the aerobic cos of saltatort y locomotio in n the …hopping is n moro e economical than running to...
TRANSCRIPT

/. exp. Biol. 136, 273-288 (1988) 2 7 3Printed in Great Britain © The Company of Biologists Limited 1988
THE AEROBIC COST OF SALTATORY LOCOMOTION INTHE FOWLER'S TOAD (BUFO WOODHOUSEI FOWLERI)
BY MICHAEL WALTON AND BRUCE D. ANDERSON
Committee on Evolutionary Biology, The University of Chicago,1025 East 57th Street, Chicago, Illinois 60637, USA
Accepted 16 December 1987
Summary
Studies of kangaroos suggest that hopping provides energy savings duringlocomotion at high speeds, although studies of small mammals suggest thathopping is no more economical than running. To obtain comparative data onanurans, we exercised Fowler's toads (Bufo woodhousei fowled, 25-8 g) ontreadmills at speeds ranging from 0-09 to 0-63 km h"1 while measuring oxygenconsumption ( V Q J , endurance or hop kinematics. The toads walked at slowspeeds and hopped at fast speeds. Steady-state VQ, (VO2 ^ increased linearly withspeed to a maximum (VQ, max) of l-17mlO2g~1h~1 at 0-27 kmh"1 and was ninetimes the average pre-exercise V O r The maximum rate of oxygen consumptionduring treadmill exercise was comparable to VQ, max previously reported for lessnatural exercise regimes. At speeds ^0-27 km h"1, VQJ ra was independent ofspeed. At speeds ^0-36 km h"1, toads moved for over lh , but endurancedecreased sharply at higher speeds. Hop rate, hop length, hop height and angle oftake-off increased with speed. Hopping in B. w. fowleri was not less costly thanrunning in other animals of similar body size and was inefficient at convertingmetabolic to mechanical energy. The present study suggests that hopping in toads,as in small mammals, is not economical during sustained locomotion and is mostimportant during short bursts of high-intensity activity.
Introduction
In most terrestrial vertebrates (for a review, see Bennett, 1985), the rate ofoxygen consumption (VOz) increases linearly with speed of locomotion to amaximum (VO2,max) and is thereafter independent of speed. The speed at whichVo2lmax occurs is called the maximal aerobic speed (MAS). Because VO2 ismaximal and constant at speeds ^MAS, these speeds are increasingly supportedby anaerobic metabolism and are generally not sustainable (Bennett, 1985).
Although saltatory locomotion occurs in several orders of vertebrates, theenergetics of saltatory transport are understood for only a few mammalian species.Surprisingly, an early study (Dawson & Taylor, 1973) of locomotor energetics inred kangaroos, Megaleia rufa (18-28 kg), showed that the aerobic cost of saltatory
Key words: locomotion, toads, anurans, energetics, aerobic metabolism.

274 M. WALTON AND B. D. ANDERSON
transport actually decreases with increased speed. This pattern obviously contrastssharply with the typical pattern of metabolic support for locomotion describedabove. Moreover, a red kangaroo hopping at rapid sustainable speeds expends lessenergy at a given speed than a running quadruped of similar body mass. Thekangaroo's economical locomotion at high sustainable speeds has been attributedto the storage and recovery of strain energy from elastic elements in the legs(Alexander & Vernon, 1975; Cavagna, Heglund & Taylor, 1977).
Subsequently, other workers examined the metabolic cost of saltatory loco-motion in small mammals (Baudinette, Nagle & Scott, 1976; Dawson, 1976;Thompson, MacMillen, Burke & Taylor, 1980). With perhaps one exception(Dawson, 1976), the small mammal data suggest that kangaroos are exceptionaland that hopping is not generally less costly than running (Thompson et al. 1980).The difference in locomotor energetics between small and large mammalianhoppers appears to be associated with morphological differences in elastic storage(Biewener, Alexander & Heglund, 1981; Emerson, 1985).
Saltatory locomotion is not restricted to mammals, but also occurs in birds,anurans and invertebrates. These other forms are useful for examining thegenerality of conclusions based on mammalian studies. Therefore, we examinedlocomotor energetics, endurance and kinematics of an anuran, Fowler's toad(Bufo woodhousei fowled), exercising on a treadmill at controlled speeds. Weaddressed the question of whether saltatory locomotion at fast sustainable speedsprovides energy savings in B. w. fowleri as it does in the red kangaroo. Subsumedin this question are two ancillary issues. First, is hopping in toads less expensivethan running or walking by animals of a similar body mass? Second, how efficientare toads at converting metabolic energy input into mechanical energy output?High efficiency may indicate a significant contribution from elastic storage.
Our data also bear upon amphibian activity metabolism in general. Manyprevious studies have attempted to measure VO2,max by shocking or overturninganurans in rotating respirometer chambers (reviewed by Taigen & Pough, 1985).Only Taigen & Beuchat (1984) related the rate of chamber rotation (presumablyspeed of movement) to VO2. Although these studies have revealed differencesamong anuran groups in activity metabolism, they have been criticized (see below)because their exercise regime did not elicit natural locomotion and may not elicitVo2,max- Therefore, an additional objective of this study was to determine iflocomotion on a treadmill, which resembles natural locomotion, yields a VO2 max
similar to that of toads in rotating respirometer jars.
Materials and methods
Animals
Fowler's toads, B. w. fowled, [body mass = 25-80 ± 1-48g ( ± S . D . ) ] wereobtained at the Indiana Dunes National Lakeshore, Porter Co., IN, and from acommercial supplier. Toads were maintained at 21 °C on a 14h:10h light: dark

Toad locomotion energetics 275
photoperiod centred on 13.00h local time. Toads were fed crickets (Achetadomestica) twice weekly until 5 days before experiments. All experimentalmeasurements were made at 21 °C.
Oxygen consumption
Measurements of VQJ during rest, sustained hopping and recovery were made ina gas-tight clear acrylic respirometer containing a variable-speed treadmill(Herreid, 1981; Full, 1986). The chamber in which the animals exercised consistedof a half-cylinder 25-4cm long, 12-7cm wide and 6-4cm tall at its highest point.Preliminary trials with unrestrained toads on a larger treadmill used for kinematicstudies (see below) indicated that the toads in the respirometer had sufficient roomto allow unencumbered hopping. The interior of the treadmill was moistened with10ml of distilled water before the toads were placed inside. Toads rested withinthe treadmill in dim light for 45min to 1 h before exercise. Resting VO2 wasmeasured during the last 10-15 min of the rest period. The toads were thenexercised for 30min at a single speed. The speeds were 0-09, 0-18, 0-27, 0-36,0-54 km h"1. An attempt was made to exercise each toad at each speed. Toadswere run only once a day and were rested for at least 2 days between trials. Trialsin which toads did not locomote consistently were not used.
Oxygen consumption was measured continuously using open-flow respirometry.An air pump drew humidified air through the system at 500 ml min"1. Incurrentfractional oxygen content (FIQJ was compared to the excurrent fractional oxygencontent ( F E O J with a dual-channel oxygen analyser (Ametek Applied Electro-chemistry, Model S-3A). Before analysis, gas was drawn through columns ofDrierite® and Ascarite® to remove water and CO2, respectively. Output from theoxygen analyser was run through an analogue-to-digital converter (Isaac 41Asystem, Cyborg) and into a microcomputer (Apple II+). Data were collected at10-s intervals.
Instantaneous V ^ was calculated from FIQ^ and FEO2 (Full & Herreid, 1983,1984; Herreid, 1981). Given any two measurements separated by a brief interval,the flow rate (V) and the 'washout' characteristics of the chamber (Feq) (theequilibrium of FEOZ inside the metabolic chamber) can be calculated andsubstituted for FEQ2 in a standard equation:
VO2 = V(Fi O 2 -F e q ) / ( ( l -F i O 2 )
to estimate instantaneous V O r Rates of oxygen consumption are reported asml O2 at STPD.
Anaerobic metabolism is known to contribute significantly to support of activityin amphibians moving at high, non-sustainable speeds (Taigen & Beuchat, 1984;Feder, 1986; Full, 1986). Inasmuch as our primary interest was in locomotion atlower, sustainable speeds, we chose not to undertake whole-body assays of lactateproduction, which necessitate killing large numbers of animals.

276 M. WALTON AND B. D. ANDERSON
Endurance
Endurance was measured by exercising toads to exhaustion on a motorizedtreadmill. Toads were placed individually on the treadmill and allowed to rest indim light for 15-30 min. The treadmill belt was moistened and humidified air wasdrawn through the chamber. Treadmill speeds ranged from 018 to 0-63 km h"1.Several toads were prodded during the experiments to encourage movement.Prodding consisted of quickly reversing the direction of the treadmill belt andcausing the toad to contact the back wall of the chamber. Prodding was requiredonly during the first few minutes of exercise to elicit steady movement and whentoads began to fatigue. At slow speeds, toads often hopped or walked indefinitelywithout fatigue; these trials ceased after 2 h. Otherwise, exhaustion was defined asthe time when a toad (1) did not keep pace with the treadmill, (2) lost balance andturned over on its back, and (3) did not respond to three successive proddings(Full, 1986). Toads were exercised once a day and rested for at least 2 daysbetween trials. Trials in which toads did not locomote consistently were discarded.
Kinematics: hop rate, length, height and angle of take-off
Hop rate (hopss"1), hop length (cm), hop height (cm) and angle of take-off(degrees) were determined from videotapes recorded while toads hopped on amotorized treadmill. The treadmill chamber consisted of an open-top rectangularbox made of transparent acrylic 41cm long, 15 cm wide and 21-6cm high. Acoordinate grid of 1-cm squares was placed on the chamber wall opposite aPanasonic WV-3230 video camera/recorder. The camera was mounted on a tripod1 m from the treadmill. The camera was focused so that the treadmill chamber andgrid filled the field of view. The toads were videotaped while moving at each of thefive speeds used in the V ^ experiments. Recordings were made consecutively atthese speeds and the order of speeds was randomized for each toad. Maximumheight and length of hops were measured against the grid as the videotapes wereadvanced in 0-03-0-04s increments. Angle of take-off was calculated usingstandard ballistics equations:
L = (v2sin20)/g ,
H = (v2sin20)/2g ,
where L is hop length, H is hop height, v is take-off velocity, g is acceleration dueto gravity, and d is angle of take-off. These two equations were solvedsimultaneously for 6 to obtain the relationship:
6 = arctan(4H/L) .
Five to ten hops were measured for each toad at each speed. Hop rates weremeasured by counting the number of hops taken during 30- to 60-s intervals ofconsecutive hopping at each speed. Two or three replicates were recorded for eachtoad at each speed and averaged to obtain the final value.

Toad locomotion energetics 277
Statistics
Results in the text are given as means ± standard deviations. Standard corre-lation and regression analyses were used to examine the responses of VQ2 and hopkinematics to changes in treadmill speed.
ResultsGait transition
Toads used two gaits: walking and hopping. At slow speeds, walking toads hadthree feet on the substrate at all times, but toads often had only two feet on thesubstrate at fast walking speeds. Hopping, as defined by Emerson (1979),consisted of short jumps (<8-9 times body length). Toads showed a gait transitionfrom walking at slow speeds to hopping at fast speeds (Fig. 1). At the slowestspeed (0-09 km rT1), all the toads tested walked for some portion of theexperimental period. Several animals walked nearly the entire time, but mostwalked and hopped intermittently. When speed was doubled (0-18kmh"1), thenumber of toads that walked halved. At faster speeds, the number of toadsobserved to walk continued to decrease until, at the fastest speeds (2sO36 km h"1),only a few toads walked during the experiments. At speeds >0-36kmh~1 walkingwas confined to one or two steps taken to regain position after landing from a hop.
Oxygen consumption
Pre-exercise Vch averaged 0-13 ± 0-01 mlO2g~1h"1. Because toads exploredthe respirometer during the resting phase of experiments, this pre-exercise VO2
may be greater than the standard rate of O2 consumption.
100
80
•cZ 60
40
20
009 018 0-27Speed (km h"1)
0-36 0-54
Fig. 1. Gait transitions in response to treadmill speed in Bufo w. fowleri. The numberof animals observed to walk during some interval of the experimental period,expressed as percentage of the total number of animals, is plotted as a function oftreadmill speed.

278 M. WALTON AND B. D . ANDERSON
During the first 5-10 min of exercise, VO2 increased to an approximate steady-state level (Vo2,ss)> after which it usually remained constant but sometimesincreased slowly, especially at higher speeds (Fig. 2). The VO2>ss values reportedhere are the average for the last 10 min of exercise.
Steady-state oxygen consumption (VO2 55) increased significantly with speedbetween 0-09 and 0-27 km h"1 (Fig. 3A). The regression equation relating speed toVoi.ss for this range of speeds was VO2 iSS (mlO2g~1h"1) = 3-87xspeed(km h~1)+0-224 (r = 0-92). The slope of this line, often termed the minimum costof transport (Cmin) (Taylor, Schmidt-Nielsen & Raab, 1970), had a 95 %confidence interval of 2-9-4-8mlO2g~1 km"1. The hypothesis of linearity acrossthis speed range was not rejected (F[l,13] =0-571, F>0-25). At speeds2*0-27 kmrT1, VO2,SS was independent of speed (b =-0-016, P>0-25) andaveraged 1-13 ±0-07mlO2g~1h~1. The greatest average VO2 ss was attained at0-27kmh"1 and was l-17±0-llmlO2g~1 h"1, nine times greater than pre-exercise VO2.
Endurance
At low speeds (=£0-27 km h"1), six toads sustained locomotion for more than 2h(Fig. 3B). At 0-36 km h"1, one of five toads was exhausted within 2 h, but even thistoad sustained hopping for longer than 1-5 h. Endurance time decreased sharply athigher speeds to <10min at 0-63 km h"1.
Fig. 2. Oxygen consumption (VO2) of an individual Bufo w. fowled (30 g) during30 min of exercise on a treadmill atO-09 (filled diamonds), 0-18 (open diamonds), 0-27(filled triangles) and 0-45 km h"1 (open squares). Data for 0-27 km h"1 exemplify theresponse of an animal that refused to move during the first few minutes of exercise (seetext).
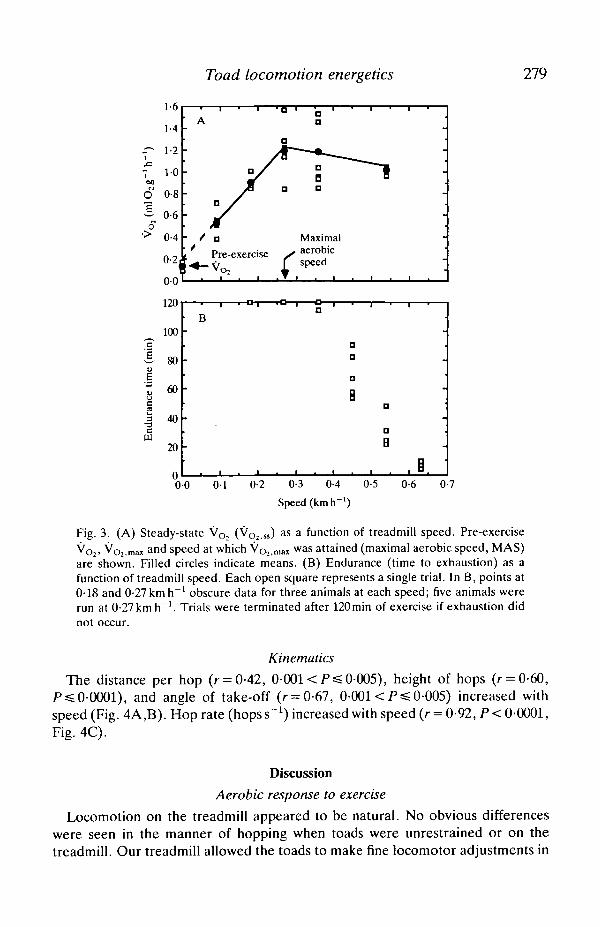
Toad locomotion energetics 279
7j=
i00
o
>
1-6
1-4
1-2
10
0-8
0-6
0-4
0 0
120
3
UJ
aa
a a
• / a Maximal' Pre-exercise / a e r o b i c
[ speed' o .
I . • I . I . I . I
120
100
80
60
40
20
n
•
B•
•
•
-
•
. i
o
aa
D
B
i • i •
-
-
-
D-
D
B
. . . 1. '00 0 1 0-2 0-3 0-4
Speed (km h"1)
0-5 0-6 0-7
Fig. 3. (A) Steady-state V o , (VO2,ss) as a function of treadmill speed. Pre-exerciseVO2, V ^ m!Ut and speed at which Voj.mai was attained (maximal aerobic speed, MAS)are shown. Filled circles indicate means. (B) Endurance (time to exhaustion) as afunction of treadmill speed. Each open square represents a single trial. In B, points at0-18 and 0-27 km h~' obscure data for three animals at each speed; five animals wererun at 0-27 km h"1. Trials were terminated after 120 min of exercise if exhaustion didnot occur.
Kinematics
The distance per hop (r = 0-42, 0-001<P=£ 0-005), height of hops (r = 0-60,Ps=0-0001), and angle of take-off (r = 0-67, 0 - 0 0 K P =£0-005) increased withspeed (Fig. 4A,B). Hop rate (hops s"1) increased with speed (/• = 0-92, P < 0-0001,Fig. AC).
Discussion
Aerobic response to exercise
Locomotion on the treadmill appeared to be natural. No obvious differenceswere seen in the manner of hopping when toads were unrestrained or on thetreadmill. Our treadmill allowed the toads to make fine locomotor adjustments in
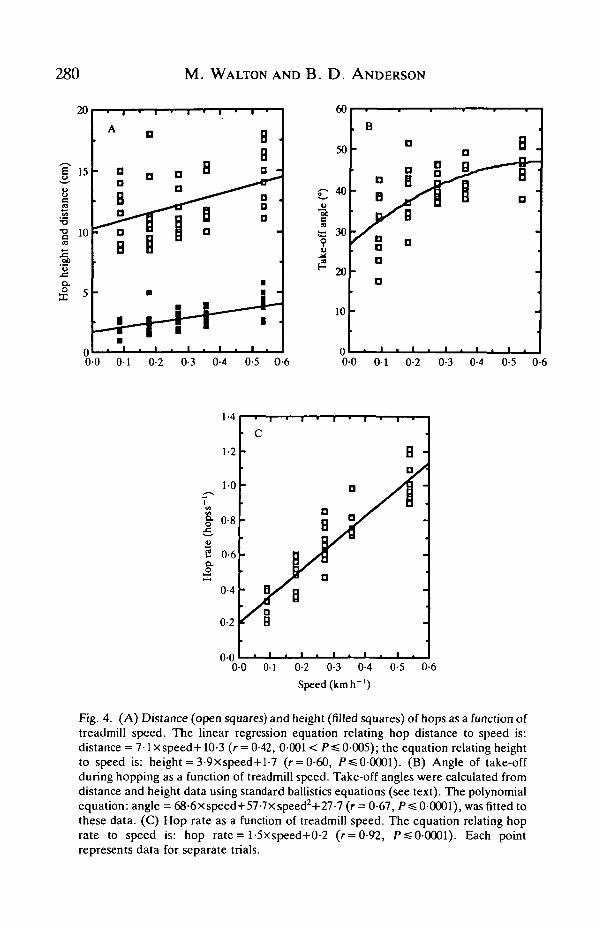
280 M. WALTON AND B. D. ANDERSON
0-0 01 0-3 0-4 0-5 0-6 0-4 0-5 0-6
0-2 0-3 0-4
Speed (km h"1)
0-5 0-6
Fig. 4. (A) Distance (open squares) and height (filled squares) of hops as a function oftreadmill speed. The linear regression equation relating hop distance to speed is:distance = 7-lXspeed+10-3 (r= 0-42, 0001 < P =£0-005); the equation relating heightto speed is: height = 3-9xspeed+l-7 (r = 0-60, PssO-0001). (B) Angle of take-offduring hopping as a function of treadmill speed. Take-off angles were calculated fromdistance and height data using standard ballistics equations (see text). The polynomialequation: angle = 68-6 x speed+57-7xspeed2+27-7 (r = 0-67, />=£ 0-0001), was fitted tothese data. (C) Hop rate as a function of treadmill speed. The equation relating hoprate to speed is: hop rate = l-5xspeed+0-2 (r=0-92, P^O-OOOl). Each pointrepresents data for separate trials.

Toad locomotion energetics 281
response to speed (i.e. changes in gait and hop kinematics). Therefore, we areconfident that our data reflect the aerobic metabolic support of normal, sustain-able locomotion in B. w. fowleri.
The response of VOj-ss to speed in B. w. fowleri resembled what may beconsidered the 'typical' pattern of locomotor energetics for vertebrates: a steady,linear increase of V ^ with speed throughout the range of sustainable speeds,above which V ^ is constant and stamina decreases (Taylor et al. 1970; Taylor,1977; Taylor, Heglund & Maloiy, 1982; Bennett, 1985). However, B. w. fowleddeparted from the typical pattern in that although stamina decreased, it remainedhigh at speeds above which VQ, max was attained. For example, toads exercised at0-45 km h"1 hopped for 50-90min (Fig. 3B). One possible explanation for thisphenomenon is that hopping provides energy savings, presumably through theelastic storage of strain energy. Hence, as in kangaroos, hopping by toads atmoderate speeds may be largely supported by aerobic metabolism supplementedby the recovery of strain energy. However, our data suggest that the contributionof strain energy is small (see below). Another possible explanation is that,although toads use anaerobic sources at these speeds, they might be able toaccumulate a large lactate load without incurring fatigue. We made no measure-ments of anaerobic metabolism, but American toads, B. americanus, showelevated lactate levels at sustainable speeds (Taigen & Beuchat, 1984). Unfortu-nately, Taigen & Beuchat (1984) exercised their animals for only a few minutes.Sustained movement may be fuelled by aerobic metabolism after an initialcontribution of anaerobiosis to support the first minutes of activity (Bennett,1985). The mechanisms of stamina, fatigue and recovery are problematic inamphibians (Bennett, 1974; Cushman, Packard & Boardman, 1976; Fitts &Holloszy, 1976; Feder & Olsen, 1978; Hutchison & Turney, 1975; Hutchison,Miller & Gratz, 1981) and merit further investigation.
Minimum cost of transport
The cost of locomotion in hopping toads was no less expensive than running byother animals of a similar body size. The minimum cost of transport (Cmin)expresses the amount of energy required to move a given distance, and isfrequently used for comparisons among taxa and modes of locomotion (Bennett,1985). The minimum cost of hopping transport in toads is considerably greaterthan in running mammals (Taylor et al. 1982) or running lizards (John-Alder,Garland & Bennett, 1986). For example, Cmin of B. w. fowleri (=3-8mlO2
g"1 km"1) is 2-4 times greater than, and outside the 95 % confidence limits of, thatpredicted for a lizard of the same body mass (John-Alder et al. 1986). Moreover,the Cmin of B. w. fowleri is greater than those of salamanders (Ambystomatigrinum, A. laterale, Desmognathus ocrophaeus, Plethodon jordani) run on atreadmill (Full, 1986; Full, Anderson, Finnerty & Feder, 1988). In fact, the Cmin ofB. w. fowleri is 12-2 times greater than that of comparably sized A. tigrinum (Fullet al. 1988).

282 M. WALTON AND B. D. ANDERSON
Although sustained hopping by toads was at least as costly as walking or runningby other animals, hopping at fast speeds may have been less costly for toads thanwalking at those speeds. This, of course, would account for the observed change ingait (Fig. 1). Unfortunately, because toads both walked and hopped at slowspeeds and rarely walked at fast speeds, we could not discern differences inmetabolic cost between the two gaits.
Efficiency of hopping
What fraction of the aerobic metabolic energy input is converted to mechanicalenergy output during locomotion in B. w. fowled! The kinetic energy required toproduce a hop can be calculated from the standard ballistics equations:
L = 2Ek(sin20/mg) ,
H = Ek(sin20/mg) ,
where m is mass, and Ek is kinetic energy (Emerson, 1985). This estimate of Ek isminimal, because it ignores the energy required to move limbs (Cavagna et al.1977). Power output can then be obtained as follows:
power output = Ek x hop rate .
In B. w. fowled, mechanical power output increased linearly across the range ofspeeds tested (r = 0-95, P< 0-0001; Fig. 5A). The energetic efficiency of loco-motion was low in B. w. fowled, ranging from 4 to 14 % (Fig. 5B) of the aerobicmetabolic power input (VO2,SS converted to units of \Vkg~1). Efficiency increasedat non-sustainable speeds (>0-27kmh~L). However, at these speeds, anaerobicsupport for activity certainly became important. Therefore, aerobic powerestimates for these speeds underestimate the total metabolic power input forhopping (aerobic+anaerobic) and overestimate efficiency. The low efficiencies atsustainable speeds are comparable to those for small mammalian hoppejs(Biewener et al. 1981).
The low efficiency estimates for B. w. fowled suggest that elastic recoil does notcontribute a substantial amount of energy to sustained hopping. The efficiency ofconverting metabolic to mechanical energy ranges from 24 to 76% in kangaroos,depending on hopping speed (Cavagna et al. 1977). The maximal efficiency for theconversion of metabolic to mechanical energy by muscle contraction alone hasbeen found to be 25 % (Hill, 1939; Gibbs & Gibson, 1972; Wendt & Gibbs, 1973).Thus, elastic recoil must provide the additional increments of energy used forhopping in kangaroos. In fact, as much as 70% of the kinetic energy that akangaroo generates at the start of a hop can be recovered upon landing(Alexander & Vernon, 1975; Cavagna et al. 1977). Toads, however, showed lowconversion efficiencies at all, and particularly sustainable, speeds (<14%,Fig. 5B), suggesting that most, if not all, of the energy required for sustainedhopping was derived directly from muscle contraction.
These findings are consistent with our observations of sustained hopping bytoads. Unlike kangaroos, they do not resemble a 'bouncing ball or the action of a
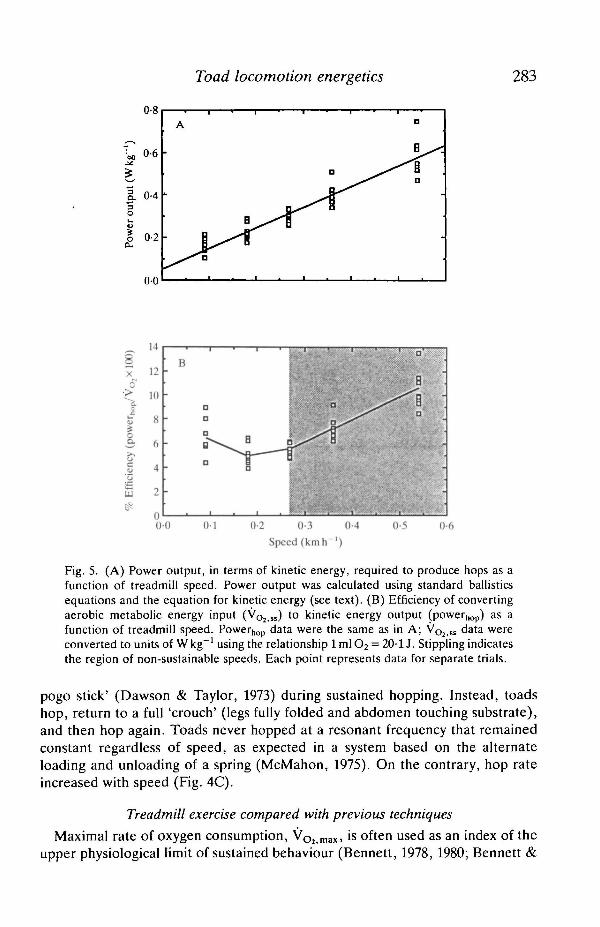
Toad locomotion energetics 283
X
6'
GC
0-1 0-2 0-3 0-4Speed (kmh ')
0-6
Fig. 5. (A) Power output, in terms of kinetic energy, required to produce hops as afunction of treadmill speed. Power output was calculated using standard ballisticsequations and the equation for kinetic energy (see text). (B) Efficiency of convertingaerobic metabolic energy input (VO2 „) to kinetic energy output (power^) as afunction of treadmill speed. Powerhop data were the same as in A; VO2,ss data wereconverted to units of Wkg"1 using the relationship lmlO2 = 20-1J. Stippling indicatesthe region of non-sustainable speeds. Each point represents data for separate trials.
pogo stick' (Dawson & Taylor, 1973) during sustained hopping. Instead, toadshop, return to a full 'crouch' (legs fully folded and abdomen touching substrate),and then hop again. Toads never hopped at a resonant frequency that remainedconstant regardless of speed, as expected in a system based on the alternateloading and unloading of a spring (McMahon, 1975). On the contrary, hop rateincreased with speed (Fig. 4C).
Treadmill exercise compared with previous techniques
Maximal rate of oxygen consumption, VO2,max> ' s often used as an index of theupper physiological limit of sustained behaviour (Bennett, 1978, 1980; Bennett &

284 M. WALTON AND B. D. ANDERSON
Ruben, 1979; Taigen, 1983). From this perspective, covariation of V ^ max withsome aspect of behaviour has been taken as evidence for the adaptive significance°f VO2 max, such that high V ^ max should potentiate higher levels of activity and,perhaps, a more complex behavioural repertoire than low VO2,max (Bennett, 1978,1980; Bennett & Ruben, 1979; Taigen, 1983). Consistent with this hypothesis,familial, species and even populational differences in VO2 max among anurans havebeen correlated with differences in mode of locomotion (Bennett & Licht, 1973,1974), foraging behaviour (Emerson, 1976; Taigen & Pough, 1983, 1985; Taigen,Emerson & Pough, 1982; Toft, 1980, 1981) and predator avoidance (Bennett &Licht, 1974; Hutchison & Miller, 1979).
Recently, these studies have been criticized because (1) the methods ofstimulation do not elicit normal locomotion (Full, 1986), (2) different methods ofstimulation may yield different values of VO2 max for the same species (Hillman,Shoemaker & Putnam, 1979; Walsberg, 1986), and (3) some anurans attain higherVQ2 levels during vocalization than during exercise in rotating chambers (Taigen &Wells, 1985; Taigen, Wells & Marsh, 1985). However, arguments advanced insupport of previous measures of V,^ max are that (1) the measures are reproduc-ible for individuals, provided that the same method of stimulation is applied to allanimals (Wells & Taigen, 1984; Walsberg, 1986; Walton, 1988), (2) VQ, increaseswith intensity of exercise, even though the exercise regime (i.e. rotation ofrespirometer jars) is not natural (Taigen & Beuchat, 1984; M. Walton, unpub-lished data), (3) V ^ during vigorous exercise covaries with the oxidativecapacities of anuran locomotor musculature (Bennett, 1974; Putnam & Bennett,1983; Taigen et al. 1985), and (4) VO2 during vigorous exercise, although notmaximal, is a good predictor of VO2,max (Taigen & Pough, 1985; but see Walsberg,1986).
Our results validate measurements of maximum VO2 during non-normallocomotion in rotating respirometers. The VO2,max reported here was similar tovalues obtained for toads that were overturned in closed respirometer jars atsimilar speeds (Table 1). However, if the cost of locomotion is at issue, our datasuggest that more natural means of locomotion are necessary to yield acceptableresults. The small cylindrical respirometers typically used in earlier studies do notallow the fine locomotor adjustments to speed found in our study. For example, inTaigen & Beuchat's (1984) study, a 'moderate' level of activity encompassedspeeds of rotation of 0-5-1-5 body lengths s-1. In our study, an equivalent speedrange (009-0-27 km h"1) produced a change in gait, a 74 % increase in hop rate, a35 % increase in hop height and a 98 % increase in VO2 (Figs 3,4).
In conclusion, sustained hopping in B. w. fowleri was not less expensive thanrunning in animals of a similar body mass, and was relatively inefficient atconverting aerobic energy input into kinetic energy output. Thus, as in smallmammals (Thompson et al. 1980), the energetic advantages associated withsustained hopping are probably small. Hopping at fast speeds may be less costlyfor toads than walking or running at those speeds. However, because toads bothwalked and hopped at all speeds and rarely walked at speeds above 0-lSkmh"1,
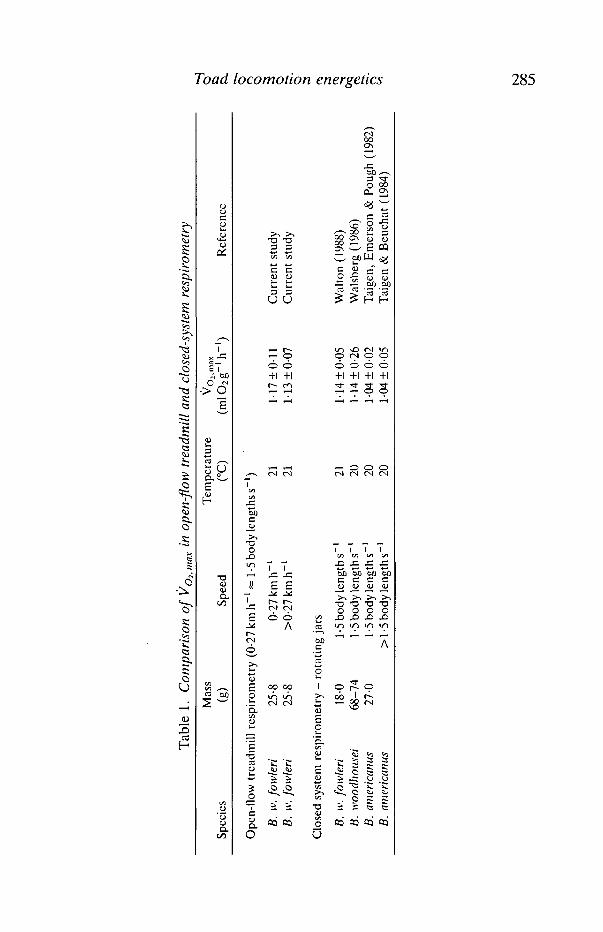
Tab
le 1
. C
ompa
riso
n of
VO
2 m
ax i
n op
en-f
low
tre
adm
ill
and
clos
ed-s
yste
m r
espi
rom
etry
Spec
ies
Ope
n-flo
w t
read
mill
B.
w.
fow
leri
B.
w.
fow
leri
Mas
s(g
)
resp
irom
etrj
25-8
25-8
Spee
dT
empe
ratu
re
/ (0
-27k
mh"
'~l-
5bod
ylen
gths
s"')
0-27
km
h"1
>0-
27km
h"'
Clo
sed
syst
em r
espi
rom
etry
- r
otat
ing
jars
B.
w.
fow
leri
B. w
oodh
ouse
iB
. am
eric
anus
B.
amer
ican
us
18-0
68-7
427
0
1-5
body
leng
ths
'1-
5 bo
dy le
ngth
s"1
1-5
body
leng
ths"
1
>1
-5 b
ody
leng
ths"
1
21 21 21 20 20 20
Vo 2
.ma X
117
+ 0
111-
13 ±
0-07
1-14
±0-
051-
14 ±
0-26
1-04
±0-
021-
04 ±
0-05
Ref
eren
ce
Cur
rent
stu
dyC
urre
nt s
tudy
Wal
ton
(198
8)W
alsb
erg
(198
6)T
aige
n, E
mer
son
& P
ough
(19
82)
Tai
gen
& B
euch
at (
1984
)
Toad locomotio 3 re lergetics 00

286 M. WALTON AND B. D. ANDERSON
the cost of walking and running transport was not quantified. The ability of toadsto sustain hopping for long periods at speeds above the apparent MAS isproblematic. Additional work concerning the anaerobic support for sustainedhopping is required to resolve this point.
Others have argued that rapid escape from predators is the major selectiveadvantage of saltatory locomotion in anurans and small mammals (Gans &Parson, 1966; Thompson etal. 1980). The anti-predator response of toads has beenclassified as 'static' (Taigen et al. 1982), but our experience has been that B. w.fowled usually hops quickly away from pursuers. Bufo w. fowleri can hop at>1 km h~', but only for about 30s (M. Walton, in preparation). Emerson (1978)suggested that anuran morphology allows the maintenance of constant acceler-ation over a range of body sizes. Hence quickness, not endurance, appears to bethe primary benefit of saltatory locomotion in anurans.
We thank Martin Feder for assistance during the research and comments on themanuscript. We also thank Robert Full and Andrew Biewener for their helpfulcomments on the manuscript. Financial support was provided by US Public HealthService Predoctoral Training Grant in Genetics and Regulation 5-T32-GM-07197to MW and by National Science Foundation Grant DCB84-16121 to M. E. Feder.
ReferencesALEXANDER, R. MCN. & VERNON, A. (1975). The mechanics of hopping by kangaroos
(Macropodidae).y. Zooi, Lond. 177,265-303.BAUDINETTE, R. V., NAGLE, K. A. & SCOTT, R. A. D. (1976). Locomotory energetics in a
marsupial {Antechrnomys spenceri) and a rodent {Notomys alexis). Experientia 32, 583.BENNETT, A. F. (1974). Enzymatic correlates of activity metabolism in anuran amphibians. Am.
J. Physiol. 226, 1149-1151.BENNETT, A. F. (1978). Activity metabolism of the lower vertebrates. A. Rev. Physiol. 400,
447-469.BENNETT, A. F. (1980). The metabolic foundations of vertebrate behavior. Biosci. 30, 452-456.BENNETT, A. F. (1985). Energetics and locomotion. In Functional Vertebrate Morphology (ed.
M. Hildebrand, D. M. Bramble, K. F. Liem & D. B. Wake), pp. 173-184. Cambridge, MA:Belknap Press.
BENNETT, A. F. & LICHT, P. (1973). Relative contributions of anaerobic and aerobic energyproduction during activity in Amphibia. J. comp. Physiol. 87, 351-360.
BENNETT, A. F. & LICHT, P. (1974). Anaerobic metabolism during activity in amphibians.Comp. Biochem. Physiol. 48A, 319-327.
BENNETT, A. F. & RUBEN, J. A. (1979). Endotheimy and activity in vertebrates. Science 206,649-654.
BIEWENER, A., ALEXANDER, R. MCN. & HEGLUND, N. (1981). Elastic energy storage in thehopping of kangaroo rats {Dipodomys spectabilis). J. ZooL, Lond. 195, 369-383.
CAVAGNA, G. A., HEGLUND, N. C. & TAYLOR, C. R. (1977). Mechanical work in terrestriallocomotion: two basic mechanisms for minimizing energy expenditure. Am. J. Physiol. 233,R243-R261.
CUSHMAN, J. R., PACKARD, G. C. & BOARDMAN, T. J. (1976). Concentrations of lactic acid inneotenic and transformed tiger salamanders (Ambysioma tigrinum) before and after activity./ . comp. Physiol. 112, 272-281.
DAWSON, T. J. (1976). Energetic cost of locomotion in Australian hopping mice. Nature, Lond.259, 305-306.

Toad locomotion energetics 287
DAWSON, T. J. & TAYLOR, C. R. (1973). Energetic cost of locomotion in kangaroos. Nature,Lond. 246, 313-314.
EMERSON, S. B. (1976). Burrowing in frogs. J. Morph. 149, 437-458.EMERSON, S. B. (1978). Allometry and jumping in frogs: helping the twain to meet. Evolution
32, 551-564.EMERSON, S. B. (1979). The ilio-sacral articulation in frogs: form and function. J. Linn. Soc.
(Zool.) 11, 153-168.EMERSON, S. B. (1985). Jumping and leaping. In Functional Vertebrate Morphology (ed.
M. Hildebrand, D. M. Bramble, K. F. Liem & D. B. Wake), pp. 58-72. Cambridge, MA:Belknap Press.
FEDER, M. E. (1986). Effect of thermal acclimation on locomotor energetics and locomotorperformance in a lungless salamander, Desmognathus ochrophaeus. J. exp. Biol. 121,271-283.
FEDER, M. E. & OLSEN, L. E. (1978). Behavioral and physiological correlates of recovery fromexhaustion in the lungless salamander Batrachoseps attenuatus (Amphibia: Plethodontidae).J. comp. Physiol. 128, 101-107.
Fnrs, R. H. & HOLLOSZY, R. H. (1976). Lactate and contractile force in frog muscle duringdevelopment of fatigue and recovery. Am. J. Physiol. 231, 430-433.
FULL, R. J. (1986). Locomotion without lungs: energetics and performance of a lunglesssalamander. Am. J. Physiol. 251, R775-R780.
FULL, R. J., ANDERSON, B. D., FINNERTY, C. M. & FEDER, M. E. (1988). Exercising with andwithout lungs. I. The effects of metabolic cost, maximal oxygen transport and body size onterrestrial locomotion in salamander species. J. exp. Biol. (in press).
FULL, R. J. & HERREID, C. F. (1983). The aerobic response to exercise of the fastest land crab.Am. J. Physiol. 244, R530-R536.
FULL, R. J. & HERREID, C. F. (1984). Fiddler crab exercise: the energetic cost of runningsideways. 7. exp. Biol. 109, 141-161.
GANS, C. & PARSONS, T. S. (1966). On the origin of the jumping mechanism in frogs. Evolution20, 92-99.
GIBBS, C. L. & GIBSON, W. R. (1972). Energy production of rat soleus muscle. Am. J. Physiol.233, 864-871.
HERREID, C. F. (1981). Energetics of pedestrian arthropods. In Locomotion and Energeticsin Arthropods (ed. C. F. Herreid & C. R. Fourtner), pp. 491-526. New York: PlenumPress.
HILL, A. V. (1939). The mechanical efficiency of frog's muscle. Proc. R. Soc. Ser. B 127,434-451.
HILLMAN, S. S., SHOEMAKER, V. H., PUTNAM, R. & WITHERS, P. C. (1979). Reassessment ofaerobic metabolism in amphibians during activity. J. comp. Physiol. 128, 169-175.
HUTCHISON, V. H. & MILLER, K. (1979). Aerobic and anaerobic contributions to sustainedactivity in Xenopus laevis. Respir. Physiol. 38, 91-103.
HUTCHISON, V. H., MILLER, K. & GRATZ, R. K. (1981). The anaerobic contributions to sustainedactivity in the giant toad Bufo marinus. Comp. Biochem. Physiol. 69A, 693-696.
HUTCHISON, V. H. & TURNEY, L. D. (1975). Glucose and lactate concentrations during activityin the leopard frog, Rana pipiens. J. comp. Physiol. 99, 287-295.
JOHN-ALDER, H. B., GARLAND, T. & BENNETT, A. F. (1986). Locomotory capacities, oxygenconsumption, and the cost of locomotion of the shingle-back lizard (Trachydosaurus rugosus).Physiol. Zool. 59, 523-531.
MCMAHON, T. A. (1975). Using body size to understand the structural design of animals:quadrapedal locomotion. J. appl. Physiol. 39, 619-627.
PUTNAM, R. W. & BENNETT, A. F. (1983). Histochemical, enzymatic, and contractile propertiesof skeletal muscles of three anuran amphibians. Am. J. Physiol. 244, R558-R567.
TAIGEN, T. L. (1983). Activity metabolism of anuran amphibians: implications for the origin ofendothermy. Am. Nat. 121, 94-109.
TAIGEN, T. L. & BEUCHAT, C. A. (1984). Anaerobic threshold of anuran amphibians. Physiol.Zool. 57, 641-647.
TAIGEN, T. L., EMERSON, S. B. & POUGH, F. H. (1982). Ecological correlates of anuran exercisephysiology. Oecologia 52, 49-56.

288 M. WALTON AND B. D. ANDERSON
TAIGEN, T. L. & POUCH, F. H. (1983). Prey preference, foraging behavior, and metaboliccharacteristics of frogs. Am. Nat. 122, 509-520.
TAIGEN, T. L. & POUGH, F. H. (1985). Metabolic correlates of anuran behavior. Am. Zool. 25,987-997.
TAIGEN, T. L. & WELLS, K. D. (1985). Energetics of vocalization by an anuran amphibian {Hylaversicolor).J. comp. Physiol. 155B, 163-170.
TAIGEN, T. L., WELLS, K. D. & MARSH, R. L. (1985). The enzymatic basis of high metabolicrates in calling frogs. Physiol. Zool. 58, 719-726.
TAYLOR, C. R. (1977). The energetics of terrestrial locomotion and body size in vertebrates. InScale Effects in Animal Locomotion (ed. T. J. Pedley), pp. 127-141. New York: AcademicPress.
TAYLOR, C. R., HEGLUND, N. C. & MALOIY, G. M. O. (1982). Energetics and mechanics ofterrestrial locomotion. I. Metabolic energy consumption as a function of speed and body sizein birds and mammals. J. exp. Biol. 97, 1-21.
TAYLOR, C. R., SCHMIDT-NIELSEN, K. & RAAB, J. C. (1970). Scaling the energetic cost of runningto body size in mammals. Am. J. Physiol. 219, 1104-1107.
THOMPSON, S. D., MACMILLEN, R. E., BURKE, E. M. & TAYLOR, C. R. (1980). The energetic costof bipedal hopping in small mammals. Nature, Lond. 287, 223-224.
TOFT, C. (1980). Feeding ecology of thirteen syntopic species of anurans in a seasonal tropicalenvironment. Oecologia 45, 131-141.
TOFT, C. (1981). Feeding ecology of Panamanian litter anurans: patterns in diet and foragingmode. J. Herp. 15, 139-144.
WALSBERG, G. E. (1986). Comparison of two techniques for estimating the maximum aerobiccapacity of amphibians. Herpetologica 42, 389-394.
WALTON, M. (1988). Relationships among metabolic, locomotor, and field measures oforganismal performance in the Fowler's toad (Bufo woodhousei fowled). Physiol. Zool. (inpress).
WELLS, K. D. & TAIGEN, T. L. (1984). Reproductive behavior and aerobic capacities of maleAmerican toads {Bufo americanus): is behavior constrained by physiology. Herpetologica 40,292-298.
WENDT, I. R. & GIBBS, C. L. (1973). Energy production of rat extensor digitorum longusmuscle. Am. J. Physiol. 224, 1081-1086.