the essential role of the intestinal microbiota in ... · the essential role of the intestinal...
TRANSCRIPT

of November 29, 2018.This information is current as
Inflammatory ResponsesMicrobiota in Facilitating Acute The Essential Role of the Intestinal
M. TeixeiraMauroVanessa Pinho, Jacques R. Nicoli, Leda Q. Vieira and
Danielle G. Souza, Angélica T. Vieira, Adriana C. Soares,
http://www.jimmunol.org/content/173/6/4137doi: 10.4049/jimmunol.173.6.4137
2004; 173:4137-4146; ;J Immunol
Referenceshttp://www.jimmunol.org/content/173/6/4137.full#ref-list-1
, 3 of which you can access for free at: cites 35 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2004 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on Novem
ber 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on N
ovember 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Essential Role of the Intestinal Microbiota in FacilitatingAcute Inflammatory Responses1
Danielle G. Souza,* Angelica T. Vieira,* Adriana C. Soares,* Vanessa Pinho,*Jacques R. Nicoli,† Leda Q. Vieira,* and Mauro M. Teixeira2*
The restoration of blood flow, i.e., reperfusion, is the treatment of choice to save viable tissue following acute ischemia of avascular territory. Nevertheless, reperfusion can be accompanied by significant inflammatory events that limit the beneficialeffects of blood flow restoration. To evaluate the potential role of the intestinal microbiota in facilitating the development oftissue injury and systemic inflammation, germ-free and conventional mice were compared in their ability to respond toischemia and reperfusion injury. In conventional mice, there was marked local (intestine) and remote (lung) edema forma-tion, neutrophil influx, hemorrhage, and production of TNF-�, KC, MIP-2, and MCP-1. Moreover, there was an increase inthe concentration of serum TNF-� and 100% lethality. In germ-free mice, there was no local, remote, or systemic inflam-matory response or lethality after intestinal ischemia and reperfusion and, in contrast to conventional mice, germ-freeanimals produced greater amounts of IL-10. Similar results were obtained after administration of LPS, i.e., little productionof TNF-� or lethality and production of IL-10 after LPS in germ-free mice. Blockade of IL-10 with Abs induced markedinflammation and lethality in germ-free mice after ischemia and reperfusion or LPS administration, demonstrating that theability of these mice to produce IL-10 was largely responsible for their “no inflammation” phenotype. This was consistentwith the prevention of reperfusion-associated injury by the exogenous administration of IL-10 to conventional mice. Thus,the lack of intestinal microbiota is accompanied by a state of active IL-10-mediated inflammatory hyporesponsiveness. TheJournal of Immunology, 2004, 173: 4137– 4146.
T he restoration of blood flow, i.e., reperfusion, is the treat-ment of choice to save viable tissue following acute isch-emia of a vascular territory. Nevertheless, reperfusion of
ischemic tissues can be accompanied by significant local, remote,and systemic inflammatory events that may limit the beneficialeffects of blood flow restoration (1–4). Thus, it is believed thatstrategies limiting the acute inflammatory response that followsreperfusion may be useful therapeutic adjuncts in the treatment ofacute ischemia (4). Factors influencing the severity of the inflam-matory response include the vascular territory, the duration of isch-emia, the duration of reperfusion, and the presence of collateralblood flow (1–4). Several mediators of the inflammatory processhave been shown to participate in the cascade of events leading toischemia and reperfusion (I/R)3 injury, including CXC chemo-kines, platelet-activating factor and cytokines, especially TNF-�and IL-10 (5–12). Treatment of animals with anti-TNF-� Abs orexperiments in TNF-� receptor 1-deficient animals (TNFR1�/�)
clearly show a central role of TNF-� in mediating tissue injury,systemic inflammation, and lethality following intestinal reperfu-sion injury (6, 7, 13). As TNF-� release and action is central in thepathogenesis of the local and systemic inflammatory response thatoccurs after intestinal reperfusion, a greater understanding of themediators that induce or modulate TNF-� production in this pro-cess is clearly of interest.
A few studies in rats have suggested that bacterial translocationfollowing disruption of intestinal epithelial lining would accountfor the increase in TNF-� production and tissue injury that occursafter intestinal I/R (14, 15). Of note, most of these studies usedshorter ischemia and prolonged reperfusion times that could allowgreater loss of the ability of epithelial cells to maintain their barrierfunction. In contrast, other studies using shorter ischemia timeshave failed to demonstrate an important role for LPS or the LPSreceptor TLR4 during I/R injury (16, 17). Thus, an aim of thepresent study was to investigate whether bacterial and/or LPStranslocation occurred and underlined the local and systemicinflammatory injury following prolonged intestinal I/R in mice.To this end the concentrations of LPS in blood and blood andliver bacterial load after intestinal I/R injury were quantified.Moreover, experiments evaluating the inflammatory injury afterI/R were also conducted in germ-free mice, which have no bac-teria (and indeed, no other known pathogen) in their gut. In thelatter animals, we wished to evaluate the relevance of the in-testinal microbiota to acute inflammation, as measured in amodel of I/R injury. As germ-free mice were shown not toinflame or die after I/R injury, additional experiments were con-ducted to evaluate a role for IL-10 in mediating the lack ofinflammatory responsiveness of these animals. For comparison,responses of germ-free and conventional mice to LPS and therole of IL-10 were also investigated.
Departaments of *Bioquımica e Imunologia e †Microbiologia, Instituto de CienciasBiologicas, Universidade Federal de Minas Gerais (UFMG), Pampulha, Belo Hori-zonte, Brazil
Received for publication May 13, 2003. Accepted for publication June 30, 2004.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by Fundacao do Amparo a Pesquisa do Estado de MinasGerais (FAPEMIG), Conselho Nacional de Desenvolvimento Cientıfico e Tecno-logico (CNPq), and Pro-Reitoria de Pesquisa da UFMG (PRPq/UFMG).2 Address correspondence and reprint requests to Dr. Mauro M. Teixeira, Imuno-farmacologia, Departamento de Bioquımica e Imunologia, Instituto de CienciasBiologicas, Universidade Federal de Minas Gerais, Avenuda Antonio Carlos,6627-Pampulha, 31270-901, Belo Horizonte, MG, Brasil. E-mail address:[email protected] Abbreviations used in this paper: I/R, ischemia and reperfusion; SMA, superiormesenteric artery.
The Journal of Immunology
Copyright © 2004 by The American Association of Immunologists, Inc. 0022-1767/04/$02.00
by guest on Novem
ber 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Materials and MethodsAnimals
Germ-free Swiss/NIH mice were derived from a germ-free nucleus (Tac-onic Farms, Germantown, NY) and maintained in flexible plastic isolators(Standard Safety Equipment, Palatine, IL) using classical gnotobiologytechniques (18). Conventional Swiss/NIH mice are derived from germ-freematrices, and considered conventional only after two generations in theconventional facility. All animals were 8- to 10-wk-old males and females.All experimental procedures in germ-free mice were conducted under asep-tic conditions to avoid infection of animals and had prior approval from thelocal animal ethics committee.
Ischemia and reperfusion
Mice were anesthetized with urethane (140 mg/kg, i.p.) and laparotomy wasperformed. The superior mesenteric artery (SMA) was isolated and ischemiawas induced by totally occluding the SMA for 60 min. For measuring per-centage of surviving mice, reperfusion was re-established, and mice were mon-itored for indicated time periods. For the other parameters, reperfusion wasallowed to occur for 40 min (I60R40) when mice were sacrificed. This time ofreperfusion (40 min) was chosen based on the presence of significant tissueinjury without unduly high mortality rates. Sham-operated animals were usedas controls. In some experiments, recombinant murine IL-10 (PeproTech,Rocky Hill, NJ) or vehicle (PBS, 100 �l/mouse) were administered s.c. at thedose of 0.5 �g/animal 45 min before reperfusion. In other experiments, anti-IL-10 polyclonal (rabbit anti-rat/murine-IL-10, 1 �l/g) or monoclonal (SCX-2clone, 100 �g/mouse) Abs were administered s.c. 45 min before reperfusion.As controls for the latter experiments, mice received preimmune rabbit serum(1 �l/g) or purified rat IgM (100 �g/mouse, clone R4-22; BD Pharmingen, SanDiego, CA).
Macrophage cultures
Resident macrophages were obtained from the peritoneal cavity of na-ive germ-free or conventional mice and were cultured in DMEM sup-plemented with 10% FBS. The cells obtained from peritoneal washeswere seeded in 96-well plates at 2 � 106 cells/ml. After 3 h of incu-bation at 37°C and 5% CO2, nonadherent cells were washed off and theadherent macrophages were incubated with or without LPS (100 ng/ml,from Escherichia coli serotype 0111:B4; Sigma-Aldrich, St. Louis,MO). After 48 h, the supernatant was removed and stored at �20°Cuntil further analysis. The concentration of TNF-� and IL-10 was mea-sured in supernatants using commercially available Abs and accordingto the procedures supplied by the manufacturer (R&D Systems,Minneapolis, MN).
Conventionalization of germ-free mice
The process of colonizing germ-free mice with microbiota from con-ventional mice is a process referred to as conventionalization and thiswas performed as previously described (18). Briefly, fecal samples re-moved from the large intestine of conventional mice were homogenizedin saline (10%) and administered by oral gavage to germ-free mice.Seven, 14, or 21 days later, intestinal I/R was conducted in these ani-mals, as described above. To assess whether there was adequate con-ventionalization of germ-free mice, fecal samples were cultured using athioglycolate test (18).
Antibiotic treatment
A combination of large spectrum antibiotics was used to sterilize the intestinalmicrobiota of conventional mice. To this end a combination of metronidazole,ciprofloxacin, vancomycin, and imipenem (all at 50 mg/kg body weight/day)was administered in the drinking water. Animal beds were changed daily andall experimental procedures were conducted under aseptic conditions to avoidreinfection of animals. To assess whether the antibiotic treatment was adequateto deplete the intestinal microbiota, fecal samples were cultured using a thio-glycolate test after 7, 14, and 21 days of antibiotic treatment (18). On day 21,the animals were submitted to intestinal I/R injury.
Measurement of bacteria and LPS after I/R
Blood was collected from the brachial plexus and the liver was harvestedand grinded in a vented hood. The homogenates and blood samples were
placed on ice, and serial 1/10 dilutions were made. One hundred microlitersof each dilution were plated on McConkey agar plates (Difco, Detroit, MI)and incubated for 24 h at 37°C and then the number of CFU was counted.The detection limit of the assay was 100 bacteria ml�1 or 100 bacteria per100 mg of tissue. LPS in plasma samples was determined using the Limulusamebocyte assay performed by Dr. S. Poole (National Institute of Biolog-ical Standard and Control, Potter Bar, U.K.).
LPS-induced lethality
LPS (1 and 10 mg/kg, from E. coli serotype 0111:B4; Sigma-Aldrich) wasadministered i.p. to conventional or germ-free mice. In these animals, lethalitywas observed at various times after injection or serum was obtained for TNF-�and IL-10 measurements. In some experiments, control Ab (rat IgM) or anti-IL-10 monoclonal (SCX-2 clone, 100 �g/mouse) Abs were administered s.c.60 min before the administration of LPS (10 mg/kg).
Evaluation of changes in vascular permeability
The extravasation of Evans blue dye into the tissue was used as an index ofincreased vascular permeability, as previously described (19, 20). Briefly,Evans blue (20 mg/kg) was administered i.v. (1 ml/kg) via a tail vein 2 minbefore reperfusion of the ischemic artery. Forty minutes after reperfusion, asegment of the duodenum (�3 cm) or the flushed left lung were cut in smallpieces and Evans blue extracted using 1 ml of formamide. The amount ofEvans blue in the tissue (micrograms of Evans Blue per 100 mg of tissue) wasobtained by comparing the extracted absorbance with that of a standard Evansblue curve read at 620 nm in an ELISA plate reader.
Myeloperoxidase concentrations
The extent of neutrophil accumulation in the intestine and right lung tissuewas measured by assaying myeloperoxidase activity, as previously de-scribed (7, 21). Briefly, a portion of duodenum and the flushed right lungsof animals that had undergone I/R injury were removed and snap frozen inliquid nitrogen. Upon thawing and processing, the tissue was assayed formyeloperoxidase activity by measuring the change in OD at 450 nm usingtetramethylbenzidine. Results were expressed as total number of neutro-phils by comparing the OD of tissue supernatant with the OD of casein-elicited murine peritoneal neutrophils processed in the same way.
Measurement of hemoglobin concentrations
The determination of hemoglobin concentrations in tissue was used as anindex of tissue hemorrhage. After washing and perfusing the intestines toremove excess blood in the intravascular space, a sample of �100 mg ofduodenum was removed and homogenized in Drabkin’s color reagent ac-cording to instructions of the manufacturer (Analisa, Belo Horizonte, Bra-zil). The suspension was centrifuged for 15 min at 3000 � g and filteredusing 0.2-�m filters. The resulting solution was read using an ELISA platereader at 520 nm and compared against a standard curve of hemoglobin.
Measurement of cytokine/chemokine concentrations in serum,intestine, and lungs
Serum was obtained from coagulated blood (15 min at 37°C, then 30 mina 4°C) and stored at �20°C until further analysis. Serum samples wereanalyzed at a 1/3 dilution in PBS. One hundred milligrams of duodenum orlung of sham-operated and reperfused animals were homogenized in 1 mlof PBS (0.4 M NaCl and 10 mM NaPO4) containing anti-proteases (0.1mM PMSF, 0.1 mM benzethonium chloride, 10 mM EDTA, and 20 Kal-likrein inhibitor units of aprotinin A) and 0.05% Tween 20. The sampleswere then centrifuged of 10 min at 3000 � g and the supernatant wasimmediately used for ELISA at a 1/3 dilution in PBS. The concentration ofTNF-�, KC, MIP-2, MCP-1, and IL-10 was measured in serum and tissueof animals using commercially available Abs and according to the proce-dures supplied by the manufacturer (R&D Systems).
Statistical analysis
Results are shown as means � SEM. Percent inhibition was calculated bysubtracting the background values obtained in sham-operated animals. Dif-ferences were compared by using ANOVA followed by Student-Newman-Keuls posthoc analysis. Results with a p � 0.05 were considered signifi-cantly different.
4138 INFLAMMATION AND THE INTESTINAL MICROBIOTA
by guest on Novem
ber 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

ResultsNo evidence of systemic bacterial translocation after I/RTo evaluate the possible role of the translocation of bacteria and/orbacterial products as a means of triggering or potentiating reperfusion-
associated systemic inflammatory response, blood and liver were cul-tured and evaluated for the presence of LPS. There were no detectableCFUs or measurable amounts of LPS in blood and liver tissue of micefollowing reperfusion of the ischemic SMA (data not shown).
FIGURE 1. Reperfusion-associated increase in neutro-phil influx, vascular permeability, and hemorrhage in theintestine and lungs of conventional (CV) and germ-free(GF) mice. Neutrophil accumulation (A and B) was as-sessed by measuring the tissue contents of myeloperoxi-dase, changes in vascular permeability (C and D) by theextravasation of Evans blue dye, and hemorrhage (E) by theintestinal levels of hemoglobin. In CV mice, recombinantmurine IL-10 (shown as IL-10) was administered s.c. at thedose of 0.5 �g/animal 45 min before reperfusion. Controlanimals (shown as IR) received IL-10 vehicle (PBS, 100�l). Results are shown as number of neutrophils, micro-grams of Evans blue dye, or micrograms of hemoglobin per100 mg of tissue and are the mean � SEM of at least sixanimals in each group. �, p � 0.01 when compared withsham-operated animals; #, p � 0.01 when compared withvehicle-treated animals submitted to I/R.
FIGURE 2. Reperfusion-associatedincrease in the concentrations of TNF-�and IL-10 in the intestine and lungs ofconventional (CV) and germ-free(GF) mice. TNF-� (A and B) andIL-10 (C and D) were measured usingspecific ELISA. In CV mice, recom-binant murine (shown as IL-10) wasadministered s.c. at the dose of 0.5�g/animal 45 min before reperfusion.Control animals (shown as IR) re-ceived IL-10 vehicle (PBS, 100 �l).Results are shown as picograms of thecytokine per 100 mg of tissue and arethe mean � SEM of at least six ani-mals in each group. �, p � 0.01 whencompared with sham-operated ani-mals; #, p � 0.01 when comparedwith vehicle-treated animals submit-ted to I/R.
4139The Journal of Immunology
by guest on Novem
ber 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Lack of tissue injury and lethality following I/R in germ-free mice
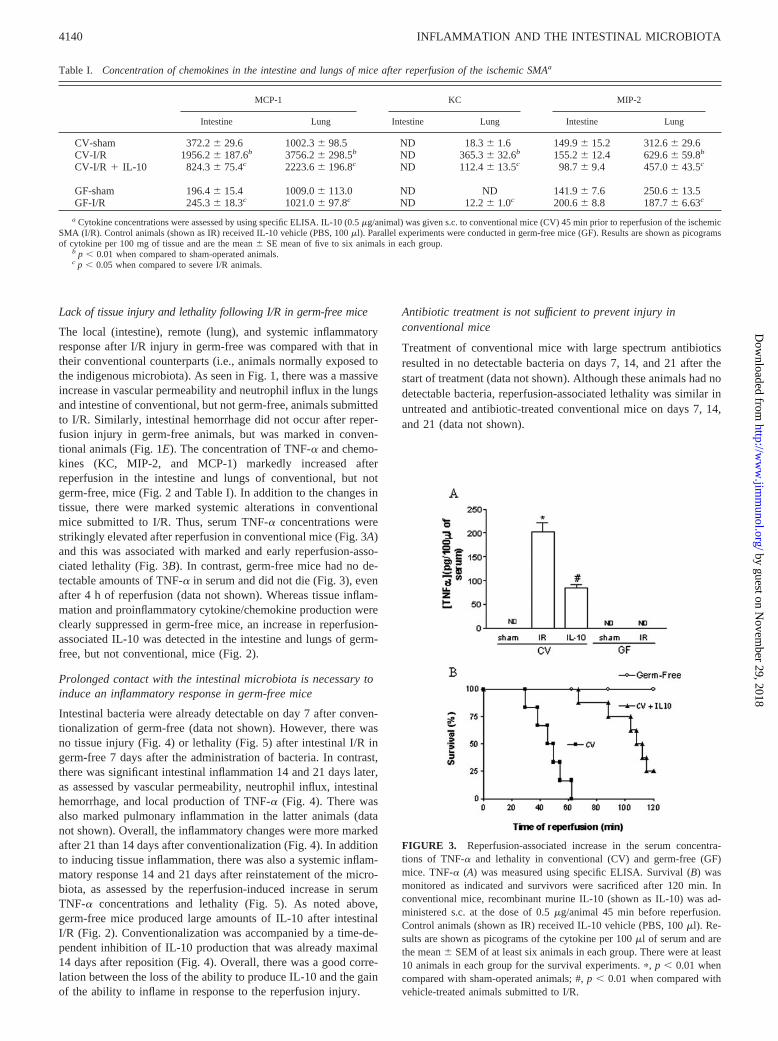
The local (intestine), remote (lung), and systemic inflammatoryresponse after I/R injury in germ-free was compared with that intheir conventional counterparts (i.e., animals normally exposed tothe indigenous microbiota). As seen in Fig. 1, there was a massiveincrease in vascular permeability and neutrophil influx in the lungsand intestine of conventional, but not germ-free, animals submittedto I/R. Similarly, intestinal hemorrhage did not occur after reper-fusion injury in germ-free animals, but was marked in conven-tional animals (Fig. 1E). The concentration of TNF-� and chemo-kines (KC, MIP-2, and MCP-1) markedly increased afterreperfusion in the intestine and lungs of conventional, but notgerm-free, mice (Fig. 2 and Table I). In addition to the changes intissue, there were marked systemic alterations in conventionalmice submitted to I/R. Thus, serum TNF-� concentrations werestrikingly elevated after reperfusion in conventional mice (Fig. 3A)and this was associated with marked and early reperfusion-asso-ciated lethality (Fig. 3B). In contrast, germ-free mice had no de-tectable amounts of TNF-� in serum and did not die (Fig. 3), evenafter 4 h of reperfusion (data not shown). Whereas tissue inflam-mation and proinflammatory cytokine/chemokine production wereclearly suppressed in germ-free mice, an increase in reperfusion-associated IL-10 was detected in the intestine and lungs of germ-free, but not conventional, mice (Fig. 2).
Prolonged contact with the intestinal microbiota is necessary toinduce an inflammatory response in germ-free mice
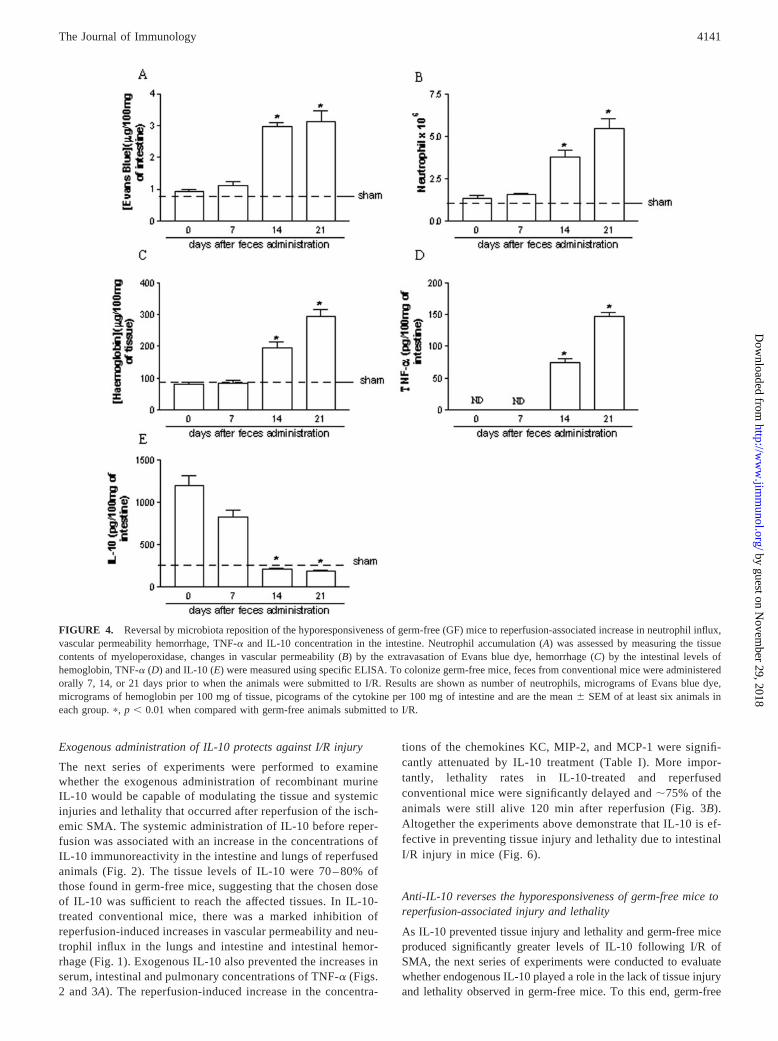
Intestinal bacteria were already detectable on day 7 after conven-tionalization of germ-free (data not shown). However, there wasno tissue injury (Fig. 4) or lethality (Fig. 5) after intestinal I/R ingerm-free 7 days after the administration of bacteria. In contrast,there was significant intestinal inflammation 14 and 21 days later,as assessed by vascular permeability, neutrophil influx, intestinalhemorrhage, and local production of TNF-� (Fig. 4). There wasalso marked pulmonary inflammation in the latter animals (datanot shown). Overall, the inflammatory changes were more markedafter 21 than 14 days after conventionalization (Fig. 4). In additionto inducing tissue inflammation, there was also a systemic inflam-matory response 14 and 21 days after reinstatement of the micro-biota, as assessed by the reperfusion-induced increase in serumTNF-� concentrations and lethality (Fig. 5). As noted above,germ-free mice produced large amounts of IL-10 after intestinalI/R (Fig. 2). Conventionalization was accompanied by a time-de-pendent inhibition of IL-10 production that was already maximal14 days after reposition (Fig. 4). Overall, there was a good corre-lation between the loss of the ability to produce IL-10 and the gainof the ability to inflame in response to the reperfusion injury.
Antibiotic treatment is not sufficient to prevent injury inconventional mice
Treatment of conventional mice with large spectrum antibioticsresulted in no detectable bacteria on days 7, 14, and 21 after thestart of treatment (data not shown). Although these animals had nodetectable bacteria, reperfusion-associated lethality was similar inuntreated and antibiotic-treated conventional mice on days 7, 14,and 21 (data not shown).
FIGURE 3. Reperfusion-associated increase in the serum concentra-tions of TNF-� and lethality in conventional (CV) and germ-free (GF)mice. TNF-� (A) was measured using specific ELISA. Survival (B) wasmonitored as indicated and survivors were sacrificed after 120 min. Inconventional mice, recombinant murine IL-10 (shown as IL-10) was ad-ministered s.c. at the dose of 0.5 �g/animal 45 min before reperfusion.Control animals (shown as IR) received IL-10 vehicle (PBS, 100 �l). Re-sults are shown as picograms of the cytokine per 100 �l of serum and arethe mean � SEM of at least six animals in each group. There were at least10 animals in each group for the survival experiments. �, p � 0.01 whencompared with sham-operated animals; #, p � 0.01 when compared withvehicle-treated animals submitted to I/R.
Table I. Concentration of chemokines in the intestine and lungs of mice after reperfusion of the ischemic SMAa
MCP-1 KC MIP-2
Intestine Lung Intestine Lung Intestine Lung
CV-sham 372.2 � 29.6 1002.3 � 98.5 ND 18.3 � 1.6 149.9 � 15.2 312.6 � 29.6CV-I/R 1956.2 � 187.6b 3756.2 � 298.5b ND 365.3 � 32.6b 155.2 � 12.4 629.6 � 59.8b
CV-I/R � IL-10 824.3 � 75.4c 2223.6 � 196.8c ND 112.4 � 13.5c 98.7 � 9.4 457.0 � 43.5c
GF-sham 196.4 � 15.4 1009.0 � 113.0 ND ND 141.9 � 7.6 250.6 � 13.5GF-I/R 245.3 � 18.3c 1021.0 � 97.8c ND 12.2 � 1.0c 200.6 � 8.8 187.7 � 6.63c
a Cytokine concentrations were assessed by using specific ELISA. IL-10 (0.5 �g/animal) was given s.c. to conventional mice (CV) 45 min prior to reperfusion of the ischemicSMA (I/R). Control animals (shown as IR) received IL-10 vehicle (PBS, 100 �l). Parallel experiments were conducted in germ-free mice (GF). Results are shown as picogramsof cytokine per 100 mg of tissue and are the mean � SE mean of five to six animals in each group.
b p � 0.01 when compared to sham-operated animals.c p � 0.05 when compared to severe I/R animals.
4140 INFLAMMATION AND THE INTESTINAL MICROBIOTA
by guest on Novem
ber 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Exogenous administration of IL-10 protects against I/R injury
The next series of experiments were performed to examinewhether the exogenous administration of recombinant murineIL-10 would be capable of modulating the tissue and systemicinjuries and lethality that occurred after reperfusion of the isch-emic SMA. The systemic administration of IL-10 before reper-fusion was associated with an increase in the concentrations ofIL-10 immunoreactivity in the intestine and lungs of reperfusedanimals (Fig. 2). The tissue levels of IL-10 were 70 – 80% ofthose found in germ-free mice, suggesting that the chosen doseof IL-10 was sufficient to reach the affected tissues. In IL-10-treated conventional mice, there was a marked inhibition ofreperfusion-induced increases in vascular permeability and neu-trophil influx in the lungs and intestine and intestinal hemor-rhage (Fig. 1). Exogenous IL-10 also prevented the increases inserum, intestinal and pulmonary concentrations of TNF-� (Figs.2 and 3A). The reperfusion-induced increase in the concentra-
tions of the chemokines KC, MIP-2, and MCP-1 were signifi-cantly attenuated by IL-10 treatment (Table I). More impor-tantly, lethality rates in IL-10-treated and reperfusedconventional mice were significantly delayed and �75% of theanimals were still alive 120 min after reperfusion (Fig. 3B).Altogether the experiments above demonstrate that IL-10 is ef-fective in preventing tissue injury and lethality due to intestinalI/R injury in mice (Fig. 6).
Anti-IL-10 reverses the hyporesponsiveness of germ-free mice toreperfusion-associated injury and lethality
As IL-10 prevented tissue injury and lethality and germ-free miceproduced significantly greater levels of IL-10 following I/R ofSMA, the next series of experiments were conducted to evaluatewhether endogenous IL-10 played a role in the lack of tissue injuryand lethality observed in germ-free mice. To this end, germ-free
FIGURE 4. Reversal by microbiota reposition of the hyporesponsiveness of germ-free (GF) mice to reperfusion-associated increase in neutrophil influx,vascular permeability hemorrhage, TNF-� and IL-10 concentration in the intestine. Neutrophil accumulation (A) was assessed by measuring the tissuecontents of myeloperoxidase, changes in vascular permeability (B) by the extravasation of Evans blue dye, hemorrhage (C) by the intestinal levels ofhemoglobin, TNF-� (D) and IL-10 (E) were measured using specific ELISA. To colonize germ-free mice, feces from conventional mice were administeredorally 7, 14, or 21 days prior to when the animals were submitted to I/R. Results are shown as number of neutrophils, micrograms of Evans blue dye,micrograms of hemoglobin per 100 mg of tissue, picograms of the cytokine per 100 mg of intestine and are the mean � SEM of at least six animals ineach group. �, p � 0.01 when compared with germ-free animals submitted to I/R.
4141The Journal of Immunology
by guest on Novem
ber 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
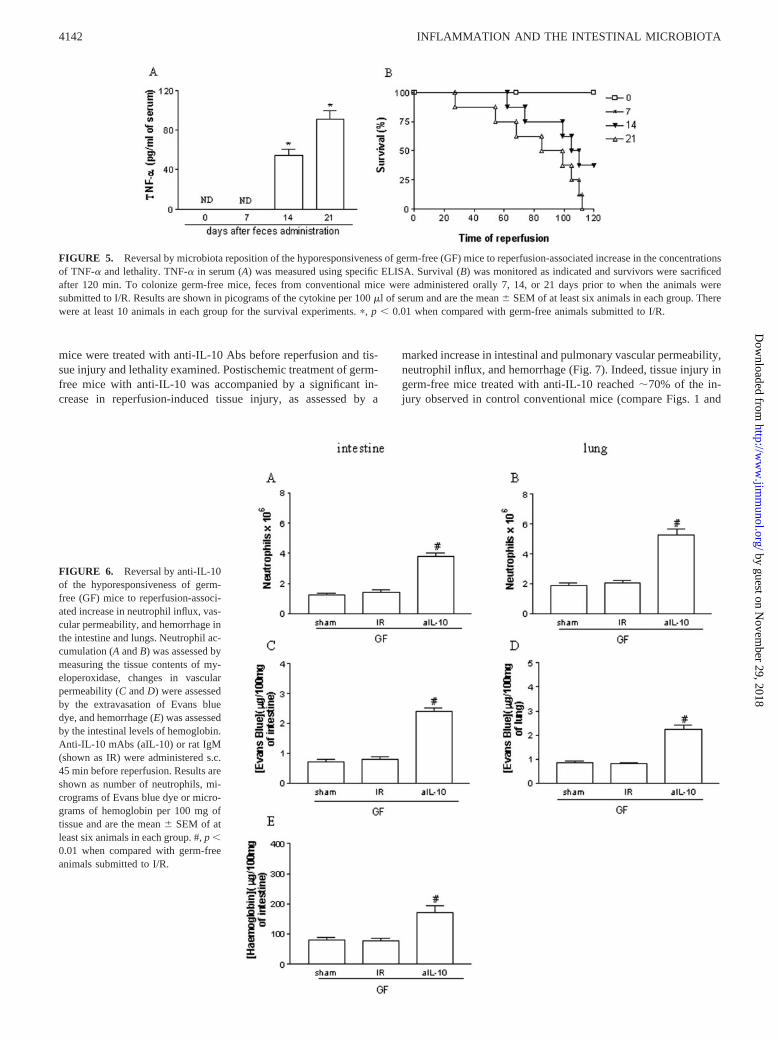
mice were treated with anti-IL-10 Abs before reperfusion and tis-sue injury and lethality examined. Postischemic treatment of germ-free mice with anti-IL-10 was accompanied by a significant in-crease in reperfusion-induced tissue injury, as assessed by a
marked increase in intestinal and pulmonary vascular permeability,neutrophil influx, and hemorrhage (Fig. 7). Indeed, tissue injury ingerm-free mice treated with anti-IL-10 reached �70% of the in-jury observed in control conventional mice (compare Figs. 1 and
FIGURE 6. Reversal by anti-IL-10of the hyporesponsiveness of germ-free (GF) mice to reperfusion-associ-ated increase in neutrophil influx, vas-cular permeability, and hemorrhage inthe intestine and lungs. Neutrophil ac-cumulation (A and B) was assessed bymeasuring the tissue contents of my-eloperoxidase, changes in vascularpermeability (C and D) were assessedby the extravasation of Evans bluedye, and hemorrhage (E) was assessedby the intestinal levels of hemoglobin.Anti-IL-10 mAbs (aIL-10) or rat IgM(shown as IR) were administered s.c.45 min before reperfusion. Results areshown as number of neutrophils, mi-crograms of Evans blue dye or micro-grams of hemoglobin per 100 mg oftissue and are the mean � SEM of atleast six animals in each group. #, p �0.01 when compared with germ-freeanimals submitted to I/R.
FIGURE 5. Reversal by microbiota reposition of the hyporesponsiveness of germ-free (GF) mice to reperfusion-associated increase in the concentrationsof TNF-� and lethality. TNF-� in serum (A) was measured using specific ELISA. Survival (B) was monitored as indicated and survivors were sacrificedafter 120 min. To colonize germ-free mice, feces from conventional mice were administered orally 7, 14, or 21 days prior to when the animals weresubmitted to I/R. Results are shown in picograms of the cytokine per 100 �l of serum and are the mean � SEM of at least six animals in each group. Therewere at least 10 animals in each group for the survival experiments. �, p � 0.01 when compared with germ-free animals submitted to I/R.
4142 INFLAMMATION AND THE INTESTINAL MICROBIOTA
by guest on Novem
ber 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

7). The concentrations of TNF-� in tissue and serum of anti-IL-10-treated germ-free mice is shown in Fig. 7. Whereas untreatedgerm-free mice have undetectable concentrations of TNF-� in tis-sues and serum after I/R injury, there was a marked and significantincrease in TNF-� in animals treated with anti-IL-10 (Fig. 7A).Similarly, whereas germ-free mice did not die after I/R injury, treat-ment with anti-IL-10 was accompanied by significant reperfusion-induced lethality, which was similar to that seen in conventional mice(Fig. 7B). Virtually identical results were obtained when germ-freemice were administered an anti-IL-10 polyclonal Ab (data notshown). Overall, our results argue that the lack of reperfusion-inducedtissue inflammation observed in germ-free mice is largely due to theirinnate ability to produce IL-10 and consequent IL-10-mediated inhi-bition of the systemic inflammatory response.
The hyporesponsiveness of germ-free mice to LPS is alsomediated by IL-10
The IL-10-dependent inability of germ-free mice to respond to such apotent inflammatory stimulus, as that triggered by the reperfusion ofan ischemic vascular, was a striking finding. It was, thus, of interest toexamine whether the lack of inflammatory responsiveness of germ-
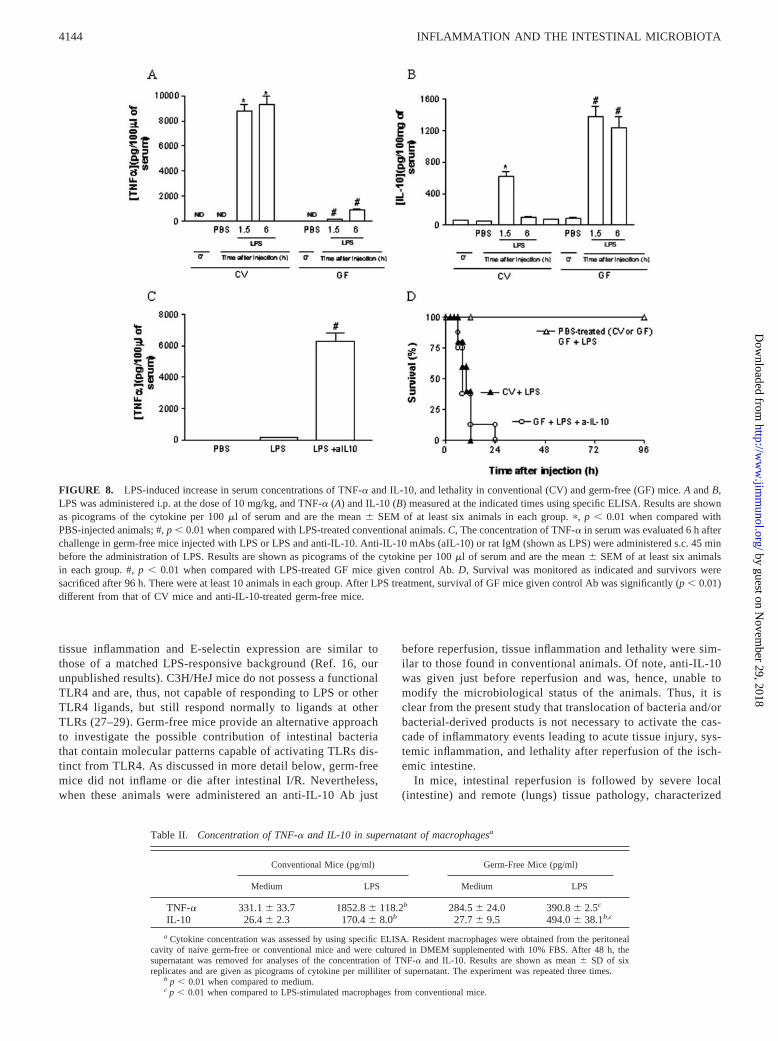
free mice would be also applicable to other stimuli capable of elicitinga systemic inflammatory response, such as those derived from mi-crobes. To this end, germ-free or conventional mice were injected i.p.with LPS (10 mg/kg) and lethality rates and cytokine (TNF-� andIL-10) concentrations examined at different time points. Concentra-tions of TNF-� rose rapidly (within 1.5 h) in the serum of conven-tional mice and were still elevated by 6 h, whereas only small amountsof TNF-� (�10% of the concentrations found in conventional mice)were detected in the serum of GF mice after systemic administrationof LPS (Fig. 8A). In contrast, concentrations of IL-10 were muchgreater and more persistent in serum of germ-free than conventionalmice (Fig. 8B). Indeed, measurable amounts of IL-10 were detectedeven 24 h after injection of LPS in germ-free (112 � 8 pg/100 mg oftissue) but not conventional (below detection limit of the assay) mice.All conventional mice injected with LPS were dead by 12 h afterinjection, whereas none of the LPS-injected germ-free or PBS-in-jected mice were dead by 96 h after LPS injection (Fig. 8C). In fact,there was no lethality even 10 days after LPS injection (data notshown).
To examine whether the endogenous production of IL-10 in re-sponse to LPS treatment in germ-free mice would also account forthe lack of responsiveness of these animals to LPS, experimentswere performed in the presence of an anti-IL-10 Ab. Similarly toour findings in the I/R injury model, germ-free treated with anti-IL-10 became very responsive to LPS administration and producedlarge amounts of TNF-� and died in a similar manner to conven-tional mice (Fig. 8, C and D).
Macrophages from germ-free mice preferentially produce IL-10
The stimulation of resident peritoneal macrophages obtained fromgerm-free or conventional mice with LPS induced a distinct profileof cytokine production (Table II). There was no difference in basalproduction of cytokines. However, whereas LPS-stimulated mac-rophages derived from conventional mice produced large amountsof TNF-� and little IL-10, macrophages from germ-free mice pro-duced large amounts of IL-10 and little TNF-� (Table II).
DiscussionThe restoration of blood flow of an ischemic vascular bed is thetreatment of choice to prevent the death of tissue cells during acuteischemic events. Nevertheless, reperfusion itself can induce a localinflammatory response that limits the benefits of blood flow res-toration. The present study was conducted to investigate the rele-vance of bacterial translocation and of the intestinal microbiota forthe inflammatory response and tissue injury that occurs afterintestinal I/R.
Inflammatory or ischemic injury to the intestine may be ofsufficient intensity to cause disruption of the intestinal epitheliallining and consequent loss of the barrier function of epithelialcells. This may occur, for example, after hypovolemic shockand facilitate the translocation of bacteria and/or bacterial prod-ucts, such as LPS, with ensuing activation of macrophages andinfection of the organism (22, 23). Whereas some investigatorshave found elevated concentrations of LPS in systemic or portalsystem blood following reperfusion of an ischemic vascular bed(14, 17, 24, 25), others, including the present study, have notbeen able to detect either LPS or significant levels of circulatingbacteria (26). The conflict between these studies is largely dueto the different durations of I/R, in such way that LPS/bacteriaare more likely to be detected after prolonged reperfusion times.However, it is still possible that sufficient amounts of bacteriaand/or LPS reach and activate inflammatory and endothelialcells within the intestinal mucosa (16). In this respect, studies inLPS-hyporesponsive C3H/HeJ mice have demonstrated that
FIGURE 7. Reversal by anti-IL-10 of the hyporesponsiveness of germ-free (GF) mice to reperfusion-associated increase in the concentrations ofTNF-� and lethality. TNF-� in serum (A) was measured using specificELISA. Survival (B) was monitored as indicated and survivors were sac-rificed after 120 min. Anti-IL-10 mAbs (a-10) or rat IgM (shown as IR)were administered s.c. 45 min before reperfusion. Results are shown aspicograms of the cytokine per 100 mg of tissue or 100 �l of serum and arethe mean � SEM of at least six animals in each group. In B, the survivalcurves of conventional mice (CV) are also shown for comparison and therewere at least 10 animals in each group. #, p � 0.01 when compared withgerm-free animals submitted to I/R.
4143The Journal of Immunology
by guest on Novem
ber 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
tissue inflammation and E-selectin expression are similar tothose of a matched LPS-responsive background (Ref. 16, ourunpublished results). C3H/HeJ mice do not possess a functionalTLR4 and are, thus, not capable of responding to LPS or otherTLR4 ligands, but still respond normally to ligands at otherTLRs (27–29). Germ-free mice provide an alternative approachto investigate the possible contribution of intestinal bacteriathat contain molecular patterns capable of activating TLRs dis-tinct from TLR4. As discussed in more detail below, germ-freemice did not inflame or die after intestinal I/R. Nevertheless,when these animals were administered an anti-IL-10 Ab just
before reperfusion, tissue inflammation and lethality were sim-ilar to those found in conventional animals. Of note, anti-IL-10was given just before reperfusion and was, hence, unable tomodify the microbiological status of the animals. Thus, it isclear from the present study that translocation of bacteria and/orbacterial-derived products is not necessary to activate the cas-cade of inflammatory events leading to acute tissue injury, sys-temic inflammation, and lethality after reperfusion of the isch-emic intestine.
In mice, intestinal reperfusion is followed by severe local(intestine) and remote (lungs) tissue pathology, characterized
FIGURE 8. LPS-induced increase in serum concentrations of TNF-� and IL-10, and lethality in conventional (CV) and germ-free (GF) mice. A and B,LPS was administered i.p. at the dose of 10 mg/kg, and TNF-� (A) and IL-10 (B) measured at the indicated times using specific ELISA. Results are shownas picograms of the cytokine per 100 �l of serum and are the mean � SEM of at least six animals in each group. �, p � 0.01 when compared withPBS-injected animals; #, p � 0.01 when compared with LPS-treated conventional animals. C, The concentration of TNF-� in serum was evaluated 6 h afterchallenge in germ-free mice injected with LPS or LPS and anti-IL-10. Anti-IL-10 mAbs (aIL-10) or rat IgM (shown as LPS) were administered s.c. 45 minbefore the administration of LPS. Results are shown as picograms of the cytokine per 100 �l of serum and are the mean � SEM of at least six animalsin each group. #, p � 0.01 when compared with LPS-treated GF mice given control Ab. D, Survival was monitored as indicated and survivors weresacrificed after 96 h. There were at least 10 animals in each group. After LPS treatment, survival of GF mice given control Ab was significantly (p � 0.01)different from that of CV mice and anti-IL-10-treated germ-free mice.
Table II. Concentration of TNF-� and IL-10 in supernatant of macrophagesa
Conventional Mice (pg/ml) Germ-Free Mice (pg/ml)
Medium LPS Medium LPS
TNF-� 331.1 � 33.7 1852.8 � 118.2b 284.5 � 24.0 390.8 � 2.5c
IL-10 26.4 � 2.3 170.4 � 8.0b 27.7 � 9.5 494.0 � 38.1b,c
a Cytokine concentration was assessed by using specific ELISA. Resident macrophages were obtained from the peritonealcavity of naive germ-free or conventional mice and were cultured in DMEM supplemented with 10% FBS. After 48 h, thesupernatant was removed for analyses of the concentration of TNF-� and IL-10. Results are shown as mean � SD of sixreplicates and are given as picograms of cytokine per milliliter of supernatant. The experiment was repeated three times.
b p � 0.01 when compared to medium.c p � 0.01 when compared to LPS-stimulated macrophages from conventional mice.
4144 INFLAMMATION AND THE INTESTINAL MICROBIOTA
by guest on Novem
ber 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

by marked neutrophil influx, edema formation, hemorrhage, andtissue destruction (shown here and in Ref. 7). Not only is theretissue damage, but also marked systemic inflammation, as as-sessed by the elevation in the serum concentration of proin-flammatory cytokines and chemokines (7). In this model,TNF-� appears to play a major pathophysiological role, as dem-onstrated by the protective effects of the administration of sol-uble TNF-� receptors or experiments in p55�/� mice (7). Sim-ilarly, other studies have clearly established a role for TNF-� inmediating both local and systemic inflammation after I/R injuryin several vascular beds (e.g., Refs. 13, 30, 31). In contrast tothese findings and to the lack of a role of the translocation ofbacteria/bacterial products, germ-free mice presented little ev-idence of local or systemic injury after intestinal I/R. Indeed,intestinal or pulmonary injury and concentrations of proinflam-matory cytokines and chemokines in germ-free mice afterreperfusion were virtually identical to those found in sham-operated mice (which were not submitted to I/R). Furthermore,there was no lethality even after 240 min of reperfusion,whereas all conventional mice were dead by 90 min. Despite theabsence of local or systemic inflammation, germ-free micewere actively sensing the inflammatory stimulation, as demon-strated by the marked increases in reperfusion-induced IL-10production.
Several studies have now reported on the ability of endogenousor exogenous IL-10 in limiting injury secondary to I/R to variousvascular beds (e.g., Refs. 12, 32, 33). In these models, IL-10 actsby limiting TNF-� production and neutrophil influx (34). In ourmodel, administration of IL-10, at a dose that led to tissue con-centrations of IL-10 similar to those found in germ-free mice, waseffective in diminishing reperfusion-associated tissue damage anddelaying lethality. More importantly, the administration of anti-IL-10 Abs before the reperfusion of the ischemic SMA in germ-free mice was followed by marked reperfusion-associated inflam-mation–roughly 70% of that observed in conventional mice–andlethality. In our experiments, the ability of endogenous or exoge-nous IL-10 to prevent lethality and tissue injury was clearly cor-related with the concentrations of local or systemic TNF-� (seeFigs. 3 and 5). The situation in LPS-injected germ-free and con-ventional mice was virtually identical. Contrary to conventionalmice, germ-free animals produced little TNF-�, did not die, andproduced large amounts of IL-10 following LPS. Moreover, ad-ministration of anti-IL-10 to germ-free mice enhanced the LPS-induced TNF-� production and was followed by markedly in-creased lethality. Thus, exogenously administered IL-10 couldmimic, and blockade of endogenous IL-10 could prevent, the ex-pression of a “no-inflammation” phenotype in two models of sys-temic inflammation in germ-free mice. The likelihood that otheranti-inflammatory molecules and/or pathways are involved isgiven by the inability of anti-IL-10 to reverse completely andIL-10 to mimic fully the phenotype of germ-free mice.
The present study did not investigate in any detail the cell typesresponsible for the production of TNF-� and/or IL-10. However,several studies have now shown that macrophages are an importantsource of the latter cytokines in the setting of I/R injury (35–37).To evaluate whether macrophages derived from germ-free andconventional mice differed in their ability to respond to inflamma-tory stimulation, resident peritoneal macrophages were obtainedfrom both strains and stimulated with LPS. As noted above, germ-free do not die after LPS injection in vivo and respond by produc-ing large quantities of IL-10. In contrast, LPS-stimulated macro-phages derived from germ-free mice responded by producing IL-10, whereas those from conventional mice produced TNF-�.Overall, the latter results show that the function of macrophages
may be fundamentally between conventional and germ-free miceand suggest that studies of macrophage function may provide cluesas to why germ-free preferentially produce IL-10.
It is interesting to note that the innate ability of germ-free toproduce IL-10 and, possibly, other molecules, was an active pro-cess that prevented the inflammatory phenotype, i.e., animals notexposed to an intestinal microbiota have an innate ability to pro-duce molecules with anti-inflammatory properties that suppress thedevelopment of an inflammatory response. In contrast, mice thathad a normal intestinal microbiota or in which the microbiota hadbeen reinstated were capable of responding to inflammatory stim-ulation, suggesting that the presence of microorganisms in the gutinduce a “state of alert” that is characterized by the loss of theinnate ability to produce IL-10 and, possibly, other molecules (e.g.,TGF-�). Indeed, colonization of the gastrointestinal tract of germ-free with gut bacteria of conventional mice was capable of pre-venting the preferential production of IL-10 and restoring inflam-matory responsiveness. It was interesting to notice that there is aneed for prolonged colonization of the gastrointestinal tract as aneffect of conventionalization was only seen after 14 days and wasmaximal at 21 days. In contrast, even prolonged treatment withlarge spectrum antibiotics was not capable of modifying the in-flammatory responsiveness of conventional mice, even if bacteriacould not be detected in feces of treated mice. The test used forbacterial detection is sensitive, suggesting that there was a de-crease of the number of bacteria below the number necessary toestablish an efficient ecological relation between host and micro-biota (usually around 107 bacteria) (18). It is not clear why theantibiotic treatment was ineffective in reversing the inflammatoryresponsiveness. It is possible that more prolonged treatment is re-quired to reverse the inflammatory reactivity of germ-free mice,but this is experimentally difficult to achieve. Alternatively, oncethe organism is programmed to inflame in response to stimulation–and not produce IL-10–it may not be possible to reverse to a “noinflammation” phenotype as that observed in germ-free mice. Fur-ther studies are clearly necessary to resolve this issue.
In conclusion, our studies demonstrate that the inability ofgerm-free mice to inflame in response to systemic LPS or reper-fusion-induced injury is largely due to the innate capacity ofthese mice to produce IL-10 and, possibly, other anti-inflammatory molecules. The mechanisms underlying the innateability of germ-free mice to produce IL-10 have not been in-vestigated here any further but our recent studies suggest thatlipoxin production underlies the greater production of IL-10 ingerm-free mice (D. G. Souza and M. M. Teixeira, unpublisheddata). The IL-10 produced then switches off proinflammatorycytokine production, inflammatory cell influx, and consequenttissue injury and lethality. The detailed understanding of themolecular interactions underlying innate IL-10 production is afundamental question that may unravel novel targets for treat-ment of acute and chronic inflammatory disorders.
AcknowledgmentsWe are grateful to Dr. Ricardo Gazzinelli for his helpful comments and toDr. Jacqueline Alvarez-Leite for the supply of conventional mice.
References1. Granger, D. N., K. Y. Stokes, T. Shigematsu, W. H. Cerwinka, A. Tailor, and
C. F. Krieglstein. 2001. Splanchnic ischaemia-reperfusion injury: mechanisticinsights provided by mutant mice. Acta Physiol. Scand. 173:83.
2. Carden, D. L., and D. N. Granger. 2000. Pathophysiology of ischaemia-reperfu-sion injury. J. Pathol. 190:255.
3. Lefer, A. M., and D. J. Lefer. 1996. The role of nitric oxide and cell adhesionmolecules on the microcirculation in ischaemia-reperfusion. Cardiovasc. Res.32:743.
4. Willerson, J. T. 1997. Pharmacologic approaches to reperfusion injury. Adv.Pharmacol. 39:291.
4145The Journal of Immunology
by guest on Novem
ber 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

5. Souza, D. G., D. C. Cara, G. D. Cassali, S. F. Coutinho, M. R. Silveira,S. P. Andrade, S. P. Poole, and M. M. Teixeira. 2000. Effects of the PAF receptorantagonist UK74505 on local and remote reperfusion injuries following isch-aemia of the superior mesenteric artery in the rat. Br. J. Pharmacol. 131:1800.
6. Souza, D. G., G. D. Cassali, S. Poole, and M. M. Teixeira. 2001. Effects ofinhibition of PDE4 and TNF-� on local and remote injuries following ischaemiaand reperfusion injury. Br. J. Pharmacol. 134:985.
7. Souza, D. G., A. C. Soares, V. Pinho, H. Torloni, L. F. L Reis, M. M. Teixeira,and A. A. M. Abalen. 2002. Increased mortality and inflammation in TSG-14transgenic mice after ischemia and reperfusion injury. Am. J. Pathol. 160:1755.
8. Sun, Z., X. Wang, A. Lasson, A. Bojesson, M. Annborn, and R. Andersson. 2001.Effects of inhibition of PAF, ICAM-1 and PECAM-1 on gut barrier failure causedby intestinal ischemia and reperfusion. Scand. J. Gastroenterol. 36:55.
9. Sun, Z., K. Olanders, A. Lasson, M. Dib, M. Annborn, K. Andersson, X. Wang,and R. Andersson. 2002. Effective treatment of gut barrier dysfunction using anantioxidant, a PAF inhibitor, and monoclonal antibodies against the adhesionmolecule PECAM-1. J. Surg. Res. 105:220.
10. Miura, M., X. Fu, Q. W. Zhang, D. G. Remick, and R. L. Fairchild. 2001. Neu-tralization of Gro � and macrophage inflammatory protein-2 attenuates renalischemia/reperfusion injury. Am. J. Pathol. 159:2137.
11. Tsuruma, T., A. Yagihashi, K. Tarumi, and K. Hirata. 1998. Anti-rat IL-8 (CINC)monoclonal antibody administration reduces ischemia-reperfusion injury in smallintestine. Transplant. Proc. 30:2644.
12. Zingarelli, B., Z. Yang, P. W. Hake, A. Denenberg, and H. R. Wong. 2001.Absence of endogenous interleukin 10 enhances early stress response duringpost-ischaemic injury in mice intestine. Gut 48:610.
13. Gilmont, R. R., A. Dardano, J. S. Engle, B. S. Adamson, M. J. Welsh, T. Li,D. G. Remick, D. J. Smith, Jr., and R. S. Rees. 1996. TNF-� potentiates oxidantand reperfusion-induced endothelial cell injury. J. Surg. Res. 61:175.
14. Cicalese, L., T. R. Billiar, A. S. Rao, and A. J. Bauer. 1997. Interaction betweenischemia/reperfusion-induced leukocyte emigration and translocating bacterialenterotoxins on enteric muscle function. Transplant. Proc. 29:1815.
15. Sorkine, P., O. Szold, P. Halpern, M. Gutman, M. Greemland, V. Rudick, andG. Goldman. 1997. Gut decontamination reduces bowel ischemia-induced lunginjury in rats. Chest 112:491.
16. Bauer, P., J. M. Russell, and D. N. Granger. 1999. Role of endotoxin in intestinalreperfusion-induced expression of E-selectin. Am. J. Physiol. 276(2 Pt. 1):G479.
17. Turnage, R. H., K. S. Guice, and K. T. Oldham. 1994. Endotoxemia and remoteorgan injury following intestinal reperfusion. J. Surg. Res. 56:571.
18. Pleasants, J. R. 1974. Gnotobiotics. In Handbook of Laboratory Animal Science.E. C. Melby, Jr. and N. H. Altman, eds. Cleveland, CRC Press, p. 119.
19. Saria, A., and J. M. Lundberg. 1983. Evans blue fluorescence: quantitative andmorphological evaluation of vascular permeability in animal tissues. J. Neurosci.Methods 8:41.
20. Souza, D. G., S. F. Coutinho, M. R. Silveira, D. C. Cara, and M. M. Teixeira.2000. Effects of a BLT receptor antagonist on local and remote reperfusion in-juries after transient ischemia of the superior mesenteric artery in rats. Eur.J. Pharmacol. 403:121.
21. Bradley, P. P., D. A. Priebat, R. D. Christensen, and G. Rothstein. 1982. Mea-surement of cutaneous inflammation: estimation of neutrophil content with anenzyme marker. J. Invest. Dermatol. 78:206.
22. Goldberg, M., and J. Fine. 1945. Traumatic shock. XI. Intestinal absorption inhemorrhagic shock. J. Clin. Invest. 24:445.
23. Grotz, M. R., J. Ding, W. Guo, Q. Huang, and E. A. Deitch. 1995. Comparisonof plasma cytokine levels in rats subjected to superior mesenteric artery occlusionor hemorrhagic shock. Shock 3:362.
24. Swank, G. M., and E. A. Deitch. 1996. Role of the gut in multiple organ failure:bacterial translocation and permeability changes. World J. Surg. 20:411.
25. Cicalese, L., W. Yacoub, J. Rogers, J. J. Fung, A. S. Rao, and T. E. Starzl. 1999.Translocation of bacteria from the gastrointestinal tract: protection afforded bylisofylline. Transplant. Proc. 31:575.
26. Nezu, Y., M. Tagawa, Y. Sakaue, Y. Hara, S. Tsuchida, and R. Ogawa. 2002.Kinetics of endotoxin concentration and tumor necrosis factor-�, interleukin-1�,and interleukin-6 activities in the systemic and portal circulation during smallintestinal ischemia and reperfusion in dogs. Am. J. Vet. Res. 63:1680.
27. Poltorak, A., X. He, I. Smirnova, M. Y. Liu, C. Van Huffel, X. Du, D. Birdwell,E. Alejos, M. Silva, C. Galanos, et al. 1998. Defective LPS signaling in C3H/HeJand C57BL/10ScCr mice: mutations in TLR4 gene. Science 282:2085.
28. Qureshi, S. T., L. Lariviere, G. Leveque, S. Clermont, K. J. Moore, P. Gros, andD. Malo. 1999. Endotoxin-tolerant mice have mutations in Toll-like receptor 4(TLR4). J. Exp. Med. 189:615.
29. Dobrovolskaia, M. A., and S. N. Vogel. 2002. Toll receptors, CD14, and mac-rophage activation and deactivation by LPS. Microbes Infect. 4:903.
30. Belosjorow, S., I. Bolle, A. Duschin, G. Heusch, and R. Schulz. 2003. TNF-�antibodies are as effective as ischemic preconditioning in reducing infarct size inrabbits. Am. J. Physiol. 284:H927.
31. Colletti, L. M., A. Cortis, N. Lukacs, S. L. Kunkel, M. Green, and R. M. Strieter.1998. Tumor necrosis factor up-regulates intercellular adhesion molecule 1,which is important in the neutrophil-dependent lung and liver injury associatedwith hepatic ischemia and reperfusion in the rat. Shock 10:182.
32. Eppinger, M. J., P. A. Ward, S. F. Bolling, and G. M. Deeb. 1996. Regulatoryeffects of interleukin-10 on lung ischemia-reperfusion injury. J. Thorac. Cardio-vasc. Surg. 112:1301.
33. Lane, J. S., K. E. Todd, M. P. Lewis, B. Gloor, S. W. Ashley, H. A. Reber,D. W. McFadden, and C. F. Chandler. 1997. Interleukin-10 reduces the systemicinflammatory response in a murine model of intestinal ischemia/reperfusion. Sur-gery 122:288.
34. Souza, D. G., R. Guabiraba, V. Pinho, A. Bristow, S. Poole, and M. M. Teixeira.IL-1-driven endogenous IL-10 production protects against the systemic and localacute inflammatory response following intestinal reperfusion injury. J. Immunol.170:4759.
35. Naidu, B. V., S. M. Woolley, A. S. Farivar, R. Thomas, C. H. Fraga, C. H. Goss,and M. S. Mulligan. 2004. Early tumor necrosis factor-� release from the pul-monary macrophage in lung ischemia-reperfusion injury. J. Thorac. Cardiovasc.Surg. 127:1502.
36. Maxey, T. S., R. I. Enelow, B. Gaston, I. L. Kron, V. E. Laubach, and A. Doctor.2004. Tumor necrosis factor-� from resident lung cells is a key initiating factorin pulmonary ischemia-reperfusion injury. J. Thorac. Cardiovasc. Surg. 127:541.
37. Frangogiannis, N. G., C. W. Smith, and M. L. Entman. 2002. The inflammatoryresponse in myocardial infarction. Cardiovasc. Res. 53:31.
4146 INFLAMMATION AND THE INTESTINAL MICROBIOTA
by guest on Novem
ber 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from