the effect of ammonium on photosynthesis and the pathway of ammonium assimilation in gymnodinium...
TRANSCRIPT

The Effect of Ammonium on Photosynthesis and the Pathway of Ammonium Assimilation inGymnodinium microadriaticum in vitro and in Symbiosis with Tridacnid Clams and CoralsAuthor(s): R. E. Summons, T. S. Boag and C. B. OsmondSource: Proceedings of the Royal Society of London. Series B, Biological Sciences, Vol. 227, No.1247 (Mar. 22, 1986), pp. 147-159Published by: The Royal SocietyStable URL: http://www.jstor.org/stable/36125 .
Accessed: 01/05/2014 09:54
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The Royal Society is collaborating with JSTOR to digitize, preserve and extend access to Proceedings of theRoyal Society of London. Series B, Biological Sciences.
http://www.jstor.org
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions

Proc. R. Soc. Lond. B 227, 147-159 (1986) Printed in Great Britain
The effect of ammonium on photosynthesis and the pathway of ammonium assimilation in Gymnodinium
microadriaticum in vitro and in symbiosis with tridacnid clams and corals
BY R. E. SUMMONSl t, T. S. BOAG2 AND C. B. OSMOND, F.R.S.1
1Department of Environmental Biology, Research School of Biological Sciences, Australian National University, P.O. Box 475, Canberra City,
A.C.T. 2601, Australia 2 Sandy Trout Laboratory, Queensland Department of Agriculture, Hamilton,
Queensland 4007, Australia
(Received 16 July 1985)
The effects of added ammonium ion (10-1000 1M) on photosynthetic 14CO2 fixation by tissues slices from the mantle of tridacnid clams, by coral tips, and by isolated zooxanthellae from clam mantle were examined. Ammonium ions stimulated photosynthesis in tissue slices but not in freshly isolated zooxanthellae. When ammonium stimulated 14CO2 fixation by coral tips an increase in water-soluble labelled compounds, especially amino acids, was observed. Even though ammonium ions did not stimulate photosynthesis in isolated zooxanthellae from clam mantle, light stimulated ammonium uptake in these cells. Studies with 15NH+ confirmed earlier observations (in zooxanthellae isolated from Hippopus) of light-stimulated transfer of ammonium from the amido-N of glutamine to the amino-N of glutamate, glutamine and other amino acids. This observation, in isolated zooxanthellae and tissue slices, suggests that the glutamine synthase-glutamate synthetase pathway of ammonium incorporation is light-driven in these systems. The possible significance of these processes during ammonium recycling by symbiotic algae in marine animals is discussed.
INTRODUCTION
Tropical marine waters contain low concentrations of available nitrogen and phosphorus which are thought to limit productivity (Thomas 1970). Coral reef ecosystems are characterized by the presence offree-living N2-fixing microorganisms (Weibe et al. 1975; Webb et al. 1975; Burris 1976; Wilkinson et al. 1984) and by symbiotic systems in which nitrogen recycling between animal and algae is thought to be a central feature of nutrient conservation (Cates & McLaughlin 1976; Muscatine I980). Although there has been a good deal of emphasis on the carbon exchanges between animal and algae in these symbioses (Lewis & Smith 1971; Trench 1979), very little is known of the biochemical pathways of nitrogen cycling in these systems.
t Present address: Baas Becking Geobiological Laboratory, Bureau of Mineral Resources, G.P.O. Box 378, Canberra City, A.C.T. 2601, Australia.
6 [ 147 ] Vol. 227 B (22 March I986)
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions

R. E. Summons, T. S. Boag and C. B. Osmond
It is clear that symbiotic zooxanthellae are important in ammonium uptake and retention from sea water (Muscatine & D'Elia 1978; Muscatine et al. 1979). In corals the absorption of ammonium is light-dependent (Muscatine & D'Elia 1978) and in the anemone Aiptasia pulchella, ammonium uptake is reduced by prolonged dark pretreatment (Wilkerson & Muscatine 1984).
These experiments suggest that ammonium uptake and incorporation in symbi- oses may either be light-dependent processes, or indirectly require the provision of carbon acceptors from photosynthetic metabolism for ammonium incorporation. There is very little evidence of direct dependence on light, beyond our preliminary studies with Gymnodinium microadriaticum from the mantle of the clam Hippopus hippopus (Summons & Osmond 1981). We found that the transfer of 15N from the most heavily labelled amino compound (the amido-N of glutamine) to the amino-N of glutamate, glutamine and other amino acids was light-stimulated. This is quite specifically indicative of the involvement of the enzyme glutamate synthase which, in many other photosynthetic systems, is driven by photochemically derived reduced ferredoxin (Lea & Miflin 1981). In non-autotrophic systems this enzyme can function with NADPH. Interestingly, Wilkerson & Muscatine (1984) were unable to demonstrate the presence of the NADPII-dependent enzyme in extracts of zooxanthellae isolated from Aiptasia.
If indirect light stimulation of ammonium uptake was involved, then we would expect that photosynthesis in the symbiotic zooxanthellae might be stimulated by ammonium ions. Once again there are very few data and these tend to be contradictory. Taylor (1978) observed that ammonium and urea stimulated photosynthesis by the coral Acropora cervicornis and by the isolated zooxanthellae, but only when the latter were growing in log phase. On the other hand Goldman et al. (1981) found no effect of ammonium ion on the rate of photosynthesis of ammonium limited phytoplankton cultures.
In this report we have addressed both aspects of the light-dependence of ammonium uptake in another symbiotic system, the mantle of the clam Tridacna. We have also sought to extend these approaches to several species of coral.
MATERIALS AND METHODS
Specimens of T. maxima (15-20 cm long), T. derasa (40-50 cm long) and T. squamosa (20-25 cm long) were collected at Davies Reef 147? 36' E, 18? 51' S) and transferred to a flowing seawater aquarium at the Australian Institute of Marine Sciences (Townsville), where they were maintained for several weeks before experiments were conducted. Experiments were repeated in 1981, 1982 and 1984 with consistent results.
The mantle was dissected free from the animal and adhering muscle tissue. The outer margin, containing the most dense populations of zooxanthellae, was then carefully dissected into small tissue slices (equivalent to 40-50 !tg chlorophyll a). These tissue slices were subsampled and maintained in flowing seawater for up to 30 min before experiments began. They were then rinsed in Millipore-filtered (0.45 gjm) seawater and transferred to the incubation mixtures. All procedures were carried out at room temperature under low illumination.
148
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions

Clam and coral photosynthetic NH+ assimilation
Zooxanthellae were isolated from the dissected mantle tissue by homogenizing 30 g mantle in 300 ml filtered seawater with a Vitris blender at 70 % line voltage. The homogenate was filtered through several layers of Miracloth and the animal tissue discarded. The filtrate was centrifuged (2000 g for 2 min) and the supernatant discarded. The pellet containing algal cells was washed four times with fresh filtered seawater by centrifugation and with subsequent resuspension. The pellet was then resuspended in 250 ml of seawater, kept at room temperature, and samples of this suspension were used for determinations of photosynthetic 14C incorporation, net 02 evolution and 15N assimilation.
Experiments with the corals Stylophora pistillata, Pocillopora damicornis and Acropora millipora were carried out in the field: either at Davies Reef, or at the Heron Island Research Station, as specified. Coral specimens were collected from a depth of 3-5 m and immediately taken to the field laboratory in shaded aquaria. Uniform samples of apical tips (equivalent to 50-100 ig chlorophyll a) were excised just before experimental incubation. Care was taken to discard coral exhibiting production of excessive mucilage, or bleaching. Experiments were repeated in 1981 and 1984, with inconsistent results.
Bicarbonate-dependent, photosynthetic 02 evolution in the light, and respiratory 02 uptake in the dark, were measured with a Hansatech 02 electrode. Cells or tissue slices were illuminated with a projector lamp and the light filtered through the water jacket of the 02 electrode chamber. Incident photosynthetically active radiation (400-700 nm) was measured with a Lambda quantum sensor.
Light-dependent 14C02 fixation was measured by incubating tissue slices, coral tips, or isolated cells with gentle shaking in labelled seawater (0.4 Ci mol-1, pH 8.2) at light saturation in sunlight (1500-2000 tmol quanta m-2 s-1) in a water bath at 23 ?C. Dark controls were wrapped in foil. In one set of experiments clam tissue slices were removed at intervals and extracted by homogenization in 5 ml of methanol:tetrahydrofuran (THF) (4:1, by volume) at 0 ?C. The extraction was repeated four times and the residual animal material solubilized in 10 ml 10 M KOH. Aliquots from both fractions were acidified, bleached in sunlight and radioactivity determined by scintillation counting. In another set of experi- ments tissue slices were removed at intervals, frozen at -80 ?C then thawed and homogenized in seawater. The homogenate was filtered through a glass fibre plug to separate zooxanthellae, which were pelleted by centrifugation and extracted in methanol: THF. The animal material was solubilized in 10 M KOH. Samples of animal tissue, supernatant incubation medium and extracted zooxanthellae were acidified, bleached and radioactivity determined by scintillation counting. Less than 2 % of acid-stable 14C was found in incubation media.
In experiments with isolated zooxanthellae, samples of the cell preparation were added to seawater containing NaH14CO3 to give 10 tg Chl a ml-l and the reaction terminated at predetermined times either by injection of 10 % (by volume) acetic acid, or by centrifugation to pellet the cells which were then extracted in ethanol: water: acetic acid (75:20:5, by volume). Samples of the supernatant were acidified with 10 % (by volume) acetic acid, and together with cell extracts, were counted in scintillant.
To determine the distribution of radioactivity, the extracts were combined and
6-2
149
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions

150 R. E. Summons, T. S. Boag and C. B. Osmond
an excess of water added. The chloroform was removed and aqueous fraction extracted twice more with chloroform. The dried and evaporated chloroform extract was counted separately as a 'lipid' fraction. A sample of the aqueous residue was passed through a column of Dowex 50 ion exchange resin (H+ form) which was washed with water. The eluate and water washings contained soluble neutral and acidic compounds. A 2 M ammonium hydroxide eluate of the Dowex column contained amino compounds. Samples of these fractions were counted and the procedure gave 92-98 % recovery of the total radioactivity in the aqueous residue.
The incorporation of 15N from N14C1, urea and NaNO3 was studied in similar
experimental protocols. In some experiments seawater was labelled with 15NH4C1 and NaH14CO3. The 15NH4C1 contained 99 at. % 15N and was added to seawater in concentrations between 10 laM and 1 mI. In some experiments the ammonium ion concentration in incubation media was monitored through time with an ammonium electrode.
Isolation of amino acids for measurements of isotope enrichment was done as described earlier (Summons & Osmond I981). The high salt content in the digest of the clam tissue homogenate precluded gas-chromatography-mass-spectrometry analysis of 15N enrichment in amino acids of that fraction. Some mass spectrometric measurements were done with a Du Pont 21-491 instrument as described earlier. Others were made with a Finnigan 4510 instrument interfaced to an Incos data
system. In this system the TMS-amino acid derivatives were chromatographed by using a 25 m x 0.3 mm bonded phase methyl silicone fused silica column (BP-1, SGE Australia) programmed from 80 to 250 ?C at 10 ?C min - with He as carrier
gas. Ionization was by electron impact at 70 eV. The complete isotope clusters for
M+'-CH3 and M+'-COOTMS ions of each amino acid were sampled by using a scan
cycle time of 0.5 s or lower and calculations of enrichment were based on the method of Biemann (1962). The statistical analysis of such results has been described (Holtum et al. X984).
Total chlorophyll a content was measured in the methanol: THF extract before acidification. It has been established (B. E. Chalker, personal communication) that
chlorophyll extinction coefficients in this solvent are almost the same as in acetone
(Jeffrey & Humphrey 1975). Chemicals were obtained from Sigma or Boehringer. Radioactive NaH14CO3 was obtained from Amersham Radiochemicals and 15N- labelled ammonium chloride was obtained from Stohler Isotope Chemicals.
RESULTS
Effect of ammonium on photosynthesis in vitro and in vivo by zooxanthellae from Tridacna spp.
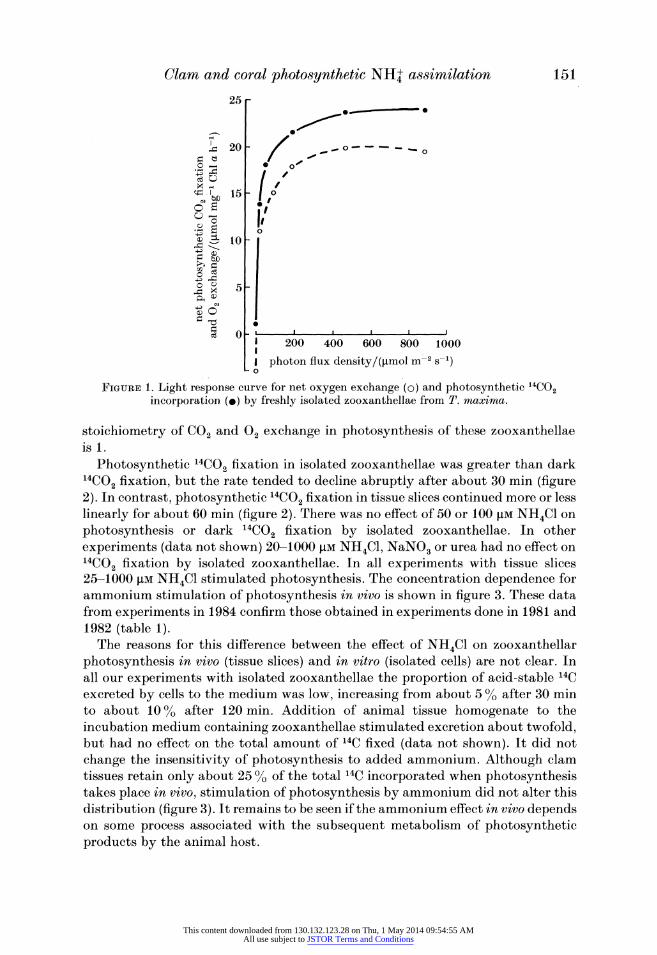
Zooxanthellae isolated from clam mantle routinely showed rates of photosyn- thesis that were 60-75 % of those measured in intact tissue slices at saturating light intensities. The light dependence of both O2 exchange and 14CO2 fixation by isolated zooxanthellae is shown in figure 1. Photosynthesis saturates at about 400 jmol quanta m-2 s-l with half saturation at only 10-20 % of this value. If it is assumed that 02 uptake owing to respiration continues in the light, then the
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions
Clam and coral photosynthetic NH+ assimilation 151
25 -
u 20 - / o - - .
15- 0o
,0- ' t o t ,
I c 5-
ce 0
! 200 400 600 800 1000
I photon flux density/(pmol m2 s-l) - o
FIGURE 1. Light response curve for net oxygen exchange (o) and photosynthetic 14C02 incorporation ()) by freshly isolated zooxanthellae from T. maxima.
stoichiometry of CO2 and 02 exchange in photosynthesis of these zooxanthellae is 1.
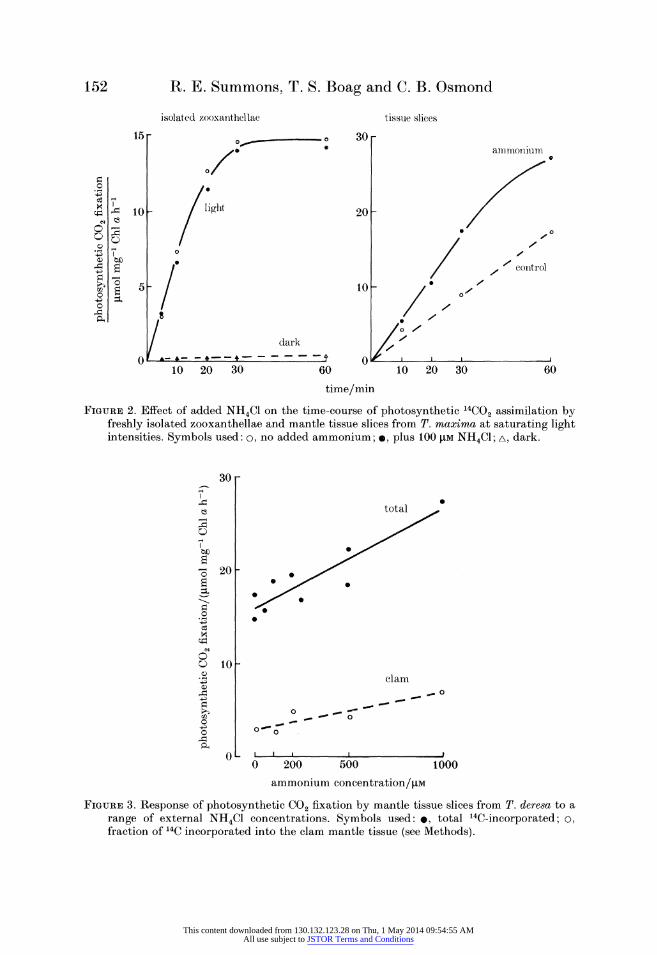
Photosynthetic 14C02 fixation in isolated zooxanthellae was greater than dark 14CO2 fixation, but the rate tended to decline abruptly after about 30 min (figure 2). In contrast, photosynthetic 14C02 fixation in tissue slices continued more or less
linearly for about 60 min (figure 2). There was no effect of 50 or 100 1M NH4C1 on
photosynthesis or dark 14C02 fixation by isolated zooxanthellae. In other
experiments (data not shown) 20-1000 gM NH4C1, NaNO3 or urea had no effect on 14CO2 fixation by isolated zooxanthellae. In all experiments with tissue slices 25-1000 LM NH4Cl stimulated photosynthesis. The concentration dependence for ammonium stimulation of photosynthesis in vivo is shown in figure 3. These data from experiments in 1984 confirm those obtained in experiments done in 1981 and 1982 (table 1).
The reasons for this difference between the effect of NH4C1 on zooxanthellar
photosynthesis in vivo (tissue slices) and in vitro (isolated cells) are not clear. In all our experiments with isolated zooxanthellae the proportion of acid-stable 14C excreted by cells to the medium was low, increasing from about 5 % after 30 min to about 10% after 120 min. Addition of animal tissue homogenate to the incubation medium containing zooxanthellae stimulated excretion about twofold, but had no effect on the total amount of 14C fixed (data not shown). It did not
change the insensitivity of photosynthesis to added ammonium. Although clam tissues retain only about 25 % of the total 14C incorporated when photosynthesis takes place in vivo, stimulation of photosynthesis by ammonium did not alter this distribution (figure 3). It remains to be seen if the ammonium effect in vivo depends on some process associated with the subsequent metabolism of photosynthetic products by the animal host.
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions
R. E. Summons, T. S. Boag and C. B. Osmond
isolated zooxanthellae tissue slices
30 r o 0
o /
light
dark 4---2 - - - - - 20 30 60
20 -
ammon ium
S 0
/ / control /-
10-
/ S - 0I
0 10 20 30 60
time/min
FIGURE 2. Effect of added NH4C1 on the time-course of photosynthetic G4C02 assimilation by freshly isolated zooxanthellae and mantle tissue slices from T. maxima at saturating light intensities. Symbols used: o, no added ammonium; , plus 100 lJM NH4CI; A, dark.
30-
total t3
Ir
St
0
0 0 0
cF:
a
CZ
0
0
-4-
01
1--1
P-
0
20 -
0
0
10 -
clam
- c --
0 a~~ 0
0 -1.
-- 0
0 0 200 500 1000
ammonium concentration/JiM
FIGURE 3. Response of photosynthetic CO2 fixation by mantle tissue slices from T. deresa to a range of external NH4CI concentrations. Symbols used: *, total 14C-incorporated; o, fraction of 14C incorporated into the clam mantle tissue (see Methods).
152
a r. 0
ece & 0 0
a
C)
-0 0
Ej
0
0
I-C
P-4
14
0
CS)
0 S
0
10
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions

Clam and coral photosynthetic NH+ assimilation 153
TABLE 1. EFFECT OF ADDED AMMONIUM ON PHOTOSYNTHETIC 14C02 FIXATION IN
ISOLATED ZOOXANTHELLAE AND TISSUE SLICES OF MANTLE FROM TRIDACNA SPP.
(In all experiments dark 14CO2 fixation was 5-10% of that in the light; control without added NH4CI.)
14C incorporation 103 x (disintegrations per minute)
experimental conditions 30 min 60 min
isolated zooxanthellae experiment 1 control 98 167
+ 100 M NH4Cl 98 159 experiment 2 control 44 92
+ 50 gM NH4CI 49 99 experiment 3 control 76
+ 100 M NH4C1 77 tissue slices
experiment 3 control 17.9 + 100 IM NH4Cl -- 24.3
experiment 4 control 1 4.2 + 50 gM NH4CI 30.9 + 100 IM NH4Cl 18.1 +250 Im NH4C1 25.3 + 500 gM NH4Cl 25.8
Effect of ammonium on photosynthesis by zooxanthellae from corals
Whereas the addition of ammonium to seawater produced a consistent stimulation of photosynthesis by tissue slices of clam mantle, results with coral tips were much more variable (table 2). It is not clear whether season or location is responsible for the different responses of corals from the two populations studied. In two
separate experiments with Stylophora in which ammonium stimulated photosyn- thesis, the response was associated with greater increase of 14C in water-soluble
TABLE 2. THE EFFECT OF ADDED AMMONIUM ON THE RATE OF PHOTOSYNTHESIS
IN CORALS
(All corals were collected from 3-5 m depth and rates are calculated from the initial slope of 60 min time courses of 14CO2 fixation; control without added NH4Cl.)
photosynthesis pmol mg-l Chl a h-l
species and source control + 25 JM NH4Cl
Stylophora pistillata Heron Island 27 35 Davies Reef 25 48a Davies Reef 30 32
Acropora millipora Davies Reef 84 75
a Chemical distribution of 14C fixed in this experiment is shown in table 3.
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions

R. E. Summons, T. S. Boag and C. B. Osmond
compounds than in the lipids (chloroform soluble), which were the principal products of photosynthesis (table 3). The presence of ammonium did not increase the proportion of water-soluble label in amino compounds and these comprised the minor fraction (25-30 %) of water-soluble label in all experiments. In experiments with this tissue there was no evidence that the stimulation of photosynthesis in
response to ammonium could be accounted for by amino acid synthesis alone. In contrast to the Tridacna tissue slices which were increasingly stimulated by ammonium concentrations up to 1000 1M, the coral tips showed the largest percentage stimulation at 25 LM with relatively little or no increase at 500 1.M.
TABLE 3. EFFECTS OF ADDED AMMONIUM ON PHOTOSYNTHESIS AND THE DISTRIBUTION
OF 14C AMONG PHOTOSYNTHETIC PRODUCTS IN THE CORAL STYLOPHORA
PISTILLA TA
(Experiment conducted in the field at Davies Reef. Percentage distribution shown in parentheses; controls without added NH4Cl.)
14C incorporation 106 x (d.p.m.a mg-1 Chl a)
X__
treatment total CHCl3-soluble H20-soluble amino acids
dark, 30 min control 0.7 0.07 (10) 0.62 (89) 0.23 (33) 25 [tM NH4C1 0.9 0.11 (13) 0.78 (87) 0.27 (31) 500 1M NH4CI 1.3 0.11 (8) 1.22(92) 0.44 (34)
dark, 60 min control 0.9 0.13 (13) 0.85 (87) 0.30 (31) 25 gM NH4C1 1.3 0.12 (9) 1.21 (91) 0.48 (36) 500 gM NH4Cl 2.4 0.32 (13) 2.10 (87) 0.83 (35)
light, 30 min control 14.1 10.50 (74) 3.6 (26) 0.89 (6) 25 pM NH4C1 23.9 18.15 (76) 5.7 (24) 1.43 (6) 500 1M NH4C1 27.5 16.91 (62) 10.5 (39) 2.50 (9)
light, 60 min control 26.3 22.6 (86) 3.73 (14) 1.02 (4) 25 pM NH4Cl 36.8 28.6 (78) 8.16 (22) 2.46 (7) 500 pM NH4Cl 35.4 24.2 (68) 11.18 (32) 3.15 (9)
a Disintegrations per minute.
Incorporation of 15NH4 by isolated zooxanthellae and tissue slices of Tridacna mantle
Ammonium uptake by zooxanthellae isolated from Tridacna mantle was stimu- lated about fourfold by light (figure 4). The time course of 15N incorporation into amino compounds confirms and extends our preliminary observations with zooxanthellae isolated from another clam, Hippopus (Summons & Osmond 1981). Figure 5 shows that the amido-N of glutamine was the most rapidly labelled
organic nitrogen moiety and that labelling was not light-stimulated. However, transfer of label to the amino-N of glutamate, glutamine, aspartate and serine was stimulated about twofold by light. Similar results were obtained when this
experiment was repeated with tissue slices (figure 6).
154
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions

Clam and coral photosynthetic NH+ assimilation 155
100
? > 80- \ ^0 I \ ^ dark
' m5 60- \ E o0
40 - 0 O ce
E S light
O?^~~~~~ ' 20 40 60
time/min
FIGURE 4. The uptake of ammonium from the incubation medium by freshly isolated zooxanthellae in either the light (o) or the dark (.).
glutamine glutamate 100 - 30 -
20- amido o/
80- - 10- / 10
^0 -
II I 0 .
60 I aspartate 60 - 30 -
I 4 o 1 20- ,
,0R; g 1 | ,1
anmino serine 2 - 20 -
- 0 10 0
0 O
2 10 20 40 60
time/min
FIGURE 5. Time-course of 15N assimilation from '5NH4CI by freshly isolated zooxanthellae from T. maxima either in the light (o) or in the dark (o).
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions

R. E. Summons, T. S. Boag and C. B. Osmond glutamine
* i-.---..-.. 0----
I amido
30 - glutamate
amino
.., 0.-... - -
.00 0
0 .0
20 - -. . 0
.10, - - 0
1 0 -
/ 0 a
ow-J iI, -
20 60 120 20 60 time/min
FIGURE 6. Time-course of 15N assimilation from 15NH4CI by mantle tissue slices of T. maxima in the light (o) or in the dark (e).
12(
DISCUSSION
These data show important interactions between light-dependent metabolic
processes in symbiotic zooxanthellae and their hosts with respect to ammonium
recycling. The addition of ammonium to seawater stimulates in vivo photosynthesis by the zooxanthellae of intact clam mantles and some corals. This stimulation confirms the studies of Taylor (I 978) with corals. The stimulation of photosynthesis by ammonium is lost after isolation of the zooxanthellae. Our freshly isolated clam zooxanthellae seem to be comparable with Taylor's cultured zooxanthellae in
stationary phase. However, Taylor (1978) reported stimulation of photosynthesis by the same concentration range of ammonium (20-200 gM) when zooxanthellae in log phase growth were used.
Zooxanthellae in corals and those extracted from an anemone have high affinities for ammonium (apparent Ks 1-10 gM) (Muscatine & D'Elia 1978; Muscatine et al. 1979; Wilkerson & Muscatine I984). It is possible that some kind of ammonium
concentrating mechanism may function in vivo, which elevates ammonium concen- trations from those in seawater to the range in which algal photosynthesis responds in our experiments (50-1000 iM). However, Wilkerson & Muscatine (1984) found that ammonium uptake in an anemone saturates at 5-6 mM. The relationships
156 100 -
80 -
60
40
--0
0-
C) ;--4
0
P-
0 C.
la
20
0
.OO - O
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions

Clam and coral photosynthetic NH+ assimilation
between ammonium uptake at very low concentrations in seawater, external ammonium concentration which stimulates in vivo photosynthesis in our experi- ments, and the internal concentration of ammonium to which zooxanthellae are exposed, are not clear (Muscatine 1980; Cates & McLaughlin 1976). Neither is it clear why isolated zooxanthellae should lose their capacity to show an increase in
photosynthesis when provided with ammonium ions. The stimulation of in vivo photosynthesis by zooxanthellae in response to added
ammonium does not seem to be related simply to increased synthesis of amino
compounds. In the coral Stylophora from Davies Reef, ammonium increased the
incorporation of label into all fractions, but the increase in the water-soluble materials was much greater. Thus ammonium reduced the proportion, but not the amount, of 14C incorporated into lipids, and stimulated the proportion and amount of label in water-soluble compounds. Although amino acid 14C incorporation was increased two- to fourfold by ammonium, these compounds comprised only 20-30 % of the water solubles. These responses differ from the effect of ammonium on photosynthesis by Chlorella and spinach cells in which there was a diversion of assimilate into amino acids at the expense of other products, without an effect on rate (Kanazawa et al. 1972; Larsen et al. 1981). However, several other studies with spinach and other plant cell systems show an effect similar to that observed here. In these, ammonium stimulation of photosynthesis could be explained by increased amino acid synthesis alone, with small effects on carbon distribution (Paul et al. 1978; Woo & Canvin 1980). Stimulation of dark carbon assimilation
processes in our experiments were quantitatively inadequate to account for the
photosynthetic response (by a factor about tenfold). Further analysis of the metabolites involved in the stimulation of photosynthesis by ammonium in these
symbiotic systems may help answer these questions. Although ammonium did not stimulate photosynthesis in isolated zooxanthellae,
a light stimulation of ammonium incorporation was still observed. As discussed
previously (Summons & Osmond 1981) the light stimulation of transfer of label from glutamine amido to glutamate and glutamine amino groups is strong evidence for participation of GS-GOGAT in the ammonium fixation system of the zooxan- thellae in vitro and in vivo. Failure to demonstrate GOGAT activity coupled to NADPH in zooxanthellae from Aiptasia (Wilkerson & Muscatine 1984) may imply an in vivo coupling of this enzyme to reduced ferredoxin in these cells. There is a useful analogy between the proposed light-stimulated ammonium recycling by zooxanthellae in the marine symbiosis and that found in cells of higher plants in the course of photorespiration. In the latter ammonium released by glycine decarboxylation in mitochondria is recovered by a chloroplast GS-GOGAT system (Keys et al. 1978; Woo et al. 1982).
The dark rate of 15NH4 incorporation found in our studies possibly reflects some concurrent activity of a glutamate dehydrogenase system in zooxanthellae. However, this enzyme has an extraordinarily low affinity for ammonium in phytoplankton as well as other organisms. In cell-free extracts of zooxanthellae from Aiptasia, Wilkerson & Muscatine (i 984) measured an apparent Km of 4-5 mM for ammonium and hence one would expect its contribution to ammonium uptake to be minor, except at very high ammonium ion concentrations.
157
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions

R. E. Summons, T. S. Boag and C. B. Osmond
In our time-course study of nitrogen labelling we again found the specific activity of glutamine amino-N to be lower than that of glutamate (cf. Summons & Osmond 1981). Glutamine amido-N remained at considerably higher specific activity than the amino-N of glutamine and glutamate. These observations are consistent with sequential dilutions of specific activity of the nitrogen label and the order in which the respective nitrogens are labelled in the GS-GOGAT
pathway. Semi-quantitative observations of the relative pool sizes were consistent with a large (possibly multiple) pool of glutamate and a smaller glutamine pool which increased relative to glutamate over the time-course. Further studies, particularly those using pulse-chase or inhibitors techniques are required to assess the alternate pathways of ammonium incorporation.
The observation of increased photosynthetic carbon assimilation in Tridacna with increased levels of ambient ammonium ion concentrations in our short-term
experiments is consistent with longer term observations in tropical communities dominated by symbiotic systems. Colal heads with resident fish schools have been shown to grow faster than those without (Meyer et al. 1983) and Kinsey & Domm
(I974) observed an increase of about 50 % in net community photosynthesis when a lagoonal patch reef was artificially fertilized to maintain 20 tLM nitrogen (as NH+ and NO3) and 2 [tM phosphate.
This work was supported by an M.S.T. (formerly A.M.S.T.A.C.) grant. We
acknowledge the help of Dr T. J. Andrews, Dr. J. Bunt and Dr C. Wilkinson, Australian Institute of Marine Science, Townsville, in collection of material and
providing laboratory facilities. We thank Dr A. A. Benson, Dr C. F. D'Elia and Dr K. C. Woo for advice and encouragement. Dixie Nott, David Everitt, Prue Kell and John Wicks also helped in different phases of the work.
REFERENCES
Biemann, K. 1962 Mass spectrometry: organic chemical applications, p. 204. New York: McGraw Hill.
Burris, R. H. 1976 Nitrogen fixation by blue-green algae of Lizard Island area of the Great Barrier Reef. Aust. J. Pl. Physiol. 3, 41-51.
Cates, N. & McLaughlin, J. J. A. 1976 Differences of ammonia metabolism in symbiotic and asymbiotic Condylactus and Cassiopea spp. J. exp. mar. Biol. Ecol. 21, 1-5.
Goldman, J. C., Dennet, M. R. & Riley, C. B. I981 Marine phytoplankton photosynthesis and transient ammonium availability. Mar. Biol. Lett. 2, 323-331.
Holtum, J. A. M., Summons, R. E., Roeske, C. A., Comins, H. N. & O'Leary, M. H. 1984 Oxygen-18 incorporation into malic acid during nocturnal carbon dioxide fixation in crassulacean acid metabolism plants. J. biol. Chem. 259, 6870-6881.
Jeffrey, S. W. & Humphrey, G. F. 1975 New spectrophotometric equations for determining chlorophylls a, b, c and cl and c, in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pfl. 167, 191-194.
Kanazawa, T., Kanazawa, K. & Kirk, M. 1972 Effect of ammonia on carbon metabolism in Chlorella pyrenoidosa during photosynthesis and respiration. Biochim. biophys. Acta 256, 565-669.
Keys, A. J., Bird, I. F., Cornelius, M. J., Lea, P. J., Wallsgrove, R. M. & Miflin, B. J. 1978 The photorespiratory nitrogen cycle. Nature, Lond. 275, 741-743.
Kinsey, D. W. & Domm, A. 1974 Effects of fertilization on a coral reef environment - primary production studies. Proc. int. 2nd Coral Reef Symp. (Brisbane) 1, 49-66.
158
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions

Clam and coral photosynthetic NH+ assimilation 159
Larsen, P. 0., Cornwell, K. L., Gee, S. L. & Bassham, J. A. 198I Amino acid synthesis in photosynthesizing spinach cells. Pi. Physiol. 68, 292-299.
Lea, P. J. & Miflin, B. J. 1979 Photosynthetic ammonia assimilation. In Encyclopaedia of plant Physiology, new series (ed. M. Gibbs & E. Latzko), vol. 6, pp. 445-456. Berlin: Springer Verlag.
Lewis D. H. & Smith, D. C. 197I The autotrophic nutrition of symbiotic marine coelenterates with special reference to hermatypic corals. I. Movement of photosynthetic products between symbionts. Proc. R. Soc. Lond. B 178, 111-129.
Meyer, J. L., Schultz, E. T. & Helfman, G. S. 1983 Fish schools: an asset to corals. Science, Wash. 220, 1047-1049.
Muscatine, L. 1980 Uptake retention and release of dissolved inorganic nutrients by marine alga-invertebrate associations. In Cellular interactions in symbiosis and parasitism (ed. C. B. Cook), pp. 229-243. Columbus: Ohio State University Press.
Muscatine, L. & D'Elia, C. F. 1978 The uptake retention and release of ammonium by reef corals. Limnol. Oceanogr. 23, 725-734.
Muscatine, L., Masuda, H. & Burnap, R. 1979 Ammonia uptake and release by symbiotic and asymbiotic reef corals. Bull. mar. Sci. 29, 572-575.
Muscatine, L., Pool, R. R. & Cernichiari, E. 1972 Some factors affecting selective release of soluble organic material by zooxanthellae from reef corals. Mar. Biol. 13, 298-308.
Paul, J. S., Cornwell, K. L. & Bassham, J. A. 1978 Effects of ammonia on carbon metabolism in photosynthesizing isolated cells from Papaver somniferum L. Planta 142, 49-54.
Summons, R. E. & Osmond, C. B. 1981 Nitrogen assimilation in the symbiotic marine alga Gymnodinium microadriaticum: direct analysis of 15N incorporation by GCMS methods. Phytochemistry 20, 575-578.
Taylor, D. L. 1978 Nutrition of algal-invertebrate symbiosis. II. Effects of exogenous nitrogen sources on growth, photosynthesis and rate of excretion by algal symbionts in vivo and in vitro. Proc. R. Soc. Lond. B 201, 401-412.
Thomas, W. H. 1970 On nitrogen deficiency in tropical Pacific Oceanic phytoplankton: photosynthetic parameters in poor and rich waters. Limnol. Oceanogr. 15, 380-385.
Trench, R. K. 1979 The cell biology of plant-animal symbiosis. A. Rev. Pi. Physiol. 30, 485-531.
Webb, K. L., DuPaul, W. D., Wiebe, W., Sottile, W. & Johannes, R. E. 1975 Enewetak Atoll: aspects of the nitrogen cycle on a coral reef. Limnol. Oceanogr. 20, 198-210.
Wiebe, W. J., Johannes, R. E. & Webb, K. L. 1975 Nitrogen fixation in a coral reef community. Science, Wash. 188, 257-259.
Wilkerson, F. P. & Muscatine, L. 1984 Uptake and assimilation of dissolved inorganic nitrogen by a symbiotic sea anemone. Proc. R. Soc. Lond. B 221, 71-86.
Wilkinson, C. R., Williams, D. McB., Sammarco, P. W., Hogg, R. W. & Trott, L. A. 1984 Rates of nitrogen fixation on coral reefs across the continental shelf of the central Great Barrier Reef. Mar. Biol. 80, 255-262.
Woo, K. C. & Canvin, D. T. 1980 Effect of ammonia on photosynthetic carbon fixation in isolated spinach cells. Can. J. Bot. 58, 506-510.
Woo, K. C., Morot-Gaudry, J. F., Summons, R. E. & Osmond, C. B I982 Evidence for the glutamine synthetase/glutamate synthase pathway during the photorespiratory nitrogen cycle in spinach leaves. Pl. Physiol. 70, 1514-1517.
This content downloaded from 130.132.123.28 on Thu, 1 May 2014 09:54:55 AMAll use subject to JSTOR Terms and Conditions