the effects of various physical and chemical agents on a
TRANSCRIPT

Butler University Botanical Studies Butler University Botanical Studies
Volume 13 Article 8
The Effects of Various Physical and Chemical Agents on a The Effects of Various Physical and Chemical Agents on a
Staphylococcus Bacteriophage Staphylococcus Bacteriophage
Donald H. Holmes
Follow this and additional works at: https://digitalcommons.butler.edu/botanical
The Butler University Botanical Studies journal was published by the Botany Department of
Butler University, Indianapolis, Indiana, from 1929 to 1964. The scientific journal featured
original papers primarily on plant ecology, taxonomy, and microbiology.
Recommended Citation Recommended Citation Holmes, Donald H. (1956) "The Effects of Various Physical and Chemical Agents on a Staphylococcus Bacteriophage," Butler University Botanical Studies: Vol. 13 , Article 8. Retrieved from: https://digitalcommons.butler.edu/botanical/vol13/iss1/8
This Article is brought to you for free and open access by Digital Commons @ Butler University. It has been accepted for inclusion in Butler University Botanical Studies by an authorized editor of Digital Commons @ Butler University. For more information, please contact [email protected].

Butler University Botanical Studies
(1929-1964)
Edited by
J. E. Potzger

The Butler University Botanical Studies journal was published by the Botany Department of Butler University, Indianapolis, Indiana, from 1929 to 1964. The scientific journal featured original papers primarily on plant ecology, taxonomy, and microbiology. The papers contain valuable historical studies, especially floristic surveys that document Indiana’s vegetation in past decades. Authors were Butler faculty, current and former master’s degree students and undergraduates, and other Indiana botanists. The journal was started by Stanley Cain, noted conservation biologist, and edited through most of its years of production by Ray C. Friesner, Butler’s first botanist and founder of the department in 1919. The journal was distributed to learned societies and libraries through exchange. During the years of the journal’s publication, the Butler University Botany Department had an active program of research and student training. 201 bachelor’s degrees and 75 master’s degrees in Botany were conferred during this period. Thirty-five of these graduates went on to earn doctorates at other institutions. The Botany Department attracted many notable faculty members and students. Distinguished faculty, in addition to Cain and Friesner , included John E. Potzger, a forest ecologist and palynologist, Willard Nelson Clute, co-founder of the American Fern Society, Marion T. Hall, former director of the Morton Arboretum, C. Mervin Palmer, Rex Webster, and John Pelton. Some of the former undergraduate and master’s students who made active contributions to the fields of botany and ecology include Dwight. W. Billings, Fay Kenoyer Daily, William A. Daily, Rexford Daudenmire, Francis Hueber, Frank McCormick, Scott McCoy, Robert Petty, Potzger, Helene Starcs, and Theodore Sperry. Cain, Daubenmire, Potzger, and Billings served as Presidents of the Ecological Society of America. Requests for use of materials, especially figures and tables for use in ecology text books, from the Butler University Botanical Studies continue to be granted. For more information, visit www.butler.edu/herbarium.

rsal, ancapical, apical and
l.efevrc. F. K. and W. A.
tification of the alga and 'ersity of British Colum
md printed macter. The
with the permission of
Iluver, British Columbia.
Cl Drouet are gratefully
THE EFFECTS OF VARIOUS PHYSICAL AND CHEMICAL AGENTS ON A STAPHYLOCOCCUS
BACTERIOPHAGE
DONALD H. HOLMES
Eli Lilly and Company
Indianapolis, Indiana
In recent years considerable attencion has been focused on a group of organ
isms known as bacterial viruses or bacteriophages. These minute virus particles
are parasitic upon baererial cells and probably most bacteria are susceptible to one or more of chem. There are several excellent reviews and symposia covering
the various steps of bacteriophage multiplication and the effeCts of inhibiting
agents (1, 29, 30). The reader is referred to them for an explanation of the
processes involved in attachment to and multiplication within the host cell.
The experimental work in this paper is divided into twO sections: (1) the
effect of physical agents and (2) the effecc of chemical agents on a staphylococ
cus bacteriophage. The physical treatments include chermal inactivation, ultra
sonic vibration, lyophilization, long term storage, effect of temperature on
adsotption rate and photoreactivation after exposure to ultraviolet light. Chem
ical treatments included suspension in various salt solutions and the effect of
several pH values on phage stability. It is apparent that some of chese procedures could be classified as eicher physical or chemical in accion or more
properly as physico-chemical. However, for the purpose of this paper the more
simple distinction will be followed beginning with che effects of physical agents. Preliminary experiments had shown that the phage used was inactivated
in 30 minutes when suspended in broth at 60° and that ic gradually loses activ
ity when stored in broth at 4°. High speed centrifugation procedures also cause rapid inactivation.
GENERAL MATERIALS AND METHODS
Media: Tryptose phosphate broch (TPB) was used to grow the host organism
and as a diluent. One percent base layer agar plates, 1.570 agar slants and
0.7% top layer agar tubes were prep.ued by adding sufficient agar to the TPB.
All media were sterilized by auroclaving 15 minutes at 15 pounds. The final pH was 7.4.
Phage (lnd Host Bacterium: The host bacterium SK9 and the phage PI are
isolates from cultures used routinely in antiphage assays at Eli Lilly and Com
pany. Lysates of SK9 by PI titer from 5 to 7 X 10 91m1. after Selas filtration.
49

l
i
Phage Titerhtg: The PI samples were titered using the method described by Adams (1). The plates were inverted, then incubated overnight at 37° and
counted on a Quebec colony counter.
ADSORPTION RATE AT VARIOUS TEMPERATURES
The first step in virus multiplication is adsorption, or attachment to the surface of the host cell prior to penetration of the cell membrane. Information regarding the adsorption phenomenon is of value since this action or a similar series of events is assumed to be universal among all plant, mammalian or bac
terial viroses as the initial step in infection of the hOSt cell.
Matl'rial and Methods: SK9 was grown in TPB to 2 x 107 organisms/mI., centrifuged and resuspended in an equal amount of physiological saline. The phage was diluted to 1 x 108 / m!. in saline and both phage and host organism were brought to the desired temperature before mixing. At t=O, one ml. of PI was added to nine m!. of SK9 and the tube was shaken. One half ml. samples were removed at intervals then diluted 1/2000 in saline at 2° to stop further adsorption. A phage titer was taken to determine the original titet and the samples were centrifuged 10 mtnutes at 5900g. The supernate was titered. Velocity constants were computed from the formula:
2.3 initial phage titer k = . 11 x log fi I ht X ce count na p age titer
Results and Discussion: Velocity constants for the attachment of PI to SK9 in saline range from 489 x 10.12 cm! min·1 at 1° to 729 X 10.11 at 45 0
(fig. 1). At a given temperature the maximum r;lte of virus adsorption is attained and additional increases in temperature do not result in a correspondingly larger velocity constant. The maximum r;lte of adsorption for this system lies between 28° and 37 0 since no increase occurs above the higher temperature. This same effect was shown by Puck, et ;II. (2) with coliphage Tl in broth or buffer with Mg++ where adsorption re;lches a maximum around 37° and falls off with decreasing or increasing temperatures. In addition to the temperature effect adsorption is influenced by the presence of certain co-factors and salts and the physiological condition of the host cell.
LONG TERM STORAGE
Information regarding the ability of microorganisms to remain viable when stored under various conditions is of prime importance. Long term storage of cultures can be beSt accomplished by first freeze-drying the material. The results found using this technique with PI are described elsewhere in this paper. However, bacteriophage suspended in liquid remains viable for some time. It is with this type of storage that the following experi~ents were concerned.
50
Material and Mel imately 4 x l05/m l. (pH 6.8). A sampl 37 0 in stoppered tes
Results and Dis
suspended in distillc
also occurs at il4 0
weeks over 99.9% pera cure in salioe aT. tivation occurs whe lost in one week at 3 pera ture. When PI in three weeks. In This phage can be sl is to be expected. F retain viable phage J
Several bacteriof tery phages were pre Fi ve of these six lost T6, grown in synth tivated. Many mam after years of storag PI was lyophilized 1
Material and Me
Selas candle and titl and rapidly shell fro one minute. The fla! dried for 21 hours. ,
form of brown £lake in a tightly stopperel or water then Jyophi
Results and Disc
ess of lyophilization but no further decr of such a large drop fresh PI Iysa tes fror in water or saline b were lyophilized anc

llcthod described by ernight at 37° and
tATURES
attachment to the nbrane. Information ; action or a similar mammalian or bac
107 0rganisms/mI.,
iological saline. The e and host organism
[=0, one ml. of PI )ne half m!. samples l 2c to stop further
~ginal titer and the pel'na te was ti tered,
ter
:u
ttachment of P 1 to 729 x 10,11 at 45°
rus adsotption is atin a correspondingly
I for this system lies higher temperature.
,hage T 1 in broth or
around 37° and falls ] to the temperature
co-factors and sal ts
remain viable when
..ang term storage of $ the material. The
where in this paper. for some time. I t is ere concerned.
MatC'Tial and Methods: A freshly prepared phag~ stock was diluted to approximately 4 x 10'/ml. in broth (pH 7.4), saline (pH 6.8) and in distilled water (pH 6.8). A sample of each was stored at 4°, room temperature (23°) :md 37° in stoppered test tubes. Titers were made at intervals.
Results and Discussion: PI was completely inactivated in three days when suspended in distilled water at room temperature or 37°. Rapid inactivation also occurs at 4° in water as only 10% was active after one week. In four weeks over 99.9% was inactive. PI was found unstable at 37° or room temperature in saline and is 60% inactivated after one week at 4°. The least inactivation occurs when PI is held in TPB. In this medium 90% of the phage is lost in one week at 37°. Only 1% remains active after two months at room temperature. \'(fhen PI is suspended in TPB and stored at 4° it loses very little titer in three weeks. In two months the titer drops to 20% of the original value, This phage can be stored in broth for short periods bu t a considerable titer drop is to be expected. Freeze-drying of stock preparations is preferable in order to
retain viable phage partieles.
LYOPHILIZATION
Several bacteriophages have been lyophilized with varying success. Six dysentery phages were prepared using this procedure by Schade and Caroline (3,4, 5).
Five of these six lost no activity after one year over a dessicanr at 37°. Coliphage T6, grown in synthetic media and lyophilized by Putnam, et al. (6) was inaetivated. Many mammalian viruses resist freeze-drying and were found viable after years of storage. Hofstadt, et a1. (7) and Scherp, et al. (8). Bacteriophage PI was lyophilized using standard procedures.
Material and Mr.thods: Freshly prepared lysates were filtered through an 02 Selas candle and titered. Twenty-five m!. were placed in a round bottom flask and rapidly shell frozen in an alcohol/C02 barh. The freezing required less than one minute. The flask was atached to a lyophilization apparatus, evacuated and dried for 21 hours. A t the conclusion of the drying process the material, in the form of brown flakes, was scraped off the sides of the flask, weighed, and placed in a tightly stoppered tube. Other phage lysatcs were first dialized at 4° in saline or water then lyophilized.
Results a'nd Disc'Ussio11: Approximately 85% of the phage is lost in the process of lyophilization. An additional 6% loss occurred in storage after 30 days but no further decrease was seen during six additional months at 4°. In spite of such a large drop in phage activity no difficulty was experienced in preparing fresh PI Iysates from the lyophilized material. Lysates which had been dialyzed in water or saline before freeze-drying lost no titer. When these preparations were lyophilized and then reconstituted in either saline or TPB over 99 % of
51

10 9
8
7
6
5
4
3
2
..... a..
0 W If) a: I
0 9 (f) 0 8
« Z
7
::J 6
5
4
3
0--0 I"J.. '" 489 X 10-12 0.1 3 M1N,-1
2 a-----D. 20" 280 X 10- 11
o---a 28" 484XIO- 1i
X--X 37" 748 X 10-11
. 45" 729 X 10-(1
I OL--'----::'=----:~-J.,---!---:':---:'----:80.L--9,L0,-----1...L0-0-,--l1-0-..J12LO-..J,3LO-...J14 0
MINUTES
FIGURE 1 The effec( of heat on (he adsorption rare of PI bacteriophage
to irs host cell S. att-reUJ SK9 in saline.
52
10
9
8
7
6
5 x
4
3
..... 2a.
(.) z -> > a:
r::J (f) 9
8fZ 7 W U 6 a: W 5Q
4
3
2
~
0
X
1,--_..1.-_ o
V:

489 X 10-IZ CM3 MIN.-I
280 X lO-"
484X 10- 11
748 X 10-11
729 X 10-11
130 140
of PI bactcriophage n saline.
10
9
8
7
6
5
4
x3
...... 2a..
l') Z -> > a: ::) I (f) 9
8f-Z 7 W U 6 a: W 5n.
4
3
2
fr---l!:,. 450 .,£= 1 I~ X 10-3 CM 3 MIN.- r
0----0 ~OO 230 x 10-3
X--x 550
19.8. 10-3
IL--.l.---'--~_-'-_....I..-_-':-.,---L_-.l._-'_---J'--_L--_.l...-_.l.-----J
o 60
MINUTES
FIGURE 2
Log of uninactivared bacteriophage PI in broth at wrious tempcratures as a function of time. 45°-55·.
53

Log
' 0
2
10
9
8
7
6
5
4
3
W
~ 2I
Il. 0 a:: w IL> <CO
~ 9
Z 8
:> 7 :>
6a:: :::) (/) 5
~ 4
3
FIGURE 4 Log of the first order specific reaction rates for inactivation
of PI bacreriophage as a function of temperature.
::l.0
MolOI,Ooo CALORIES/MOLE
1.0
4.0,-----------,-----,
t ABSOLUTE TEMPERATURE" 10-3
t'l, o
"~ 20
o o .J
54
SECONDS
30 60 90 120 150 180
60·
.k~ 1307 X 10·'CM' MIN-'
1 0
2
FIGURE 3 Log of uninactivated bacteriophage
PI in brorh at 50° as a function of time.
10 9
8
7
6
5
4
3
-Q. 2
0 Z
>->a: :) tI)
~ Z W u a: w Q.

1412 13II10987
~ - T6}k=o 768 0 ... 3 MIN-'0--0 Pi .
55
MINUTES
FIGURE 5
Log of uninactivated bactc:'riophages PI and T6 in broth treared wirh radio frequency oscillation as a funcrion of time.
10 9
8
7
6
5
4
3
W ~ <{
2I Cl 0 a::: w IU <{ 1iI
~ Z > :> a::: => (/)
~
UTE TEMPERATURE. 10-3
~.IOI,OOO
CALORIES/MOLE
FIGURE 4 g£ che first order specific ion rates for inaaivacjoo PI bacteriophage as a ccion of cemperature.

{he phage was found to be inacr.ivared. If PI were lyophilized in ampoules under
vacuum or dry nin'ogen assuring the complete absence of moisture no doubt its
srability would be greatly increased.
ULTRA-SONIC VIBRATION
Bacteriophages and other viruses have been shown to be very vulnerable to
the effects of high frequency vibration. Such inactivation usually proceeds as a
first order reaction.
Materials and Metbods: The phage was diluted in TPB and 50 ml. placed
in the water cooled Raytheon Magnetostriction sonic oscillatOr (Model DF-l 01) .
The frequency produced by this instrument is 10,000kcs. One tenth ml. samples
were removed at intervals, diluted and plated. A sample of T6 coliphage was also
prepared and trea ted in the same manner in order to compare the sonic effects
on both phages. Velocity constants were determined using the formula,
k = ~ x log ~~~ial phage titer t final phage titer
cited in Pollard and Reaume (12).
Remits and Discussion: PI was found to be inacrivared at approximately the
same rate as the coliphages T2, T4, T5 and T6 and megatherium phages M2
and M3. In one minute one half of the phage was destroyed and only 2%
remained active after Dve minutes exposure (fig. 5). All of the T series of
coliphages have been subjected to this form of inactivation by Anderson, et al.
(9). He found T2, T4, T5 and T6 to be more rapidly inactivated than the
smaller phages Tl, T3 and T7, and suggested that tbe larger, more complex
phages were more susceptible to sonic action. Friedman and Cowles (10), work
ing with a group of five B. megatberiu.m phages could find no relationship
berween size and relative sensitivity to sonoration. A staphylococcus phage was
reported by Krueger (11) to be over 99% inactivated after 10 minutes exposure
ro high frequency vibration. Velocity constants for both PI and T6 were
0.77cm 'min"'.
THERMAL INACTIVAnON
Velocity constants for the hear inactivation of various phages have been
determined. Cherry (13) investigated the effect of heat on a Streptococcus lactis phage at temperatures from 30 to 65 0. Other workers, Chang, et aI., (14), Pol
lard and Reaume (15) and Adams (16) observed the effects of heat on the
coliphage series.' Friedman and Cowles (17) ran heat inactivation curves on
their group of B. meglltheriu111 phages. In this paper a series of experiments were
performed with Pl at temperatures of from 45 to 60°.
56
MatCl'wls lind ml. was added to
brought to the dl
The tube was Sf chilled TPB, then
inactiva tion curv
constant equatior
plotted against th
manner developec reaction rate and
Remits and I at 45° for 180 m
for the same leng'
of the phage wa indicate that over
not prepared for
sampling. Velocit 1307 x 10-) at 60'
with those found
temperarure chan
for all phages su~
linea l' over its eni
p. value of 11,00 calories from 55
a non-linear cun
100,000 calories
Krueger (17) wi
A description
bacteriophages wa
all of the T coliph:
ble host cells, wo
tirers than similar
of the host cells PHTR and irradi,
hosr cell. Hill and
showed that no PI
PHTR was depen to ultraviolet light
in phage wbich I: phage was shown 1

1 ampoules under
lure no doubt its
~ry vulnerable to
IIIy proceeds as a
Id fO ml. placed
:Model DF-101). :enth m!. samples
oliphage was also the sonic effects
19 the formula,
Ipproximately the
~rium phages M2
~d and only 2 % : the T series of
Anderson, et al.
:tivated than the
:r, mote complex
wles (10), worki no relationship
~occus phage was
minutes exposure ~1 and T 6 were
phages have been 'rcptococcus lactis
tal., (14), Pol
of heat on the
ation curves on
xperimen ts were
},iraterials and Methods: P 1 was diluted in broth to 5 x 10",1 m!. One tenth
ml. was added to a tube containing 9.9 m!. of TPB which had been pteviously
brought to the desired temperature in a thermostatically controlled water bath.
The tube was shaken, samples removed at intervals, diluted immediately in
chilled TPB, then plated. The rate of inactivation (k) was calculated from the
inactivation curves (figs. 2 and 3) for each temperature using the velocity constant equation. The logarithms of the k values for each temperatures were
plotted against the reciprocals of the corresponding absolute temperatures in the
manner developed by Arrhenius in order to illustrate the relationship between
reaction rate and temperature.
Results ami Discussion: Seventeen percent of the phage PI was inactivated
at 45° for 180 minutes and approximately twice that amount was dead at 50°
for the same length of time. When PI was treated at 55 0 for 180 minutes 97%
of the phage was destroyed (fig. 2 and 3). E:xperiments performed at 65°
indicate that over 99% was lost in less than 30 seconds. Velocity constants were
not prepared for this temperature due to the difficulty in performing accurate sampling. Velocity constants (k) range from 1.15 x 10-JcmJmin-' at 45° to
1307 x 10'; at 60°. All thermal inactivation studies for this phage compare well
with those found by Krueger (17) in 1932 for a staphylococcus phage. The
temperature characteristic of thermal inactivation (fL) has not been determined
for all phages subjected to heat inactivation. In some cases the curve "'as not
linear over its entire length. Cherry (13) found his S. lactis phage to have a
fL value of 11,000 calories between 30 and 55° with an increase to 76,000
calories from 55 to 65 0 • The coliphage studies by Chang et al. (14) also had
a non-linear curve with two fL values. The value of fL found for PI was
100,000 calories and the curve was linear. This figure was also found by
Krueger (17) with a staphylococcus phage (fig. 4).
PHOTOREACTIVAnON
A description of the phenomonon known as photoreactivation (PHTR) of bacteriophages was first published in 1949 by Dulbecco (18). He found that
all of the T coliphages, after exposure to ultraviolet light and plating on suscepti
ble host cells, would, if incubated under strong visible light, result in higher
titers than similarly treated coliphages incubated in the dark. Pre-illuminaticn
of the host cells or of the irradiated phage with visible light resulted in no
PHTR and irradiated phage could only be reactivated after adsorption to the
host cell. Hill and Rossi (19) working with a dried preparation of phage Tl,
showed that no PHTR rook place with the dry material. They concluded that
PHTR was dependent upon the state of the phage at the time of exposure
to ultraviolet light. Dulbecco (18) and Watson (20) found no PHTR to occur
in phage which had been treated with x-rays. PHTR in PI staphylococcus phage was shown ro occur in the following experiments.
57

Materials and Methods: PI was irradiated using a 15 watt GE germicidal lamp with a filter giving maximum emission at 2570 A. Samples of phage diluted in TPB were exposed for periods of from three to three and one half minute~
so as to cause approximately 80% inactivation. The irradiated phage was titered on 12 plates. Four were immediately placed in total darkness, four were placed beneath one 40 watt fluorescent lamp at a distance of 26 em. and four beneath two 40 watt lamps at the same distance. All plates were incubated at 26° for 18 hours and then counted.
Results and Discussion: In these experiments reactivation titers amounted to twice the titer of the irradiated phage not treated with visible light. Titers of irradiated phage not reactiva ted averaged 15 % of the original untreated phage
but exposure to two fluorescent lamps for 18 hours resulted in titers equal to 29% of the untreated original phage. This effect has not previously been demonstrated using a staphylococcus phage. It is interesting to note that exposure of
the irradiated phage to the visible light produced by one lamp did not cause the reactivation that two lamps produced in the same length of time. Also, if phage plates previously exposed to one lamp received additional visible light after the initial 18 hour period no additional reactivation occurred (table III). The maximum PHTR occurs in 18 hours or less but the total PHTR for a given period is dependent upon the intensity of the illumination for that period.
Perhaps, and this possibility was not investigated, the only time when PHTR actually takes place is during the adsorption of the irradiated phage particle immediately after plating and all subsequent visible light treatment has no effect
on phage titer.
HYDROGEN ION CONCENTRATION
Friedman and Cowles (10) found two of five B. megatberium phages to be stable for one hour at 37° in broth at pH 6 to 9 and one each stable from pH 5 to 9, 5 to 10 and 6 to 10. Purified coliphage Tl is most stable at pH 6
but Ii tde loss of titer occurs a t pH values of from 4.3 to 7 after 24 hours in broth (Pollard and Reaume, 15). T7 is most stable from pH 6 to 8 (Kerby, et al. 21) and T6 is stable from pH 4.9 to 8.6 (Putnam, 22). No information regarding the stability of staphylococcus phages over a range of pH was found
in the literature.
Materials and Met/:JOds: Baero tryptose and NaCl were dissolved in water,
brought to the desired pH, and sterilized in the autoclave. Glucose and Na~HP04
were dissolved separately in water and sterilized, cooled and added to the tryptose-NaCl solution. Hydrochloric acid (O.lM) and NaOH (O.lM) were
employed where necessary in adjusting to the exact pH. The phage was diluted in the appropriate pH broth, then held in a 37° water bath or stored at 4° for 24 hours. Samples were removed at one and 24 hours for titering.
58
C'l 0 ......
0'""' '""'
0 0..c;
'""'8 ...0
c:: a> tn M
""t1 V
-e; c:: v CL; ...... " Q. to V> -= " ::l :>V>
v :z: Po0() t- oo
oj ..q '"Q. 0.;:
'" .0 ~ U '" oj
...0~ ......>-l p., L'>J:Q <Xl'" -< 0c:::
/:-< V> v
.",. 0::l-;
;>
~ Q. ~ 0 V> ::l
·3 0004;':
;> .... '" ..... ~~ ::l 0 ~ .. 0
Au ~: v ........... ".-Il.":..... v .. v
..q !-< :g
A'"til
A.'" S oj
rn
tt GE germicidal
s of phage diluted
one half minu tes
phage was titered
i, four were placed
. and four benea th
ubated at 26 0 for N 0 0 0 0 ..
n titers amounted .... 0 0 0 0 rl ':
isible light. Titers S <0 lD ~
OIl untreated phage ..c 0 0 0 0 0 0
0 .... r-in titers equal to .... en ~
.D ~
0 "<l' "<l'
ously been demon.... t-
C <7l lD lD CQ .... vI -v C'? <0 lD !>Jl , that exposure of os
OJ .D
mp did -v p.. 6not cause: c
~ 0 <0 <0 v .; oc ....
""" t oo 'i:: .... t-
of time. Also, if c. lD """ <0
"""'" OJ
~ ;;. tJ .....
ional visible light os U
v :: .D '" <I
'" Z 6t.() t- eo N lD eo >-< N "<l'
:lIrred (table III). '" oo e- en oo p.. ~ .... eo N ..c 0
otal PHTR for P- c "a 0 0 Q
';:; >-< :;;
on for that period. ~ "" lD 0 0 <0 ~
oo <0 0 G> U ~ 6 eo eo-. u .... -. os " .... eo N os -.
time when PHTR ~ .D Z .. u
......l ii: ~ ...... " phage particle im ~'" <0 <0 0 ......l 0 0
~ <0 U eo .... 0 en ~ c<: c .... ~ .... 00
""" ent has no effect 0 <: = 00 N t-< '" t-<
.g '0 v ~ :;; ::J ~ 0 0 <0 oo ....
os C'l e- C ;> v N ~ t
::r: u oo Nc a. '" 0 0 0 0 0
u '" '"::l 0 ::l
ium phages to be .;:: 0 '"
lD ~ .;:: oo N
'" 1""4;': r/l '" os each sta ble from
;> Il.~ ... ... g ... ;> ...... ..~ ::> ::l ::l ......0 "'" 0 0 0 0
ost stable at pH 6 ~~ ..c ,.r:: ..c ..c 0
tJ ~
"'" "<l' N
7 after 24 hours J:: .. ' .... ~ .... """ u en N
...... 1>.<: N N J:: OJ = ......
H 6 to 8 (Kerby, v
OJ v ~ ..
..c .D ~ ~). No information I-< "0 ;:s I-< " '" ~ ~ '"
.. .. ..c ..c of pH was found ..c ,.r:: u
~0 .. .... """ '" e '" 0 ~
~ N 0.> ~ OJ """ Il. .. 0. ..., 0. ..., S ol S
ol
dissolved in water, ol til lZl lZl
ose and Na2 HP04
added to the tryp
t! (O.lM) were
The phage was fcr bath Or stored
hours for titering.
59

,....; 0. ...
OS) oj
.r:: 0. ,g
-~ v oj
.D- v OJ -
(..l.l .~ >--l v Q::I 2 ...-<}-< '0
c 0 .~
.2: v ...'"... 8 0
.r:: 0.
0]...... c .......... u ~ ... ~ ~.. .. " ll.~
'" .;. ~
W
... ." 0 ..
~~~ ~~~ ~;:J
,;, ~
W
... ." 0 ...... ~ ceo .. ~~~
~=~ ~~
~ fil'"
~ 0C'l ..... MC'l
S 0 6 b ..... ..... .......... >< >< ><><
C> 0 LO 00 C'-J ° ~ oq>t:i -<'"
M 0C'l..... MC'l
<:, 6 0 0 ..... ..... ..... -<
>0: >< >0: >< 0 ~ M
M C'-JC'-J ~ "" tt:i -<"" '"
to 00 ~ -< ..... C'l
<:, 0 Q 0 ..... -< ..... ..... >< >< >< >< ~ LO rL) C'l ~ ..... oq 0
C'l ~ .,..:""
>:: 'n .....8 '" v v.... ol Q,) Q,)
oj p. .... .... c;l oj'":.a ,.Qi=. ,.QP<
c;l "'~ .... ::sa ::sS'" ,n Uoj Val,~ .... '" ::s '" .=,.-4 .S-I:: >:: .... .... '~..o.: '" "' .... "' ....a S ... "' .... "'oj.... al'H ol -:;;~~~ ~"tj..0'" p,o p.~rol::.... ~..". v-a....'" ~~
'.p "'I:: 2~.... '" -0",
._oj"'"al -:;So oj+'
bD v..o ,--0",
.... ;.o~'" oj oj ::s oj-o c;l'"
.... U .... -0.r:: .... I:: t: I:: .... >::p.., .......- H::; ...... ::s
60
Results IlI1d DisC1 5 to 7 for one hour ,
at pH 9. All the ph.
A 2570 drop In titer
at 37°,
PI In TPB at 4° loss occurred at pH's
occurred at pH 3 an 4° in pH S to 7 in ~
general, both mamm,
held Hl suspensions al
however, vanes over
except for short perio
Several phage syst
Ions on adsorption an
classifica tion of these
In the presence of Cit
Ts is inhibited in the
24) . None of the ad studied a large group
variation in regard to
staphylococcal phages
megatherium phages
to varying degrees.
Material a11d Metl were prepared contair
In TPB to 5 x lO} /
titered on standard TIl
Results and Disc! not inhibited by any
Burnet and McKiE
tion on a group of
sodium, potassium or
was heated at 60°, 1
calcium, magnesium (

Results and Discussio11: PI was found to be relatively stable from pH
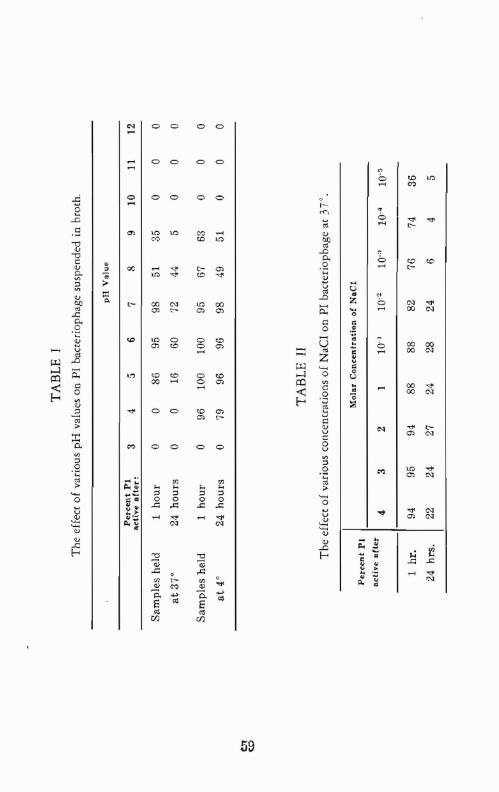
5 to 7 for one hour at J 7° dropping off sharply at pH 4 and down to 35% J[ pH 9. All the phage was inactivated at pH values below 5 and above 9.
A 25% drop in titer was seen when PI was held in pH 7 TPB for 24 hours at 37°.
PI in TPB at 4° was inactivated at an expected slower rate. Little titer loss occurred at pH's from 4 to 7 aher one hour but complete inactivarion occurred at pH 3 and pH 10. The phage remained stable after 24 hours at 4° in pH 5 to 7 in TPB. Fifty percent was destroyed at pH 8 (table I). In
general, bOth mammalian viruses and bacteriophages are most Hable when held in suspensions at pH values near neutrality. The zone of pH stability, however, varies over a range of pH 4 to 10 with very few viruses surviving except for short periods at pH's above 10 or below 4.
CITRATE ION
Several phage systems have been examined regarding the effect of citrate ions on adsorption and multiplication. It was suggested at one time that the classification of these organisms could be partially based on their behavior in the presence of citrate (Burnet, 2J). Multiplication of coliphages Tl and T5 is inhibited in the presence of citnte but adsorption is uneffected (Adams, 24). None of the other T series phages are effected. Burnet and McKie (25) studied a large group of dysentery-Salmonelb phages and found considerable variation in regard to citrate sensitivity. Rountree (26) divided a number of staphylococcal phages into sensitive and insensitive groups. AU five of the megatherium phages examined by Friedman and Cowles (10) were sensitive to varying degrees.
Mer/erial (Iud Methods: Top layer agar tubes and bottom layer agar plates
were prepared containing from 10" to 10"M sodium citrate. PI was diluted in TPB to 5 x 1oj / ml. then titered using the citrate agar. Controls were titered on standard TP agar.
Results and Discussion: The multiplication of Pion the host cell was not inhibited by any concentration of sodium citrate tested.
DIVALENT SALTS
Burnet and McKie (25) investigated the effect of heat and salt concenttation on a group of Salmonella and dysenrery phages and found dilu tion in sodium, potassium or ammonium salts resulted in rapid loss when the phage
was heated at 60°. The addition of small quantities of divalent salts such as calcium, magnesium or barium partially or completely prevented this inactiva
61

tion. The addition of anyone of four divalent salts at 10'; M prevents phage loss
at 37° for one hour on threc megatherium phages (Friedman, 27). The other two megatherium phages were only partially stabilized by the same salts. All except Hg++ and Pb++ had a definite protective effect on coliphage T5 in saline (Adams, 24).
Material and Methods: All divalent salts were prepared in 0.15 M NaCI and brought to pH 7. The phage was dlluted and added to the salt concentrations then incubated at 37° in a water bath. Titers were made at one and 24 hours.
Results a'nd Discussion: P I was completely inactiva ted in one hour at 37 0
In the presence of 10.2 or 10.3 M Cu++, Pb++, or Fe++ ions and lost
80% in 10.2 M Cd++. None of the other salts used at these concentrations prevented inactivation any more than the 0.15 M N.ICI control. The stability of PI is not increased when diluted in divalent salt solutions.
SODIUM CHLORIDE
Only one group of phages has been examined for the effect of different NaCl concentrations on phage stability. This group consisted of the five B. megathen'um phages studied by Friedman (18). Four of these phages were inactivated completely in NaCl concentrations of from 10" to 5 X 10.2 M after one hour at 37°. All were stable for the same length of time in 1 M solutions.
Material and Methods: Salt solutions were prepared using reagent grade NaCl dissolved in deionized water. The phage was dihlted in deionized water to the desired concentration then 0.1 ml. was added to the tubes containing 9.9 mt. of each salt solution at 37°. Incubation was continued at 37° and titers were made at one and 24 hours.
Results and Discussi011: In one hour at 37° from 5% to 26% of the PI had been inactivated in the solutions containing from 4 M to 10.4 M NaCl. The least inactivation occurred at NaCI concentrations of 2 M to 4 M.
Approximately 75 % of the phage was lost in 24 hours at salt concentrations of 4 M to 10-2 M and ovet 90% at 10-l M to 10· j M (table II).
SUMMARY
A staphylococcus bacteriophage lysate was treated with various physical and chemical agents and the following results were obtained:
Bacteriophage PI is inactivated by heat at a logarithmic rate and possesses a temperature characteristic (Ik) of 100,000 calories/mole. Inactivation by
62
high f requenc)
of 0.77 cm'm exposed to stn to SK9 range
most stable at none' contribul
citrate ions d( TPB little loss bu t there is C(
Sincere apl of the research
critical readin~
on preparation reports and of for gift of su their equipmen
l. ADAMS, M, ieal Resear'
2. PUCK, THl'
ism of \'iru J. ExpeL N
3. SCHADE, A valent dyse tions and ~
4. dry and st:
ophage dUJ
L
dry and st~
iry, age, aJ
6. PUTNAM, I chemical SI
khia coli I
7. HOFFSTADl
of certain' Dis. 78:18:

LO-) M prevents phage loss
~riedman. 27). The other
eo by the same salts. All
effect on coliphage Ts in
rtpared in 0.15 M NaCl
led to the salt concentra
were made at one and 24
ated in one hour at 37°
or Fe+ + ions and lost
d at these concentrations
ICI control. The stability
:llutions.
r the effect of different
consisted of the five B.
ur of these phages were
om ID-' to 5 x 10.2 M _ length of time in 1 M
ed using reagent grade
. uted in deionized water
to the tubes contain Lng
s continued at 37° and
m 5% to 26% of the
from 4 M to 10.4 M (rations of 2 M to 4 M.
s at salt concentrations
(table II).
d with varLous physical
ined:
. thmic ra te and possesses
/mole. Inactivation by
high frequency oscillation is a first order reaction with a velocity constant
of 0.77 cmJmin". Photo-reactivation occurs when ultraviolet treated PI is
exposed to strong light in the visible spectrum. Adsorption conStants of PI
to SK9 range from 489 x 10·'2 at 10 to 748 x 10·JlcmJmin·! at 37°. PI is
most stable at pH values from 5 to 7. Of the mono- or divalent ions tested,
none contributed significantly to the StabilitY of the phage; the presence of
citrate ions does nOt prevent phage multiplication. If PI is held at 4° in
TPB little loss of titer occurs in one month. PI can be successfully lyophilized
but there is considerable loss in the process.
ACKNOWLEDGMENTS
Sincere appreciation is expressed to Dr. Rex N. WebSter for superviswn
of the research, for hLs constant interest in the progress of the work, and for
critical reading of the manuscript; to the late Dr. J. E. Potzger for suggestions
on preparation of the manuscript; to my wife, Shirley, for typing of progress
reports and of the thesis. Thanks is also expressed to the Eli Lilly and Company
for gift of supplies used in the research and for permission to use some of
their equipment for experimentation.
LITERATURE CITED
1. ADAMS, MARK. 1950. Methods of study of bacterial viruses. Methods In Medical Research, Vol. 2. The Yearbook Publishers, Chicago, Illinois.
2. PUCK, THEODORE T., ALAN GAREN, AND JEWELL CLINE. 1951. The mechanism of virus attachment to host cells, 1. The role of ions in the primary reaction. J. Exper. Med., 93:65-88.
~. SCHADE, ARTHUR L., AND LEONA CAROLINE. 194.1. The preparation of a polyvalent dysentery bacteriophage in a dry and stable form. I. Preliminary invenigations and general procedure. J. Bacr. 46:46~-4n.
4. 1944. The preparation of a polyvalent dysentery bacteriophage in a dry and stable form. II. Factors affecting the stabilization of dysentery bacteriophage during lyophilization. J. BaCt. 48:179-190.
5. 1944. The prepararion of a polyvalent dysentery bacteriophage in a dry and srable form. Ill. Stability of rhe dried bacteriophage tOward hear, humidity, age, and acidiry. J. Bact. 48:24~-251.
6. PUTNAM, FRANK W., LLOYD M. KOZLOFF, AND JANtCE C. NEAL. 1949. Biochemical studies of virus reproduction. I. Purification and properties of Escherkhill coli bacteriophage T6. J. BioI. Chern. 179:303-323.
7. HOFFSTADT, RACHEL E.. AND HELEN B. TRIPI. 1946. A srudy of the survival of certain strains of viruses after lyophilizarion and prolonged stor~ge. ]. Infect. Dis. n:18~-189.
63

8. SCHER!', H. \'(1., E. W. FLOSDORF, AND D. R. SHAW. J938. Survival of influ 24. ADAMS, MAR". enZa virus under various conditions. J. Immuno!. 34:447-454. Gen. Physio!' 32:
9. ANDERSON, THOMAS F., SHIELA BOGGs, AND BETTY C. WINTERS. 1948. The 25. BURNETT, F. M. relative sensitivities of bacterial viruses to intense sonic vibration. Science fesred in bacreric J08: 18. 200.
10. FRIEDMAN, MURRAY, AND P. B. CoWLES. 1953. The bacteriophages of Bacillus 111-ega/!Je·r;ufII. 1. Serological, physical and biochemical properties. J. Bact.
66:379-385.
IJ. KRUEGER, ALBERT P., B. B. BROWN, .~NO E. J. SCRIBNER. 1941. The effect of sonic vibrations on phage, precursor and the bacterial substrate. J. Gen. Physio!.
24 :691-698.
12. POLLARD, ERNEST. 1953. The physics of VIruses. Academic Press, Inc. New
York, N. Y.
13. CHERRY, WILLIAM B., AND D. WATSON. 1949. The Streptococcus lac/is host
virus system. I. Factors influencing quantitative measurement of the virus. J.
Bact. 58:601-610.
14. CHANG, SHIH 1., MARY \'(1ILLNER, AND LOIS TEGARDEN. 1950. Studies on the destruction of bacterial virus. II. Kinetics in the thermodestruction of bac terial virus (bacteriophage against E. coli) in water. Am. J. Hyg. 52: 194-201.
15. POLLARD, ERNEST, AND MARJORIE REAUME. 1951. Thermal inactivation of bacterial viruses. Arch. Biochem. and Biophys. 32:278-287.
16. ADAMS, MARK. 1951. Classification of bacrerial viruses: characteristics of the 1'5 species and of the 1'2, C16 species. ]. Bact. 64:387-396.
17 KRUEGER, ALBERT P. 1932. The heat inaCtivation of antistaphylocaccus bacreriophage. ]. Gen. Physio!' 15:363-368.
18. DUi..BECCO, RENATO. 1949. Reactivation of ul t ra - violet -inactiva ted bacreri oph;l.ge by visible light. Nature 163 :949-9 50.
19. HILL, RUTH F., AND H,\ROLD Ross!. 1952. Absence of photo-reactivation in 1'1 bacteriophage irradiated with ultraviolet in the dried state. Sci~nce 116:425.
20. \'(1 AnON, JAMES D. 1950. The properties of x-ray inactivated bacteriophage. 1. Inactivation by direct effect. ]. Bact. 60:697-718.
21. KERBY, G. P., R. A. GOWDY, M. 1. DILLON, D. G. SHARP, E. S. Dll..I.ON, T. Z. CSAK Y, AND J. W. BEARD. 1949. Purification, pH stability, and sedimentation properties of the 1'7 hacteriophage of E. coli. J. Immuno!. 63:93-107.
22. PUTNAM, FRANK W. 1951. Physiological properties of bacteriophages. II. Sedimentation of bacteriophage 1'6. ]. BioI. Chern. 190:61-74.
2:;. BURNETT, F. M. 1933. The classifica~ion of dysentery-coli baCteriophages. III. A correlation of the serological classification with cerrain biochemical tests. J. Path. and Bacr. 37:179-l84.
64
26. ROUNTREE, PHYI
of sr"ph ylococca
27. FRIJ::OMAN. MUR Stability In mon
28. SY:>"fPOSliI. ON QL ing Company, Ba.
29. SECOND SYMpOSI nature of vu:us

1938. Survival of influ14:447-454.
. C. WIKTERS. 1948. The ;>e sonic vibration. Science
e bacteriophages of Bacillus mical properries. J. Bact.
NER. 1941. Thc effecr of ,I substrate. J. Gen. Physiol.
Academ.ic Press, Inc. New
C Streptococcus lactis hostasurement of the virus. J.
fDEN. 1950. Studies on the hermodesrruction of bactcrial • J. Hyg. 52:194-201.
I. Thcrmal macrivarion of -278-287.
iruses: characteristics of the :387-396.
n of antistaphylococCtls hac
ra-violet-inacrivared bacteri
ce of photO-reactivarion in ried srare. Scicnce 116:425.
y inactivated bacteriophage.
SHARP, E. S. DILLON, T. Z. stabiliry, and sedimenr;lrion
mOluno!. 63 :93-107.
II. Scdi
tery-coli bacteriophages. III. ccrrain biochemical tests. J.
24. AOAJl.IS, MARIe 1949. The stability of baC[erial viruses in solutiom of salt~. J. Gen. Physio!' 32 :579-594 .
25. BURNETT, F. M., AND M-'l\GET McKIE. 1930. Bala'nced salt action as m:2I1J·
fcsted in bacteriophage phenoOlcn.1. Australian J. ExpeL BioI. 'lnd M. Sc. 7: 183200.
26. ROUNTREE, PHYLLIS M. 1951. The role of certain electrolytes in the adsorption of staphj·lococcal bacc.eriophages. J. Gen. Microbiol. 5 :673-680.
27. FRIEDMAN, MURRAY. 1953. The bacteriophages of Bacill1/.S 1J1.egatberjltm. II. Stability in mono- and divalent ions. J. Bact. 66:501-502.
28. SYMPOSIA ON QUA NTlTATIVE BIOLOGY. 1953. Vo!. XVlII Monumemal Publishing Company, Balcimore, Maryland.
29. SECOND SYMPOSIUM OF THE SOCll':n' FOR GENERAL MICROBIOLOGY. 1953. The nature of virus multiplication. University Press, Cambridge, EngLtnd.
65