the growth o mixef d populations o chilomonasf...
TRANSCRIPT

251
The Growth of Mixed Populations of Chilomonasparamecium and Tetrahymena patula
By SMILJA MUCIBABIC"
(From the Philosophical Faculty, Sarajevo, Yugoslavia)
With one plate (fig. i)
SUMMARY
In mixed cultures, where Tetrahymena patula preys on Chilomonas paramecium,the growth of the Chilomonas population does not initially differ significantly from thatin single cultures. Later, however, the decrease in numbers of Chilomonas is morerapid in mixed cultures, where it dies out after two months.
The composition of the polymorphic Tetrahymena patula population depends onage and on the presence or absence of Chilomonas. Slit-mouthed forms are absent atthe beginning of growth in both mixed and single cultures, and only appear sub-sequently in mixed cultures. The percentage of these forms increases in older cultures.The percentage of microstomatous forms is larger, however, at the beginning ofpopulation growth, both in mixed and single cultures. The percentage of macrosto-matous forms increases from the beginning, reaches a maximum, and then decreases.Cannibalism has been observed in old mixed cultures.
Significant differences in the size of individual organisms of Chilomonas betweensingle and mixed cultures appeared from the beginning of the logarithmic phase ofgrowth, and were pronounced up to the end of observations. Significant differencesin the size of Tetrahymena patula between single and mixed cultures appeared some-what later, from the sixth day onwards.
The differences in size and shape between organisms in cultures of the same kindbut of different ages were also significant. Variation in size and shape of both specieswas greater in mixed than in single cultures.
INTRODUCTION
EXPERIMENTAL studies of the relations between predator and preyhave not been as numerous, nor have they been made on as many different
species, as have studies on competition between organisms. Gause (1934,1935a, 19356) showed that their interaction did not invariably lead to periodicoscillations of population density, as predicted in the mathematical theoriesof Lotka (1925) and Volterra (1926), but that the occurrence of oscillationdepended on certain characters of the species under investigation, and onenvironmental factors. He observed (Gause, 19356) oscillations with Para-mecium bursaria and Saccharomyces pombe, provided that yeast was addedto the medium from time to time, and that the population of the predatorwas decreased by taking samples. In other experiments (Gause, 1935a),where Bursaria truncatella was predator and Paramecium bursaria prey,,periodic fluctuations did not appear, however, and Bursaria died out beforethe paramecia were destroyed. Gause does not mention that the relationship'was more complex here than in his previous experiments on Paramecium[Quarterly Journal of Microscopical Science, Vol. 98, part 2, pp. 251-63, June 1957.]

252 Mucibabid—Mixed Populations of
bursaria and Saccharomyces pombe; but evidently the food chain of Bursaria-Paramecium-Saccharomyces was more complex than the simple relation ofpredator to prey. According to Sandon (1932), Bursaria truncatella feeds onciliates as well as taking other food. In Gause's experiments Bursaria mayhave preyed both on Paramecium and on yeast. According to Lund's obser-vations (1914a, 19146), Bursaria truncatella is selective in its feeding, since itaccepted fragments of hard-boiled yolk stained with dyes insoluble in water,but refused fragments of yolk stained with dyes soluble in water; but Schaeffer(1917), was unable to agree that Bursaria had any food-selecting mechanismwhatsoever; the occasional refusal of food was only part of a general reactionto more or less injurious stimuli. If Bursaria is not selective it is probablethat it was indeed feeding on both paramecia and yeast in Gause's experi-ments.
Gause (1934) reported that Didinium nasutum, a voracious predator,destroyed Paramecium caudatum and then died out; fluctuations in thedensities of their populations did not normally appear. Only when theirinteraction was modified by the presence of a 'refuge' (a sediment of yeast)or when, as in other experiments, he introduced both species into the cultureat regular intervals of time, did fluctuations occur.
Brown (1940) observed the growth of mixed populations of Leucophryspatula preying on Glaucoma pyriformis. He was interested in the logarithmicphase of the growth of Leucophrys and he prepared both theoretical andobserved growth curves for that phase, but followed the growth of thecultures for 4 days only, so that his experiments give very limited informationabout predator-prey relations. In his equations for rates of growth, Browntook the values for Leucophrys from the graph representing its growth insingle populations, but he did not define the conditions under which it wasgrown in single populations. At that time L. patula (Tetrahymena patulaaccording to Corliss, 1952) had not yet been established in bacteria-freecultures. It is doubtful, therefore, whether he was justified in taking valuesfor its growth from graphs based on figures derived from growth under quitedifferent conditions. Nor is it clear why Brown ascribed the end of theexponential phase to the high concentration of predator in the one instance,and to the exhaustion of food in the other. The density of Leucophrys was ashigh in the second instance as in the first, or even higher, as is shown by thecurves. He does not state from how many cultures or samples data wereobtained.
The study made by Dewey and Kidder (1940) of the growth of Perispiraovum with Euglena gracilis gives more details and covers a longer period thanBrown's data. It is interesting to note that Perispira continued multiplyingfor some time after the extinction of its prey, and at the expense of the size ofindividuals.
Lilly's study (1942) of the nutrition of Stylonychia pustulata and Pleuro-tricha lanceolata is also important as a contribution to knowledge of thegrowth of mixed populations. It reveals how small details gathered from

s.
S. MUClBABlC

Chilomonas and Tetrahymena 253
observation of the species may be of value in explaining the growth of thepopulation. At the same time, it implies how many variables are necessarilyneglected, if the growth of mixed populations—for example, the interactionbetween predator and prey—is approached purely mathematically. Lillyconfirmed that the length of the lag phase for these ciliates depended on theage of the inoculum. He also found that the size of the organisms is importantin relation to the duration of the lag phase, which lasted longer if the inoculumconsisted of small organisms, because these are not able to ingest large foodorganisms until they have increased in size. Lilly compared the phases ofgrowth of both predators, and reported that they were characteristic for eachspecies. Differences between the growth of their populations also appearedwhen the food was exhausted: Pleurotricha continued to grow, whereasStylonychia did not.
According to this review of previous work, the growth of mixed populationsof two protozoan species which are related as predator to prey, has previouslybeen studied either in small communities composed of three species at least,or on two species in bacteria-free cultures but without control experiments.It seemed desirable, therefore, to examine the predator-prey relationship ina community of two species with control cultures of the single species.
MATERIAL AND METHODS
In preliminary experiments, two species of Protozoa were sought, able tolive separately in pure axenic culture in the same organic medium, but which,when brought together, interact as predator and prey. Chilomonas parameciumserves as prey to many ciliates, but since most of these are carnivorous formsthat cannot live without food organisms, even in an organic medium, choicewas restricted to those few ciliates which have been obtained in bacteria-freecultures. Among the flagellates, Peranema trichophorum feeds upon Chilomonas(Chen, 1950), but it cannot live with Chilomonas in the same organic medium.An attempt was first made to use Tetrahymena vorax (strain V2) as predator.Mixed cultures were inoculated with macrostomatous forms only, becausethese are known to be carnivorous. They fed on Chilomonas, but after 3 to 4days all macrostomatous forms had disappeared, and the population wascomposed of microstomatous forms only, which are not predaceous (Kidder,Lilly, and Claaff, 1940). This species, therefore, was abandoned. Subse-quently, Tetrahymena patula (Miiller) Corliss was found to be suitable forthe present experiments; it takes Chilomonas as food (fig. r, A) and can also
FIG. I (plate). A, Tetrahymena patula with ingested Chilomonas paramecium, from a culture6 days old.
B, macrostomatous, microstomatous, intermediate, and slit-mouthed forms of Tetrahymenapatula.
C, cannibalistic forms of T. patula.D, forms of T. patula with re-entrant posterior end.E, forms of T. patula showing longitudinal folds.Each smaller division of the scale represents iOju.

254 Mucibabid—Mixed Populations of
live as a saprophyte. Subcultures were obtained from the Culture Collectionof Algae and Protozoa, Botany School, Cambridge.
Maupas (1888) wrote on Tetrahymena patula under the name Leucophryspatula, giving a description of the. species and many observations on its lifecycle. Faure-Fremiet (1948) extended Maupas's description, retaining thename Leucophrys. Recently, however, Corliss (1952, 1953) gave a full accountof the history, systematics, and morphology of this species as Tetrahymenapatula. T. patula was isolated before 1942 by Faure-Fremiet (Corliss, 1952).
It was in principle desirable to perform experiments with the same mediumas had been used in the experiments on T. pyriformis (Mucibabic, 1957) butthis was not possible because in the medium of o-i% proteose peptone ando-i% sodium acetate, T. patula did not continue to multiply after one or twoinitial divisions. The medium became satisfactory, however, when theconcentration of proteose peptone was increased from o-i% to 1%.
Cultures in 1% proteose peptone were inoculated with 10 Chilomonas and10 Tetrahymena patula, and control cultures were inoculated with 10 organismsof each species separately. All cultures were maintained at 22-5° C. Thenumber of organisms was counted every day as in previous studies (1956);but since the density of population of Chilomonas differed greatly from thatof Tetrahymena patula from the third day onwards, the size of the twopopulations could not be determined in the same way. Five samples fromeach culture were counted in order to determine the size of the Chilomonaspopulation; while the entire population of Tetrahymena patula in each culturewas counted throughout the period of the experiment. As the culture had tobe diluted before the population of Chilomonas could be counted, Tetra-hymena patula was counted before dilution. The counting-procedure was asfollows.
One drop of culture was placed on a slide. Since during the counting ofT. patula it was not possible to pick up T. patula alone, the organisms collectedin the capillary micro-pipette during counting were not discarded, but weredelivered into a test-tube containing 5 to 10 ml of fresh medium. The nextdrop was placed on the same spot on the slide, and so on, till the whole culturehad been examined. When the last drop had been taken, 0-3 ml of the mediumwas pipetted into the culture test-tube. This was also examined, in order topick up any organisms that might have remained on the wall of the test-tube.The individuals of Chilomonas remaining in the drop on the slide were washedfrom the slide into the large test-tube in which the whole culture had beendiluted. For washing, 5 ml of the fresh medium was used. Finally, the neces-sary quantity of medium was added to complete the required dilution. Fromthis diluted culture 5 samples were then counted, in order to determine thesize of the Chilomonas population.
Growth of single and mixed populations of Chilomonas paramecium andTetrahymena patula in terms of total number of organismsPopulation growth of Ch. paramecium and Tetrahymena patula in terms of

Chilomonas and Tetrahymena 255
total number of organisms is shown in fig. 2 and table 1 (p. 256). The tableshows mean size of the population, standard deviation, and number ofobservations for both species in single and mixed cultures. Table 1 and fig. 2
>monas. single
12 14 16 18 20 22 24 26 28 30 32 34Population aqe in days
FIG. 2. Graph showing the growth of populations of Chilomonas paramecium and Tetrahymenapatula in single and mixed cultures, in terms of total number of organisms.
reveal that the growth rate of the Chilomonas population is very similar insingle and mixed cultures. A very pronounced lag-phase appeared in bothsingle and mixed cultures in this medium. It lasted for 2 days. The maximumnumbers of Chilomonas in single and mixed cultures do not differ significantly.
At the beginning, the growth of the population of Tetrahymena patula isvery similar in single and mixed cultures. Differences appear from the 3rdday onwards and later become greater. From the 6th day, the population ofTetrahymena in single cultures begins to degenerate: the organisms becomevery opaque, flattened, and abnormal, and their number decreases rapidly.In mixed cultures, however, tetrahymenae were multiplying until the 32ndday. On the 6th day they began to take Chilomonas more noticeably thanbefore, and organisms with ingested individuals of Chilomonas were frequent(fig. 1, A). The following day the population of Tetrahymena patula wasnearly twice as great. This outbreak of division of Tetrahymena after increasedpredatory activity on the 6th day was observed in both series of experiments.The population of Tetrahymena increased from then onwards with smallfluctuation till the end of experiment. Since the single culture of Tetrahymenahad died out by the 18th day, counting of mixed cultures was not continuedafter the 20th day, but occasional observations were made from time to time.On the 32nd day the entire population of Chilomonas and of Tetrahymena
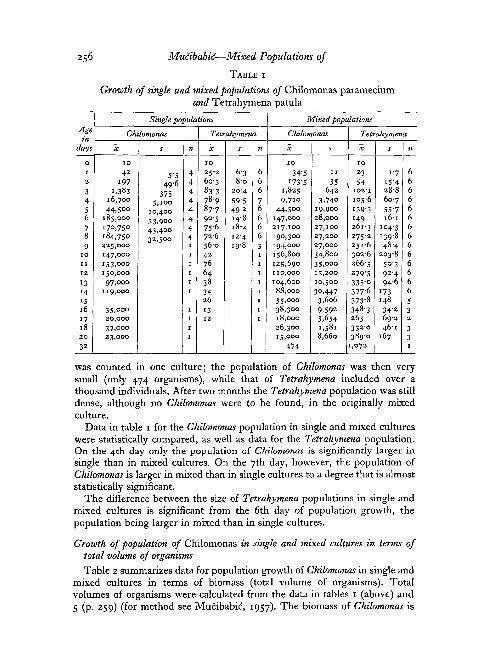
256 Mucibabid—Mixed Populations of
TABLE I
Growth of single and mixed populations of Chilomonas parameciumand Tetrahymena patula
Agein
days
0
1
2
3456789
1 0
1 1
1 2
13' 41516
»7182 0
3Z
Single populations
Chilomonas
X
1 0
42197
1,38316,70044.5OO
185,000170,750!84.75O225,000147,000153,000150,00097,000
119,000
35,ooo20,00037,00023,000
j
5'549'6
3755,100
10,40033,90043,40032,500
n
44444444
Tetrahymena
X
1 0
25-260-383-378-987-79 0 575-672-65 6 04 2
76643834261 3
1 2
s
6 38-o
20-459-549'21481 8 2
12-4198
n
666766663
Mixed populations
Chilomonas
X
1 0
34-5173-5
1.8259,710
44,5oo147,000217,100190,300194,000156,800125,690110,000104,60088,00055,ooo38,30018,00026,30015,000
474
1 1
55642
3,74O19,90028,00027,10027,20027,00034,80035,00015,20010,5003o,4473,6069,5925,6541,5818,660
Tetrahymena
X
1 0
23
54102-1105-6139-31 4 9
2613275-22516302-626652795335-O377-6373-8348-3265332-03890
1,072
1-7
iS'428860755"716-1
104-5139-84 8 4
203-85O-392-49 4 6
17314834-269-246-1
167
n
66666666666666532
331
was counted in one culture; the population of Chilomonas was then verysmall (only 474 organisms), while that of Tetrahymena included over athousand individuals. After two months the Tetrahymena population was stilldense, although no Chilomonas were to be found, in the originally mixedculture.
Data in table 1 for the Chilomonas population in single and mixed cultureswere statistically compared, as well as data for the Tetrahymena population.On the 4th day only the population of Chilomonas is significantly larger insingle than in mixed cultures. On the 7th day, however, the population ofChilomonas is larger in mixed than in single cultures to a degree that is almoststatistically significant.
The difference between the size of Tetrahymena populations in single andmixed cultures is significant from the 6th day of population growth, thepopulation being larger in mixed than in single cultures.
Growth of population of Chilomonas in single and mixed cultures in terms oftotal volume of organisms
Table 2 summarizes data for population growth of Chilomonas in single andmixed cultures in terms of biomass (total volume of organisms). Totalvolumes of organisms were calculated from the data in tables 1 (above) and5 (p. 259) (for method see Mucibabid, 1957). The biomass of Chilomonas is

Chilotnonas and Tetrahymena
TABLE a
Growth of population of Chilomonas in single cultures and in mixedcultures with Tetrahymena patula in terms of total volume of
organisms {in thousands of cubic /x)
257
Age indays
0
2468
1 0
14
Singlepopulation
1 6 9282
18,003
193.249215,776
Mixedpopulation
16-9257
12,118175,224
123,40283,512
initially smaller in mixed than in single cultures. Later, for a short time, itsurpasses the amount in single cultures; but as soon as it reaches the maximumstationary phase, the Chilomonas population in mixed cultures begins todecrease in total volume.
Changes in composition of population of Tetrahymena patula in single andmixed cultures
Tetrahymena patula is known to be a polymorphic species (fig. 1, B). Formswith a large peristome+mouth (macrostomatous), with a small mouth(microstomatous), and with a small slit instead of an open mouth, havepreviously been observed in bacteria-free cultures, as well as in cultures withbacteria. Transitional forms between the microstomatous and macrostomatousforms have also been described. In the present work, during the first seriesof experiments, it was noticed that the composition of the Tetrahymenapopulation changes during population growth. For this reason, in the secondseries of experiments the number of each form of Tetrahymena present wasrecorded. This prolongs and increases the difficulty of counting, because onehas to wait until the organisms are in such a position that the mouth can beseen. Occasionally it was not possible to determine by observation in life towhich type an organism belonged. Transitional forms between microstomatousand macrostomatous forms were counted as macrostomatous forms.
The percentage of each form of organism from the whole population wascalculated, in order to make the results comparable. These are shown intable 3 (p. 258). The table reveals that slit-mouthed forms never appear insingle populations, while microstomatous forms are more frequent during thelogarithmic phase of population growth than at other times, both in singleand in mixed cultures. Their number in mixed populations increases on the7th day, at the time when the rate of population growth suddenly increases.
The proportion of macrostomatous forms increases during populationgrowth. Their percentage is greatest when the single cultures are in the

258 Mucibabic"—Mixed Populations of
TABLE 3
Composition of population of Tetrahymena in single and mixed cultures
2,
2
IIIItI2C
3 2
Single cultures
X
%
87-I3t86-8695-19
83-0646-35
S
s
31
4
I.I576 1
0
b 1§ 1
X
%
4411
13
5
0 311
' 4
7688
s
2-575-624 6 1
6-8o5'44
uthe
d
.11
.V
•A
&
%
2-46I-S6
o-i847-77
n
313
3
to-
lli l
X
%
?8i8J?
58
707771
H7375
740181
o i t351466688 03 2
77-O672272966
s
7-10
3 2 46 6 2
1-48
3-56
Mixed cultur
to-
icro
sU
ous
§ S
X
%
4 12 7
9
*5I I10
85341
3
z67491
4 1o i0 8
I S9 76 2
578 498
2 9 9
s
7-41I-6I1-71
1-35
es
uthe
d
I |S5-S,
/o
o- 8
69
122 0
2 02 22 0
102 2
68586 2
782 0
3S57I I
0 0
3 367-35
|
•la
*%
4
0
2 1
86
2 7
1-42
n
3133
3413133312
1
signific t highly significant
stationary phase. At the same time their percentage is also maximal in mixedpopulations; later their number decreases when the slit-mouthed forms appearin greater number. These macrostomatous forms are known to be cannibalistic(fig. 1, c). They prey on the slit-mouthed forms as well as on Chilomonas, sothat the percentage of both macrostomatous and slit-mouthed forms oscillates.In cultures a month and two months old, slit-mouthed forms are the mostnumerous; the percentage of macrostomatous forms is smaller than before,but they are large and healthy; microstomatous forms are poorly represented.
This division of forms of Tetrahymena patula into three groups is onlyapproximately correct; it does not reflect the variety of shapes in whichmacrostomatous forms may appear in the same culture. Besides the formsthat are rounded at the posterior end, as mentioned by previous authors, thereare also forms with a re-entrant posterior end (fig. 1, D), forms with longitu-dinal folds (fig. 1, E), and forms with an obtusely pointed end. The last werementioned by Maupas (1888). In cultures 8 days old or more, macrostomatousforms were observed in a striking and characteristic attitude, as if looking forfood: the body tilted with the mouth downwards, apparently examining thesubstratum, while the organism slowly advanced.
On the whole, there are no statistically significant differences between thepercentages of microstomatous or macrostomatous forms present in singleand mixed populations. The most important difference is the occurrence ofslit-mouthed forms in mixed populations only.

Chilomonas and Tetrahymena 259
Changes in size and shape of Chilomonas paramecium and Tetrahymenapatula during the growth of their single and mixed populations
Tables 4 and 5 contain data on changes in size and shape of Chilomonasparamecium and Tetrahymena patula during the growth of their single andmixed populations. These data were obtained in the same way as in previous
TABLE 4
Changes in size and shape of Chilomonas paramecium during populsyiongrowth in single and mixed cultures
Days
2468
14
Single population
L
X
219
21-7
c.v.
12-44
7-05
w
97
9-6
C.V.
5-8o
LIW
X
2-27
C.V.
8-77817
V
X
1,078
I,O46
„16
30
Mixed population
L
X
22-8199
20'I
C.v.
6
4
567270
57
W
X
8-88995
c.v.
1294
9153'45
272
LIW
X
2
51
2828
II
C.V.
1454
1263100910-75n-4910-14
V
X
1,6901,4821,2481,192
804767949
n
16332637403028
TABLE 5
Changes in size and shape of Tetrahymena patula during populationgrowth in single and mixed cultures
ma
ml
ma
ma
cro
outhed
Days
2
4
68
101314
c.v. coefficient of va
Single population
L
X
638463785972
9505759
c.v.
7'749-98
12-78
1-6715 49
W
X
34'9
37-25
39°406
c.v.
u-66
1279
9591650
LIW
X
•83
• 7 1
•54•81
c.v.
4-04
13-45
8901238
n
1 0
8
414
nation
Mixed population
L
X
63
64
81
5995
78
S4
0
8
8
Z
4
5
c.v.
7'74
5'77
1628
1870
21-29
6-O9
wX
34
4 1
44
3953
63
9
8
5
0
3
6
3 0 0
c.v.
n-66
i i - S i
I2-8S
15-61
22-47
2 7 2
LIW
X
1
I
I
I
8 3
57
85
5181
67
1-82
c.v.
4 0 4
10649 47
15-66
20-17
1114
7 6 4
n
1 0
17
32
I183
192
4
experiments on the growth of mixed populations of Chilomonas parameciumand Tetrahymena pyriformis (1957). The volume of individual organisms ofT. patula, however, cannot be calculated from the data obtained from thephotographs, because of the irregular shape of the animal, and for this reasondata for the volume of T. patula do not appear in table 5.
Length, width, and volume of individual organisms of Chilomonas, both insingle and mixed cultures, decrease during population growth. The size of

260 Mucibabid—Mixed Populations of
individual organisms increases only when the numbers of the Chilomonaspopulation in mixed cultures begin to decrease.
The ratio of length to width of Chilomonas shows a decrease after inocula-tion, in single and mixed cultures, during the lag phase. This means that theorganisms become more plump. In other phases of population growth,changes in shape of Chilomonas are not great. In mixed cultures only is therea pronounced decrease in the ratio at the end of the stationary phase, when thepopulation of Chilomonas starts decreasing.
The length of Tetrahymena patula decreases, both in single and mixedcultures, after inoculation. Later, the length increases slightly in single cul-tures, though the organisms never regain the length of freshly inoculatedorganisms. In mixed cultures, however, an increase in length is very markedon the day following the increased ingestion of Chilomonas. A further increasein length was recorded when the organisms become cannibalistic; at thebeginning of population growth they are not cannibalistic. The width oforganisms also increases in mixed cultures at the same time.
The ratio of length to width of Tetrahymena patula decreases after inocu-lation, and the organisms are plump at the beginning of the logarithmicphase. Later, however, they become slender, even more slender than freshlyinoculated organisms. Data for microstomatous forms are scarce, but suchas they are, they are shown in table 5 (p. 259) under the data for macro-stomatous forms. The lowest row (on the 14th day) comprises data for theslit-mouthed forms.
The variation in shape of Chilomonas is greater in mixed than in singlecultures, as can be seen from the coefficients of variation of the ratio length:width (coefficient of variation = relative standard deviation, that is, thestandard deviation expressed as a percentage of the mean). The variation insize and shape of Tetrahymena patula is very great in mixed cultures. At thebeginning of growth, variation is smaller than later.
Using the Mest (Fisher, 1950), the data from tables 4 and 5 have beenstatistically compared, in order to determine the significance of difference ofmeans between single and mixed cultures. Significant differences in lengthbetween Chilomonas from single and mixed cultures exist from the beginningof the logarithmic phase of growth till the end of observations. Differences inwidth of organisms from single and mixed populations, as well as differencesin the ratio of length to width, are usually not significant.
The comparison of results for Tetrahymena patula shows that differencesin width of organisms from single and mixed cultures are always significantor nearly significant. Differences in length of Tetrahymena are significantfrom the 6th day, but differences in the ratio of length to width are lesssignificant than those for length or width.
The mean maximal and minimal values for length, width, and ratio oflength to width, for members of the single populations were compared. Thiscomparison was also made for members of the mixed populations. Values oft for Chilomonas for the difference between mean maximal value at 2 days and

Chilomonas and Tetrahymena 261
minimal value at 10 days, from single populations, were: 3-167 for length;5-681 for width; and 1-756 for the ratio of length to width. Correspondingvalues for Chilomonas from mixed cultures were: for length (2 and 10 daysold), 13-022; for width (2 and 8 days old), 7-547; for the ratio length : width(L : W) (6 and 14 days old), 2-737. These values show that the differences arehighly significant, with the exception of the difference in the ratio of length towidth in organisms in single population which, however, is nearly significant;that is to say, the organisms in single culture change significantly in size duringpopulation growth, while those in mixed culture change significantly both insize and in shape. The corresponding values of t for Tetrahymena patula insingle population were: for length (2 and 6 days old), 3-40; for width (2 and6 days old), 5-70; for the ratio L : W (4 and 6 days old), 3-506. In mixedpopulations the values were: for length (4 and 14 days old), 5-276; for width(6 and 14 days old), 5-568; for the ratio L : W (2 and 8 days old), 5-416.These are all highly significant; so that both size and shape of Tetrahymenapatula change significantly during population growth, both in single and inmixed cultures.
DISCUSSION
According to the mathematical theories of Lotka (1925) and Volterra(1926), the interaction between predator and prey is characterized by cyclicalvariation in numbers of both species. In the present experiments, however,such periodic oscillations in population numbers have not appeared. Afterreaching the stationary phase, the population of the prey decreased continu-ously until it was exterminated, while the population of the predator increasedsteadily. At the beginning of population growth, the size of the Chilomonaspopulation in single and mixed cultures does not differ significantly; for ashort time, however, the population of Chilomonas is greater in mixed thanin single cultures. This is, surprisingly enough, at the time when Tetrahymenaingests Chilomonas more readily than it did before. This must mean that themultiplication rate of Chilomonas in mixed cultures is greater at that timethan it is in single cultures.
It has been shown that the composition of the population of Tetrahymenapatula is affected by the presence or absence of Chilomonas; it also changeswith the age of population. Slit-mouthed forms do not appear in singlecultures, while in mixed cultures their percentage increases in old cultures.Corliss (1953) shows slit-mouthed forms as a transitory stage betweenmicrostomatous and macrostomatous forms; but in the present experiments,slit-mouthed forms did not appear in single cultures, though both macro-stomatous and microstomatous forms were present. It was observed that theslit-mouthed forms swim quickly, and Maupas (1888) also recorded a greaterrate of swimming in slit-mouthed forms. He found them in cultures with asmall quantity of food. In the present experiments also, the slit-mouthedforms do not appear from the beginning, but only from the seventh day. Inone culture only, a single slit-mouthed form was noticed on the 4th day.

262 Mucibabid—Mixed Populations of
Maupas ascribed to them the role of the distribution of species. Faure-Fremiet (1948) observed slit-mouthed forms in cultures of Tetrahymena{'Leucophrys') patula, previously fed with Colpidium. They appeared only instarving cultures. He observed a progressive reduction in the size of themouth of microstomatous forms; indeed, in some forms the mouth becamevestigial, and such organisms were unable to ingest bacteria. He mentionsthat these forms became the prey of the macrostomatous forms; this was alsoobserved in the present experiments. Faure-Fremiet and Mugard (1949)have described macrostomatous and microstomatous forms of Espejoiamucicola. The microstomatous forms are migratory and were observed totransform themselves to macrostomatous forms as soon as they entered themucilage covering the thallus of Batrachospermum. In Espejoia the occurrenceof microstomatous and macrostomatous forms does not depend on the ageof population, as observed in the present experiments on Tetrahymena patula,but on the presence or absence of the appropriate mucilage.
It has been observed that T. patula does not prey continuously on Chilo-monas. After a day of voracious feeding, they seem to 'rest', since on thefollowing day Tetrahymena could not be seen to take Chilomonas. Volterraand Lotka's equations for the interaction of organisms are based on thesupposition that the rate of feeding is constant, and that the finding of preyby predator is a matter of chance. In old cultures, with 'clouds' of degeneratingChilomonas, Tetrahymena patula was observed to gather in these clouds. Itis questionable whether either this aggregation or the discontinuity in feedingis the result of chance.
I would like to express my gratitude to the Heads of the Botany School andof the Department of Zoology, Cambridge University, for research facilities,and to many assistants who have spared no trouble in helping me. Sincerethanks are due to Professor E. G. Pringsheim, Dr. G. Salt, Professor S.Stankovic, and Professor G. E. Briggs for stimulating discussions and foruseful suggestions in relation to my work. In particular I am indebted toDr. L. E. R. Picken for constant help and encouragement.
REFERENCESBROWN, M. G., 1940. 'Growth of protozoan cultures. II. Leucophrys patula and Glaucoma
pyrifonnis in a bacteria-free medium.' Physiol. Zool., 13, 277.CHEN, Y. T., 1950. 'Investigations of the biology of Peranema trichophorum.' Quart. J. micr..
Sci., 91, 279.CORLISS, J. O., 1952. 'Comparative studies on holotrichous ciliates in the Colpidium-Glaucoma-
Leucophrys-Tetrahymena group. I. General consideration of history of strains in pureculture.' Trans. Amer. micr. Soc, 71, 159.
'953- 'Comparative studies on holotrichous ciliates in the Colpidium-Glaucoma-Leuco-phrys-Tetrahymena group. II. Morphology, life cycles and systematic status of strains in.pure culture.' Parasitology, 43, 49.
DEWEY, V. C, and KIDDER, G. W., 1940. 'Growth studies in ciliates. VI. Diagnosis, sterili-zation and growth characteristics of Perispira ovum.' Biol. Bull., 79, 255.
FAURE-FREMIET, E., 1948. 'Doublets homopolaires et regulation morphogenetique chez lecili6 Leucophrys patula.' Arch. Anat. micr., 37, 183., et MUGARD, H., 1949. 'Le dimorphisme de Espejoia mucicola.' Hydrobiologia, 1, 379.

Chilomonas and Tetrahymena 263
FISHER, R. A., 1950. Statistical methods for research workers, pp. 114-19. Edinburgh (Oliver& Boyd).
GAUSE, G. F., 1934. 'Untersuchungen iiber den Kampf urns Dasein bei Protisten.'" Biol.Zbl., 55, 536., 1935a. 'Verifications experimentales de la theorie mathematique de la lutte pour lavie.' Actualites sci. industr., 277, ix, 62.1 !935&- 'Experimental demonstration of Volterra's periodic oscillations in the numbersof animals.' J. exp. Biol., 12, 44.
KIDDER, G. W., LILLY, D. M., and CLAAFF, C. L., 1940. 'Growth studies onciliates. IV. Theinfluence of food on the structure and growth of Glaucoma vorax sp. nov.' Biol. Bull.,78.9.
LILLY, D. M., 1942. 'Nutritional and supplementary factors in the growth of carnivorousciliates.' Physiol. Zool., 15, 146.
LOTKA, A. J., 1925. Elements of physical biology, p. 64-99. Baltimore (Williams & Wilkins).LUND, E. J., 1914a. 'The relations of Bursaria to food. I. Selection in feeding and in extrusion.'
J. exp. Zool., 16, 1., 1914b. 'The relation of Bursaria to food. II. Digestion and resorption in the foodvacuole, and further analysis of the process of extrusion.' Ibid., 17, 1.
MAUPAS, E., 1888. 'Recherches experimentales sur la multiplication des infusoires cili^s.Arch. Zool. exp. gen., 6, 165.
MUCIBABI<5, S., 1956. 'Some aspects of the growth of single and mixed populations offlagellates and ciliates. The effect of temperature on the growth of Chilo?nonas para-mecium.' J. exp. Biol., 33, 627.I957- 'The growth of mixed populations of Chilomonas paramecium and Tetrahymena
pyriformis.' J. gen. Microbiol. (in press).SANDON, H., 1932. The food of Protozoa, p. 121. Cairo (Misr-Sokkar Press).SCHAEFFER, A. A., 1917. 'Choice of food in Ameba.' J. Anim. Behavior, 7, 220.VOLTERRA, V., 1926. 'Variazioni e fiuttuazioni del numero d'individui in specie animali
conviventi.' R. Com. Talass. Ital. Mem., 131, 142.