telomere dysfunction in breast differentiation, aging and...
TRANSCRIPT

Telomere dysfuydifferentiation, a
David GiDavid GiIndiana University
Department of MedicalDepartment of Medical
unction in breast aging and cancer
illey Ph Dilley Ph.D. School of Medicineand Molecular Geneticsand Molecular Genetics

Part I: The
Consequences of telqin cancer: an
capCapped telome
Uncapping event
Telomere fusion
Breakage-fusion-bridge (BFB) cyc
e hypothesis
lomere dysfunction yn early event?
ere
Genomic instability
cle - McClintock, Genetics,1941

Telomere dysfunTelomere dysfun
Knowledge gap: no ditelomere dysfunction y
A novel assay to detecunderstand and moni
nction in cancersnction in cancers
irect evidence that occurs in human cancers
ct telomere fusions toitor disease progressionp g

Detection and analysNormal
chromosomes
Dicentric chromosomeDicentric chromosome
FusionTAR Fusion PCR
primer
TAR Fusion PCR
telomere adjacent seq.
p
PCR pr
Detec
sis of fusion junction
TelomereTelomeredysfunction
point
primer
telomere adjacent seq.Multiplex: primer sets
p
roduct
ction

TAR-Fusion PCR p-arm(CCCTAA)n
Chr 1
Chr 2
TAR1
TAR1
Chr 4
Chr 5
Chr 7
TAR1
TAR1
Chr 9
Chr 10
Chr 11
TAR1
Chr 12
Chr 16
Chr 15
TAR1
Chr 17
Chr 18
Chr 19
TAR1
Chr 21
Chr X or Y
multiplex primersq-arm (TTAGGG)n
TAR1
TAR1
TAR1
TAR1
TAR1
TAR1
NOTE:TAR-PCR primers cover
40% of ends but onlyTAR1
TAR1
~40% of ends but only…~15% of possible
end-to-end combinations
TAR1
TAR1
TAR1

Telomere fusioni b t t
Primer Mix: A B
5 0(kb)
A B A B A B A
in breast tu
1.01.5
2.03.04.05.0
1 2 3 1
DCIS InvDCIS Inv
40%
50%
P=0.004
usio
n m
ors
10%
20%
30%
Telo
mer
e fu
posi
tive
tum
0%
10%
Normal D(N=24) (N
ns present early i i
A B A B A B A B A B
morigenesis
2 1 23 3
vasive Normalvasive NormalP=0.016
1. ~40% of tumor tissue
2. Only 15% of possible fusionscovered by TAR fusion PCR
3. Stabilization at or before
DCIS InvasiveN=25) (N=23)
DCIS stage

Summary: fusioybreast tum
TAR1 (TTAGGG)n (CCTelomere to Telomere
TAR1 (TTAGGG)nTelomere to subtelomere
Complex TAR1 (TTAGGG)n I
0.6 ~ 2 kb
*Hallmark of sshortened telomeres ans o te ed te o e es a
on junctions in jmor tissue
7.7% (1/13) 6.7% (1/15)
DCIS Invasive
CCTAA)n TAR1
92.3% (12/13)
0% (0/13)
80.0% (12/15)
13.3% (2/15)nsertion
TAR1
TAR1
solid tumors: nd activated telomerased act vated te o e ase

Short insertions of retrt i i b t tat invasive breast tumor
Chr.4p q
Chr.2q p
Chr.17p q
pq
Chr. X or Yp q
XpYp4q
Chr.2q:211 bp Chr.17p:374 bp
SINEXpYp telomere
4q telomere SINE
rotransposon elements ti f i j ti
p q
r tissue fusion junctions
Chr.10
p q
Chr.4
Chr.4
p q
q p
4p10q LTR
Chr.4q: 447 bp
q
4p telomere10q telomere

Conclusions: fusioConclusions: fusio Telomere fusions (dy Telomere fusions (dy
early in human bre
Potentially a highly pmarker for tumoribreast, prostate*, obreast, prostate , o
Tanaka, Abe, Huda, Tu, Beam, Gin early human breas
on junction resultson junction resultsysfunction) foundysfunction) foundeast tumors (DCIS).
prevalent genetic genesis:
ovarian*, CLL, others?ovarian , CLL, others?
Grimes and Gilley, Telomere fusionst carcinoma. PNAS (2012)

Par
Telomere plastTelomere plasthuman mammaryy
Note1) Critical to determine
repopulating subpcomparing to tum
2) Uncovering possibdisease resistance
rt II:
ticity in normalticity in normaly repopulating cellsy p p g
e: the biology of normal
populations before mor subpopulations
ble mechanisms of e and initiation

Human mammary
MammaryMammarystem cell (BC)
Lprog
Bipotentprogenitor (BC)
p g
MyoeProgen
cellular hierarchyCell of origin:
Luminal progenitors??
Luminal genitor (LP) Mature Luminal g ( )
Cells (LC)
epithelialnitors (BC)
Mature Myoepithelial Cells (BC)
(Lim et al., 2009 Nat. Med.)(Molyneux et al., (2010) Cell Stem Cell)

Subpopulatif d tifrom reduction
Enzymaticdissociation
trypsin/dispase/DNase
Si l C llSingle Cells
ion isolationl tn mammoplasty
FACS purification A
M P
E LPLC
CD31- CD45-
EpC
A
BCSC
CD49f APCs

Telomere length plasticity20 ld20 year old
Mammarystem cell (BC)
Bipotprogenito
T
9
10
11
12
ngth
(kb)
Tp <
p < 0.001
5
6
7
8
Telo
mer
e le
n
4
T
BC L
y in isolated breast subsets
Luminal M t L i l
tentor (BC)
progenitor (LP) Mature Luminal Cells (LC)
MyoepithelialProgenitors (BC)
Mature Myoepithelial Cells (BC)
TRF qPCR
9
10
11
12
ngth
(kb)
TRF qPCR0.001
p = 0.004
p = 0.004
5
6
7
8
Telo
mer
e le
n
4
TLP SCLC BC LP SCLC

Age-dependent telomerin the mature lu
2620463146
STLCLPBC
TCEEC
TCEEC
HT1
080
ladd
er
STLCLPBC
STLCLPBC
23.1
(kb)
8.07.0
9.0
6.0
5.0
4.34.0
3.0
2.4
2.0
1.5
1.0
re length shortening uminal cells .
8
10
r 2 = 0.50
b)
6
8
C T
RF
(kb
40 20 40 60 80
age (years)LC
age (years)

Telomerase activation
BC
26
BC
LP LC ST
24
HeL
a
HI CH
APS
HT1
080
BC
LP LC ST BC
LP LC ST
48
TRF2 hTERT merge
BC
LP
in luminal progenitors
LP LC ST
58
p < 0.001
4
8
12
16
RTA
(%)
IC
BC LP SCLC0
R
100o-fo
ci
hTERT / TRF2
0
20
40
60
80
quen
cy o
f co
0
Freq BC LP

Reduced activation of progenitor cprogenitor c
25 (%)
r 2 = 0.54
15
20
lom
eras
e
0
5
10
0 20 40 60 80
elat
ive
Tel
0 20 40 60 80R
age (year)
Telomerase activity decreases with age ofluminal progenitor cells
co
f telomerase in luminal cells with agecells with age
10
8
r 2 = 0.50
F (k
b)4
6
LC T
RF
0 20 40 60 80
age (year)
Telomere lengths decreases with age of differentiated luminal cellsrrelates with:

Telomere-associated DNnormal luminalnormal luminal
LP fraction
BP Fraction
T l iTelomere-associ
NA damage response in progenitor cellsprogenitor cells
MRN complexMRE11Rad50
i t d t i NBS1
RT-PCR confirmation
iated proteins
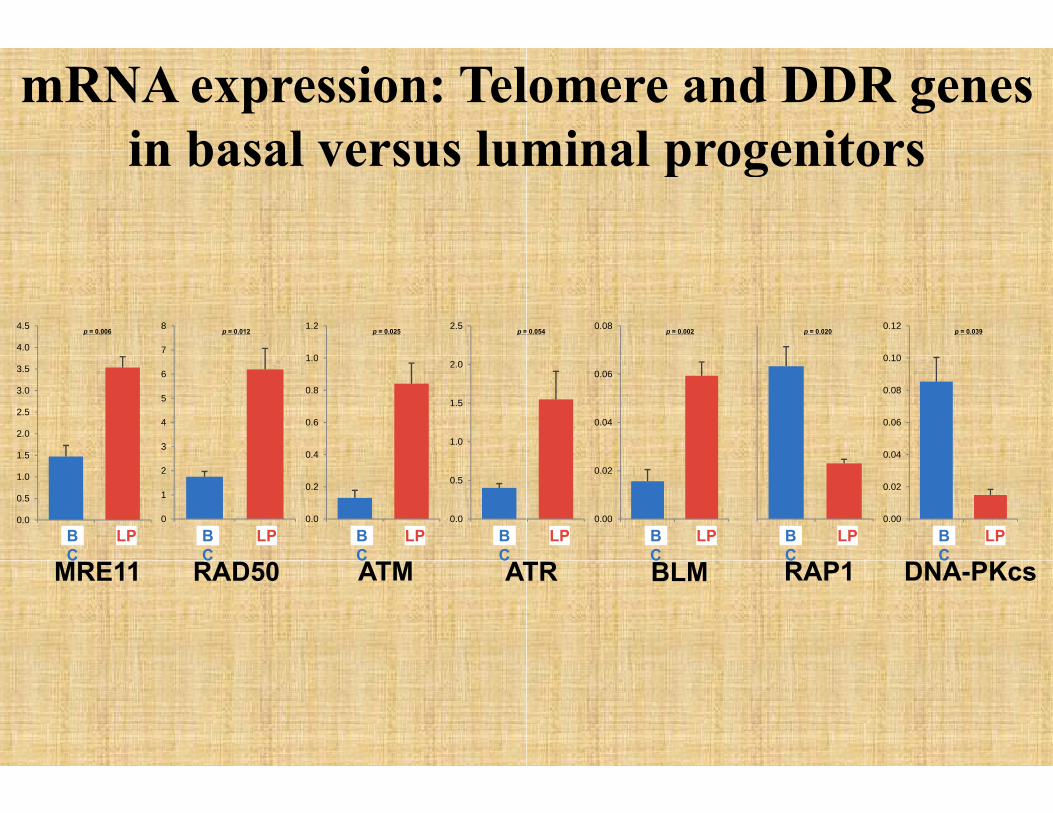
mRNA expression: Telin basal versus lumin basal versus lum
4.0
4.5
7
8
1 0
1.2 2.5p = 0.006 p = 0.012 p = 0.025 p =
1.5
2.0
2.5
3.0
3.5
3
4
5
6
0.4
0.6
0.8
1.0
1.0
1.5
2.0
0.0
0.5
1.0
1.5
0
1
2
0.0
0.2
0.0
0.5
BC
LP BC
LP BC
LP BCC C C C
MRE11 RAD50 ATM AT
lomere and DDR genesminal progenitorsminal progenitors
0.08
0 10
0.12 0.054 p = 0.002 p = 0.020 p = 0.039
0.04
0.06
0.04
0.06
0.08
0.10
0.00
0.02
0.00
0.02
LP BC
LP BC
LP BC
LPC C C
TR BLM RAP1 DNA-PKcs

Telomere dysfunction-indnormal human
TRF2 H2A
normal human
TRF2 γ-H2A
BCBC
LP
Si il lt / 53BP1 NSimilar results w/ 53BP1, N
duced DNA damage foci in mammary LPs
AX M
mammary LPs.
AX Merge
NBS1 MRE11 d RAD50NBS1, MRE11 and RAD50

Telomere-dysfunctyresponse in normal l
100
% c
ells
50
0 3 4 6 7 9 10 1
%
0
0 - 3 4 - 6 7 - 9 10 - 1
# of TIF
tion DNA damage gluminal progenitors
BC LPTRF2 / NBS1
TRF2 / MRE11TRF2 / γH2AXTRF2 / 53BP1TRF2 / RAD5053BP1 / γH2AX
2 13 15 16 18 19 21 22 24
53BP1 / γH2AX
2 13 - 15 16 - 18 19 - 21 22 - 24
Fs/nucleus

stemcell
Nd
bipotentprogenitor
dmature luminal
Longer telomeresdecreasing with age
mature
luminalprogenitor
proliferation
shortened
mature luminal
Telomerase negative
luminaltelomeres
activated telomerase decreasing
with age
Telomere dysfunctioninduced foci
Very
tumorinitiation
yRareCommon
shortentumor
suppression
Normal breast diff ti tidifferentiation:
a doublea double edged sword?edged sword?*Hallmark of solid tumors:
ed telomeres and activated telomerase

Disease consequences ofqCancer: Blood-borne and solid tum
C di l t l h t i
i f
Cardiovascular: telomere shortenin
Environmental factors &(-via telomere shortening, oxidative dama
Exogenous: smoking, obesity, socio-
Endogenous: stress, meditation, exe
Now include
g
f telomere dysfunctionymors (CLL, breast, prostate*, ovarian*)
i t d ith h t i CVD
f i
ng associated with hypertension, CVD
& telomere dysfunctionage, epigenetic remodeling, others)
-economic group, etc…
ercise, etc….
: normal cellular differentiation

Ack
David GilleyDavid Gilley(IUSM)Hiromi TanakaNazmul HudaSatoshi AbeLiRen TuMatthew BeamMatthew RehmelMatthew RehmelGabriela FigueroaKimberly SmithAmanda CampbellAlli B tAllison BatesKimberly Ho A LimZahir Sheikh
nowledgements
Breast subpopulations:Connie Eaves (Terry Fox Lab)Nagarajan Kannang j
Cancer stem
S llStem cell
Progenitor
Differentiated progeny
cell hypothesis
Adapted from Cheng L et al. Toxicol Pathol 2009;38:62-71
