiology 187, 169–190
36. Rietschel, E. T., Kirikae, T., Schade, F. U., Mamat, U., Schmidt, G., Loppnow,H., Ulmer, A. J., Zahringer, U., Seydel, U., Di Padova, F., Schreier, M., andBrade, H. (1994) FASEB J. 8, 217–225
37. Schromm, A. B., Brandenburg, K., Loppnow, H., Moran, A. P., Koch, M. H. J.,Rietschel, E. T., and Seydel, U. (2000) Eur. J. Biochem. 267, 2008–2013
38. Bengoechea, J. A., Brandenburg, K., Seydel, U., Diaz, R., and Moriyon, I.(1998) Microbiology 144, 1517–1526
39. Stein, M. A., McAllister, S. A., Torian, B. E., and Diedrich, D. L. (1992) J.Bacteriol. 174, 2858–2864
40. Raetz, C. H. (1996) in Escherichia coli and Salmonella (Neidhardt, C. R. H., ed)2nd Ed., Vol. 1, pp. 1035–1063, ASM Press, Washington, D. C.
41. Rund, S., Lindner, B., Brade, H., and Holst, O. (1999) J. Biol. Chem. 274,16819–16824
42. Cotter, T. W., and Thomashow, M. F. (1992) J. Bacteriol. 174, 6018–602443. Grafe, U. (ed) (1992) Biochemie der Antibiotika, pp. 268, Spektrum Akademis-
cher Verlag, Heidelberg44. Mingeot-Leclercq, M. P., Glupczynski, Y., and Tulkens, P. M. (1999) Antimi-
crob. Agents Chemother. 43, 727–73745. Ploetz, B. M., Lindner, B., Stetter, K. O., and Holst, O. (2000) J. Biol. Chem.
275, 11222–11228
Structure of B. bacteriovorus Lipid A27512
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from

Hermann Moll, Mareike Müller, Lothar Brecker, Sabine Gronow and Buko LindnerDominik Schwudke, Michael Linscheid, Eckhard Strauch, Bernd Appel, Ulrich Zähringer,
HOST-INDEPENDENT DERIVATIVE HI100OF THE WILD TYPE STRAIN B. BACTERIOVORUS HD100 AND ITS
DIFFERENCES BETWEEN THE LIPID As AND THE LIPOPOLYSACCHARIDES -D-Mannoses That Replace Phosphate Residues: SIMILARITIES ANDαContaining
Possesses a Neutral Lipid ABdellovibrio bacteriovorusThe Obligate Predatory
doi: 10.1074/jbc.M303012200 originally published online May 12, 20032003, 278:27502-27512.J. Biol. Chem.
10.1074/jbc.M303012200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/278/30/27502.full.html#ref-list-1
This article cites 41 references, 22 of which can be accessed free at
by guest on July 10, 2018http://w
ww
.jbc.org/D
ownloaded from