tetraspanins in viral infections: a fundamental role in viral biology?
TRANSCRIPT

JOURNAL OF VIROLOGY, Sept. 2005, p. 10839–10851 Vol. 79, No. 170022-538X/05/$08.00�0 doi:10.1128/JVI.79.17.10839–10851.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
MINIREVIEW
Tetraspanins in Viral Infections: a Fundamental Role in Viral Biology?F. Martin,1 D. M. Roth,2,3 D. A. Jans,2 C. W. Pouton,3 L. J. Partridge,4 P. N. Monk,1 and
G. W. Moseley2*Academic Neurology Unit, Division of Genomic Medicine, University of Sheffield, Sheffield, United Kingdom,1 Department ofBiochemistry and Molecular Biology, Monash University, Clayton, Victoria 3800, Australia,2 Victorian College of Pharmacy,
Monash University, 381 Royal Parade, Parkville, Victoria 3052, Australia,3 and Department of Molecular Biology andBiotechnology, University of Sheffield, Sheffield, United Kingdom4
The tetraspanins are a broadly expressed superfamily oftransmembrane glycoproteins with over 30 members found inhumans and with homologues conserved through distantly re-lated species, including insects, sponges, and fungi. Membersof this family appear to form large integrated signaling com-plexes or tetraspanin-enriched microdomains (TEMs) by theirassociation with a variety of transmembrane and intracellularsignaling/cytoskeletal proteins (49). These interactions link tet-raspanins to an array of physiological functions and, in conse-quence, to numerous endogenous pathologies, including can-cer development and inherited disorders (Table 1).
Tetraspanins are also known to have roles in the pathologyof infectious diseases such as diphtheria, malaria, and numer-ous viral infections (Table 1). The literature currently indicatesthat specific tetraspanin family members are selectively asso-ciated with specific viruses and affect multiple stages of infec-tivity, from initial cellular attachment to syncytium formationand viral particle release. Thus, the relationship of tetraspaninswith viruses appears to be particularly complex.
Here, we will consider this data in the context of recentdevelopments in tetraspanin biology, particularly in our under-standing of the architecture and function of TEMs. With thebenefit of recent insights into tetraspanin function in cell fu-sion events and intracellular trafficking, we discuss commonfeatures of tetraspanin/viral associations which indicate a fun-damental role for TEMs in a number of viral infections. Wewill also consider the existing therapeutic strategies for humanimmunodeficiency virus (HIV), hepatitis C virus (HCV), andhuman T-cell lymphotropic virus type 1 (HTLV-1), focusing onthe potential therapeutic value of targeting TEMs, using pep-tide reagents based on tetraspanin extracellular regions.
THE TETRASPANIN SUPERFAMILY
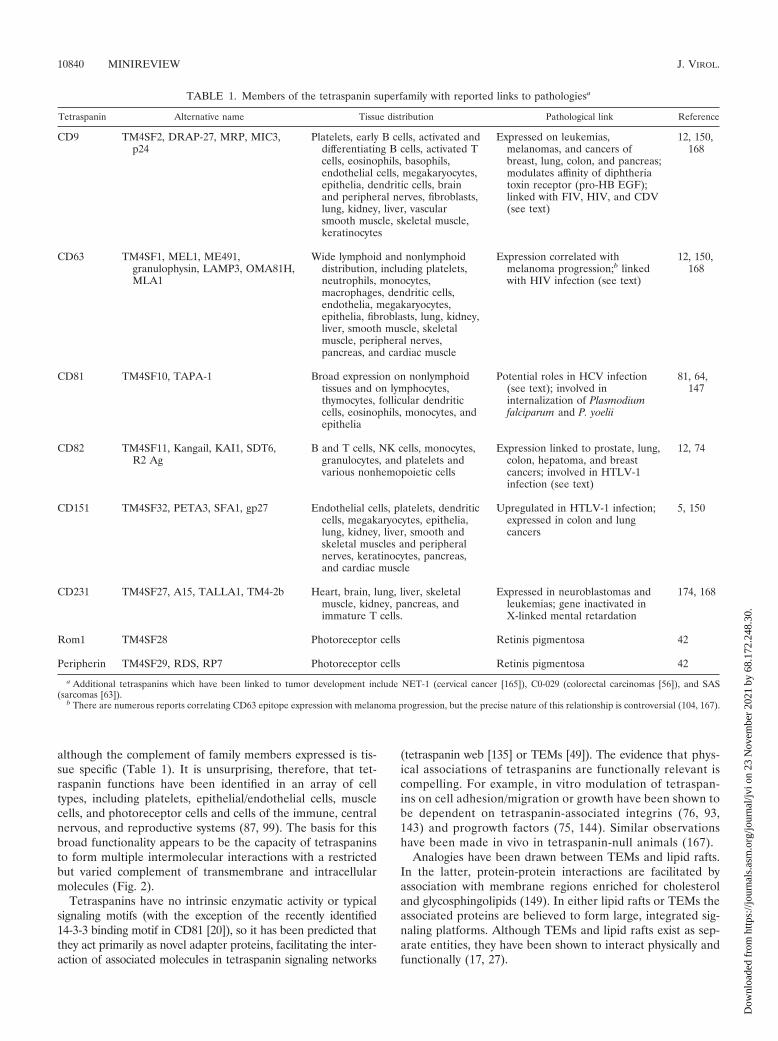
Structural features of the tetraspanin superfamily. Tet-raspanins are type III membrane glycoproteins which span theplasma membrane four times, producing two extracellularloops and short intracellular regions (Fig. 1). A defining struc-
tural signature of the superfamily is the “tetraspanin fold” ofthe larger extracellular loop (LEL). In this region, disulfidebonding of four absolutely conserved cysteines forms a subloopstructure containing a region which is hypervariable betweenfamily members and between species homologues of the sametetraspanin (Fig. 1) (81, 142). The cysteines are present inthree variously conserved motifs (CysCysGly, ProXSerCys[where X � any amino acid], and GluGlyCys) (Fig. 1), whereinthe flexibility and constraint imparted by the conserved Glyresidue in CysCysGly and by Pro in ProXSerCys contribute tosubloop formation (68). Many members of the tetraspaninfamily have one or two additional pairs of Cys residues in thetetraspanin fold region, potentially allowing the formation ofcomplex subloop structures. The region of the LEL outside ofthis subloop shows greater structural conservation, formingthree �-helices which not only form a structural platform topresent the tetraspanin fold but may also contribute indepen-dently to tetraspanin function (see below) (68, 152).
Conservation of the canonical LEL Cys, Gly, and otherresidues, including charged transmembrane amino-acids, dis-tinguishes members of the tetraspanin superfamily properfrom other “tetraspan” proteins (168). Modification of tet-raspanins includes O- and N-linked glycosylation. N glycosyl-ation sites are largely located in the LEL, and glycosylation hasbeen shown to regulate specific tetraspanin functions (111). Nglycosylation is generally assumed to occur at classical AsnXSer/Thr sites, but the LELs of some tetraspanins, includingCD9 and CD81, contain the rarer AsnXCys N-glycosylationmotif (138) which, interestingly, incorporates canonical cys-teines. Mutation of the only AsnXSer sites in CD9 does notprevent N glycosylation, indicating that the AsnXCys site maybe utilized in some cases (G. W. Moseley, L. J. Partridge, andP. N. Monk, unpublished data). Palmitoylation of tetraspaninshas also been shown to be functionally significant, as it appearsto participate in the formation of heterotetraspanin associa-tions (14) and to regulate association with lipid rafts (seebelow) (16).
Tetraspanin-enriched microdomains. Tetraspanins affectmultiple events in vitro, including cellular signaling, migration,adhesion, fusion, cytoskeletal reorganization, and prolifera-tion, which appear relevant to altered expression patterns seenduring cellular activation, differentiation, proliferation, andmalignant transformation in vivo (reviewed in references 87and 168). Tetraspanins are expressed by all mammalian tissues,
* Corresponding author. Mailing address: Nuclear Signalling Labo-ratory, Department of Biochemistry and Molecular Biology, MonashUniversity, Building 13D, Monash, Victoria 3080, Australia. Phone:61-3-99051220. Fax: 61-3-99053726. E-mail: [email protected].
10839
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.
although the complement of family members expressed is tis-sue specific (Table 1). It is unsurprising, therefore, that tet-raspanin functions have been identified in an array of celltypes, including platelets, epithelial/endothelial cells, musclecells, and photoreceptor cells and cells of the immune, centralnervous, and reproductive systems (87, 99). The basis for thisbroad functionality appears to be the capacity of tetraspaninsto form multiple intermolecular interactions with a restrictedbut varied complement of transmembrane and intracellularmolecules (Fig. 2).
Tetraspanins have no intrinsic enzymatic activity or typicalsignaling motifs (with the exception of the recently identified14-3-3 binding motif in CD81 [20]), so it has been predicted thatthey act primarily as novel adapter proteins, facilitating the inter-action of associated molecules in tetraspanin signaling networks
(tetraspanin web [135] or TEMs [49]). The evidence that phys-ical associations of tetraspanins are functionally relevant iscompelling. For example, in vitro modulation of tetraspan-ins on cell adhesion/migration or growth have been shown tobe dependent on tetraspanin-associated integrins (76, 93,143) and progrowth factors (75, 144). Similar observationshave been made in vivo in tetraspanin-null animals (167).
Analogies have been drawn between TEMs and lipid rafts.In the latter, protein-protein interactions are facilitated byassociation with membrane regions enriched for cholesteroland glycosphingolipids (149). In either lipid rafts or TEMs theassociated proteins are believed to form large, integrated sig-naling platforms. Although TEMs and lipid rafts exist as sep-arate entities, they have been shown to interact physically andfunctionally (17, 27).
TABLE 1. Members of the tetraspanin superfamily with reported links to pathologiesa
Tetraspanin Alternative name Tissue distribution Pathological link Reference
CD9 TM4SF2, DRAP-27, MRP, MIC3,p24
Platelets, early B cells, activated anddifferentiating B cells, activated Tcells, eosinophils, basophils,endothelial cells, megakaryocytes,epithelia, dendritic cells, brainand peripheral nerves, fibroblasts,lung, kidney, liver, vascularsmooth muscle, skeletal muscle,keratinocytes
Expressed on leukemias,melanomas, and cancers ofbreast, lung, colon, and pancreas;modulates affinity of diphtheriatoxin receptor (pro-HB EGF);linked with FIV, HIV, and CDV(see text)
12, 150,168
CD63 TM4SF1, MEL1, ME491,granulophysin, LAMP3, OMA81H,MLA1
Wide lymphoid and nonlymphoiddistribution, including platelets,neutrophils, monocytes,macrophages, dendritic cells,endothelia, megakaryocytes,epithelia, fibroblasts, lung, kidney,liver, smooth muscle, skeletalmuscle, peripheral nerves,pancreas, and cardiac muscle
Expression correlated withmelanoma progression;b linkedwith HIV infection (see text)
12, 150,168
CD81 TM4SF10, TAPA-1 Broad expression on nonlymphoidtissues and on lymphocytes,thymocytes, follicular dendriticcells, eosinophils, monocytes, andepithelia
Potential roles in HCV infection(see text); involved ininternalization of Plasmodiumfalciparum and P. yoelii
81, 64,147
CD82 TM4SF11, Kangail, KAI1, SDT6,R2 Ag
B and T cells, NK cells, monocytes,granulocytes, and platelets andvarious nonhemopoietic cells
Expression linked to prostate, lung,colon, hepatoma, and breastcancers; involved in HTLV-1infection (see text)
12, 74
CD151 TM4SF32, PETA3, SFA1, gp27 Endothelial cells, platelets, dendriticcells, megakaryocytes, epithelia,lung, kidney, liver, smooth andskeletal muscles and peripheralnerves, keratinocytes, pancreas,and cardiac muscle
Upregulated in HTLV-1 infection;expressed in colon and lungcancers
5, 150
CD231 TM4SF27, A15, TALLA1, TM4-2b Heart, brain, lung, liver, skeletalmuscle, kidney, pancreas, andimmature T cells.
Expressed in neuroblastomas andleukemias; gene inactivated inX-linked mental retardation
174, 168
Rom1 TM4SF28 Photoreceptor cells Retinis pigmentosa 42
Peripherin TM4SF29, RDS, RP7 Photoreceptor cells Retinis pigmentosa 42
a Additional tetraspanins which have been linked to tumor development include NET-1 (cervical cancer [165]), C0-029 (colorectal carcinomas [56]), and SAS(sarcomas [63]).
b There are numerous reports correlating CD63 epitope expression with melanoma progression, but the precise nature of this relationship is controversial (104, 167).
10840 MINIREVIEW J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.
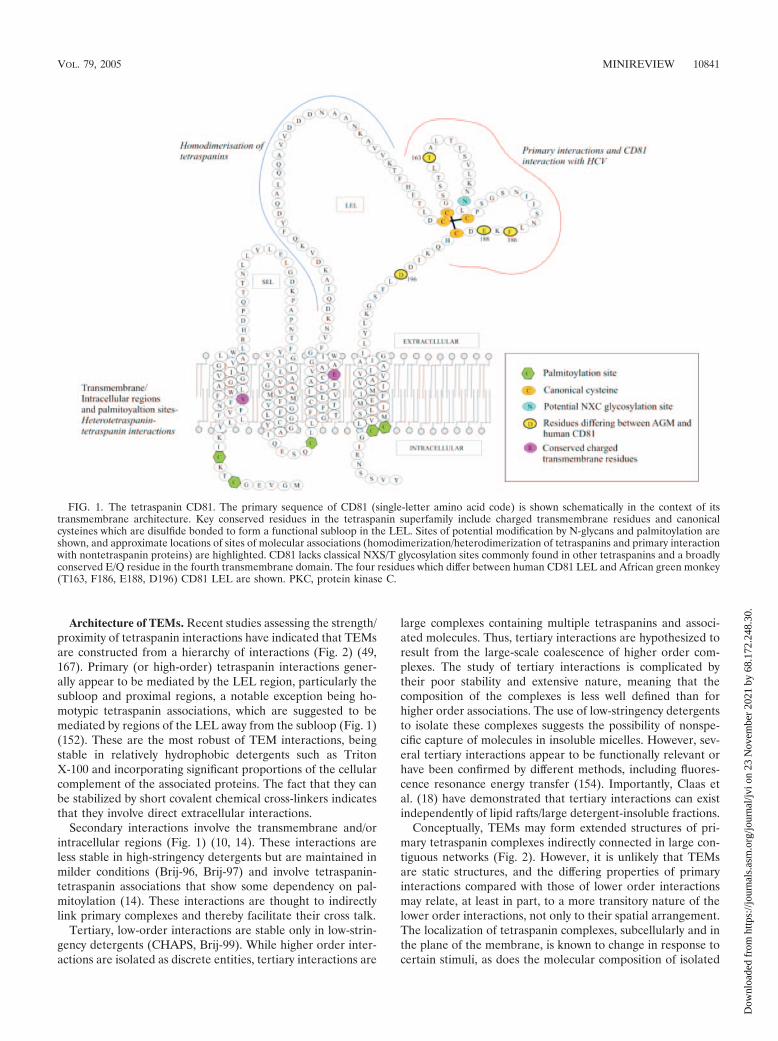
Architecture of TEMs. Recent studies assessing the strength/proximity of tetraspanin interactions have indicated that TEMsare constructed from a hierarchy of interactions (Fig. 2) (49,167). Primary (or high-order) tetraspanin interactions gener-ally appear to be mediated by the LEL region, particularly thesubloop and proximal regions, a notable exception being ho-motypic tetraspanin associations, which are suggested to bemediated by regions of the LEL away from the subloop (Fig. 1)(152). These are the most robust of TEM interactions, beingstable in relatively hydrophobic detergents such as TritonX-100 and incorporating significant proportions of the cellularcomplement of the associated proteins. The fact that they canbe stabilized by short covalent chemical cross-linkers indicatesthat they involve direct extracellular interactions.
Secondary interactions involve the transmembrane and/orintracellular regions (Fig. 1) (10, 14). These interactions areless stable in high-stringency detergents but are maintained inmilder conditions (Brij-96, Brij-97) and involve tetraspanin-tetraspanin associations that show some dependency on pal-mitoylation (14). These interactions are thought to indirectlylink primary complexes and thereby facilitate their cross talk.
Tertiary, low-order interactions are stable only in low-strin-gency detergents (CHAPS, Brij-99). While higher order inter-actions are isolated as discrete entities, tertiary interactions are
large complexes containing multiple tetraspanins and associ-ated molecules. Thus, tertiary interactions are hypothesized toresult from the large-scale coalescence of higher order com-plexes. The study of tertiary interactions is complicated bytheir poor stability and extensive nature, meaning that thecomposition of the complexes is less well defined than forhigher order associations. The use of low-stringency detergentsto isolate these complexes suggests the possibility of nonspe-cific capture of molecules in insoluble micelles. However, sev-eral tertiary interactions appear to be functionally relevant orhave been confirmed by different methods, including fluores-cence resonance energy transfer (154). Importantly, Claas etal. (18) have demonstrated that tertiary interactions can existindependently of lipid rafts/large detergent-insoluble fractions.
Conceptually, TEMs may form extended structures of pri-mary tetraspanin complexes indirectly connected in large con-tiguous networks (Fig. 2). However, it is unlikely that TEMsare static structures, and the differing properties of primaryinteractions compared with those of lower order interactionsmay relate, at least in part, to a more transitory nature of thelower order interactions, not only to their spatial arrangement.The localization of tetraspanin complexes, subcellularly and inthe plane of the membrane, is known to change in response tocertain stimuli, as does the molecular composition of isolated
FIG. 1. The tetraspanin CD81. The primary sequence of CD81 (single-letter amino acid code) is shown schematically in the context of itstransmembrane architecture. Key conserved residues in the tetraspanin superfamily include charged transmembrane residues and canonicalcysteines which are disulfide bonded to form a functional subloop in the LEL. Sites of potential modification by N-glycans and palmitoylation areshown, and approximate locations of sites of molecular associations (homodimerization/heterodimerization of tetraspanins and primary interactionwith nontetraspanin proteins) are highlighted. CD81 lacks classical NXS/T glycosylation sites commonly found in other tetraspanins and a broadlyconserved E/Q residue in the fourth transmembrane domain. The four residues which differ between human CD81 LEL and African green monkey(T163, F186, E188, D196) CD81 LEL are shown. PKC, protein kinase C.
VOL. 79, 2005 MINIREVIEW 10841
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.

complexes (59, 118). The dynamic subcellular localization oftetraspanins has been suggested to relate to roles in traffickingof web-associated molecules or as novel chaperone proteins inintegrin maturation (9, 33, 60, 89, 134, 173). Moreover, in Bcells the CD81/CD21/CD19 coreceptor complex interacts dy-namically with lipid rafts, modulated by palmitoylation ofCD81 (16), and regulated association of CD82, lipid rafts, andthe cytoskeleton has been reported (27). Cytoskeletal associa-tion of TEMs is likely to be involved in their subcellular/membrane distribution. Thus, TEMs, like lipid rafts, are likelyto be highly dynamic and regulable structures.
Tetraspanin function in vivo. A major feature of the tet-raspanin web hypothesis (135) was that it provided an expla-nation for the similar functional effects of different members ofthe superfamily observed in vitro. This hypothesis has receivedsupport from studies of tetraspanin-null mice, where similar
effects of knockouts of CD151, CD81, CD37, Tssc-6 have beenseen in T cells (167) and in the CD9-knockout mouse, whereCD81 expression in vitro can compensate for deficiencies insperm-egg fusion (65). Moreover, tetraspanin-knockout miceare largely healthy, generally displaying only mild phenotypicabnormalities. This is surprising, as quite dramatic in vivophenotypes result from mutation of the primary partners ofsome tetraspanins (69, 166). Thus, it appears that there is acertain degree of functional redundancy between tetraspanins.
While single-tetraspanin knockouts display mild phenotypes,phenotypic abnormalities are widespread and are found in Tcells, platelets, and keratinocytes of CD151-null mice (77, 166),in the central nervous system, B and T cells, and retinal pig-ment epithelial cells of CD81-null mice (39, 100, 151), and ingametes, smooth muscle cells, and the peripheral nervous sys-tem of CD9-null mice (58, 99, 140). It would appear likely,
FIG. 2. Possible architecture of the tetraspanin web. The principal interactions of TEMs are robust primary interactions which involve directextracellular tetraspanin-protein interactions. Secondary interactions are less robust and include heterotetraspanin interactions which facilitateindirect interaction of primary tetraspanin complex proteins. Tertiary interactions are the least-robust complexes and are thought to reflectlarge-scale coalescence of the higher order interactions into large signaling domains. Based on the known structure of the tetraspanin-richurothelial plaque, this order of interactions is shown as interlinked hexagons (98). Interactions within TEMs include homotypic and heterotypictetraspanin associations and association with adhesion molecules (integrins [9]), immune cell receptors (e.g., Fc receptors and T-cell receptor(TCR), CD4/CD8, and MHC proteins [103, 167]), growth factors and their receptors (pro-HB-EGF [136], pro-transforming growth factor � [144],and epidermal growth factor receptor [110]), intracellular signaling molecules (protein kinases hck, lyn, and PKC [167, 176] and phosphatidyl-inositol 4-kinase [172]), and cytoskeletal components (27), as well as members of a novel transmembrane immunoglobulin superfamily (EWI-2,EWI-F) (152).
10842 MINIREVIEW J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.

therefore, that TEMs per se have important roles in the phys-iology of most, if not all, cell types. The removal of a singletetraspanin shows some redundancy, owing to the extensivenature of TEMs and the expression of close homologues. How-ever, the clear phenotypic abnormalities observed indicate thatnot all functions of a single tetraspanin can be replicated bytheir counterparts (as is suggested by the evolutionary conser-vation of the members of this large family). The production ofknockout mice lacking multiple tetraspanins may lead to adelineation of the overlapping and unique functions of evolu-tionarily close members of the tetraspanin family.
As described above, studies of the structure of TEMs haverelied heavily on detergent solubility studies, which are clearlylimited and relatively insensitive. Similarly, with the data cur-rently available, the functional/physical relationship of tet-raspanin associations remains inferred, as it has not formallybeen correlated through mutagenesis of tetraspanins. The re-expression of mutated tetraspanins in null mice may thereforepermit more intricate and physiologically relevant investiga-tions of the structure and functional architecture of TEMs.
TETRASPANINS IN VIRAL INFECTIONS
Given the broad physiological importance of tetraspanins, itis not surprising that tetraspanins have been commonly iden-tified in cellular disorders (Table 1). There is increasing evi-dence for the involvement of tetraspanins in infections by var-ious microbes, but they have been most commonly identified asplaying a role in viral infections. The remainder of this reviewwill focus on these associations.
FLAVIVIRUSES: HCV AND CD81
Interaction of CD81 and HCV-E2. The most exhaustivelydocumented involvement of a tetraspanin in viral infection isof CD81 in HCV infection. HCV is a small, enveloped RNAvirus whose genome encodes a single polyprotein that is pro-cessed by viral/cellular proteases into structural and nonstruc-tural proteins. Structural proteins include the envelope glyco-proteins E1 and E2, which mediate viral binding and entry intohost cells (78).
CD81 was the first protein ligand identified for HCV, spe-cifically for E2 protein. The interaction was identified using acDNA library from a subclone of a human T-cell lymphomaline with high E2 binding capacity and was confirmed by thedemonstration that monoclonal antibodies to CD81-LEL caninhibit the binding of recombinant E2 to Epstein-Barr virus-transformed B cells (122) and that human CD81-LEL specif-ically binds E2 from HCV-infected plasma. The chimpanzee isthe only species other than humans that shows susceptibility toHCV (132), and vaccination with E2 glycoproteins can protectchimps from the virus. The physiological significance ofCD81/E2 association was implied by the demonstration thatthis protection correlated with the ability of sera to block theirinteraction (122).
The sequence in the LEL essential for binding E2 is con-tained within residues 164 to 201, a region that forms theepitope for antibody 1D6, which inhibits E2 binding to CD81(Fig. 1) (50). Inhibition of E2 binding by a small peptideanalogue of CD81 LEL has highlighted the role of residues 176
to 189 (30). African green monkey (AGM; Cercopithecus ae-thiops) CD81 differs from human CD81 LEL by only fourresidues (Fig. 1), but AGM-CD81 is unable to bind to E2 andAGMs are not susceptible to HCV. Reciprocal mutations ofhuman and AGM CD81 have shown that the key residue inthis interaction is CD81 Phe186 (50). The importance ofPhe186 was confirmed when tamarin monkey (Saguinus im-perator) CD81 was shown to be able to bind to E2 with a higheraffinity than human CD81 (1). Phe186 is conserved in tamarinCD81, and mutation of this residue can inhibit E2 binding (97).Mutational analysis has determined that the E2 binding site onCD81 also comprises residues Ile182, Asn184, and Leu162,forming a hydrophobic E2 binding surface of approximately806 Å2 (31). It has also been suggested that the SEL domain isimportant during HCV infection, although this appears to re-late primarily to its role in promoting surface expression ofCD81 by masking an intracellular retention signal (91).
E2 binds to cells of human origin but not to cell lines derivedfrom AGM, rat, and rabbit (36). Thus, the interaction appearsto be dependent on human (h)CD81 and not to require othermolecules in human cells, as rat cell lines can bind E2 proteinwhen transfected with hCD81. Binding to this particular tet-raspanin is also specific, as E2 does not bind to hCD9-, hCD63-or hCD151-transfected rat cells (36).
Based on this evidence, it has been suggested that CD81 maybe a cellular receptor for HCV. However, recent data indicatesthat CD81 is not sufficient for HCV entry, and the relevance ofthe CD81-HCV E2 association for viral entry remains unclear.One study has reported that binding of HCV to three hepato-cyte-derived cell lines was not blocked by anti-CD81 antibody(137). It has also been shown that hepatocarcinoma cells thathave lost the expression of CD81 mRNA still show a small butdetectable level of HCV binding (120). Tamarins are not sus-ceptible to HCV infection, but tamarin CD81 binds to E2 withgreat affinity (1), while hCD81 transgenic mice cannot be in-fected with HCV (91) and certain human liver cell lines arenonpermissive to infection by HCV-HIV pseudotypes despiteCD81 expression. Thus, it appears that formation of theCD81-E2 complex does not necessarily confer susceptibility tothe virus. It has also been observed that different HCV strainsutilize CD81 to different extents (70, 94, 131); E2 subtype 1b,for instance, shows a lower binding affinity to CD81-LEL thanE2 subtype 1a. The CD81-LEL Phe186Leu mutation has beenshown to abolish the binding of CD81 LEL to soluble E2 strainH but not to strain Con 1, even though this mutation has noeffect on the ability of transfected HepG2 cells to supportinfection by pseudotype viruses expressing either Con 1 orstrain H E2 (175). Another study using a more diverse range ofHCV glycoproteins revealed that the CD81-LEL Asp196Glumutation reduced the infectivity of HCV pseudotypes express-ing CH35, HJC4, and C6a1 glycoproteins by 70 to 96% (94).These data suggest that all HCV pseudotypes require CD81for infection of HepG2 cells but that the importance of theinteractions between CD81 LEL and HCV glycoproteins dif-fers between viral strains. The available evidence suggests thatCD81 represents an important component in HCV cellularreceptor activity but that this process is complex and likely toinvolve multiple receptor complexes. Further research will berequired to establish the in vivo relevance of these in vitroobservations.
VOL. 79, 2005 MINIREVIEW 10843
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.

CD81 in viral entry and release. There are other candidatereceptors for HCV, namely the scavenger receptor class B type1 (SR-B1) (139), dendritic cell-specific intracellular adhesionmolecule 3-grabbing nonintegrin (DC-SIGN) (84, 126), andlow-density lipoprotein receptor (LDLR) (170). One study hasestablished a requirement for both CD81 and SR-B1 for bind-ing and infectivity of HCV pseudoviruses, indicating a role forCD81 as a coreceptor in pseudotype infections (8). Similarly,pseudotype infections can be inhibited or enhanced by CD81silencing or expression, respectively (175), indicating a role forCD81 in viral entry. However, CD81 alone may be insufficientfor HCV entry, as the antibody-mediated internalization rateof CD81 is extremely low compared with that of other surfaceproteins such as CD4 and CD71 (120). Chimeras consisting ofCD81 and the cytoplasmic domains of two receptors that fre-quently undergo endocytosis (LDLR and the transferrin re-ceptor [TfR]) have been constructed (156); both chimeras sig-nificantly increase the endocytosis of recombinant HCV-E2protein, anti-CD81 antibody, and E2-enveloped viral particlesfrom the serum of HCV-infected patients. Expression of thechimeras in the Huh7 hepatoma cell line also resulted in in-creased viral replication, suggesting that additional factors arerequired for efficient endocytosis of wild-type CD81/virus com-plexes. Thus the mechanism of receptor-HCV complex inter-nalization is still poorly understood. Some lines of in vitroevidence indicate the involvement of receptor complexes in-corporating CD81, but further research will be required toidentify additional receptor components which contribute toefficient endocytosis
As discussed above, TEMs may have a general role in reg-ulating the trafficking of proteins associated with tetraspanins.There is precedent of a pathogenic exploitation of these pro-cesses: CD81 has been linked to entry of plasmodium intohepatocytes in the context of a parasitophorus vacuole (147).Thus, it is possible that viral proteins which interact with tet-raspanins become incorporated into TEMs, facilitating theirtransport into or out of the cell. In the absence of humanCD81, mammalian cells transfected with HCV E1-E2 cDNAalmost completely retain these proteins in the endoplasmicreticulum. However, when cotransfected with hCD81, a smallfraction of these proteins pass through the Golgi and are foundassociated with extracellular exosomes (92). The CD81Asp196Glu mutation decreases the susceptibility to infection(see above), despite having only a minimal effect on binding ofE2. It is possible that this mutation decreases infectivity byaltering the intracellular trafficking characteristics of CD81(50), lending further support to the notion that CD81-E2 in-teractions are involved in facilitating trafficking of HCV par-ticles within the cell. Thus, CD81 may play a role in facilitatingviral glycoprotein translocation into exosomes during viral exitand not exclusively in cell entry.
The possible involvement of CD81 in facilitating vacuoletrafficking/fusion/release is consistent with hypothesized rolesfor other tetraspanins in physiological processes (167, 169) andsimilar to events described for the role of CD63 in HIV-1infection (see below). In vitro studies showing that anti-CD81antibody delayed fusion of a mouse myogenic cell line supporta role for CD81 in membrane fusion events (155). Anti-CD9antibody induced the same effect, suggesting that TEMs con-taining different tetraspanins may be involved in this event.
Further evidence for TEM activity includes the demonstrationthat antibodies to CD9 or CD81 delayed syncytia formation ina rhabdomyosarcoma cell line (155). CD9 is also involved insperm-egg fusion (15, 79, 99), and significantly, the impairedfusion seen in CD9-null animals can be compensated in vitroby CD81 expression (65). Also, recent data suggest that solubleLEL domains of both CD9 and CD63 can inhibit concanavalinA-induced fusion of human blood monocytes, suggesting that arange of tetraspanins may be involved in the mechanism of cellfusion (V. Parthasarathy, G. W. Moseley, A. Higginbottom,P. N. Monk, R. C. Read, and L. J. Partridge, unpublisheddata). Therefore, TEMs containing multiple tetraspanins areinvolved in membrane fusion events which are likely to relateto roles in viral infection.
Potential roles of CD81 and E2 in modulation of the im-mune response. Although HCV-infected patients show bothcellular and humoral immune responses, over 85% are unableto eliminate the virus. In common with other leukocyte-ex-pressed tetraspanins, CD81 has roles in modulating immunecell function (17, 28, 86). An intriguing possibility is that theinteraction of CD81 with E2 could modify immune responsesin HCV-infected individuals. It has been shown that cross-linking of natural killer (NK) CD81 with immobilized E2 in-hibits NK cell response to interleukins-2, -12 and -15, whichmay affect early innate immune response to HCV infections(21, 159), while E2 cross-linking of CD81 on B- and T-cell linesalters proliferation and aggregation (36). E2 engagement ofCD81 also causes the secretion of the chemokine RANTES byCD8� T cells and the subsequent down-regulation of its re-ceptor, CCR5, a mechanism by which HCV might interferewith lymphocyte migration (109). These events may providestrategies for immune evasion or, alternatively, may relate tothe immune dysregulation effects of chronic HCV infections,including type 2 diabetes mellitus, glomerulonephritis, arthri-tis, and non-Hodgkin’s lymphoma (157). As CD81 is widelyexpressed and involved in an array of functions, there are anumber of potential biological effects of E2-CD81 interaction.Flint et al. (36) hypothesized that interactions between E2 oninfected hepatocytes and CD81 on uninfected cells couldprime the latter for infection. These are certainly intriguingpossibilities, and the genuine physiological significance of thebiological effects of E2 interactions with CD81 awaits furtherresearch.
RETROVIRUSES: HTLV-1, HIV, AND FIV
HTLV-1. HTLV-1 is a type C retrovirus that infects CD4�
lymphocytes in vivo (130), although CD8 T cells may also serveas a reservoir (107). This tropism contrasts with the in vitroability of viral Env protein to bind and enter multiple cell types(13, 45, 67, 153, 158). The receptor for HTLV-1 is unknown,but a likely candidate is the human GLUT1 glucose trans-porter (88). Molecules involved in syncytium formation, virusadsorption, and infection include ICAM-1, ICAM-3,VCAM-1, and tetraspanins (22, 52, 123, 124). Cell-cell fusionvia syncytium formation is thought to be one of the maintransmission mechanisms of HTLV-1 (19). The tetraspaninCD82 was identified as an antigen recognized by several anti-bodies found to inhibit HTLV-1-mediated syncytium forma-tion (38, 57). Coexpression of CD82 with HTLV-1 Env glyco-
10844 MINIREVIEW J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.

protein in COS-1 cells had an inhibitory effect on syncytiumformation and HTLV-1 infectivity, although the incorporationof Env into nascent virions was unaffected (124).
CD82 is expressed on the surfaces of most types of periph-eral blood mononuclear cells (PBMCs) and granulocytes, al-though CD16� NK cells show very low levels of expression andCD4� T cells express higher levels than CD8� T cells (57).CD82 is heterogeneously glycosylated in T cells, with apparentmolecular mass ranging from 35 to 50 kDa (57). Activation ofT cells (e.g., using phytohemagglutinin) and/or infection withHTLV-1 results in up-regulation of CD82 expression and alsoincreases the amount of glycosylation (38, 57). The glycosyla-tion state of CD82 has been shown to be essential for itscorrect functioning (111, 112), possibly by altering the affinityof CD82 for membrane proteins such as integrins. Coimmu-noprecipitation experiments show that HTLV-1 Env glycopro-teins interact with highly glycosylated forms of CD82, althoughinteractions with immature forms of the tetraspanin suggestthat these interactions occur early in the secretory pathway(124). The tetraspanin CD151 has also been shown to be up-regulated in HTLV-1-infected cells (47, 48), and this has beenlinked to increased adhesion of infected cells to the extracel-lular matrix. CD82 also appears to be involved in cellularadhesion and motility events (112), and it is possible thatCD151 and CD82 have common functions during HTLV-1infection. CD82 has recently been proposed to act as an adap-tor protein, linking lipid rafts and the actin cytoskeleton in Tcells (26). In this role, it is likely to control the function ofmolecules such as ICAM-1 and �1 integrins that are directlyinvolved in the cell-cell interactions that lead to syncytiumformation (22). Thus, although CD82 does not act as a cellularreceptor for HTLV-1 or affect expression levels of HTLV-1proteins, it is likely to function in concert with CD151 andother tetraspanins in TEM-dependent cell fusion events.
HIV-1. Entry of the retrovirus HIV (genus, Lentivirus) intocells occurs in three distinct steps: the attachment of virions tocell surface CD4� by envelope gp120 protein, the subsequentinterplay of the conformationally altered gp120 with cellularCCR5 and CXR4 coreceptors, and, finally, envelope gp41 pro-tein-mediated fusion to target cells (102). The HIV genomeconsists of two copies of RNA which are reverse transcribed toDNA to form the preintegration complex containing the viralmatrix (MA), integrase, and Vpr proteins as well as severalhost proteins, which is imported into the nucleus and incorpo-rated into the host chromosomes, where it can become latentfor 3 to 10 years (44). The subtypes of HIV, HIV-1 and HIV-2,have the same modes of transmission and are associated withsimilar opportunistic infections, but immunodeficiency seemsto develop more slowly with HIV-2 (90, 148).
A role for the tetraspanin CD63 in HIV-1 infection was firstsuggested when it was shown to be up-regulated from intracel-lular vesicles to the surfaces of HIV-1-infected cells and selec-tively incorporated into budding structures and newly synthe-sized virus particles (95, 96). Consistent with a role in viralrelease, vesicle fractions in infected H9 T cells contain in-creased levels of CD63 compared with noninfected cells (41).
Macrophages are sources of virus reservoirs during HIV-1infection: previous work has described organelles within mac-rophages as preferential sites for virus accumulation in mac-rophages (114). Expression analysis revealed high levels of
CD63 and major histocompatibility complex type II (MHC II)and low levels of Lamp 1 in these organelles, resembling thecompartments where MHC II molecules undergo the finalstages of maturation. CD63 is also enriched on the surface ofmultivesicular bodies, and it is presumed that HIV-1 particlesmimic the mechanisms utilized in the production of naturalinternal vesicles (128). Thus it is possible that CD63, like CD81and CD82 (see above), may be involved in viral traffickingand/or association with intracellular vesicles for fusion andrelease.
It has recently been shown that HIV-1 infection is inhibitedby anti-CD63 antibodies, but not by antibodies to CD9 orCD81. This was specific for macrophages and HIV-1 strainswhich use the CCR5 coreceptor, as X4 virus and T-cell infec-tion were unaffected by anti-CD63 antibodies (161). Prelimi-nary data also shows that recombinant tetraspanin-LEL pro-teins can also specifically inhibit macrophage infection but, incontrast to antibody inhibitors, can also inhibit CXCR4-tropicvirus; significantly, the effect is seen with a range of tet-raspanin-LELs (including CD9 and CD81) (S. H. Ho, F. Mar-tin, G. W. Moseley, L. J. Partridge, C. Cheng-Mayer, and P. N.Monk, unpublished data). These experiments indicate a rolefor multiple tetraspanins, probably organized in TEMs, in theinfection of macrophages by HIV. The differences observedbetween the studies using tetraspanin LELs and antibodiesmay be due to a number of factors, as neither approach isphysiological. However, tetraspanin LELs are small proteinsand are not prone to the same artifacts as antibodies such as Fcreceptor coengagement, known to suppress HIV replication inmacrophages (119). Alternatively, the differing efficacy of an-tibodies may relate to epitope-specific events, as described foranti-CD82 in syncytium formation (57). It is also possible thatanti-CD63 antibodies bind directly to the viral envelope,known to contain higher levels of CD63 than other tetraspan-ins, thus inhibiting infection by steric hindrance (117). How-ever, the apparent involvement of CD9 and CD81 in HIV-1infection implies that TEMs are involved in HIV-1 infectionand that this role may involve vacuolar targeting and mem-brane fusion events in viral release.
FIV. Feline immunodeficiency virus (FIV) infection also ap-pears to involve tetraspanins, in this case CD9 (164). In 104-C1PBMC cells transfected to express feline (f)CD9, the anti-CD9antibody vpg15 was shown to delay reversibly, but not com-pletely block, reverse transcriptase activity following infection(29). As the inhibitory effect was enhanced upon addition ofantibody postinfection, it appeared that CD9 can affect eventsafter cellular entry. Treatment of chronically infected cells withvpg15 reduced the numbers of budding particles at the plasmamembrane and inhibited FIV spread in cell culture, so it ap-pears likely that viral assembly and/or release (29) is inhibitedby anti-CD9 antibody, indicating a role for fCD9 similar to thatof other tetraspanins in viral infections. FIV can infect CD9-negative 3201 cells (55), implying that CD9 is not essential forFIV infection or at least that other tetraspanins can take overthis role. However, ectopic expression of CD9 in 3201 cellsenhances FIV infectivity, suggesting that CD9 plays a directrole in infection rather than an indirect role via cell signalingpathways affected by anti-CD9 antibody ligation (163).
VOL. 79, 2005 MINIREVIEW 10845
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.

PARAMYXOVIRUS
Canine distemper virus (CDV) (a morbillivirus of theParamyxoviridae family) is a negative-stranded RNA virus, sim-ilar to human measles virus, for which the host cell surfacereceptor(s) are currently not identified (46). As for FIV, ananti-CD9 antibody has been shown to inhibit viral infection(83) and transfection of CD9 into cell lines increases viralproduction, leading to a greater number of infectious centersand larger syncytia, consistent with the effects of CD9 expres-sion on syncytium formation by a rhabdomyosarcoma cell line(155). CD9 does not appear to be a receptor for CDV (83), andanti-CD9 antibody does not block CDV attachment to cells(141). The involvement in subsequent events (83), includingmembrane fusion/syncytium formation, is comparable to thatof other tetraspanins in human viral diseases and CD9 in FIV.
TETRASPANINS AS POTENTIAL THERAPEUTICTARGETS FOR VIRUSES THAT INFECT HUMANS
HCV infects 170 million people worldwide, causing liverdiseases that include hepatitis, cirrhosis, hepatocellular carci-noma, and disorders relating to immune dysregulation (125,157). HIV/AIDS is responsible for more than 25 million deaths
since 1984, with an estimated 37.8 million individuals currentlyinfected (Joint United Nations Program on HIV/AIDS, UN-AIDS, 2004). HTLV-1 presently infects 20 million individualsworldwide (32) and is associated with adult T-cell-leukemia/lymphoma and myelopathy/tropical spatic paraparesis. Treat-ments for these viruses include antiviral drugs such as inter-ferons, viral enzyme inhibitors, and viral-cell receptorinteraction inhibitors (24, 25, 35, 62, 71, 100, 157), which cansubstantially lower the viral load (115) but cannot eliminate thevirus. Further problems associated with such treatments in-clude toxicity and side effects, high cost, development of viralresistance, and low efficacy owing to viral genotype specificity/patient variables (11, 43, 61, 101, 108, 121, 145, 157, 171).Efforts to develop conventional vaccines for HIV have beenhampered by potential pathogenicity and poor capacity to elicitprotective immune responses (6, 54, 80, 162). Several new vacci-nation approaches for HIV (e.g., DNA and viral vectors) are inclinical trials (3, 4, 7, 23, 82, 105, 133, 146). However, in commonwith conventional vaccines, most of these strategies have beenshown to be ineffective due either to their poor capacity to elicitimmune protection (40, 85) or to the various mechanisms of HIVimmune evasion, such as high rate of mutations and host genomeinsertion (34, 53, 72, 73, 106, 116, 127).
FIG. 3. Model for the role of TEMs in viral infection. Current evidence (represented in inset) indicates that viral proteins may associate withTEMs by forming direct primary interactions with a tetraspanin (black arrows) which, via secondary/tertiary interactions of tetraspanins (greyarrow), incorporate the viral protein into the tetraspanin web, permitting indirect interaction with tetraspanin-associated molecules (dotted arrow).Various data indicate that, through these interactions, TEMs may be involved in initial viral attachment (no. 1) and entry (no. 2) into the cell. Somedata also indicate that viral interaction with TEMs may couple to intracellular signaling pathways, priming cells for infection or altering immunecell functions (no. 3). TEMs may interact with new viral proteins after infection/transcription in the biosynthesis/secretion pathway (no. 4). Multiplelines of evidence also indicate that TEM involvement in membrane fusion events may be exploited for viral exit from cells whether via direct releaseby secretory vesicles or in exosomes (no. 5 and 6) or via cell-cell fusion to form syncytia (no. 7). In the latter, tetraspanin-associated cell adhesionmolecules (e.g., ICAM, integrins) are likely to mediate intercellular interactions in the fusion process.
10846 MINIREVIEW J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.

As described above, various groups have identified tet-raspanins as the targets of antibodies which inhibit the infec-tivity of a range of viruses but have generally identified aspecific tetraspanin in each of these studies. However, it ap-pears that at least two tetraspanins may be involved in HTLV-1infection and at least four in HIV-1 infection. Thus, we hy-pothesize that (i) several tetraspanins will be involved in someor all of the viral life cycles here mentioned, and (ii) thattetraspanins are likely to be involved in many other viral in-fections as yet undiscovered. While definition of the preciseroles of TEMs in viral infection awaits further research, it hasalready been shown that peptides based on tetraspanin-LELscan inhibit tetraspanin functions such as sperm/oocyte fusion(51). A peptide inhibitor of only 14 residues, designed to mimicthe region 176 to 189 of CD81 has been shown to inhibit theCD81-E2 interaction (30). With a greater understanding of theassociation of TEMs with viral proteins, particularly the pri-mary ligand interactions, it should become possible to selec-tively target TEMs for therapy. The E2 binding site on CD81has been mapped to specific amino acids in the LEL subloop(31, 160). Based on these findings, several bi-imidazole-basedcompounds that can also inhibit E2 binding to CD81 have beenidentified (160). These resemble the solvent-exposed face ofthe helix of CD81 LEL that contains Phe186, thus mimickingthe E2 binding site on CD81. Two of the compounds have 50%inhibitory concentration values below 100 �M and representan important proof of the concept that tetraspanin-based drugscan disrupt biological function. Thus, targeting TEMs mayprovide a novel strategy to inhibit critical processes of viralinfection.
CONCLUDING REMARKS
The precise nature of the relationship of tetraspanins withviruses is presently poorly defined and is complicated by theapparently disparate functions of specific tetraspanins in infec-tions by different viruses (summarized in Fig. 3). However,much of the data describing this relationship has been derivedfrom in vitro studies. Thus, it is unclear whether modulation ofimmune cell line function by LEL-CD81 ligation, for example,has any genuine significance in vivo. Similarly, while CD81 isknown to interact with HCV coat proteins in vitro, the stage ofinfection at which this interaction occurs in vivo is unknownand need not necessarily occur during cell entry. Indeed theinteraction of CD82 with HTLV-1 proteins occurs postinfec-tion, while CD9 and CD63 appear to affect FIV/CDV andHIV-1 late in infection. Of particular significance is the recentdemonstration that soluble LEL proteins from several differenttetraspanins can inhibit HIV-1 infection and the overlappingeffects of certain antitetraspanin antibodies in syncytium for-mation by rhabdomyosarcoma cells. This suggests that there isfunctional overlap between different tetraspanins involved inviral infection processes, which implies the involvement ofTEMs.
Thus, rather than being passive targets for opportunisticviral attachment to cells, tetraspanins are likely to play a morefundamental role in viral infectivity through the involvement ofTEMs in viral protein maturation/trafficking, in cell-cell fusionevents or in other membrane fusion events involved in viralrelease, processes which have clear parallels in normal TEM
physiology. In this context, the selective primary interaction ofviral protein with a specific tetraspanin (e.g., CD81 with HCVE2) to connect it via secondary/tertiary interactions to tet-raspanin complexes involved in these processes would be typ-ical of the incorporation of nontetraspanin proteins into theTEMs (Fig. 3). Targeting TEMs for viral therapies may there-fore target a principal mechanism in the infectious cycles ofHIV-1, HTLV-1, and HCV which viral mutation would notreadily circumvent, providing new antiviral therapies so ur-gently required by an ever-increasing percentage of the world’spopulation.
REFERENCES
1. Allander, T., X. Forns, S. U. Emerson, R. H. Purcell, and J. Bukh. 2000.Hepatitis C virus envelope protein E2 binds to CD81 of tamarins. Virology277:358–367.
2. Reference deleted.3. Amara, R. R., F. Villinger, J. D. Altman, S. L. Lydy, S. P. O’Neil, S. I.
Staprans, D. C. Montefiori, Y. Xu, J. G. Herndon, L. S. Wyatt, M. A.Candido, N. L. Kozyr, P. L. Earl, J. M. Smith, H. L. Ma, B. D. Grimm, M. L.Hulsey, J. Miller, H. M. McClure, J. M. McNicholl, B. Moss, and H. L.Robinson. 2001. Control of a mucosal challenge and prevention of AIDS bya multiprotein DNA/MVA vaccine. Science 292:69–74.
4. Andino, R., D. Silvera, S. D. Suggett, P. L. Achacoso, C. J. Miller, D.Baltimore, and M. B. Feinberg. 1994. Engineering poliovirus as a vaccinevector for the expression of diverse antigens. Science 265:1448–1451.
5. Ashman, L., S. Fitter, P. M. Sincock, L. Nguyen, and A. C. Cambareri. 1997.CD151 (PETA-3) workshop summary report, p. 681–683. In T. Kishimotoet al. (ed.), Leukocyte typing VI. Garland Publishing, New York, N.Y.
6. Baba, T. W., Y. S. Jeong, D. Pennick, R. Bronson, M. F. Greene, and R. M.Ruprecht. 1995. Pathogenicity of live, attenuated SIV after mucosal infec-tion of neonatal macaques. Science 267:1820–1825.
7. Barouch, D. H., S. Santra, M. J. Kuroda, J. E. Schmitz, R. Plishka, A.Buckler-White, A. E. Gaitan, R. Zin, J. H. Nam, L. S. Wyatt, M. A. Lifton,C. E. Nickerson, B. Moss, D. C. Montefiori, V. M. Hirsch, and N. L. Letvin.2001. Reduction of simian-human immunodeficiency virus 89.6P viremia inrhesus monkeys by recombinant modified vaccinia virus Ankara vaccina-tion. J. Virol. 75:5151–5158.
8. Bartosch, B., A. Vitelli, C. Granier, C. Goujon, J. Dubuisson, S. Pascale, E.Scarselli, R. Cortese, A. Nicosia, and F. L. Cosset. 2003. Cell entry ofhepatitis C virus requires a set of co-receptors that include the CD81tetraspanin and the SR-B1 scavenger receptor. J. Biol. Chem. 278:41624–41630.
9. Berditchevski, F. 2001. Complexes of tetraspanins with integrins: more thanmeets the eye. J. Cell Sci. 114:4143–4151.
10. Berditchevski, F., E. Gilbert, M. R. Griffiths, S. Fitter, L. Ashman, and S. J.Jenner. 2001. Analysis of the CD151-�3�1 integrin and CD151-tetraspanininteractions by mutagenesis. J. Biol. Chem. 276:41165–41174.
11. Besson, C., Y. Plumelle, B. Arnulf, C. Gonin, G. Panelatti, A. Bazarbachi,and O. Hermine. 2001. Adult T-cell leukemia/lymphoma. Therapeutic as-pects. Presse Med. 30:243–245. (In French.)
12. Boucheix, C., G. H. Duc, C. Jasmin, and E. Rubinstein. 2001. Tetraspaninsand malignancy. Expert Rev. Mol. Med. 31:1–17.
13. Cavrois, M., A. Gessain, S. Wain-Hobson, and E. Wattel. 1996. Prolifera-tion of HTLV-1 infected circulating cells in vivo in all asymptomatic carriersand patients with TSP/HAM. Oncogene 12:2419–2423.
14. Charrin, S., S. Manie, M. Oualid, M. Billard, C. Boucheix, and E. Rubin-stein. 2002. Differential stability of tetraspanin/tetraspanin interactions:role of palmitoylation. FEBS Lett. 516:139–144.
15. Chen, M. S., K. S. Tung, S. A. Coonrod, Y. Takahashi, D. Bigler, A. Chang,Y. Yamashita, P. W. Kincade, J. C. Herr, and J. M. White. 1999. Role of theintegrin-associated protein CD9 in binding between sperm ADAM 2 andthe egg integrin �6�1: implications for murine fertilization. Proc. Natl.Acad. Sci. USA 96:11830–11835.
16. Cherukuri, A., R. H. Carter, S. Brooks, W. Bornmann, R. Finn, C. S. Dowd,and S. K. Pierce. 2004. B cell signaling is regulated by induced palmitoyl-ation of CD81. J. Biol. Chem. 279:31973–31982.
17. Cherukuri, A., T. Shoham, H. W. Sohn, S. Levy, S. Brooks, R. Carter, andS. K. Pierce. 2004. The tetraspanin CD81 is necessary for partitioning ofcoligated CD19/CD21-B cell antigen receptor complexes into signaling-active lipid rafts. J. Immunol. 172:270–280.
18. Claas, C., C. S. Stipp, and M. E. Hemler. 2001. Evaluation of prototypetransmembrane 4 superfamily protein complexes and their relation to lipidrafts. J. Biol. Chem. 276:7974–7984.
19. Clapham, P., K. Nagy, R. Cheingsong-Popov, M. Exley, and R. A. Weiss.1983. Productive infection and cell-free transmission of human T-cell leu-kemia virus in a nonlymphoid cell line. Science 222:1125–1127.
20. Clark, K., A. Oelke, M. E. Johnson, K. D. Eilert, P. C. Simpson, and S. C.
VOL. 79, 2005 MINIREVIEW 10847
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.

Todd. 2004. CD81 associates with 14-3-3 in a redox-regulated palmitoyla-tion-dependent manner. J. Biol. Chem. 279:19401–19406.
21. Crotta, S., A. Stilla, A. Wack, A. D’Andrea, S. Nuti, U. D’Oro, M. Mosca, F.Filliponi, R. M. Brunetto, F. Bonino, S. Abrignani, and N. M. Valiante.2002. Inhibition of natural killer cells through engagement of CD81 by themajor hepatitis C virus envelope protein. J. Exp. Med. 195:35–41.
22. Daenke, S., S. A. McCracken, and S. Booth. 1999. Human T-cell leukaemia/lymphoma virus type 1 syncytium formation is regulated in a cell-specificmanner by ICAM-1, ICAM-3 and VCAM-1 and can be inhibited by anti-bodies to integrin beta2 or beta7. J. Gen. Virol. 80:1429–1436.
23. Davis, N. L., I. J. Caley, K. W. Brown, M. R. Betts, D. M. Irlbeck, K. M.McGrath, M. J. Connell, D. C. Montefiori, J. A. Frelinger, R. Swanstrom,P. R. Johnson, and R. E. Johnston. 2000. Vaccination of macaques againstpathogenic simian immunodeficiency virus with Venezuelan equine en-cephalitis virus replicon particles. J. Virol. 74:371–378.
24. Dean, M., M. Carrington, C. Winkler, G. A. Huttley, M. W. Smith, R.Allikmets, J. J. Goedert, S. P. Buchbinder, E. Vittinghoff, E. Gomperts, S.Donfield, D. Vlahov, R. Kaslow, A. Saah, C. Rinaldo, R. Detels, and S. J.O’Brien. 1996. Genetic restriction of HIV-1 infection and progression toAIDS by a deletion allele of the CKR5 structural gene. Hemophilia Growthand Development Study, Multicenter AIDS Cohort Study, MulticenterHemophilia Cohort Study, San Francisco City Cohort, ALIVE Study. Sci-ence 273:1856–1862.
25. De Clercq, E. 2004. Antiviral drugs in current clinical use. J. Clin. Virol.30:115–133.
26. Delaguillaumie, A., J. Harriague, S. Kohanna, G. Bismuth, E. Rubinstein,M. Seigneuret, and H. Conjeaud. 2004. Tetraspanin CD82 controls theassociation of cholesterol-dependent microdomains with the actin cytoskel-eton in T lymphocytes: relevance to co-stimulation. J. Cell Sci. 117:5269–5282.
27. Delaguillaumie, A., Harriague, J., Kohanna, S., Bismuth, G., Rubinstein,E., Seigneuret, M., Conjeaud, H. 2004. Tetraspanin CD82 controls theassociation of cholesterol-dependent microdomains with the actin cytoskel-eton in T lymphocytes: relevance to co-stimulation. J. Cell Sci. 117:5269–5282.
28. Deng, J., R. H. Dekruyff, G. J. Freeman, D. T. Umetsu, and S. Levy. 2002.Critical role of CD81 in cognate T-B cell interactions leading to Th2responses. Int. Immunol. 14:513–523.
29. de Parseval, A., D. L. Lerner, P. Borrow, B. J. Willett, and J. H. Elder. 1997.Blocking of feline immunodeficiency virus infection by a monoclonal anti-body to CD9 is via inhibition of virus release rather than interference withreceptor binding. J. Virol. 71:5742–5749.
30. Dhanasekaran, M., P. W. Baures, S. VanCompernolle, S. Todd, and O.Prakash. 2003. Structural characterization of peptide fragments fromhCD81-LEL. J. Pept. Res. 61:80–89.
31. Drummer, H. E., K. A. Wilson, and P. Poumbourios. 2002. Identification ofthe hepatitis C virus E2 glycoprotein binding site on the large extracellularloop of CD81. J. Virol. 76:11143–11147.
32. Edlich, R. F., J. A. Arnette, and F. M. Williams. 2000. Global epidemic ofhuman T-cell lymphotropic virus type-I (HTLV-I). J. Emerg. Med. 18:109–119.
33. Engering, A., and J. Pieters. 2001. Association of distinct tetraspanins withMHC class II molecules at different subcellular locations in human imma-ture dendritic cells. Int. Immunol. 13:1713–1723.
34. Finzi, D., and R. F. Siliciano. 1998. Viral dynamics in HIV-1 infection. Cell93:665–671.
35. Flexner, C. 1998. HIV-protease inhibitors. N. Engl. J. Med. 338:1281–1292.36. Flint, M., C. Maidens, L. D. Loomis-Price, C. Shotton, J. Dubuisson, P.
Monk, A. Higginbottom, S. Levy, and J. A. McKeating. 1999. Character-ization of hepatitis C virus E2 glycoprotein interaction with a putativecellular receptor, CD81. J. Virol. 73:6235–6244.
37. Reference deleted.38. Fukudome, K., M. Furuse, T. Imai, M. Nishimura, S. Takagi, Y. Hinuma,
and O. Yoshie. 1992. Identification of membrane antigen C33 recognized bymonoclonal antibodies inhibitory to human T-cell leukemia virus type 1(HTLV-1)-induced syncytium formation: altered glycosylation of C33 an-tigen in HTLV-1-positive T cells. J. Virol. 66:1394–1401.
39. Geisert, E. J., R. W. Williams, G. R. Geisert, L. Fan, A. M. Asbury, H. T.Maecker, J. Deng, and S. Levy. 2002. Increased brain size and glial cellnumber in CD81-null mice. J. Comp. Neurol. 453:22–32.
40. Giri, M., K. E. Ugen, and D. B. Weiner. 2004. DNA vaccines against humanimmunodeficiency virus type 1 in the past decade. Clin. Microbiol. Rev.17:370–389.
41. Gluschankof, P., I. Mondor, H. R. Gelderblom, and Q. J. Sattentau. 1997.Cell membrane vesicles are a major contaminant of gradient-enriched hu-man immunodeficiency virus type-1 preparations. Virology 230:125–133.
42. Goldberg, A., and R. S. Molday. 1996. Defective subunit assembly underliesa digenic form of retinitis pigmentosa linked to mutations in peripherin/rdsand rom-1. Proc. Natl. Acad. Sci. USA 26:13726–13730.
43. Gout, O., A. Gessain, M. Iba-Zizen, S. Kouzan, F. Bolgert, G. de The, andO. Lyon-Caen. 1991. The effect of zidovudine on chronic myelopathy asso-ciated with HTLV-1. J. Neurol. 238:108–109.
44. Greene, W. C. 2004. The brightening future of HIV therapeutics. Nat.Immunol. 5:867–871.
45. Hanon, E., J. C. Stinchcombe, M. Saito, B. E. Asquith, G. P. Taylor, Y.Tanaka, J. N. Weber, G. M. Griffiths, and C. R. Bangham. 2000. Fratricideamong CD8� T lymphocytes naturally infected with human T cell lympho-tropic virus type I. Immunity 13:657–664.
46. Harder, T. C., and A. D. Osterhaus. 1997. Canine distemper virus[em]amorbillivirus in search of new hosts? Trends Microbiol. 5:120–124.
47. Hasegawa, H., T. Nomura, K. Kishimoto, K. Yanagisawa, and S. Fujita.1998. SFA-1/PETA-3 (CD151), a member of the transmembrane 4 super-family, associates preferentially with �5�1 integrin and regulates adhesionof human T cell leukemia virus type 1-infected T cells to fibronectin.J. Immunol. 161:3087–3095.
48. Hasegawa, H., Y. Utsunomiya, K. Kishimoto, K. Yanagisawa, and S. Fujita.1996. SFA-1, a novel cellular gene induced by human T-cell leukemia virustype 1, is a member of the transmembrane 4 superfamily. J. Virol. 70:3258–3263.
49. Hemler, M. 2003. Tetraspanin proteins mediate cellular penetration, inva-sion, and fusion events and define a novel type of membrane microdomain.Annu. Rev. Cell Dev. Biol. 19:397–422.
50. Higginbottom, A., E. R. Quinn, C. C. Kuo, M. Flint, L. H. Wilson, E.Bianchi, A. Nicosia, P. N. Monk, J. A. McKeating, and S. Levy. 2000.Identification of amino acid residues in CD81 critical for interaction withhepatitis C virus envelope glycoprotein E2. J. Virol. 74:3642–3649.
51. Higginbottom, A., Y. Takahashi, L. Bolling, S. A. Coonrod, J. M. White,L. J. Partridge, and P. N. Monk. 2003. Structural requirements for theinhibitory action of the CD9 large extracellular domain in sperm/oocytebinding and fusion. Biochem. Biophys. Res. Commun. 311:208–214.
52. Hildreth, J. E., A. Subramanium, and R. A. Hampton. 1997. Human T-celllymphotropic virus type 1 (HTLV-1)-induced syncytium formation medi-ated by vascular cell adhesion molecule-1: evidence for involvement of celladhesion molecules in HTLV-1 biology. J. Virol. 71:1173–1180.
53. Ho, D. D., A. U. Neumann, A. S. Perelson, W. Chen, J. M. Leonard, and M.Markowitz. 1995. Rapid turnover of plasma virions and CD4 lymphocytesin HIV-1 infection. Nature 373:123–126.
54. Hofmann-Lehmann, R., J. Vlasak, A. L. Williams, A. L. Chenine, H. M.McClure, D. C. Anderson, S. O’Neil, and R. M. Ruprecht. 2003. Liveattenuated, nef-deleted SIV is pathogenic in most adult macaques afterprolonged observation. AIDS 17:157–166.
55. Hohdatsu, T., H. Hirabayashi, K. Motokawa, and H. Koyama. 1996. Com-parative study of the cell tropism of feline immunodeficiency virus isolatesof subtypes A, B and D classified on the basis of the env gene V3-V5sequence. J. Gen. Virol. 77:93–100.
56. Huerta, S., D. M. Harris, A. Jazirehi, B. Bonavida, D. Elashoff, E. H.Livingston, and D. Heber. 2003. Gene expression profile of metastatic coloncancer cells resistant to cisplatin-induced apoptosis. Int. J. Oncol. 22:663–670.
57. Imai, T., K. Fukudome, S. Takagi, M. Nagira, M. Furuse, N. Fukuhara, M.Nishimura, Y. Hinuma, and O. Yoshie. 1992. C33 antigen recognized bymonoclonal antibodies inhibitory to human T cell leukemia virus type1-induced syncytium formation is a member of a new family of transmem-brane proteins including CD9, CD37, CD53, and CD63. J. Immunol. 149:2879–2886.
58. Ishibashi, T., L. Ding, K. Ikenaka, Y. Inoue, K. Miyado, E. Mekada, and H.Baba. 2004. Tetraspanin protein CD9 is a novel paranodal componentregulating paranodal junctional formation. J. Neurosci. 24:96–102.
59. Israels, S., E. M. McMillan-Ward, J. Easton, C. Robertson, and A. McNi-col. 2001. CD63 associates with the alphaIIb beta3 integrin-CD9 complexon the surface of activated platelets. Thromb. Haemostasis 85:134–141.
60. Ivaska, J., R. D. Whelan, R. Watson, P. J. Parker. 2002. PKC epsiloncontrols the traffic of beta1 integrins in motile cells. EMBO J. 21:3608–3619.
61. Izumo, S., I. Goto, Y. Itoyama, T. Okajima, S. Watanabe, Y. Kuroda, S.Araki, M. Mori, S. Nagataki, S. Matsukura, T. Akamine, M. Nakagawa, I.Yamamoto, and M. Osame. 1996. Interferon-alpha is effective in HTLV-I-associated myelopathy: a multicenter, randomized, double-blind, controlledtrial. Neurology 46:1016–1021.
62. Jacobson, J. M., R. J. Israel, I. Lowy, N. A. Ostrow, L. S. Vassilatos, M.Barish, D. N. Tran, B. M. Sullivan, T. J. Ketas, T. J. O’Neill, K. A.Nagashima, W. Huang, C. J. Petropoulos, J. P. Moore, P. J. Maddon, andW. C. Olson. 2004. Treatment of advanced human immunodeficiency virustype 1 disease with the viral entry inhibitor PRO 542. Antimicrob. AgentsChemother. 48:423–429.
63. Jankowski, S. A., D. S. Mitchell, S. H. Smith, J. M. Trent, and P. S. Meltzer.1994. SAS, a gene amplified in human sarcomas, encodes a new member ofthe transmembrane 4 superfamily of proteins. Oncogene 9:1205–1211.
64. Jones, M., and D. Y. Mason. 1997. CD81 workshop panel report, p. 187–188. In T. Kishimoto et al. (ed.), Leukocyte typing VI. Garland Publishing,New York, N.Y.
65. Kaji, K., S. Oda, S. Miyazaki, and A. Kudo. 2002. Infertility of CD9-deficient mouse eggs is reversed by mouse CD9, human CD9, or mouse
10848 MINIREVIEW J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.

CD81; polyadenylated mRNA injection developed for molecular analysis ofsperm-egg fusion. Dev. Biol. 247:327–334.
66. Reference deleted.67. Kim, F. J., N. Manel, Y. Boublik, J. L. Battini, and M. Sitbon. 2003. Human
T-cell leukemia virus type 1 envelope-mediated syncytium formation can beactivated in resistant mammalian cell lines by a carboxy-terminal truncationof the envelope cytoplasmic domain. J. Virol. 77:963–969.
68. Kitadokoro, K., D. Bordo, G. Galli, R. Petracca, F. Falugi, S. Abrignani, G.Grandi, and M. Bolognesi. 2001. CD81 extracellular domain 3D structure:insight into the tetraspanin superfamily structural motifs. EMBO J. 20:12–18.
69. Kreidberg, J., M. J. Donovan, S. L. Goldstein, H. Rennke, K. Shepherd,R. C. Jones, and R. Jaenisch. 1996. Alpha 3 beta 1 integrin has a crucial rolein kidney and lung organogenesis. Development 122:3537–3547.
70. Kronenberger, B., C. Sarrazin, W. P. Hofmann, M. von Wagner, E. Herr-mann, C. Welsch, R. Elez, B. Ruster, A. Piiper, and S. Zeuzem. 2004.Mutations in the putative HCV-E2 CD81 binding regions and correlationwith cell surface CD81 expression. J. Viral Hepatol. 11:310–318.
71. Kuritzkes, D. R., J. Jacobson, W. G. Powderly, E. Godofsky, E. DeJesus, F.Haas, K. A. Reimann, J. L. Larson, P. O. Yarbough, V. Curt, and W. R.Shanahan, Jr. 2004. Antiretroviral activity of the anti-CD4 monoclonalantibody TNX-355 in patients infected with HIV type 1. J. Infect. Dis.189:286–291.
72. Kwong, P. D., M. L. Doyle, D. J. Casper, C. Cicala, S. A. Leavitt, S. Majeed,T. D. Steenbeke, M. Venturi, I. Chaiken, M. Fung, H. Katinger, P. W.Parren, J. Robinson, D. Van Ryk, L. Wang, D. R. Burton, E. Freire, R.Wyatt, J. Sodroski, W. A. Hendrickson, and J. Arthos. 2002. HIV-1 evadesantibody-mediated neutralization through conformational masking of re-ceptor-binding sites. Nature 420:678–682.
73. Kwong, P. D., R. Wyatt, J. Robinson, R. W. Sweet, J. Sodroski, and W. A.Hendrickson. 1998. Structure of an HIV gp120 envelope glycoprotein incomplex with the CD4 receptor and a neutralizing human antibody. Nature393:648–659.
74. Lagaudriere-Gesbert, C., and H. Conjeaud. 1997. CD82 workshop panelreport, p. 189–190. In T. Kishimoto (ed.), Leukocyte typing VI. GarlandPublishing, New York, N.Y.
75. Lagaudriere-Gesbert, C., F. Le Naour, S. Lebel-Binay, M. Billard, E.Lemichez, P. Boquet, C. Boucheix, H. Conjeaud, and E. Rubinstein. 1997.Functional analysis of four tetraspans, CD9, CD53, CD81, and CD82,suggests a common role in costimulation, cell adhesion, and migration: onlyCD9 upregulates HB-EGF activity. Cell. Immunol. 182:105–112.
76. Lammerding, J., A. R. Kazarov, H. Huang, R. T. Lee, and M. E. Hemler.2003. Tetraspanin CD151 regulates �6�1 integrin adhesion strengthening.Proc. Natl. Acad. Sci. USA 100:7616–7621.
77. Lau, L., J. L. Wee, M. D. Wright, G. W. Moseley, P. M. Hogarth, L. K.Ashman, and D. E. Jackson. 2004. The tetraspanin superfamily memberCD151 regulates outside-in integrin alphaIIbbeta3 signaling and plateletfunction. Blood 104:2368–2375.
78. Lauer, G. M., and B. D. Walker. 2001. Hepatitis C virus infection. N. Engl.J. Med. 345:41–52.
79. Le Naour, F., E. Rubinstein, C. Jasmin, M. Prenant, and C. Boucheix. 2000.Severely reduced female fertility in CD9-deficient mice. Science 287:319–321.
80. Levine, A. M., S. Groshen, J. Allen, K. M. Munson, D. J. Carlo, A. E. Daigle,F. Ferre, F. C. Jensen, S. P. Richieri, R. J. Trauger, J. W. Parker, P. L. Salk,and J. Salk. 1996. Initial studies on active immunization of HIV-infectedsubjects using a gp120-depleted HIV-1 immunogen: long-term follow-up. J.Acquir. Immune Defic. Syndr. Hum. Retrovirol. 11:351–364.
81. Levy, S., S. C. Todd, and H. T. Maecker. 1998. CD81 (TAPA-1): a moleculeinvolved in signal transduction and cell adhesion in the immune system.Annu. Rev. Immunol. 16:89–109.
82. Liu, X. L., K. R. Clark, and P. R. Johnson. 1999. Production of recombinantadeno-associated virus vectors using a packaging cell line and a hybridrecombinant adenovirus. Gene Ther. 6:293–299.
83. Loffler, S., F. Lottspeich, F. Lanza, D. O. Azorsa, V. ter Meulen, and J.Schneider-Schaulies. 1997. CD9, a tetraspan transmembrane protein, ren-ders cells susceptible to canine distemper virus. J. Virol. 71:42–49.
84. Lozach, P. Y., H. Lortat-Jacob, A. de Lacroix de Lavalette, I. Staropoli, S.Foung, A. Amara, C. Houles, F. Fieschi, O. Schwartz, J. L. Virelizier, F.Arenzana-Seisdedos, and R. Altmeyer. 2003. DC-SIGN and L-SIGN arehigh affinity binding receptors for hepatitis C virus glycoprotein E2. J. Biol.Chem. 278:20358–20366.
85. Lu, S., J. Arthos, D. C. Montefiori, Y. Yasutomi, K. Manson, F. Mustafa, E.Johnson, J. C. Santoro, J. Wissink, J. I. Mullins, J. R. Haynes, N. L. Letvin,M. Wyand, and H. L. Robinson. 1996. Simian immunodeficiency virus DNAvaccine trial in macaques. J. Virol. 70:3978–3991.
86. Maecker, H. 2003. Human CD81 directly enhances Th1 and Th2 cell acti-vation, but preferentially induces proliferation of Th2 cells upon long-termstimulation. BMC Immunol. 4:1.
87. Maecker, H., S. C. Todd, and S. Levy. 1997. The tetraspanin superfamily:molecular facilitators. FASEB J. 11:428–442.
88. Manel, N., F. J. Kim, S. Kinet, N. Taylor, M. Sitbon, and J. L. Battini. 2003.
The ubiquitous glucose transporter GLUT-1 is a receptor for HTLV. Cell115:449–459.
89. Mantegazza, A., M. M. Barrio, S. Moutel, L. Bover,M. Weck, P. Brossart,J. L. Teillaud, and J. Mordoh. 2004. CD63 tetraspanin slows down cellmigration and translocates to the endosomal-lysosomal-MIICs route afterextracellular stimuli in human immature dendritic cells. Blood 104:1183–1190.
90. Marlink, R., P. Kanki, I. Thior, K. Travers, G. Eisen, T. Siby, I. Traore,C. C. Hsieh, M. C. Dia, and E. H. Gueye. 1994. Reduced rate of diseasedevelopment after HIV-2 infection as compared to HIV-1. Science 265:1587–1590.
91. Masciopinto, F., S. Campagnoli, S. Abrignani, Y. Uematsu, and P. Pileri.2001. The small extracellular loop of CD81 is necessary for optimal surfaceexpression of the large loop, a putative HCV receptor. Virus Res. 80:1–10.
92. Masciopinto, F., C. Giovani, S. Campagnoli, L. Galli-Stampino, P. Colom-batto, M. Brunetto, T. S. Yen, M. Houghton, P. Pileri, and S. Abrignani.2004. Association of hepatitis C virus envelope proteins with exosomes.Eur. J. Immunol. 34:2834.
93. Masellis-Smith, A., and A. R. Shaw. 1994. CD9-regulated adhesion. Anti-CD9 monoclonal antibody induce pre-B cell adhesion to bone marrowfibroblasts through de novo recognition of fibronectin. J. Immunol. 152:2768–2777.
94. McKeating, J. A., L. Q. Zhang, C. Logvinoff, M. Flint, J. Zhang, J. Yu, D.Butera, D. D. Ho, L. B. Dustin, C. M. Rice, and P. Balfe. 2004. Diversehepatitis C virus glycoproteins mediate viral infection in a CD81-dependentmanner. J. Virol. 78:8496–8505.
95. Meerloo, T., H. K. Parmentier, A. D. Osterhaus, J. Goudsmit, and H. J.Schuurman. 1992. Modulation of cell surface molecules during HIV-1infection of H9 cells. An immunoelectron microscopic study. AIDS 6:1105–1116.
96. Meerloo, T., M. A. Sheikh, A. C. Bloem, A. de Ronde, M. Schutten, C. A. vanEls, P. J. Roholl, P. Joling, J. Goudsmit, and H. J. Schuurman. 1993. Hostcell membrane proteins on human immunodeficiency virus type 1 after invitro infection of H9 cells and blood mononuclear cells. An immuno-electron microscopic study. J. Gen. Virol. 74:129–135.
97. Meola, A., A. Sbardellati, B. Bruni Ercole, M. Cerretani, M. Pezzanera, A.Ceccacci, A. Vitelli, S. Levy, A. Nicosia, C. Traboni, J. McKeating, and E.Scarselli. 2000. Binding of hepatitis C virus E2 glycoprotein to CD81 doesnot correlate with species permissiveness to infection. J. Virol. 74:5933–5938.
98. Min, G., M. Stolz, G. Zhou, F. Liang, P. Sebbel, D. Stoffler, R. Glockshuber,T. T. Sun, U. Aebi, and X. P. Kong. 2002. Localization of uroplakin Ia, theurothelial receptor for bacterial adhesin FimH, on the six inner domains ofthe 16 nm urothelial plaque particle. J. Mol. Biol. 317:697–706.
99. Miyado, K., G. Yamada, S. Yamada, H. Hasuwa, Y. Nakamura, F. Ryu, K.Suzuki, K. Kosai, K. Inoue, A. Ogura, M. Okabe, and E. Mekada. 2000.Requirement of CD9 on the egg plasma membrane for fertilization. Science287:321–324.
100. Miyazaki, T., U. Muller, and K. S. Campbell. 1997. Normal developmentbut differentially altered proliferative responses of lymphocytes in micelacking CD81. EMBO J. 16:4217–4225.
101. Mondal, D., L. Pradhan, M. Ali, and K. C. Agrawal. 2004. HAART drugsinduce oxidative stress in human endothelial cells and increase endothelialrecruitment of mononuclear cells: exacerbation by inflammatory cytokinesand amelioration by antioxidants. Cardiovasc. Toxicol. 4:287–302.
102. Moore, J. P., and R. W. Doms. 2003. The entry of entry inhibitors: a fusionof science and medicine. Proc. Natl. Acad. Sci USA 100:10598–10602.
103. Moseley, G. W. 2005. Tetraspanin-Fc receptor interactions. Platelets 16:3–11.
104. Moseley, G. W., J. Elliott, M. D. Wright, L. J. Partridge, and P. N. Monk.2003. Interspecies contamination of the KM3 cell line: implications forCD63 function in melanoma metastasis. Int. J. Cancer 105:613–616.
105. Murphy, C. G., W. T. Lucas, R. E. Means, S. Czajak, C. L. Hale, J. D.Lifson, A. Kaur, R. P. Johnson, D. M. Knipe, and R. C. Desrosiers. 2000.Vaccine protection against simian immunodeficiency virus by recombinantstrains of herpes simplex virus. J. Virol. 74:7745–7754.
106. Myszka, D. G., R. W. Sweet, P. Hensley, M. Brigham-Burke, P. D. Kwong,W. A. Hendrickson, R. Wyatt, J. Sodroski, and M. L. Doyle. 2000. Ener-getics of the HIV gp120-CD4 binding reaction. Proc. Natl. Acad. Sci. USA97:9026–9031.
107. Nagai, M., M. B. Brennan, J. A. Sakai, C. A. Mora, and S. Jacobson. 2001.CD8� T cells are an in vivo reservoir for human T-cell lymphotropic virustype I. Blood 98:1858–1861.
108. Nakamura, T., K. Shibayama, K. Nagasato, H. Matsuo, M. Tsujihata, andS. Nagataki. 1990. The efficacy of interferon-alpha treatment in humanT-lymphotropic virus type-I-associated myelopathy. Jpn. J. Med. 29:362–367.
109. Nattermann, J., H. D. Nischalke, B. Kupfer, J. Rockstroh, L. Hess, T.Sauerbruch, and U. Spengler. 2003. Regulation of CC chemokine receptor5 in hepatitis G virus infection. AIDS 17:1457–1462.
110. Odintsova, E., T. Sugiura, and F. Berditchevski. 2000. Attenuation of EGF
VOL. 79, 2005 MINIREVIEW 10849
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.

receptor signaling by a metastasis suppressor, the tetraspanin CD82/KAI-1.Curr. Biol. 10:1009–1012.
111. Ono, M., K. Handa, D. A. Withers, and S. Hakomori. 2000. Glycosylationeffect on membrane domain (GEM) involved in cell adhesion and motility:a preliminary note on functional �3, �5-CD82 glycosylation complex inIdID 14 cells. Biochem. Biophys. Res. Commun. 279:744–750.
112. Ono, M., K. Handa, D. A. Withers, and S. Hakomori. 1999. Motility inhi-bition and apoptosis are induced by metastasis-suppressing gene productCD82 and its analogue CD9, with concurrent glycosylation. Cancer Res.59:2335–2339.
113. Reference deleted.114. Orenstein, J. M., M. S. Meltzer, T. Phipps, and H. E. Gendelman. 1988.
Cytoplasmic assembly and accumulation of human immunodeficiency virustypes 1 and 2 in recombinant human colony-stimulating factor-1-treatedhuman monocytes: an ultrastructural study. J. Virol. 62:2578–2586.
115. Palella, F. J., Jr., K. M. Delaney, A. C. Moorman, M. O. Loveless, J. Fuhrer,G. A. Satten, D. J. Aschman, and S. D. Holmberg. 1998. Declining mor-bidity and mortality among patients with advanced human immunodefi-ciency virus infection. N. Engl. J. Med. 338:853–860.
116. Parren, P. W., J. P. Moore, D. R. Burton, and Q. J. Sattentau. 1999. Theneutralizing antibody response to HIV-1: viral evasion and escape fromhumoral immunity. AIDS 13(Suppl. A):S137–S162.
117. Pelchen-Matthews, A., B. Kramer, and M. Marsh. 2003. Infectious HIV-1assembles in late endosomes in primary macrophages. J. Cell Biol. 162:443–455.
118. Penas, P., A. Garcia-Diez, F. Sanchez-Madrid, and M. Yanez-Mo. 2000.Tetraspanins are localized at motility-related structures and involved innormal human keratinocyte wound healing migration. J. Investig. Derma-tol. 114:1126–1135.
119. Perez-Bercoff, D., A. David, H. Sudry, F. Barre-Sinoussi, and G. Pancino.2003. Fc� receptor-mediated suppression of human immunodeficiency virustype 1 replication in primary human macrophages. J. Virol. 77:4081–4094.
120. Petracca, R., F. Falugi, G. Galli, N. Norais, D. Rosa, S. Campagnoli, V.Burgio, E. Di Stasio, B. Giardina, M. Houghton, S. Abrignani, and G.Grandi. 2000. Structure-function analysis of hepatitis C virus envelope-CD81 binding. J. Virol. 74:4824–4830.
121. Phillips, A. 2001. Will the drugs still work? Transmission of resistant HIV.Nat. Med. 7:993–994.
122. Pileri, P., Y. Uematsu, S. Campagnoli, G. Galli, F. Falugi, R. Petracca, A. J.Weiner, M. Houghton, D. Rosa, G. Grandi, and S. Abrignani. 1998. Bindingof hepatitis C virus to CD81. Science 282:938–941.
123. Pinon, J. D., P. J. Klasse, S. R. Jassal, S. Welson, J. Weber, D. W. Brighty,and Q. J. Sattentau. 2003. Human T-cell leukemia virus type 1 envelopeglycoprotein gp46 interacts with cell surface heparan sulfate proteoglycans.J. Virol. 77:9922–9930.
124. Pique, C., C. Lagaudriere-Gesbert, L. Delamarre, A. R. Rosenberg, H.Conjeaud, and M. C. Dokhelar. 2000. Interaction of CD82 tetraspaninproteins with HTLV-1 envelope glycoproteins inhibits cell-to-cell fusionand virus transmission. Virology 276:455–465.
125. Poduri, C. D. 2003. Hepatitis C virus (HCV)[em]a review molecular biologyof the virus, immunodiagnostics, genomic heterogeneity and the role ofvirus in hepatocellular carcinoma. Indian J. Exp. Biol. 41:549–562.
126. Pohlmann, S., J. Zhang, F. Baribaud, Z. Chen, G. J. Leslie, G. Lin, A.Granelli-Piperno, R. W. Doms, C. M. Rice, and J. A. McKeating. 2003.Hepatitis C virus glycoproteins interact with DC-SIGN and DC-SIGNR.J. Virol. 77:4070–4080.
127. Preston, B. D. 1997. Reverse transcriptase fidelity and HIV-1 variation.Science 275:228–231.
128. Raposo, G., M. Moore, D. Innes, R. Leijendekker, A. Leigh-Brown, P.Benaroch, and H. Geuze. 2002. Human macrophages accumulate HIV-1particles in MHC II compartments. Traffic 3:718–729.
129. Rice, C. M. 1999. Is CD81 the key to hepatitis C virus entry? Hepatology29:990–992.
130. Richardson, J. H., A. J. Edwards, J. K. Cruickshank, P. Rudge, and A. G.Dalgleish. 1990. In vivo cellular tropism of human T-cell leukemia virustype 1. J. Virol. 64:5682–5687.
131. Roccasecca, R., H. Ansuini, A. Vitelli, A. Meola, E. Scarselli, S. Acali, M.Pezzanera, B. B. Ercole, J. McKeating, A. Yagnik, A. Lahm, A. Tramon-tano, R. Cortese, and A. Nicosia. 2003. Binding of the hepatitis C virus E2glycoprotein to CD81 is strain specific and is modulated by a complexinterplay between hypervariable regions 1 and 2. J. Virol. 77:1856–1867.
132. Rosa, D. 1996. E2 binds to chimpanzee mononuclear cells, whereas it doesnot bind to rat, rabbit, or green monkey mononuclear cells or hepatocytes.Proc. Natl. Acad. Sci. USA 93:1759.
133. Rose, N. F., P. A. Marx, A. Luckay, D. F. Nixon, W. J. Moretto, S. M.Donahoe, D. Montefiori, A. Roberts, L. Buonocore, and J. K. Rose. 2001.An effective AIDS vaccine based on live attenuated vesicular stomatitisvirus recombinants. Cell 106:539–549.
134. Rubinstein, E., F. Le Naour, M. Billard, M. Prenant, C. Boucheix. 1994.CD9 antigen is an accessory subunit of the VLA integrin complexes. Eur.J. Immunol. 24:3005–3013.
135. Rubinstein, E., F. Le Naour, C. Lagaudriere-Gesbert, M. Billard, H. Con-
jeaud, C. Boucheix. 1996. CD9, CD63, CD81, and CD82 are components ofa surface tetraspan network connected to HLA-DR and VLA integrins.Eur. J. Immunol. 26:2657–2665.
136. Sakuma, T., S. Higashiyama, S. Hosoe, S. Hayashi, and N. Taniguchi. 1997.CD9 antigen interacts with heparin-binding EGF-like growth factorthrough its heparin-binding domain. J. Biochem. 122:474–480.
137. Sasaki, M., K. Yamauchi, T. Nakanishi, Y. Kamogawa, and N. Hayashi.2003. In vitro binding of hepatitis C virus to CD81-positive and -negativehuman cell lines. J. Gastroenterol. Hepatol. 18:74–79.
138. Satomi, Y., Y. Shimonishi, and T. Takao. 2004. N-glycosylation at Asn491 inthe Asn-Xaa-Cys motif of human transferrin. FEBS Lett. 576:51–56.
139. Scarselli, E., H. Ansuini, R. Cerino, R. M. Roccasecca, S. Acali, G. Filo-camo, C. Traboni, A. Nicosia, R. Cortese, and A. Vitelli. 2002. The humanscavenger receptor class B type I is a novel candidate receptor for thehepatitis C virus. EMBO J. 21:5017–5025.
140. Scherberich, A., G. Giannone, E. Perennou, K. Takeda, C. Boucheix, E.Rubinstein, F. Lanza, and A. Beretz. 2002. FAK-mediated inhibition ofvascular smooth muscle cell migration by the tetraspanin CD9. Thromb.Haemostasis 87:1043–1050.
141. Schmid, E., A. Zurbriggen, U. Gassen, B. Rima, V. ter Meulen, and J.Schneider-Schaulies. 2000. Antibodies to CD9, a tetraspan transmembraneprotein, inhibit canine distemper virus-induced cell-cell fusion but not vi-rus-cell fusion. J. Virol. 74:7554–7561.
142. Seigneuret, M., A. Delaguillaumie, C. Lagaudriere-Gesbert, and H. Con-jeaud. 2001. Structure of the tetraspanin main extracellular domain. Apartially conserved fold with a structurally variable domain insertion.J. Biol. Chem. 276:40055–40064.
143. Shaw, A., A. Domanska, A. Mak, A. Gilchrist, K. Dobler, L. Visser, S.Poppema, L. Fliegel, M. Letarte, and B. J. Willett. 1995. Ectopic expressionof human and feline CD9 in a human B cell line confers beta 1 integrin-dependent motility on fibronectin and laminin substrates and enhancedtyrosine phosphorylation. J. Biol. Chem. 270:24092–24099.
144. Shi, W., H. Fan, L. Shum, and R. Derynck. 2000. The tetraspanin CD9associates with transmembrane TGF-alpha and regulates TGF-alpha-in-duced EGF receptor activation and cell proliferation. J. Cell Biol. 148:591–602.
145. Shibayama, K., T. Nakamura, K. Nagasato, S. Shirabe, M. Tsujihata, andS. Nagataki. 1991. Interferon-alpha treatment in HTLV-I-associated my-elopathy. Studies of clinical and immunological aspects. J. Neurol. Sci.106:186–192.
146. Shiver, J. W., T. M. Fu, L. Chen, D. R. Casimiro, M. E. Davies, R. K. Evans,Z. Q. Zhang, A. J. Simon, W. L. Trigona, S. A. Dubey, L. Huang, V. A.Harris, R. S. Long, X. Liang, L. Handt, W. A. Schleif, L. Zhu, D. C. Freed,N. V. Persaud, L. Guan, K. S. Punt, A. Tang, M. Chen, K. A. Wilson, K. B.Collins, G. J. Heidecker, V. R. Fernandez, H. C. Perry, J. G. Joyce, K. M.Grimm, J. C. Cook, P. M. Keller, D. S. Kresock, H. Mach, R. D. Troutman,L. A. Isopi, D. M. Williams, Z. Xu, K. E. Bohannon, D. B. Volkin, D. C.Montefiori, A. Miura, G. R. Krivulka, M. A. Lifton, M. J. Kuroda, J. E.Schmitz, N. L. Letvin, M. J. Caulfield, A. J. Bett, R. Youil, D. C. Kaslow,and E. A. Emini. 2002. Replication-incompetent adenoviral vaccine vectorelicits effective anti-immunodeficiency-virus immunity. Nature415:331–335.
147. Silvie, O., E. Rubinstein, J. F. Franetich, M. Prenant, E. Belnoue, L. Renia,L. Hannoun, W. Eling, S. Levy, C. Boucheix, and D. Mazier. 2003. Hepa-tocyte CD81 is required for Plasmodium falciparum and Plasmodium yoeliisporozoite infectivity. Nat. Med. 9:93–96.
148. Simon, F., S. Matheron, C. Tamalet, I. Loussert-Ajaka, S. Bartczak, J. M.Pepin, C. Dhiver, E. Gamba, C. Elbim, and J. A. Gastaut. 1993. Cellularand plasma viral load in patients infected with HIV-2. AIDS 7:1411–1417.
149. Simons, K., and E. Ikonen. 1997. Functional rafts in cell membranes.Nature 387:569–572.
150. Sincock, P., G. Mayrhofer, and L. K. Ashman. 1997. Localization of thetransmembrane 4 superfamily (TM4SF) member PETA-3 (CD151) in nor-mal human tissues: comparison with CD9, CD63, and �5�1 integrin. J. His-tochem. Cytochem. 45:515–525.
151. Song, B., S. Levy, and E. E. Geisert, Jr. 2004. Increased density of retinalpigment epithelium in cd81�/� mice. J. Cell. Biochem. 92:1160–1170.
152. Stipp, C., T. V. Kolesnikova, and M. E. Hemler. 2003. Functional domainsin tetraspanin proteins. Trends Biochem. Sci. 28:106–112.
153. Sutton, R. E., and D. R. Littman. 1996. Broad host range of human T-cellleukemia virus type 1 demonstrated with an improved pseudotyping system.J. Virol. 70:7322–7336.
154. Szollosi, J., V. Horejsi, L. Bene, P. Angelisova, and S. Damjanovich. 1996.Supramolecular complexes of MHC class I, MHC class II, CD20, andtetraspan molecules (CD53, CD81, and CD82) at the surface of a B cell lineJY. J. Immunol. 157:2939–2946.
155. Tachibana, I., and M. E. Hemler, M. E. 1999. Role of transmembrane 4superfamily (TM4SF) proteins CD9 and CD81 in muscle cell fusion andmyotube maintenance. J. Cell Biol. 146:893–904.
156. Tan, Y. J., S. P. Lim, P. Ng, P. Y. Goh, S. G. Lim, Y. H. Tan, and W. Hong.2003. CD81 engineered with endocytotic signals mediates HCV cell entry:implications for receptor usage by HCV in vivo. Virology 308:250–269.
10850 MINIREVIEW J. VIROL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.

157. Teoh, N. C., and G. C. Farrell. 2004. Management of chronic hepatitis Cvirus infection: a new era of disease control. Intern. Med. J. 34:324–337.
158. Trejo, S. R., and L. Ratner. 2000. The HTLV receptor is a widely expressedprotein. Virology 268:41–48.
159. Tseng, C. T., and G. R. Klimpel. 2002. Binding of the hepatitis C virusenvelope protein E2 to CD81 inhibits natural killer cell functions. J. Exp.Med. 195:43–49.
160. VanCompernolle, S. E., A. V. Wiznycia, J. R. Rush, M. Dhanasekaran,P. W. Baures, and S. C. Todd. 2003. Small molecule inhibition of hepatitisC virus E2 binding to CD81. Virology 314:371–380.
161. von Lindern, J. J., D. Rojo, K. Grovit-Ferbas, C. Yeramian, C. Deng, G.Herbein, M. R. Ferguson, T. C. Pappas, J. M. Decker, A. Singh, R. G.Collman, and W. A. O’Brien. 2003. Potential role for CD63 in CCR5-mediated human immunodeficiency virus type 1 infection of macrophages.J. Virol. 77:3624–3633.
162. Watanabe, M. E. 2003. Skeptical scientists skewer VaxGen statistics. Nat.Med. 9:376.
163. Willett, B., M. Hosie, A. Shaw, and J. Neil. 1997. Inhibition of felineimmunodeficiency virus infection by CD9 antibody operates after virusentry and is independent of virus tropism. J. Gen. Virol. 78:611–618.
164. Willett, B. J., M. J. Hosie, O. Jarrett, and J. C. Neil. 1994. Identification ofa putative cellular receptor for feline immunodeficiency virus as the felinehomologue of CD9. Immunology 81:228–233.
165. Wollscheid, V., R. Kuhne-Heid, I. Stein, L. Jansen, S. Kollner, A. Schnei-der, and M. Durst. 2002. Identification of a new proliferation-associatedprotein NET-1/C4.8 characteristic for a subset of high-grade cervical intra-epithelial neoplasia and cervical carcinomas. Int. J. Cancer. 99:771–775.
166. Wright, M., S. M. Geary, S. Fitter, G. W. Moseley, L. M. Lau, K. C. Sheng,V. Apostolopoulos, E. G. Stanley, D. E. Jackson, and L. Ashman. 2004.Characterization of mice lacking the tetraspanin superfamily memberCD151. Mol. Cell. Biol. 24:5978–5988.
167. Wright, M., G. W. Moseley, and A. B. VanSpriel. 2004. Tetraspanin mi-crodomains in immune cell signalling and malignant disease. Tissue Anti-gens 64:533–542.
168. Wright, M., and M. Tomlinson. 1994. The ins and outs of the transmem-brane-4 superfamily. Immunol. Today 15:588–594.
169. Wubbolts, R., R. S. Leckie, P. T. Veenhuizen, G. Schwarzmann, W. Mobius,J. Hoernschemeyer, J. W. Slot, H. J. Geuze, and W. Stoorvogel. 2003.Proteomic and biochemical analyses of human B cell-derived exosomes.Potential implications for their function and multivesicular body formation.J. Biol. Chem. 278:10963–10972.
170. Wunschmann, S., J. D. Medh, D. Klinzmann, W. N. Schmidt, and J. T.Stapleton. 2000. Characterization of hepatitis C virus (HCV) and HCV E2interactions with CD81 and the low-density lipoprotein receptor. J. Virol.74:10055–10062.
171. Yamasaki, K., J. Kira, Y. Koyanagi, Y. Kawano, N. Miyano-Kurosaki, M.Nakamura, E. Baba, J. Suzuki, A. Yamamoto, N. Yamamoto, and T. Koba-yashi. 1997. Long-term, high dose interferon-alpha treatment in HTLV-I-associated myelopathy/tropical spastic paraparesis: a combined clinical, vi-rological and immunological study. J. Neurol. Sci. 147:135–144.
172. Yauch, R., and M. E. Hemler. 2000. Specific interactions among transmem-brane 4 superfamily (TM4SF) proteins and phosphoinositide 4-kinase. Bio-chem. J. 351:629–637.
173. Yauch, R., A. R. Kazarov, B. Desai, R. T. Lee, and M. E. Hemler. 2000.Direct extracellular contact between integrin �3�1 and TM4SF proteinCD151. J. Biol. Chem. 275:9230–9238.
174. Zemni, R., T. Bienvenu, M. C. Vinet, A. Sefiani, A. Carrie, A., P. Billuart,N. McDonell, P. Couvert, F. Francis, P. Chafey, F. Fauchereau, G. Frio-court, V. des Portes, A. Cardona, S. Frints, A. Meindl, O. Brandau, N.Ronce, C. Moraine, H. van Bokhoven, H. H. Ropers, R. Sudbrak, A. Kahn,J. P. Fryns, C. Beldjord, and J. Chelly. 2000. A new gene involved inX-linked mental retardation identified by analysis of an X;2 balanced trans-location. Nat. Genet. 24:167–170.
175. Zhang, J., G. Randall, A. Higginbottom, P. Monk, C. M. Rice, and J. A.McKeating. 2004. CD81 is required for hepatitis C virus glycoprotein-mediated viral infection. J. Virol. 78:1448–1455.
176. Zhang, X., A. L. Bontrager, and M. E. Hemler. 2001. Transmembrane-4superfamily proteins associate with activated protein kinase C (PKC) andlink PKC to specific �1 integrins. J. Biol. Chem. 276:25005–25013.
VOL. 79, 2005 MINIREVIEW 10851
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
vi o
n 23
Nov
embe
r 20
21 b
y 68
.172
.248
.30.