the cosmetic dye quinoline yellow causes dna dmage in vitro
TRANSCRIPT

7/26/2019 The Cosmetic Dye Quinoline Yellow Causes DNA Dmage in VITRO
http://slidepdf.com/reader/full/the-cosmetic-dye-quinoline-yellow-causes-dna-dmage-in-vitro 1/8
Mutation Research 777 (2015) 54–61
Contents lists available at ScienceDirect
Mutation Research/Genetic Toxicology andEnvironmental Mutagenesis
journal homepage: www.elsevier .com/ locate /gentox
Commun i ty address : www.elsevier .com/ locate /mutres
The cosmetic dye quinoline yellow causes DNA damage in vitro
Farah Maria Drumond Chequera,b,∗, Vinícius de Paula Venâncioa,Maíra Rocha de Souza Prado a, Luiz Raimundo Campos da Silva e Cunha Junior c,Thiago Mescoloto Lizier d, Maria Valnice Boldrin Zanoni d, Rommel Rodríguez Burbanoc,Maria Lourdes Pires Bianchia, Lusânia Maria Greggi Antunes a
a Departamento de Análises Clínicas, Toxicológicas e Bromatológicas, Faculdadede Ciências Farmacêuticas de Ribeirão Preto,Universidadede SãoPaulo,
USP, Ribeirão Preto, SP 14040-903, Brazilb Departamento de Análises Clínicas e Toxicológicas, Faculdade Federal de Minas Gerais, UFMG, Belo Horizonte, MG 31270-901, Brazilc Laboratório de CitogenéticaHumana, Instituto de Ciências Biológicas, Universidade Federal do Pará, Belém, PA, Brazild Instituto de Química. Departamento de Química Analítica, Universidade Estadual Paulista – UNESP, Quitandinha 14800-900,Araraquara/SP, Brazil
a r t i c l e i n f o
Article history:
Received 29 June 2014
Received in revised form 6 November 2014
Accepted 11 November 2014
Available online18 November 2014
Keywords:
Genotoxicity
Cosmetic dye
Micronucleus
Comet assay
Oxidation
a b s t r a c t
Quinoline yellow (QY) is a chinophthalon derivative used in cosmetic compositions for application to
the skin, lips, and/or body surface. However, regulatory data about the genotoxicity and/or mutageni-
city of this compound are still controversial. Therefore, this work evaluated the genotoxicity of QY using
the comet assay and the cytokinesis-block micronucleus cytome assay (CBMN-Cyt) in the metabolically
competent cell line HepG2, which closely mimics phase I metabolism. This research also identified the
products formed after electrochemical oxidation of the QY dye, which simulates hepatic biotransforma-
tion. The primary products generated after the oxidation process were analyzed by High Performance
Liquid Chromatography coupled with a Diode Array Detector (HPLC/DAD), which detected the production
of 4,4-diaminodiphenylmethane, 2-methoxy-5-methylaniline and 4,4-oxydianiline. The results demon-
strated that low (from 0.5to 20g mL −1) QY concentrations were genotoxic in HepG2 cells on both assays
and those harmful compounds were detected after the oxidation process. Our findings suggest that this
colorant could cause harmful effects to humans if it is metabolized or absorbed through the skin.© 2014 Elsevier B.V. All rights reserved.
1. Introduction
Synthetic dyes are used extensively in many industries,
including the cosmetics, textile, pharmaceutical, food, plastics,
photography and paper industries [1–4]. It is estimated that over
10,000 different dyes and pigments are used industrially and that
over 7×105 tons of synthetic dyes are produced annually world-
wide [4–6]. However, there is insufficient information about their
potential health risks for humans and the environment [7,8]. The
available toxicological data about cosmetics dyes have shown
effects that range from contact allergies to different types of
genetic damages, including genotoxicity, mutagenicity and early
age leukemia [9–13].
∗ Corresponding author at: Departamento de Análises Clínicas, Toxicológicas e
Bromatológicas, Faculdade de Ciências Farmacêuticas de Ribeirão Preto, Universi-
dade deSão Paulo,USP,Ribeirão Preto, SP14040-903, Brazil.Tel.: +5516 36024186;
fax: +55 16 3602 4725.
E-mail address: [email protected] (F.M.D. Chequer).
The dye quinoline yellow (QY) is a chinophthalon derivative
used in cosmetic compositions for application to the skin, lips,
and/or body surface [14]. This dye (also known as D&C Yellow no.
11) was found to induce allergic contact dermatitis; in a human
maximization test,15 of 20 subjects became sensitized[15,16]. The
regulatory data regarding QY genotoxicity and/or mutagenicity are
still controversial [14,17]. Therefore, we studied the dye QY in this
research. Theaim of this investigation was to evaluate thegenotox-
icity of QY using the alkaline comet assay andthe cytokinesis-block
micronucleus cytome assay (CBMN-Cyt) in the metabolically com-
petent cell line HepG2, which closely mimics phase I metabolism.
Micronuclei (MN) were also analyzedusingthe fluorescence in situ
hybridization (FISH) technique for further hazard characterization.
In addition, it is known that aromatic amines can be produced
during oxidative and/or reductive processes [3,18], and the forma-
tion of these aromatic amine byproducts could be important for
understanding the chemical transformation of dyes. Therefore, the
present work also aimed to determine if 16 aromatic amines used
as standard models of amines classified by IARC [19] could be pro-
duced during theoxidative and/or reductiveconditions.In addition,
http://dx.doi.org/10.1016/j.mrgentox.2014.11.003
1383-5718/© 2014 Elsevier B.V. All rightsreserved.

7/26/2019 The Cosmetic Dye Quinoline Yellow Causes DNA Dmage in VITRO
http://slidepdf.com/reader/full/the-cosmetic-dye-quinoline-yellow-causes-dna-dmage-in-vitro 2/8

7/26/2019 The Cosmetic Dye Quinoline Yellow Causes DNA Dmage in VITRO
http://slidepdf.com/reader/full/the-cosmetic-dye-quinoline-yellow-causes-dna-dmage-in-vitro 3/8
56 F.M.D. Chequer et al. / Mutation Research 777 (2015) 54–61
Fig. 2. The effects of 0.5, 1.0, 2.0, 5.0, 10.0, 15.0 or 20.0g mL −1 of quinoline yellow (QY) for 4h on the tail intensity (A) and tail moment (B) of HepG2 cells evaluated by
the comet assay. The values shown represent the mean±SD, and the data are based on three independent experiments. Vehicle control, 1.0% dimethylsulfoxide; positive
control, 0.3g mL −1
of doxorubicin. *: Significantly different from vehicle control group.
Table 1
Assessment of themutagenic effects of quinolineyellow (QY) on HepG2 cells using thecytokinesis-block micronucleus cytomeassay (CBMN-Cyt).
Treatment
(g mL −1)
CBMN-Cyt NDI FISH
Total no. in 1000 BN cells C + MN(%)/C−MN(%)
MNi NPBs NBUDs
Vehicle control 19 ± 3 2 ± 2 5 ± 3 1.6 ± 0.1 57.1/42.9
Positive control 99 ± 14* 16 ± 5 10 ± 7 1.6 ± 0.1 60.5/39.5
0.5 QY 53 ± 1* 3 ± 4 3 ± 3 1.5 ± 0.1 n/a
1.0 QY 46 ± 8* 2 ± 3 3 ± 2 1.5 ± 0.1 n/a
2.0 QY 50 ± 15* 2 ± 3 4 ± 3 1.5 ± 0.1 n/a
5.0 QY 63 ± 13* 1 ± 1 6 ± 4 1.6 ± 0.1 n/a
10.0 QY 65 ± 14* 2 ± 3 6 ± 2 1.5 ± 0.1 n/a
15.0 QY 69 ± 7* 2 ± 2 7 ± 2 1.5 ± 0.1 57.9/42.120.0 QY 72 ± 4* 3 ± 2 4 ± 1 1.4 ± 0.1 57.2/42.8
Valuesshown arethemean±SD; BN, binucleatedcell; MNi,micronuclei; NPBs, nucleoplasmic bridges; NBUDs, nuclear buds;NDI, nuclear division index; C + MN, centromere
positive (i.e., MN containing one or more whole chromosome signals); C−MN, centromere negative (i.e., MN containing acentric chromosome fragment signals). The data
shown are based on three independent experiments. Vehicle control, 1.0% dimethylsulfoxide; positive control, 0.03g mL −1 doxorubicin. *: Significantly different from the
control group ( p< 0.05).
The conditions used for monitoring the degradation of the dye consisted of a
MeOH/water 80:20 mobile phase, 1.0mL min−1 flow rate, column temperature of
45 ◦C, Phenomenex Luna C18 column (250×4, 6 m m, 5m) and analysis = 450nm.
The analysis time was 10min, and all of theanalyseswere conductedin triplicate.
All of these methodologies were conducted based on chromatographic param-
e ter s s uch as r et ent ion t ime (t R ), retention constant factor (k), selectivity (˛),
resolution between peaks (Rs) and theoretical plate number (N ). Standard curves
anda quantitativeanalysisof thetarget amineswereobtainedusing a linearregres-
sion of thepeak area vs.concentration.Further comparisons were performedusing
the standardadditionmethodin whichaliquotsof theworkingstandard dissolvedin
methanol were spiked into thesamples.The procedurewas conductedin triplicatefor each sample.
2.8. Statistical analysis
Allof thedatashownare expressed asthe mean value±SD of three independent
experiments.The results were analyzed using one-way ANOVA with post hocDun-
nett’s tests (ata significancelevel p< 0.05)in GraphPadPrism 5 (GraphPadSoftware,
USA).
3. Results
All of the QY concentrations used in this study resulted in at
least 80% cell viability prior to cell harvesting, as determined by
the trypan-blue exclusionmethod,and concentrations greater than
20g mL −1
were excluded due to the low cell viability.
The results of the comet assay are shown in Fig. 2. The sensi-
tivity of this in vitro assay was demonstrated by the response to
0.3g mL −1 DXR, which induced a statistically significant increase
in tail moment and tail intensity compared with the vehicle con-
trol group (i.e., cells treated with1.0% DMSO). Moreover, compared
with the vehicle control, QY was genotoxic to HepG2 cells at con-
centrations ranging from 2.0 to 20g mL −1.
In the CBMN-Cyt assay, the increased MNi frequencies (Table 1)
indicated that this dye promoted genotoxic effects at each of theconcentrations tested (0.5–20g mL −1). The frequencies of NPB
and NBUD and the NDI calculation (also shown in Table 1) were
not significantly different between the experimental and control
groups. TheFISH assay also showedthatalthough the MNifrequen-
cies were higher in the treated groups, there was no difference in
the C+ MN/C−MN ratio between the treated groups and the nega-
tive control group ( p< 0.05). No difference between aneugenic and
clastogenic effects was observed across the treatments. DXR was
used as a positive control, and 0.03g mL −1 DXR increased the MN
frequency compared with the vehicle control group.
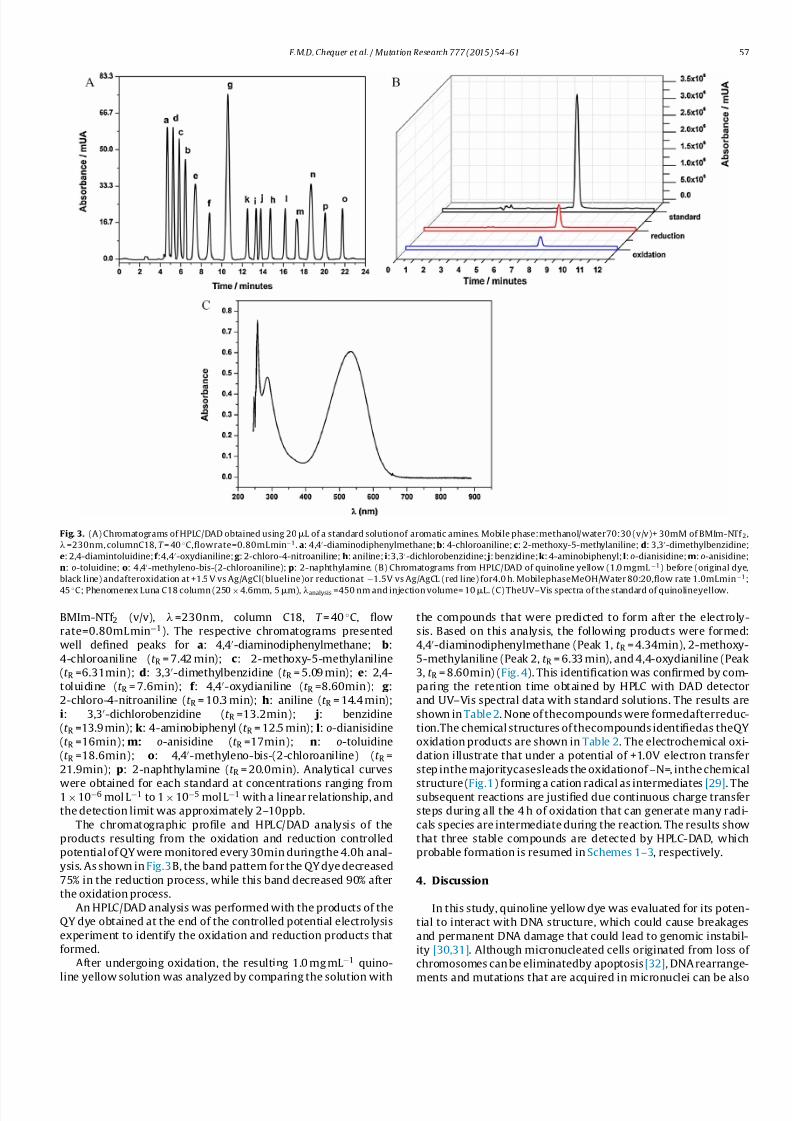
Fig. 3A shows representative chromatograms of the HPLC-
DAD data obtained for a 20L solution containing 50ppm
of the standard aromatic amine of interest (mobile phase:
methanol/water 70:30 (v/v) containing 30mM of liquid ionic
7/26/2019 The Cosmetic Dye Quinoline Yellow Causes DNA Dmage in VITRO
http://slidepdf.com/reader/full/the-cosmetic-dye-quinoline-yellow-causes-dna-dmage-in-vitro 4/8
F.M.D. Chequer et al. / Mutation Research 777 (2015) 54–61 57
Fig. 3. (A) Chromatograms of HPLC/DAD obtained using 20L of a standard solutionof aromatic amines. Mobile phase:methanol/water70:30 (v/v)+ 30mM of BMIm-NTf 2,
=230nm, columnC18, T = 40 ◦C,flowrate=0.80mL min−1. a: 4,4-diaminodiphenylmethane;b: 4-chloroaniline;c: 2-methoxy-5-methylaniline;d: 3,3-dimethylbenzidine;
e: 2,4-diamintoluidine;f :4,4-oxydianiline;g : 2-chloro-4-nitroaniline;h: aniline;i:3,3-dichlorobenzidine; j: benzidine;k: 4-aminobiphenyl;l:o-dianisidine;m: o-anisidine;
n: o-toluidine; o: 4,4-methyleno-bis-(2-chloroaniline);p: 2-naphthylamine. (B) Chromatograms from HPLC/DAD of quinoline yellow (1.0 mgmL −1) before (original dye,
black line) andafteroxidation at +1.5 V vs Ag/AgCl(blueline)or reductionat −1.5V vs Ag/AgCL (red line) for4.0 h. MobilephaseMeOH/Water 80:20,flow rate 1.0mLmin−1;
45 ◦C; Phenomenex Luna C18 column (250×4.6mm, 5m), analysis =450 nm and injection volume= 10L. (C) TheUV–Vis spectra of the standard of quinolineyellow.
BMIm-NTf 2 (v/v), =230nm, column C18, T = 40 ◦C, flow
rate=0.80mL min−1). The respective chromatograms presented
well defined peaks for a: 4,4-diaminodiphenylmethane; b:
4-chloroaniline (t R = 7.42 min); c: 2-methoxy-5-methylaniline
(t R =6.31min); d: 3,3-dimethylbenzidine (t R = 5.09 min); e: 2,4-
toluidine (t R = 7.6min); f : 4,4-oxydianiline (t R =8.60min); g :
2-chloro-4-nitroaniline (t R = 10.3 min); h: aniline (t R = 14.4 min);
i: 3,3-dichlorobenzidine (t R =13.2min); j: benzidine
(t R =13.9 min); k: 4-aminobiphenyl (t R = 12.5 min); l: o-dianisidine
(t R =16min); m: o-anisidine (t R =17min); n: o-toluidine
(t R =18.6min); o: 4,4-methyleno-bis-(2-chloroaniline) (t R =
21.9min); p: 2-naphthylamine (t R = 20.0min). Analytical curves
were obtained for each standard at concentrations ranging from1×10−6 mol L −1 to 1×10−5 mol L −1 with a linear relationship, and
the detection limit was approximately 2–10ppb.
The chromatographic profile and HPLC/DAD analysis of the
products resulting from the oxidation and reduction controlled
potential of QY were monitored every 30min duringthe 4.0h anal-
ysis. As shown in Fig.3B, the band pattern for the QY dye decreased
75% in the reduction process, while this band decreased 90% after
the oxidation process.
An HPLC/DAD analysis was performed with the products of the
QY dye obtained at the end of the controlled potential electrolysis
experiment to identify the oxidation and reduction products that
formed.
After undergoing oxidation, the resulting 1.0 mg mL −1 quino-
line yellow solution was analyzed by comparing the solution with
the compounds that were predicted to form after the electroly-
sis. Based on this analysis, the following products were formed:
4,4-diaminodiphenylmethane (Peak 1, t R = 4.34min), 2-methoxy-
5-methylaniline (Peak 2, t R = 6.33 min), and 4,4-oxydianiline (Peak
3, t R = 8.60min) (Fig. 4). This identification was confirmed by com-
paring the retention time obtained by HPLC with DAD detector
and UV–Vis spectral data with standard solutions. The results are
shown in Table 2. None of thecompounds were formedafterreduc-
tion.The chemical structures of thecompounds identifiedas theQY
oxidation products are shown in Table 2. The electrochemical oxi-
dation illustrate that under a potential of +1.0V electron transfer
step inthe majoritycasesleads the oxidationof –N=, inthe chemical
structure (Fig.1) forming a cation radical as intermediates [29]. Thesubsequent reactions are justified due continuous charge transfer
steps during all the 4 h of oxidation that can generate many radi-
cals species are intermediate during the reaction. The results show
that three stable compounds are detected by HPLC-DAD, which
probable formation is resumed in Schemes 1–3, respectively.
4. Discussion
In this study, quinoline yellow dye was evaluated for its poten-
tial to interact with DNA structure, which could cause breakages
and permanent DNA damage that could lead to genomic instabil-
ity [30,31]. Although micronucleated cells originated from loss of
chromosomes can be eliminatedby apoptosis [32], DNA rearrange-
ments and mutations that are acquired in micronuclei can be also

7/26/2019 The Cosmetic Dye Quinoline Yellow Causes DNA Dmage in VITRO
http://slidepdf.com/reader/full/the-cosmetic-dye-quinoline-yellow-causes-dna-dmage-in-vitro 5/8
58 F.M.D. Chequer et al. / Mutation Research 777 (2015) 54–61
Fig. 4. (A) Chromatogram from HPLC/DAD of quinolineyellow (1.0 mg mL −1) before and after oxidationand reduction at controlled potential at +1.0 and−1.0V vs Ag/AgCl,
respectively. The black line corresponds to the standard (original QY dye); red line: after 4 h of oxidation; and blue line: 4 h after the reduction process. Chromatographic
conditions: mobile phase MeOH/water +30 mM of BMIm [NTf 2] 70:30, flow rate 0.8mL min−1; column temperature= 45◦ C; Phenomenex Luna C18 column (250×4, 6mm,
5m), analysis =230 nm and injectionvolume= 10L. (B)The UV–Vis spectra of theoxidation products of quinoline yellow.
incorporated intothe genomes of developing cancer cells, andthese
micronuclei can persist for many generations [33].The alkaline version of comet assay was used once this method-
ology can detect DNA single-strand breaks, alkali-labile sites,
DNA–DNA and DNA–protein cross-linking and single strand breaks
associated with incomplete excision repair sites [21]. CBMN-Cyt
was chosen as an important complementary technique because
in addition to the DNA damage evaluation, this methodology can
detect dicentric chromosomes and gene amplification [24], provid-
ing a broad chrosomosome instability screening. In metabolically
competent HepG2 cells, QY was genotoxic by both the comet assay
and CBMN-Cyt. FISH assays revealed that QY induces DNA damage
through both aneugenic and clastogenic processes.
HepG2 cells are often used in toxicological investigations and
gene expression studies because they express metabolic enzymes
that can oxidize or reduce xenobiotics, closely mimicking thein vivo activity of hepatocytes [34–37]. These cells have retained
the inducibility and activities of several phase I and phase II xeno-
biotic metabolizing enzymes, and have been shown to be suitable
for the detection of different classes of indirect-acting genotoxic
agents [38]. In addition, this cell line is considered useful for avoid-
ing false negative results in the detection of genotoxic carcinogens
[35,39,40], such as synthetic dyes and other chemical compounds
that can be oxidized or reduced to becomeeither more or less toxic
[18,41]. HepG2cells also express wild-typetumor suppressorTP53,
making them an appropriate model for studying P53-regulated
responses to DNA damage at the level of gene transcription and
translation [42,43].
Here, HepG2 cells were treated with concentrations of QY
that were determined based on its solubility and low ADI(0–0.5 mg kg−1) [44]. The concentrations tested were not capable
of inhibiting the cell cycle (no significant differences in the NDI
between the experimental and control groups were observed) and
resulted in cell viability levels of greater than 80% based on the
trypan-blue exclusion method, ensuring the consistency of our
results. In addition, the percentage of binucleated cells observed
using CBMN-Cyt was greater than 35% in all of the treatments.
In addition to QY, other cosmetic dyes have also been found to
be potentially genotoxic in mammalian cells. Mpountoukas et al.
[12] evaluated the genotoxic, cytotoxic and cytostatic potential of
the synthetic dyes amaranth, erythrosine and tartrazine in human
peripheral blood cells in vitro. These dyes are used in food and cos-
metic products,and theresults of this research indicatedthat these
colorants werepotentially toxicto human lymphocytes in vitro andcould possibly bind directly to DNA [12]. However, research in syn-
thetic dyes are often controversial andsome results of in vitro tests
do not show the same effects in vivo assays. For instance, accord-
ing to Poul et al. [45], acute oral exposure to food dye additives
amaranth, tartrazine and sunset yellow as well as to the hepa-
tocarcinogen azo dye dimethylaminoazobenzene (DAB) did not
induce genotoxiceffect in the gut using micronucleus assayin mice.
However, the DNA damage induced by amaranth and tartrazine,
previously noted in the in vivo comet assay in mouse colon [46],
was not corroborate in the gut micronucleus assay in mice [45].
Additionally, it is known that several biotransformation reac-
tions may occur after the absorption of a xenobiotic, and the
Table 2
Oxidation products obtained from QY, as determined by HPLC/DAD.
Compound HPLC/DAD
Structure CAS number t R (min)
Peak 1: 4,4-diaminodiphenylmetane 101-77-9 4.34
Peak 2: 2-methoxy-5-methylaniline 120-71-8 6.33
Peak 3: 4,4-oxydianiline 101-80-4 8.60
7/26/2019 The Cosmetic Dye Quinoline Yellow Causes DNA Dmage in VITRO
http://slidepdf.com/reader/full/the-cosmetic-dye-quinoline-yellow-causes-dna-dmage-in-vitro 6/8
F.M.D. Chequer et al. / Mutation Research 777 (2015) 54–61 59
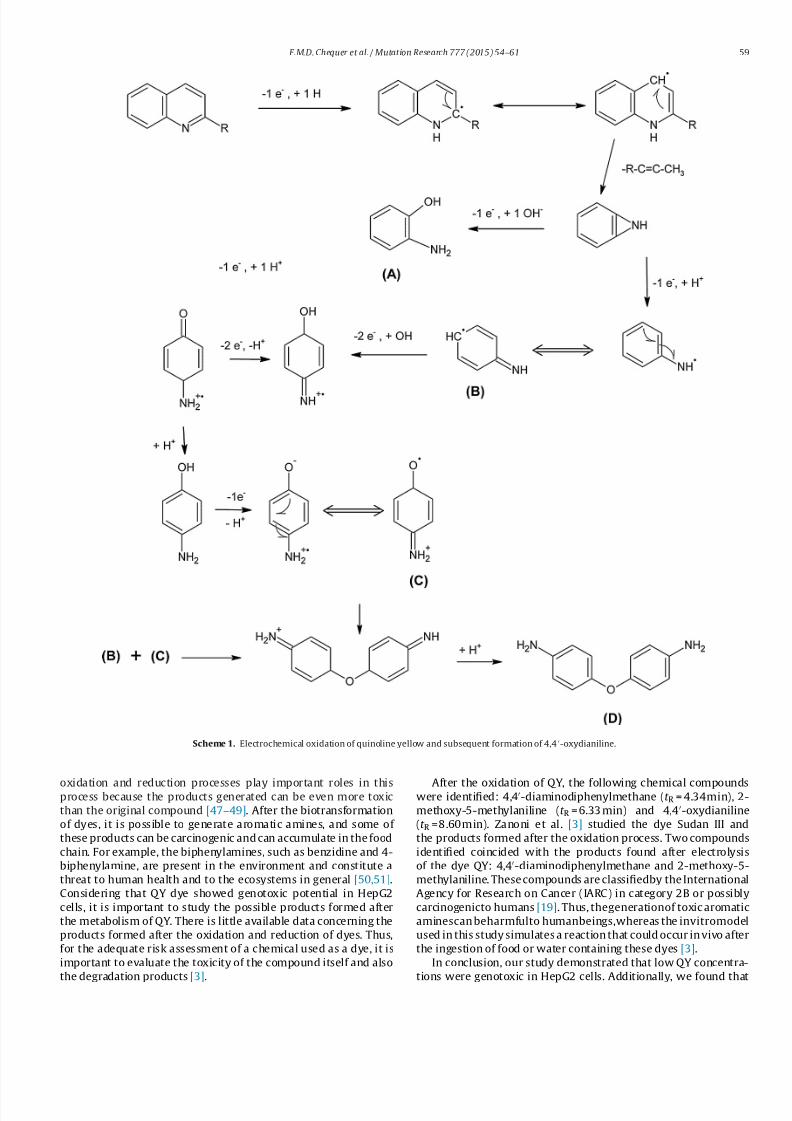
Scheme 1. Electrochemical oxidation of quinoline yellow and subsequent formation of 4,4-oxydianiline.
oxidation and reduction processes play important roles in thisprocess because the products generated can be even more toxic
than the original compound [47–49]. After the biotransformation
of dyes, it is possible to generate aromatic amines, and some of
these products can be carcinogenic and can accumulate in the food
chain. For example, the biphenylamines, such as benzidine and 4-
biphenylamine, are present in the environment and constitute a
threat to human health and to the ecosystems in general [50,51].
Considering that QY dye showed genotoxic potential in HepG2
cells, it is important to study the possible products formed after
the metabolism of QY. There is little available data concerning the
products formed after the oxidation and reduction of dyes. Thus,
for the adequate risk assessment of a chemical used as a dye, it is
important to evaluate the toxicity of the compound itself and also
the degradation products [3].
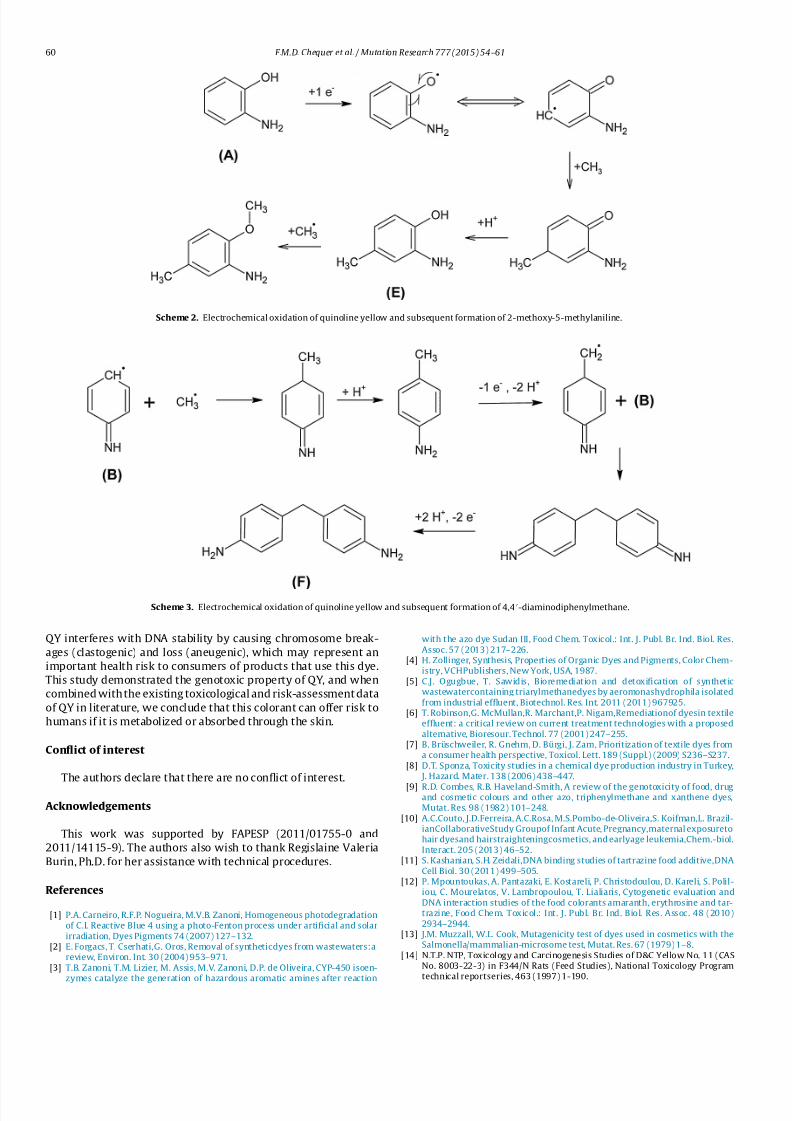
After the oxidation of QY, the following chemical compoundswere identified: 4,4-diaminodiphenylmethane (t R = 4.34min), 2-
methoxy-5-methylaniline (t R = 6.33 min) and 4,4-oxydianiline
(t R =8.60min). Zanoni et al. [3] studied the dye Sudan III and
the products formed after the oxidation process. Two compounds
identified coincided with the products found after electrolysis
of the dye QY: 4,4-diaminodiphenylmethane and 2-methoxy-5-
methylaniline. These compounds are classifiedby the International
Agency for Research on Cancer (IARC) in category 2B or possibly
carcinogenicto humans [19]. Thus, thegenerationof toxic aromatic
aminescan beharmfulto humanbeings,whereas the invitromodel
used in this study simulates a reaction that could occur in vivo after
the ingestion of food or water containing these dyes [3].
In conclusion, our study demonstrated that low QY concentra-
tions were genotoxic in HepG2 cells. Additionally, we found that
7/26/2019 The Cosmetic Dye Quinoline Yellow Causes DNA Dmage in VITRO
http://slidepdf.com/reader/full/the-cosmetic-dye-quinoline-yellow-causes-dna-dmage-in-vitro 7/8
60 F.M.D. Chequer et al. / Mutation Research 777 (2015) 54–61
Scheme 2. Electrochemical oxidation of quinoline yellow and subsequent formation of 2-methoxy-5-methylaniline.
Scheme 3. Electrochemical oxidation of quinoline yellow and subsequent formation of 4,4-diaminodiphenylmethane.
QY interferes with DNA stability by causing chromosome break-
ages (clastogenic) and loss (aneugenic), which may represent an
important health risk to consumers of products that use this dye.
This study demonstrated the genotoxic property of QY, and when
combined with the existing toxicological and risk-assessment data
of QY in literature, we conclude that this colorant can offer risk to
humans if it is metabolized or absorbed through the skin.
Conflict of interest
The authors declare that there are no conflict of interest.
Acknowledgements
This work was supported by FAPESP (2011/01755-0 and
2011/14115-9). The authors also wish to thank Regislaine Valeria
Burin, Ph.D. for her assistance with technical procedures.
References
[1] P.A. Carneiro, R.F.P. Nogueira, M.V.B. Zanoni, Homogeneous photodegradationof C.I. Reactive Blue 4 using a photo-Fenton process under artificial and solarirradiation, Dyes Pigments 74 (2007) 127–132.
[2] E. Forgacs, T. Cserhati,G. Oros, Removal of syntheticdyes from wastewaters:areview, Environ. Int. 30 (2004) 953–971.
[3] T.B. Zanoni, T.M. Lizier, M. Assis, M.V. Zanoni, D.P. de Oliveira, CYP-450 isoen-
zymes catalyze the generation of hazardous aromatic amines after reaction
with the azo dye Sudan III, Food Chem. Toxicol.: Int. J. Publ. Br. Ind. Biol. Res.Assoc. 57 (2013) 217–226.
[4] H. Zollinger, Synthesis, Properties of Organic Dyes and Pigments, Color Chem-istry, VCHPublishers, New York, USA, 1987.
[5] C.J. Ogugbue, T. Sawidis, Bioremediation and detoxification of syntheticwastewatercontaining triarylmethanedyes by aeromonashydrophila isolatedfrom industrial effluent, Biotechnol. Res. Int. 2011 (2011) 967925.
[6] T. Robinson,G. McMullan,R. Marchant,P. Nigam,Remediationof dyesin textileeffluent: a critical review on current treatment technologies with a proposedalternative, Bioresour. Technol. 77 (2001) 247–255.
[7] B. Brüschweiler, R. Gnehm, D. Bürgi, J. Zarn, Prioritization of textile dyes froma consumer health perspective, Toxicol. Lett. 189 (Suppl.) (2009) S236–S237.
[8] D.T. Sponza, Toxicity studies in a chemical dye production industry in Turkey, J. Hazard. Mater. 138 (2006) 438–447.
[9] R.D. Combes, R.B. Haveland-Smith, A review of the genotoxicity of food, drugand cosmetic colours and other azo, triphenylmethane and xanthene dyes,Mutat. Res. 98 (1982) 101–248.
[10] A.C.Couto, J.D.Ferreira, A.C.Rosa, M.S.Pombo-de-Oliveira,S. Koifman,L. Brazil-ianCollaborativeStudy Groupof Infant Acute, Pregnancy,maternal exposuretohair dyesand hairstraighteningcosmetics, and earlyage leukemia,Chem.-biol.Interact. 205 (2013) 46–52.
[11] S. Kashanian, S.H. Zeidali,DNA binding studies of tartrazine food additive,DNACell Biol. 30 (2011) 499–505.
[12] P. Mpountoukas, A. Pantazaki, E. Kostareli, P. Christodoulou, D. Kareli, S. Polil-iou, C. Mourelatos, V. Lambropoulou, T. Lialiaris, Cytogenetic evaluation andDNA interaction studies of the food colorants amaranth, erythrosine and tar-trazine, Food Chem. Toxicol.: Int. J . Publ. Br. Ind. Biol. Res. Assoc. 48 (2010)2934–2944.
[13] J.M. Muzzall, W.L. Cook, Mutagenicity test of dyes used in cosmetics with theSalmonella/mammalian-microsome test, Mutat. Res. 67 (1979) 1–8.
[14] N.T.P. NTP, Toxicology and Carcinogenesis Studies of D&C Yellow No. 11 (CASNo. 8003-22-3) in F344/N Rats (Feed Studies), National Toxicology Programtechnical reportseries, 463 (1997) 1-190.

7/26/2019 The Cosmetic Dye Quinoline Yellow Causes DNA Dmage in VITRO
http://slidepdf.com/reader/full/the-cosmetic-dye-quinoline-yellow-causes-dna-dmage-in-vitro 8/8
F.M.D. Chequer et al. / Mutation Research 777 (2015) 54–61 61
[15] E. Jerschow, J.J. Hostynek, H.I. Maibach, Allergic contact dermatitis elicitationthresholds of potent allergens in humans, Food Chem. Toxicol.: Int. J. Publ. Br.Ind. Biol. Res. Assoc. 39 (2001) 1095–1108.
[16] S. Kita, T. Kobayashi, H. Kutsuna, A.M. Kligman, Human maximization test-ing of D&C Yellow no. 10 and Yellow no. 11, Contact Dermat. 1 1 (1984)210–213.
[17] E. Zeiger, B. Anderson,S. Haworth, T. Lawlor, K. Mortelmans, Salmonella muta-genicity tests: IV. Results from the testing of 300 chemicals, Environ. Mol.Mutagen. 11 (Suppl. 12) (1988) 1–157.
[18] F.M. Chequer, T.M. Lizier,R. deFelicio,M.V.Zanoni, H.M. Debonsi, N.P. Lopes,R.Marcos, D.P. de Oliveira, Analyses of thegenotoxic and mutagenic potential of
the products formed after the biotransformation of the azo dye Disperse Red1, Toxicol. In Vitro:Int. J. Publ. Assoc. BIBRA 25 (2011) 2054–2063.
[19] I.A.R.C. IARC, IARC Monographs on the Evaluation of Carcinogenic Risks toHumans.Some AromaticAmines,OrganicDyes,and RelatedExposures(volume99), 2010.
[20] N.P. Singh,M.T. McCoy,R.R. Tice, E.L. Schneider, A simpletechnique forquanti-tationof lowlevels ofDNA damagein individual cells,Exp.Cell Res. 175(1988)184–191.
[21] R.R. Tice, E. Agurell, D. Anderson, B. Burlinson, A. Hartmann, H. Kobayashi, Y.Miyamae, E. Rojas, J.C. Ryu, Y.F. Sasaki, Single cell gel/comet assay: guidelinesfor in vitro and in vivo genetic toxicology testing, Environ. Mol. Mutagen. 35(2000) 206–221.
[22] M. Fenech, Cytokinesis-block micronucleus cytome assay, Nat.Protoc. 2 (2007)1084–1104.
[23] A.T. Natarajan, F. Darroudi,Use of human hepatoma cells forin vitro metabolicactivation of chemical mutagens/carcinogens, Mutagenesis 6 (1991) 399–403.
[24] M. Fenech, Cytokinesis-block micronucleus assay evolves into a cytome assayof chromosomal instability, mitotic dysfunction and cell death, Mutat.Res. 600(2006) 58–66.
[25] D.A. Eastmond, J.D. Tucker, Identification of aneuploidy-inducing agents usingcytokinesis-blocked human lymphocytes and an antikinetochore antibody,Environ. Mol. Mutagenesis 13 (1989) 34–43.
[26] A.C.Guimaraes, L.M. Antunes, H.F. Ribeiro, A.K. dosSantos, P.C. Cardoso, P.L. deLima, A.D. Seabra, T.B. Pontes, C. Pessoa, M.O. de Moraes, B.C. Cavalcanti, C.M.Sombra, O. Bahia Mde, R.R. Burbano, Cytogeneticbiomonitoring of inhabitantsof a large uranium mineralization area: the municipalities of Monte Alegre,Prainha, and Alenquer, in the State of Para, Brazil, Cell Biol. Toxicol. 26 (2010)403–419.
[27] L. Kapka,A. Baumgartner,E. Siwinska,L.E. Knudsen,D. Anderson,D. Mielzynska,Environmental lead exposure increases micronuclei in children, Mutagenesis22 (2007) 201–207.
[28] T.M. Lizier, M.V. Boldrin Zanoni, Effect of ionic liquid on the determinationof aromatic amines as contaminants in hair dyes by liquid chromatographycoupled to electrochemical detection, Molecules 17 (2012) 7961–7979.
[29] M.M. Baizer, H. Lund, Organic Electrochemistry – An Introductionand a Guide,Second ed., Marcel Dekker Inc., New York, 1983.
[30] S. Bonassi, L. Hagmar, U. Stromberg, A.H. Montagud, H. Tinnerberg, A. Forni, P.
Heikkila,S. Wanders, P. Wilhardt,I.L. Hansteen,L.E. Knudsen, H. Norppa, Chro-mosomal aberrations in lymphocytes predict human cancer independently of exposure to carcinogens. European Study Group on Cytogenetic Biomarkersand Health, CancerRes. 60 (2000) 1619–1625.
[31] L. Hagmar, S. Bonassi, U. Stromberg, A. Brogger, L.E. Knudsen, H. Norppa, C.Reuterwall, Chromosomal aberrations in lymphocytes predict human cancer:a report fromthe EuropeanStudy Group on Cytogenetic Biomarkers andHealth(ESCH), Cancer Res. 58 (1998) 4117–4121.
[32] L. Luzhna, P. Kathiria, O. Kovalchuk, Micronuclei in genotoxicity assessment:from genetics to epigenetics and beyond, Front. Genet. 4 (2013) 131.
[33] K. Crasta, N.J. Ganem, R. Dagher, A.B. Lantermann, E.V. Ivanova, Y. Pan, L. Nezi,A. Protopopov, D. Chowdhury, D. Pellman, DNA breaks and chromosome pul-verization from errorsin mitosis, Nature 482 (2012) 53–58.
[34] L. Huc, A. Lemarie, F. Gueraud, C. Helies-Toussaint, Low concentrations of bisphenol A induce lipid accumulation mediated by the production of reac-tive oxygenspecies in themitochondria of HepG2 cells,Toxicol.In Vitro:Int. J.Publ. Assoc. BIBRA 26 (2012) 709–717.
[35] S. Knasmuller, W. Parzefall, R. Sanyal, S. Ecker, C. Schwab, M. Uhl, V. Mersch-Sundermann, G. Williamson, G. Hietsch, T. Langer, F. Darroudi, A.T. Natarajan,Use of metabolically competent human hepatoma cells for the detection of mutagens and antimutagens, Mutat. Res. 402 (1998) 185–202.
[36] M. Pezdirc, B. Zegura, M. Filipic, Genotoxicity and induction of DNA damageresponsive genes by food-borne heterocyclic aromatic amines in human hep-atoma HepG2 cells,Food Chem. Toxicol.:Int. J. Publ. Br.Ind. Biol. Res. Assoc.59(2013) 386–394.
[37] J.M. Serpeloni, G.R. Barcelos, J.P. Friedmann Angeli, A.Z. Mercadante, M. Lour-des Pires Bianchi, L.M. Antunes, Dietary carotenoidlutein protects against DNAdamage and alterations of the redox status induced by cisplatin in humanderived HepG2 cells Toxicol, In Vitro: Int. J . Publ. Assoc. BIBRA 26 (2012)288–294.
[38] S.Knasmuller,V. Mersch-Sundermann, S. Kevekordes, F.Darroudi,W.W. Huber,C. Hoelzl, J. Bichler, B.J. Majer, Use of human-derived liver cell lines for thedetectionof environmental and dietary genotoxicants;current stateof knowl-edge, Toxicology 198 (2004) 315–328.
[39] W. Q. Lu, X .N. Chen , F . Yue , C. J en ter , R. G minski, X. Y. Li, H. X ie, V. Mers ch-Sundermann, Studies on the in vivo and in vitro mutagenicity and the lipidperoxidation of chlorinated surface (drinking) water in rats and metabolicallycompetent human cells, Mutat. Res. 513 (2002) 151–157.
[40] M. Uhl,C. Helma, S. Knasmuller, Evaluationof thesingle cellgel electrophoresisassay with human hepatoma (Hep G2)cells, Mutat. Res. 468 (2000) 213–225.
[41] M. Ohno, Y. Ikenaka, M. Ishizuka, SudanIII dye strongly induces CYP1A1mRNAexpression in HepG2 cells, J. Biochem. Mol. Toxicol. 26 (2012) 16–22.
[42] B.Bressac,K.M.Galvin, T.J. Liang, K.J. Isselbacher,J.R.Wands, M. Ozturk, Abnor-mal structure and expressionof p53 gene in humanhepatocellular carcinoma,Proc. Natl. Acad. Sci. U. S. A. 87 (1990) 1973–1977.
[43] J.H. van Delft, E. van Agen, S.G. van Breda, M.H. Herwijnen, Y.C. Staal, J.C. Klein- jans, Discrimination of genotoxic from non-genotoxic carcinogens by geneexpression profiling, Carcinogenesis 25 (2004) 1265–1276.
[44] E.F.S.A., EFSA, Scientific opinion on the re-evaluation of quinoline yellow (E104) as food additive, EFSA J. 7 (2009) 1329.
[45] M. Po ul, G. J ar ry , M.O. Elhkim , J .M. P oul, Lack of geno to xic e ff ect of f oo ddyes amaranth, sunset yellow and tartrazine and their metabolites in the gutmicronucleus assay in mice, Food Chem. Toxicol.: Int. J. Publ. Br. Ind. Biol. Res.Assoc. 47 (2009) 443–448.
[46] Y.F. Sasaki, S. Kawaguchi, A. Kamaya, M. Ohshita, K. Kabasawa, K. Iwama, K.Taniguchi, S. Tsuda, The comet assay with 8 mouse organs: results with 39currently used food additives, Mutat. Res. 519 (2002) 103–119.
[47] R.S. Chhabra, Intestinal absorption and metabolism of xenobiotics, Environ.
Health Perspect. 33 (1979) 61–69.[48] B. Meunier,Metalloporphyrinsas versatilecatalysts foroxidationreactionsandoxidative DNA cleavage, Chem. Rev. 92 (1992) 1411–1456.
[49] P.R. Ortiz de Montellano, J.J. De Voss, Oxidizing species in the mechanism of cytochrome P450, Nat. ProductRep. 19 (2002) 477–493.
[50] G. Choudhary,Human health perspectives on environmental exposure to ben-zidine: a review, Chemosphere 32 (1996) 267–291.
[51] K. Chung, T.J. Hughes, L.D. Claxton, Comparison of the mutagenic speci-ficity induced by four nitro-group-containing aromatic amines in Salmonellatyphimurium hisgenes, Mutat. Res. 465 (2000) 165–171.