the development and function of regulatory b cells ... · the development and function of...
TRANSCRIPT

of May 23, 2012.This information is current as
TLR SignalsRequires Antigen Receptor Diversity andB Cells Expressing IL-10 (B10 Cells) The Development and Function of Regulatory
Takeshi Tsubata and Thomas F. TedderKoichi Yanaba, Jean-David Bouaziz, Takashi Matsushita,
http://www.jimmunol.org/content/182/12/7459doi: 10.4049/jimmunol.0900270
2009; 182:7459-7472; ;J Immunol
Referenceshttp://www.jimmunol.org/content/182/12/7459.full#ref-list-1
, 23 of which you can access for free at: cites 56 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2009 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

The Development and Function of Regulatory B CellsExpressing IL-10 (B10 Cells) Requires Antigen ReceptorDiversity and TLR Signals1
Koichi Yanaba,2* Jean-David Bouaziz,2* Takashi Matsushita,2* Takeshi Tsubata,†
and Thomas F. Tedder3*
Autoimmunity and inflammation are controlled in part by regulatory B cells, including a recently identified IL-10-competentCD1dhighCD5� B cell subset termed B10 cells that represents 1–3% of adult mouse spleen B cells. In this study, pathways thatinfluence B10 cell generation and IL-10 production were identified and compared with previously described regulatory B cells.IL-10-competent B cells were predominantly CD1dhighCD5� in adult spleen and were the prevalent source of IL-10, but not othercytokines. B10 cell development and/or maturation in vivo required Ag receptor diversity and intact signaling pathways, but notT cells, gut-associated flora, or environmental pathogens. Spleen B10 cell frequencies were significantly expanded in aged mice andmice predisposed to autoimmunity, but were significantly decreased in mouse strains that are susceptible to exogenous autoan-tigen-induced autoimmunity. LPS, PMA, plus ionomycin stimulation in vitro for 5 h induced B10 cells to express cytoplasmicIL-10. However, prolonged LPS or CD40 stimulation (48 h) induced additional adult spleen CD1dhighCD5� B cells to express IL-10following PMA plus ionomycin stimulation. Prolonged LPS or CD40 stimulation of newborn spleen and adult blood or lymph nodeCD1dlow and/or CD5� B cells also induced cytoplasmic IL-10 competence in rare B cells, with CD40 ligation uniformly inducingCD5 expression. IL-10 secretion was induced by LPS signaling through MyD88-dependent pathways, but not following CD40ligation. LPS stimulation also induced rapid B10 cell clonal expansion when compared with other spleen B cells. Thereby, bothadaptive and innate signals regulate B10 cell development, maturation, CD5 expression, and competence for IL-10production. The Journal of Immunology, 2009, 182: 7459–7472.
I mmunological tolerance exemplifies the capacity of the im-mune system to down-modulate immune responses. B cellsare generally considered to positively regulate immune re-
sponses by producing Ag-specific Ab and helping induce optimalCD4� T cell activation (1). However, B cells and specific B cellsubsets can also negatively regulate immune responses in mice,validating the existence of regulatory B cells (2–8). The absence orloss of negative regulatory B cells exacerbates disease symptomsin contact hypersensitivity (CHS),4 experimental autoimmune en-cephalomyelitis (EAE), chronic colitis, and collagen-induced ar-thritis (CIA) models of autoimmunity and inflammation (9–18).
Furthermore, IL-10-producing B cells down-regulate autoimmunedisease initiation, onset, or severity in EAE (13, 19), CIA (15),CHS (18), and inflammatory bowel disease (14). Although IL-10can augment immune responses, it can also suppress both Th1 andTh2 polarization and inhibit Ag presentation and proinflammatorycytokine production by monocytes and macrophages (20). IL-10production and/or regulatory B cell activities have been variablyattributed to all B cells, CD5� B-1a cells, or cells withCD21�CD23� marginal zone (MZ) or CD1d�CD21�CD23�
T2-MZ precursor B cell phenotypes (17, 21, 22). B cells can alsocontribute to immunoregulation through the production of IL-4,IL-6, IFN-�, and TGF-� (23, 24). Altogether, these studies haveestablished that phenotypically diverse regulatory B cells signifi-cantly influence immune responses.
A potent subset of regulatory B cells was recently found toregulate T cell-dependent CHS and EAE responses in an IL-10-dependent manner (18, 19). This phenotypically uniqueCD1dhighCD5�CD19high subset of regulatory B cells sharesoverlapping cell surface markers with the CD5� B-1a,CD21�CD23� MZ, and CD1d�CD21�CD23� T2-MZ precur-sor B cell subsets (17, 18, 21, 22), and is found within thespleens of naive wild-type mice at frequencies of 1–2% (18).These CD1dhighCD5� B cells are induced to express cytoplas-mic IL-10 following 5-h in vitro stimulation with LPS, PMA,ionomycin, plus monensin (L�PIM), and are called B10 cells todistinguish them from other regulatory B cell subsets that mayalso exist and to identify them as the predominant source of Bcell IL-10 production (25). B10 cell regulatory functions are Agrestricted in vivo (18, 19), implying a requirement for Ag-spe-cific BCR signaling. The adoptive transfer of Ag-primed B10cells reduces inflammation during CHS responses and reduces
*Department of Immunology, Duke University Medical Center, Durham, NC 27710;and †Department of Immunology, Medical Research Institute, Tokyo Medical andDental University, Tokyo, Japan
Received for publication January 23, 2009. Accepted for publication April 20, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grants CA105001,CA96547, AI56363, and AI057157. J.-D.B. is supported by grants from Associationpour la Recherche contre le Cancer, the Fondation Rene Touraine, and the PhilippeFoundation.2 K.Y., J.-D.B., and T.M. contributed equally to these studies and share firstauthorship.3 Address correspondence and reprint requests to Dr. Thomas F. Tedder, Box 3010,Department of Immunology, Duke University Medical Center, Durham, NC 27710.E-mail address: [email protected] Abbreviations used in this paper: CHS, contact hypersensitivity; CD40L/BTg,CD40L/B transgenic; CIA, collagen-induced arthritis; EAE, experimental autoim-mune encephalomyelitis; hCD19Tg, human CD19 transgenic; int, intermediate;L�PIM, LPS, PMA, ionomycin, plus monensin; MZ, marginal zone; NZB/W, NewZealand Black/White.
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0900270
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

EAE severity during disease onset (18, 19), and may also reg-ulate other autoimmune diseases (25).
Regulatory B cells are commonly stimulated in vitro to inducetheir capacity to inhibit immune responses in mice with autoim-mune disease. For example, the activation of arthritogenic spleno-cytes with collagen alone (17) or collagen plus agonistic CD40mAb in vitro gives rise to B cells that produce high IL-10 levelsand prevent arthritis (15). Transfusions of BCR-activated B cellsalso protect NOD mice from type 1 diabetes in an IL-10-dependentmanner (26). LPS-activated B cells are also reported to preventdiabetes in NOD mice (24). Mouse B cells express TLR-4 andRP-105 that bind LPS (27), and B10 cells produce IL-10 in re-sponse to LPS stimulation in vitro (18). BCR and CD40 engage-ment also appear to be required for IL-10-dependent regulatory Bcell functions in CIA, CHS, and EAE models (13, 17–19). SpleenB cells with a CD1dhighCD21�CD23� MZ phenotype can alsoproduce IL-10 in response to CpG stimulation in mice with lupus-like autoimmune disease (22). Based on these findings, Mizoguchiand Bhan (2) have postulated that distinct regulatory B cell subsetsmight arise from separate follicular and MZ B cells through innatetype (polyclonal stimulus-induced IL-10 production) or acquiredtype (Ag-specific IL-10 production) developmental pathways, re-spectively, or from B1 cells.
Identification of B10 cells as a phenotypically distinct, IL-10-producing B cell subset (18, 19) facilitates their characterization aseither innate type or acquired type regulatory B cells. Whether B10cells share the physiologic triggers that lead to their expansion andfunction as described for other regulatory B cells is unknown. It isalso unknown whether B10 cells produce cytokines other than IL-10. Therefore, these issues and the factors that influence B10 cellgeneration in vivo and in vitro were examined in the current studywith the finding that both adaptive and innate signals facilitate B10cell generation, maturation, and optimal IL-10 production.
Materials and MethodsMice
Wild-type C57BL/6 (B6), IL-10�/� (B6.129P2-Il10tmlCgn/J), NOD (NOD/Lt), DBA/1J, SJL/J, New Zealand Black/White (NZB/W) F1 (NZBWF1/J),CD40�/� (B6.129P2-CD40tm1Kik/J), MRL/lpr (MRL/MpJ-Faslpr/J), andMD4 (C57BL/6-Tg(TghelMD4)4Ccg/J) that express IgM and IgD specificfor HEL (28), and nude (C57BL/6-Hfh11�) mice were from The JacksonLaboratory. MHC-I/II�/� (B6.129-H2-Ab1tm1GruB2mtmJaeN17) from Tac-onic Farms mice were as described (29) and were provided by Y. Zhuang(Duke University, Durham, NC). MyD88�/� mice (30) were provided byY. Yang (Duke University) with the permission of S. Akira (Osaka Uni-versity, Osaka, Japan). CD22�/�, CD21�/�, CD19�/�, and human CD19transgenic (hCD19Tg) (h19-1 line) mice on a B6 genetic background wereas described (31–34). CD40L/B transgenic (CD40L/BTg) mice with Bcells expressing cell surface CD40L were as described (35). CD40L/BTg/CD22�/� double-mutant mice were generated by crossing CD40L/BTgmice with CD22�/� mice. B6 neonates were 3–10 days old. All mice werehoused in a specific pathogen-free barrier facility and used at 12–16 wk ofage, unless otherwise specified. All studies were approved by the DukeUniversity Animal Care and Use Committee. Tissues from 6-mo-old gno-tobiotic and specific pathogen-free 129S6/SvEv mice were provided by S.Plevy and the University of North Carolina Center for GastrointestinalBiology and Disease Gnotobiotic Core (Chapel Hill, NC).
Antibodies
Anti-mouse mAbs included the following: B220 mAb RA3-6B2 (providedby R. Coffman, DNAX, Palo Alto, CA), and CD19 (1D3), CD5 (53-7.3),CD1d (1B1), CD40 (HM40-3), CD21/35 (7G6), CD23 (B3B4), CD24(M1/69), CD43 (S7), and CD93 (AA4.1) mAbs from BD Pharmingen.Anti-mouse IgM Ab was from Jackson ImmunoResearch Laboratories. PE-conjugated anti-mouse IL-10 mAb (JES5-16E3) was from eBioscience.
B cell isolation, immunofluorescence analysis, and cell sorting
Blood mononuclear cells were isolated from heparinized blood after cen-trifugation over a discontinuous Lymphoprep (Axis-Shield) gradient. Sin-
gle-cell splenocyte suspensions were generated by gentle dissection with�90% cell viability as determined by trypan blue exclusion. Cell numberswere quantified using a hemocytometer, with relative lymphocyte percent-ages among viable cells (based on scatter properties) determined by flowcytometry analysis. B220 or CD19 mAb-coated microbeads (Miltenyi Bio-tec) were used to purify spleen B cells by positive selection following themanufacturer’s instructions. When necessary, the cells were enriched asecond time using a fresh MACS column to obtain �99% purities.
Single-cell leukocyte suspensions were stained on ice using predeter-mined optimal concentrations of each Ab for 20–60 min, and fixed asdescribed (32). Cells with the light scatter properties of lymphocytes wereanalyzed by two- to four-color immunofluorescence staining and FACScanor FACSCalibur flow cytometers (BD Biosciences). Dead cells were ex-cluded from the analysis based on their forward and side light scatter prop-erties and the use of LIVE/DEAD Fixable Dead Cell Stain Kits (Invitro-gen-Molecular Probes). All histograms are shown on a 4-decadelogarithmic scale, with gates shown to indicate background isotype-matched control mAb staining set with �2% of the cells being positive.Background staining was determined using unreactive isotype-matchedcontrol mAbs (Caltag Laboratories) with gates positioned to exclude�98% of unreactive cells. Spleen CD1dhighCD5�, and CD1dlowCD5� Bcells were isolated using a FACSVantage SE flow cytometer (BD Bio-sciences) with �75–95% purities.
Analysis of IL-10 production
Intracellular IL-10 analysis by flow cytometry was as described (18).Briefly, isolated leukocytes or purified cells were resuspended (2 � 106
cells/ml) in complete medium (RPMI 1640 medium containing 10% FCS,200 �g/ml penicillin, 200 U/ml streptomycin, 4 mM L-glutamine, and 5 �10�5 M 2-ME (all from Life Technologies)) with LPS (10 �g/ml, Esche-richia coli serotype 0111:B4; Sigma-Aldrich), PMA (50 ng/ml; Sigma-Aldrich), ionomycin (500 ng/ml; Sigma-Aldrich), and monensin (2 �M;eBioscience) for 5 h, in 24-well flat-bottom plates. In some experiments,the cells were incubated for 48 h with LPS (10 �g/ml), and/or anti-mouseCD40 mAb (1 �g/ml), and/or anti-mouse IgM Ab (10 �g/ml; JacksonImmunoResearch Laboratories). For analysis of cell proliferation, leuko-cytes were stained with CFSE Vybrant CFDA SE fluorescent dye (0.1 �M;CFSE; Invitrogen-Molecular Probes), according to the manufacturer’s in-structions. For IL-10 detection, FcRs were blocked with mouse FcR mAb(2.4G2; BD Pharmingen) with dead cells detected by using a LIVE/DEADFixable Green Dead Cell Stain Kit (Invitrogen-Molecular Probes) beforecell surface staining. Stained cells were fixed and permeabilized using aCytofix/Cytoperm kit (BD Pharmingen), according to the manufacturer’sinstructions, and stained with PE-conjugated mouse anti-IL-10 mAb. Leu-kocytes from IL-10�/� mice served as negative controls to demonstratespecificity and to establish background IL-10-staining levels.
Secreted IL-10 was quantified by ELISA. Purified B cells (4 � 105)were cultured in 0.2 ml of complete medium in 96-well flat-bottom tissueculture plates. Culture supernatant fluid IL-10 concentrations were quan-tified using IL-10 OptEIA ELISA kits (BD Pharmingen) following themanufacturer’s protocols. All assays were conducted using triplicatesamples.
B cell cytokine transcript expression analysis
Purified spleen B cells were cultured for 5 h with LPS plus PMA plusionomycin. IL-10-secreting spleen B cells were identified using an IL-10secretion detection kit (Miltenyi Biotec) with subsequent staining for CD19expression before cell sorting into IL-10�CD19� and IL-10�CD19� pop-ulations. Total RNA was extracted from the purified B cells using TRIzol(Invitrogen-Molecular Probes), with relative cytokine transcripts quantifiedby GeneChip analysis (Affymetrix Mouse Genome 430 2.0 GeneChips;Affymetrix). All quality parameters for the arrays were confirmed to be inthe range recommended by the manufacturer.
Statistical analysis
All data are shown as means (�SEM). Significant differences betweensample means were determined using Student’s t test.
ResultsIL-10-producing B cells preferentially secrete IL-10
Spleen B cells that are competent to express cytoplasmic IL-10following a 5 h L�PIM stimulation were predominantly foundwithin the CD1dhighCD5�CD19� subset in wild-type B6 mice(Fig. 1A), as described (18, 19). By contrast, IL-10-expressing Bcells were significantly less common within the CD1dhighCD5�,
7460 REGULATORY B10 CELL DEVELOPMENT
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

CD1dlowCD5�, or CD1dlowCD5� B cell subsets ( p � 0.01), withB cells from IL-10�/� mice used as negative controls for back-ground IL-10 staining. We have previously shown that 5-h L�PIMstimulation does not influence the phenotype of these B cellsubsets (18). IL-10�CD19� B10 cells were predominantlyCD21int/high, CD23low, CD24high, CD43�/�, and CD93� (AA4.1)(Fig. 1B). Thereby, spleen B10 cells are relatively rare and share
some overlapping phenotypic markers with the B-1a, MZ, andT2-MZ precursor B cell subsets, but are nonetheless phenotypi-cally distinct (18, 19).
Determining whether spleen B10 cells purified from wild-typemice produce only IL-10 was problematic due to the inherent tech-nical difficulties when purifying such low-frequency cells and thepredominantly low-level induction of most cytokines by B cells
FIGURE 1. B10 cells preferentiallysecrete IL-10. A, IL-10-producing Bcells were predominantly found withinthe CD1dhighCD5�CD19� B cell sub-set. Splenocytes from wild-type and IL-10�/� mice were cultured with L�PIMfor 5 h, then stained with CD1d, CD5,and CD19 mAb before permeabiliza-tion and staining using IL-10 mAb. Per-centages and bar graphs indicate mean(�SEM) B cell subset frequencies andnumbers among CD19� splenocytes orIL-10� cell frequencies among the in-dicated B cell subsets (a, CD1dhigh
CD5�; b, CD1dhighCD5�; c,CD1dlowCD5�; d, CD1dlowCD5�)from three mice as determined by flowcytometry analysis. Values signifi-cantly different from background fre-quencies or numbers for IL-10�/� miceare indicated, as follows: �, p � 0.05;��, p � 0.01. B, CD21, CD23, CD24,CD43, and CD93 expression by IL-10-producing (thick line) and IL-10� (thinline) CD19� spleen B cells from wild-type mice cultured with L�PIM for5 h, and then stained for cell surfaceAgs before permeabilization andcytoplasmic IL-10 staining. Gray histo-grams represent isotype-matched con-trol mAb staining. Results are represen-tative of those obtained with B cellsfrom �3 mice as determined by flowcytometry analysis. C, IL-10-producingB cells from hCD19Tg mice arepredominantly found within theCD1dhighCD5�CD19� B cell subset.Staining and analysis were as describedin A. D, Representative isolation of IL-10-secreting B cells. Splenic B220�
cells purified from three hCD19Tgmice were pooled and cultured withLPS plus PMA plus ionomycin for 5 hbefore staining for CD19 and secretedIL-10 capture (left panel). IL-10� andIL-10� B cells were isolated by cellsorting using the indicated gates, andsubsequently reassessed for IL-10 se-cretion and CD19 expression (rightpanels). E, Cytokine gene expressionby IL-10-secreting and nonsecreting Bcells purified as in D. Mean fold differ-ences (�SEM) in cytokine transcriptlevels (IL-10�/IL-10� cells) from threeindependent experiments are shown.Values of 1 (dashed line) indicate nodifference in cytokine expression be-tween the IL-10� and IL-10� B cells,with significant differences indicated,as follows: ��, p � 0.005.
7461The Journal of Immunology
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

(data not shown). However, spleen B10 cell frequencies and num-bers are expanded in hCD19Tg mice (Fig. 1C) (18). Within theCD1dhighCD5� B cell subset in hCD19Tg mice, 58% of the cellswere induced to express cytoplasmic IL-10 following L�PIMstimulation for 5 h, but were significantly less common withinthe CD1dhighCD5�, CD1dlowCD5�, or CD1dlowCD5� B cellsubsets ( p � 0.01). Whether IL-10-competent B cells representa heterogeneous population capable of producing other cyto-kines was therefore examined by purifying IL-10-secretingCD19� B cells from hCD19Tg mice (Fig. 1D). IL-10 transcriptswere expressed at �6-fold higher frequencies in IL-10-secret-ing B cells when compared with B cells that did not secretedetectable IL-10 (Fig. 1E). Furthermore, IL-10� B cells did notproduce transcripts for 31 additional cytokines at levels higherthan IL-10� B cells under these culture conditions. Thus, theIL-10-secreting CD1dhighCD5� B10 cell subset was phenotyp-ically and functionally unique.
B10 cell numbers during development
To characterize B10 cell development, the frequencies andnumbers of spleen CD1dhighCD5� B cells and IL-10-producingB cells were assessed in neonatal, 2-mo-old, and 6-mo-old wild-type B6 mice. CD1dhighCD5� B cells were virtually absent inneonatal spleen, with 5-fold lower frequencies than in 2-mo-oldmice (Fig. 2A). Remarkably, neonatal spleen had 6.8-foldhigher frequencies of IL-10-producing B cells than the 1–2%frequency induced in 2-mo-old wild-type spleen B cells follow-ing 5-h L�PIM stimulation (Fig. 2B). Nonetheless, the majority
of IL-10� B cells in neonates had a CD1dlowCD5� phenotype,with 6.5-fold higher CD5 expression levels than IL-10� B cells( p � 0.001; Fig. 2C). Conversely, the frequencies and numbersof CD1dhighCD5� B cells were 1.4- and 1.8-fold higher in6-mo-old mice than in 2-mo-old mice. IL-10� B cell frequen-cies and numbers were also 1.8- and 1.6-fold higher, respec-tively, in 6-mo-old mice compared with 2-mo-old mice ( p �0.05). Spleen IL-10� B cells from 2- and 6-mo-old mice werepredominantly CD1dhighCD5�. Thus, neonatal IL-10-producingCD1dlowCD5� B cells were present at relatively high frequen-cies and numbers, whereas CD1dhighCD5� B10 cells expandedwith age in the spleens of adult mice.
B10 cell development is T cell and pathogen independent
To identify factors that influence B10 cell development,CD1dhighCD5� and IL-10-producing B cells were assessed in Tcell-deficient nude mice and in gnotobiotic mice. CD1dhighCD5�
B cell frequencies and numbers were �5-fold higher in adult nudemice than in age-matched wild-type mice ( p � 0.05; Fig. 3A).Cytoplasmic IL-10� B cell frequencies and numbers were also�4.5-fold higher in L�PIM-stimulated splenocytes from nudemice when compared with wild-type mice ( p � 0.05; Fig. 3B).The majority of IL-10� B cells in nude and wild-type mice had aCD1dhighCD5� phenotype, whereas IL-10� B cells wereCD1dlowCD5� (Fig. 3C). Whether B cell IL-10 production in vitrowas influenced by the presence of T cells was also assessed byculturing whole splenocytes or purified B cells alone with L�PIMfor 5 h. The frequency of B cells that expressed cytoplasmic IL-10
FIGURE 2. B10 cell development in neonatal and 2- or 6-mo-old wild-type B6 mice. A, Representative CD1d and CD5 expression by CD19� B cells.Splenocytes were stained with CD1d, CD5, and CD19 mAbs with flow cytometry analysis of cells. Results represent one mouse indicating the frequencyof CD1dhighCD5� B cells among total B cells within the indicated gates. Bar graphs indicate mean (�SEM) percentages and numbers of CD1dhighCD5�
B cells in one of two independent experiments with three mice in each group. B, IL-10 production by B cells. Splenocytes were cultured with L�PIM for5 h, then stained with CD19 mAb to identify B cells, permeabilized, and stained using IL-10 mAb with flow cytometry analysis. Representative resultsdemonstrate the frequency of IL-10-producing cells among total CD19� B cells within the indicated gates. Bar graphs indicate mean (�SEM) percentagesand numbers of B cells that produced IL-10 in one of two independent experiments with three mice in each group. A and B, Significant differences betweensample means are indicated, as follows: �, p � 0.05; ��, p � 0.01. C, Representative CD1d and CD5 expression by IL-10� or IL-10� B cells from neonatalmice. Horizontal and vertical gates delineate background staining using unreactive isotype-matched control mAbs.
7462 REGULATORY B10 CELL DEVELOPMENT
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

among all B cells was comparable in both cultures (Fig. 3D). Thus,spleen B10 cell development does not require the presence of Tcells in nude mice.
To determine whether environmental factors influence B10cell development, germfree mice were assessed. CD1dhigh
CD5� B cell frequencies and numbers were similar, if not
FIGURE 3. B10 cell development in T cell-defi-cient and gnotobiotic mice. A, CD1d and CD5 ex-pression by spleen CD19� B cells from 2-mo-oldwild-type and nude mice. Results represent onemouse indicating the frequency of CD1dhighCD5� Bcells within the indicated gates among total B cells.Bar graphs indicate mean (�SEM) percentages andnumbers of CD1dhighCD5� B cells in one of twoindependent experiments with three mice in eachgroup. B, IL-10 production by B cells from wild-type and nude mice. Splenocytes were cultured withL�PIM for 5 h, stained with CD19 mAb, perme-abilized, and stained using IL-10 mAb with flowcytometry analysis. Representative results demon-strate the frequency of IL-10-producing cells withinthe indicated gates among total CD19� B cells. Bargraphs indicate mean (�SEM) percentages andnumbers of B cells that produced IL-10 in one oftwo independent experiments with three mice ineach group. C, CD1d and CD5 expression by IL-10� or IL-10� B cells from wild-type and nudemice. Data are representative of two independentexperiments with three mice in each group. Hori-zontal and vertical gates delineate background stain-ing using unreactive isotype-matched control mAbs.D, The presence of T cells during in vitro culturesdoes not influence B cell IL-10 production. Wild-type splenocytes or purified B220� B cells were cul-tured with L�PIM for 5 h, then stained with CD19mAb, permeabilized, and stained using IL-10 mAbwith flow cytometry analysis. Representative resultsdemonstrate the frequency of IL-10-producing cellswithin the indicated gates among total CD19� Bcells. Bar graphs indicate mean (�SEM) percent-ages and numbers of B cells that produced IL-10 inone of two independent experiments with three micein each group. E, CD1d and CD5 expression byspleen CD19� B cells from specific pathogen-free(SPF) and gnotobiotic mice. Bar graphs indicatemean (�SEM) percentages and numbers ofCD1dhighCD5� B cells in three mice. F, IL-10 pro-duction by B cells from specific pathogen-free(SPF) and gnotobiotic mice, cultured as in B. Bargraphs indicate mean (�SEM) percentages andnumbers of B cells that produced IL-10 in threemice. A, B, and D–F, Significant differences be-tween sample means are indicated, as follows:�, p � 0.05; ��, p � 0.01.
7463The Journal of Immunology
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

identical, in age-matched mice reared in gnotobiotic and spe-cific pathogen-free colonies (Fig. 3E). Cytoplasmic IL-10� Bcell frequencies and numbers were also similar (Fig. 3F), andthe majority of IL-10� B cells had a CD1dhighCD5� phenotype(data not shown). Thus, environmental flora and gut-associatedbacteria are not required for spleen B10 cell development.
Autoimmunity promotes B10 cell development
The influence of autoimmunity on B10 cell development was as-sessed in the NOD, NZB/W F1, MRL/lpr, DBA/1, and SJL mousestrains. NOD mice are a spontaneous model of type 1 diabetes(36). DBA/1 mice develop CIA after collagen immunization (37).SJL mice are susceptible to EAE after myelin proteolipid proteinimmunization (38). MRL/lpr and NZB/W mice spontaneously de-velop lupus-like disease (39). Most B cells in NOD (85 � 2%, n�3), MRL/lpr (80 � 12%, n � 3), and SJL (94 � 1%, n � 3) miceexpressed cell surface CD5 at levels that were significantly higherthan background control mAb staining in comparison with B cells
from B6 (25 � 2%, n � 3), NZB/W (28 � 1%, n � 3), and DBA/1(14 � 1%, n � 3) mice in side-by-side comparisons (Fig. 4A).Nonetheless, the frequency of CD1dhighCD5� B cells was limited,but 3- to 9-fold higher in NZB/W, MRL/lpr, NOD, and SJL micethan in 2-mo-old B6 mice. CD1dhighCD5� B cell numbers werealso 3.8- to 5.9-fold increased in NZB/W, MRL/lpr, and NODmice. Thus, the CD1dhighCD5� B cell subset increased in fre-quency in mice predisposed to autoimmunity.
The numbers of cytoplasmic IL-10� B cells were 2- to 4-foldhigher in NZB/W, MRL/lpr, and NOD mice than in B6 wild-typemice after L�PIM stimulation (Fig. 4B). By contrast, IL-10-pro-ducing B cell numbers were 49 and 55% lower in DBA/1 and SJLmice, respectively, relative to wild-type mice ( p � 0.01). In allcases, the majority of cytoplasmic IL-10� B cells also retained aCD1dhighCD5� phenotype (Fig. 4C). Thus, B10 cell numbers weresignificantly higher in diabetes- and lupus-prone mice, but signif-icantly below wild-type levels in DBA/1 and SJL mice that are sus-ceptible to exogenous autoantigen-induced autoimmune disease.
FIGURE 4. Autoimmunity promo-tes B10 cell expansion. A, CD1d andCD5 expression by spleen B cellsfrom 2-mo-old wild-type B6, DBA/1,and SJL/J mice, 3-mo-old NOD mice,and 4-mo-old NZB/W F1 and MRL/lpr mice. Representative results dem-onstrate the frequency of CD1dhigh
CD5� B cells within the indicatedgates among total CD19� B cells.Horizontal and vertical gates are setto delineate the CD1dhighCD5� B cellsubset. Bar graphs indicate mean(�SEM) percentages and numbers ofCD1dhighCD5� B cells in one of twoindependent experiments with threemice in each group. B, IL-10 produc-tion by B cells. Splenocytes were cul-tured with L�PIM for 5 h, thenstained with CD19 mAb, permeabil-ized, and stained using IL-10 mAbwith flow cytometry analysis. Repre-sentative results demonstrate the fre-quency of IL-10-producing cellswithin the indicated gates among totalB cells. Bar graphs indicate mean(�SEM) percentages and numbers ofB cells that produced IL-10 in one oftwo independent experiments withthree mice in each group. A and B,Significant differences between sam-ple means are indicated, as follows: �,p � 0.05; ��, p � 0.01. C, CD1d andCD5 expression by IL-10� or IL-10�
B cells. Horizontal and vertical gatesare set to delineate the CD1dhigh
CD5� B cell subset, as in A. Data arerepresentative of two independent ex-periments with three mice in eachgroup.
7464 REGULATORY B10 CELL DEVELOPMENT
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

Receptors that regulate B10 cell development in vivo
B cell development is regulated through the BCR and other mol-ecules that inform B cells of their extracellular microenvironment,including CD19, CD21, CD22, and CD40 (40). Whether cell sur-face signals influence B10 cell development was determined byassessing CD1dhighCD5� and IL-10-producing B cell develop-ment in IL-10�/�, MD4, CD19�/�, CD21�/�, CD40�/�, MHC-I/II�/�, hCD19Tg, CD22�/�, CD40L/BTg, and CD40L/BTg/CD22�/� mice. MD4 transgenic mice have a fixed BCR specificfor hen egg lysozyme (28). MHC-I/II�/� mice are deficient in cellsurface MHC class II, and most MHC class I and CD1 moleculesdue to combined disruption of the H2-Ab1 and �2-microglobulingenes (29, 41). B cells from CD40L/BTg mice express ectopic cellsurface CD40L constitutively, with some mice developing lupus-like disease (35).
CD1dhighCD5� B cells were present at similar frequencies andnumbers in IL-10�/�, wild-type, and MD4 mice (Fig. 5A). How-ever, both the frequencies (65% decrease, p � 0.01) and numbers(90% decrease, p � 0.01) of L�PIM-induced cytoplasmic IL-10�
B cells were reduced in MD4 mice when compared with wild-typemice (Fig. 5B). In CD19�/� mice, the frequency and number ofCD1dhighCD5� B cells were 87–92% lower than in wild-type lit-termates, whereas L�PIM-induced IL-10� B cell frequencies andnumbers were 73 and 89% lower, respectively ( p � 0.01). Bycontrast, CD21 or CD40 deficiencies did not affect the frequenciesor numbers of CD1dhighCD5� or IL-10-producing B cells.CD1dhighCD5� B cell frequencies could not be assessed in MHCI/II�/� mice that do not express CD1d, but IL-10-producing spleenB cell frequencies and numbers were normal. SpleenCD1dhighCD5� B cell frequencies and numbers were normal inMyD88�/� mice (Fig. 5A), whereas L�PIM-induced cytoplasmicIL-10� B cell frequencies and numbers were reduced by 40 and46%, respectively, in 5-h assays (Fig. 5B). Thus, BCR diversityand CD19- and MyD88-generated signals were critical for normalIL-10-producing CD1dhighCD5� B10 cell development and/or pe-ripheral expansion in vivo, or visualization in vitro.
The frequencies and numbers of CD1dhighCD5� B cells were5.8- and 1.5-fold higher in hCD19Tg mice than in wild-type lit-termates, respectively (Fig. 5A). IL-10-producing B cell frequen-cies and numbers were 7.9- and 2.1-fold higher in hCD19Tg mice,respectively (Fig. 5B). Similarly, the frequency and number ofCD1dhighCD5� B cells were 2.7- and 1.9-fold higher in CD22�/�
mice than in wild-type mice, whereas the frequency and number ofIL-10-producing B cells were 4.1- and 2.8-fold higher, respec-tively. The frequency and number of CD1dhighCD5� B cells were1.4- and 3.9-fold higher in CD40L/BTg mice than in wild-typemice, whereas the frequency and number of IL-10-producing Bcells were 1.4- and 3.7-fold higher, respectively. Thus, CD19 over-expression, CD22 deficiency, and ectopic CD40L expression on Bcells significantly enhanced B10 cell numbers in vivo.
Combined CD22 deficiency and CD40L expression dramaticallyexpanded the B10 cell subset in CD40L/BTg/CD22�/� mice (Fig. 5).The frequency and number of CD1dhighCD5� B cells were 7.0- and16-fold higher in CD40L/BTg/CD22�/� mice, whereas the frequencyand number of IL-10-producing B cells were 11- and 26-foldhigher in CD40L/BTg/CD22�/� mice than in wild-type mice, re-spectively ( p � 0.01). Thus, the absence of CD22 regulation com-bined with CD40L expression by B cells dramatically increasedB10 cell numbers in vivo. In all mouse lines except MHC-I/II�/�
mice, L�PIM-induced IL-10� B cells maintained aCD1dhighCD5� phenotype when present (data not shown). Thus,spleen B10 cell development or expansion in vivo is not intrinsic,but depends in part on transmembrane signals.
LPS and CD40 stimulation induce B cell cytoplasmic IL-10production in vitro
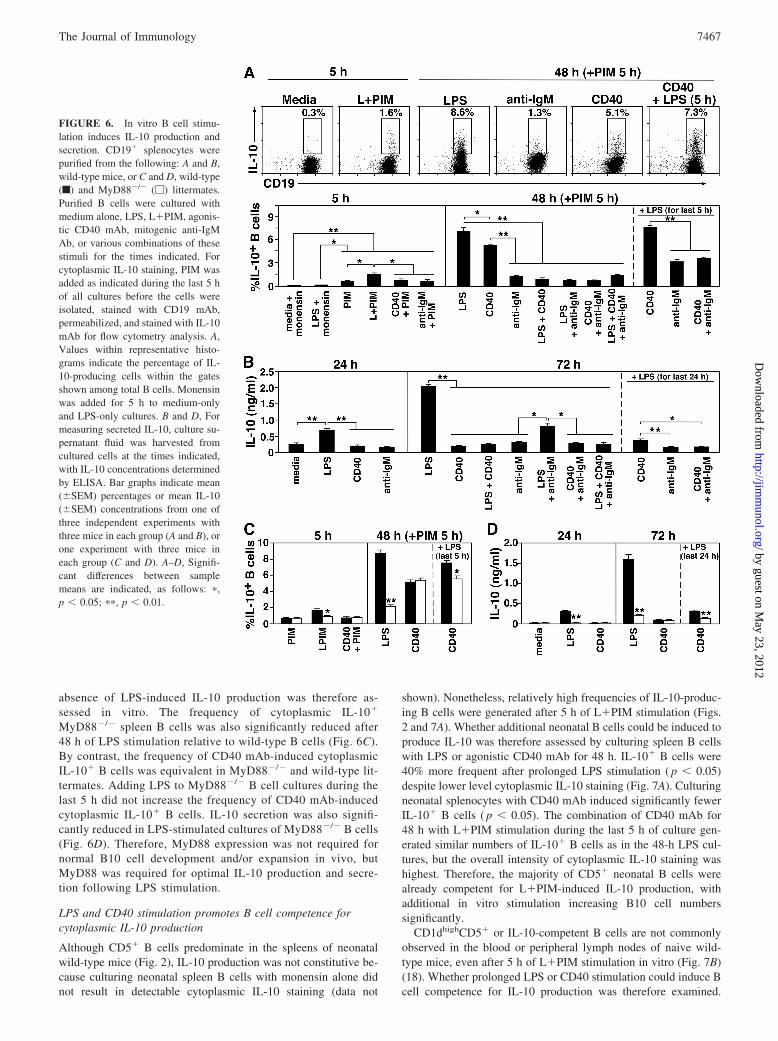
Signals that regulate B cell IL-10 production were assessed byculturing wild-type spleen B cells with LPS, agonistic CD40 mAb,or mitogenic anti-IgM Ab at predetermined optimal concentrations(data not shown). PMA, ionomycin, and monensin (PIM) stimu-lation for 5 h induced cytoplasmic IL-10 expression by 0.5–2% ofB cells, which was 8- to 13-fold higher than for medium alone and�5-fold higher than for LPS alone (Fig. 6A). The addition ofCD40 mAb or anti-IgM Ab to PIM-stimulated cultures did notsignificantly increase IL-10� B cell frequencies. However,L�PIM stimulation for 5 h induced �2-fold higher frequencies ofIL-10� B cells than PIM, or CD40 mAb plus PIM, or anti-IgM Abplus PIM ( p � 0.01). Thus, L�PIM stimulation induced optimalB cell cytoplasmic IL-10 expression in 5-h assays.
Culturing B cells with LPS or CD40 mAb for 48 h with PIMadded during the last 5 h of culture induced significantly higherfrequencies of cytoplasmic IL-10� B cells than anti-IgM Ab withPIM added during the last 5 h of culture (Fig. 6A). LPS stimulationwas also significantly more robust than CD40 mAb stimulation.Unexpectedly, however, the combination of LPS plus CD40 mAbfor 48 h, or anti-IgM Ab plus either LPS or CD40 mAb, or all threetogether with PIM stimulation during the last 5 h did not increaseIL-10� B cell frequencies significantly beyond what was normallyobserved with 5-h PIM stimulation alone. Thus, culturing B cellswith LPS or CD40 mAb for 48 h before PIM stimulation inducedthe highest numbers of B cells with cytoplasmic IL-10 expression.
Spleen B cells stimulated with CD40 mAb for 48 h plus L�PIMfor 5 h did not induce significantly higher numbers of cytoplasmicIL-10� B cells than LPS for 48 h plus PIM for 5 h (Fig. 6A).However, this sequential combination of stimuli induced the mostrobust levels of cytoplasmic IL-10 expression when comparedwith independent LPS or CD40 mAb stimulation. By contrast,adding L�PIM during the last 5 h of anti-IgM Ab, or CD40 mAbplus anti-IgM Ab cultures only induced �2-fold higher numbers ofIL-10� B cells than anti-IgM Ab or CD40 mAb alone. Thus, CD40ligation with subsequent 5-h L�PIM stimulation was the mostpotent strategy for inducing the highest numbers of cytoplasmicIL-10� B cells with the highest levels of cytoplasmic IL-10.
LPS, but not BCR or CD40 ligation induces B cell IL-10secretion in vitro
Signals that regulate B cell IL-10 secretion were assessed by cul-turing spleen B cells with LPS, agonistic CD40 mAb, or mitogenicanti-IgM Ab, with culture supernatant fluid IL-10 levels deter-mined by ELISA. LPS stimulation of spleen B cells for 24 h in-duced 3.5- to 3.8-fold more IL-10 than unstimulated cells, or cellscultured with CD40 mAb or anti-IgM Ab ( p � 0.01; Fig. 6B). LPSstimulation alone for 72 h induced significant B cell IL-10 secre-tion in contrast to CD40 mAb, anti-IgM Ab, or CD40 mAb plusanti-IgM Ab ( p � 0.01). In fact, simultaneous CD40 mAb oranti-IgM Ab treatment reduced LPS-induced IL-10 secretion by�68%. Furthermore, B cells cultured with CD40 mAb, anti-IgMAb, and CD40 mAb plus anti-IgM Ab did not secrete significantlymore IL-10 when LPS was added during the last 24 h of culture.Thus, LPS was the most potent stimulus for inducing both IL-10production and secretion, whereas CD40-generated signals pro-moted cytoplasmic IL-10 generation, but inhibited its secretion.
Normal B10 cell development in MyD88�/� mice
L�PIM-induced cytoplasmic IL-10� B cell frequencies andnumbers were reduced in MyD88�/� mice (Fig. 5B). Whetherthis represented a developmental defect in vivo or reflected the
7465The Journal of Immunology
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from
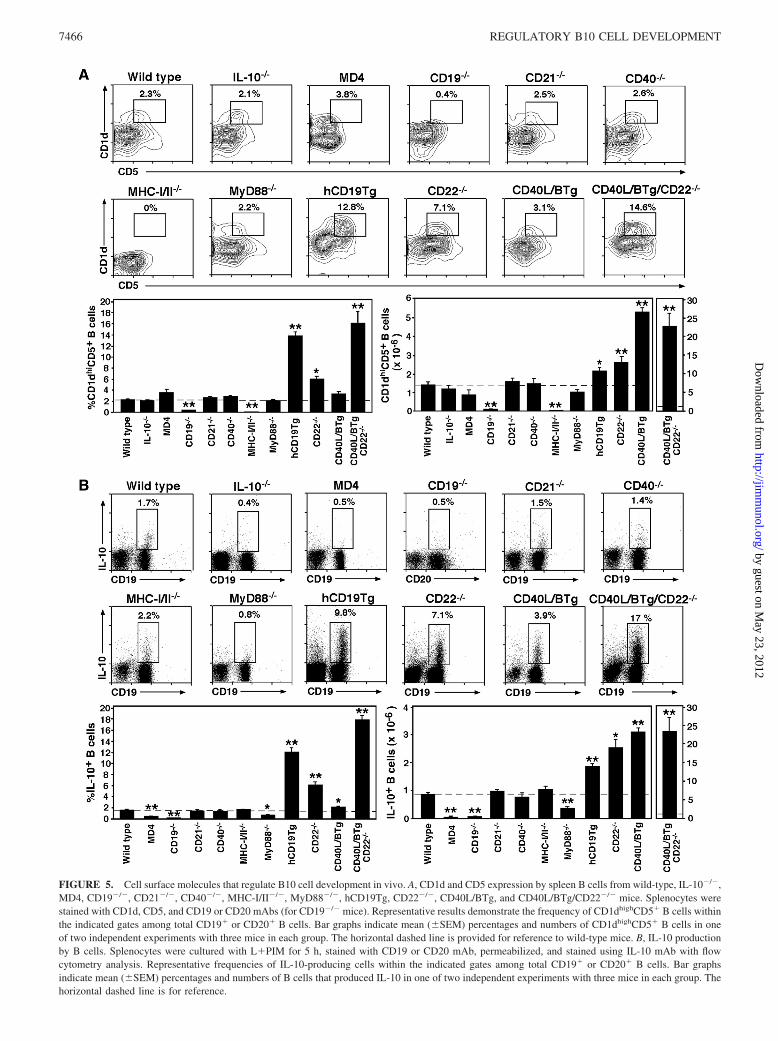
FIGURE 5. Cell surface molecules that regulate B10 cell development in vivo. A, CD1d and CD5 expression by spleen B cells from wild-type, IL-10�/�,MD4, CD19�/�, CD21�/�, CD40�/�, MHC-I/II�/�, MyD88�/�, hCD19Tg, CD22�/�, CD40L/BTg, and CD40L/BTg/CD22�/� mice. Splenocytes werestained with CD1d, CD5, and CD19 or CD20 mAbs (for CD19�/� mice). Representative results demonstrate the frequency of CD1dhighCD5� B cells withinthe indicated gates among total CD19� or CD20� B cells. Bar graphs indicate mean (�SEM) percentages and numbers of CD1dhighCD5� B cells in oneof two independent experiments with three mice in each group. The horizontal dashed line is provided for reference to wild-type mice. B, IL-10 productionby B cells. Splenocytes were cultured with L�PIM for 5 h, stained with CD19 or CD20 mAb, permeabilized, and stained using IL-10 mAb with flowcytometry analysis. Representative frequencies of IL-10-producing cells within the indicated gates among total CD19� or CD20� B cells. Bar graphsindicate mean (�SEM) percentages and numbers of B cells that produced IL-10 in one of two independent experiments with three mice in each group. Thehorizontal dashed line is for reference.
7466 REGULATORY B10 CELL DEVELOPMENT
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from
absence of LPS-induced IL-10 production was therefore as-sessed in vitro. The frequency of cytoplasmic IL-10�
MyD88�/� spleen B cells was also significantly reduced after48 h of LPS stimulation relative to wild-type B cells (Fig. 6C).By contrast, the frequency of CD40 mAb-induced cytoplasmicIL-10� B cells was equivalent in MyD88�/� and wild-type lit-termates. Adding LPS to MyD88�/� B cell cultures during thelast 5 h did not increase the frequency of CD40 mAb-inducedcytoplasmic IL-10� B cells. IL-10 secretion was also signifi-cantly reduced in LPS-stimulated cultures of MyD88�/� B cells(Fig. 6D). Therefore, MyD88 expression was not required fornormal B10 cell development and/or expansion in vivo, butMyD88 was required for optimal IL-10 production and secre-tion following LPS stimulation.
LPS and CD40 stimulation promotes B cell competence forcytoplasmic IL-10 production
Although CD5� B cells predominate in the spleens of neonatalwild-type mice (Fig. 2), IL-10 production was not constitutive be-cause culturing neonatal spleen B cells with monensin alone didnot result in detectable cytoplasmic IL-10 staining (data not
shown). Nonetheless, relatively high frequencies of IL-10-produc-ing B cells were generated after 5 h of L�PIM stimulation (Figs.2 and 7A). Whether additional neonatal B cells could be induced toproduce IL-10 was therefore assessed by culturing spleen B cellswith LPS or agonistic CD40 mAb for 48 h. IL-10� B cells were40% more frequent after prolonged LPS stimulation ( p � 0.05)despite lower level cytoplasmic IL-10 staining (Fig. 7A). Culturingneonatal splenocytes with CD40 mAb induced significantly fewerIL-10� B cells ( p � 0.05). The combination of CD40 mAb for48 h with L�PIM stimulation during the last 5 h of culture gen-erated similar numbers of IL-10� B cells as in the 48-h LPS cul-tures, but the overall intensity of cytoplasmic IL-10 staining washighest. Therefore, the majority of CD5� neonatal B cells werealready competent for L�PIM-induced IL-10 production, withadditional in vitro stimulation increasing B10 cell numberssignificantly.
CD1dhighCD5� or IL-10-competent B cells are not commonlyobserved in the blood or peripheral lymph nodes of naive wild-type mice, even after 5 h of L�PIM stimulation in vitro (Fig. 7B)(18). Whether prolonged LPS or CD40 stimulation could induce Bcell competence for IL-10 production was therefore examined.
FIGURE 6. In vitro B cell stimu-lation induces IL-10 production andsecretion. CD19� splenocytes werepurified from the following: A and B,wild-type mice, or C and D, wild-type(f) and MyD88�/� (�) littermates.Purified B cells were cultured withmedium alone, LPS, L�PIM, agonis-tic CD40 mAb, mitogenic anti-IgMAb, or various combinations of thesestimuli for the times indicated. Forcytoplasmic IL-10 staining, PIM wasadded as indicated during the last 5 hof all cultures before the cells wereisolated, stained with CD19 mAb,permeabilized, and stained with IL-10mAb for flow cytometry analysis. A,Values within representative histo-grams indicate the percentage of IL-10-producing cells within the gatesshown among total B cells. Monensinwas added for 5 h to medium-onlyand LPS-only cultures. B and D, Formeasuring secreted IL-10, culture su-pernatant fluid was harvested fromcultured cells at the times indicated,with IL-10 concentrations determinedby ELISA. Bar graphs indicate mean(�SEM) percentages or mean IL-10(�SEM) concentrations from one ofthree independent experiments withthree mice in each group (A and B), orone experiment with three mice ineach group (C and D). A–D, Signifi-cant differences between samplemeans are indicated, as follows: �,p � 0.05; ��, p � 0.01.
7467The Journal of Immunology
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

LPS or agonistic CD40 mAb stimulation induced 6- to 9-foldhigher frequencies of cytoplasmic IL-10� B cells in 48-h culturesthan in 5-h L�PIM cultures ( p � 0.01; Fig. 7B). The combinationof CD40 mAb for 48 h with L�PIM stimulation during the last 5 hof culture also generated high numbers of IL-10� B cells with thehighest intensity of cytoplasmic IL-10 staining. Similar resultswere obtained using peripheral lymph node B cells (data notshown). These results suggest that prolonged LPS or CD40 stim-ulation can promote the maturation of CD5� progenitor B10 cellsinto an IL-10-competent state.
Whether LPS- or CD40-generated signals induce B cells to ex-press a CD1dhighCD5� phenotype was therefore assessed. Neona-tal spleen, and adult blood and spleen B cells were cultured withLPS or agonistic CD40 mAb for 48 h and examined for CD1d andCD5 expression by immunofluorescence staining. CD40 mAb, butnot LPS stimulation induced markedly higher CD5 expression onmost B cells (Fig. 7C). By contrast, B cell CD1d expression was
not induced or changed by LPS or CD40 mAb stimulation or thecombination of both treatments for 48 h. Thus, CD5 was an in-duced marker for CD40-stimulated B10 cells.
IL-10 production by adult spleen B cells is restricted to theCD1dhighCD5� B cell subset
Splenic B10 cells that express cytoplasmic IL-10 after L�PIMstimulation localize primarily within the CD1dhighCD5� subset(Fig. 1A). It was therefore determined whether the increased fre-quency of IL-10� B cells in LPS- or CD40-stimulated culturesresults from the maturation of B10 cell progenitor cells within theCD1dhighCD5� subset or other B cell populations. SpleenCD1dhighCD5� or non-CD1dhighCD5� B cells from wild-typemice were purified and cultured with LPS for 48 h, or with ago-nistic CD40 mAb for 48 h with LPS added during the last 5 h ofculture. The CD1dhighCD5� B cell subset from B6 mice normallycontains �9–18% IL-10� B cells after 5 h of L�PIM stimulation
FIGURE 7. LPS and CD40 signalsinduce the maturation of B10 progen-itor cells. LPS and CD40 mAb induceIL-10 production by A, neonatalspleen or B, adult blood B cells fromwild-type mice. A and B, Cells werecultured with LPS, agonistic CD40mAb, or both for the times indicated,with PIM added during the last 5 h ofeach culture. The cultured cells wereisolated, stained with CD19 mAb,permeabilized, and stained usingIL-10 mAb with flow cytometry anal-ysis. Values within representative his-tograms indicate the percentage of IL-10-producing cells among CD19� Bcells within the gates shown. Bargraphs indicate mean (�SEM) per-centages of IL-10-producing B cellsin one of two independent experi-ments with three mice in each group.Significant differences between sam-ple means are indicated, as follows: �,p � 0.05; ��, p � 0.01. C, CD40stimulation induces B cell CD5 ex-pression. Cell surface CD1d and CD5expression by wild-type CD19� cellswas determined by immunofluores-cence staining. Neonatal splenocytes,or adult blood and spleen B cells werefreshly isolated, or cultured for 48 hwith LPS or agonistic CD40 mAb(plus or minus LPS for the last 5 h ofculture). Values indicate the percent-age of CD1dhighCD5� B cells amongtotal B cells within the indicatedgates. Single-color histograms arerepresentative of two independent ex-periments with three mice in eachgroup.
7468 REGULATORY B10 CELL DEVELOPMENT
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

(Fig. 1A) (18). However, 33–43% of the CD1dhighCD5� B cellsexpressed cytoplasmic IL-10 after 48-h LPS or CD40 mAb stim-ulation, whereas �3% of CD1dlowCD5� B cells produced IL-10(Fig. 8A). Thus, splenic B cells capable of producing IL-10 afterprolonged LPS or CD40 mAb stimulation predominantly derivefrom the CD1dhighCD5� subset.
To determine whether the increased frequency of IL-10� B cellsafter LPS or CD40 stimulation results from the clonal expansion ofexisting IL-10-competent B cells or maturation of progenitor B10cells, IL-10� B cell proliferation was assessed by labeling purified
spleen B cells with CFSE before LPS or CD40 mAb stimulation invitro. LPS stimulation for 48 h induced IL-10� and IL-10� B cellproliferation, although IL-10� B cells proliferated more than IL-10� B cells, as measured by reduced CFSE staining (Fig. 8B). Bycontrast, CD40 mAb stimulation for 48 h plus LPS treatment forthe last 5 h of culture only induced modest IL-10� or IL-10� Bcell proliferation during these 48-h cultures. CD40 mAb stimula-tion predominantly induces B cell clonal expansion between 72and 96 h, as described (31, 42). Thus, LPS stimulation induces andexpands the IL-10� B cell subset during 48-h cultures, whereasCD40 ligation induces B cell competence for cytoplasmic IL-10production (Fig. 8C).
DiscussionThe majority of adult spleen B cells that were competent for IL-10production after 5-h L�PIM stimulation were found within theCD1dhighCD5� subset (Fig. 1A). IL-10� B10 cells preferentiallyproduced IL-10 transcripts relative to other B cells, but did notappear to preferentially produce other known cytokines (Fig. 1E).IL-10-competent B cells were also found within theCD1dhighCD5� and CD1dintCD5� subsets, but at significantlylower ( p � 0.05) frequencies and numbers than in theCD1dhighCD5� subset. Spleen CD1dhighCD5� B cells also existthat could acquire IL-10 competence in vitro after 48-h stimulationwith LPS or agonistic CD40 mAb (Figs. 6A and 8A), potentiallyreflecting their maturation. By contrast, spleen CD1dlowCD5� Bcells were not rendered IL-10 competent after 48-h stimulationwith LPS or agonistic CD40 mAb (Fig. 8A). Progenitor B10 cellsmay also exist that do not express CD5 or CD1d, yet can be in-duced to express IL-10 in vitro. Specifically, the vast majority ofblood and lymph node B cells in adult mice were CD1dlowCD5�
and did not express IL-10 after 5-h L�PIM stimulation (Fig. 7B).However, a small subset of blood and lymph node B cells acquiredIL-10 competence after 48-h CD40 ligation and/or LPS exposure.Neonatal spleen B cells predominantly expressed CD5 and werealmost exclusively CD1dlow, but �14% were induced to expresscytoplasmic IL-10 after 5-h L�PIM exposure (Fig. 2). Consistentwith this, neonatal and adult B cells uniformly up-regulated CD5expression after CD40 ligation in vitro (Fig. 7C). Thereby, L�PIMstimulation may induce IL-10 production in small subsets of Bcells that have received appropriate competence-inducing signalsin vivo or in vitro regardless of their maturation. Alternatively,CD1dlowCD5� progenitor B10 cells may be induced to mature,express CD5, and acquire competence for activation-induced cy-toplasmic IL-10 production as proposed in the maturation schemeoutlined in Fig. 8C. Factors that regulate or induce CD1d expres-sion by some spleen B cells are unknown. Thus, IL-10 competenceand the CD1dhighCD5� phenotype define the spleen B10 cell sub-set, but may also reflect their maturation, activation status, subsetcommitment, and/or tissue localization.
Development, maturation, and/or expansion of the spleen B10cell subset required specific external signals. BCR specificity sig-nificantly influenced B10 cell development, with B10 cell numbersreduced by 90% in transgenic mice expressing a fixed Ag receptor(Fig. 5B). In contrast, B10 cell development did not require thepresence of T or NKT cells (Fig. 3). Furthermore, CD1, MHCclass I and class II, CD21, or CD40 expression were not requiredfor normal B10 cell development or IL-10 induction (Fig. 5).Nonetheless, CD40 ligation induced cytoplasmic IL-10 productionby B cells in vitro (Fig. 6), and ectopic CD154 expression by Bcells in CD40L/BTg mice increased B10 cell numbers by 3- to4-fold (Fig. 5). Thereby, CD40:CD154 interactions may facilitateB10 cell maturation under some conditions, but were not required
FIGURE 8. Effect of LPS or CD40 ligation on IL-10 production, pro-liferation, and the phenotype of CD1dhighCD5� B cells. A, LPS and CD40mAb-induced cytoplasmic IL-10 production are restricted toCD1dhighCD5� B cells. CD1dhighCD5� or CD1dlowCD5� B220� B cellswere purified from pooled splenocytes of three wild-type mice by cellsorting and reassessed for CD1d and CD5 expression (middle panels). Thepurified B cell subsets were cultured with LPS or CD40 mAb for 48 h, withL�PIM added for the last 5 h of culture before permeabilization, stainingfor IL-10, and flow cytometry analysis (right panels). The frequencies ofIL-10� cells among the sorted CD1dhighCD5� or CD1dlowCD5� B cellsubsets are shown for one of two independent experiments. B, Clonal ex-pansion of IL-10-producing B cells after LPS, but not CD40 stimulation invitro for 48 h. Wild-type CD19� splenocytes were labeled with CFSE andcultured with LPS or CD40 mAb for 48 h, with L�PIM added for the last5 h of culture. Histograms (right) represent CFSE expression by the IL-10�
or IL-10� B cell subsets. Dashed lines represent CFSE staining of un-stimulated B cells. A and B, Data are representative of two independentexperiments. C, Potential B10 developmental pathway leading to the gen-eration of the IL-10-secreting B10 cell subset. Dashed arrows and questionmarks represent potential maturation steps based on CD5 and CD1d ex-pression patterns. LN, lymph node; B10pro, progenitor B10 cell.
7469The Journal of Immunology
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

for B10 cell acquisition of IL-10 competence in vivo. TLR sig-naling was also critical for B10 cell effector function because LPSinduced B10 cells to both produce and secrete IL-10 in vitro,whereas CD40 ligation only induced cytoplasmic IL-10 production(Fig. 6). B10 cell development was normal in MyD88�/� mice(Fig. 6, C and D), but LPS-induced IL-10 production and secretionwere significantly reduced in MyD88�/� B cells (Fig. 6D). A needfor MyD88 in LPS-induced B10 cell function may explain whymice containing only MyD88�/� B cells develop chronic EAE(43). Thus, intertwined innate and adaptive signals may regulateB10 cell maturation and effector function rather than indepen-dently regulating distinct follicular, MZ, and B-1a regulatory Bcell subsets.
The B10 cell subset expanded significantly in response to en-hanced B cell signaling in vivo, while retaining theirCD1dhighCD5� phenotype. B10 cell numbers were significantlyexpanded in hCD19Tg mice, but were dramatically reduced inCD19�/� mice (Fig. 5) (18). B10 cell numbers were also increased2- to 3-fold in CD22�/� mice (Fig. 5). CD19 regulates a Lynkinase amplification loop (44, 45) that enhances transmembranesignals (27, 46, 47), whereas CD22 dampens B cell and CD19signal transduction through the recruitment of Src homology re-gion 2 domain-containing phosphatase 1 and SHIP phosphatases(44, 46), resulting in elevated cell surface CD5 expression by Bcells in CD22�/� B6 mice (48). Spleen B10 cells were also sig-nificantly expanded in CD40L/BTg mice, with a 26-fold increasein CD22�/�CD40L/BTg mice, in which up to 20% of spleen Bcells were B10 cells (Fig. 5). Because CD22 negatively regulatesCD40 signaling (31, 48), enhanced CD40 function may drive B10cell expansion and/or survival in CD22�/�CD40L/BTg mice (35,49). Although spleen B1a cells were also expanded in hCD19Tg(3-fold), CD40L/BTg (4.2-fold), and CD22�/�CD40L/BTg (3-fold) mice (data not shown), these frequencies did not parallel B10cell expansion. Thus, the B10 cell subset responds significantly totransmembrane signals in vivo.
Spleen B10 cell numbers were increased in mice predisposed todevelop autoimmunity. B10 cell numbers expanded significantlyin the NZB/W F1 and MRL/lpr mouse models of lupus and theNOD model of diabetes even before obvious autoantibodies andsigns of disease were apparent (Fig. 4; data now shown). B10 cellnumbers are significantly expanded in CD40L/BTg mice (Fig. 5),although some develop lupus-like disease (35). Spleen B10 cellnumbers were also significantly higher in 6-mo-old C57BL/6 micerelative to 2-mo-old mice (Fig. 2), which may combat the devel-opment of autoimmunity with age. By contrast, B10 cell numberswere significantly lower in the DBA/1 and SJL mouse models ofautoantigen-inducible autoimmunity, in which the relative paucityof B10 cells may prevent effective tolerance induction. Thereby,B10 cell expansion may suppress autoimmunity, in contrast to B1acells that contribute to autoimmune disease (50). As a result, theseautoimmune diseases may be worse in the absence of B10 cells, asoccurs when all B cells are depleted during CHS and EAE (18, 19).Because B10 cell numbers are dynamic, change during develop-ment, and increase with age and autoimmunity, alterations in thebalance between B10 cell-negative regulation and B cell-positivecontributions to immune responses are likely to vary in differentdiseases and during the course of disease (4).
Spleen B10 cells and their potential progenitors (Fig. 8C) canaccount for many of the in vivo activities previously attributed toregulatory B cells (4, 18, 19). Specifically, BCR and CD40 en-gagement are required for regulatory B cell functions in CIA,CHS, and EAE models (13, 17–19), and functional B10 cells re-quired diverse BCRs (Fig. 5) and in vivo Ag sensitization (18, 19)for their generation. Stimulating naive or autoimmune spleen B
cells in vitro with LPS or agonistic CD40 mAb also gives rise toregulatory B cells that inhibit or prevent autoimmunity (15, 24).That CD40 ligation induced IL-10 competence in bothCD1dhighCD5� and some CD1dintCD5� B cells (Fig. 6) may alsoexplain how agonistic CD40 mAbs reduce inflammation in theCIA model of rheumatoid arthritis (51). LPS induction of B10 cellcompetence for IL-10 production and secretion (Fig. 6C) may alsoexplain why LPS pretreatment modulates the course of disease inEAE (52). Similarly, B cells activated with LPS in vitro can pro-tect NOD mice in vivo, although this effect was not attributed to Bcell IL-10 production (24). Thus, B10 cells and regulatory B cellsidentified in previous studies were similar in their responses topolyclonal stimuli such as LPS and CD40.
That BCR diversity was required for B10 cell development invivo (Fig. 5) supports observations that B10 cell and regulatory Bcell function is Ag specific (13, 18, 19). The activation of arthri-togenic splenocytes with collagen alone (17) or collagen plus ag-onistic CD40 mAb in vitro gives rise to IL-10-producing B cellsthat prevent arthritis (15). Autoreactive B cell production of IL-10during EAE also requires simultaneous autoantigen and CD40stimulation (13). Transfusions of BCR-activated B cells also pro-tect NOD mice from type 1 diabetes in an IL-10-dependent manner(26). However, BCR ligation using mitogenic Ab in vitro nega-tively regulated cytoplasmic and secreted IL-10 production whencombined with LPS or CD40 mAb during in vitro cultures, al-though BCR ligation alone induced some B cells to express IL-10at higher than background levels (Fig. 6). These results contrastwith the findings of others that BCR ligation using anti-Ig� Abdoes not affect simultaneous LPS-induced IL-10 secretion bysplenic transitional, follicular, and MZ B cells; B1 B cells from theperitoneal cavity; or lymph node B cells (43). However, thestrength, nature, or timing of BCR-generated signals required forevoking B10 cell development or function may be specifically reg-ulated in vivo. For example, BCR engagement by potent foreignAgs may inhibit B10 cell clonal expansion or divert B10 progen-itor cells along a distinct functional pathway, whereas BCR signalsgenerated by self Ags may promote their expansion. Thereby, LPSor other signals may optimally induce B10 cell effector function(IL-10 secretion) after Ag selection or CD40-induced maturationin vivo.
It remains difficult to distinguish the relationships betweenspleen B10, B-1a, and MZ B cells due to their shared phenotypicmarkers and potentially overlapping developmental pathways. Forexample, microbial colonization and conventional T cells were notrequired for spleen B10, B-1a, or CD1dhigh MZ B cell develop-ment, and all three subsets require CD19 expression (Figs. 3 and 5)(53–55). However, spleen CD5� and IL-10-competent B cellswere present at high frequencies in newborns, whereas the splenicCD1dhigh subset was not detectable in newborns (Figs. 1–2), butdevelops between 3 and 7 wk after birth (53). Spleen B10 cellproliferation was also more robust following LPS stimulation thanfor IL-10� B cells (Fig. 8B). MZ B cells also expand and provideprotection early during pathogen challenge (56). Furthermore,some IL-10-producing cells can be induced within the spleenCD1dhighCD5� and CD1dlowCD5� subsets (Fig. 1A), but it is hardto discern whether these cells represent contaminating B10 cells orare progenitor B10 cells that have not fully up-regulated CD1d orCD5 expression (Fig. 8C). Therefore, it is likely that spleen B-1aand MZ B cells represent subsets of mixed origins, with B10 cellsrepresenting either a distinct subset with shared phenotypic mark-ers, or a subset representing different branches of a commonlineage.
7470 REGULATORY B10 CELL DEVELOPMENT
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

These studies address the ambiguity regarding a major B cellsubset that regulates inflammation and autoimmune disease. Evi-dence for the existence of a distinct natural B10 cell subset thatgenerally suppresses immune responses was not uncovered.Rather, the current data indicate that BCR and other signals arecentral to B10 cell generation and that polyclonal signals such asCD40 and LPS can induce their maturation and/or regulatory func-tions. Thereby, immature CD5�/� progenitor B10 cells may beinduced to mature and express CD5 and CD1d through Ag selec-tion, potentially involving CD40 signals that induced CD5 expres-sion (Fig. 7C). BCR ligation is also well characterized to induceCD5 expression (57). That CD40 ligation induces cytoplasmicIL-10 production, but not significant cytokine secretion is likely torepresent another critical regulatory checkpoint in B10 cell func-tion. Although regulatory B cells and B10 cells have been pre-dominantly described in mouse models in which autoantigen plusTLR agonist-containing adjuvants induce autoimmunity, B10cells also significantly influence CHS inflammation, in whichdisease is independent of adjuvant challenge (18). Thus, stimuliin addition to LPS are likely to also regulate IL-10 secretion byB10 cells. Although B10 cell development and tolerance regu-lation are undoubtedly more complex, the current results pro-vide a potential framework (Fig. 8C) for further characterizingB10 cell development.
AcknowledgmentsWe thank Drs. Susan Harless-Smith, Damian Maseda, David DiLillo, andDavid Gray for their help and influential comments, and Dr. Scott Plevy forproviding gnotobiotic mouse tissues.
DisclosuresT.F.T. is a paid consultant for MedImmune and a paid consultant andshareholder for Angelica Therapeutics. The remaining authors have nofinancial conflict of interest.
References1. LeBien, T. W., and T. F. Tedder. 2008. B-lymphocytes: how they develop and
function. Blood 112: 1570–1579.2. Mizoguchi, A., and A. K. Bhan. 2006. A case for regulatory B cells. J. Immunol.
176: 705–710.3. Serra, P., and P. Santamaria. 2006. To ‘B’ regulated: B cells as members of the
regulatory workforce. Trends Immunol. 27: 7–10.4. Bouaziz, J.-D., K. Yanaba, and T. F. Tedder. 2008. Regulatory B cells as inhib-
itors of immune responses and inflammation. Immunol. Rev. 224: 201–214.5. Mauri, C., and M. R. Ehrenstein. 2008. The ‘short’ history of regulatory B cells.
Trends Immunol. 29: 34–40.6. Lund, F. E. 2008. Cytokine-producing B lymphocytes: key regulators of immu-
nity. Curr. Opin. Immunol. 20: 1–7.7. Fillatreau, S., D. Gray, and S. M. Anderton. 2008. Not always the bad guys: B
cells as regulators of autoimmune pathology. Nat. Rev. Immunol. 8: 391–397.8. Rieger, A., and A. Bar-Or. 2008. B-cell-derived interleukin-10 in autoimmune
disease: regulating the regulators. Nat. Rev. Immunol. 8: 486–487.9. Neta, R., and S. B. Salvin. 1974. Specific suppression of delayed hypersensitivity:
the possible presence of a suppressor B cell in the regulation of delayed hyper-sensitivity. J. Immunol. 113: 1716–1725.
10. Katz, S. I., D. Parker, and J. L. Turk. 1974. B-cell suppression of delayed hy-persensitivity reactions. Nature 251: 550–551.
11. Wolf, S. D., B. N. Dittel, F. Hardardottir, and C. A. Janeway, Jr. 1996. Experi-mental autoimmune encephalomyelitis induction in genetically B cell-deficientmice. J. Exp. Med. 184: 2271–2278.
12. Mizoguchi, A., E. Mizoguchi, R. N. Smith, F. I. Preffer, and A. K. Bhan. 1997.Suppressive role of B cells in chronic colitis of T cell receptor � mutant mice.J. Exp. Med. 186: 1749–1756.
13. Fillatreau, S., C. H. Sweenie, M. J. McGeachy, D. Gray, and S. M. Anderton.2002. B cells regulate autoimmunity by provision of IL-10. Nat. Immunol. 3:944–950.
14. Mizoguchi, A., E. Mizoguchi, H. Takedatsu, R. S. Blumberg, and A. K. Bhan.2002. Chronic intestinal inflammatory condition generates IL-10-producing reg-ulatory B cell subset characterized by CD1d up-regulation. Immunity 16:219–230.
15. Mauri, C., D. Gray, N. Mushtaq, and M. Londei. 2003. Prevention of arthritis byinterleukin 10-producing B cells. J. Exp. Med. 197: 489–501.
16. Watanabe, R., M. Fujimoto, N. Ishiura, Y. Kuwano, H. Nakashima, N. Yazawa,H. Okochi, S. Sato, T. F. Tedder, and K. Tamaki. 2007. CD19 expression in B
cells is important for suppression of contact hypersensitivity. Am. J. Pathol. 171:560–570.
17. Evans, J. G., K. A. Chavez-Rueda, A. Eddaoudi, A. Meyer-Bahlburg,D. J. Rawlings, M. R. Ehrenstein, and C. Mauri. 2007. Novel suppressive func-tion of transitional 2 B cells in experimental arthritis. J. Immunol. 178:7868–7878.
18. Yanaba, K., J.-D. Bouaziz, K. M. Haas, J. C. Poe, M. Fujimoto, and T. F. Tedder.2008. A regulatory B cell subset with a unique CD1dhiCD5� phenotype controlsT cell-dependent inflammatory responses. Immunity 28: 639–650.
19. Matsushita, T., K. Yanaba, J.-D. Bouaziz, M. Fujimoto, and T. F. Tedder. 2008.Regulatory B cells inhibit EAE initiation in mice while other B cells promotedisease progression. J. Clin. Invest. 118: 3420–3430.
20. Asadullah, K., W. Sterry, and H. D. Volk. 2003. Interleukin-10 therapy: reviewof a new approach. Pharmacol. Rev. 55: 241–269.
21. Spencer, N. F., and R. A. Daynes. 1997. IL-12 directly stimulates expression ofIL-10 by CD5� B cells and IL-6 by both CD5� and CD5� B cells: possibleinvolvement in age-associated cytokine dysregulation. Int. Immunol. 9: 745–754.
22. Brummel, R., and P. Lenert. 2005. Activation of marginal zone B cells from lupusmice with type A(D) CpG-oligodeoxynucleotides. J. Immunol. 174: 2429–2434.
23. Harris, D. P., L. Haynes, P. C. Sayles, D. K. Duso, S. M. Eaton, N. M. Lepak,L. L. Johnson, S. L. Swain, and F. E. Lund. 2000. Reciprocal regulation ofpolarized cytokine production by effector B and T cells. Nat. Immunol. 1:475–482.
24. Tian, J., D. Zekzer, L. Hanssen, Y. Lu, A. Olcott, and D. L. Kaufman. 2001.Lipopolysaccharide-activated B cells down-regulate Th1 immunity and preventautoimmune diabetes in nonobese diabetic mice. J. Immunol. 167: 1081–1089.
25. Yanaba, K., J. D. Bouaziz, T. Matsushita, C. M. Magro, E. W. St. Clair, andT. F. Tedder. 2008. B-lymphocyte contributions to human autoimmune disease.Immunol. Rev. 223: 284–299.
26. Hussain, S., and T. L. Delovitch. 2007. Intravenous transfusion of BCR-activatedB cells protects NOD mice from type 1 diabetes in an IL-10-dependent manner.J. Immunol. 179: 7225–7232.
27. Yazawa, N., M. Fujimoto, S. Sato, K. Miyake, N. Asano, O. Takeuchi,K. Takeda, S. Akira, T. F. Tedder, and K. Tamaki. 2003. CD19 regulates innateimmunity by the Toll-like receptor RP105 signaling in B lymphocytes. Blood102: 1374–1380.
28. Goodnow, C. C., J. Crosbie, S. Adelstein, T. B. Lavoie, S. J. Smith-Gill,R. A. Brink, H. Pritchard-Briscoe, J. S. Wotherspoon, R. H. Loblay, K. Raphael,et al. 1988. Altered immunoglobulin expression and functional silencing of self-reactive B lymphocytes in transgenic mice. Nature 334: 676–682.
29. Grusby, M. J., H. Auchincloss, Jr., R. Lee, R. S. Johnson, J. P. Spencer,M. Zijlstra, R. Jaenisch, V. E. Papaioannou, and L. H. Glimcher. 1993. Micelacking major histocompatibility complex class I and class II molecules. Proc.Natl. Acad. Sci. USA 90: 3913–3917.
30. Adachi, O., T. Kawai, K. Takeda, M. Matsumoto, H. Tsutsui, M. Sakagami,K. Nakanishi, and S. Akira. 1998. Targeted disruption of the MyD88 gene resultsin loss of IL-1- and IL-18-mediated function. Immunity 9: 143–150.
31. Poe, J. C., Y. Fujimoto, M. Hasegawa, K. M. Haas, A. S. Miller, I. G. Sanford,C. B. Bock, M. Fujimoto, and T. F. Tedder. 2004. CD22 regulates B lymphocytefunction in vivo through both ligand-dependent and ligand-independent mecha-nisms. Nat. Immunol. 5: 1078–1087.
32. Sato, S., N. Ono, D. A. Steeber, D. S. Pisetsky, and T. F. Tedder. 1996. CD19regulates B lymphocyte signaling thresholds critical for the development of B-1lineage cells and autoimmunity. J. Immunol. 157: 4371–4378.
33. Sato, S., D. A. Steeber, P. J. Jansen, and T. F. Tedder. 1997. CD19 expressionlevels regulate B lymphocyte development: human CD19 restores normal func-tion in mice lacking endogenous CD19. J. Immunol. 158: 4662–4669.
34. Haas, K. M., M. Hasegawa, D. A. Steeber, J. C. Poe, M. D. Zabel, C. B. Bock,D. R. Karp, D. E. Briles, J. H. Weis, and T. F. Tedder. 2002. Complementreceptors CD21/35 link innate and protective immunity during Streptococcuspneumoniae infection by regulating IgG3 antibody responses. Immunity 17:713–723.
35. Higuchi, T., Y. Aiba, T. Nomura, J. Matsuda, K. Mochida, M. Suzuki,H. Kikutani, T. Honjo, K. Nishioka, and T. Tsubata. 2002. Cutting edge: ectopicexpression of CD40 ligand on B cells induces lupus-like autoimmune disease.J. Immunol. 168: 9–12.
36. Anderson, M. S., and J. A. Bluestone. 2005. The NOD mouse: a model of im-mune dysregulation. Annu. Rev. Immunol. 23: 447–485.
37. Courtenay, J. S., M. J. Dallman, A. D. Dayan, A. Martin, and B. Mosedale. 1980.Immunization against heterologous type II collagen induces arthritis in mice.Nature 283: 666–668.
38. Dal Canto, M. C., R. W. Melvold, B. S. Kim, and S. D. Miller. 1995. Two modelsof multiple sclerosis: experimental allergic encephalomyelitis (EAE) and Thei-ler’s murine encephalomyelitis virus (TMEV) infection: a pathological and im-munological comparison. Microsc. Res. Tech. 32: 215–229.
39. Theofilopoulos, A. N., ed. 1992. Murine Models of Lupus. Churchill Livingston,Edinburgh.
40. Tedder, T. F. 1998. Response-regulators of B lymphocyte signaling thresholdsprovide a context for antigen receptor signal transduction. Semin. Immunol. 10:259–265.
41. Brutkiewicz, R. R., J. R. Bennink, J. W. Yewdell, and A. Bendelac. 1995. TAP-independent, �2-microglobulin-dependent surface expression of functionalmouse CD1.1. J. Exp. Med. 182: 1913–1919.
42. Hokazono, Y., T. Adachi, M. Wabl, N. Tada, T. Amagasa, and T. Tsubata. 2003.Inhibitory coreceptors activated by antigens but not by anti-Ig heavy chain an-tibodies install requirement of costimulation through CD40 for survival and pro-liferation of B cells. J. Immunol. 171: 1835–1843.
7471The Journal of Immunology
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from

43. Lampropoulou, V., K. Hoehlig, T. Roch, P. Neves, E. Calderon Gomez,C. H. Sweenie, Y. Hao, A. A. Freitas, U. Steinhoff, S. M. Anderton, andS. Fillatreau. 2008. TLR-activated B cells suppress T cell-mediated autoimmu-nity. J. Immunol. 180: 4763–4773.
44. Fujimoto, M., A. P. Bradney, J. C. Poe, D. A. Steeber, and T. F. Tedder. 1999.Modulation of B lymphocyte antigen receptor signal transduction by a CD19/CD22 regulatory loop. Immunity 11: 191–200.
45. Fujimoto, M., Y. Fujimoto, J. C. Poe, P. J. Jansen, C. A. Lowell, A. L. DeFranco,and T. F. Tedder. 2000. CD19 regulates Src-family protein tyrosine kinase acti-vation in B lymphocytes through processive amplification. Immunity 13: 47–57.
46. Poe, J. C., M. Hasegawa, and T. F. Tedder. 2001. CD19, CD21 and CD22:multifaceted response regulators of B lymphocyte signal transduction. Int. Rev.Immunol. 20: 739–762.
47. Tedder, T. F., J. C. Poe, M. Fujimoto, K. M. Haas, and S. Sato. 2005. TheCD19-CD21 signal transduction complex of B lymphocytes regulates the balancebetween health and autoimmune disease: systemic sclerosis as a model system.Curr. Dir. Autoimmun. 8: 55–90.
48. Poe, J. C., K. M. Haas, J. Uchida, Y. Lee, M. Fujimoto, and T. F. Tedder. 2004.Severely-impaired B lymphocyte proliferation, survival and induction of the c-Myc:Cullin 1 ubiquitin ligase pathway resulting from CD22 deficiency on theC57BL/6 genetic background. J. Immunol. 172: 2100–2110.
49. Van Kooten, C., and J. Banchereau. 2000. CD40-CD40 ligand. J. Leukocyte Biol.67: 2–17.
50. Hayakawa, K., R. R. Hardy, and L. A. Herzenberg. 1986. Peritoneal Ly-1 B cells:genetic control, autoantibody production, increased light chain expression. Eur.J. Immunol. 16: 450–456.
51. Mauri, C., L. T. Mars, and M. Londei. 2000. Therapeutic activity of agonisticmonoclonal antibodies against CD40 in a chronic autoimmune inflammatory pro-cess. Nat. Med. 6: 673–679.
52. Buenafe, A. C., and D. N. Bourdette. 2007. Lipopolysaccharide pretreatmentmodulates the disease course in experimental autoimmune encephalomyelitis.J. Neuroimmunol. 182: 32–40.
53. Makowska, A., N. N. Faizunnessa, P. Anderson, T. Midtvedt, and S. Cardell.1999. CD1high B cells: a population of mixed origin. Eur. J. Immunol. 29:3285–3294.
54. Haas, K. M., J. C. Poe, D. A. Steeber, and T. F. Tedder. 2005. B-1a and B-1b cellsexhibit distinct developmental requirements and have unique functional roles ininnate and adaptive immunity to S. pneumoniae. Immunity 23: 7–18.
55. Lalor, P. A., and G. Morahan. 1990. The peritoneal Ly-1 (CD5) B cell repertoireis unique among murine B cell repertoires. Eur. J. Immunol. 20: 485–492.
56. Martin, F., A. M. Oliver, and J. F. Kearney. 2001. Marginal zone and B1 B cellsunite in the early response against T-independent blood-borne particulate anti-gens. Immunity 14: 617–629.
57. Cong, T., E. Rabin, and H. Wortis. 1991. Treatment of CD5� B cells with anti-Igbut not LPS, induces surface CD5: two B cell activation pathways. Int. Immunol.3: 467.
7472 REGULATORY B10 CELL DEVELOPMENT
by guest on May 23, 2012
http://jimm
unol.org/D
ownloaded from