testing the limits of paleozoic chronostratigraphic
TRANSCRIPT

1700
ABSTRACT
The resolution and fi delity of global chronostratigraphic correlation are direct functions of the time period under consid-eration. By virtue of deep-ocean cores and astrochronology, the Cenozoic and Meso-zoic time scales carry error bars of a few thousand years (k.y.) to a few hundred k.y. In contrast, most of the Paleozoic time scale carries error bars of plus or minus a few million years (m.y.), and chronostrati-graphic control better than ±1 m.y. is con-sidered “high resolution.” The general lack of Paleozoic abyssal sediments and paucity of orbitally tuned Paleozoic data series com-bined with the relative incompleteness of the Paleozoic stratigraphic record have proven historically to be such an obstacle to inter-continental chronostratigraphic correlation that resolving the Paleozoic time scale to the level achieved during the Mesozoic and Cenozoic was viewed as impractical, impos-sible, or both.
Here, we utilize integrated graptolite, cono-dont, and carbonate carbon isotope (δ13Ccarb) data from three paleocontinents (Baltica, Avalonia, and Laurentia) to demonstrate chronostratigraphic control for upper Llan-do very through middle Wenlock (Telychian–Sheinwoodian , ~436–426 Ma) strata with a resolution of a few hundred k.y. The interval surrounding the base of the Wenlock Series can now be correlated globally with preci-sion approaching 100 k.y., but some intervals (e.g., uppermost Telychian and upper Shein-woodian) are either yet to be studied in suffi -cient detail or do not show suffi cient biologic speciation and/or extinction or carbon isotopic features to delineate such small time slices. Although producing such resolution during the Paleozoic presents an array of challenges unique to the era, we have begun to demon-strate that erecting a Paleozoic time scale com-parable to that of younger eras is achievable.
INTRODUCTION
Historically, the Silurian has been considered to be an interval of relative climatic stability between the Late Ordovician icehouse and the Devo nian terrestrial revolution, during which high sea levels and a greenhouse climate com-
bined to produce an ~30-m.y.-long episode of little interest (Fischer, 1983; Bassett and Edwards, 1991). Over the past two decades, however, it has become increasingly clear that the Silurian is in fact among the climatically least stable periods of the Phanerozoic because it contains at least four major positive carbonate carbon isotope (δ13Ccarb) excursions >+4‰ (Samtleben et al., 1996, 2000; Saltzman, 2002; Eriksson and Calner, 2005; Calner and Eriksson , 2006; Cramer et al., 2006a; Kaljo et al., 2007), includ-ing the largest positive excursion of the Phanero-zoic with values exceeding +10‰ (Talent et al., 1993; Wigforss-Lange, 1999; Jeppsson et al., 2007). The dramatic global climate events that must have occurred during the Silurian in or-der to produce such variability in the δ13Ccarb record demonstrate the importance of Silurian research to our understanding of the evolution of the global climate system. However, one fundamental feature of Paleozoic stratigraphy has limited the direct applicability of deep-time studies to investigations of more recent time in-tervals and vice versa. The nearly complete lack of pre-Mesozoic deep-sea strata adds a degree of diffi culty to intercontinental chronostrati-graphic correlation that scientists working on post-Paleozoic strata seldom encounter; rarely can the Paleozoic record of climate change be
For permission to copy, contact [email protected]© 2010 Geological Society of America
GSA Bulletin; September/October 2010; v. 122; no. 9/10; p. 1700–1716; doi: 10.1130/B26602.1; 10 fi gures; Data Repository item 2010130.
†Current address: Kansas Geological Survey, Depart ment of Geology, University of Kansas, 1930 Constant Ave., Lawrence, Kansas 66047, USA; e-mail: [email protected]
Testing the limits of Paleozoic chronostratigraphic correlation via high-resolution (<500 k.y.) integrated conodont, graptolite, and carbon isotope (δ13Ccarb) biochemostratigraphy across the Llandovery–Wenlock
(Silurian) boundary: Is a unifi ed Phanerozoic time scale achievable?
Bradley D. Cramer1,†, David K. Loydell2, Christian Samtleben3, Axel Munnecke4, Dimitri Kaljo5, Peep Männik5, Tõnu Martma5, Lennart Jeppsson6, Mark A. Kleffner1, James E. Barrick7, Craig A. Johnson8, Poul Emsbo8, Michael M. Joachimski9, Torsten Bickert10, and Matthew R. Saltzman1
1Division of Earth History, School of Earth Sciences, The Ohio State University, 125 S. Oval Mall, Columbus, Ohio 43210, USA2School of Earth and Environmental Sciences, University of Portsmouth, Burnaby Building, Burnaby Road, Portsmouth P01 3QL, United Kingdom3Institut für Geowissenschaften, Universität Kiel, Olshausenstraße 40-60, 24118 Kiel, Germany4GeoZentrum Nordbayern, Fachgruppe Paläoumwelt, Universität Erlangen, Loewenichstraße 28, Erlangen, D-91054, Germany5Institute of Geology at Tallinn University of Technology, Ehitajate tee 5, 19086 Tallinn, Estonia6Department of Geology, GeoBiosphere Science Centre, Lund University, Sölvegatan 12, SE-223-62 Lund, Sweden7Department of Geosciences, Texas Tech University, Lubbock, Texas 79409, USA8U.S. Geological Survey, Box 25046, MS 963, Denver, Colorado 80225, USA9GeoZentrum Nordbayern, Fachgruppe Krustendynamik, Universität Erlangen, Schlossgarten 5, Erlangen, D-91054, Germany10Fachbereich Geowissenschaften, Universität Bremen, 28334 Bremen, Germany

Testing the limits of Paleozoic chronostratigraphy
Geological Society of America Bulletin, September/October 2010 1701
correlated globally with accuracy and precision better than one million years. The coarser reso-lution of the Paleozoic time scale limits direct application of modern global climate studies to Paleozoic problems because they typically con-centrate on much shorter (Milankovitch, millen-nial, etc.) time scales and events. There is no a priori reason why time scales and events of such short duration should not or could not be inves-tigated during the Paleozoic.
Largely independent efforts toward a high-resolution Silurian chronostratigraphy over the past decade (e.g., Jeppsson, 1997; Jeppsson et al., 2006; Männik, 1998, 2007a; Loydell, 1998; Loydell et al., 1998, 2003; Kaljo et al., 1998, 2003, 2007; Munnecke et al., 2003; Porębska et al., 2004; Cramer et al., 2006a, 2006b; Kaljo and Martma, 2006) have made signifi cant improvements to global Silurian chronostratigraphic correlation. Here, for the fi rst time, we integrate all of the available cono dont, graptolite, and carbonate carbon isotope data from seven of the chronostrati-graphically best constrained Silurian regions, outcrops, and cores worldwide (Fig. 1): the Robert Moses Power Plant S-1 core, New York, USA; the Banwy River section, Wales, UK; Gotland, Sweden; the Ohesaare core, Estonia; the Ruhnu core, Estonia; the Viki core, Esto-nia; and the Aizpute-41 core, Latvia; in order to demonstrate the global correlation potential of multiproxy Paleozoic chronostratigraphy.
The carbonate carbon isotope (δ13Ccarb) data from the Banwy River section and the Aizpute-41 core were analyzed for this inves-tigation. The δ13Ccarb data from diagenetically unaltered brachiopods from Gotland, Sweden, have been published by Samtleben et al. (1996, 2000, 2001) and Munnecke et al. (2003), and a generalized curve of this data set was published by Calner et al. (2004). In the present study new data from Gotland are added to this data set. All available conodont, graptolite, and carbon-ate carbon isotope data from the three Estonian cores (Ruhnu, Ohesaare, and Viki) are shown together for the fi rst time here, and they are combined with new data from the Ohesaare and Viki cores. In an effort to provide a chronostrati-graphic standard against which future research can be correlated, below we discuss the current state of early Silurian chronostratigraphy. We provide clarifi ed defi nitions of biostratigraphic and chemostratigraphic terms in order to mini-mize nomenclatural issues common to inter-disciplinary research.
Although the remainder of this paper deals with the technical aspects of Telychian–Shein-woodian global biochemostratigraphic corre-lation, this Silurian example can serve more broadly to demonstrate several general and
specifi c empirical warnings for future inte-grated, high-resolution, biochemostratigraphic correla tion. The scale of investigation is criti-cal. Whereas it should be expected that paleo-biological events are broadly synchronous with global climate events, at what scale of investigation are paleobiological and paleo-climatic events diachro nous? High-resolution chemostratigraphic data have the potential to provide extremely accurate and precise global correlations but still must be broadly verifi ed by biostratigraphy fi rst. Finally, there are strati-graphic and diagenetic factors that can limit the chronostratigraphic utility of chemostrati-graphic data, and in such cases, biostratigraphy must be integrated with chemostratigraphy to achieve the desired certainty of correlation.
SILURIAN CHRONOSTRATIGRAPHY
The Silurian Period represents a smaller amount of time than any other geologic period except the Neogene (Geologic Time Scale 2004 [GTS2004], Gradstein et al., 2004). Likewise,
Silurian epochs and ages are also comparatively short, so much so that the Wenlock, Ludlow, and Pridoli (the second, third, and fi nal of the four Silurian epochs, respectively) combined repre-sent less time (~12 m.y.) than the Famennian Age of the Devonian alone (~15 m.y.). The re-sult is a detailed chronostratigraphic framework making the Silurian a natural target for high-resolution chronostratigraphy.
Base of the Wenlock Series
The Wenlock Series was formally ratifi ed (Martinsson et al., 1981) as the second Silu-rian Series following a review of the classical strata around Wenlock Edge, Shropshire, UK (Bassett et al., 1975). A stream bank along Hughley Brook, no longer well exposed and subsequently shown to be faulted, was chosen as the stratotype section for the global boundary stratotype section and point (GSSP) of the base of the Wenlock Series. The GSSP was stated to coincide with the fi rst appearance of the grapto-lite Cyrtograptus centrifugus and therefore the
N
SWEDEN
LITHUANIA
LATVIA
ESTONIA
GOTLAND
GotlandComposite
BalticSea
Aizpute-41
RuhnuOhesaare
Viki
0 200 km
A
B
Niagara
Banwy River
Gotland
Viki
OhesaareAizpute-41
Ruhnu
Equator
15°
15°
30°
45°
15°
30°
45°
15°
Equator
N
Laurentia
Baltica
Figure 1. (A) Wenlock paleogeographic map after Woodcock (2000), Cocks (2001), and Johnson et al. (2001) modifi ed from Cramer and Saltzman (2005) to show Telychian–Sheinwoodian δ13Ccarb sample localities included in this investigation. (B) Present-day map of the Baltic Sea and surrounding countries showing the distribution of East Baltic sample localities.

Cramer et al.
1702 Geological Society of America Bulletin, September/October 2010
base of the Cyrtograptus centrifugus graptolite Biozone. Unfortunately the global stratotype section is devoid of graptolites in the criti-cal interval: they are absent from 10 m below to 3 m above the GSSP, requiring alternative biostratigraphic controls (i.e., conodonts and chitinozoans) to be used around this boundary (Mabillard and Aldridge, 1985). At present, global stratigraphic sections can be correlated to the GSSP using conodont biostratigraphy be-cause extinction Datum 2 of the Ireviken Event (Jeppsson, 1987, 1997) lies within a few cen-timeters (probably millimeters) of the GSSP (Mabillard and Aldridge, 1985; Jeppsson, 1997; Loydell, 2008a). It should be noted, however, that neither Apsidognathus nor Ozarkodina polinclinata polinclinata, the LAD (last appear-ance datum) of which are Datum 2, have yet been recovered from the stratotype section (see discussion in Jeppsson, 1997). David Loydell, who was asked by the International Subcom-mission on Silurian Stratigraphy (ISSS) to serve as “leader and organizer” of the task group for the base of the Wenlock Series, has detailed the problems regarding the formal defi nition of the boundary in Loydell (2008a) and the fi nd-ings of the task group are summarized below.
Because conodonts and chitinozoans are available at the global stratotype section, lo-calities that contain both conodonts and chitino-zoans together with graptolites became essential to defi ne the placement of the GSSP within the graptolite biostratigraphic zonation. Pri-marily through the study of Baltic cores and outcrops (Männik, 1998, 2007a, 2007b; Kaljo et al., 1998, 2003, 2007; Loydell et al., 1998, 2003; Loydell and Nestor, 2005) that contain conodonts, chitinozoans, and graptolites, it has been demonstrated that Datum 2 of the Ireviken Event (the de facto position of the GSSP) cor-relates to a position either at the base of or (more likely) within the Cyrtograptus murchi-soni graptolite Biozone (Loydell et al., 2003; Männik, 2007a, 2007b), which is at least a full zone higher than the original defi nition of the GSSP (coincident with the fi rst appearance of C. centrifugus) presented in Martinsson et al. (1981). This is consistent with the conclusions of Mullins and Aldridge (2004), who studied the chitinozoans of the Hughley Brook sec-tion and concluded, based on correlation with the graptolite- and chitinozoan-bearing Banwy River section, that the base of the Wenlock Series correlates to a position within the upper C. centrifugus to lower C. murchisoni graptolite biozone interval. Research on the base of the Wenlock Series is ongoing (Loydell, 2008a). Details of the graptolite and conodont species used to correlate the base of the Wenlock Series herein are provided below.
Telychian–Homerian Biostratigraphy
The graptolite biozonation of Loydell (1998) is used in this investigation. The base of each biozone is defi ned by the FAD (fi rst appearance datum) of the eponymous species (Fig. 2). This differs from other published biozonations (e.g., Melchin et al., 2004) by the use of the “mid-Wenlock” interval for the upper Sheinwoodian. This refl ects diffi culties in precise inter national correlation in this interval, resulting from (1) the provincialism of graptolites during this interval; (2) the need for taxonomic revision of some late Sheinwoodian graptolite taxa (e.g., “Monograptus” antennularius); and (3) the fact that in some areas (e.g., Wales; Zalasiewicz and Williams, 1999) the guide species Cyrtograp-tus rigidus and Monograptus fl exilis appear in the order C. rigidus then M. fl exilis, whereas in other areas (e.g., Poland; Teller, 1969), M. fl ex-ilis appears before C. rigidus. The guide species Cyrtograptus perneri has a similarly problem-atic fi rst occurrence as well (signifi cantly above the fi rst appearance of C. lundgreni, guide spe-cies of the lowest Homerian graptolite biozone,
in some regions; Zalasiewicz and Williams, 1999) and is another of the previously utilized zones included here within the “mid-Wenlock” interval. Throughout this paper, when grapto-lites have been used to correlate the position of the base of the Wenlock Series, we have used the fi rst appearance of C. murchisoni.
The conodont biozonations of Jeppsson (1997) and Männik (1998, 2007a) are used in this investigation (Fig. 3). The Telychian portion (Männik, 1998, 2007a, 2007b) has the base of each biozone defi ned by the FAD of the epony-mous species, whereas the interval surrounding the base of the Wenlock has biozones defi ned by the LAD of important, but not necessarily epon-ymous species (Jeppsson, 1997). Many of the Sheinwoodian biozones above the boundary in-terval are defi ned by the FAD of the eponymous species as well (Jeppsson, 1997). Until more research can be done, we suggest that use of the “Middle Kockelella walliseri,” “K. patula,” and “Uppermost K. walliseri range” biozones be suspended for the upper Sheinwoodian in favor of the Upper K. walliseri Superzone, due to the co-occurrence of K. patula with K. ortus
C. lundgreni
C. lapworthi
C. insectus
C. centrifugus
C. murchisoni
M. firmus
M. riccartonensis
C. lundgreni
Tely
chia
n
Llan
dove
ryW
enlo
ck
She
inw
oodi
anH
om.
SeriesStage Graptolite
zones Selected graptolite ranges
O. spiralis
Mcl. crenulata
Mcl. griestoniensis
Str. crispusStr. crispus
Str. sartorius Str. sartorius
Mcl. grieston-iensis
Sp. turriculatus
Mcl. crenulata
O. spiralis
C. lapworthi
C. insectus
C. centrifugus
C. murchisoni
M. firmus
M. riccartonensis
‘mid-Wenlock’ C. rigidusM. flexilis
‘M.’ antennularius
Sti. vesiculosusStr. wimani
P. dubius
Figure 2. Range chart of selected graptolite species (after Loydell, 1998) used to defi ne the graptolite biozonation utilized herein. Biozones are shown here as equal duration for illustrative purposes only. Abbreviations: Sti.—Stimulograptus; Str.—Streptograptus; Sp.—Spirograptus; Mcl.—Monoclimacis; O.—Oktavites; C.—Cyrtograptus; M.—Monograptus; P.—Pristiograptus.

Testing the limits of Paleozoic chronostratigraphy
Geological Society of America Bulletin, September/October 2010 1703
ortus in southern Laurentia (Oklahoma, Clarita Formation, Haragan Creek Section, Sample #5, Barrick and Klapper, 1976). If the presence of K. patula in a latest Sheinwoodian fauna con-taining K. ortus ortus is not the result of re-working, contamination, or sampling across an unconformity, this co-occurrence would re-quire a range extension to K. patula and make the “Uppermost K. walliseri range” an invalid zone (cf. Jeppsson, 1997). Although the FAD of K. patula remains unchanged by the informa-tion above, indicating that the Middle K. wal-liseri Zone may still be a valid zone; due to its exceptional rarity in most regions, the fact that it has no known lineage prior to the late Shein-woodian, and until the range of K. patula can be studied in more detail, the use of the Upper
K. walliseri Superzone is suggested for the in-terval between the LAD of Ozarkodina sagitta rhenana and the LAD of K. walliseri (Fig. 3).
Detailed comparisons of the sequence stra-tigraphy, carbon isotope stratigraphy, and cono-dont biostratigraphy of North America with the classi cal area of Gotland, Sweden, have produced an important observation regard-ing the paleobiogeographic emergence of the cono dont Kockelella walliseri (Jeppsson, 1997; McLaughlin et al., 2008). On Gotland, the fi rst occurrence of K. walliseri is within the basal Slite Formation (Jeppsson, 1997, 2008; Sequence VII of McLaughlin et al., 2008; see below) where δ13Ccarb values are below +2.0‰ (Samtleben et al., 1996; Calner et al., 2004), whereas in parts of North America K. walliseri fi rst occurs
where δ13Ccarb values are still >+5.0‰ (Cramer et al., 2006a [conodont zonation modifi ed from Kleffner , 1991]; Sequence VI of McLaughlin et al., 2008; Bancroft, 2008) prior to deposition of the Slite Formation on Gotland, thus indicating that we are likely seeing a paleobiogeographic difference in the fi rst occurrence of K. walliseri.
Even older occurrences of K. walliseri have been reported from North America (co-occurring with Pterospathodus within Sequence IV) by Helfrich (1980), Over and Chatterton (1987), and Uyeno (1990), but until these collec-tions can be restudied in detail to eliminate concerns about identifi cation, reworking of Pterospathodus into younger strata containing K. walliseri, and laboratory contamination, or be recollected using modern sampling methods,
K. patula
O. sagitta rhenana
Pt. p. procerus
Pt. am.amorpho-
gnathoides
D. stauro-gnathoides
K. walliseri
O. martinssoni
O. sagittasagitta K. ortus
ortus
O. p.polinclinata
O. para-confluens
Pt. am. amorpho-gnathoides Zone
Lower Ps.bicornis Zone
Upper Ps.bicornis Zone
Lower Pt.procerus Zone
Upper Pt.procerus Zone
Lower K.ranuliformis Zone
Upper K.ranuliformis Zone
O. sagittarhenana Zone
Lower K.walliseri Zone
Middle K.walliseri Zone
K. patula Zone
Uppermost K.walliseri range
post K. walliseriinterregnum
K. ortusortus Zone
O. sagittasagitta Zone
Pt. am.amorpho-
gnathoidesZonalGroup
Pt. p.procerus
SuperZoneUpperPtero-
spatho-dus
ZonalGroup
K.ranuliformisSuperZone
O. sagittarhenana
SuperZone
UpperK. walliseriSuperZone
K. walliseriZonalGroup
K. ortusortus
SuperZone
Tely
chia
n
Llan
dove
ryW
enlo
ck
She
inw
oodi
anH
om.
1
2
3
4
6
8
SeriesStage
Conodontsuperzones and
Zonal groups
Conodontzones Selected conodont ranges
?
?Pt. am.
lithuanicus ZonePt. am.
lennarti ZonePt. am.
angulatus ZonePt. eopennatus ssp. n. 2 Zone
Pt. eopennatus ssp. n. 1 Zone
LowerPtero-
spatho-dus
ZonalGroup
Pt. celloniSuper-Zone
Pt.eopennatusSuperZone
Pt. am. lithuanicus
Pt. am. lennarti
Pt. am. angulatus
Pt. eop. ssp. n. 2
Pt. eop. ssp. n. 1
Pt.celloni O. p.
estonica
K. ranuli-formis
?
Au.kuenhi
N. sensitiva
?
Aps.ruginosus
Aps.walmsleyi
Panderodussp. nov. N
Panderoduspanderi
Ps. linguicornis
Panderoduspanderi
Figure 3. Range chart of selected conodont species used to defi ne the conodont biozonation utilized herein; modifi ed from Jeppsson (1997) and Männik (1998, 2007a) to account for the co-occurrence of Kockelella patula with K. ortus ortus in southern Laurentia (see text). Biozones are shown here as equal duration for illustrative purposes only. Abbreviations: Pt.—Pterospathodus; N.—Nudibelodina; eop.—eopennatus; Au.—Aulacognathus; O. p.—Ozarkodina polinclinata; Pt. p.—Pterospathodus pennatus; am.—amorphognathoides; Aps.—Apsidognathus; K.—Kockelella; D.—Distomodus; Ps.—Pseudooneotodus.

Cramer et al.
1704 Geological Society of America Bulletin, September/October 2010
these reports remain diffi cult to include in the current biostratigraphic zonation. Reports of K. walliseri in the literature are problematic in that some specimens identifi ed as K. walliseri are instead K. ranuliformis and whether or not these two species represent a lineage remains an open question (see discussion in Jeppsson, 1997). In all recent studies conducted by Peep Männik, Lennart Jeppsson, Mark Kleffner, and Jim Barrick, including the sections investigated here, K. walliseri fi rst occurs in the presence of O. s. rhenana, which limits the range of K. walliseri to the middle-upper Sheinwoodian as used in the present zonation (Fig. 3). Further biochemostratigraphy needs to be done to con-fi rm these fi ndings, and until then, care should be taken when utilizing the Lower K. walliseri Biozone (base of zone = the co-occurrence of K. walliseri with O. sagitta rhenana).
The datum points of Jeppsson (1997) utilized in the text are: Datum 1—LAD Nudibelodina sensitiva; Datum 2—LAD Ozarkodina polin-clinata polinclinata and/or Apsidognathus (i.e., A. walmsleyi and A. ruginosus); Datum 3—LAD Pterospathodus amorphognathoides amor-phognathoides and/or Panderodus sp. nov. N, whichever is stratigraphically higher (see Männik , 2007a, 2007b); Datum 6—LAD Ptero-spathodus pennatus procerus. Throughout this paper, when conodonts have been used to cor-relate the position of the base of the Wenlock Series, we have used Datum 2, the last appear-ance of O. polinclinata polinclinata, and/or Apsidognathus.
Telychian–Homerian Carbon Isotope (δ13Ccarb) Stratigraphy
Two positive δ13Ccarb excursions are recorded in the Wenlock Series (e.g., Talent et al., 1993); one in the Sheinwoodian Stage (Samtleben et al., 1996; Wenzel and Joachimski, 1996; Kaljo et al., 1997, 2007; Munnecke et al., 2003), with an onset correlating to a position within the Ireviken Event as defi ned by Jeppsson (1987, 1997) and another in the Homerian Stage (Corfi eld et al., 1992; Samtleben et al., 2000; Porębska et al., 2004; Cramer et al., 2006b), with an onset correlating to a position within the Mulde Event as defi ned by Jeppsson (1997) and Jeppsson et al. (2006) immediately follow-ing the bulk of the conodont extinctions and coincident with “der Grossen Krise” (The Great Crisis; Jaeger, 1959), the most signifi cant grap-tolite extinction event during the Silurian. These chronostratigraphic markers make the Wenlock Series an ideal candidate for high-resolution chronostratigraphic study due to the presence of two protracted positive carbonate carbon (δ13Ccarb) isotope excursions during an estimated
six million years (GTS2004, Gradstein et al., 2004). These δ13Ccarb excursions have been shown to be valuable tools for global chrono-strati graphic correlation, particularly when combined with the equally high-resolution biostratigraphy of conodonts and graptolites (Kaljo et al., 1998, 2003; Munnecke et al., 2003; Calner et al., 2004; Porębska et al., 2004; Cramer et al., 2006a, 2006b; Kaljo and Martma , 2006). The early Sheinwoodian (Ireviken) δ13Ccarb excursion is the best documented of the Silurian excursions and has now been identifi ed in more than 20 localities worldwide (Talent et al., 1993; Samtleben et al., 1996; Wenzel, 1997; Kaljo et al., 1997, 1998, 2003, 2007; Saltzman, 2001; Munnecke et al., 2003; Martma et al., 2005; Noble et al., 2005; Cramer et al., 2006a, 2006c; Kaljo and Martma, 2006; Cramer and Saltzman, 2007) spanning the paleocontinents of Lau rentia, Avalonia, Baltica, and Gondwana.
At present, there is no standard defi nition of the beginning or end of an isotope excursion. Herein, we defi ne the onset of the excursion as the infl ection point immediately prior to the onset of the maximum positive rate of change in δ13Ccarb that defi nes the increasing limb of the excursion, not necessarily the onset of in-creasing isotope values. Likewise, we defi ne the end of the excursion as the point where δ13Ccarb values are consistently at or below the values immediately prior to the onset as defi ned above. Defi ning an excursion in this manner potentially allows the practical ap-plication of such terms to any excursion, but individual isotopic records will always be in-fl uenced by the continuity, completeness, and diagenetic history of the given section, all of which can potentially obscure the positions defi ned above. An open circle is used in all data fi gures (Figs. 4–8) to demonstrate the position we have determined to be the onset of the early Sheinwoodian (Ireviken ) δ13Ccarb excursion. All data tables are available in the GSA Data Repository (Tables DR1–DR91).
METHODS
The Banwy River section was analyzed for δ13Ccarb stratigraphy using samples collected by Loydell and Cave (1996), together with additional samples collected by DKL in June, 2006 (see Loydell and Frýda, 2007), which permits direct correlation between the δ13Ccarb data and the generally excellent graptolite bio-
stratigraphic control of this section. Samples were cut to remove any weathered surfaces, polished to expose the area to be sampled, and microdrilled to generate a few decigrams of powder with a preference for fi ne-grained carbonate (Saltzman, 2002). In the Aizpute-41 core, biostratigraphically useful early Silurian conodonts and graptolites occur together in abundance, and they permit the biostratigraphic positions of the isotope samples to be well constrained (Loydell et al., 2003). Carbonate samples from the Aizpute-41, Ohesaare, Viki, and Ruhnu cores were prepared and analyzed using the methods as described in Kaljo et al. (1997). Brachiopod samples from Gotland, Sweden, were prepared and analyzed as de-scribed in Samtleben et al. (2001). Gotland is remarkable in that it is suffi ciently fossiliferous to allow brachiopods to be used for semicon-tinuous, high-resolution chemostratigraphy. Carbonate samples from Gotland come directly from the conodont samples used to construct the Jeppsson (1997) and Jeppsson et al. (2006) conodont biozonations providing exceptional biostratigraphic control.
All values are reported in per mil notation rela-tive to the Vienna Pee Dee belemnite (VPDB) standard by assigning a δ13Ccarb value of +1.95‰ and a δ18O value of –2.20‰ to National Bureau of Standards (NBS)-19. Reproducibility was checked by replicate analysis of laboratory standards and is better than ±0.10‰ for δ13Ccarb and ±0.10‰ for δ18O of all samples analyzed from all laboratories. Carbonate samples from the Banwy River section were prepared by Brad Cramer and analyzed by Michael Joachimski at the University of Erlangen, Germany, and Craig Johnson at the U.S. Geological Survey, Denver, Colorado, USA. Carbonate samples from Gotland were prepared by Brad Cramer and analyzed by Tõnu Martma at the Tallinn University of Technology, Estonia. Brachio-pod samples from Gotland were prepared by Christian Samtleben and Axel Munnecke and were analyzed by Torsten Bickert at the Univer-sity of Bremen, Germany. Samples from New York, USA, were prepared by Brad Cramer and analyzed by Tim Prokopiuk at the University of Saskatchewan, Saskatoon, Canada. Carbonate samples from the East Baltic cores were col-lected, prepared, and analyzed by Dimitri Kaljo, Peep Männik, and Tõnu Martma at the Tallinn University of Technology, Estonia.
RESULTS
Banwy River, Wales
Data from the Banwy River section are shown (Fig. 4) with the organic carbon isotope data of Loydell and Frýda (2007). Stable isotope
1GSA Data Repository item 2010130, Telychian–Sheinwoodian biochemostratigraphy and Swedish National Grid reference data, is available at http://www.geosociety.org/pubs/ft2010.htm or by request to [email protected].

Testing the limits of Paleozoic chronostratigraphy
Geological Society of America Bulletin, September/October 2010 1705
S
120
100
X
140
130
110
+3
+7
+12+14
+27+28.5
+5
+8, 8.5
+12+14+15.5
+18
–24.5, –24–23
+22.5, 23, 23.5
–19
O.s
pira
lisC
. lap
w.
C. i
nsec
tus
Tara
nnon
Sha
les
For
mat
ion
180
170
160
150 –14–16
–7
–2, –1.5–1, –0.5C+2+3
+4, 4.5, 5, 5.5, 6, 6.5, 7
+11.75+12.25+13, 13.5+14.5+15+16, 16.5, 17
+30
190
200
210
220
230
240
250
C. c
entr
ifugu
sC
. mur
ch.
M. f
irm.
M. r
icca
rton
ensi
s
Nan
t-ys
gollo
n S
hale
s F
orm
atio
nB
anw
y B
urro
wed
Mem
ber
Llan
dove
ryW
enlo
ck
δ13Ccarb (‰VPDB)
0
+47
+59 (C. rigidus)
+79 (M. flexilis)
+23
Banwy River, Wales, United Kingdom
Tely
chia
nS
hein
woo
dian
Series StageLitho-Strat.
GraptoliteZone m
δ13Corg (‰)
–1–2–3–4–5–6–7 –27.0–28.0–29.0–30.0 0.80.60.40.20
TOC (%C)
1
2
3
g
Laminated hemipelagite
Bioturbated turbiditic mud-stones and graptolitic shalesPale green and red mud-stones and graptolitic shales
Figure 4. Carbonate carbon (δ13Ccarb) isotope data from the Banwy River section, Wales, combined with graptolite data from Loydell and Cave (1996) and organic carbon isotope data from Loydell and Frýda (2007). The italic lower-case “g” bisected by a thin gray line repre-sents the base of the Wenlock Series as defi ned in this section upon the fi rst occurrence of the graptolite species C. murchisoni. See text for details regarding the numbered gray intervals (1, 2, and 3). Numbers and arrows to the right of the stratigraphic column refer to graptolite samples from Loydell and Cave (1996). The sole occurrences of the graptolites M. fl exilis and C. rigidus are shown. VPDB—Vienna Pee Dee belemnite; TOC—total organic carbon.

Cramer et al.
1706 Geological Society of America Bulletin, September/October 2010
data (δ13Ccarb and δ18O) are directly correlated (Table DR1 [see footnote 1]) to the graptolite biostratigraphy of Loydell and Cave (1996). The δ13Ccarb values are low at the base of the section (–7.0‰) and gradually increase to an initial high of –0.48‰ within the Monograptus fi rmus Biozone. A decline of roughly –2.0‰ oc-curred low in the M. riccartonensis Biozone be-fore recovering to values >–0.5‰ higher in the M. riccartonensis Biozone. The highest values recovered in the section, +0.09‰ and +0.24‰, were recorded within the “mid-Wenlock” grap-tolite interval of Loydell (1998) at 228 and 231 m, respectively. Carbonate carbon isotope
values decline at the top of the section as values fall below –1.0‰.
Banwy River δ13Ccarb values are generally low, even in the lower Sheinwoodian (Ireviken) excursion, perhaps because it represents one of the deepest marine (although still only shelf edge) and complete sections yet recovered from this interval. Diminished absolute values of δ13Ccarb away from Silurian epicontinental shelves toward basinal sections have been dis-cussed previously (Kaljo et al., 1998; Mun-necke et al., 2003; Melchin and Holmden, 2006; Loydell, 2007, 2008b; Cramer and Munnecke, 2008). The most analogous Telychian strata yet
investigated for δ13Ccarb are the graptolitic shales of Cornwallis Island, Canada, of Melchin and Holmden (2006) which show average lower Telychian (Spirograptus guerichi and Sp. tur-riculatus graptolite biozones) δ13Ccarb values of –5.0‰. The lowermost samples from the Banwy River section show similar low δ13Ccarb values in the Oktavites spiralis Biozone.
Low δ13Ccarb values are often taken as indica-tors of meteoric diagenetic overprinting of the primary marine δ13Ccarb signal (e.g., Algeo et al., 1992). The infl uence of late-stage, postdeposi-tional diagenesis is visible when the isotopic values of carbon and oxygen are plotted against each other (Banner and Hanson, 1990) due to the fact that meteoric diagenesis typically pro-duces low δ18O values as well. Therefore, a posi-tive correlation between low δ13Ccarb values and low δ18O values is an indicator of diagenetic re-setting of original δ13Ccarb values. Carbonate car-bon and oxygen isotope values from the Banwy River section were plotted against each other (Fig. DR1 [see footnote 1]) and show no such correlation. In fact, the lowest δ13Ccarb values are typically associated with the highest δ18O values, which provide further evidence that the δ13Ccarb signal recorded from the Banwy River section were not altered by meteoric diagenesis. The fact that this section contains graptolitic shales and mudstones suggests that these strata were more enriched in organic carbon than they are at present (Fig. 4). Remineralization of organic carbon and therefore contribution of isotopically light carbon during the diagenetic stabilization and/or cementation of the carbon-ates likely affected the δ13Ccarb values of the lower part of the section (Scholle and Arthur, 1980; Weissert et al., 2008).
The data from the Banwy River section do not show a clear infl ection point but instead show a more gradual increase during the upper Telychian. As a result, the onset of the early Sheinwoodian (Ireviken) δ13Ccarb excursion re-corded in this section remains diffi cult to de-fi ne. Prior to this study, the early Sheinwoodian (Ireviken ) δ13Ccarb excursion had been recorded in graptolite-bearing strata from the Ohesaare and Ruhnu cores, Estonia, and suggested a placement for the onset somewhere near the top of the C. murchisoni graptolite Biozone (Kaljo et al., 2007). Both the δ13Corg and total organic carbon (TOC) data of Loydell and Frýda (2007) show a major infl ection point at this level (gray band #1 in Fig. 4). This likely represents the on-set of the early Sheinwoodian (Ireviken) δ13Ccarb excursion (open circle in Fig. 4).
It is extremely important to note, however, that rarely are records of δ13Ccarb and δ13Corg excursions precisely synchronous and occasion-ally show opposing trends (Patzkowsky et al.,
Aizpute-41 Core, LatviaSeriesConodont
ZoneGraptolite
Zone mL
lan
do
very
Jurm
ala
For
mat
ion
960
950
940
Pt. eop.ssp. n. 2
930
920
910
Pt. am.ang-
ulatusMcl.
crenulata
O.spiralis
C.lapworthi
Pt.
amor
ph.
amor
phog
nath
oide
sZ
.G.
Mcl. griestoniensis
C. m
urch
ison
i
M. firmus
M. riccart-onensis
proc. S.Z.
K. ran.S.Z.W
enlo
ckP
t. ce
lloni
S.Z
.3
1
g
Str. crispusStr. sartorius
Pt. e. ssp. n. 1
+2+10–1 +4+3
δ13Ccarb (‰ VPDB)
Panderodussp. nov. N
Pt. am.amorph.
Green, gray and brown marlstoneswith calcareous marlstones
Greenish and brownish gray marlstones
Red-bedsGreenish gray claystones
Rig
a F
orm
atio
n
6
Aps.sp.
N. sens.+
K. ranuliformis
Figure 5. Carbonate carbon (δ13Ccarb) isotope data from the Aizpute-41 core, Latvia, combined with biostratigraphic data from Loydell et al. (2003). The italic lower-case “g” bisected by a thin gray line represents the base of the Wenlock Series as defi ned by the fi rst appearance datum of C. murchisoni. The last appearance datum (LAD) of Nudibelodina sensitiva and LAD of Apsidognathus sp. are shown to the right of the stratigraphic column but are un-likely to be the true LADs of these species (see text). This is the only section known to contain both conodont and graptolite species used to defi ne the base of the Wenlock Series, but as described in the text, they co-occur immediately above a suspected unconformity that removes more than two graptolite zones (upper part of the C. lapworthi, the C. insectus, and the C. centrifugus biozones). See text for details regarding the numbered gray interval. LADs of Pt. am. amorphognathoides and Panderodus sp. nov. N, and the level of (re)appearance of Kockelella ranuliformis are shown. The onset of the early Sheinwoodian (Ireviken) δ13Ccarb excursion (open circle) occurs in this core below Datum 3 and above Datum 1 of the Ireviken Event. VPDB—Vienna Pee Dee belemnite.

Testing the limits of Paleozoic chronostratigraphy
Geological Society of America Bulletin, September/October 2010 1707
1997; Cramer and Saltzman, 2007; Young et al., 2008) meaning that determination of the timing of the onset of a δ13Ccarb excursion via δ13Corg data (or vice versa) should be avoided when-ever possible. Here, the δ13Ccarb record from the Banwy River section did not provide suffi cient evidence to place the onset of the excursion, and we were forced to make the determination based upon the organic data alone. Without prior confi rmation that the δ13Ccarb excursion be-gan within the C. murchisoni graptolite Biozone (Kaljo et al., 2007; see discussion in Loydell and Frýda, 2007), we would not have had suffi -cient justifi cation to determine the placement of the onset of the early Sheinwoodian (Ireviken) δ13Ccarb excursion within this section.
Aizpute-41, Latvia
Carbonate carbon isotope (δ13Ccarb) data from the Aizpute-41 core, Latvia, are shown in Fig-ure 5 and Table DR2 (see footnote 1) together with graptolite and conodont data from Loydell et al. (2003). This represents one of the most intensively studied Llandovery to lower Wen-lock sections worldwide. The δ13Ccarb data show a gentle increase of +0.75‰ (from +0.50‰ to roughly +1.25‰) over the lower 40 m sampled before a rapid increase of >+1‰ (from +1.34‰ to +2.36‰) during the uppermost fi ve meters of strata, signaling the onset of the early Shein-woodian (Ireviken) positive carbon isotope ex-cursion. The onset of the excursion (open circle in Fig. 5) in this core occurs above Datum 1 and below Datum 3 (Fig. 5; Table DR2 [see footnote 1]).
At least one signifi cant break in deposition is likely present in the Aizpute-41 core, and it is most easily recognized by graptolite biostra-tigraphy. Just below the 930 m mark, grapto-lite biostratigraphy suggests the presence of an unconformity (Loydell et al., 2003): the upper part of the C. lapworthi, the C. insectus, and the C. centrifugus graptolite biozones are not rep-resented (Fig. 5; Table DR2 [see footnote 1]). Immediately superjacent strata contain both the fi rst appearance of the graptolite C. murchisoni as well as the last appearance of the conodont Apsidognathus sp. making this one of the only localities known to contain both conodont and graptolite guide fossils necessary to correlate the base of the Wenlock Series. Unfortunately, the two species co-occur in strata above a prob-able unconformity, which means the fi rst ap-pearance of C. murchisoni in this section may not represent the true base of the C. murchisoni graptolite Biozone. Similarly, the conodont Apsidognathus sp. is rare in the Aizpute-41 core, and the highest recovery of that species in this core evidently does not indicate the
–10
0
10
20
30
40
50
m
+4+3+2+10 +6+5
δ13Ccarb (‰ VPDB)
Lland.
Wenlock
Series
GraptoliteZone
L. V
isby
ConodontZone
3
6
Pt.
am
.am
orph
. Z.G
.K
. ran
uli-
form
is S
.Z.
O. s
agitt
a rh
enan
aL.
K.
wal
l.
Tofta
Han
gvar
Slit
eM
. fir.
/ric
cart
.
U. V
isby
Hög
klin
t
Gotland, SwedenP
t. p.
proc
.
1
2
3
c
Limestone-marlalternationsReef, flank, and coarsecrinoidal grainstones
Crinoidal limestone
–1–2
60
Mid
dle
K.
wal
liser
i
Micritic sample
Brachiopod sample
Figure 6. Carbonate carbon isotope (δ13Ccarb) data from Gotland, Sweden, combined with biostratigraphic data from Jeppsson (1997, 2008), and Jeppsson et al. (2006). The black line represents mean values from brachiopods (Table DR3 [see footnote 1], from Samtleben et al., 1996, 2000; Munnecke et al., 2003; and new data). The gray line represents values from selected carbonate components and micritic rocks (Table DR4 [see footnote 1]). Each brachiopod data point represents the average of three or more brachiopod sam-ples. The italic lower-case “c” bisected by a thin gray line represents the base of the Wenlock Series as defi ned by the last appearance datum of O. polinclinata polinclinata. This composite fi gure is highly synthetic in its representation of unit thickness because the thick-ness of each stratigraphic unit varies across the island and data were collected from over 50 localities with a maximum distance of over 60 km. We have, however, used a formational contact as a reference point (Lower to Upper Visby contact), and the thicknesses refer to a standardized section (see text for details). The onset of the early Sheinwoodian (Ireviken) δ13Ccarb excursion (open circle) occurs in this composite between Datum 3 and Datum 6 of the Ireviken Event (precisely at Datum 4). See text for details regarding the numbered gray intervals. VPDB—Vienna Pee Dee belemnite.
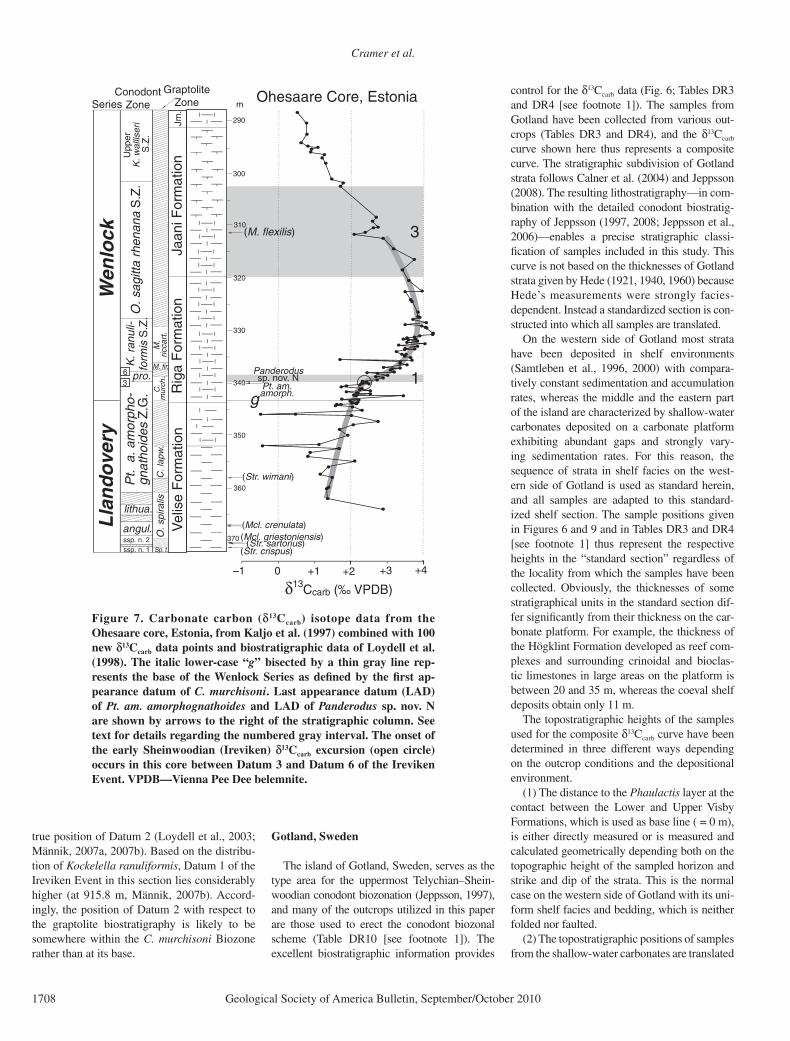
Cramer et al.
1708 Geological Society of America Bulletin, September/October 2010
true position of Datum 2 (Loydell et al., 2003; Männik , 2007a, 2007b). Based on the distribu-tion of Kockelella ranuliformis, Datum 1 of the Ireviken Event in this section lies considerably higher (at 915.8 m, Männik, 2007b). Accord-ingly, the position of Datum 2 with respect to the graptolite biostratigraphy is likely to be somewhere within the C. murchisoni Biozone rather than at its base.
Gotland, Sweden
The island of Gotland, Sweden, serves as the type area for the uppermost Telychian–Shein-woodian conodont biozonation (Jeppsson, 1997), and many of the outcrops utilized in this paper are those used to erect the conodont biozonal scheme (Table DR10 [see footnote 1]). The excellent biostratigraphic information provides
control for the δ13Ccarb data (Fig. 6; Tables DR3 and DR4 [see footnote 1]). The samples from Gotland have been collected from various out-crops (Tables DR3 and DR4), and the δ13Ccarb curve shown here thus represents a composite curve. The stratigraphic subdivision of Gotland strata follows Calner et al. (2004) and Jeppsson (2008). The resulting lithostratigraphy—in com-bination with the detailed conodont biostratig-raphy of Jeppsson (1997, 2008; Jeppsson et al., 2006)—enables a precise stratigraphic classi-fi cation of samples included in this study. This curve is not based on the thicknesses of Gotland strata given by Hede (1921, 1940, 1960) because Hede’s measurements were strongly facies-dependent . Instead a standardized section is con-structed into which all samples are translated.
On the western side of Gotland most strata have been deposited in shelf environments (Samtleben et al., 1996, 2000) with compara-tively constant sedimentation and accumulation rates, whereas the middle and the eastern part of the island are characterized by shallow-water carbonates deposited on a carbonate platform exhibiting abundant gaps and strongly vary-ing sedimentation rates. For this reason, the sequence of strata in shelf facies on the west-ern side of Gotland is used as standard herein, and all samples are adapted to this standard-ized shelf section. The sample positions given in Figures 6 and 9 and in Tables DR3 and DR4 [see footnote 1] thus represent the respective heights in the “standard section” regardless of the locality from which the samples have been collected. Obviously, the thicknesses of some stratigraphical units in the standard section dif-fer signifi cantly from their thickness on the car-bonate platform. For example, the thickness of the Högklint Formation developed as reef com-plexes and surrounding crinoidal and bioclas-tic limestones in large areas on the platform is between 20 and 35 m, whereas the coeval shelf deposits obtain only 11 m.
The topostratigraphic heights of the samples used for the composite δ13Ccarb curve have been determined in three different ways depending on the outcrop conditions and the depositional environment.
(1) The distance to the Phaulactis layer at the contact between the Lower and Upper Visby Formations, which is used as base line ( = 0 m), is either directly measured or is measured and calculated geometrically depending both on the topographic height of the sampled horizon and strike and dip of the strata. This is the normal case on the western side of Gotland with its uni-form shelf facies and bedding, which is neither folded nor faulted.
(2) The topostratigraphic positions of samples from the shallow-water carbonates are translated
+4+3+2+10–1
O. s
pira
lisC
. lap
w.
C.
mur
ch.
M.
ricca
rt.
Rig
a F
orm
atio
n
Lla
nd
ove
ryW
enlo
ckSeries
GraptoliteZone
M. fir.
angul.
lithua.
Pt.
a. a
mor
pho-
gnat
hoid
es Z
.G.
Vel
ise
For
mat
ion
mConodont
Zone
370
360
350
340
330
320
36
δ13Ccarb (‰ VPDB)
K. r
anul
i-fo
rmis
S.Z
.
ssp. n. 2
Ohesaare Core, Estonia
Jaan
i For
mat
ion
310
300
pro.
ssp. n. 1
1
3
g
(M. flexilis)
(Mcl. crenulata)
(Str. crispus)
(Mcl. griestoniensis)(Str. sartorius)
Sp. t.
(Str. wimani)
O. s
agitt
a rh
enan
a S
.Z.
Pt. am.amorph.
Panderodussp. nov. N
290Jm.
Upp
erK
. wal
liser
i S
.Z.
Figure 7. Carbonate carbon (δ13Ccarb) isotope data from the Ohesaare core, Estonia, from Kaljo et al. (1997) combined with 100 new δ13Ccarb data points and biostratigraphic data of Loydell et al. (1998). The italic lower-case “g” bisected by a thin gray line rep-resents the base of the Wenlock Series as defi ned by the fi rst ap-pearance datum of C. murchisoni. Last appearance datum (LAD) of Pt. am. amorphognathoides and LAD of Panderodus sp. nov. N are shown by arrows to the right of the stratigraphic column. See text for details regarding the numbered gray interval. The onset of the early Sheinwoodian (Ireviken) δ13Ccarb excursion (open circle) occurs in this core between Datum 3 and Datum 6 of the Ireviken Event. VPDB—Vienna Pee Dee belemnite.

Testing the limits of Paleozoic chronostratigraphy
Geological Society of America Bulletin, September/October 2010 1709
B
C
410
400
390
380
370
m 480
470
460
450
440
430
420
+4
+3
+2
+1
0+
6+
5
Jaani Formation Velise Formation
O.s
agitt
arh
enan
a
Pt. a. amorpho-gnathoides Z.G.
proc
.
K.walliseri Z.G.
Pt.
a. li
th.
Pt.
a.an
g.
6 3Wenlock Llandovery
Ruh
nu C
ore,
Est
onia
S
erie
sCon
odon
tZ
one
13
g
Pt.
a. le
n.
K. ranuli-formis S.Z.
eop.
s.z
.
Riga Fm.Jamaja Formation
M. f
irm.
M. ric-carton-ensis
C.
mur
ch-
ison
i
Mcl
. gri.O.
spiralis
Gra
ptol
iteZ
one
Pt.
eop.
ssn
. 1P
t. eo
p. s
sn. 2
(Sti.
ves
icul
osus
)
Pan
dero
dus
sp. n
ov. N
Pt.
am.
amor
ph.
D
+
+4
+3
+2
+1
0–1
O. spiralisC. lapw.C.murch.
M.riccart.
Riga Formation
LlandoveryWenlock
Ser
ies
Gra
ptol
iteZ
one
M. f
ir.
angu
l.
lithu
a.
Pt. a. amorpho-gnathoides Z.G.
Velise Formation
mC
onod
ont
Zon
e
37036
0
350
340
330
320
36
δ13C
carb
(‰
VP
DB
)
K. ranuli-formis S.Z.
ssp.
n. 2
Ohe
saar
e C
ore,
Est
onia
Jaani Formation
310
300
pro.
ssp.
n. 1
13
g(M. f
lexi
lis)
(Mcl
. cre
nula
ta)
(Str.
cris
pus)
(Mcl
. grie
ston
iens
is)
(Str.
sar
toriu
s)S
p. t.
(Str.
wim
ani)
O. sagitta rhenana S.Z.
Pt.
am.
amor
ph.
Pan
dero
dus
sp. n
ov. N
290
Jm.
UpperK. walliseri
S.Z.
Aiz
pute
-41
Cor
e, L
atvi
aS
erie
sCon
odon
tZ
one
Gra
ptol
iteZ
one
m
Llandovery
Jurmala Formation
960
950
940
Pt. eop.ssp. n. 2
930
920
910
Pt. am.angul.
Mcl.crenul.
O.spiralis
C.lapworthi
Pt. amorph. amorpho-gnathoides Z.G. M
cl. g
rie.C. murchisoni
M. f
irm.
M.
ricc.
proc
.
K. r
an.
WenlockPt. celloni S.Z.
31
g
Str.
sart
oriu
s
ssp.
n. 1
+2
+1
0–1
+4
+3
δ13C
carb
(‰
VP
DB
)
δ13C
carb
(‰
VP
DB
)
Pan
dero
dus
sp. n
ov. N
Pt.
am.
amor
ph.
Riga Formation
6
Aps
.sp
.
N. s
ens.
Str.
cris
pus
A
100
90807060m 180
170
160
150
140
130
120
110
+4
+3
+2
+1
0+
6+
5
δ13C
carb
(‰
VP
DB
)
Jaani Formation Velise Formation
Pt. a.amorphognathoides Z.G.
proc
.
K.ranuliformis
S.Z.
Pt.
a.lit
h. Pt. a.angulatus
Pt. eop.ssn. 2
6 3Wenlock Llandovery
Vik
i Cor
e, E
ston
ia
Ser
iesC
onod
ont
Zon
e
13
c
O. sagittarhenana S.Z.
K.
wal
liser
i
Pt.
am. a
mor
ph.
Pan
dero
dus
sp. n
ov. N
+
Mustjala MemberNinase
Mbr.Jaagarahu Formation
Upper K.walliseri
S.Z.
Fin
e gr
aine
d do
lom
itic
grai
nsto
nes
Blu
eish
-gra
y do
lom
itic
mar
lsto
nes
+
K. r
anul
iform
is
Fig
ure
8. S
ynop
tic
pres
enta
tion
of
carb
onat
e ca
rbon
(δ13
Cca
rb)
isot
ope
data
fro
m t
he (
A)
Vik
i; (
B)
Ohe
saar
e; (
C)
Ruh
nu;
and
(D) A
izpu
te-4
1 co
res
usin
g id
enti
cal m
etri
c an
d is
otop
ic s
cale
s fo
r ea
ch s
ecti
on s
how
n (T
able
s D
R2
and
DR
4–D
R7
[see
fo
otno
te 1
]). T
he it
alic
low
er-c
ase
“g”
repr
esen
ts t
he b
ase
of t
he W
enlo
ck S
erie
s as
defi
ned
by
the
fi rst
app
eara
nce
datu
m o
f C
. mur
chis
oni.
The
ital
ic lo
wer
-cas
e “c
” re
pres
ents
the
bas
e of
the
Wen
lock
Ser
ies
as d
efi n
ed b
y th
e la
st a
ppea
ranc
e da
tum
(L
AD
) of
O. p
olin
clin
ata
polin
clin
ata
in t
he V
iki c
ore.
(A
) C
onod
ont
data
fro
m t
he V
iki c
ore,
Est
onia
, aft
er M
änni
k (2
007a
).
Isot
ope
data
from
the
Vik
i cor
e af
ter
Kal
jo e
t al.
(200
3) a
nd n
ew d
ata
pres
ente
d he
rein
. The
LA
Ds
of P
t. am
. am
orph
ogna
thoi
des
and
Pan
dero
dus
sp. n
ov N
are
sho
wn
wit
h th
e so
le p
osit
ive
iden
tifi c
atio
n of
K. w
allis
eri.
The
ons
et o
f th
e ea
rly
Shei
nwoo
dian
(I
revi
ken)
δ13
Cca
rb e
xcur
sion
(op
en c
ircl
e) o
ccur
s in
thi
s co
re a
t D
atum
2 o
f th
e Ir
evik
en E
vent
. (B
) O
hesa
are
data
app
ear
as
pres
ente
d in
Fig
ure
7. (
C)
Bio
stra
tigr
aphi
c da
ta f
rom
the
Ruh
nu c
ore
afte
r P
õldv
ere
(200
3). I
soto
pe d
ata
from
the
Ruh
nu
core
fro
m K
aljo
et
al.
(199
8).
The
sol
e oc
curr
ence
s of
the
gra
ptol
ite
spec
ies
Stim
. ve
sicu
losu
s an
d th
e co
nodo
nt s
peci
es
Pt.
eope
nnat
us s
sp. n
ov. 1
, Pt.
eope
nnat
us s
sp. n
ov. 2
, and
Pt.
am. l
ithua
nicu
s ar
e sh
own.
The
LA
Ds
of P
t. am
. am
orph
ogna
thoi
des
and
Pan
dero
dus
sp. n
ov. N
are
als
o sh
own
wit
h th
e on
ly p
osit
ive
iden
tifi c
atio
n of
O. s
agitt
a rh
enan
a in
the
Ruh
nu c
ore.
The
on
set
of t
he e
arly
She
inw
oodi
an (
Irev
iken
) δ13
Cca
rb e
xcur
sion
(op
en c
ircl
e) o
ccur
s in
thi
s co
re b
elow
Dat
um 3
of
the
Irev
iken
E
vent
. (D
) Aiz
pute
-41
data
app
ear
as p
rese
nted
in F
igur
e 5.
All
litho
stra
tigr
aphi
c sy
mbo
ls a
re t
hose
uti
lized
in F
igur
e 5
un-
less
oth
erw
ise
indi
cate
d. V
PD
B—
Vie
nna
Pee
Dee
bel
emni
te.

Cramer et al.
1710 Geological Society of America Bulletin, September/October 2010
into the standard section by linear interpolation. For example, a sample taken 15 m above the base of a 30-m-thick formation on the carbonate platform is placed at 7.5 m above the base of the corresponding 15-m-thick shelf formation. Because of the strongly varying sediments on the platform this method bears some uncertain-ties, however, the isotope values from such sam-ples fi t well into the curve reconstructed by the “category-1” samples from shelf facies.
(3) As a result of the facies distribution on Gotland, the determination of the topostrati-graphic height of samples from the eastern part
of the island is more diffi cult. Here, the verti-cal distance from specifi c widespread marker beds, e.g., the Rhipidium tenuistriatum layer, is used. In some cases isolated samples had to be positioned solely due to their biostratigraphic placement within the conodont biozonation (Jeppsson, 1997; Jeppsson et al., 2006). The heights of these samples are given in Tables DR3 and DR4 [see footnote 1] and are marked with a double dagger (‡). Although the topo-strati graphic position of some of these samples may deviate in the order of meters, the relative succession of the samples is correct.
On Gotland, the onset of the early Shein-woodian (Ireviken) positive carbonate carbon isotope excursion has been precisely correlated to Datum 4 of the Ireviken Event by Mun-necke et al. (2003) and is shown by an open circle (Fig. 6). The Gotland δ13Ccarb record be-gins with values averaging +1.5‰ through the Lower Visby Formation and rapidly increasing to >+4.0‰ within the Upper Visby Formation. A decline > –1.0‰ is recorded within the lower portion of the Högklint Formation before once again reaching values >+5.0‰ at the top of the formation.
120
100
140
130
110
O. s
pira
lisC
. lap
wor
.C
. ins
ectu
s
Tara
nnon
Sha
les
For
mat
ion
180
170
160
150
190
200
210
220
230
240
250
C. c
entr
ifugu
sC
. mur
chis
oni
M. f
irm.
M. r
icca
rton
ensi
s
Nan
t-ys
gollo
n S
hale
s F
orm
atio
nB
anw
y B
urro
wed
Mem
ber
Lla
nd
ove
ryW
enlo
ck
δ13Ccarb (‰ VPDB)
0–1–2–3–4–5–6–7
SeriesLitho-Strat.
GraptoliteZone m Banwy River, Wales
1
2
3
g
(C. rigidus)
(M. flexilis)
A
+4+3+2+1 +6+5
δ13Ccarb (‰ VPDB)
Wen
lock
R.
Roc
hest
er
Decew
Gas
-po
rtG
oat
Isla
nd
P. a. a.
K. r
anul
i-fo
rmis
S.Z
.O
. s.
rhen
ana
Low
er K
.w
allis
eri
Llan. 0
10
20
30
40
m
Iron.
Niagara River Power ProjectS-1 Core, New York
SeriesConodont
Zone
3
21
+0
CCalcareous gray shale
Crinoidal and finegrained dolomite
Crinoidal limestone
Laminated and argill-aceous dolomite
B
–10
0
10
20
30
40
50
m
+4+3+2+10 +6+5
δ13Ccarb (‰ VPDB)
Lla
nd
.W
enlo
ck
Series
GraptoliteZone
L. V
isby
ConodontZone
3
6
Pt.
am
.am
orph
. Z.G
.K
. ran
uli-
form
is S
.Z.
O. s
agitt
a rh
enan
aL.
K.
wal
l.
Tofta
Han
gvar
Slit
eM
. fir.
/ric
cart
.
U. V
isby
Hog
klin
t
Gotland, Sweden
Pt.
p.pr
oc.
1
2
3
c
–1–2
60
Mid
dle
K.
wal
liser
i
Figure 9. Synoptic presentation of carbonate carbon (δ13Ccarb) isotope data from (A) Banwy River, Wales; (B) Gotland, Sweden; and (C) New York, USA, using identical metric and isotopic scales for each section shown (Tables DR1, DR3, and DR8 [see footnote 1]). The italic lower-case “g” bisected by a thin gray line represents the base of the Wenlock Series as defi ned by the fi rst appearance datum of C. murchisoni. The italic lower-case “c” represents the last appearance datum of O. polinclinata polinclinata. See text for details regarding the numbered gray intervals. (A) Banwy River data appear as presented in Figure 4. (B) Gotland data appear as presented in Figure 7. (C) Isotope and biostratigraphic data from New York after Cramer et al. (2006a). By virtue of an unconformity at the Rockway and Irondequoit contact, Datum 3, Datum 6, and the onset of the early Sheinwoodian (Ireviken) δ13Ccarb excursion (open circle) all occur at the same point in this section (at the Rockway Irondequoit contact). Note the difference in position of the base of the Lower K. walliseri Zone with respect to the early Sheinwoodian (Ireviken) δ13Ccarb excursion between Gotland and New York. VPDB—Vienna Pee Dee belemnite.

Testing the limits of Paleozoic chronostratigraphy
Geological Society of America Bulletin, September/October 2010 1711
There are two noticeable breaks in the iso-tope curve of Gotland (Fig. 6; Tables DR3 and DR4 [see footnote 1]); the fi rst is at the Högklint Formation–Tofta Formation contact; and the other at the Hangvar Formation–Slite Formation contact. Both of these lithostratigraphic contacts are disconformable (Hede, 1940; Eriksson and Calner, 2005; Jeppsson, 2008). The δ13Ccarb val-ues return to pre-excursion levels below +2.0‰ in the basal Slite Formation. Skoglund (1979) reported the occurrence of Monograptus cf. riccartonensis from the Högklint Formation (Fig. DR2 [see footnote 1]). Reexamination of this specimen (DKL) shows that it is a distal fragment of a Monograptus that may be either M. fi rmus or M. riccartonensis, but the incom-plete specimen does not allow identifi cation to species level.
Ohesaare-Viki-Ruhnu, Estonia
The Ohesaare core, Estonia, was studied in one of the fi rst attempts at a directly integrated, multiproxy, high-resolution biostratigraphy through the Llandovery and Wenlock Series (Loydell et al., 1998), and some of the carbon-ate carbon isotope data have been published previously (Kaljo et al., 1997). Here, 100 new data points are added to defi ne the onset of the early Sheinwoodian (Ireviken) δ13Ccarb excur-sion (Fig. 7; Tables DR5 and DR6 [see foot-note 1]). The Ohesaare δ13Ccarb record shows signifi cant instability during the late Telychian, suggesting that at least two disconformable sur-faces are likely present (cf. Algeo et al., 1992): one near the 352.00 m mark within the Velise Formation, and the other near the base of the Riga Formation (345.8 m). Both disconformi-ties are within the Pt. amorphognathoides amorphognathoides Zonal Group in the cono-dont biostratigraphy and between the last appearance of C. lap worthi and the fi rst ap-pearance of C. murchisoni in the graptolite biostratigraphy. This is consistent with a latest Llandovery age for the unconformity inferred by Loydell et al. (2003) from the Aizpute-41 core. The δ13Ccarb values increase in the lower-most Sheinwoodian and reach values >+4.0‰ within the Riga Formation but never exceed +5.0‰, before a long steady decline back to pre-excursion values within the Jaani Forma-tion. In the Ohesaare core, the onset of the ex-cursion occurs between Datum 3 and Datum 6 (Fig. 7; Tables DR5 and DR6 [see footnote 1]).
The Viki core, Estonia, serves as one of the primary reference sections for the Telychian portion of the conodont biostratigraphic scheme (Männik, 2007a). Most of the δ13Ccarb data presented here (Fig. 8A; Table DR7 [see foot-note 1]) are from Kaljo et al. (2003) except for
a handful of new data points near the base of the Wenlock Series. Similar to Gotland, the Viki δ13Ccarb record shows values averaging +1.5‰ in the Pt. amorphognathoides amorphognathoides Zonal Group and a rapid increase to above +4.0‰ during the earliest Wenlock. Carbonate carbon isotope values reach >+5.0‰ but then show a rapid decline back to pre-excursion val-ues. In the Viki core, the onset of the excursion occurs at Datum 2 (Fig. 8A; Table DR7 [see footnote 1]).
The Ruhnu core, Estonia, has long served as a regional reference for Baltic Silurian stratig-raphy (Põldvere, 2003). No new samples from the Ruhnu core were included in this investiga-tion, but a synoptic presentation of all presently available graptolite, conodont, and δ13Ccarb data is shown here for the fi rst time (Fig. 8C; Table DR8 [see footnote 1]). The δ13Ccarb record shows values averaging +1.5‰ during the uppermost Telychian and rapidly increasing during the lowermost Sheinwoodian (Riga Formation), eventually exceeding +4.0‰ in the Jaani For-mation. Similar to the Ohesaare δ13Ccarb record, Ruhnu values never exceeded +5.0‰ and show a slow, steady decline to pre-excursion values in the Jamaja Formation. In the Ruhnu core, the onset of the excursion occurs below Datum 3 (Fig. 8C; Table DR8 [see footnote 1]).
Robert Moses Power Plant S-1 Core, New York, USA
The Robert Moses Power Plant S-1 core, New York, represents the most detailed and extensive record of the early Sheinwoodian (Ireviken) positive carbon isotope excursion yet recovered from North America. The δ13Ccarb data from the S-1 core are shown in Figure 9C and Table DR9 (see footnote 1). Conodont biostratigraphic in-formation is modifi ed from Kleffner (1991) and Cramer et al. (2006a). Carbonate carbon isotope values increase by nearly +2.0‰ (from +3.13‰ to +4.98‰) across the Rockway Irondequoit contact, and except for a decrease of roughly –2.0‰ in the lowermost Rochester Forma-tion, δ13Ccarb values remain near +5.0‰ into the Gasport Formation before declining to below pre-excursion values near the top of the Goat Island Formation. Due to the unconformity at the Rockway Irondequoit contact, the strata that contain Datum 3, Datum 6, and the onset of the excursion are all missing at this level (Fig. 9C; Table DR9 [see footnote 1]).
Comparison of the S-1 core to summaries of the stratigraphy of New York and the U.S. Midcontinent shows that the S-1 core exhibits an atypical record. In the core, the Rockway Formation is comparatively thin, and the black phosphatic interval below it, questionably as-
signed to the Williamson Formation by Cramer et al. (2006a), is even thinner. Typically, the Rockway Formation is as thick, or thicker, than the Irondequoit Formation (Brett et al., 1995), and the lack of an extensive Williamson Forma-tion immediately below it suggests a sizeable disconformity in the Telychian portion of this core (Brett et al., 1995; Loydell et al., 2007; McLaughlin et al., 2008). The lithostratigraphic and biostratigraphic information provided (Fig. 9C; Table DR9 [see footnote 1]) has been modi-fi ed from Cramer et al. (2006a) to refl ect these observations (McLaughlin et al., 2008).
CHRONOSTRATIGRAPHIC CORRELATION
The Silurian System is typical of much of the lower Paleozoic where both conodont and graptolite biozonations are available: one used primarily in shale basins and the other primar-ily on carbonate platforms. Correlating between these two types of biostratigraphy, however, has been challenging. Carbonate carbon isotope (δ13Ccarb) stratigraphy provides an opportunity to correlate between independent biostratigraphic schemes and is particularly useful when the fossil groups used to construct the zonations are not found together in abundance. Here, we have compiled δ13Ccarb data from: two of the best studied Telychian through earliest Sheinwood-ian graptolite localities (Banwy River and the Aizpute-41 core); the Telychian and Sheinwood-ian reference sections for conodont biostratig-raphy (Viki core and Gotland); the type area of the Niagaran (North American) Provincial Series (New York); as well as two of the most intensively studied Silurian cores in the world (Ruhnu and Ohesaare cores). As a result, we can verify and/or improve correlation between the graptolite and conodont biozonations, dem-onstrate the fi delity of δ13Ccarb stratigraphy as a chronostratigraphic tool, as well as begin to see the paleobiogeographic implications of inte-grated high-resolution chronostratigraphy.
Biochemostratigraphy
The compilation diagrams presented herein (Figs. 8 and 9) illustrate a remarkable degree of consistency between all three methods of chronostratigraphic correlation (grapto-lite and conodont biostratigraphy and δ13Ccarb chemostratigraphy). However, some impor-tant variability exists between the timing of biologic versus chemical events among the sampled sections. Rather than being a demon-stration of the unreliability of biostratigraphy or the inconsistency of δ13Ccarb chemo stratig-raphy, such variability should be expected

Cramer et al.
1712 Geological Society of America Bulletin, September/October 2010
when working at such a fi ne temporal scale. The data presented herein were collected at cm-scale resolution and the temporal variabil-ity we are seeing is likely to be measured in thousands of years, not millions of years (see below), but such variability forces us to ad-dress a signifi cant chronostratigraphic ques-tion: Is the fact that conodont fi rst and last appearances occur at different positions with respect to the early Sheinwoodian (Ireviken) δ13Ccarb excursion: (1) a result of the excursion beginning and ending at different times in dif-ferent places; or (2) a result of FADs and LADs occurring at different times in different places?
Both choices are likely correct because nei-ther the biostratigraphic nor chemostratigraphic markers are truly isochronous horizons; how-ever, there is a difference in potential between the two choices. Because the global ocean mix-ing time is typically considered to be on the scale of a few k.y., even for poorly ventilated Paleozoic oceans (Kiehl and Shields, 2005), a clearly global chemostratigraphic event such as the early Sheinwoodian (Ireviken) δ13Ccarb excur-sion is likely to be a reliable chronostratigraphic marker to within plus or minus a few thousand years (a few tens of thousands at most). Biologi-cal speciation and migration, or extinction and Lazarus taxa, on the other hand, are likely to be consistent within tens to hundreds of thousands of years (cf. Ma and Day, 2003); but global Paleozoic fi rst and last appearances, combined with the likelihood of actually sampling the true moment of fi rst or last appearance (Barrick and Männik, 2005), are unlikely to be able to ap-proach the chronostratigraphic potential offered by δ13Ccarb stratigraphy.
The global limit to the chronostratigraphic resolving power provided by origination and/or extinction of any single species is highly vari-able. Because species typically develop from small or isolated populations in local areas, there is a geographic component to the rate of distribution of any given species. That is, it will take time to migrate across the geography of the planet. In addition, because speciation is often a response to changing environmental conditions, where new conditions provide a selective ad-vantage for a new character possessed by a new species, there is also an environmental compo-nent to the rate of distribution of new species. Whereas local conditions may favor a new spe-cies, it may not be until global conditions are similarly advantageous that the new species can colonize the remainder of the planet. Therefore, geographic confi guration, oceanographic circu-lation, as well as global and local environmental conditions, can all signifi cantly impact the rate of expansion of new species. Similarly, the truly global last appearance of a species can be geo-
logically instantaneous (such as has been pro-posed for the end-Cretaceous event), but more often, species tend to disappear fi rst in, what were to them, areas of marginal environmental conditions. A species may survive long after its interval of global cosmopolitan status, in small isolated areas of refugia, prior to its fi nal truly global last appearance. All of these factors limit the potential resolving power of single-taxon biostratigraphic zones, and some examples have been demonstrated by the data presented here.
The paleobiogeographical difference in the fi rst appearance of the conodont K. walliseri, fi rst appearing in North America at least a full sequence earlier than in the Baltic (Sequence VI versus Sequence VII of Brett et al., 1998), and the varying positions of fi rst and last appear-ances with respect to the δ13Ccarb record (Figs. 8 and 9) likely demonstrate the fact that we are operating at the functional temporal limit of single-taxon biostratigraphic zonations. Until paleobiogeographic origination and migration histories are better documented, we have chosen herein to consider the δ13Ccarb record to be the chronostratigraphic tool of choice. Signifi cant future research is necessary to fully determine which will be the fi nal chronostratigraphic tool of record (isotopes, biostratigraphy, or some-thing else), and our choice to use the δ13Ccarb record was made primarily in order to simplify discussion. As a result we discuss the changing positions of the datum points of the Ireviken Event with respect to the early Sheinwoodian (Ireviken) δ13Ccarb excursion below, not the other way around.
Onset of the Early Sheinwoodian (Ireviken) δ13Ccarb Excursion
Although conodont biostratigraphy had been well correlated with the early Sheinwood-ian (Ireviken) δ13Ccarb excursion from Gotland (Munnecke et al., 2003), the data presented here demonstrate important new information regard-ing the correlation of conodont and graptolite biostratigraphic zones with the onset of the early Sheinwoodian (Ireviken) δ13Ccarb excur-sion. Brand et al. (2006) correlated the onset of the excursion with the base of the C. centrifugus Biozone, and Noble et al. (2005) correlated it to a position below a combined C. centrifugus–C. insectus Biozone. Neither study contained graptolite biostratigraphic data to support these positions (see discussion in Loydell, 2007). Without exception, the sections studied herein demonstrate that the onset of the early Shein-woodian (Ireviken) δ13Ccarb excursion correlates to a position from high in the C. murchisoni graptolite Biozone to near the base of the M. fi r-mus graptolite Biozone.
The gray band labeled 1 in Figures 5–9 (ex-cept Fig. 9A) was drawn with a lower boundary at Datum 3 of the Ireviken Event and an upper boundary at Datum 6 effectively highlighting the Pt. pennatus procerus Superzone (Fig. 3). In Gotland, New York, and the Ohesaare core, this interval (Pt. pennatus procerus Superzone) con-tains the infl ection point in the δ13Ccarb curve and the onset of the early Sheinwoodian (Ireviken) δ13Ccarb excursion (open circle within gray band 1 in Figs. 6, 7, and 9C). In the Aizpute-41, Viki and Ruhnu cores, however, Datum 3 of the Ireviken Event is above the onset of the early Sheinwoodian (Ireviken) δ13Ccarb excursion (open circle below gray band labeled 1 in Figs. 5, 8A, and 8C).
The presence of Datum 3 above the onset of the δ13Ccarb excursion is not the result of sampling a drill core (i.e., an effect of small sample size) because it is the presence, not the absence, of certain conodont species that is signifi cant here. It is the presence of Pt. amorphognathoides amorphognathoides and Panderodus sp. nov. N above the onset of the early Sheinwoodian (Ireviken ) δ13Ccarb excursion that extends Datum 3 to this level, and therefore likely represents ac-tual variability in the last appearances of those species. The onset of the early Sheinwoodian (Ireviken) δ13Ccarb excursion correlates to a position ranging from Datum 2 (Viki core) to Datum 4 (Gotland) of the Ireviken Event, rather than exclusively to Datum 4 as was previously thought (Munnecke et al., 2003).
The onset of the early Sheinwoodian (Ireviken ) δ13Ccarb excursion correlates to an interval from the upper part of the C. murchi-soni graptolite Biozone to near the base of the M. fi rmus graptolite Biozone and from Datum 2 to Datum 4 of the Ireviken Event. The onset of the early Sheinwoodian (Ireviken) δ13Ccarb excursion is therefore restricted to the lower-most Sheinwoodian based upon either biostrati-graphic correlation for the base of the Wenlock Series (graptolites or conodonts).
Decline of δ13Ccarb Following Initial Peak
In sections that have been sampled at a suf-fi cient resolution and are comparatively strati-graphically complete, a consistent negative shift in the δ13Ccarb curve of roughly –1.0‰ to –2.0‰ slightly above the initial peak has been recorded in Sweden (Samtleben et al., 1996; Calner et al., 2004), Iowa (Cramer et al., 2006c), New York (Cramer et al., 2006a), and now in the Banwy River section (Figs. 4 and 9). On Got-land (Fig. 6), this negative feature is recorded in the Högklint Formation, which, as discussed above, belongs to either the M. fi rmus or M. ric-cartonensis graptolite Biozone (Fig. DR2 [see

Testing the limits of Paleozoic chronostratigraphy
Geological Society of America Bulletin, September/October 2010 1713
footnote 1]). Recognition of this δ13Ccarb feature in the M. riccartonensis Biozone in the Banwy River section (Fig. 4) is consistent with these observations.
The gray band labeled 2 was drawn based upon the isotopic feature itself (Figs. 4, 6, and 9), and is consistently located within the K. ranuli-formis Superzone. This feature can be located more precisely within the Upper K. ranuliformis Zone; however, detailed biostratigraphic con-trol is required to subdivide the superzone. In each conodont-bearing section included herein, we could defi ne the K. ranuliformis Superzone using available biostratigraphic data, but defi n-ing separate Lower and Upper K. ranuliformis biozones was possible only in a few localities.
This isotopic feature is restricted to the M. ric-carto nensis graptolite Biozone and the Upper K. ranuliformis conodont Biozone within the K. ranuli formis Superzone. It should be noted that this negative δ13Ccarb feature is not associ-ated with a sequence boundary or signifi cant disconformity (cf. Algeo et al., 1992), but it occurs within a sequence (Sequence V of Brett et al., 1998; McLaughlin et al., 2008), is defi ned by multiple isotope samples over several me-ters of strata (not a single data point or a single stratigraphic horizon), and is also recognizable in the well-preserved brachiopod data from Got-land (Fig. 6). Most importantly, this feature is biostratigraphically consistent from Laurentia, Avalonia, and Baltica (Fig. 9).
End of the Early Sheinwoodian (Ireviken) δ13Ccarb Excursion
We have defi ned the end of the early Shein-woodian (Ireviken) δ13Ccarb excursion as the point where δ13Ccarb values are consistently at or below the values immediately preceding the onset of the excursion. The gray band labeled 3 has been placed to highlight the descending limb of the excursion as δ13Ccarb values drop from near peak levels to below starting values, effectively placing the end of the excursion near the top of this box. In the Banwy River section, all three sets of data recovered (δ13Ccarb, δ13Corg, TOC; Fig. 4) indicate that the beginning of the end of the excursion is present in the top of the section within the “mid-Wenlock” grapto-lite interval (graptolites from the top of the sec-tion include Monograptus fl exilis).
The end of the early Sheinwoodian (Ireviken) δ13Ccarb excursion is restricted to the Lower K. walliseri Biozone of the O. sagitta rhenana Superzone. The conodont K. walliseri appears to have differing paleobiogeographic appear-ances. It fi rst appears in parts of North Amer-ica during the early Sheinwoodian (Ireviken ) δ13Ccarb excursion, when δ13Ccarb values were still
>+5.0‰ (Fig. 9C) at the base of Sequence VI of Brett et al. (1998). This position is equivalent to the base of the Tofta Formation on Gotland (McLaughlin et al., 2008). Kockelella walliseri fi rst occurs on Gotland (and apparently through-out the Baltic) where δ13Ccarb values are just below +2.0‰ at the very end of the early Shein-woodian (Ireviken) δ13Ccarb excursion (Fig. 6). In either case, δ13Ccarb values begin to be con-sistently below pre-excursion levels within the Lower K. walliseri Biozone (high in the biozone in North America, and near the base of the biozone in Baltica).
Temporal Control for Telychian–Sheinwoodian Chronostratigraphy
Radiometric age control for the Silurian is generally poor (Gradstein et al., 2004), making it diffi cult to determine the scale of temporal resolution achieved in the present study. If the Sheinwoodian is only two million years long, as suggested by Gradstein et al. (2004), then the entire early Sheinwoodian (Ireviken) δ13Ccarb ex-cursion perhaps represents one million years. If this is the case, then the conodont and graptolite biozones that correlate to positions within the excursion are each on the order of 500 k.y. or less, and the interval near the base of the Wen-lock Series and the onset of the δ13Ccarb excursion is defi ned by biozones (particularly conodont biozones) of signifi cantly shorter duration.
Recently, a method for identifi cation and cali bration of orbital signals within the deep-time sedimentary record has been developed (Meyers and Sageman, 2007) and applied to the Gotland sequence (Cramer, 2009), and it provides the fi rst portion of an orbitally tuned Silurian time scale. Results (Cramer, 2009) indicate that the Upper Visby Formation was deposited at a rate of roughly 4.5 cm/k.y., meaning that the Pt. pennatus procerus Super-zone represents a time interval of ≤100 k.y., and that the individual datum points during the Ireviken Event are separated by only tens of thousands of years. As a result, even though we cannot precisely correlate the onset of the early Sheinwoodian (Ireviken) δ13Ccarb excur-sion with graptolite or conodont biostratigraphy and are instead left with a range of correlation (upper part of the C. murchisoni to the base of the M. fi rmus graptolite Biozone or Datum 2 to Datum 4 of the Ireviken Event), such a range likely represents less than 100 k.y. Therefore, regardless of which proves to be the most reli-able chronostratigraphic marker—conodonts, graptolites, or the δ13Ccarb record—the integra-tion of all three chronostratigraphic tools has produced a Silurian time scale comparable to that of more recent time periods.
CONCLUSIONS
The early Sheinwoodian (Ireviken) δ13Ccarb ex-cursion was entirely limited to the Wenlock, with an onset at a level from high within the C. mur-chisoni graptolite Biozone to the base of the M. fi rmus graptolite Biozone and from Datum 2 to Datum 4 of the Ireviken Event. A small de-cline in δ13Ccarb values occurs, slightly above the initial peak, within the M. riccartonensis grap-tolite Biozone and the Upper K. ranuliformis conodont Biozone (within the K. ranuliformis Superzone) before a return to a sustained inter-val of peak values. The δ13Ccarb values decline to a level consistently below pre-excursion values within the “mid-Wenlock” graptolite interval and within the Lower K. walliseri conodont Biozone (although where within the Lower K. walliseri Biozone appears to depend on the paleoconti-nent under investigation). The seven conodont biozones and four graptolite biozones between the base of the Wenlock Series and the top of the early Sheinwoodian (Ireviken) δ13Ccarb excur-sion, representing roughly one million years of time, demonstrate that these biozones (Fig. 10) have a maximum duration of <500 k.y. each and even shorter durations near the base of the Wenlock Series. By achieving intercontinental chronostratigraphic correlation on such a scale, we have begun to demonstrate that it is possible to erect a Paleozoic time scale comparable to that of younger eras.
The present study results from more than a decade of intense research surrounding the base of the Wenlock Series and the early Shein woodian (Ireviken) δ13Ccarb excursion and the contributions of numerous stratigraphic disciplines from biostra-tigraphy to sedimentology to isotope geochemis-try to basic core logging and fi eld mapping. The improvements made to global Silurian chrono-strati graphic correlation during the past decade have had a signifi cant impact on the age and global correlation of the Silurian strata of North America (Kleffner et al., 2005; Metzger, 2005; Cramer et al., 2006a, 2006b, 2006c; Melchin and Holmden, 2006; Loydell et al., 2007; McLaughlin et al., 2008), while the growth of δ13Ccarb stratig-raphy as a chronostratigraphic tool independent of biostratigraphy has begun to provide a new opportunity to investigate the paleobiogeographic migration of certain taxa. It is only through the in-tegration of various means of chronostratigraphic correlation that the recent advances in Silurian global chronostratigraphy could have taken place. We hope that this paper may serve to demonstrate not only what chronostratigraphic resolution is possible during the Paleozoic but also how much basic stratigraphic research still remains to be done in order to produce a Phanerozoic time scale of equal resolution and fi delity throughout.

Cramer et al.
1714 Geological Society of America Bulletin, September/October 2010
ACKNOWLEDGMENTS
We thank Richard Cave, Tony Butcher, and Bob Loveridge for help with sample collection at the Banwy River locality, Loren Babcock for improving an earlier version of this manuscript, and the U.S. Geological Survey Mineral Team for support. Insight-ful and constructive comments from Charles Mitchell , Peter Sheehan, and Peter Sadler helped make this a stronger manuscript and were greatly appreciated. This work is a contribution to the International Geo-
science Programme (IGCP) 503 project and was partially funded by Natural Environment Research Council (NERC) small grant GR9-1129 to D.K.L.; German Research Foundation (DFG) grant Mu-2352-1 to A.M.; Estonian Science Foundation (ETF) grant 7138 to P.M., and Estonian target fi nanced projects SF-0140020s08 and SF-0320080s07; grants from the Swedish Research Council (VR) and its predeces-sor the Swedish National Science Research Council (NFR) to L.J.; and National Science Foundation (NSF) grant EAR-0517929 to M.A.K. and J.E.B.
REFERENCES CITED
Algeo, T.J., Lohmann, K.C., and Wilkinson, B.H., 1992, Meteoric-burial diagenesis of middle Pennsylvanian limestones in the Orogrande Basin, New Mexico: Water/rock interactions and basin geothermics: Journal of Sedimentary Petrology, v. 62, p. 652–670.
Bancroft, A.M., 2008, Silurian conodont biostratigraphy and carbonate carbon isotope stratigraphy of the Eramosa of Southwestern Ontario, Canada [M.S. thesis]: Colum bus, The Ohio State University, 45 p.
Banner, J.L., and Hanson, G.N., 1990, Calculation of simul-taneous isotopic and trace element variations during water-rock interaction with applications to carbonate diagenesis: Geochimica et Cosmochimica Acta, v. 54, p. 3123–3137, doi: 10.1016/0016-7037(90)90128-8.
Barrick, J.E., and Klapper, G., 1976, Multielement Silurian (late Llandoverian–Wenlockian) conodonts of the Clarita Formation, Arbuckle Mountains, Oklahoma, and phylogeny of Kockelella: Geologica et Palaeonto-logica, v. 10, p. 59–100.
Barrick, J.E., and Männik, P., 2005, Silurian conodont biostratigraphy and palaeobiology in stratigraphic se-quences, in Purnell, M.A., and Donoghue, P.C.J., eds., Conodont Biology and Phylogeny—Interpreting the Fossil Record: The Palaeontological Association Spe-cial Papers in Palaeontology, v. 73, p. 103–116.
Bassett, M.G., and Edwards, D., eds., 1991, The Murchison Symposium—Proceedings of an International Confer-ence on the Silurian System: Special Papers in Palaeon-tology, v. 44, 397 p.
Bassett, M.G., Rickards, R.B., and Warren, P.T., 1975, The type Wenlock Series: Report of the Institute of Geo-logical Sciences, v. 75, p. 1–19.
Brand, U., Azmy, K., and Veizer, J., 2006, Evaluation of the Salinic I tectonic, Cancañiri glacial, and Ireviken biotic events: Biochemostratigraphy of the Lower Silurian succession in the Niagara Gorge area, Canada and U.S.A: Palaeogeography, Palaeoclimatol-ogy, Palaeoecology, v. 241, p. 192–213, doi: 10.1016/j.palaeo.2006.03.004.
Brett, C.E., Tepper, D.H., Goodman, W.M., Loduca, S.T., and Eckert, B.-Y., 1995, Revised stratigraphy and cor-relation of the Niagaran Provincial Series (Medina, Clinton, and Lockport Groups) in the type area of western New York: U.S. Geological Survey Bulletin 2086, 66 p.
Brett, C.E., Baarli, B.G., Chowns, T.M., Cotter, E., Driese, S., and Johnson, M.E., 1998, Early Silurian condensed horizons, ironstones, and sequence stratigraphy in the Appalachian Foreland Basin, in Landing, E., and John-son, M.E., eds., Silurian Cycles: Linkages of Dynamic Stratigraphy with Atmospheric, Oceanic, and Tectonic Changes: New York State Museum Bulletin, v. 491, p. 265–282.
Calner, M., and Eriksson, M.E., 2006, Silurian research at the crossroads: GFF (Geologiska Föreningens i Stock-holm Förhandlingar), v. 128, p. 73–74, doi: 10.1080/11035890601282073.
Calner, M., Jeppsson, L., and Munnecke, A., 2004, The Silurian of Gotland—Part 1: Review of the stratigraphic framework, event stratigraphy, and stable carbon and oxygen isotope development: Erlanger Geologische Abhandlungen, Sonderband, v. 5, p. 113–131.
Cocks, L.R.M., 2001, Ordovician and Silurian global geog-raphy: Journal of the Geological Society of London, v. 158, p. 197–210.
Corfi eld, R.M., Siveter, D.J., Cartlidge, J.E., and McKerrow , W.S., 1992, Carbon isotope excursion near the Wenlock–Ludlow (Silurian) boundary in the Anglo-Welsh area: Geology, v. 20, p. 371–374, doi: 10.1130/0091-7613(1992)020<0371:CIENTW>2.3.CO;2.
Cramer, B.D., 2009, Chapter 2: Extending astrochronology into the lower Paleozoic, in Application of Integrated High-Resolution Biochemostratigraphy to Paleozoic Chronostratigraphic Correlation: Recalibrating the Silurian System [Ph.D. thesis]: Columbus, The Ohio State University, p. 37–46.
Cramer, B.D., and Munnecke, A., 2008, Early Silurian δ13C excursions and their relationship to glaciations, sea-level changes and extinction events: Geological Jour-nal, v. 43, no. 4, p. 517–519, doi: 10.1002/gj.1112.
Figure 10. Revised Telychian and Sheinwoodian chrono-stratigraphic correlation chart based upon Jeppsson (1997), Männik (1998, 2007a, 2007b), Loydell et al. (2003), Munnecke et al. (2003), Cramer et al. (2006a), McLaughlin et al. (2008), and the integrated data presented herein. Numbers in boxes in the conodont biozona-tion column refer to the datum points of the Ireviken Event (Jeppsson, 1997). The gray area in the series and stage col-umns highlights the confusion regarding the graptolite ver-sus conodont correlation for the base of the Wenlock Series (see text and Loydell, 2008a). Although the best known local-ity that contains both conodont and graptolite index fossils for the base of the Wenlock Series shows both indicators at the same horizon (immediately above an unconformity), grow-ing evidence (Männik et al., 2002; Männik, 2007a, 2007b) suggests that the position of Datum 2 (conodont correla-tion to the global boundary stratotype and point) corre-lates to a position within the murchisoni graptolite Biozone, not at the base of the biozone as in the Aizpute-41 core (see text). Chitinozoan evidence from Hughley Brook (Mul-lins and Aldridge, 2004) lim-its the possible level of the Global Boundary Stratotype Section and Point (GSSP) to the upper C. centrifugus to lower C. murchisoni Biozone inter-val. Although the current lack of other sections containing both index fossils remains a vexing problem, in either case, the early Sheinwoodian (Ireviken) positive δ13Ccarb excur-sion is restricted to the Wenlock.
Sp. turriculatus
Sp. guerichi
Sti. halli
C. murchisoni
C. centrifugus
C. insectus
C. lapworthi
O. spiralis
Mcl. crenulata
Mcl. griestoniensis
Str. sartorius
Str. crispus
K. ranuliformisS.Z.
Pt. a. amorpho-gnathoides Z.G.
M. firmus
M. riccartonensis
‘Mid-Wenlock’(C. perneri,
C. rigidus/M. flexilis,‘M.’ antennularius)
C. lundgreni
Pt. eop. ssp. nov. 1
Pt. eopennatus ssp. nov. 2
Pt. amorph.angulatus
Pt. a. lennarti
Pt. a. lithuanicus
Pt. p. procerus S.Z.
O. sagittarhenana
K. ortusortus
O. sagittasagitta
D. stauro-gnathoides
Lower K. walliseri
Upper K. walliseriSuperzoneLl
ando
very
Wen
lock
Tely
chia
nS
hein
woo
dian
Hom
.A
er.
? ? ?
Series Stage Graptolite zone Conodont zone
23 46
8
Post K. walliseriInterregnum

Testing the limits of Paleozoic chronostratigraphy
Geological Society of America Bulletin, September/October 2010 1715
Cramer, B.D., and Saltzman, M.R., 2005, Sequestration of 12C in the deep ocean during the early Wenlock (Silurian) positive carbon isotope excursion: Palaeo-geography, Palaeoclimatology, Palaeoecology, v. 219, p. 333–349, doi: 10.1016/j.palaeo.2005.01.009.
Cramer, B.D., and Saltzman, M.R., 2007, Early Silurian paired δ13Ccarb and δ13Corg analyses from the Midcontinent of North America: Implications for paleocean og raphy and paleoclimate: Palaeogeography, Palaeo climatology, Palaeoecology, v. 256, p. 195–203, doi: 10.1016/j.palaeo.2007.02.032.
Cramer, B.D., Saltzman, M.R., and Kleffner, M.A., 2006a, Spatial and temporal variability in organic carbon burial during global positive carbon isotope δ13Ccarb excursions: New insight from high-resolution δ13Ccarb stratigraphy from the type area of the Niagaran (Silu-rian) Provincial Series: Stratigraphy, v. 2, p. 327–340.
Cramer, B.D., Kleffner, M.A., and Saltzman, M.R., 2006b, The Late Wenlock Mulde positive carbon isotope ex-cursion in North America: GFF (Geologiska Förenin-gens i Stockholm Förhandlingar), v. 128, p. 85–90, doi: 10.1080/11035890601282085.
Cramer, B.D., Kleffner, M.A., and Saltzman, M.R., 2006c, Chemostratigraphic correlation of lower Silurian deposits in eastern Iowa: Placing the Llandovery–Wenlock boundary in the mid-continent: Iowa Geo-logical Survey Guidebook Series, v. 25, p. 103–109.
Eriksson, M.E., and Calner, M., eds., 2005, The Dynamic Silurian Earth: Sveriges geologiska undersökning Rap-porter och meddelanden, v. 121, p. 1–99.
Fischer, A.G., 1983, Long-term climate oscillations recorded in stratigraphy, in Berger, W.H., ed., Climate and Earth History: National Academy of Sciences, p. 97–104.
Gradstein, F.M., Ogg, J.G., and Smith, A.G., eds., 2004, A Geologic Time Scale 2004: Cambridge, UK, Cam-bridge University Press, 589 p.
Hede, J.E., 1921, Gotlands Silurstratigrafi : Sveriges Geolo-giska Undersökning, C 305, 100 p.
Hede, J.E., 1940, Berggrunden, in Lundqvist, G., Hede, J.E., and Sundius, N., eds., Beskrivning till kartbladet Visby and Lummelunda: Sveriges Geologiska Undersökning, Aa 183, 167 p.
Hede, J.E., 1960, The Silurian of Gotland, in Regnéll, J.E., and Hede, J.E., eds., The Lower Palaeozoic of Scania. The Silurian of Gotland: Stockholm, Inter-national Geological Congress XXI, Session Norden, Guidebook Sweden, 89 p. [Also available as: Publi-cations of the Institutes of Mineralogy, Palaeontol-ogy, and Quaternary Geology of the University of Lund, v. 91.]
Helfrich, C.T., 1980, Late Llandovery–early Wenlock cono-donts from the upper part of the Rose Hill and the basal part of the Miffl intown formations, Virginia, West Vir-ginia, and Maryland: Journal of Paleontology, v. 54, p. 557–569.
Jaeger, H., 1959, Graptolithen und Stratigraphie des jüngsten Thüringer Silurs: Berlin, Abhandlungen der Deutschen Akademie der Wissenschaften Berlin, Klasse Chemie, Geologie und Biologie v. 2, 197 p.
Jeppsson, L., 1987, Lithological and conodont distributional evidence for episodes of anomalous oceanic conditions during the Silurian, in Aldridge, R.J., ed., Palaeobiol-ogy of Conodonts: Chichester, UK, Ellis Horwood, p. 129–145.
Jeppsson, L., 1997, A new latest Telychian, Sheinwoodian and early Homerian (early Silurian) standard conodont zonation: Transactions of the Royal Society of Edin-burgh, Earth Sciences, v. 88, p. 91–114.
Jeppsson, L., 2008, The lower Wenlock Hangvar Formation—A sequence previously split between the Högklint and Slite beds (Silurian, Gotland, Sweden): GFF (Geolo-giska Föreningens i Stockholm Förhandlingar), v. 130, p. 31–40, doi: 10.1080/11035890801301031.
Jeppsson, L., Eriksson, M.E., and Calner, M., 2006, A lat-est Llandovery to latest Ludlow high-resolution bio-stratigraphy based on the Silurian of Gotland—A summary: GFF (Geologiska Föreningens i Stockholm Förhandlingar), v. 128, p. 109–114, doi: 10.1080/11035890601282109.
Jeppsson, L., Talent, J.A., Mawson, R., Simpson, A.J., Andrew, A.S., Calner, M., Whitford, D.J., Trotter, J.A., Sandström, O., and Caldon, H.-J., 2007, High-
resolution Late Silurian correlations between Gotland, Sweden, and the Broken River region, NE Australia: Lithologies, conodonts and isotopes: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 245, p. 115–137, doi: 10.1016/j.palaeo.2006.02.032.
Johnson, M.E., Rong, J., Wang, C., and Wang, P., 2001, Conti-nental island from the Upper Silurian (Ludfordian Stage) of Inner Mongolia: Implication for eustasy and paleo-geography: Geology, v. 29, p. 955–958, doi: 10.1130/0091-7613(2001)029<0955:CIFTUS>2.0.CO;2.
Kaljo, D., and Martma, T., 2006, Application of carbon isotope stratigraphy to dating the Baltic Silurian rocks: GFF (Geologiska Föreningens i Stockholm Förhandlingar), v. 128, p. 123–129, doi: 10.1080/11035890601282123.
Kaljo, D., Kiipli, T., and Martma, T., 1997, Carbon isotope event markers through the Wenlock–Přídolí sequence in Ohesaare (Estonia) and Priekule (Latvia): Palaeo-geography, Palaeoclimatology, Palaeoecology, v. 132, p. 211–224, doi: 10.1016/S0031-0182(97)00065-5.
Kaljo, D., Kiipli, T., and Martma, T., 1998, Correlation of carbon isotope events and environmental cyclicity in the East Baltic Silurian, in Landing E., and Johnson, M.E., eds., Silurian Cycles: Linkages of Dynamic Stratigraphy, with Atmospheric, Oceanic, and Tectonic Changes: New York State Museum Bulletin, v. 491, p. 297–312.
Kaljo, D., Martma, T., Männik, P., and Viira, V., 2003, Implications of Gondwana glaciations in the Baltic Late Ordovician and Silurian and a carbon isotopic test of environmental cyclicity: Bulletin de la Société Géologique de France, v. 174, p. 59–66, doi: 10.2113/174.1.59.
Kaljo, D., Grytsenko, V., Martma, T., and Mõtus, M.-A., 2007, Three global carbon isotope shifts in the Silu-rian of Podolia (Ukraine): Stratigraphical implications: Estonian Journal of Earth Sciences, v. 56, p. 205–220, doi: 10.3176/earth.2007.02.
Kiehl, J.T., and Shields, C.A., 2005, Climate simulation of the latest Permian: Implications for mass extinction: Geology, v. 33, p. 757–760, doi: 10.1130/G21654.1.
Kleffner, M.A., 1991, Conodont biostratigraphy of the up-per part of the Clinton Group and the Lockport Group (Silurian) in the Niagara Gorge region, New York and Ontario: Journal of Paleontology, v. 65, p. 500–511.
Kleffner, M.A., Cramer, B.D., and Saltzman, M.R., 2005, First documentation of the Early Wenlock (Ireviken) positive δ13Ccarb excursion in the type area of the Niagaran Provincial Series: Irondequoit Limestone, Rochester Shale, Decew Dolomite, Gasport Dolomite, and Goat Island Dolomite are all Sheinwoodian (early Wenlock) in age: Geological Society of America Ab-stracts with Programs, v. 37, no. 5, p. 79.
Loydell, D.K., 1998, Early Silurian sea-level changes: Geo-logical Magazine, v. 135, p. 447–471, doi: 10.1017/S0016756898008917.
Loydell, D.K., 2007, Early Silurian positive δ13C excursions and their relationship to glaciations, sea-level changes and extinction events: Geological Journal, v. 42, p. 531–546, doi: 10.1002/gj.1090.
Loydell, D.K., 2008a, The base of the Wenlock Series: Silu-rian Times, v. 15, p. 10–12.
Loydell, D.K., 2008b, Early Silurian δ13C excursions and their relationship to glaciations, sea-level changes and extinction events: Geological Journal, v. 43, p. 511–515, doi: 10.1002/gj.1110.
Loydell, D.K., and Cave, R., 1996, The Llandovery–Wenlock boundary and related stratigraphy in eastern mid-Wales with special reference to the Banwy River section: Newsletters on Stratigraphy, v. 34, p. 39–64.
Loydell, D.K., and Frýda, J., 2007, Carbon isotope stratig-raphy of the upper Telychian and lower Sheinwood-ian (Llandovery–Wenlock, Silurian) of the Banwy River section, Wales: Geological Magazine, v. 144, p. 1015–1019.
Loydell, D.K., and Nestor, V., 2005, Integrated graptolite and chitinozoan biostratigraphy of the upper Telychian (Llandovery, Silurian) of the Ventspils D-3 core, Lat-via: Geological Magazine, v. 142, p. 369–376, doi: 10.1017/S0016756805000531.
Loydell, D.K., Kaljo, D., and Männik, P., 1998, Integrated biostratigraphy of the lower Silurian of the Ohesaare
core, Saaremaa, Estonia: Geological Magazine, v. 135, p. 769–783, doi: 10.1017/S0016756898001423.
Loydell, D.K., Männik, P., and Nestor, V., 2003, Integrated biostratigraphy of the lower Silurian of the Aizpute-41 core, Latvia: Geological Magazine, v. 140, p. 205–229, doi: 10.1017/S0016756802007264.
Loydell, D.K., Kleffner, M.A., Mullins, G.L., Butcher, A., Matteson, D.K., and Ebert, J.R., 2007, The lower Williamson Shale (Silurian) of New York: A bio-stratigraphical enigma: Geological Magazine, v. 144, p. 225–234, doi: 10.1017/S0016756806003049.
Ma, X., and Day, J.E., 2003, Revision of selected North American and Eurasian Late Devonian (Frasnian) spe-cies of Cyrtospirifer and Regelia (Brachiopoda): Jour-nal of Paleontology, v. 77, p. 267–292, doi: 10.1666/0022-3360(2003)077<0267:ROSNAA>2.0.CO;2.
Mabillard, J.E., and Aldridge, R.J., 1985, Microfossil distri-bution across the base of the Wenlock Series in the type area: Palaeontology, v. 28, p. 89–100.
Männik, P., 1998, Evolution and taxonomy of the Silurian conodont Pterospathodus: Palaeontology, v. 41, p. 1001–1050.
Männik, P., 2007a, An updated Telychian (late Llandovery, Silurian) conodont zonation based on Baltic faunas: Lethaia, v. 40, p. 45–60.
Männik, P., 2007b, Some comments on Telychian–Early Sheinwoodian conodont faunas, events and stratigra-phy: Acta Palaeontologica Sinica, v. 46, supplement, p. 305–310.
Männik, P., Kallaste, T., and Martma, T., 2002, Dating of the Ireviken Event and the problem of the Llandovery–Wenlock boundary, some possible developments, in Satkünas, J., and Lazauskienė, J., eds., The Fifth Bal-tic Stratigraphical Conference “Basin Stratigraphy—Modern Methods and Problems,” September 22–27, 2002: Vilnius, Lithuania, Extended Abstracts, Geologi-cal Survey of Lithuania, p. 114–116.
Martinsson, A., Bassett, M.G., and Holland, C.H., 1981, Ratifi cation of standard chronostratigraphical divisions and stratotypes for the Silurian System: Lethaia, v. 14, p. 168, doi: 10.1111/j.1502-3931.1981.tb01923.x.
Martma, T., Brazauskas, A., Kaljo, D., Kaminskas, D., and Musteikis, P., 2005, The Wenlock–Ludlow carbon iso-tope trend in the Vidukle core, Lithuania, and its rela-tions with oceanic events: Geological Quarterly, v. 49, p. 223–234.
McLaughlin, P.I., Cramer, B.D., Brett, C.E., and Kleffner, M.A., 2008, Silurian high-resolution stratigraphy on the Cincinnati Arch: Progress on recalibrating the layer-cake, in Maria, A.H., and Counts, R.C., eds., From the Cincinnati Arch to the Illinois Basin: Geo-logical Field Excursions along the Ohio River Valley: Geological Society of America Field Guide, v. 12, p. 119–180, doi: 10.1130/2008.fl d012(08).
Melchin, M.J., and Holmden, C., 2006, Carbon isotope chemostratigraphy of the Llandovery in Arctic Can-ada: Implications for global correlation and sea-level change: GFF (Geologiska Föreningens i Stockholm Förhandlingar), v. 128, p. 173–180, doi: 10.1080/11035890601282173.
Melchin, M.J., Cooper, R.A., and Sadler, P.M., 2004, The Si-lurian Period, in Gradstein, F.M., Ogg, J.G., and Smith, A.G., eds., A Geologic Time Scale 2004: Cambridge, UK, Cambridge University Press, p. 188–201.
Metzger, R.A., 2005, Conodont biostratigraphy of the Scotch Grove and Laporte City Formations (late Llandovery–early Wenlock, Silurian) in eastern Iowa: Bulletins of American Paleontology, v. 369, p. 93–104.
Meyers, S.R., and Sageman, B.B., 2007, Quantifi cation of deep-time orbital forcing by average spectral misfi t: American Journal of Science, v. 307, p. 773–792, doi: 10.2475/05.2007.01.
Mullins, G.L., and Aldridge, R.J., 2004, Chitinozoan biostra-tigraphy of the basal Wenlock Series (Silurian) global stratotype section and point: Palaeontology, v. 47, p. 745–773, doi: 10.1111/j.0031-0239.2004.00387.x.
Munnecke, A., Samtleben, C., and Bickert, T., 2003, The Ireviken Event in the lower Silurian of Gotland, Swe-den—Relation to similar Palaeozoic and Protero zoic events: Palaeogeography, Palaeoclimatology, Palaeo-ecology, v. 195, p. 99–124, doi: 10.1016/S0031-0182(03)00304-3.

Cramer et al.
1716 Geological Society of America Bulletin, September/October 2010
Noble, P.J., Zimmerman, M.K., Holmden, C., and Lenz, A.C., 2005, Early Silurian (Wenlockian) δ13C profi les from the Cape Phillips Formation, Arctic Canada and their relation to biotic events: Canadian Journal of Earth Sciences, v. 42, p. 1419–1430, doi: 10.1139/e05-055.
Over, D.J., and Chatterton, D.E., 1987, Silurian conodonts from the southern Mackenzie Mountains, Northwest Territories, Canada: Geologica et Palaeontologica, v. 21, p. 1–49.
Patzkowsky, M.E., Slupik, L.M., Arthur, M.A., Pancost, R.D., and Freeman, K.H., 1997, Late Middle Ordovi-cian environmental change and extinction: Harbinger of the Late Ordovician or continuation of Cambrian patterns?: Geology, v. 25, p. 911–914, doi: 10.1130/0091-7613(1997)025<0911:LMOECA>2.3.CO;2.
Põldvere, A., ed., 2003, Ruhnu (500) Drill Core: Estonian Geological Sections, Bulletin 5, 76 p.
Porębska, E., Kozłowska-Dawidziuk, A., and Masiak, M., 2004, The lundgreni event in the Silurian of the East European Platform, Poland: Palaeogeography, Palaeo-climatology, Palaeoecology, v. 213, p. 271–294.
Saltzman, M.R., 2001, Silurian δ13C stratigraphy: A view from North America: Geology, v. 29, p. 671–674, doi: 10.1130/0091-7613(2001)029<0671:SCSAVF>2.0.CO;2.
Saltzman, M.R., 2002, Carbon isotope δ13C stratigraphy across the Silurian–Devonian transition in North Amer-ica: Evidence for a perturbation of the global carbon cycle: Palaeogeography, Palaeoclimatology, Palaeo-ecology, v. 187, p. 83–100, doi: 10.1016/S0031-0182(02)00510-2.
Samtleben, C., Munnecke, A., Bickert, T., and Pätzold, J., 1996, The Silurian of Gotland (Sweden): Facies in-terpretation based on stable isotopes in brachiopod
shells: Geologische Rundschau, v. 85, p. 278–292, doi: 10.1007/BF02422234.
Samtleben, C., Munnecke, A., and Bickert, T., 2000, Devel-opment of facies and C/O-isotopes in transects through the Ludlow of Gotland: Evidence for global and local infl uences on a shallow-marine environment: Facies, v. 43, p. 1–38, doi: 10.1007/BF02536983.
Samtleben, C., Munnecke, A., Bickert, T., and Pätzold, J., 2001, Shell succession, assemblage and species de-pendent effects on the C/O-isotopic composition of brachiopods—Examples from the Silurian of Gotland: Chemical Geology, v. 175, p. 61–107, doi: 10.1016/S0009-2541(00)00364-8.
Scholle, P.A., and Arthur, M.A., 1980, Carbon isotope fl uc-tuations in Cretaceous pelagic limestones: Potential stratigraphic and petroleum exploration tool: American Association of Petroleum Geologists Bulletin, v. 64, p. 67–87.
Skoglund, R., 1979, Graptolites: Sveriges Geologiska Under-sökning, v. C762, p. 213–215.
Talent, J.A., Mawson, R., Andrew, A.S., Hamilton, P.J., and Whitford, D.J., 1993, Middle Palaeozoic extinction events: Faunal and isotopic data: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 104, p. 139–152, doi: 10.1016/0031-0182(93)90126-4.
Teller, L., 1969, The Silurian biostratigraphy of Poland based on graptolites: Acta Geologica Polonica, v. 19, p. 393–502.
Uyeno, T.T., 1990, Biostratigraphy and conodont faunas of upper Ordovician through middle Devonian rocks, Eastern Arctic Archipelago: Geological Survey of Canada Bulletin, v. 401, p. 1–211.
Weissert, H., Joachimski, M.M., and Sarnthein, M., 2008, Chemostratigraphy: Newsletters on Stratigraphy, v. 42, p. 145–179, doi: 10.1127/0078-0421/2008/0042-0145.
Wenzel, B., and Joachimski, M.M., 1996, Carbon and oxygen isotopic composition of Silurian brachiopods (Gotland/Sweden): Palaeoceanographic implications: Palaeo-geography, Palaeoclimatology, Palaeoecology, v. 122, p. 143–166, doi: 10.1016/0031-0182(95)00094-1.
Wenzel, B.C., 1997, Isotopenstratigraphische Untersuchungen an silurischen Abfolgen und deren paläozeanogra-phische Interpretation: Erlanger geologische Abhand-lungen, v. 129, p. 1–117.
Wigforss-Lange, J., 1999, Carbon isotope 13C enrichment in Upper Silurian (Whitcliffi an) marine calcareous rocks in Scania, Sweden: GFF (Geologiska Föreningens i Stockholm Förhandlingar), v. 121, p. 273–279, doi: 10.1080/11035899901214273.
Woodcock, N.H., 2000, Introduction to the Silurian: Geo-logic Conservation Review Series, v. 19, p. 3–22.
Young, S.A., Saltzman, M.R., Bergström, S.M., Leslie, S.A., and Xu, C., 2008, Paired δ13Ccarb and δ13Corg records of the Upper Ordovician (Sandbian-Katian) carbonates in North America and China: Implications for paleocean-ographic change: Palaeogeography, Palaeoclimatol-ogy, Palaeoecology, v. 270, p. 166–178, doi: 10.1016/j.palaeo.2008.09.006.
Zalasiewicz, J., and Williams, M., 1999, Graptolite bio-zona tion of the Wenlock Series (Silurian) of the Builth Wells district, central Wales: Geological Magazine, v. 136, p. 263–283, doi: 10.1017/S0016756899002599.
MANUSCRIPT RECEIVED 22 DECEMBER 2008REVISED MANUSCRIPT RECEIVED 4 JUNE 2009MANUSCRIPT ACCEPTED 18 JUNE 2009
Printed in the USA