the influence of motor sequence learning on stroop effectessay.utwente.nl/64848/1/lam, w.l. -...
TRANSCRIPT

The influence of motor sequence learning on Stroop effect
Wai Ling Lam (s1100572)
Faculty of Behavioral Sciences, University of Twente, the Netherlands
In fulfillment of the Bachelor‘s thesis requirement
Psychology
1st Supervisor: Dr. Rob van der Lubbe
2nd Supervisor: Dr. Marit Ruitenberg
January 23, 2014

2
Contents
Abstract............................................................................................................................................... 3
Samenvatting....................................................................................................................................... 4
Introduction......................................................................................................................................... 5
Method................................................................................................................................................ 10
Subjects..................................................................................................................................... 10
Apparatus.................................................................................................................................. 10
Task and Procedure................................................................................................................... 10
Results................................................................................................................................................. 13
Training phase........................................................................................................................... 13
Test phase.................................................................................................................................. 16
Discussion............................................................................................................................................. 21
References............................................................................................................................................ 24

3
Abstract
The aim of this study was to find out whether motor sequence learning could contribute to less
perceptual attention, which in turn caused reduced Stroop conflict. Thirty two students of the
University of Twente completed a computer task that consisted of the Discrete Sequence Production
(DSP) Task and the Stroop Task. Subjects had to react to the ink color of a color word, while
disregarding the meaning of the word. In this study, subjects of one group practiced two fixed motor
sequences (each sequence consisted of six key presses) across six blocks while the subjects of the
other group did not. After the training phase, the sequence training group was presented with two
familiar and two unfamiliar sequences in the test phase. Results showed that subjects in the
sequence training group showed smaller Stroop effect when they performed motor sequences that
they previously had learned compared to when they were exposed to unfamiliar sequences. But this
was only the case when reaction times of the first key press were omitted. Furthermore, this group
did not overcome the Stroop effect completely when confronted with familiar sequences. The results
were in line with the Dual Processor Model of sequencing skill. The findings suggested that motor
sequence learning reduced Stroop effect. Future research could focus on whether repeated exposure
to the task (without fixed sequences being presented) would reduce Stroop conflict, which would
suggest increased cognitive control.

4
Samenvatting
Het doel van deze studie was om achter te komen of motorisch sequentieleren kon bijdragen aan
verminderde perceptuele aandacht, wat op zijn beurt verminderde Stroopconflict kon veroorzaken.
Tweeëndertig studenten van Universiteit Twente voerden een computertaak uit die bestond uit de
Discrete Sequentie Productie (DSP) taak en de Stroop taak. Participanten moesten reageren op de
inktkleur van het kleurwoord, terwijl ze de betekenis van het kleurwoord moesten negeren. In dit
experiment hadden participanten van de ene groep geoefend met vaste motorsequenties (elke
sequentie bestond uit zes key toetsen) over de zes trainingsblokken, terwijl participanten van de
andere groep niet hadden geoefend. Na de trainingsfase kreeg de groep met sequentietraining twee
bekende en twee onbekende sequenties te zien. De uitkomsten lieten zien dat participanten van de
groep met sequentieleren kleinere Stroopeffecten hadden wanneer ze bekende motorsequenties
moesten uitvoeren, vergeleken met wanneer ze blootbesteld werden aan onbekende sequenties.
Maar dit resultaat gold alleen wanneer de reactietijden van de eerste key toets waren weggelaten in
de analyse. Verder kon deze groep niet het Stroopeffect helemaal weerstaan wanneer ze werden
blootgesteld aan bekende sequenties. De resultaten waren in lijn met het Dual Processor Model of
sequencing skill. De bevindingen suggereerden dat motorisch sequentieleren het Stroopeffect
verminderde. Toekomstig onderzoek zou kunnen focussen op de vraag of herhaaldelijke blootstelling
aan deze taak (zonder vaste sequenties) zou kunnen zorgen voor verlaging van het stroopconflict, die
dan zou wijzen op verhoogde cognitieve controle.

5
Introduction
Human behavior consists of automatic and controlled actions and a significant proportion of
human behavior appears to be automatic. Automatic processes can generally be seen as rapid,
smooth, involuntary and cognitively effortless. These processes also need little or no conscious
attention or explicit monitoring which makes them suitable for behavioral guidance (Langer, 1978). A
controlled action on the other hand does require some degree of attention in order to reach goals.
The limitation of our cognitive resources forces us to concentrate on just one object, dimension,
situation, or task. The other aspects are ignored, inhibited or deferred for later processing. The
exertion of selective attention makes this possible (Deroost, Vandenbossche, Zeischka, Coomans &
Soetens, 2012).
According to Lavie, Hirst, de Fockert and Viding (2004), selective attention consists of two
mechanisms. There is a passive perceptual system which can be seen as a passive mechanism,
whereby possible interference caused by irrelevant distractors is avoided because the distractors are
not perceived. There is also an active mechanism of cognitive control that is used for rejecting
unimportant distractors even when these distractors are perceived. According to Deroost,
Vandenbossche, Zeischka, Coomans and Soetens (2012), it is important to monitor and regulate our
ongoing action, because by doing so we make it possible to flexibly adapt our behavior to our ever-
changing environment. The ability of monitoring and regulating is referred to as cognitive control.
A well-known task for investigating cognitive control is the Stroop task (Stroop, 1935). In this
task, subjects have to determine the ink color in which a word is printed and at the same time, they
are asked to ignore the meaning of the word. When the color and the meaning of the word are
congruent, it means that the written word of the color matches with the color itself (e.g., GREEN
written in green). Moreover, the color-meaning pair can be incongruent (e.g., BLUE written in green).
Macleod (1991) concluded that the reaction time (RT) is the shortest for congruent trials while RT for
incongruent trials is the longest. According to Macleod and MacDonald (2000), the executive aspects
of attentional control can be investigated with the help of the Stroop task. The Stroop Task may be
considered a difficult task because people cannot just turn off the automatic habit of reading words,
while the point of this task is to focus on the color and not on the meaning of words. The executive
aspects of attentional control are required to inhibit automatic word reading and at the same time
making the word color recognition, which is less automatic, the top priority during the Stroop task.
When word meaning and word color are incongruent, a cognitive conflict appears. As a result of that,
the cognitive system has to increase the level of control to guarantee successful behavior, which is
naming the right color of the word while ignoring the word meaning, thus causing a slowed response.
The slowed response in the incongruent trials compared to the congruent trials is known as the
Stroop effect (Jensen & Rohwer, 1966). According to McBride, Boy, Husain and Sumner (2012),

6
stimuli are processed by two routes at the same time. The processing of information via the direct
processing route occurs quickly and automatically and happens irrespective of whether task
instructions were given (e.g., reading Stroop words). At the same time, the task-relevant target
attribute (e.g., target color in the Stroop task) is processed via the slower indirect processing route. If
test subjects have to deal with congruent trials, then the same response will be activated by the
indirect and the direct processing routes, which will produce responses that are fast and correct.
However, if the test subjects have to deal with incongruent trials, then different responses will be
activated by the direct and indirect processing route, causing slower response times and increased
error rates. This is because the conflict is being resolved between the competing responses (e.g.
meaning and color of the Stroop words that are incompatible).
Deroost et al. (2012) examined the influence of sequence learning on the experience of the
Stroop effect. They utilized a so-called sequential Stroop task, in which the subjects learned a
perceptual sequence, expressed in terms of the color arrangements of the Stroop words (e.g., GREEN
or BLUE written in green). During the sequence learning task, the order of colors was determined by
sequential regularity. The experimenters observed that subjects had faster RTs over training,
indicating that better anticipation of the color was achieved over sequential training. So the faster
RTs in the sequenced condition were attributed to sequence learning effects (in other words, the
main effect of training condition showed that RTs in the sequence condition were lower than in the
random condition). Moreover, they found a decrease of the Stroop effect over training. Toward the
end of the experiment, the subjects were not affected any more by the Stroop effect (i.e., the RTs
derived from congruent word-color pairs did not differ significantly from the RTs derived from
incongruent pairs) although the Stroop effect was fully restored when the regularity was replaced by
random color sequences. The RTs of incongruent pairs became much higher than the RTs of
congruent pairs.
In a second experiment of Deroost et al. (2012), they examined whether subjects with
sequence knowledge would show smaller Stroop effects than subjects without that knowledge. What
was different from the first experiment was that during the training phase, all subjects were
confronted with neutral words (e.g., BIKE written in green) instead of Stroop color words. During the
training, half of the subjects were presented with a color sequence and the other half with colors
that randomly changed. After the training, subjects ended up in the testing phase in which Stroop
conditions were applied to see whether subjects who acquired sequence knowledge would be better
at handling conflict than subjects who did not receive sequence training. During this testing, the
same color sequence of the Stroop words (as the sequence presented in the sequenced training
condition) was used for both training groups. To see whether sequence learning had taken place, the
experimenters examined the difference in performance in the testing phase between the two trained

7
groups. The experimenters came to the conclusion that the same sequence learning effects were
found (just like in the first experiment of Deroost et al.) and that during the testing phase, subjects
with sequenced training experienced as much conflict as the subjects with random training. In other
words, the difference in RTs between congruent trials and incongruent trails in the sequence training
group was similar to the difference in RTs in the random training group. These results illustrated that
sequence learning had no beneficial effect on Stroop effect reduction. Subjects who acquired
sequence knowledge did not handle the Stroop conflict any better than subjects who did not go
through sequenced training.
In both experiments of Deroost et al., a 4-stimulus-to-2-response (4S-to-2R) mapping was
used in the serial reaction time (SRT) task to learn the color sequences. Two response keys were used
to specify the four colors (the C-key was used for a red or green word and the N-key for a blue or
yellow word. They investigated perceptual sequence learning in which probabilistic sequences were
used that were not entirely fixed beforehand. Subjects were asked to react to a fixed set of stimuli
over and over again in which each cue (i.e., ink color of the word) indicated that one of the two
keyboard keys (i.e., C-key or N-key) had to be pressed. Probabilities governed the transition between
the cues, so the appropriate key presses following one cue had a certain degree of predictability,
which could result in faster RTs to the cues because the test subjects could pick up and use these
probabilities of transition.
In this current study, not only perceptual sequencing (fixed color sequences were displayed
on the screen) was applied but also motor sequencing, which was relevant to examine because the
ability to perform motor sequences is a very important part of our life, as most actions we perform
on a daily basis consist of sequences of basic movements (such as lacing a shoe, playing piano,
changing clothes, etc.). Abrahamse, Ruitenberg, de Kleine and Verwey (2013) explained that motor
sequence learning is about acquiring the competence to quickly generate a sequence of movements
with limited effort and attentional monitoring. A task that is well-suited for studying motor sequence
learning is the discrete sequence production (DSP) task (Abrahamse et al., 2013), because the
development of automated skill could be monitored in a controlled environment. Moreover, this task
allowed test subjects to acquire sequencing skill very quickly (Ruitenberg et al., 2012). In this task,
two sequences of two to seven stimuli were displayed to subjects in a specific order that was
constructed beforehand. The intention was that they respond to the stimuli by pressing keys on a
keyboard. The more the test subjects practiced with the DSP task, the better they learned and
executed the sequences.
On the basis of research within the field of the discrete sequence production (DSP) task, the
Dual Processor Model of sequencing skill has been proposed (see Verwey, 2001). According to this
model, there are two separate processors also known as a cognitive processor and a motor

8
processor. These processors contribute to sequencing performance according to the degree of
practice, which ultimately leads to different forms of sequence execution. In the DSP task, a motor
sequence can be performed in different modes. In the reaction mode - usually displayed at the
beginning of a motor sequence performance – the execution of each key press of the sequence
occurs independently of the other key presses. The conversion of each stimulus into the appropriate
response is regulated by the cognitive processor. The motor processor ensures the production of the
actual motor response. When subjects have practiced their discrete sequence skills thoroughly, the
execution of the motor sequence can change into the chunking mode. In this mode, a motor
sequence is performed as one or more motor chunks. The selection, preparation and execution of all
separate elements of the sequence are no longer necessary as the execution of the complete
sequence become automatic. The whole sequence can be completed on the basis of the first
stimulus which functions as the trigger of the sequence that should be executed (Ruitenberg,
Abrahamse & Verwey, 2013). In this chunking mode, “(…) the cognitive processor selects motor
chunks and loads them into a temporary motor buffer, from which the motor processor then executes
these motor chunks in a relatively automatic—that is, autonomous—fashion (i.e., without the need
for cognitive involvement once initiated) (Ruitenberg et al. 2013, pp. 209)”.
In this study, a combination of the Stroop task and the DSP task will be used to examine the
role of motor sequence learning on Stroop effect. In brief, subjects will be randomly assigned to one
of two conditions: the sequenced training condition and the random training condition. During the
training phase, both groups have to react to the ink color of the so-called neutral words (i.e., words
that do not indicate a certain color, like the word BIKE presented in green ink) by pressing keyboard
keys. One group will be exposed to various neutral words that are presented in a certain ink color
sequence which is constructed beforehand. In the other group, the ink colors of the neutral words
occur in random order. The four different colors were mapped into four different response keys (4S-
to-4R) and it was expected that motor sequence learning would take place. In the test phase, Stroop
stimuli will be presented (Color words will be used instead of neutral words, e.g., the word GREEN in
blue ink) to see whether subjects who learned a color sequence in the training phase show a smaller
Stroop effect compared to subjects who did not learn a sequence.
The first hypothesis is that repeated execution of fixed response sequences assists in
sequence learning, because motor chunks could be used in the task. It is expected that subjects in
the sequence training group will show a larger decrease in RTs across the training blocks than
subjects in the random training group, because the former group is exposed to both task-specific
learning and sequence learning. The random training group is only exposed to task-specific learning,
which means that the potential gains in performance (e.g., faster RTs) are only due to repeated
practice on a task.

9
The second hypothesis is that the sequence training group will show smaller Stroop effects in
the test phase when they are exposed to color sequences that they had learned during the training
phase (familiar color sequences), compared to the exposure to unfamiliar color sequences. Subjects
in this group can react to these fixed color sequences by performing corresponding fixed motor
sequences. The moment subjects become familiar with these motor sequences, they would pay less
attention to the stimuli on the screen (e.g., meaning of the color words). In other words, the
sequence training group would experience less interference (i.e., Stroop conflict), because they could
use motor chunks in the test phase when Stroop stimuli are presented. When unfamiliar sequences
(which are identical to familiar sequences in terms of complexity) are presented in the test phase, it
is expected that the people in the sequence training group will show large Stroop effects, due to the
fact that they cannot use the motor chunks they previously had developed. Moreover, people in the
random training group will show large Stroop effects throughout the task, because of the same
reason that they cannot develop motor chunks which they could use in the test phase.
During the training phase and in one block of the test phase, each subject of the sequence
training group were presented with two different fixed color sequences (each sequence contained six
colors). It should be noted that the first stimulus of the color sequence was different from the
subsequent stimuli of the sequence, in terms of attention that need to be paid. On the basis of the
first stimulus (and the corresponding first key press), these subjects knew which of the two motor
sequences had to be performed, so they still had to pay attention to the first stimulus. After that, it
became less important to attend to the subsequent stimuli of the color sequence (2nd to 6th stimuli),
in order to perform the corresponding motor sequence, because then motor chunks could be used.
In this context, it was assumed that the 2nd to 6th key presses of a fixed motor sequence were the
most sensitive to motor sequence learning. The characteristics of the first key press and the last five
key presses of a motor sequence would be kept in mind in the data analysis.

10
Method
Subjects
Thirty two students from the university of Twente participated in the experiment (26 female
and 6 male, M age = 21.94 years, SD = 2.56 years). In order to participate in this study, students were
required to have German as their native language and had to be eighteen years or older. Moreover,
they should not have motoric problems in their arms and hands and needed normal eyesight (though
the use of glasses or lenses was acceptable). Last but not least, they had to report that they were not
colorblind in order to be eligible for this experiment. As compensation for their participation,
subjects were rewarded with course credits. The ethics committee at the faculty of behavioral
sciences of University Twente approved this experiment.
Apparatus
The task was conducted on a personal computer located in the laboratory of University
Twente. The PC had an Intel Core i7-3770 CPU @ 3.40 GHz processor with 8GB RAM and was running
a 64-bit version of Windows 7. In order to maximize computing resources for the E-Prime 2.0
software, which was used for the presentation of stimulus and collection of data, Windows was
started in a so-called lean mode (i.e., only the basic and essential Microsoft processes were allowed
to run).That way, the E-Prime 2.0 software was maximally responsive to the key presses on the
(qwerty) keyboard, so in turn accurate reaction times can be measured. The stimuli were presented
on a 22” LG Flatron E2210 monitor.
Task and Procedure
Before the actual experiment, the subjects were asked to give written informed consent if
they agreed with the conditions of the experiments. After they gave their permission, the experiment
could start. To give them a little impression of the experiment, information was given briefly about
the procedure of the study.
After that, specific instructions were given about the placement of their fingers on the
computer keyboard in order to perform the computer task correctly. Subjects were instructed to
place their index and middle fingers above the keyboard keys C, V, B and N. Each key represented a
certain color which subjects could press - one key at a time - to indicate what ink color of the word
was shown on the screen. For their convenience, a piece of paper was taped on the bottom of the
computer screen to help them remember what colors the keys stood for. After these instructions
were clear for the subjects, the computer program could start.
Firstly, they were presented with instructions on the screen in German to make sure they
really understand the verbal instructions. Then, subjects proceeded to the training phase that
consisted of six blocks in which neutral words were presented in a certain color in the middle of the

11
screen. The subjects had to react to the color by pressing the right key on the keyboard. The words
were displayed in Courier New and had a font size of 18. The sequence training group was presented
with two fixed predetermined color sequences. It was intended that they would react to these color
sequences by performing fixed predetermined motor sequences accordingly. The subjects learned
two sequences across six training blocks and each motor sequence consisted of 6 key presses. The
key sequences were counterbalanced across subjects in each group, which means that the middle
fingers and index fingers of every subject were used equally across the training- and test blocks.
Thus, each subject was presented with two different sequences that had to be initiated with two
different kinds of fingers (so one sequence began with the index finger and one sequence with the
middle finger) given that one sequence began with a finger from the left hand and the other
sequence with a finger from the right hand.
After the two groups completed the training phase, they proceeded to the test phase in
which only Stroop stimuli (i.e., color words in a certain ink color which could be either congruent or
incongruent; the distribution of congruence/incongruence was 50/50) were introduced. Each
congruent word-color pair (e.g., the word RED in red ink color) had to be shown three times as much
as each related incongruent word-color pair (i.e., RED in green, RED in yellow, RED in blue), in order
to keep the distribution of 50% congruent pairs and 50% incongruent pairs intact. The test phase
consisted of two blocks: test block X and test block Y (see table 1 for the design of the experiment).

12
Table 1
Design of the Experiment
Condition Training phase Test phase
Sequence training 6x training blocks with
neutral words
following fixed color
sequences
1x test block X with
color words
(or test block Y as first
test block)
1x test block Y with
color words
(or test block X as first
test block)
Random training 6x training blocks with
neutral words
presented in random
colors
1x test block X with
color words
(or test block Y as first
test block)
1x test block Y with
color words
(or test block X as first
test block)
Note. When subjects in the sequence training conditon were exposed to test block X, the same color
sequences were presented as in the training phase. Test block Y contained color sequences which
were unfamiliar for them. Subjects in the random training condition were presented with unfamiliar
sequences in both test block X and testblock Y. Subjects in the sequence training condition were
exposed to the same color sequences as the subjects in the random training condition during the test
phase.
In condition one, subjects were presented with the sequences “ncbvbc” and “vnbcnv” across
the six training blocks and in block X of the test phase. In condition two, three and four, subjects
were presented with the sequences “bnvcvn” and “cbvnbc”, “vbcncb” and “nvcbvn”, “cvnbnv” and
“bcnvcb”, respectively. In block Y of the test phase, subjects got the same sequences from the other
conditions. Specifically, subjects in condition one got the two sequences from condition two in the
unfamiliar block. In condition two, three and four, subjects got the two sequences from condition
three, four and one in the unfamiliar block, respectively.
The random training group was presented with random colors, so no fixed sequence was
used. Note that the 16 subjects in the random training group were presented with the exact same
test block X and test block Y as the 16 subjects in the sequence training group (e.g., subject #8 in the
random training group got the same sequences in test block X and Y as subject #8 in the sequence
training group). Furthermore, the order of test blocks was counterbalanced across subjects in each
group in the test phase.

13
If subjects could identify the color of the word by pressing the appropriate key, then it was
considered a correct answer. Specifically, in the training phase, they had to react to the ink color of
the neutral words (“Boot” = boat, “Müll” = trash, “Held” = hero and “Frau” = woman), and during the
test phase, they had to react to the ink color of the color words (Rot” = red, “Grün” = green, “Gelb” =
yellow, and “Blau” = blue). If the subjects pressed the wrong key, then they got a message on the
screen indicating that they got the wrong answer (“Falsch”). Subjects could also react too quickly by
accident (i.e., before the actual colored word was presented). If this was the case, they got the
message “Zufrüh” which means “Too early” in German. In the sequence training group, each block
had 48 trials and each trial consisted of 6 key presses (288 key presses in total). The order of
executing sequence 1 or sequence 2 is randomly generated, given that each motor sequence was
presented 24 times per block. In the random training group, 288 individual color-word combinations
were presented. Halfway through each block there was a break of 30 seconds. After each block,
subjects had the opportunity to take a break of a couple of minutes.
Results
Training phase
In figure 1, mean RTs of the sequence training group of each key of the fixed motor sequence
are presented for each training block. It seemed that all key presses improved over the training
blocks, especially for key press 2 to 6. According to figure 2, it seemed that in the random training
group, RTs of all key presses did not become lower over the training blocks. To test whether these
assumptions were true, a mixed factorial repeated measures ANOVA with Block (6; training blocks 1 -
6) and Key Press (6; key 1 - 6) as within-subject factors and Group (2; sequence training vs. random
training) as a between-subject factor was performed.
.

14
Figure 1. Mean RTs (in milliseconds) per key of the sequence for each training block
(block 1 – 6 indicated as B1 – 6) of the sequence training group

15
Figure 2. Mean RTs (in milliseconds) per key of the sequence for each training block
(block 1 – 6 indicated as B1 – 6) of the random training group. Compared to figure 1,
different scales and a different starting point of the Y-axis were used.
A main effect of Block showed that the mean RTs differed across the training blocks,
F(5, 150) = 45.74, p < .001. Furthermore, a main effect of Key Press showed that some key presses
were executed faster than others, F(5, 150) = 58.23, p < .001. There was also a significant Block x Key
Press interaction effect, F(25, 750) = 3.09, p < .001, which indicated that the RT differences between
the training blocks varied across the key presses. Moreover, a Block x Group interaction effect, F(5,
150) = 13.55, p < .001, showed that there is more difference in RTs across the practice blocks in one
of the two training groups. It was decided not to explore these main effects and two-way interaction
effects mentioned above any further, because these effects varied with a third variable: there was
also a significant Key Press (6) x Block (6) x Group (2) interaction effect.
This three-way interaction effect indicated that the RT differences between the training
blocks that varied across the key presses were not the same in the two training groups. It was

16
assumed that the Block x Key Press interaction effect of the sequence training group would be
different than the Block x Key Press interaction effect of the random training group, since the former
group was not only exposed to task-specific learning, but also sequence learning (also figures 1 and 2
point in this direction). To test whether this was true, two separate repeated measures ANOVA’s
(one for the sequence training group and one for the random training group) with Block (6) and Key
Press (6) as within-subject factors were conducted. Results showed that there was a Block x Key
Press interaction effect in the sequence training group, F(25, 375) = 3.71, p < .001, but not in the
random training group, F(25, 375) = 1.12, p = .32, indicating that the RT differences between the
training blocks varied across the different key presses in the sequence training group (see figure 1).
However, this was not the case in the random training group (see figure 2), which showed that the
improvement of key presses over the training blocks (due to task-specific learning) was not
significant. (In other words, the RT differences between key presses varied across the training blocks
in the sequence training group, but not in the random training group).
It was assumed that the RT difference between block 6 and block 1 would capture the full
effects of motor sequence learning and task-specific learning. Figure 1 gave the impression that the
differences in RT between block 1 and 6 were significant for all keys. To examine whether this was
actually true, separate independent-samples t-tests were conducted for the sequence training group
to see whether RTs of block 6 were significantly higher than RTs of block 1 for each key press. Results
showed that the RTs of block 1 were not significantly different from the RTs of block 6 (754 ms vs.
650 ms), for key press 1, t(30) = 1.97, p = .06. However, RTs of block 1 were significantly higher than
RTs of block 6 for key press 2 (592 ms vs. 350 ms), key press 3 (576 ms vs. 358 ms), key press 4 (507
ms vs. 279 ms), key press 5 (656 ms vs. 344 ms), and key press 6 (545 ms vs. 288 ms), t(30) > 3.43, p <
.01. These t-tests results together with the significant Block x Key Press interaction effect found in
the sequence training group and figure 1, suggested that there was a gradual decrease in RTs for key
press 2 to 6 over the training blocks. Note that these t-tests above specifically showed that the RT
difference between block 1 and 6 was significant for key press 2 to 6.
Test phase
Important results of the test phase are displayed in figures 3 to 5. Results of a mixed factorial
repeated measures ANOVA with Test Block (2), Congruency (2; congruent trials vs. incongruent trials)
and Key Press (6) as within-subject factors and Group (2) as a between-subject factor showed a main
effect of Test Block which indicated that sequences were performed faster in the familiar block than
in the unfamiliar block (544 ms vs. 651 ms), F(1, 30) = 33.93, p < .001. A main effect of Congruency
showed that RTs were faster in the congruent trials than in the incongruent trials (574 ms vs. 620
ms), F(1, 30) = 54.83, p < .001. Furthermore, a main effect of Key Press showed that some key
presses were executed faster than others, F(5, 150) = 39.48, p < .001. Moreover, there was a Test

17
Block x Group interaction effect that showed that the difference in reaction times between the
familiar and unfamiliar blocks was larger in one of the two training groups, F(1, 30) = 34.68, p < .001.
A Key Press x Group interaction effect showed that the difference in reaction times between key
presses was not the same in the two training groups, F(5, 150) = 9.30, p < .001. Furthermore, a Test
Block x Key Press interaction effect indicated that the RT differences between key presses are not the
same across the test blocks, F(5,150) = 3.02, p < .01. It was decided not to examine the main effect of
Key Press and the two-way interaction effects mentioned above any further, because these effects
varied with a third variable: there was also a significant Test Block (2) x Key Press (6) x Group (2)
interaction effect.
This three-way interaction effect indicated that the differences in RTs between the test
blocks that varied the key presses were not the same in the two training groups, F(5,150) = 2.89, p <
.05. It was expected that the Test Block x Key Press interaction effect of the sequence training group
would differ from the Test Block x Key Press interaction effect of the random training group, because
the former group was exposed to sequence learning. To test whether this was true, two separate
repeated measures ANOVA’s (one for the sequence training group and one for the random training
group) with Test Block (2) and Key Press (6) as within-subject factors were performed. Results
showed that there was a Test Block x Key Press interaction effect in the sequence training group, F(5,
75) = 3.92, p < .01, but not in the random training group, F(5, 75) = .60, p = .70, indicating that the
differences in RTs between the test blocks varied across the key presses in the sequence training
group, but not in the random training group. According to figure 3, it seemed that the RTs of test
block Y were significantly higher than the RTs of test block X for all key presses in the sequence
training group. To test whether this was true, separate independent-samples t-tests were conducted
for the sequence training group for each key press. Results showed that the RTs of test block X were
not significantly different from the RTs of test block Y (804 ms vs. 710 ms), for key press 1, t(30) = -
1.05, p = .30. However, RTs of test block Y were significantly higher than RTs of test block X for key
press 2 (619 ms vs. 386 ms), key press 3 (573 ms vs. 381 ms), key press 4 (525 ms vs. 306 ms), key
press 5 (660 ms vs. 340 ms), and key press 6 (512 ms vs. 271 ms), t(30) < -2.80, p < .01, in the
sequence training group.

18
Figure 3. Mean RTs (in milliseconds) per key for the two test blocks in the sequence training group
Moreover, a Congruency x Key Press interaction effect showed that the RT difference
between incongruent trials and congruent trails (i.e., RT of incongruent trials minus RT of congruent
trials) varied across the key presses, F(5, 150) = 3.58, p < .01. According to figure 4, it seemed that
the RTs incongruent trials were significantly higher than the RTs of congruent trials for all key
presses. To test whether this was true, separate independent-samples t-tests were conducted to
compare the RTs of incongruent trials with RTs of congruent trials for each key press. Results showed
that RTs of incongruent trials were significantly higher than RTs of congruent trials for key press 1
(810 ms vs. 714 ms), t(62) = -2.25, p < .05, but this was not the case for key press 2 (612 ms vs. 572
ms), key press 3 (579 ms vs. 543 ms), key press 4 (527 ms vs. 496 ms), key press 5 (631 ms vs. 590
ms), and key press 6 (562 ms vs. 517 ms), t(30) > -.93, p > .35.

19
Figure 4. Mean RTs (in milliseconds) per key for the congruent and incongruent trials
The Test Block (2) x Congruency (2) x Group (2) interaction effect was not significant, F(1, 30)
= 3.26, p = .08, when all key presses were considered in the analysis. It was expected that the
influence of key press 1 may prevent this 3-way interaction to be significant, since key press 1 was
not as sensitive to motor sequence learning as the subsequent key presses. That was the reason why
another repeated measures ANOVA with Test Block (2), Congruency (2), Key Press (5) as within-
subject factors and Group (2) as a between-subject factor was carried out (RTs of key press 1 were
omitted). Results showed that Test Block x Congruency x Group interaction effect was still not
significant, F(1, 30) = 3.25, p = .08.
To specifically test the hypothesis that the sequence training group would show smaller
Stroop effects in test block X (with familiar sequences), than in test block Y (with unfamiliar
sequences), a focused repeated measures ANOVA with Test Block (2), Congruency (2) and Key Press
(5) as within-subject factors was performed for the sequence training group (RTs of key press 1 were
omitted in this analysis). A Test Block x Congruency interaction effect was found in the sequence
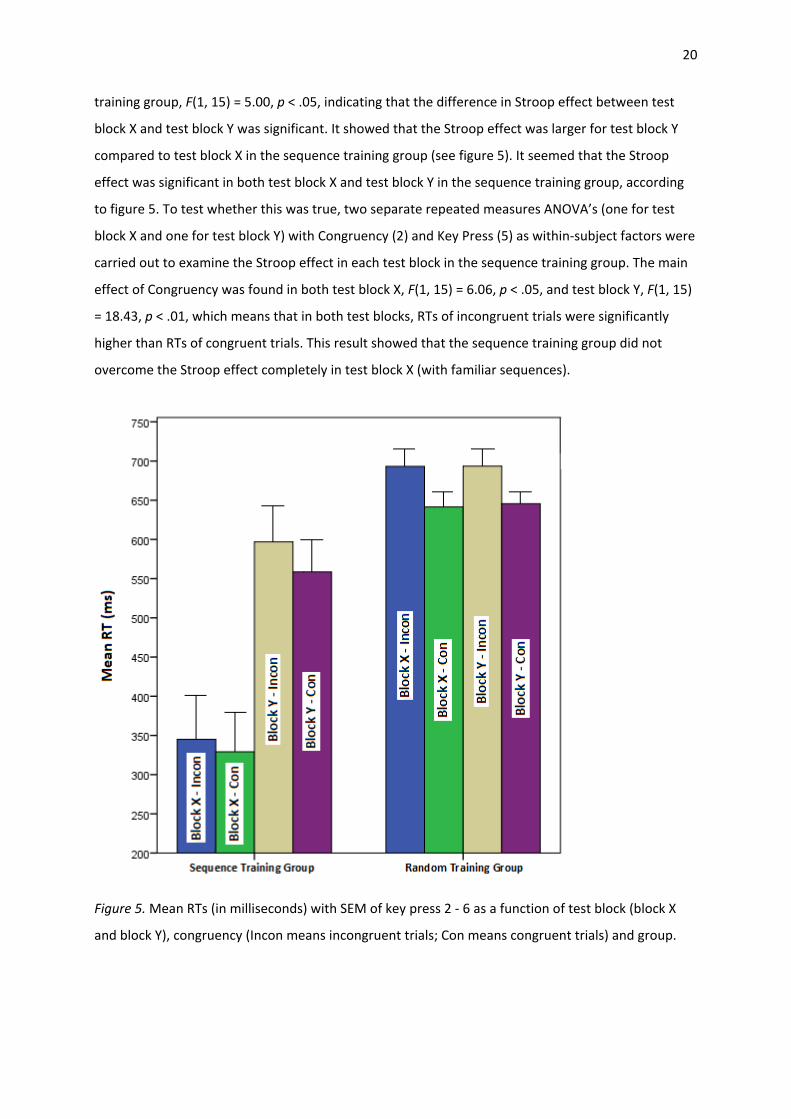
20
training group, F(1, 15) = 5.00, p < .05, indicating that the difference in Stroop effect between test
block X and test block Y was significant. It showed that the Stroop effect was larger for test block Y
compared to test block X in the sequence training group (see figure 5). It seemed that the Stroop
effect was significant in both test block X and test block Y in the sequence training group, according
to figure 5. To test whether this was true, two separate repeated measures ANOVA’s (one for test
block X and one for test block Y) with Congruency (2) and Key Press (5) as within-subject factors were
carried out to examine the Stroop effect in each test block in the sequence training group. The main
effect of Congruency was found in both test block X, F(1, 15) = 6.06, p < .05, and test block Y, F(1, 15)
= 18.43, p < .01, which means that in both test blocks, RTs of incongruent trials were significantly
higher than RTs of congruent trials. This result showed that the sequence training group did not
overcome the Stroop effect completely in test block X (with familiar sequences).
Figure 5. Mean RTs (in milliseconds) with SEM of key press 2 - 6 as a function of test block (block X
and block Y), congruency (Incon means incongruent trials; Con means congruent trials) and group.

21
Discussion
The current study investigated whether motor sequence learning can help in the reduction of
Stroop effect. Subjects performed a computer task in which words were presented in a certain ink
color. They were instructed to only react to the ink color of the words while ignoring word meanings.
The sequence training group was presented with fixed color sequences in the training phase (with
neutral words), so they had to react to these sequences by performing fixed motor sequences (i.e.,
fixed sequences of key presses). Subjects of the random training group were presented with neutral
words displayed in random ink colors, so no sequence learning took place. In the test phase, subjects
had to react to Stroop stimuli (i.e., color words displayed in a certain ink color). The reaction times
between congruent trials and incongruent trials would indicate the degree of Stroop effect.
Results of the study showed that the sequence training group showed a larger decline in RTs
over the training blocks than the random training group. In other words, the former group learned
more of the fixed motor sequences than the latter group. Furthermore, the sequence training group
would show significant smaller Stroop effect when these subjects performed familiar motor
sequences compared to when they performed unfamiliar sequences, but this was only true when RTs
of key press 1 were omitted. This was not surprising because it was expected that only key press 2 to
6 were responsive to motor sequence learning.
In regard to the Dual Processor Model of sequencing skill (Verwey, 2001), the subjects in the
sequence training group could use motor chunks to help them perform the task. During the training
phase, subjects of this group had practiced fixed motor sequences. The execution of these sequences
could develop into the so-called chunking mode, which made it possible that an entire key press
sequence could be executed as one (or more) motor chunks. On the basis of the first key press, the
whole motor sequence could be performed as this key press functioned as the trigger of the fixed
key press sequence that had to be completed. A possible reason why the subjects of the sequence
training group showed decreased Stroop effects in the test phase, was because after the
presentation of the first color stimulus (and the first key press), they paid less attention to the 2nd to
6th stimuli of the fixed color sequence presented on the screen (because this was not necessary for
completing the 2nd to 6th stimuli of a fixed motor sequences). So the subjects of this group paid less
attention to the word meaning of the 2nd to 6th stimuli, which resulted in reduced experience of
Stroop conflict for these stimuli.
The results (regarding Stroop effect) of this study were not in line with the results of the
second experiment of Deroost et al. (2012) which showed that sequence learning had no positive
impact on Stroop effect reduction. It was possible that the kind of task (Serial Reaction Time task vs.
DSP task) and the kind of sequence learning (perceptual sequence learning vs. motor sequence
learning) test subjects had encountered during the experiment plays an important role in whether

22
Stroop effect reduction would take place. In contrast to this study in which motor sequence learning
was examined, Deroost et al. (2012) investigated perceptual sequence learning in which probabilistic
sequences were used that are not entirely fixed beforehand. This seemed to be the main reason why
the results were not in line. Subjects were asked to react to a fixed set of stimuli over and over again
in which each cue (i.e., ink color of the word) indicated that one of the two keyboard keys (i.e., C-key
or N-key) had to be pressed. Probabilities governed the transition between the cues, so the
appropriate key presses following one cue have a certain degree of predictability, which could result
to faster RTs to the cues because the test subjects can pick up and use these probabilities of
transition. These perceptual sequences did not have a clear starting and ending point of the
sequence, unlike the discrete motor sequences used in this study. Furthermore, subjects in the study
of Deroost may not learn the sequences as well as in this study. The findings of this study had
implications for the theory: it seemed that sequences which were completely fixed beforehand and
which subjects can learn and utilize relatively easy (each motor sequence only consisted of 6 keys)
resulted in the above mentioned reduced Stroop effects.
Future research could focus on whether there is a reduction of Stroop effect when subjects
had to perform this task for multiple times (for example, they were instructed to complete this task
once every day for one month). They would get the same training blocks and test blocks as if they
were in the random training condition in this study (no fixed motor sequences would be presented
during the training phase). Every day, they should receive different (unfamiliar) fixed sequences in
the test phase, to avoid the possibility that they would learn the fixed sequences. If significant Stroop
effect reduction is found over time, this would suggest increased cognitive control, because they
would have to identify each stimulus, in order to press the right keys. In this case unimportant
distractors (meaning of the color word) were perceived and were successfully rejected.
There were certain limitations in this study. Only a small Stroop effect reduction was
observed between the two test blocks in the sequence training group. Maybe this is due to the
number of blocks in the training phase. A stronger decrease of Stroop effect could be found when
more training blocks were used in this task and subjects of the sequence training group may even
overcome the Stroop effect in the test block in which familiar sequences were presented.
Furthermore, even though the question was asked in the beginning of the experiment whether
subjects had color-blindness (all subjects said they do not), the possibility cannot be excluded that
some subjects may have various degrees of (partial) color-blindness without them knowing. It would
help to actually test all subjects on color-blindness with the help of the Test for Color Blindness by
Ishihara (1917). Moreover, some of the subjects reported that the colors green and yellow were
difficult to distinguish from each other. If the intensities of these two colors were higher, then it
would be easier to tell them apart.

23
In summary, this present study showed that subjects in the sequence training group showed
smaller Stroop effects when they performed motor sequences that they previously had learned
compared to when they were exposed to unfamiliar sequences. This result can be explained by
motor chunks from the Dual Processor Model of sequencing skill. Future research could focus on
similar tasks in a repeated setting, without the use of fixed sequences. In sum, this current study
showed that motor sequence learning can contribute to less perceptual attention, which in turn can
cause reduced Stroop conflict.

24
References
Deroost, N., Vandenbossche, J., Zeischka, P., Coomans, D., & Soetens, E. (2012). Cognitive Control: A
Role for Implicit Learning. Journal of Experimental Psychology: Learning, Memory,
and Cognition, 38, 1243-1258. doi: 10.1037/a0027633
Ishihara, S. (1917). Tests for color-blindness. Handaya, Tokyo, Hongo Harukicho.
Jensen, A. R., Rohwer, W. D. (1966). The Stroop color-word test: a review. Acta psychologica, 25(1):
36–93. doi:10.1016/0001-6918(66)90004-7
Langer, E. J. (1978). Rethinking the role of thought in social interaction. Hillsdale, NJ: Erlbaum.
Lavie, N., Hirst, A., de Fockert, J., & Viding, E. (2004). Load theory of selective attention and
cognitive control. Journal of Experimental Psychology: General, 133(3), 339-354.
Lavie, N. (1995). Perceptual load as a necessary condition for selective attention. Journal of
Experimental Psychology: Human Perception and Performance, 21, 451–468.
MacLeod, C. M. (1991). Half a century of research on the Stroop effect: An integrative review.
Psychological Bulletin, 109, 163–203. doi:10.1037/0033-2909.109.2.163
MacLeod, C. M., & MacDonald, P. A. (2000). Interdimensional interference in the Stroop effect:
Uncovering the cognitive and neural anatomy of attention. Trends in Cognitive Sciences, 4,
383–391. doi:10.1016/S1364-6613(00)01530-8
McBride, J., Boy, F., Husain, M., & Sumner, P. (2012). Automatic motor activation in the executive
control of action. Frontiers in human neuroscience, 6, doi: 10.3389/fnhum.2012.00082
Ruitenberg, M. F. L., Abrahamse, E. L., de Kleine, E., & Verwey, W. B. (2012). Context-dependent
motor skill: perceptual processing in memory-based sequence production. Experimental
Brain Research, 222, 31-40. doi: 10.1007/s00221-012-3193-6
Ruitenberg, M. F. L., Abrahamse, E. L., & Verwey, W. B. (2013). Sequential motor skill in
preadolescent children: The development of automaticity. Journal of Experimental Child
Psychology, 115, 607-623. doi: http://dx.doi.org/10.1016/j.jecp.2013.04.005
Stroop, J. (1935). Studies of interference in serial verbal reaction. Journal of Experimental Psychology,
18, 643-662.
Verwey, W. B. (2001). Concatenating familiar movement sequences: the versatile cognitive
processor. Acta psychologica, 106, 69–95.