the effects of neurosteroids on acquisition and retention of a modified passive-avoidance learning...
TRANSCRIPT

Ž .Brain Research 791 1998 108–116
Research report
The effects of neurosteroids on acquisition and retention of a modifiedpassive-avoidance learning task in mice
D.S. Reddy, S.K. Kulkarni )
Department of Pharmacology, UniÕersity Institute of Pharmaceutical Sciences, Panjab UniÕersity, Chandigarh-160 014, India
Accepted 20 January 1998
Abstract
Ž . Ž .This study examined the effects of neurosteroids, pregnenolone sulfate PS and dehydroepiandrosterone sulfate DHEAS , on learningand memory processes in a modified passive-avoidance task in mice. The two parameters measured are number of passive-avoidancestep-down descents and the active escape latency to reach shock-free zone. Each neurosteroid was administered 60 min before orimmediately after the training session, or 60 min before the retention test given 24 h after acquisition. Pretraining injection of PSŽ . Ž .0.125–10 mgrkg, s.c. and DHEAS 0.125–10 mgrkg, s.c. decreased the number of mistakes committed on training day but had no
Ž . Ž .effect on the latency measure. Both PS 0.125–10 mgrkg, s.c. and DHEAS 0.125–10 mgrkg, s.c. decreased the number of mistakesand latency to reach shock-free zone, in a dose-dependent and bell-shaped manner, following pretraining and posttraining administration
Žschedules. Neurosteroids failed to improve memory performance when administered 60 min before retention testing. Injection of PS 0.5. Ž .and 1 mgrkg or DHEAS 1 and 5 mgrkg before both the training and test sessions, however, also significantly facilitated memory
Ž . Ž .retention. In addition, the memory-facilitating effects of PS 0.5 mgrkg, s.c. or DHEAS 1 mgrkg when administered posttraining areŽ .blocked by concurrent administration of haloperidol 0.25 mgrkg, i.p. , a prototype s receptor antagonist. These results confirm that
both PS and DHEAS facilitate retention of a modified learning task when given either pretraining or posttraining, but not prior toretention test. The pretraining neurosteroid-induced memory modulation do not involve state-dependent effects. These results suggest arole for central s receptor in the memory-modulating effects of neurosteroids. q 1998 Elsevier Science B.V.
Keywords: Neurosteroid; Pregnenolone sulfate; Dehydroepiandrosterone sulfate; Learning and memory; s Receptor; Passive-avoidance performance;Haloperidol; Mouse
1. Introduction
Neurosteroids such as allopregnanolone, pregnenolone,Ž .progesterone, pregnenolone sulfate PS and dehy-
Ž .droepiandrosterone sulfate DHEAS are characterized inw xthe central nervous system 14,34 and a role for mitochon-
drial diazepam binding inhibitor receptors in the regulationw xof neurosteroidogenesis proposed 2 . Neurosteroids have
been shown to affect the activity of various neurotrans-mitter systems, including that involved in learning andmemory processes. PS and DHEAS are both allosteric
Ž .antagonists of g-aminobutyric acid type A GABA-Aw xreceptor 16,33 and positive modulators of N-methyl-D-Ž . w xaspartate NMDA receptor-mediated responses 11,40 .
Recently, neurosteroids are reported to inhibit the bindingw3 xŽ . w3 xof s receptor ligands H q -SKF1007 and H halo-
w xperidol 18,21,38,39 and modulate NMDA-evoked
) Corresponding author. Fax: q91-172-541142.
w3 xH norepinephrine from preloaded hippocampal slicesw x25 . Furthermore, increasing evidence indicates that neu-rosteroid sulfates act as agonists at central s receptors andexert a facilitatory action on NMDA-mediated glutamater-
w xgic neurotransmission 20,21 .Pregnenolone sulfate infused into the basal magnocellu-
lar nucleus enhanced memory performance, whereas allo-w xpregnanolone disrupted memory 22 . Immediate posttrain-
ing administration of DHEAS improved retention in aT-maze footshock active avoidance paradigm in both young
w xand aged mice 4,6 . PS markedly enhanced posttrainingmemory processes when injected into cerebral ventriclesw x w x4 and amygdala 5 . Neurosteroids had pronounced acti-vational affects on spatialrreference, working, and long-
w xterm memory in female rats 7 . Neurosteroid sulfates alsocompletely counteracted the amnesic effects of muscariniccholinergic receptor agonists and NMDA receptor antago-nists in animal paradigms of learning and memory pro-
w xcesses 15,17,19 . In many of these studies, however,
0006-8993r98r$19.00 q 1998 Elsevier Science B.V. All rights reserved.Ž .PII S0006-8993 98 00085-7
( )D.S. Reddy, S.K. KulkarnirBrain Research 791 1998 108–116 109
dosages employed were supraphysiological. Furthermore,the use of pretraining amnesic procedures limits the inter-pretation of steroid effects with direct interactions withlearning and memory processes.
Because learning and memory consist of at least threeŽ .stages i.e., acquisition, consolidation and retrieval phases
w x26 , the above findings suggest that neurosteroids modu-late retention performance when administered immediatelyafter training. However, the effects of other administrationschedules of neurosteroids on learning and memory pro-cesses have not been examined. Before attempting todelineate mechanisms of actions, it is necessary to deter-mine the time of administration of exogenous steroidtreatments. Test substances may affect acquisition whenadministered before training. Retention can be altered by
Žtreatments given after learning trials modulate memory. Ž .storage or before retention test affect memory retrieval .
Therefore, in the present study, we minutely examined theŽ .modulatory effects of pretraining acquisition phase , post-
Ž . Žtraining consolidation phase or preretention retrieval.phase administration of neurosteroids PS and DHEAS on
the memory performance in mice. Memory processes wereevaluated using a modified passive-avoidance learning taskthat has been extensively validated in our laboratoryw x26,36 . The putative implication of s receptors was testedby using the prototype s receptor antagonist haloperidolw x13,18,35 .
2. Materials and methods
2.1. Animals
ŽMale albino mice of Laka strain Central Animal House,.Panjab University, Chandigarh , weighing 20–25 g, were
housed five per cage with free access to food and water atthe laboratory conditions. The animals were used follow-ing at least a 4-day period of adaptation to the laboratoryconditions. Mice were only administered drug or vehicleonce. The experiments were carried out between 0900 and1700 h at the ambient temperature. All the experimentshave been conducted according to the NIH Guidelines forthe Care and Use of Laboratory Animals.
2.2. Modified passiÕe-aÕoidance task
Learning and memory processes were evaluated in amodified two-task learning paradigm that has been vali-
w xdated in our laboratory 26,36 . In brief, this is a modifiedversion of the classic passive-avoidance model to assessboth passive and active avoidance behavior in a singletask. The apparatus consisted of a Plexiglass rectangular
Ž . Žbox 22 cm=22 cm=30 cm featuring a grid floor 3.mm stainless-steel rods set 7.5 mm apart , with a wooden
Ž .platform 10 cm=7 cm=1 cm in the center of the grid
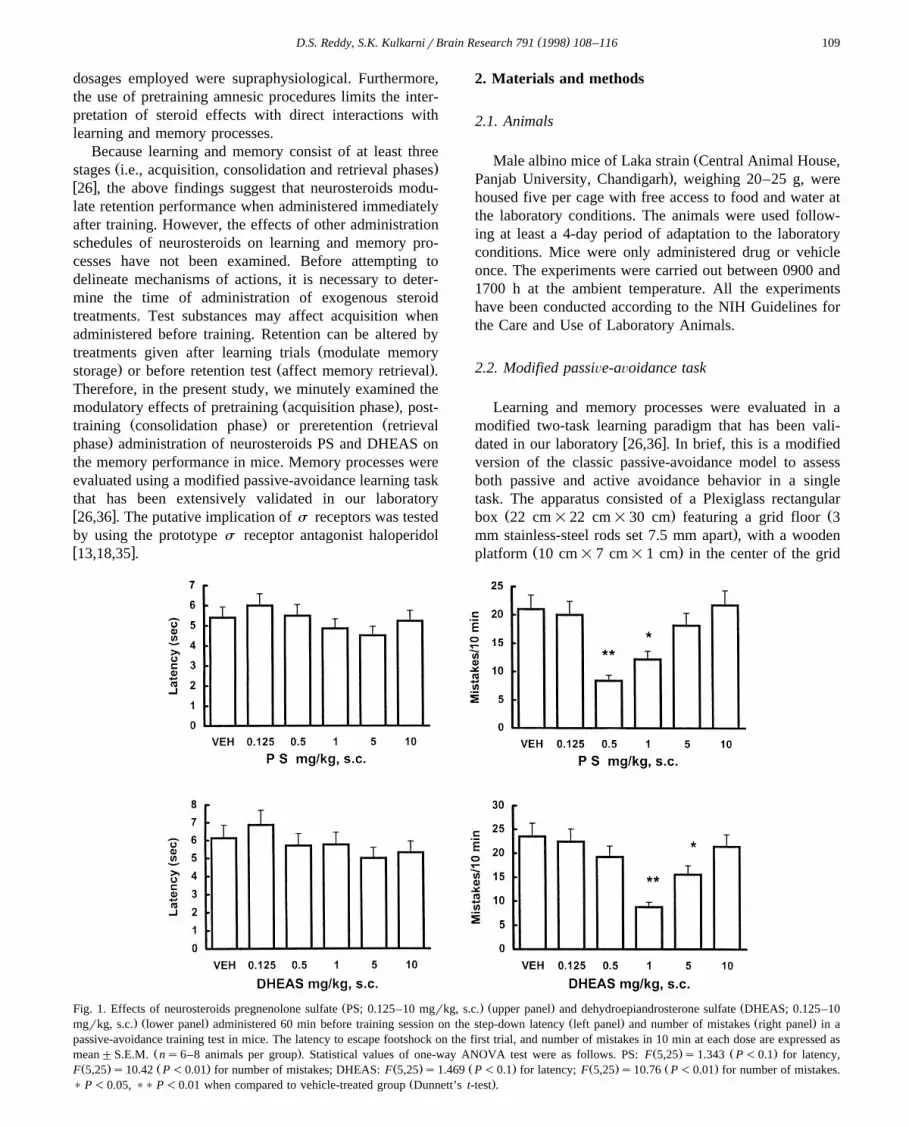
Ž . Ž . ŽFig. 1. Effects of neurosteroids pregnenolone sulfate PS; 0.125–10 mgrkg, s.c. upper panel and dehydroepiandrosterone sulfate DHEAS; 0.125–10. Ž . Ž . Ž .mgrkg, s.c. lower panel administered 60 min before training session on the step-down latency left panel and number of mistakes right panel in a
passive-avoidance training test in mice. The latency to escape footshock on the first trial, and number of mistakes in 10 min at each dose are expressed asŽ . Ž . Ž .mean"S.E.M. ns6–8 animals per group . Statistical values of one-way ANOVA test were as follows. PS: F 5,25 s1.343 P-0.1 for latency,
Ž . Ž . Ž . Ž . Ž . Ž .F 5,25 s10.42 P-0.01 for number of mistakes; DHEAS: F 5,25 s1.469 P-0.1 for latency; F 5,25 s10.76 P-0.01 for number of mistakes.Ž .) P-0.05, )) P-0.01 when compared to vehicle-treated group Dunnett’s t-test .
( )D.S. Reddy, S.K. KulkarnirBrain Research 791 1998 108–116110
Ž .floor. Electric shocks 20 V DC were delivered to the gridfloor with a stimulator. The wooden platform served as
Ž .shock-free zone SFZ during training and retention testingw x36 . A prescreening session and training trial was carriedout under similar conditions. Each mouse was gently placedon the wooden platform and allowed to explore for about 1min. When the mouse stepped off the platform and placedall its paws on the grid floor, a 20-V electric footshockwas given and the latency to reach SFZ was measured.Animals showing latency to reach SFZ within the criterion
Ž .range 5–60 s during the screening session were used for
training and retention tests. The training session was car-ried out 60 min after the prescreening session. Each animalwas placed on the electric grid floor, and the latency to
Ž .reach SFZ and number of mistakes descents made in 10min were recorded. A mistake was counted whenever themouse stepped down from the platform and touched the
Ž .electric grid floor at shock mode 20 V DC . Retentionwas tested 24 h after the training session. Each mouse wasagain placed on the grid floor, and the latency to reachSFZ and the number of mistakes made in 10 min wererecorded.
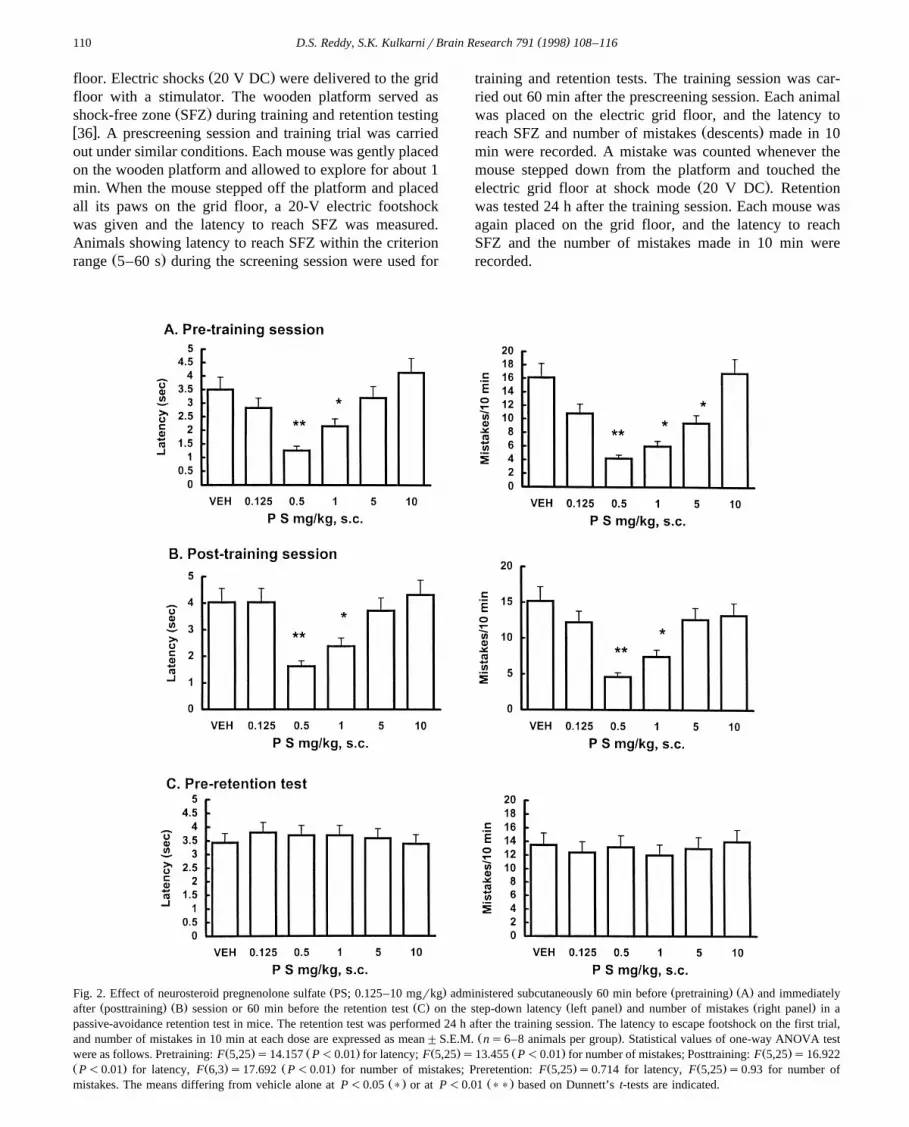
Ž . Ž . Ž .Fig. 2. Effect of neurosteroid pregnenolone sulfate PS; 0.125–10 mgrkg administered subcutaneously 60 min before pretraining A and immediatelyŽ . Ž . Ž . Ž . Ž .after posttraining B session or 60 min before the retention test C on the step-down latency left panel and number of mistakes right panel in a
passive-avoidance retention test in mice. The retention test was performed 24 h after the training session. The latency to escape footshock on the first trial,Ž .and number of mistakes in 10 min at each dose are expressed as mean"S.E.M. ns6–8 animals per group . Statistical values of one-way ANOVA test
Ž . Ž . Ž . Ž . Ž .were as follows. Pretraining: F 5,25 s14.157 P-0.01 for latency; F 5,25 s13.455 P-0.01 for number of mistakes; Posttraining: F 5,25 s16.922Ž . Ž . Ž . Ž . Ž .P-0.01 for latency, F 6,3 s17.692 P-0.01 for number of mistakes; Preretention: F 5,25 s0.714 for latency, F 5,25 s0.93 for number of
Ž . Ž .mistakes. The means differing from vehicle alone at P-0.05 ) or at P-0.01 )) based on Dunnett’s t-tests are indicated.
( )D.S. Reddy, S.K. KulkarnirBrain Research 791 1998 108–116 111
2.3. Drug administration
Ž . ŽDHEAS and PS Sigma, USA and haloperidol Searle,.Skokie, IL, USA were dissolved in 0.1% Tween 80 and
diluted with saline. All drugs were given at a dose of 0.1mlr10 g body weight. Neurosteroids were given following
Žthree administrative schedules i.e., pretraining, posttrain-. Žing and preretention administration . PS 0.125, 0.5, 1, 5
. Žand 10 mgrkg or DHEAS 0.125, 0.5, 1, 5 and 10. Žmgrkg were injected subcutaneously 60 min before pre-. Žtraining or immediately after the training session post-
. Ž .training or 60 min before the retention test preretention ,respectively. To examine the possible involvement of s
receptors in the memory-enhancing effect of neurosteroids,we used haloperidol, the central s receptor antagonistw x Ž .18,20 . Haloperidol 0.25 mgrkg was administered i.p.
Ž . Žsimultaneously with PS 0.5 mgrkg, s.c. or DHEAS 1.mgrkg, s.c. immediately after the training session. In a
Ž .separate group, haloperidol 0.25 mgrkg, i.p. was admin-istered alone immediately after training session. Controlanimals were treated at the corresponding times with vehi-cle instead of drugs.
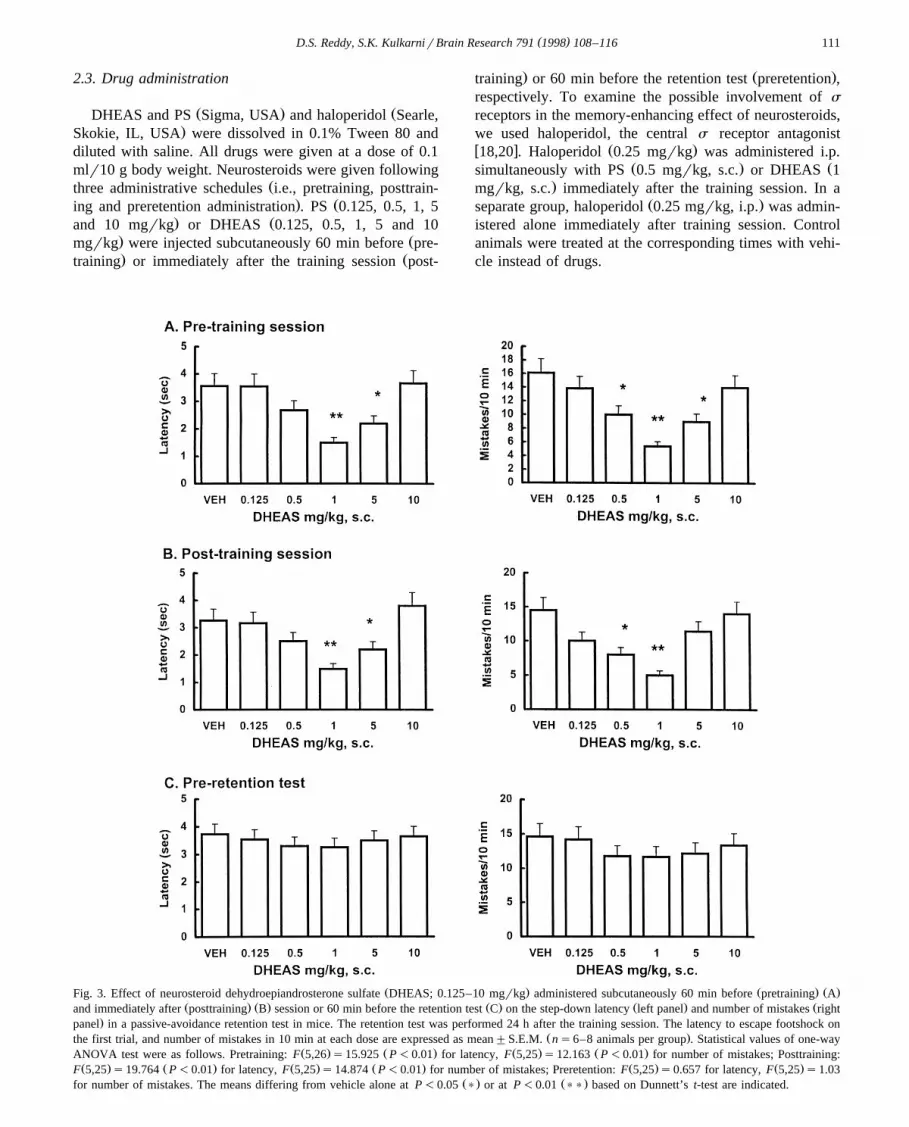
Ž . Ž . Ž .Fig. 3. Effect of neurosteroid dehydroepiandrosterone sulfate DHEAS; 0.125–10 mgrkg administered subcutaneously 60 min before pretraining AŽ . Ž . Ž . Ž . Žand immediately after posttraining B session or 60 min before the retention test C on the step-down latency left panel and number of mistakes right
.panel in a passive-avoidance retention test in mice. The retention test was performed 24 h after the training session. The latency to escape footshock onŽ .the first trial, and number of mistakes in 10 min at each dose are expressed as mean"S.E.M. ns6–8 animals per group . Statistical values of one-way
Ž . Ž . Ž . Ž .ANOVA test were as follows. Pretraining: F 5,26 s15.925 P-0.01 for latency, F 5,25 s12.163 P-0.01 for number of mistakes; Posttraining:Ž . Ž . Ž . Ž . Ž . Ž .F 5,25 s19.764 P-0.01 for latency, F 5,25 s14.874 P-0.01 for number of mistakes; Preretention: F 5,25 s0.657 for latency, F 5,25 s1.03
Ž . Ž .for number of mistakes. The means differing from vehicle alone at P-0.05 ) or at P-0.01 )) based on Dunnett’s t-test are indicated.
( )D.S. Reddy, S.K. KulkarnirBrain Research 791 1998 108–116112
2.4. Statistical analysis
The results are expressed as mean"S.E.M. The pas-sive-avoidance latencies and number of mistakes showed anormal distribution. Consequently, data were analysed us-
Ž .ing one-way analysis of variance ANOVA followed byDunnetts’ t-test or Duncan’s multiple range test. In alltests, P-0.05 was used as the criterion for statisticalsignificance.
3. Results
3.1. Effects of neurosteroids PS and DHEAS on modifiedpassiÕe-aÕoidance training performance
Vehicle-treated mice that received an electric shockduring the training session exhibited a latency of 5.8"0.62s to reach SFZ and showed 21.8"1.68 mistakes in 10 mintest period on the modified passive-avoidance task. As
Ž .shown in Fig. 1, PS 0.125–10 mgrkg, s.c. or DHEASŽ .0.125–10 mgrkg, s.c. when administered 60 min beforethe training session did not affect the latency to SFZ whencompared to vehicle controls. The s.c. pretraining adminis-
tration of PS or DHEAS, in the 0.125–10 mgrkg doserange, significantly decreased the number of mistakes intraining session when compared to vehicle controls. Vehi-cle control and animals assigned into neurosteroid post-, orpreretention groups did not differ in the mean latency andnumber of mistakes in the modified passive-avoidance
Ž .training session data not shown .
3.2. Effects of neurosteroids PS and DHEAS on modifiedpassiÕe-aÕoidance retention performance
Vehicle-treated mice showed a latency of 3.5"0.32 sand made 16.1"1.57 mistakes in the passive-avoidance
Žretention test. Fig. 2 shows that PS 0.125–10 mgrkg,.s.c. -treated mice, in pretraining or posttraining administra-
tion schedule, required shorter latencies to escape fromfootshock than vehicle-treated group. The number of mis-
Žtakes made by mice administered PS 0.125–10 mgrkg,.s.c. 60 min before or immediately after the training
sessions were significantly low relative to vehicle controlsŽ .Fig. 2 . Maximum modulation of retention test perfor-mance in pretraining or posttraining administration of PSwas observed at a dose of 0.5 mgrkg. On the other hand,
Ž .PS 0.125–10 mgrkg, s.c. did not affect either latency to
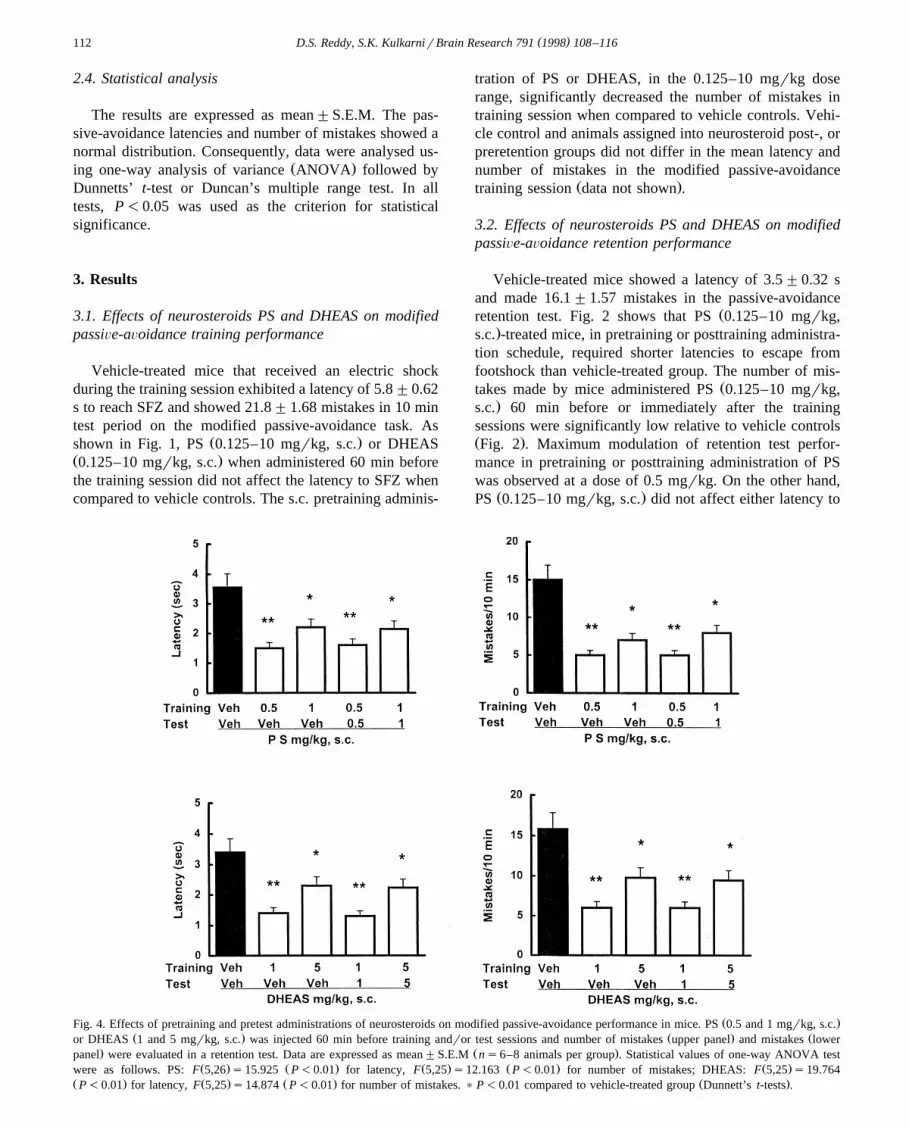
Ž .Fig. 4. Effects of pretraining and pretest administrations of neurosteroids on modified passive-avoidance performance in mice. PS 0.5 and 1 mgrkg, s.c.Ž . Ž . Žor DHEAS 1 and 5 mgrkg, s.c. was injected 60 min before training andror test sessions and number of mistakes upper panel and mistakes lower
. Ž .panel were evaluated in a retention test. Data are expressed as mean"S.E.M ns6–8 animals per group . Statistical values of one-way ANOVA testŽ . Ž . Ž . Ž . Ž .were as follows. PS: F 5,26 s15.925 P-0.01 for latency, F 5,25 s12.163 P-0.01 for number of mistakes; DHEAS: F 5,25 s19.764
Ž . Ž . Ž . Ž .P-0.01 for latency, F 5,25 s14.874 P-0.01 for number of mistakes. ) P-0.01 compared to vehicle-treated group Dunnett’s t-tests .
( )D.S. Reddy, S.K. KulkarnirBrain Research 791 1998 108–116 113
reach SFZ or number of mistakes on the modifiedpassive-avoidance retention test when administered in pre-
Žretention schedule as compared to vehicle controls Fig. 2,. Ž .lower panel . DHEAS 0.125–10 mgrkg, s.c. dose-de-
pendently reduced the latency to reach SFZ and number ofmistakes in the retention test when administered in pre-
Ž .training and posttraining schedules Fig. 3 . Maximal en-hancement of retention test performance was obtained with1 mgrkg DHEAS in pretraining or posttraining adminis-tration schedule. However, preretention test administrationof DHEAS, in the 0.125–10 mgrkg dose range, did notaffect the latency to reach SFZ when compared to vehiclecontrols, nor the number of mistakes on the modified
Ž .passive-avoidance retention test Fig. 3, lower panel .
3.3. Effects of neurosteroids PS and DHEAS on state-de-pendent learning on modified passiÕe-aÕoidance perfor-mance
Mice that received a training session on a task undersome drugs often show a failure of learning performance
w xwhen they are tested in the absence of the drugs 26 . Inorder to determine such state-dependent effects, separategroups of mice received the neurosteroids both beforetraining as well as before testing. As shown in Fig. 4,
Ž . Žadministration of PS 0.5 and 1 mgrkg or DHEAS 1 and.5 mgrkg 60 min before both training and test sessions
also significantly reduced the latency to reach SFZ anddecreased the number of mistakes as compared to vehicle-treated controls, indicating that state-dependent learningdoes not contribute to the memory-facilitating effects ofneurosteroids in the pretraining group.
3.4. Influence of haloperidol on the neurosteroid-inducedenhancement of modified passiÕe-aÕoidance retention per-formance
In order to explore whether the memory enhancingeffects of neurosteroids involve central s receptor, theeffect of haloperidol, a s receptor antagonist, was studied.
Ž . Ž .The PS 0.5 mgrkg, s.c. -, or DHEAS 1 mgrkg, s.c. -in-duced facilitation of retention of the modified passive-
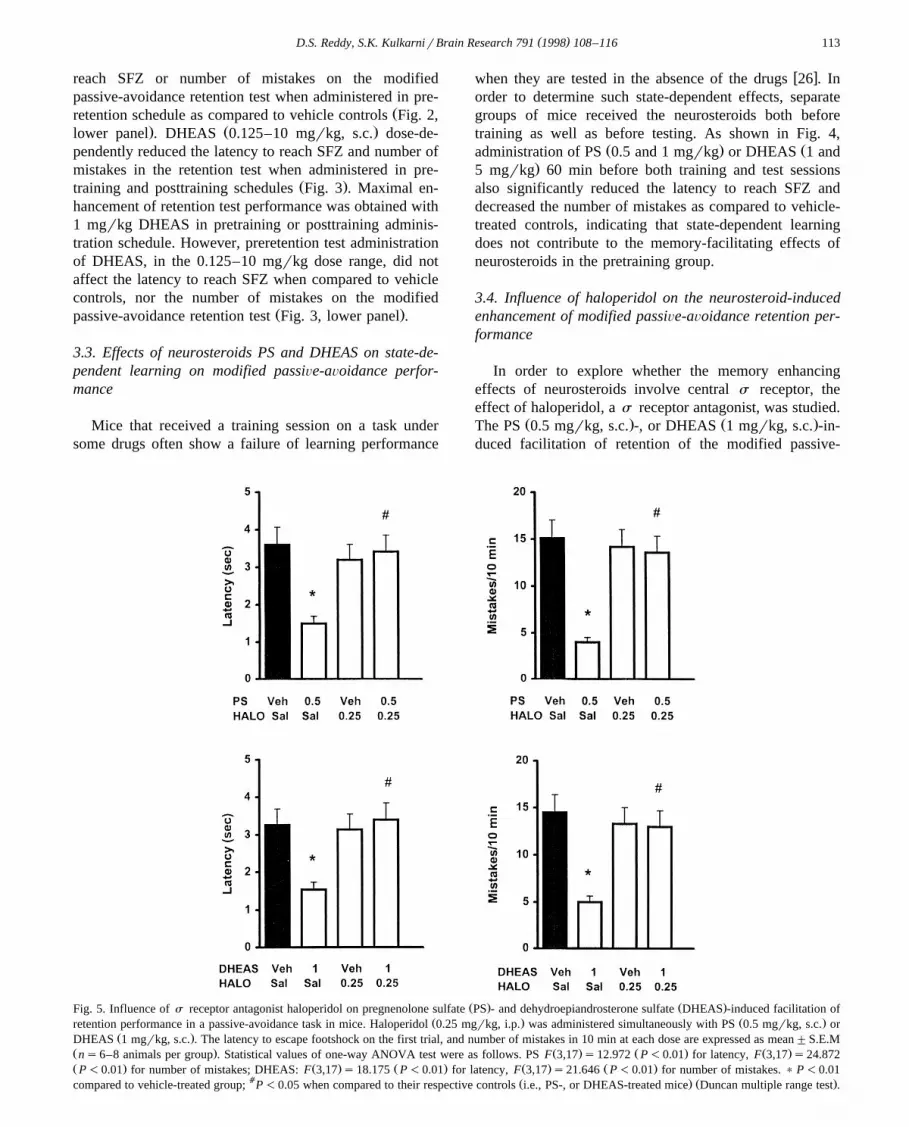
Ž . Ž .Fig. 5. Influence of s receptor antagonist haloperidol on pregnenolone sulfate PS - and dehydroepiandrosterone sulfate DHEAS -induced facilitation ofŽ . Ž .retention performance in a passive-avoidance task in mice. Haloperidol 0.25 mgrkg, i.p. was administered simultaneously with PS 0.5 mgrkg, s.c. or
Ž .DHEAS 1 mgrkg, s.c. . The latency to escape footshock on the first trial, and number of mistakes in 10 min at each dose are expressed as mean"S.E.MŽ . Ž . Ž . Ž .ns6–8 animals per group . Statistical values of one-way ANOVA test were as follows. PS F 3,17 s12.972 P-0.01 for latency, F 3,17 s24.872Ž . Ž . Ž . Ž . Ž .P-0.01 for number of mistakes; DHEAS: F 3,17 s18.175 P-0.01 for latency, F 3,17 s21.646 P-0.01 for number of mistakes. ) P-0.01
a Ž . Ž .compared to vehicle-treated group; P-0.05 when compared to their respective controls i.e., PS-, or DHEAS-treated mice Duncan multiple range test .

( )D.S. Reddy, S.K. KulkarnirBrain Research 791 1998 108–116114
avoidance behavior when administered immediately afterthe training session was significantly antagonized by the
Žconcurrent administration of haloperidol 0.25 mgrkg,. Ž . Ž .s.c. Fig. 5 . Haloperidol 0.25 mgrkg, s.c. alone had no
effect on the latency to reach SFZ or number of mistakesin the retention test when administered immediately after
Ž .the training session Fig. 5 .
4. Discussion
The present study was aimed to explore the effects ofneurosteroids in yet another test of learning and memory,the modified passive-avoidance paradigm. The learning
w xparadigm used in the present study 26,36 is a modifiedversion of the classic passive-avoidance model where thestep-down latency has been indirectly assessed by measur-
Ž .ing the number of descents mistakes made by the animalŽby stepping down from the wooden platform shock-free
. Ž .zone to the grid shock zone in a 10-min test period.Hence the number of descents is a parameter of negativereinforcement behavior. In addition, it also estimates anactive-avoidance behavior in the form of latency to reachshock-free zone. According to our training procedures, amouse made one of two types of responses in this taskduring the first and second training and retention testing. Aresponse latency longer than 5 s was classed as an escape
Žfrom the footshock occurs only during screening and.training sessions . A response latency less than or equal to
5 s was considered an avoidance, since the mouse avoidedreceiving a footshock for a considerable duration. Thus,we also applied exclusion criteria to reduce learning vari-ability among mice, as mice with escape latencies greaterthan 60 s and less than 5 s were discarded from the study.Thus, the term ‘modified passive-avoidance task’ can bejustifiably used to describe the model.
In the present study, training was carried out in twodifferent sessions. In actuality, the screening represents thefirst training and a second training was performed 60 minafter the first, and retention was examined 24 h after thesecond training. The latency to reach shock-free zonebetween different groups during the first training sessionwas similar. During the second training session, the vehi-cle- and neurosteroid-treated groups exhibited a significant
Ž .decrease in latency data not shown , indicating that themice acquired the task and thus avoided receiving shocks.However, this only implies the registration of the memoryprocesses, but is too less a time for the consolidationprocess to take place to consider the second training as aretention test. Further, the possible conditioned fear-likeeffects or changes in pain perception may confound thebehavior on second training. However, this is unlikelybecause of the absence of any conditioned fear such a lightor sound, and that neurosteroids do not affect pain sensitiv-
w xity 30 .
The present study demonstrated that the pretraining orposttraining session administration of neurosteroids PS andDHEAS significantly facilitated retention, while prereten-tion administration was generally without any effect on themodified passive-avoidance retention performance. Thepretraining neurosteroid-induced memory modulation donot involve a state-dependent effects. In addition, neuros-teroid-induced facilitation of memory retention may in-volve central s receptors, because the effect of neuros-teroids was completely antagonized by s receptor antago-nist haloperidol.
The design of the present experiment permitted to ana-lyze the direct effect of neurosteroids on acquisition, mem-ory consolidation or retrieval processes. The effects ofpretraining administration of neurosteroids on the acquisi-tion of the passive-avoidance response depended on theparameter evaluated. Neurosteroids significantly decreasedthe number of mistakes, a parameter of negative reinforce-ment behavior, while latency to reach SFZ, an activeescape response, was not affected by either steroids. How-ever it is difficult to directly compare our results to other
w xstudies in the field 4,7 , because of the different treatmentschedules andror learning tasks used. For example, neu-rosteroids have activational effects on short-term andlong-term memory processes in the Morris water-maze and
w x w xY-maze tests 7 . However, Flood et al. 4 has found noevidence for an improved acquisition of the T-maze foot-shock avoidance task when DHEAS was given for 2 weeksbefore training. Perhaps the important distinction is thedifferent behavior task or acute vs. chronic neurosteroidadministration protocol. Extended administration ofDHEAS has recently been shown to not only increasecirculating levels of DHEAS, but also of DHEA, which
w xhas opposite actions on learning and memory processes 3 ,so these effects may have canceled out one another in the
w xmore prolonged administration study of Flood et al. 4 .Further, long-term administration of neurosteroids pro-
w xduces changes in neuronal structure and function 27,31 .Although the data in the present study suggest that pre-training administration neurosteroids may at least partlyaffect the acquisition of the modified learning task, theprocedural differences raise concern in the interpretation ofneurosteroids’ effects on acquisition.
In the present study, pretraining and posttraining admin-istration of PS and DHEAS in doses 25–50 mgrmouseŽ .0.5–1 mgrkg that corresponds to 0.05–0.2 mM en-hanced memory retention of passive-avoidance training. Inboth treatments, an inverted U dose–response curve wasobtained covering 2- to 5-fold dose range in a mannertypical for memory-enhancing substances. However, prere-tention administration of either neurosteroid did not resultin any improvement in retention of the passive-avoidancetraining. This finding supports the time-dependency ofpostacquisition interventions in altering memory storage or
w xretrieval processes 23 . Enhancement of retention was alsofound to be facilitated by immediate or within 60 min

( )D.S. Reddy, S.K. KulkarnirBrain Research 791 1998 108–116 115
posttraining administration of pregnenolone and itsw xmetabolites 4,12 . Although preretention administration of
excitatory agents amphetamine and epinephrine enhancedw xretention of a passive-avoidance task 37 , many estab-
lished memory-enhancing agents failed to facilitate reten-tion when given before retention test or 3–24 h after
w xtraining with retention testing occurring 3 days after 24 .In the present study, we found that there was no state-de-pendent effect in mice injected with both neurosteroids onsubsequent retention. Thus, the pretraining neurosteroid-in-duced facilitation of memory processes are not confoundedby their effects on state-dependent learning.
The exact mechanism, and even specificity, of thememory enhancing effect of neurosteroids is currently notknown. It is thought that steroid hormones bind to specificreceptors in the cytoplasm, which translocate into thenucleus and finally regulate gene transcription or affecttranslational efficiency and protein stability, which arepostulated to be involved in long-lasting learning andmemory storage. Alternately, it has been hypothesized that
Ž .a cell membrane receptor mechanism s may also involvein the effects of neurosteroids in learning and memoryprocesses. Neurosteroids have been shown to affect theactivity of neurotransmitter systems, which are involved inthe learning and memory processes. PS and DHEAS arenegative allosteric modulators of the GABA-A receptorw x18,33 and positive modulators of NMDA receptor-media-
w xted responses 11,40 . Recently, increasing evidence indi-cate that these neurosteroids acts as agonists at central s
receptors and exerts a facilitatory action on NMDA-media-w xted glutamatergic neurotransmission 20 .
In the present study, neurosteroid effects may involve atleast partly an interaction with s receptors, since it couldbe significantly antagonized by a simultaneous administra-tion of the s receptor antagonist, haloperidol. In supportof these findings, haloperidol has previously been reported
w xto block s receptors 13 , antagonize s receptor agonistŽ .q -SKF-10,047-induced activation of the central cholin-
w xergic system 18 , and prevent the attenuating effect of s
receptor agonists in learning impairment induced by scopo-w xlamine or dizocilpine 19,20 . Apart from s receptors,
haloperidol has been reported to interact with dopamine D2w xreceptor 13 . Therefore, there is a possibility that the
memory-facilitating effect of PS or DHEAS is involved inthe D dopaminergic system. However, dopamine D re-2 2
Ž .ceptor agonists such as y -apomorphine and RU-24213w xinduce memory impairment 9,10 , while dopamine D2
Ž .receptor antagonist y -sulpiride enhances passive-avoi-w xdance learning 9 , which suggests that the activation of the
dopamine D receptor results in memory dysfunction.2
However, in the present study, haloperidol treatment alonedid not affect the passive-avoidance retention, but signifi-cantly antagonized the memory-enhancing effect of PS andDHEAS. Moreover, binding studies showed that haloperi-dol bound to both s and s receptor subtypes with the1 2
w xsame potency 1,8 . Neurosteroid-induced memory-enhanc-
ing effect may be mediated by s receptor, because the1
effect of PS and DHEAS was antagonized by NE-100, aw x Žselective s receptor antagonist 1 unpublished observa-1
.tions . Taken together, these data indicate a role for s
receptors in the memory-facilitating effects of neuros-teroids.
In addition, it could be possible that administration ofneurosteroids modulated diverse behaviors that might af-fect performance in passive-avoidance situations. To pre-vent this ambiguity in the interpretation of results, steroideffects on learning and memory processes need to bedissociated from other behaviors. We previously showedthat administration of PS and DHEAS, at doses that im-proved memory performance in the present study, do notresult in significant differences in spontaneous locomotor
wactivity, rota-rod performance, anxiety behavior 27–x w x29,31,32 or pain sensitivity 30 when compared to vehi-
cle controls. These data, taken together with the lack ofactivational effects in preretention administration schedule,indicate that the effects of neurosteroids in the passive-avoidance task are memory specific, and not secondary toother behavior.
In conclusion, the present study confirms the modulat-ing role of neurosteroids in learning and memory processesand showed that neurosteroids, PS and DHEAS, facilitatedthe modified passive-avoidance memory retention withoutaffecting retrieval phase. In addition, this study suggest arole for central s receptors in the memory-modulatingeffect of neurosteroids. Further studies would confirmthese convergent signaling mechanisms in the neurosteroidmodulation of learning and memory processes.
References
w x1 S. Chaki, M. Tanaka, M. Muramatsu, S. Otomo, NE-100, a novelpotent s ligand, preferentially binds to s binding sites in guinea1
Ž .pig brain, Eur. J. Pharmacol. 251 1994 R1–R2.w x2 E. Costa, J. Auta, A. Guidotti, A. Korneyev, E. Romeo, The
pharmacology of neurosteroidogenesis, J. Steroid Biochem. Mol.Ž .Biol. 49 1994 385–389.
w x Ž .3 Fleshner et al., Behav. Neurosci. 111 1997 512–517.w x4 J.F. Flood, J.E. Morley, E. Roberts, Memory-enhancing effects in
male mice of pregnenolone and steroids metabolically derived fromŽ .it, Proc. Natl. Acad. Sci. U.S.A. 89 1992 1567–1571.
w x5 J.F. Flood, J.E. Morley, E. Roberts, Pregnenolone sulfate enhancesposttraining memory processes when injected in very low doses intolimbic system structures: The amygdala is by far the most sensitive,
Ž .Proc. Natl. Acad. Sci. U.S.A. 92 1995 10806–10810.w x6 J.F. Flood, E. Roberts, Dehydroepiandrosterone sulfate improves
Ž .memory in aging mice, Brain Res. 448 1988 178–181.w x7 C.A. Frye, J.D. Sturgis, Neurosteroids affect spatialrreference,
working, and long-term memory of female rats, Neurobiol. Learn.Ž .Mem. 64 1995 83–96.
w x8 S.B. Hellewell, W.D. Bowen, A sigma-like binding site in ratŽ . Ž .pheochromocytoma PC12 cells: decreased affinity for q -benzo-
morphans and lower molecular weight suggest a different sigmaŽ .receptor form from that of guinea pig brain, Brain Res. 527 1990
244–253.w x9 K. Ichihara, T. Nabeshima, T. Kameyama, Opposite effects induced

( )D.S. Reddy, S.K. KulkarnirBrain Research 791 1998 108–116116
by low and high doses of apomorphine on single-trial passive-avoi-Ž .dance learning in mice, Pharmacol. Biochem. Behav. 30 1988
107–113.w x10 K. Ichihara, T. Nabeshima, T. Kameyama, Effects of dopamine
receptor agonists on passive-avoidance learning in mice: interactionŽ .of dopamine D and D receptors, Eur. J. Pharmacol. 213 19921 2
243–250.w x11 R.P. Irwin, S.Z. Lin, M.A. Rogawski, R.H. Purdy, S.M. Paul,
Steroid potentiation and inhibition of N-methyl-D-aspartatereceptor-mediated intracellular Ca2q responses: structure-activity
Ž .studies, J. Pharmacol. Exp. Ther. 271 1994 677–682.w x12 R.C. Isaacson, J.A. Varner, J.-M. Baars, D. de Wied, The effects of
pregnenolone sulfate and ethylestrenol on retention of a passive-Ž .avoidance task, Brain Res. 689 1995 79–84.
w x13 Y. Itzhak, S. Alerhand, Differential regulation of s and PCPreceptors after chronic administration of haloperidol and phencycli-
Ž .dine in mice, FASEB J. 3 1989 1868–1872.w x14 S.K. Kulkarni, D.S. Reddy, Neurosteroids: A new class of neuro-
Ž .modulators, Drugs Today 31 1995 433–455.w x15 P.-K. Li, M.E. Rhodes, S. Jagannathan, D.A. Johnson, Reversal of
scopolamine induced amnesia in rats by the steroid sulfatase in-Ž .hibitor estrone-3-O-sulfamate, Cogn. Brain. Res. 2 1995 251–254.
w x16 M.D. Majewska, Neurosteroids: Endogenous bimodal modulators ofthe GABA-A receptor. Mechanism of action and physiological
Ž .significance, Prog. Neurobiol. 38 1992 379–395.w x17 C. Mathis, S.M. Paul, J.N. Crawley, The neurosteroid pregnenolone
sulfate blocks NMDA antagonist-induced deficits in a passive-avoi-Ž .dance memory task, Psychopharmacology 11 1994 201–206.
w x18 K. Matsuno, K. Matsunaga, T. Senda, S. Mita, Increase in extracel-lular acetylcholine level by sigma ligands in rat frontal cortex, J.
Ž .Pharmacol. Exp. Ther. 265 1993 851–859.w x19 T. Maurice, J.-L. Junien, A. Privat, Dehydroepiandrosterone sulfate
attenuates dizocilpine-induced learning impairment in mice via s -1Ž .receptors, Behav. Brain Res. 83 1997 159–164.
w x20 T. Maurice, B.P. Lockhart, Neuroprotective and anti-amnesic poten-Ž .tials of sigma s receptor ligands, Prog. Neuro-Psychopharmacol.
Ž .Biol. Psychiatry 21 1997 69–102.w x21 T. Maurice, F.J. Roman, A. Privat, Modulation by neurosteroids of
Ž . w3 xthe in vivo q - H SKF-10,047 binding to s receptors in the1Ž .mouse forebrain, J. Neurosci. Res. 46 1996 734–743.
w x22 W. Mayo, F. Dellu, P. Robel, J. Cherkaoui, M. Le Moal, E.-E.Baulieu, H. Simon, Infusion of neurosteroids into the nucleus basalismagnocellularis affects cognitive processes in the rat, Brain Res. 607Ž .1993 324–328.
w x23 J.L. McGough, Preserving the presence of the past: HormonalŽ .influences on memory storage, Am. Psychologist 38 1983 161–174.
w x24 C. Mondadori, T. Ducret, J. Borkowski, How long does ‘memoryconsolidation’ take? New compounds can improve retention perfor-mance, even if administered upto 24 hours after the learning experi-
Ž .ence, Brain Res. 555 1991 107–111.
w x25 F.P. Monnet, V. Mahe, P. Robel, E.E. Baulieu, Neurosteroids, via s
w3 xreceptors, modulate the H norepinephrine release evoked by N-methyl-D-aspartate in the rat hippocampus, Proc. Natl. Acad. Sci.
Ž .U.S.A. 92 1995 3774–3778.w x26 D.S. Reddy, Assessment of nootropic and amnestic activity of
Ž .centrally acting agents, Indian J. Pharmacol. 29 1997 208–221.w x27 D.S. Reddy, S.K. Kulkarni, Role of GABA-A and mitochondrial
diazepam binding inhibitor receptors in the antistress activity ofŽ .neurosteroids in mice, Psychopharmacology 128 1996 280–292.
w x28 D.S. Reddy, S.K. Kulkarni, Differential anxiolytic effects of neuros-teroids in the mirrored chamber behavior test in mice, Brain Res.
Ž .752 1997 61–71.w x29 D.S. Reddy, S.K. Kulkarni, Neurosteroid coadministration prevents
development of tolerance and augments recovery from benzodi-azepine withdrawal anxiety and hyperactivity in mice, Methods
Ž .Find. Exp. Clin. Pharmacol. 19 1997 395–405.w x30 D.S. Reddy, S.K. Kulkarni, Chronic neurosteroid treatment prevents
development of morphine tolerance and attenuates abstinence behav-Ž .ior in mice, Eur. J. Pharmacol. 337 1997 19–25.
w x31 D.S. Reddy, S.K. Kulkarni, Neuroprotective effects of neurosteroidsagainst hypoxic stress-induced neurotoxicity in naive and FG 7142-
Ž .treated mice, Indian J. Pharmacol. 29 1997 381–392.w x32 D.S. Reddy, S.K. Kulkarni, Reversal of benzodiazepine inverse
agonist FG 7142-induced anxiety syndrome by neurosteroids inŽ . Ž .mice, Methods Find. Exp. Clin. Pharmacol. 19 1997 in press .
w x33 E. Roberts, Pregnenolone: From Selye to Alzheimer and a model ofthe pregnenolone sulfate binding site on the GABA-A receptor,
Ž .Biochem. Pharmacol. 49 1995 1–16.w x34 M. Schumacher, P. Robel, E.-E. Baulieu, Development and regener-
ation of the nervous system: A role for neurosteroids, Dev. Neurosci.Ž .18 1996 6–21.
w x35 T. Senda, M. Kiyoshi, T. Kobayashi, S. Mita, Reduction of thescopolamine-induced impairment of passive-avoidance performance
Ž .by s receptor agonist in mice, Physiol. Behav. 61 1997 257–264.w x36 A.C. Sharma, S.K. Kulkarni, Effects of MK-801 and Ketamine on
short-term memory deficits in passive-avoidance step-down taskŽ .paradigm in mice, Methods Find. Exp. Clin. Pharmacol. 13 1991
155–159.w x37 W.S. Stone, R.J. Rudd, P.E. Gold, Amphetamine, epinephrine and
Ž .glucose enhancement of memory retrieval, Psychobiology 18 1990227–230.
w x38 T.P. Su, E.D. London, J.H. Jaffe, Steroid binding at s receptorssuggests a link between endocrine, nervous and immune systems,
Ž .Science 240 1988 219–221.w x39 J.M. Walker, W.D. Bowen, F.O. Walker, R.R. Matsumoto, B. De
Costa, K.C. Rice, Sigma receptors: biology and function, Pharmacol.Ž .Rev. 42 1990 355–402.
w x40 F.S. Wu, T.T. Gibbs, D.H. Farb, Pregnenolone sulfate: a positiveallosteric modulator at the N-methyl-D-aspartate receptor, Mol.
Ž .Pharmacol. 40 1991 333–336.