tandem restoration of diadema antillarum and …
TRANSCRIPT

1
TANDEM RESTORATION OF DIADEMA ANTILLARUM AND ACROPORA CERVICORNIS
FOR ENHANCED REEF RECOVERY
By
KAYLA J. RIPPLE
A THESIS PRESENTED TO THE GRADUATE SCHOOL
OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
UNIVERSITY OF FLORIDA
2017

2
© 2017 Kayla J. Ripple

3
ACKNOWLEDGMENTS
I would like to thank all of the organizations and agencies who supported this project.
Without funding and resources provided by the Florida Fish and Wildlife Conservation
Commission, Coral Restoration Foundation, University of Florida, and staff at the Florida
Aquarium’s Center for Conservation, this project would not have been possible.
I would like to thank the members of my committee, Dr. Mark Flint, Dr. Don Behringer,
and Dr. Scott Winters. They have provided me with the resources and support necessary to carry
out this project. Their diverse backgrounds and joint caliber taught me the process and inner
workings of the scientific process. I would like to thank most of all my Committee Chair, Dr.
Mark Flint, for his patience, continuous check-ins, and calming tactics to get me through every
moment of this degree. Mark has taught me the fun side of science, how to tell a story through
my research, and imparted on me his care and kindness with a side of wit.
I would also like to thank my family and close friends for their unwavering support. To
my mother and father, who have supported my dreams and ambitions no matter how crazy. To
my brother and sister who always remind me life should be taken a little less seriously. And to
Jessica and Jana, who kept me sane, listened to my rants, and rejoiced in my eureka moments
and happy times. This was not just a degree or a research project, it was a life lesson and
transforming journey.

4
TABLE OF CONTENTS
page
ACKNOWLEDGMENTS ...............................................................................................................3
LIST OF TABLES ...........................................................................................................................6
LIST OF FIGURES .........................................................................................................................7
ABSTRACT ....................................................................................................................................8
CHAPTER
1 A REVIEW OF THE BIOLOGY, ECOLOGY, AND JUSTIFICATION FOR CORAL-
URCHIN RESTORATION ....................................................................................................10
Introduction .............................................................................................................................10 Biology and Ecology of the Long-Spined Sea Urchin ...........................................................11 The Diadema Die-Off .............................................................................................................14 Rationale for Coral-Urchin Restoration ..................................................................................20 General Hypotheses and Objectives .......................................................................................22
2 HEALTH ASSESSMENTS OF EX SITU EXPERIMENTATION OF DIADEMA
ANTILLARUM AND TRIPNUESTES VENTRICOSUS .........................................................26
Introduction .............................................................................................................................26 Methods ..................................................................................................................................27 Results .....................................................................................................................................29 Discussion ...............................................................................................................................30
3 UNDERSTANDING THE RELATIONSHIP BETWEEN ACROPORA CERVICORNIS
CORAL DENSITY AND JUVENILE DIADEMA ANTILLARUM SURVIVORSHIP .........36
Introduction .............................................................................................................................36 Methods ..................................................................................................................................39 Results .....................................................................................................................................43 Discussion ...............................................................................................................................44
4 PREDICTING SUCCESSFUL DIADEMA ANITLLARUM RELOCATION SITES IN
THE FLORIDA KEYS ...........................................................................................................54
Introduction .............................................................................................................................54 Methods ..................................................................................................................................55 Results .....................................................................................................................................57 Discussion ...............................................................................................................................58
5 DISCUSSION .........................................................................................................................66

5
Diadema antillarum Health and Behavior Parameters Ex Situ: Applications for In Situ
Monitoring and Relocation Efforts .....................................................................................67 Acropora cervicornis Densities to Promote Diadema antillarum Retention .........................70 Applications for Management ................................................................................................71 Future Work ............................................................................................................................74
APPENDIX A: Coral Clusters Present for Urchin Relocations from 2017-2020 .........................76
LIST OF REFERENCES ...............................................................................................................80
BIOGRAPHICAL SKETCH .........................................................................................................91

6
LIST OF TABLES
Table page
2-1 Urchin Health Day 2 ..........................................................................................................35
2-2 Urchin Health Day 3 ..........................................................................................................35
2-3 Shelter Seeking Behavior in D. antillarum and T. Ventricosus .........................................35
3-1 Averages of urchins present within cages during experiment. ..........................................53
4-1 Proportion of Clusters Present for Each Category for Years 2017-2020...........................65
A-1 Clusters Present for Each Category of Cluster-Biomass at Restoration Sites in 2017. .....76
A-2 Clusters Present for Each Category of Cluster-Biomass at Restoration Sites in 2018. .....77
A-3 Clusters Present for Each Category of Cluster-Biomass at Restoration Sites in 2019. .....78
A-4 Clusters Present for Each Category of Cluster-Biomass at Restoration Sites in 2020. .....79

7
LIST OF FIGURES
Figure page
1-1 Urchin grazing effects. .......................................................................................................24
1-2 The spread of Diadema antillarum mortality sightings. ....................................................25
2-1 Grid and design of experimental pools. .............................................................................33
2-3 Urchin abundances across times spent in pools. ................................................................34
3-1 Layout of coral density experimental grid. ........................................................................49
3-2 Cage Design. ......................................................................................................................50
3-3 Average urchins present for time across all treatments. ....................................................50
3-4 Urchins present over time for control cages. .....................................................................51
3-5 Urchins present over time for 10 corals. ............................................................................51
3-6 Urchins present over time for 25 corals. ............................................................................52
3-7 Urchins present over time for 40 corals. ............................................................................52
3-8 Urchins present over time for 55 corals. ............................................................................53
4-1 “Diadema Retention Model” for A. cervicornis restoration sites. .....................................62
4-2 A. cervicornis restoration sites identified in the Florida Keys for preliminary D.
antillarum relocations in 2017. ..........................................................................................63
4-3 A. cervicornis restoration sites identified in the Florida Keys for preliminary D.
antillarum relocations in 2018. ..........................................................................................63
4-4 A. cervicornis restoration sites identified in the Florida Keys for preliminary D.
antillarum relocations in 2019. ..........................................................................................64
4-5 A. cervicornis restoration sites identified in the Florida Keys for preliminary D.
antillarum relocations in 2020. ..........................................................................................64

8
Abstract of Thesis Presented to the Graduate School
of the University of Florida in Partial Fulfillment of the
Requirements for the Degree of Master of Science
TANDEM RESTORATION OF DIADEMA ANTILLARUM AND ACROPORA CERVICORNIS
FOR ENHANCED REEF RECOVERY
By
Kayla J. Ripple
May 2017
Chair: Mark Flint
Major: Fisheries and Aquatic Science
The long-spined sea urchin, Diadema antillarum, is often considered a keystone species
on coral reefs throughout the Caribbean. In 1983, a disease epidemic resulted in the mass
mortality of D. antillarum, correlating with a phase shift of coral to macroalgae dominated reefs.
Since the epidemic, few reefs have shown signs of urchin recovery. Efforts to reverse reefs to
coral dominance have taken place in the form of coral population enhancement programs and
urchin relocation efforts, but the two have yet to be combined in earnest.
This study aimed to understand multi-species reef-restoration techniques, specifically that
of coral-urchin restoration, to promote recovery of both urchin and coral populations in tandem,
and promote reef health. To achieve this, experiments were conducted to assess the general
hypotheses that: (1) relocation of D. antillarum to coral restoration sites does not affect health of
the urchins; (2) with increasing Acropora cervicornis density, D. antillarum retention within A.
cervicornis clusters increases; and (3) with multiple restoration efforts taking place, suitable
habitat currently exists to support D. antillarum relocation.
Urchin health was addressed through the adoption of protocols outlined by Francis-Floyd
et al. (in press) for studies in situ, and showed there were no changes to urchin health when
relocated from one site to another. Coral density manipulation experiments showed that there

9
appears to be an apparent threshold for coral density where certain cluster-biomass is needed to
support >75% urchin retention within A. cervicornis coral clusters. A Diadema Retention Model
was developed to predict cluster-biomass needed to promote urchin retention, and sites were
identified that to begin preliminary coral-urchin restoration efforts in as soon as 2017 and
incrementally until 2020.

10
CHAPTER 1
A REVIEW OF THE BIOLOGY, ECOLOGY, AND JUSTIFICATION FOR CORAL-URCHIN
RESTORATION
Introduction
Coral reefs are an ecosystem of great importance both biologically and economically.
Ecosystem services coral reefs provide globally, have been estimated at $9.9 trillion (Costanza et
al., 2014). Often referred to as the rainforests of the sea, the system fosters great biodiversity
with a multitude of species filling various niches. Populations of different species act in various
roles within the community to promote checks and balances within the system that contribute to
overall health and resiliency of the biome (Tuya et al., 2004; Bellwood 2004; Graham and Nash,
2013). Large reef-building corals are essential to provide continuous habitat and shelter over
time for the diverse group of organisms. In the Florida Keys and Caribbean, the branching corals
Acropora cervicornis and A. palmata, were once the most abundant of reef-builders, providing
three-dimensional habitat and shelter for reef fishes and invertebrates. The intricate morphology
of the thickets they form, complimented with the morphology of massive mounds of boulder and
pillar corals, provides opportunity for a wide range of biodiversity (Gratwicke and Speight,
2005).
Current reef decline debates target climate change as the present critical stressor affecting
reefs today. However, fishing pressures are considered to be the source of early reef decline
(Hay, 1984; Jackson, 2001). In the 1970s, hard coral cover, specifically A. cervicornis and A.
palmata, drastically decreased due to an outbreak of coral disease (Aronson and Precht, 2001;
Miller et al., 2002). Periods of coral bleaching and consistently poor water quality compounded
the issues, as greater coral numbers began to decline.
The long-spined sea urchin, Diadema antillarum, is often considered a keystone species
on coral reefs throughout the Caribbean (Lessios, 1988, Knowlton, 2001; Tuya et al., 2004),

11
contributing to the control of macroalgae abundances and allowing the growth of Caribbean
scleractinian coral species. A mass mortality event of D. antillarum in the 1980’s is thought to
have contributed to the ultimate demise of Caribbean coral reefs (Carpenter, 1990; Knowlton,
1992; Hughes, 1994). The following review provides greater detail for the biology of D.
antillarum, factors contributing to reef decline, management tools for recovery, and rationale to
promote a holistic approach to reef recovery by combining the restoration of both coral and
urchins.
Biology and Ecology of the Long-Spined Sea Urchin
D. antillarum are distinguished from other herbivorous urchins by their dark bodies, long,
ubiquitous spines, and iridescent receptors that allow them to detect differences in light and dark
(Randall et al., 1964). D. antillarum are light sensitive (Millot, 1953, 1954) and exhibit diel
activity patterns preferring dark, sheltered habitats and exhibiting nocturnal behavior to lessen
predation pressures (Thornton, 1956; Randall et al., 1964; Ogden et al., 1973). A number of
anecdotal observations describe the urchins’ spines as exhibiting a diverse array of color
patterns, with juveniles often possessing black and white variegated spines and adults with all
black, or gray spines. Spines are used as protection, and respond by vigorous movement when
exposed to higher light intensities and potential threats such as predators (Millot, 1954).
D. antillarum can be found in multiple tropical habitats including Thalassia testudinum
beds, mangrove propagation roots, sand flats, and, most notably, on coral reefs and inshore
rubble habitats (Randall et al., 1964). In years leading up to the catastrophic die-off of D.
antillarum, urchin populations were considered to be the most abundant herbivorous species on
Caribbean reefs, often deleterious to recreational activities for tourists on reefs (Randall et al.,
1964). At the Virgin Islands Marine Park in St. Thomas, a management plan was even enacted to
remove urchins to improve the experience of visiting sightseers (Kumpf and Randall, 1961).

12
The density of D. antillarum has been shown to be positively correlated with reef
complexity (Lee, 2006; Clemente and Hernandez, 2008; Dame, 2008; Bodmer et al., 2015).
Historically, high D. antillarum densities were correlated with shallow reef areas represented by
high reef complexity comprised of multiple coral species including A. cervicornis, A. palmata,
Orbicella annularis, and Millepora complanta (Weil et al., 1984). Urchins even have the ability
to evaluate crevice quality (Carpenter, 1984) relative to their test size to avoid predator attacks
(Carpenter, 1988), where individuals often exhibit homing behavior to their selected reef crevice
(Tuya at el., 2003). Consequently, high structural complexity is an important characteristic to
maintain urchin populations at a reef site.
D. antillarum are observed as an aggregative species (Randall et al., 1964; Bauer, 1976;
Levitan, 1988), where they can exist individually, or in aggregations of up to 100 individuals
(Randall et al., 1964). They are broadcast spawners expelling their gametes for fertilization
within the water column (Randall et al., 1964; Bauer, 1976). Increased numbers in urchin
aggregations are thought to be correlated with reproductive purposes where urchins form denser
aggregations around the full moon and new moon (Bauer, 1975). Gonadal index has been
positively correlated for females with the lunar cycle where egg development is highest around
the new moon and decreases after the new moon (Bauer, 1975), however, spawning tendencies
appear to be asynchronous where males and females have been documented as spawning across
different time periods (Levitan, 1988).
Population structure for multiple organisms is often determined by food availability and
predation pressure (Holt, 1977; Anholt and Werner, 1995). Like many other species, the local
population of D. antillarum and their habitat preferences are heavily influenced by predator
abundance. With increasing predator abundance, D. antillarum populations tend to decrease,

13
resulting in smaller populations of urchins (Ogden, 1973; Carpenter, 1984). High predation
pressure within reef sites can result in D. antillarum exhibiting high crevice fidelity where
urchins remain in the same crevice during the day, leave the crevice at night to graze, and return
to the same crevice before dusk (Carpenter, 1984; Tuya et al., 2004). This homing behavior may
be an indirect result of the individual to avoid predation pressure and increase their chances of
survival (Carpenter, 1984). When predation pressure is reduced in an area, D. antillarum are less
likely to exhibit nocturnal behavior and crevice fidelity and increase total grazing time (Frike,
1974). This type of behavior has been observed across many Caribbean reefs including reefs of
Jamaica and St. Croix (Miller et al., 2003; Carpenter and Edmunds, 2006), where declines in D.
antillarum predators have been recorded (Hughes, 1994).
D. antillarum, in concert with other herbivorous fish species, consume macroalgae and
expose rock suitable for the formation of crustose coralline algae (CCA) needed to promote
recruitment of reef fish and invertebrate species whose larval metamorphosis and settlement are
queued by CCA (Macintyre et al., 2005; Carpenter and Edmunds, 2006). Their intense grazing
effects can be seen by the clear delineation of “clean” reef substrate where urchins have grazed
and adjacent macroalgae covered substrate where urchins have not grazed (Figure 1.1, Idjadi et
al., 2010). D. antillarum grazing effects can also be seen from satellite images as halo’s around
patch reefs (Ogden et al., 1973).
D. antillarum were once the most abundant reef herbivore in the Caribbean (Lessios et
al., 2001). Their large populations were thought to be attributed to an already declining reef fish
trajectory of both predatory fish which decreased numbers of urchins, and herbivorous fishes
which competed for macroalgae resources on the reef (Hay, 1984). Reef fish populations were
reported as declining before the 20th century due in part to the development of new commercial

14
and recreational fishing techniques (Jackson, 2001). While sportfish were the main target,
multiple herbivorous reef species also dwindled (Paddock et al., 2009). As fish populations
began falling, D. antillarum faced less predation pressure and increased food resources, resulting
in a Type II predator-prey functional response where the urchins took over the role as
predominant herbivore on the reef (Hay, 1984; Carpenter, 1984; McClanahan and Muthiga,
1988; McClanahan et al., 1996). Large populations of D. antillarum kept macroalgal abundances
low, allowing corals to thrive even in the absence of normal herbivorous fish abundances (Ogden
1973; Sammarco, 1980; Carpenter, 1981; Sammarco, 1982). This changed with the mass die-off
in 1983, and the decline has not yet been corrected along the Caribbean reef tract.
The Diadema Die-Off
In 1983, a plague swept through D. antillarum populations resulting in the largest
documented near mass extinction event recorded for a marine species (Figure 1.2; Lessios,
1988). In one year, the plague spread from Panama throughout the Caribbean over an area of
approximately 3.5 million km2 (Lessios, 1988). Up to 93% of the long-spined urchins perished in
most areas of the Caribbean (Lessios, 1988), and there appeared to be no populations untouched
by the plague (Lessios, 2016). The cause of the mortality remains unknown, but is suspected to
be a species-specific water-borne pathogen, whose transmission was facilitated by currents
throughout the Caribbean (Lessios, 1988). In 1987, Bauer and Agerter (1987) received infected
D. antillarum where they were able to isolate and culture Clostridium spp. bacteria. When this
bacterium was injected into healthy D. antillarum, results were very similar to the disease
symptoms that plagued wild populations, possibly implying the bacteria as an agent for the 1983
mass mortality. However, Bauer and Agerter caution highly that these findings did not fulfill
Koch’s Postulates thus do not indisputably demonstrate cause of the mortality event.

15
Before the D. antillarum epidemic, urchin densities were recorded in high abundance
across various areas of the Caribbean. Urchin densities varied across regions from 3-64
individuals/m2 (Bauer, 1980). In the Florida Keys, densities were lower than other areas of the
Caribbean around 4-5 individuals/m2 (Kier and Grant, 1965; Bauer, 1976, 1980), but higher than
present densities at maximums of 0.33 individuals/m2 (Chiappone et al., 2009). It is possible
these high densities were a result of overfishing of predatory and competitive fish species
(Jackson, 2001), however, analysis of the mtDNA region of D. antillarum in the Caribbean,
Atlantic, and of D. mexicanum in the Pacific, suggests that pre-mortality populations were likely
high in abundance even 100,000 years ago (Lessios et al., 2001). It is unclear from the literature,
whether these high population densities contributed to the mass dissemination of the fatal
pathogen in D. antillarum populations, or if similar epidemics occurred in evolutionary history
of the population (Lessios, 1988).
Regardless of cause, the devastating reduction of D. antillarum populations facilitated a
spike in macroalgae abundance across Caribbean reefs, which is strongly correlated with mass
mortality of already failing Caribbean Acroporid populations (Hughes, 1994; Jackson, 2001).
Today, D. antillarum populations are 25 times less dense than populations on reefs before the
mass mortality event (Hughes et al., 2010). This drastic decline in herbivore abundance, coupled
with an increase in macroalgae abundance, signified a possible phase shift on Caribbean reefs
from a coral dominated to macroalgae dominated state (Carpenter, 1990; Knowlton, 1992;
Hughes, 1994). There were no populations left untouched by the die-off (Lessios, 2016), but
recent improvements for populations in Jamaica and Honduras show a decrease in macroalgae
and increase in scleractinian coral recruits (Edmunds and Carpenter, 2010, Bodmer et al., 2015).

16
This evidence suggests that D. antillarum are key in promoting health of the reef as no other
species has been able to fill the niche of D. antillarum for removing macroalgae.
It is increasingly accepted that coral reefs throughout the Caribbean have undergone
phase shifts from a coral-dominated state to a macroalgae-dominated state (Knowlton, 1992;
Hughes, 1994; Jackson, 2001; Bellwood et al., 2004; Maliao et al., 2008). Environmental
conditions can vary overtime, and stochastic events such as hurricanes or disease outbreaks can
cause direct effects to a system, often affecting the populations of an organism (Scheffer et al.,
2001). Factors causing these shifts can allow for simple or difficult reversal based on the type of
perturbation or event that caused the change (Beisner et al., 2003). If the system only has one
stable state it exists in, it is expected to settle back to original parameters after the event,
however, if the system has multiple stable states, it may shift to an alternative stable state
(Scheffer et al., 2001). The prolonged period of a macroalgae dominated state on most Caribbean
reefs has led some scientists to believe reefs now exist in an “alternative stable state”, where
effects of hysteresis may require drastic management measures to overcome its effects and revert
back to coral dominant states (Mumby et al., 2007; Hughes et al., 2010; Fung et al., 2011;
Graham et al., 2013).
As coral cover declines and macroalgae grows, positive feedback loops keep reefs in an
algae-dominant state (Scheffer et al., 2001; Norstrom et al., 2009; Hoey and Bellwood, 2011).
Macroalgae causes direct interference with coral growth, often times overshadowing slower
growing corals and secreting allelopathic chemicals (Lirman, 2001; McCook et al., 2001). Some
algae species competing for space on substrate create unsuitable habitat for coral recruitment,
smothering new coral recruits, or providing no space for fragments to settle and cement
themselves to the substrate (Lirman, 2001). Further, significantly smaller coral populations,

17
suffer from allee effects, decreasing the opportunity for coral species to successfully spawn and
to expand populations (Knowlton, 1992; Williams et al., 2008).
Although declines in coral cover have slowed since the 1980’s, recovery back to a coral-
dominated state is not recorded for most regions of the Caribbean and in the Florida Keys
(Gardner et al., 2003). It is thought that the resilience of these ecosystems has been weakened by
the multiple stressors they face, making them more susceptible to pulse perturbations and
resulting in difficulty reversing back to coral-dominated states (Bellwood et al., 2004). Models
and empirical studies provide considerable evidence that the Florida Keys and areas of the
Caribbean have currently shifted to this alternate state as a direct result of the Diadema die-off
(Mumby et al., 2007; Lessios, 2016).
A decrease in reef-building corals and increase in reef erosion, lessens reef rugosity and
complexity (Alvarez-Filip et al., 2009). This “flattening” of reefs, along with other
anthropogenic stressors such as overfishing, has been linked to a decrease in economically
important fish species and overall deterioration of the biodiverse food web that once existed in
coral-dominant ecosystems (Alvarez-Filip, 2009; Dixson et al., 2014). This current state of
macroalgae dominated substrate offers little to no refuge for D. antillarum individuals, inhibiting
the recovery of populations (Bodmer et al., 2015; Roger and Lorenzen, 2016).
These observations may not encourage optimism for reversion of reefs back to coral
dominance. Demonstrated shifts in other ecosystems however, prove it can be done. Urchin
barrens of kelp forests in California provide a prime example. Sea otters control populations of
the sea urchin Strongylocentrotus droebachiensis which graze on kelp holdfasts releasing
floating kelp into the water column where it then drifts away (Estes and Palmisano, 1974). In the
1800’s, sea otter populations were decimated by fisherman harvesting sea otters for fur-trade,

18
and an explosion of the sea urchin population occurred, resulting in near decimation of kelp
forests and a shift from kelp-dominated state to a barren urchin dominated state, or “urchin
barrens” (1974). Regulations put in place to assist recovery of sea otter populations, and also
increase farming pressure on sea urchin populations has resulted in an increase in the kelp beds
to begin to come back where the distribution of kelp forest habitats has now expanded to one-
third of the habitat it once was (Estes and Palmisano, 1974; Filbee-Dexter and Scheibling, 2014).
Evidence from this shift, show that with proper regulation and conservation of natural resources,
ecosystems are able to revert to a more biodiverse state to support original productivity and
function of the ecosystem.
Overall, D. antillarum populations have shown little recovery since their ecological demise
in the 1980’s with the exception of a few reefs (Hughes, 2010; Lessios, 2016). Their inability to
recover to pre-mortality densities is likely attributed to a multitude of compounding factors. Five
main theories exist for the poor recovery of D. antillarum (Bodmer et al., 2015) including: (1)
increased competition from vertebrate reef herbivores, (2) suppressed recruitment resulting from
natural asynchronous spawning and allee effects, (3) predation pressure driving high mortality,
and (4) loss of reef structural complexity removing microhabitat provision (5) ecological
interactions with heterospecifc echinoids.
D. antillarum have historically interacted with other reef echinoid species either by
complementing grazing activities or competing with urchins for macroalgae resources. The
specifically includes the East-Indian sea egg, Tripnuestes ventricosus (Haley and Solandt, 2001),
and the rock-boring urchin, Echinometra viridis (Shulman, 1990). The sea urchin, T. ventricosus,
feeds on larger macroalgae on reefs, that allows for juvenile macroalgae to recruit to reef
substrate (Haley and Solandt, 2001). This type of algae is the preferred food source for D.

19
antillarum (Haley and Solandt, 2001; Bechtel et al., 2006). Because of great increases in
macroalgae, it is possible T. ventricosus cannot keep up with current levels of macroalgae,
leaving an inadequate food sources for D. antillarum (Liddel and Ohlhorst, 1986; Carpenter,
2005). E. viridis and D. antillarum have been observed as aggressive towards one another, and
can compete for space on a reef (Shulman, 1990). However, there have been no population
spikes in E. viridis since the Diadema die-off, and this is not expected to impact local D.
antillarum population recovery (McClanahan, 1999).
D. antillarum and herbivorous reef fishes are essential for the removal of macroalgae
from reefs to maintain hard coral cover and recruitment. It is suspected that recovery may also be
inhibited by the competition between herbivorous fishes and D. antillarum for food (Robertson,
1991). However, the two can exist harmoniously, and grazing pressure exerted by both are
needed together for maximum macroalgae reduction (Carpenter, 1986). Moreover, studies have
even shown that large populations of D. antillarum can drive down local herbivorous fish
populations (Hay and Taylor, 1985), suggesting urchin populations are able to stabilize and can
be unaffected by normal abundances of grazers in the area.
Despite high fecundity in females who can produce up to one million eggs in a single
spawning event (Levitan, 1989), recruitment of D. antillarum still appears low (Lessios, 2010)
suggesting possible recruitment limitation may also be an issue (Karlson and Levitan, 1989).
Sparse adult populations may contribute to allee effects, resulting in low fertilization success and
the inability to produce enough larvae to overcome typically high mortality in the larval stage
(Levitan, 1995). In the Florida Keys, current D. antillarum distributions are thought to have a
<1% fertilization success rates as opposed to >96% before 1983 (Feehan et al., 2016).

20
D. antillarum have >15 known predators (Randall et al., 1964). Juvenile urchins (20-
30mm maximum test diameter) are prone to increased predation pressure at reef sites, but after
reaching sizes >40mm, D. antillarum are still vulnerable, but less prone to predator attacks
(Clemente et al., 2007). Harborne et al. (2008) found that increased fish biomass of species
known to feed on D. antillarum is linked with smaller urchin populations inside marine protected
areas on Bahamian reefs, demonstrating how predation pressure can impact urchin abundances.
Greatest recovery of D. antillarum populations have been documented in sheltered
lagoonal areas and back reef habitat where these populations were also once abundant (Miller et
al., 2003, Debrot and Nagelkerken, 2006; Steiner and Williams, 2006; Vermeij et al., 2010). This
is likely a result of the increased habitat complexity offered in these areas (Rogers & Lorenzen,
2016). Greater habitat complexity is linked to larger D. antillarum populations (Clemente and
Hernandez, 2008). Increasing habitat structure at a reef site can also increase retention of
relocated D. antillarum (Dame, 2008). With decreased hard coral populations, there is a
documented decrease in reef structural complexity throughout Florida and the Caribbean
(Alvarez-Filip et al., 2009). It is possible, predation pressure, coupled with lack of structural
complexity is a major rate-limiting step in recovery of D. antillarum populations (Bodmer et al.,
2015).
Rationale for Coral-Urchin Restoration
The benefits of healthy D. antillarum populations are abundant. Their ability to reduce
macroalgae abundance at reef sites is important for promoting the dominance of coral cover,
which in turn, promotes reef biodiversity. Recovery of D. antillarum is potentially the largest
driving factor in coral reef recovery (Edmunds & Carpenter, 2001). Scarids and other
herbivorous fishes are responsible for only a proportion of grazing pressure on reefs (Mumby,
2006). Increased herbivory from Scarids may even cause harmful effects to existing coral

21
populations by inhibiting the healing of hard corals from parrotfish scrapes in areas with poor
water quality and eutrophication on reefs from agricultural runoff (Zaneveld et al., 2016). D.
antillarum proves to be beneficial for removing fast-growing macroalgae enhancing coral growth
on reefs (Hernandez et al., 2008). Increasing abundance of D. antillarum at reef sites may break
the positive feedback loop by decreasing the abundance of macroalgae at reef sites (Chiappone et
al., 2001; Burdick, 2008; Nedimyer and Moe, 2011) increasing coral cover (Edmunds and
Carpenter, 2001; Idjadi et al., 2010), which in turn can increase reef complexity necessary to
increase D. antillarum abundances (Sammarco, 1982; Clemente and Hernandez, 2008; Bodmer
et al., 2015).
Since their die-off in the 1980’s, D. antillarum population enhancement efforts have
taken place in the form of preliminary relocation trials at reef sites (Burdick, 2008; Nedimyer
and Moe, 2011), development of comprehensive strategies for relocation (Hunt and Sharp,
2014), and workshops to promote ex situ production for relocation efforts (Diadema Workshop,
2017). Preliminary relocation efforts were successful in reducing macroalgae cover at reef sites,
but these population numbers ultimately disappeared (Burdick, 2008; Nedimyer and Moe, 2011).
Low urchin retention rates at relocation sites were attributed to the low complexity and predation
pressure.
Coral restoration efforts may provide a solution for low reef complexity. In 2012, over 60
programs were in place to restore degraded reef sites at 14 countries (Young et al., 2012). These
programs focus on several threatened species of corals, including Acropora species which, grow
quickly in nursery programs. As coral restoration programs become well established, best
practices are better understood, as well as management strategies for reef recovery. The NOAA
Acropora Recovery Plan provides an outline of criteria needed for the species to recover over the

22
long-term (2015). It is clear that without addressing environmental concerns such as climate
change, overfishing, poor water quality, and increased macroalgae abundance, coral reefs cannot
recover to their full potential. While not historically associated with thickets of A. cervicornis, in
the absence of more favorable coral species like A. palmata, Orbicella annularis, and Millepora
complanta (Weil et al., 1984), A. cervicornis, when configured in the right densities, may act as
an alternate habitat for D. antillarum. Coupling the relocation of D. antillarum individuals with
already existing A. cervicornis restoration sites could possibly lead to enhanced reef recovery,
where A. cervicornis thickets can provide shelter to urchins and urchins can reduce macroalgae
to increase coral growth and health at the restoration site.
General Hypotheses and Objectives
This study aims to support the necessary steps to generate an effective coral-urchin
restoration strategy for coral reefs in the Florida Keys. To achieve this, experiments were
conducted to assess the following general hypotheses and objectives for each:
Hypothesis 1: Relocation of D. antillarum to coral restoration sites does not affect health
of the urchins and its objectives (a) determine the parameters necessary to promote D. antillarum
health in an ex situ environment; (b) understand potential habitat usage and behavior of D.
antillarum and T. ventricosus in a controlled environment when compared to expected normal
behavior; (c) understand proper health metrics for monitoring D. antillarum health for in situ
relocations and studies.
Hypothesis 2: With increasing A. cervicornis density, D. antillarum retention within coral
clusters increases and its objective (a) determine the coverage of A. cervicornis coral needed to
promote urchin retention over time when urchins were relocated to coral clusters.
Hypothesis 3: With multiple restoration efforts taking place, suitable habitat currently
exists to support D. antillarum relocation and its objectives (a) predict and identify current

23
restoration efforts that may facilitate the recovery of urchins by selecting suitable relocation to
reefs with appropriate coral infrastructure (coverage); (b) create a tool for managers charged with
implementing coral restoration programs in conjunction with D. antillarum recovery programs.
For the first hypothesis, experiments were conducted at The Florida Aquarium’s Center for
Conservation in Apollo Beach, FL. Habitat preference and health were assessed during these
experiments. Objectives 1a and 1b were partially achieved where we observed parameters that
may potentially lead to demised urchin health ex situ, and observed abnormal shelter-seeking
behavior. Results from urchin behavior were inconclusive however, as abnormal behavior may
have been attributed to urchin health. Objective 1c was achieved through the adoption of health
assessment protocols outlined by Francis-Floyd et al. (in press), for aquacultured urchin release
onto reef sites, and demonstrated applicability in the field during in situ trials.
Caging experiments in the Coral Restoration Foundation coral nursery offshore
Tavernier, FL tested the second hypothesis. Objective 2a was achieved by manipulating coral
densities from low to high (0-55 corals/0.25m2). A trend was discovered for apparent thresholds
of A. cervicornis cluster-biomass that can promote stable urchin retention rates >75%, and
provide a baseline for coral-urchin restoration projects to test in future work.
Coral densities and their associated urchin retention rates gathered from coral density
manipulations were used to generate a Diadema Retention Model to achieve Objective 3b, which
was used to predict coral densities needed to promote 75-100% urchin retention when urchins
were relocated to A. cervicornis clusters to achieve Objective 3a. The model allowed the
identification of coral clusters that currently exist on reefs of the Florida Keys from 2017-2020
so that preliminary urchin relocation trials may be tested in future work to achieve.

24
Figures
Figure 1-1: Urchin grazing effects. The delineation between urchin grazing zones and their
boundary limits on a reef in Discovery Bay, Jamaica (After Idjadi et al., 2010).

25
Figure 1-2: The spread of Diadema antillarum mortality sightings. The spread of Diadema
antillarum mass mortality through the Caribbean in relation to surface currents. The
direction of spread was deduced from the timing of outbreaks at each locality. 1:
Panama, 2: Curacao, 3: Tobago, 4: Barbados, 5: Jamaica, 6: Flower Garden Banks, 7:
St. Croix, 8: St. Thomas. 9: Bermuda (After Lessios, 1988).

26
CHAPTER 2
HEALTH ASSESSMENTS OF EX SITU EXPERIMENTATION OF DIADEMA ANTILLARUM
AND TRIPNUESTES VENTRICOSUS
Introduction
Current plans to contribute to the recovery of Diadema antillarum are widespread and
include the collection and relocation of natural D. antillarum recruits on reefs, and aquaculture
for restocking efforts. While the ex situ aquaculture provides a potential source of tens of
thousands of larvae each batch, the health risk of introducing these animals to natural reefs and
the effect artificial environments have on natural feeding and predator avoidance behaviors of
the urchins is unknown. To evaluate viability of aquaculture raised urchins, a health assessment
was developed by Francis-Floyd et al. (in press) to provide some degree of confidence that
cultured sea urchins are not introducing pathogens to already compromised reef ecosystems. This
work serves as an assessment tool for management agencies as efforts move forward in on-land
mass production.
Breeding programs that exist for population enhancement of terrestrial species have
demonstrated that animals raised in captivity must undergo an imprinting stage where they learn
behaviors needed for survival when released into the wild. Breeding, imprinting, and release
techniques are well documented for avian species like the Andean Condor (Wallace and Temple,
1987). Sport fish restoration programs also demonstrate the need to initiate wild instincts and
homing behavior for fish bred in aquaculture programs and released for re-stocking efforts
(Hendricks et al., 2002). The effect aquaculture environments have on the feeding and predator
avoidance behavior of sea urchins for use in population enhancement programs has not yet been
comprehensively assessed. It has been anecdotally reported that long-spined sea urchins held in
captivity for prolonged periods of time become habituated to their environment and actively seek
food from handlers and do not seek shelter to avoid threats (Sharp and Delgado, pers. comm.).

27
As such, there is a concern that breeding sea urchins in artificial environments will remove their
basic instincts to forage and seek shelter. Equally important, there is a risk that land-based
experimentation to determine home range, feeding behavior, sheltering patterns, and threat
response are not reliable with habituated subjects.
Objectives of this study were to determine the parameters necessary to promote D.
antillarum health in an ex situ environment, understand habitat usage and behavior of D.
antillarum in a controlled environment when compared to expected wild behavior, and
understand proper health metrics for monitoring D. antillarum health for in situ relocations and
studies.
Methods
Experiments took place at The Florida Aquarium’s Center for Conservation (CFC)
greenhouse in Apollo Beach, FL over the winter of 2015. Tripnuestes ventricosus were cultured
by Mr. Martin Moe in Islamorada, FL, and D. antillarum were caught on patch reefs off
Marathon, FL by the Florida Fish and Wildlife Research Institute and obtained under the
authorization of Florida Keys National Marine Sanctuary.
Sixteen pools were established in a greenhouse, filled with clean seawater imported from
the Gulf of Mexico, and aerated using individual bubbler systems. All pools were independent of
one another to ensure separation of replicates. Water quality was assessed by The Florida
Aquarium to ensure water was approximating natural sea water parameters and was free of
pathogens. In twelve pools, one staghorn structure (~50cm in diameter), and one planter pot
structure were placed on one end of the pool each with a food source (Gracilaria tikvahiae and
Acanthophora spicifera) (Figure 2-1). Four pools were left empty as controls. Twenty adult T.
ventricosus and six adult D. antillarum were available at CFC for study. Ten adult T. ventricosus
and six adult D. antillarum were added separately to each pool at the beginning of the study (one

28
urchin per pool). Urchins were assigned randomly to a pool and placed at the opposite end of the
coral and pot structures. As T. ventricosus began exhibiting signs of ill health, they were replaced
with a new urchin for a total of 24 urchins used throughout the study (18 T. ventricosus and 6 D.
antillarum).
Routine water changes were performed daily, and urchin fecal pellets were removed
through siphoning. Food was provided to urchins on the second day of experiments and secured
in the middle of the pools by use of a clothes peg tethered to a dive weight. Food was replaced
every day regardless of grazing or no grazing to ensure stable water quality and an equal fresh
supply of food across pools and treatments. Temperatures were recorded daily using a point and
shoot laser thermometer and a data-logger (LabJack, Digit-TL) submerged in one pool.
Shelter seeking behavior was recorded as “uncovered” if urchins were found in open
space of the pool with no surrounding cover, “under coral” if urchins were under the coral
structure, and “shaded” if urchins were within the flower pot structure, or on shaded edges of the
pool. The average percent of time and standard deviation for time spent in each location for Days
1, 2, and combined averages were compared in a table.
Urchin health criteria was adopted from the protocol established by Francis-Floyd et al.
(in press). Health categories were recorded as spine loss, test lesions, and urchin appearance
(alert and active vs. lethargic and minimally responsive). Urchins were removed as signs of poor
health became apparent. Appearance was classified as “normal”, “poor”, or “very poor”.
“Normal” appearance was defined as normal spine position, no spine loss, or normal body
position. “Poor” appearance was identified through spine loss or drooping spines, and “very
poor” appearance was assigned when the urchin exhibited multiple spine loss (>30%), test
lesions, or had abnormal body position (upside-down, or on their side). Observed urchin health

29
and expected urchin health was analyzed using a chi-square test of contingency for Day 1, Day 2,
and Day 3 of the experiment.
Urchins were observed the first hour that they were introduced to the tank and
observations recorded. On Days 1-2, urchins were observed at 800, 1200, and 1600 for their
selected habitat in the pool (uncovered, under coral, or shaded). Urchin location was recorded
again on Days 5-6, and on Day 7, the trials were ended due to poor urchin health.
Results
Temperatures of the pools fluctuated between 2-4ºC every 24-hours throughout the
experiment and steadily declined over the seven days of the trial (Figure 2-2). Urchin
replacements took place as of Day 3 of the experiment (Figure 2-2), and very few urchins
remained in pools more than 3 days. The count of urchins remaining in pools at 24h time
intervals decreased as the number of days increased (Figure 2-3). D. antillarum numbers
remained stable at six urchins until Day 6 when all urchins were removed and 0 remained. T.
ventricosus urchins were replaced on Day 3, and abundances dropped by 61% for urchins held in
the pools for 4 or more days. No urchins survived 7 days in the pools, and only 11% of total T.
ventricosus survived to Day 6 (Figure 2-3).
Urchin shelter seeking behavior was variable across all treatments where mean percent of
time spent overlapped across treatments (Table 2-3). D. antillarum spent an average of 31.1%
30.2 of time uncovered, 6.7% 22.1 under coral, and 61.7% 34.8 of time in shaded habitat
(Table 2-1). T. ventricosus spent 32.3% 30.4 of time uncovered, 8. 6% 24.3 under coral, and
58.4% 36.3 in shaded habitat (Table 2-3).
On Day 1 all urchins appeared in normal health. On Day 2, 1 urchin exhibited signs of
very poor health, but there was not a significant difference in health amongst all urchins

30
(2=1.011, p= 0.3148) (Table 2-1). The first urchin removal event took place on Day 3 where 6
urchins exhibited signs of poor health. On Day 3, urchins exhibited significant differences in
expected normal health and those observed with normal health, where 6 urchins appeared to be
in poor health, and 10 urchins remained healthy (2= 7.385, p= 0.0066) (Table 2-2).
Discussion
We were able to partially fulfill objectives to determine the parameters necessary to
promote D. antillarum health in an ex situ environment, understand habitat usage of D.
antillarum in a controlled environment when compared to expected normal behavior, and
understand proper health metrics for monitoring D. antillarum health for in situ relocations and
studies. Due to a sudden decline in urchin health on Day 3 of the study, conclusions drawn from
analysis are speculative, and require further investigation to draw substantive conclusions.
Techniques for monitoring urchin health were adopted from Francis-Floyd et al. (in press) and
were used in later field experiments demonstrated in the following chapter. This validated that
these adopted protocols for ex situ assessment can serve as a standardized data collection tool for
coral-urchin restoration programs monitoring urchin health in field studies.
D. antillarum shelter-seeking behavior is typically associated with shaded habitats
(Randall et al., 1964; Carpenter, 1984). This shelter-seeking behavior can be altered in the
absence of predators, where urchins remain in the open taking advantage of grazing time
(Carpenter, 1984). Upon movement to a new environment in situ, D. antillarum aggressively
seek shaded shelter (pers. obs.). Urchins in this study showed differences in percent of time spent
in habitats, which were highly variable, suggesting expected natural behaviors are not being
exhibited in an artificial environment. Urchin behavior in artificial environments has been
observed as dissimilar to wild urchin behavior (Sharp and Delgado, pers. comm.). All urchins

31
used in this study had been long-term residents of artificial environments. Their shelter-seeking
behavior was not consistent nor showed patterns of preferred habitat throughout the trial. It is
possible they did not seek shaded shelter in the absence of predators, but evidence to support this
is minimal, and therefore cannot be conclusive. Observed abnormal behavior may also be a
product of compromised urchin health, so these conclusions are suggestions for further studies.
These experiments subjected urchins to multiple parameters that did not align with natural
environment measurements. Urchins are sensitive to harmful effects of metals and other
toxicants in the environment (Kobayashi, 1980; Bielmyer et al., 2005). Pools were washed
thoroughly before use in experiments, however, it is possible that chemicals from the lining of
the pool may have contributed to poor urchin health on Day 3 of the experiment as well.
Experimentation outside of normal temperature ranges and stark artificial environments did not
produce normal behaviors for both T. ventricosus and D. antillarum and may induce health
issues not expected in a natural environment. It is suggested that great care must be taken when
designing artificial holding tanks for either D. antillarum experimentation aiming to predict
animal response out on the reef, and when developing mass production aquaculture plans.
Effects of temperature on urchin health ex situ are well documented for the development
of gonads within many urchin species (McBride et al., 1997; Spirlet et al., 2000; Lawrence et al.,
2009), however very little peer-reviewed literature exists to document culture of D. antillarum
and effects of sharp temperature changes in the environment. Lawrence et al. (2009) found that
temperature has insignificant effects on the survival and metabolic functions of the urchin
species Strongylocentrotus intermedius, in culture. These experiments were conducted using
gradually adjusted temperatures however, dissimilar to the drastic lulls in temperature
experienced during the cold snap during this experiment. Average water temperature of tropical

32
coral reef ecosystems ranges from 23-29ºC annually and does not typically fluctuate more than
1-2ºC in a 24h period, rather fluctuates over longer periods of time in weeks or months (Lee and
Williams, 1999). While temperatures in the pool remained within the normal tropical reef
temperature ranges, they did undergo greater than natural diurnal fluctuations. Despite attempts
to warm the greenhouse with external heaters to counteract a cold snap experienced during the
trial, pool temperatures fluctuated between 2-4ºC in less than a 24h period each day of the
experiment. It is suspected that this great range in temperature fluctuations may have contributed
to possible health deterioration; for example- hypothermic shock. Attempts to stabilize water
temperatures of the ex situ environment be a priority in maintaining urchin health. Multiple
variables exist on reefs where urchin relocations may take place, and a comprehensive
understanding of factors affecting the survivorship of urchins is essential when designing land
based experiments and aquaculture facilities for these species.
This experiment highlighted the need to closely approximate the target natural
environment of the Florida Keys when considering ex situ rearing efforts to enhance D.
antillarum stock for use in coral-urchin restoration programs. Further, it highlighted the health
assessment protocols established for release of D. antillarum onto restoration sites (Francis-
Floyd et al., in press) are a viable source for monitoring health of experimental animals. These
monitoring protocols can also serve as a baseline for monitoring urchin health at restoration sites.
Outcomes of this experiment demonstrate the vital nature of documenting health in any scenario
where urchin relocations are tested.

33
Figures
Figure 2-1: Grid and design of experimental pools.

34
Figure 2-2: Temperature of urchin holding pools during experiment. Temperature of pools
fluctuated between 2-4ºC over a 24h period and declined overtime.
Figure 2-3: Urchin abundances across times spent in pools. 96-100% of total urchins remained in
pools for 3 days. Urchin abundances dropped at Day 4, and no urchins survived in
pools for 7 days.
0
5
10
15
20
25
30
1 2 3 4 5 6 7
Num
ber
of
Urc
hin
s
Days in Pool
Total Tripnuestes Diadema

35
Table 2-1: Urchin Health Day 2
Expected Observed Total
Normal 16 15 31
Very Poor 0 1 1
Total 16 16 32
Chi-squared table of observed urchin health versus expected urchin health (2=1.011, p=
0.3148).
Table 2-2: Urchin Health Day 3
Expected Observed Total
Normal 16 10 26
Poor 0 6 6
Total 16 16 32
Chi-squared table of observed urchin health versus expected urchin health (2=7.385, p=
0.0066).
Table 2-3: Shelter Seeking Behavior in D. antillarum and T. Ventricosus
Species Day Uncovered Under Coral Shaded
D. anti
llaru
m
Day 1 28.6 (22.10) 0 (0) 72.9 (22.68)
Day 2 31.0 (38.0) 11.9 (31.0) 55.0 (43.1)
Total 31.1 (30.2) 6.7 (22.1) 61.7 (34.8)
T. ve
ntr
icosu
s
Day 1 31.1 (23.5) 2.2 (8.6) 67.3 (26.6)
Day 2 35.6 (36.7) 8.9 (23.5) 53.3 (42.0)
Total 32.3 (30.4) 8.6 (24.3) 58.4 (36.3)
This table presents the average percent and standard deviation in parentheses of time urchins
spent across three habitat options within pools.

36
CHAPTER 3
UNDERSTANDING THE RELATIONSHIP BETWEEN ACROPORA CERVICORNIS CORAL
DENSITY AND JUVENILE DIADEMA ANTILLARUM SURVIVORSHIP
Introduction
The 1980s mass mortality event of Diadema antillarum coincided with a stark decline in
scleractinian coral cover and a sharp increase in macroalgae on reefs around the Caribbean
(Jackson, 2001). As hard coral cover declined and macroalgae increased, bare reef structure was
eroded away, resulting in a shift from high structural complexity reefs to more homogenous, low
complexity reefs dominated by macroalgae. Many reefs have been “flattened” through decrease
in structural complexity (Alvarez-Fillip et al., 2009) leaving poor habitat for fish and invertebrate
populations (Dixson et al., 2014). Over three decades after the mass mortality of D. antillarum,
many countries have reported slow or no recovery of the populations to pre-mortality densities.
Amongst other theories, reduced habitat complexity and opportunistic predation pressure may be
linked to slow recovery (Bodmer et al., 2015).
D. antillarum require enough reef rugosity to provide crevices or ledges to escape from
predators (Levitan and Genovese, 1989). D. antillarum are preyed upon by many fish and
invertebrate species including triggerfish, parrotfish, wrasses, and lobsters (Randall et al., 1964).
Numerous studies suggest that sea urchin populations are heavily structured by predation
pressures within the system, where increased predation pressure can decrease urchin populations
and vice versa (McClanahan and Muthiga 1989; Hereu et al., 2005, Harborne et al., 2009).
Decline in fish populations, due to overfishing, have shown to be correlated with an increase in
D. antillarum abundance and an increase in grazing at reef sites (Carpenter, 1984). It is possible
that predation pressure coupled with lack of reef habitat is attributed as one of the bottlenecks in
D. antillarum recovery.

37
While recovery is minimal in many areas of the Caribbean, some countries have
documented recent increases of D. antillarum individuals recolonizing their reefs. Jamaica and
the Bay Islands of Honduras have reported increased densities of D. antillarum in areas such as
Discovery Bay, Jamaica and Banco Capiro, Honduras (Edmunds and Carpenter, 2001; Bodmer
et al., 2015). Lee (2006) found that habitat complexity and D. antillarum density were positively
correlated on reefs in Jamaica and suggested that low habitat complexity may be one explanation
for the failure of D. antillarum populations to recover to pre-mortality densities. However, in
Discovery Bay, Jamaica, densities of D. antillarum have increased 10-fold with an inversely
correlated decrease in macroalgae, and resultant correlation of an increase in juvenile corals up
to 11-fold (Carpenter, 2001). Recovery of D. antillarum populations in this region are likely
attributed to depauperate fish populations from overfishing, releasing urchins of the need to seek
shelter and avoid predation allowing for exploitation of resources and increased urchin densities
(Hughes et al., 1994).
Bodmer et al. (2015) used the isolated population boom of D. antillarum individuals off
Banco Capiro, Honduras to study factors that may have contributed to an increase in D.
antillarum individuals at depths of 10m and 15m. On Banco Capiro, D. antillarum individuals
were found at 225.67 ± 26.06 100/m2 and 73.56 ± 19.11/100m2respectively, while at a nearby
reef off Utila, abundances ranged from 1.22 ± 0.43 and 0.33 ± 0.17/100 m2. Banco Capiro was
more rugose, suggesting that amongst other factors, structural complexity may be a driving
factor in D. antillarum adult population recovery.
Increased reef complexity may protect smaller D. antillarum from local predation
pressures and increase their likelihood of surviving to less predator prone sizes. D. antillarum
with a test diameter <20mm are most prone to predation while larger urchins with a test diameter

38
>40mm seem to be less affected by predation (Clemente et al., 2007). Bodmer et al. (2015)
recorded large abundances of juvenile D. antillarum individuals across two Honduran reefs, but
no resultant significant increase in adult populations the following years. This suggests that
recruitment is occurring and is not a significant bottleneck for population recovery, but rather
smaller urchins, not surviving into adulthood because poor complexity and high predation
pressure is a major factor in slow recovery of populations.
In the Florida Keys, D. antillarum populations experienced mass mortalities from 1983-
84, and another disease outbreak in 1991 (Forcucci, 1994). These recurring outbreaks
contributed to slow recovery rates (1994). Little data was collected before 1983 in the Florida
Keys, but what was collected, was able to be reconstructed and analyzed to show that urchins
abundances were high across reefs in the Florida Keys, similar to the rest of the Caribbean
(Kissling et al., 2014). In some cases, D. antillarum densities were greater than 7.9
individuals/m2 (Kier and Grant, 1965; Bauer, 1980; Forcucci, 1994; Kissling et al., 2014), but
these dropped and remain since 2011 <1 individual/m2 post-mortalities (Chiappone et al., 2011).
Furthermore, Chiappone et al. (2009) reported a predominance of smaller D. antillarum test sizes
between 1999-2007, which are similar to the findings of Bodmer et al. (2015) for Honduran
reefs, supporting the theory there is poor survivorship of juveniles into adulthood for D.
antillarum populations in the Florida Keys.
Increasing habitat complexity may increase the survivorship of juveniles into adulthood,
helping to re-establish adult population sizes. While decreased habitat complexity is common
throughout the Caribbean (Alvarez-Filip et al., 2009), coral restoration efforts may provide
opportunity to improve habitat complexity by adding coral to sites where it has diminished.
When structural complexity was enhanced at reef sites, urchins relocated to the site showed

39
greater retention rates and decreased the amount of macroalgae in the area (Macia et al., 2007).
Establishing a targeted threshold of habitat complexity would be desirable for reef managers
when planning A. cervicornis coral restoration efforts may help enhance D. antillarum
populations when relocated to these sites.
Coral Restoration Foundation (CRF) has outplanted over 40,000 A. cervicornis clusters to
>20 reef sites in the Florida since 2000. CRF’s outplanting method involves outplanting ten or
more A. cervicornis corals over an area of approximately 1m2 (CRF, unpublished data). Multiple
clusters are outplanted to the same general area of a restoration site to increase coral cover of the
area. Anecdotal results reported by CRF suggest this method of outplanting provides opportunity
for multiple coral thickets to develop and persist over time, increasing the structural complexity
of the area, and possibly allowing re-establishment of habitat suitable for D. antillarum
relocation. The objective of this experiment was to determine the coverage of A. cervicornis
coral needed to promote urchin retention over time when urchins were relocated to coral clusters.
Methods
To assess the relationship between coral density and the retention of juvenile D. antillarum
(20-30mm test diameter) over time when relocated to coral plots, experiments were performed in
a moderately controlled environment at the Coral Restoration Foundation coral nursery offshore
Tavernier, Florida (N24°58'55.60, W080°26'12.11) approximately 30ft in depth. Preliminary
trials were conducted to understand optimal cage design that would allow urchins to inhabit the
cages and reduce risk of escape. Cages were installed in a 3 x 5 grid and spaced 2m apart on each
side (Figure 3-1) and constructed of double-sided chicken wire panels to decrease hole size. Each
cage was 50cm x 50cm, constructed on land and taken to the nursery, cages were zip-tied to 1m
rebar rods that had been installed in a sandy bottom area just outside the CRF nursery
approximately 30ft deep (Figure 3-2). Cages were constructed of chicken wire on four sides and

40
left open at the top to allow predator access to urchins, but limit urchins from dispersal away
from the coral cluster. In preliminary trials, corals experienced high mortality, and urchins
immediately sought to escape cages in the sandy bottom habitat. To elevate corals and urchins
off the sand, cement blocks, previously used for coral propagation, were added to each cage
(Figure 3-2A). The blocks were originally created for coral propagation and growth, but were
since retired. The platform of the blocks was made of cement with twelve 15cm PVC pipes
cemented into the concrete. The blocks were placed with the PVC side down creating a table-like
platform for corals. Coral density treatments were randomly assigned a cage number, and the
assigned coral density was added to each cage using medium sized A. cervicornis corals (~30cm
TLE) from the CRF coral nursery.
Four coral density treatments (10, 25, 40, and 55 corals/0.25m2) were replicated three times.
These densities are visually assessed at approximately 30, 50, 80 and 95% ground coverage. An
additional three control treatments were included in the experiment with a coral density of 0
corals/0.25m2. Coral densities selected for this experiment began at 10 corals and were scaled up
to test for the optimal A. cervicornis cluster density needed to enhance retention of small (2-3cm
test diameter) D. antillarum when relocated to A. cervicornis coral clusters at established
restoration sites. Minimum densities were based on current restoration practices of planting 10 A.
cervicornis fragments of 30cm TLE, clustered approximately over an area ~1m2 to form dense
continuous thickets of A. cervicornis over time.
Urchins were collected from a rubble patch inshore of Pickles Reef off Tavernier, Florida
(N2459.180’, W08025.126’) under permits designated by the Fish and Wildlife Conservation
Commission (SAL-16-1722-SCRP) and Florida Keys National Marine Sanctuary (FKNMS-
2015-028-A2). All urchins were collected and relocated to the experimental cages on the same

41
day. General characteristics of the collection site were taken including site substrate (rubble,
sand, hard bottom), depth, tidal flow (incoming, outgoing, high tide, low tide), and surface and
water temperature as outlined in the health assessment protocol for release of D. antillarum onto
reefs (Francis-Floyd et al., in press). All urchins were assessed for tissue loss, spine loss, normal
spine movement and normal body position before they were collected to ensure that urchins were
“healthy” upon relocation to the nursery (Francis-Floyd et al., in press). Each urchin was
measured using a plastic ruler, and only those that were 2-3cm in test diameter were included in
the study. Four urchins were added to each cage and monitored over 20 days when weather was
appropriate, for health condition, behavior, movement and urchin counts within coral, under
blocks, and total present within the cage on Days 1-20.
Health of the urchins was assessed at the time of collection and throughout the trial to
understand how urchin health was affected by collection and relocation methods. Categories of
health assessment criteria were selected from the D. antillarum Health Assessment developed for
FWC by Francis-Floyd et al. (in press). The following criteria were visually assessed, any
urchins exhibiting abnormal behavior or appearance were not selected for the trials: 1) Spine
position: healthy urchins should have spines erect and extended, “drooping” spines were
considered abnormal, as they are indicative of a disease. 2) Spine movement: spine movement
simply assessed by the movement or non-movement of spines when approached for capture.
Abnormal spine movement was considered slow spine movement or no spine movement. 3)
Spine loss: spine loss was assessed by observing the surrounding area the urchin inhabited for
whole spines and a visual assessment of the test. Spines that were dropped from the test and
spines that were broken, were distinguishable by the type of lesion left on the test. If spines are
dropping from the test, skeleton is exposed at the band and socket joint where the spine is

42
attached to the test. 4) Spine breakage: spine breakage does not leave visual lesions to the test,
but can be seen by shortened spines on the urchin and indicated by broken spines in the
surrounding area. Urchins with 5% or more spine breakage were considered abnormal, and not
selected for the trials. 5) Test lesions: a healthy urchin test should exhibit no signs of tissue
exposure. If test was exposed, urchins were not selected for trials.
The criteria outlined here, were used in visual assessments of the urchins while in coral
plots at the CRF nursery. The number of urchins exhibiting abnormal signs within each plot was
counted during the experiment and analyzed for differences between collection site, and the end
of the experiment, as well as differences amongst treatments.
A one-way analysis of variance was used to compare the average of urchins present in
cages across treatments and a Tukey HSD test was used to determine differences amongst
treatments in R version 3.2.2 ‘stats’ package (2016). We used descriptive statistics (averages and
standard deviations) to demonstrate trends in the average number of urchins for treatments at
each monitoring point to evaluate trends in urchin abundance in plots and within coral overtime.
For each treatment, the average number of urchins present within the plot, within coral, and
under blocks was plotted on a graph where the relationship between time and urchin abundance
could be evaluated for trends.
Frequencies of abnormal health parameters were recorded for each plot by counting the
number of urchins exhibiting abnormal signs. The health of urchins did not change over time,
with the exception of one event of spine breakage for one urchin on Day 2. Variance in spine
loss across treatments for Day 2 was evaluated using one-way analysis of variance in R version
3.2.2 ‘stats’ package (2016) for spine breakage against treatment.

43
Results
The average abundance of urchins across treatments was significantly different (p<0.01).
further analysis with a Tukey HSD test, cages with 0 and 10 corals were not significantly
different from one another (p=0.95), and cages >25 corals were significantly different from cages
with 0 corals (p<0.05). Over time, there was a decreasing abundance of urchins within cages <25
corals per cage (Figure 3-3). In cages with >25 corals, there was an inverse relationship between
urchins in corals and urchins in clusters- as more urchins sheltered under blocks, less were found
in corals, and vice versa. Within increasing coral cover above 25 corals per structure, urchin
numbers remained relatively stable. Overall, counts of urchins found in each cluster per
treatment showed that after Day 7, there was a decrease in urchin abundance in cages with less
than 25 corals. Plots with >25 corals showed stability in urchin abundance throughout the
experiment where 75% of urchins added to plots were retained in treatments (Figure 3-3).
An average of 2.7 1.7 urchins persisted over time within plots of 10 corals. Plotted
averages of urchins within 10 coral plots decreased over the 20 days of the experiment, notably
on day 7 where averages dropped from 4 urchins to 2 urchins and ended with an average of 1
urchin per plot (Figure 3-5).
Treatments with 25 corals showed a variable response in urchins in corals and urchins
under blocks compared to control and 10 coral treatments (Figure 3-6). Overall, an average 1.6
1.5 urchins were present within coral structures, while 2.2 1.5 urchins were found under
blocks, while the average urchins present within the plot throughout the experiments was 3.5
0.5 (Table 3-1). Urchins present within corals and under blocks with 25 corals per unit appeared
to have an inverse relationship (Figure 3-6). As urchin abundances rose in corals, urchin
abundances decreased under blocks, and vice versa.

44
In treatments with 40 corals, an average of 2.4 1.3 urchins were found in coral clusters,
whereas an average of 1.51.25 urchins were found under blocks (Table 3-1). These plots
showed a stable trend in overall urchin presence over time. For plots with 40 corals, urchin
abundances were inversely related, with a trend of greater urchin presence within coral plots
(Figure 3-7).
Treatments with 50 corals per plot had an average of 2.9 1.5 urchins present in corals
versus an average of 0.9 1.3 urchins found under blocks (Table 3-1). Similar to the 40-coral
treatment, 50 coral plots showed a stable trend in urchin abundance over the duration of the
experiment (Figure 3-8). Recruitment occurred in two of the examined plots (one 40 coral and
one 55 coral treatment), with 5 urchins present on Day 17.
Urchin health parameters were recorded on each monitoring trip. Urchins did not exhibit
signs of abnormalities or poor health throughout the experiment. There were no significant signs
of ill health for urchins relocated to plots within the nursery or across coral density treatments.
Urchins were either present or absent from plots and there were no signs of predation or poor
health (broken spines, test lesions, or loss of spines) with one exception where one urchin
showed signs of broken spines. All urchins exhibited normal spine movement and normal spine
position.
Discussion
This study demonstrated a potential threshold for A. cervicornis coral density of 25
corals/0.25m2 needed for small D. antillarum to have retention rates greater than 75% of
translocated urchins after 20 days. Urchin abundance was significantly different across
treatments, where cages with <10 corals had smaller urchin abundances overtime and cages >25
corals had greater urchin abundances. This provides an important piece of information for

45
managers involved in coral restoration activities and D. antillarum translocation with the purpose
of restoring reef biodiversity and habitat complexity.
Cages were used in this experiment to keep urchins from wandering away and specifically
address the relationship of coral density and the shelter it provided urchins as protection from
predation. Caging artifacts are a concern in any caging experiment, where cage and mesh size
can affect water flow, sedimentation, light attenuation amongst many other parameters (Stocker,
1986; Hall et al. 1990; Como et al., 2006; Miller and Gaylord, 2007). Cages can also increase the
abundance of other biofauna that may be attracted to increased structure at the experiment site
and lead to unexpected artifacts of predation on the organism of study (Stocker, 1986). Because
the aim of this study was to understand which A. cervicornis coral densities could promote
urchin retention over time, cages were left open at the top to keep urchins within the cage, but
still allow predator access to the urchins. It is possible that these cages attracted predator
abundance to the area as it is well established that reef fishes and other invertebrates are attracted
to increased artificial or natural complex structures in the marine environment. Since one of the
goals was to understand how corals provided protection from predation, this was not a concern
for results.
D. antillarum exhibit photosensitive behaviors, showing preference for habitats with lower
light levels (Millot, 1953, 1954; Woodley, 1982). Cement blocks at the bottom of cages were
somewhat elevated from sand where urchins retreated under blocks when added to plots. Over
time, urchins inhabited corals or remained underneath blocks. As strong winds brought greater
wave action and turbidity, sand began to build up under blocks. Plots with less than 25 corals
experienced a loss of urchins, some plots ending the experiment with no urchins present. It
appears as habitat became unavailable under the blocks, coral cover became a major contributing

46
factor for the ability of the urchin to stay within the plot. Multiple factors contributing to a
declining trend in D. antillarum abundances within cages may have occurred including predation
from surrounding fish populations or urchins leaving to seek better shelter (Carpenter, 1984).
With unsuitable coral cover, urchin abundances could not be sustained. This is important for
relocation site considerations. Sites with greater reef rugosity and more coral cover may increase
the urchin retention rate. If reef rugosity is insufficient, greater coral densities may be required to
maintain urchin retention rates.
There was a general trend that as coral density increased, urchin retention also increased,
indicating a positive linear correlation between coral density and urchin retention rates.
However, there was an apparent threshold of coral density that promoted stable urchin retention
rates >75% where coral clusters with >25 corals/0.25m2 retained 75% or more of urchins within
the plot, and clusters with <10 corals/0.25m2, were similar in retention rates (<25%). To further
understand if this is a true linear relationship, future studies should increase coral density
replicates and the number of urchins relocated to coral clusters. These findings of a threshold
however, do indicate there is a “tipping point” where urchin retention is promoted therefore, it is
recommended that urchins are only relocated to clusters with enough structural complexity that
will promote high retention rates, which is possibly coral densities >25 corals/0.25m2.
While cautiously interpreted, two events of urchin recruitment occurred during the
experiment, showing that with greater coral density, there is potential for recruitment of urchins
to coral clusters. On day 17, one plot each with 40 and 55 corals, had five urchins present (one
more than initially added). It is unclear if these urchins were from another cage or from a wild
source. Because of the great distance to a wild population, it is speculated that the increase in
urchins was from urchins migrating from one cage to another. Unfortunately, on Day 20, these

47
cages had been dismantled by strong wind and waves, and urchin abundances could not be
recorded a second time. While this only occurred twice during the experiment, it may be
important for future studies. As coral cover increases, there may be the potential to promote and
foster aggregates of urchins within clusters that can chemically attract other urchins. Aggregates
of urchins may have greater impact on surrounding macroalgae cover (Hernandez et al., 2007)
and contribute to natural re-population of urchins to reseed downstream reefs (Bauer, 1976;
Karlson and Levitan, 1989). This idea should be developed and further explored.
Throughout the experiment, there were no signs of poor health. All urchins retained normal
spine position and movement and did not show signs of spine loss, spine breakage, or test
lesions, indicating current practices for urchin relocation were not detrimental to the health of the
urchin where ample coral cover was provided. Therefore, relocation is not expected to have an
impact on relocated populations through this management strategy if enough coral cover is
provided.
Because of limitations to the study, it is important to address some of the findings with
further experiments. Greater replication and investigation is warranted. Thresholds found in this
study should be identified on reef sites and preliminary D. antillarum relocations should be
conducted and monitored to understand translation of nursery retention to reef site retention
rates. Urchin densities have been shown to affect the abundance of urchins within a cluster or
artificial habitat (Sharp and Delgado, pers. comm.). To understand retention rates when above
four urchins per plot, studies should be repeated with various coral and urchin densities. This
type of information could be used in management plans to enhance success of coral-urchin
restoration projects.

48
The implications of these findings are important to consider when combining coral and
urchin restoration projects into reef recovery programs. They corroborate previous studies that
suggest increased structural complexity can increase D. antillarum abundances, and lay the
ground work for additional urchin relocation experiments on reef habitats. With coral cover of 25
corals/0.25m2, urchin retention can be stably maintained and possibly >75%. Providing habitat to
maintain D. antillarum populations could enhance both coral and urchin restoration success,
providing opportunity for reversal of the currently dominated macroalgae ecosystem. Stocking
restoration sites with multiple juvenile individuals could provide D. antillarum with the
necessary habitat to outgrow predation vulnerability and form breeding populations to increase
recruitment of urchins to the reef site.

49
Figures
Figure 3-1: Layout of coral density experimental grid. Cages were constructed of chicken wire
with an area of 0.25m2 and installed in a sandy bottom area on the edge of the CRF
coral nursery. Cages were 2m apart from each other. Coral density treatments were
randomly assigned to each cage for densities of 0, 10, 25, 40, and 55 corals shown
here.
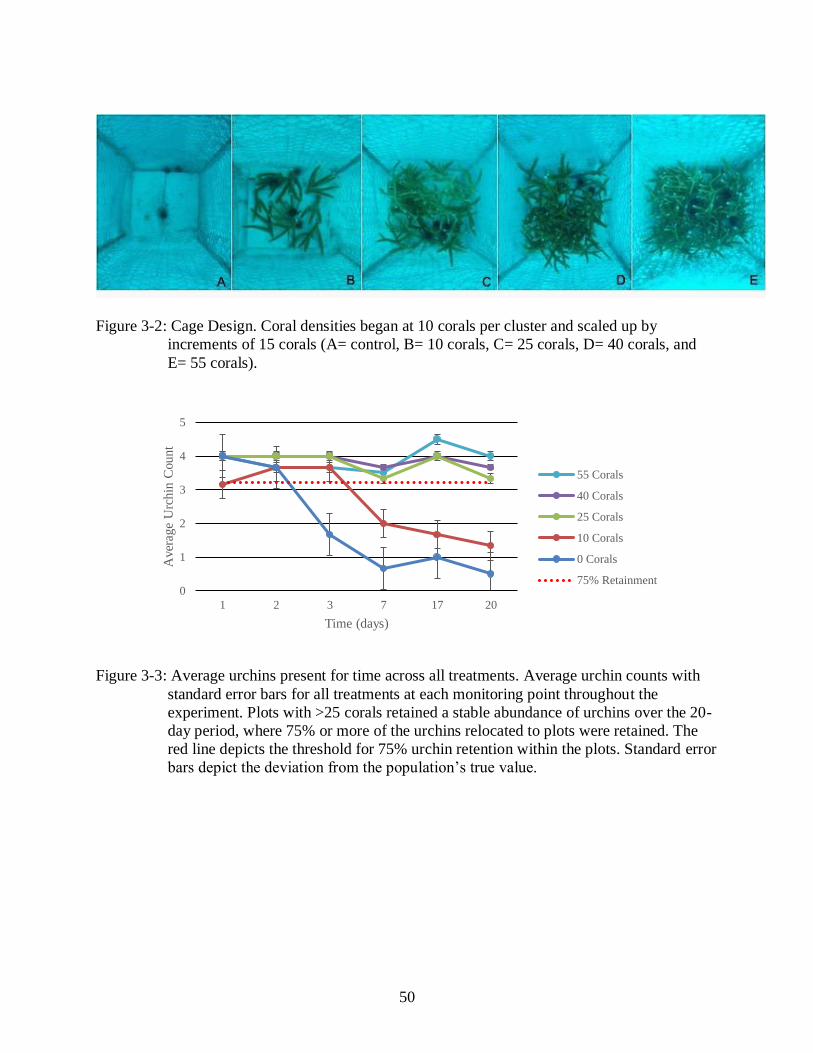
50
Figure 3-2: Cage Design. Coral densities began at 10 corals per cluster and scaled up by
increments of 15 corals (A= control, B= 10 corals, C= 25 corals, D= 40 corals, and
E= 55 corals).
Figure 3-3: Average urchins present for time across all treatments. Average urchin counts with
standard error bars for all treatments at each monitoring point throughout the
experiment. Plots with >25 corals retained a stable abundance of urchins over the 20-
day period, where 75% or more of the urchins relocated to plots were retained. The
red line depicts the threshold for 75% urchin retention within the plots. Standard error
bars depict the deviation from the population’s true value.
0
1
2
3
4
5
1 2 3 7 17 20
Aver
age
Urc
hin
Count
Time (days)
55 Corals
40 Corals
25 Corals
10 Corals
0 Corals
75% Retainment

51
Figure 3-4: Urchins present over time for control cages. Average counts with standard error bars
for urchins present at each monitoring point during the experiment for control plots
across all potential locations within the plot, which includes “in coral” and “under
blocks”. The grey line represents overall average of total urchins present within the
plot. In control plots, overall urchin retention dropped quickly over a short-term
period.
Figure 3-5: Urchins present over time for 10 corals. Average counts with standard error bars for
urchins present at each monitoring point during the experiment for plots with 10
corals across all potential locations within the plot, which includes “in coral” and
“under blocks”. The grey line represents overall average of total urchins present
within the plot. In 10 coral plots, overall urchin retention dropped quickly over a
short-term period.
0
1
2
3
4
5
1 2 3 7 17 20
Aver
age
Urc
hin
Count
Time (Days)
Urchins in Coral
Urchins Under Blocks
Total Urchins
0
1
2
3
4
5
1 2 3 7 17 20
Aver
agde
Urc
hin
Count
Time (Days)
Urchins in Coral
Urchins Under Block
Total Urchins

52
Figure 3-6: Urchins present over time for 25 corals. Average counts with standard error bars for
urchins present at each monitoring point during the experiment for plots with 25
corals across all potential locations within the plot, which includes “in coral” and
“under blocks”. The grey line represents overall average of total urchins present
within the plot. In 25 coral plots, stable urchin retention was maintained over the 20-
day period.
Figure 3-7: Urchins present over time for 40 corals. Average counts with standard error bars for
urchins present at each monitoring point during the experiment for plots with 40
corals across all potential locations within the plot, which includes “in coral” and
“under blocks”. The grey line represents overall average of total urchins present
within the plot. In 40 coral plots, stable urchin retention was maintained over the 20-
day period.
0
1
2
3
4
5
1 2 3 7 17 20
Aver
gae
Urc
hin
Count
Time (days)
Urchins in Coral
Urchins Under Blocks
Total Urchins
0
1
2
3
4
5
1 2 3 7 17 20
Aver
age
Urc
hin
Count
Time (days)
Urchins in Coral
Urchins Under Blocks
Total Urchins

53
Figure 3-8: Urchins present over time for 55 corals. Average counts with standard error bars for
urchins present at each monitoring point during the experiment for plots with 55
corals across all potential locations within the plot, which includes “in coral” and
“under blocks”. The grey line represents overall average of total urchins present
within the plot. In 55 coral plots, stable urchin retention was maintained over the 20-
day period.
Table 3-1: Averages of urchins present within cages during experiment.
Treatment Urchins in Coral (avg std dev)
Urchins Under Blocks (avg std dev)
Urchins Present (avg std dev)
0 corals 0 0 2.2 1.7 2.3 1.7
10 corals 0 0.8 2.4 1.7 2.7 1.7
25 corals 1.6 1.5 2.2 1.5 3.8 0.5
40 corals 2.4 1.3 1.5 1.3 3.9 0.5
55 corals 2.9 1.5 0.9 1.3 3.9 0.5
Table of the average ( standard deviations) abundance of urchins present within experimental
plots for the number of urchins “in coral”, “under blocks”, and the total urchins
present with standard deviation values.
0
1
2
3
4
5
1 2 3 7 17 20
Aver
age
Urc
hin
Count
Time (days)
Urchins in Coral
Urchins Under Blocks
Total Urchins

54
CHAPTER 4
PREDICTING SUCCESSFUL DIADEMA ANITLLARUM RELOCATION SITES IN THE
FLORIDA KEYS
Introduction
When done on a large scale, coral restoration efforts can create a positive trajectory for
threatened Acroporid populations (Miller et al., 2016). Despite positive trajectories, corals
outplanted to reefs are still subject to various local and global stressors including climate change,
rising sea surface temperatures, and the persistence of macroalgae which makes habitat
unsuitable for coral recruitment and recovery (Jackson et al., 2001; Knowlton, 2001; Bellwood et
al., 2004; Brown et al., 2014; Dixson et al., 2014). The National Oceanic and Atmospheric
Administration (NOAA) Acropora Recovery Plan (2015), notes that for coral restoration efforts
to be successful, populations must be present across approximately 5% of consolidated reef
habitat and sustained for 20 or more years. Abating local stressors, such as poor water quality,
eutrophication, and decreasing surrounding macroalgae may help recovering coral populations
become resilient in the face of global climatic pressures (Mumby et al., 2007; Anthony et al.,
2015) and decrease requirements for continued restoration.
Diadema antillarum are critical to the removal of macroalgae on Caribbean coral reefs. In the
absence of D. antillarum, as has been the case since their epidemic mortality in 1983, coral reefs
have become dominated by fast-growing macroalgae which outcompetes and smothers the
scleractinian corals (Lirman, 2001; McCook et al, 2001). Recovery of D. antillarum has the
potential to reverse reef states, currently existing as macroalgae dominated, back to coral
dominated states (Edmunds and Carpenter, 2001). Many studies have demonstrated the
significant positive impacts reintroduction of D. antillarum can have to surrounding macroalgae
(Nedimyer & Moe, 2003; Burdick, 2008; Dame, 2008). Natural recovery of D. antillarum on
reefs in Jamaica have corresponded with a remarkable increase in scleractinian coral cover

55
(Edmunds and Carpenter, 2001; Carpenter and Edmunds, 2006; Idjadi et al., 2010). D.
antillarum are in need of enhanced coral cover for protection from predators (Levitan and
Genovese, 1989), as such, the recovery of coral may promote the recovery urchins, and the
recovery of urchins may in turn promote the recovery of corals.
The objectives of this study were to predict and identify current restoration efforts that
may facilitate the recovery of urchins by selecting suitable translocation reefs with appropriate A.
cervicornis coral coverage, and create a tool for managers charged with implementing coral
restoration programs in conjunction with D. antillarum recovery programs, so that preliminary
coral-urchin restoration trials may be tested using the information presented in this study.
Methods
Datasets used to generate information for this study were obtained from previous coral
density experiments in the CRF nursery (Chapter 3), annual A. cervicornis coral density and
growth (Lirman et al., 2014), and outplanting data with permission from Coral Restoration
Foundation (CRF, unpublished data). Information regarding restoration site name, coordinates,
number of clusters at the restoration site, number of corals outplanted within the cluster, and time
on reef were extracted from the CRF outplanting database. The average size of each coral
fragment outplanted to the reef was ~30cm total linear extension (TLE) (CRF, unpublished data)
with the exception of ECO sites, where average coral size was ~60cm TLE. The average annual
growth rate of the corals was set at 4cm per initial size of the coral according to Lirman et al.
(2014). Using the datasets above, we generated a total “cluster-biomass” for each coral cluster
and predicted from 2017-2020 which sites would be suitable for coral-urchin restoration projects
based off the cluster-biomass of outplanted A. cervicornis to reef sites in the Florida Keys.
A. cervicornis were outplanted to reefs in one of two ways. The first method, which is the
protocol used for the majority of outplant sites, involved clearing the substrate of algae and

56
calcareous algae to the clean limestone rock underneath with a hammer. Corals were attached to
the reef using a two-part marine epoxy at three points of attachment. Once attached, the epoxy
hardened and the coral grew over the epoxy on to the substrate, and eventually fused with other
surrounding A. cervicornis colonies within the cluster. Corals outplanted with this first method
were on average 30cm TLE. The second method, which is still being refined, is an expedited
coral outplanting (ECO) strategy where large corals (avg. 60cm TLE) were taken to patch reefs,
wedged into crevices, and then overlaid on one another to create a coral thicket. The latter
method was first deployed in September 2015, and has been used experimentally to increase the
amount of coral at a reef site in a short amount of time.
Diadema Retention Model. The calculated cluster-biomass of coral clusters used in this
study was defined as the cumulative TLE (cm) of all corals outplanted in an area of 1m2 (cluster-
biomass TLE cm/m2). First, all coral density treatments in Chapter 3, were converted to units of
cluster-biomass using the “initial cluster biomass” equation:
𝐵 = 𝑆𝑖 × 𝑛
𝐴
Where Si is the initial coral size, n is the number of coral fragments within the designated cluster,
and A is the area over which the corals were placed.
The converted cluster-biomass for coral density treatments (0, 1200, 3000, 4800, and
6600cm TLE/m2) and their associated Day 20 urchin retention rates (.25, .25, .75, 1.0, 1.0) were
fed into R Version 3.2.2. using the ‘stats’ Package (2016) to generate a Diadema Retention
Model (DRM) with cluster-biomass as a predictor of urchin retention. The model was analyzed
using regression analysis.
Identifying Suitable Sites. To identify sites where coral-urchin relocations could take
place from 2017-2020, values were generated for cluster-biomass that would support urchin

57
retention >75% for at least 20 days. Outputs from the DRM for desired thresholds were assigned
one of three categories based on the percent of urchin retention each cluster could maintain:
“Good” (75-84.9%), “Better” (85-94.9%), and “Best” (>95%). These values were used to
suggest sites where necessary cluster-biomass currently existed from 2017-2020.
We calculated the yearly cluster-biomass (BY) for each cluster in relation to the amount of
time on the reef for years 2017-2020 using the following equation:
𝐵𝑌 = 𝐵 ×(𝐴𝑃)𝑡 × (𝐴𝑆)𝑡
Where B is the beginning cluster-biomass (calculated above), t is the time the cluster has been on
the reef in years, AP is the annual productivity of the corals (4cm/year; Lirman et al., 2014), and
AS is the annual survivorship of outplanted corals (0.80; CRF, unpublished data).
This calculation assumes the following parameters: (i) corals outplanted to reef sites have
an annual survivorship rate of 0.80, (ii) corals >30cm TLE have an annual productivity rate of
4cm annually per initial size of the coral, (iii) coral clusters are outplanted over an area ~1m2,
and (iv) coral growth is exponential.
Coordinates of clusters identified as good, better, and best for urchin relocations were
imported into ESRI ArcGIS. Sites where specific ranges of corals categorized as good, better, or
best were plotted using yellow, blue, and green dots respectively for years 2017-2020.
Results
The Diadema Retention Model (DRM) was significantly correlated between cluster-biomass
and urchin retention rates (p=0.01604, F= 25.25, R2=0.8532) and is linear between 0-6000cm
TLE/m2 units of cluster-biomass where urchin retention is between 0-100% (Figure 4-1). Input
for desired urchin retention rates for “Good” (75-84.9%), “Better” (85-94.9%), and “Best”
(>95%) were specified as requiring 3782.6 - 4438.6cm TLE/m2, 4445.2 - 5101.2cm TLE/m2, and
>5107.8cm TLE/m2 respectively, to retain urchins for at least 20 days within A. cervicornis coral

58
clusters outplanted to restoration sites. A table of restoration sites with the corresponding number
of clusters was generated as a reference for optimal coral densities for urchin relocation trials
2017-2020 to be used by managers for easy identification of reef sites suitable for coral-urchin
restoration (Appendix A).
Over 3,000 A. cervicornis coral clusters have been outplanted across 40 restoration sites
along the Florida Keys Reef Tract since 2014 and were incorporated into this model. In 2017,
very few clusters suited necessary parameters needed for successful urchin retention. Currently
in 2017, 6.77%, 4.21%, and 15.5% of A. cervicornis clusters are considered good, better, and
best respectively for urchin relocations (Figure 4-5). Based on growth rates in 2018, 3.58%,
4.73%, and 61.68% of coral clusters will be considered, good, better, or best for urchin retention
respectively (Table 4-5). In 2019, 0.22%, 1.40%, and 98.27% will be considered good, better,
and best (Table 4-5). In 2020, 100% of coral clusters used in this study are expected to be
suitable for urchin relocation efforts (Table 4-5). Restoration sites clustered around areas of the
Upper Keys and spread out overtime as coral-biomass increased along the Florida reef tract
(Figure 4-2 to 4-5).
Discussion
The DRM predicted which reefs sites are suitable for urchin relocation now and will
become so over the next three years. It serves as a baseline for predicting the amount of A.
cervicornis cluster-biomass needed to support urchin retention at restoration sites and may offer
explanations for past D. antillarum relocation efforts that were unsuccessful as a result of poor
reef complexity (Nedimyer & Moe, 2003; Burdick, 2008). As coral-urchin restoration is scaled
up, additional data can be incorporated into the DRM for enhanced results to predict restoration
sites that are suitable for urchin relocation efforts.

59
Based on the applied information from the DRM, <30% of A. cervicornis clusters
outplanted to reef sites are currently able to support a target minimum of 75% urchin retention
for 20 days. Relocating urchins at present to A. cervicornis clusters may result in mediocre
retention rates, but provide opportunity to begin preliminary relocation trials to test the results of
these studies further. Corals outplanted in clusters of 10 with an average size of 30cm TLE
appear to reach cluster-biomass necessary to promote >95% urchin retention around 3 years of
growth. In 2018, the total proportion of clusters suitable for potential urchin relocation trials
increases to 69.99%, then 99.89% and finally 100% in years 2019 and 2020, where all corals
have been on the reef for 3 or more years.
Scaling up current coral restoration programs in the Florida Keys and Caribbean is of
great emphasis for many reef managers (NOAA Workshop, Nov. 2016). Increasing the
abundance of coral on degraded reef sites and promoting their long-term recovery is a priority
outlined in the NOAA Acropora Recovery Plan (2015) for recovery of the species.
Unprecedented amounts of corals have been outplanted across the Florida Keys since 2014, but
may take a few years to reach optimal sizes for urchin relocations. Recently, an effort to scale-up
restoration projects has taken place using ECO methods. These methods have been deployed at
targeted patch reef restoration sites, where more than 50 large corals can be placed in an area
~1m2. In combination with current methods, this ECO strategy produced 7 restoration sites in
less than one year that are suitable to promote >75% urchin retention if relocated to clusters at
these restoration sites. As more reef area becomes available overtime, it is timely to begin
consideration of alternative strategies to increase D. antillarum production and collections. Ex
situ D. antillarum rearing may increase numbers of urchins available for relocation trials and
speed recovery of the species (Chandler et al., in press).

60
Although promising, the DRM has limitations in design and available data that warrant
further investigation. The DRM was targeted towards urchins relocated to a caged coral cluster.
It is assumed that with a greater number of suitable coral clusters, you may add more urchins to a
restoration site. However, it is important to understand the relationship between urchin retention
and coral cover over a greater area of the reef. Carrying capacity of urchins on reefs is currently
an unknown factor and one recently identified as a potential critical factor in future management
decisions (Diadema Workshop, Feb. 2017).
Data used to create the DRM was based on caged experiments in a semi-controlled
environment. It is imperative to replicate experiments on restoration sites and compare against
nursery experiments to better understand the relationship of D. antillarum retention more
practically. The sites suggested for preliminary urchin relocations can serve as a viable starting
point for these experiments.
It is known that with increasing reef complexity, D. antillarum densities can increase
(Clemente and Hernandez, 2008; Dame 2008; Rogers and Lorenzen, 2016). As A. cervicornis
clusters become larger and more complex suitable area for urchin relocations can increase. The
DRM currently predicts the sites where cluster-biomass will become suitable over the period of a
few years using an A. cervicornis annual productivity rate (Lirman et al., 2014) and average A.
cervicornis survivorship across multiple reef sites in the Florida Keys (CRF, unpublished data).
The equations used to predict this, assume exponential growth, which is impractical for a biotic
organism that can experience multiple stressors that can cause mortality. Multiple variables
including differences in reef habitat (e.g. Dizon and Yap, 2005; Mieog et al., 2009), coral
genotype (e.g. Baums, 2008; Williams et al., 2014), and bleaching or disease events (e.g.
Aronson and Precht, 2001; Eakin et al., 2010) have the ability to alter survivorship of coral

61
clusters. If known, specific survivorship percentages for each reef site or region should be
incorporated in to the DRM for greater predictive accuracy of cluster-biomass over sequential
years. Consequently, it is important to consider in recovery plans which sites may have seen
greater mortalities of restored corals in the sequential years that are presented in tables for reef
site identification so that urchins are not relocated to sites where mass coral mortalities have
taken place.
Finally, the DRM only used A. cervicornis to model cluster-biomass and urchin retention. A.
cervicornis can rapidly form densely branching thickets that can provide juvenile D. antillarum
with natural habitat as they grow into adults and work to re-establish local populations. While
other coral species and growth forms, such as Orbicella spp., A. palmata, and Millepora
complanata, may provide more suitable habitat for larger urchins (Weil et al., 1984), past
declines in coral cover coupled with their slower growth rates may not provide viable options for
timely restoration of corals and urchins. As A. palmata or other coral species become more
abundant in coral restoration programs, perhaps they too can begin to provide increased habitat
complexity for urchins.
A holistic approach to reef restoration combining coral and urchin recovery may lead to
enhanced results, where corals provide greater habitat complexity for urchins, and urchins reduce
the amount of macroalgae in the area increasing coral survivorship by reducing competition.
Some sites currently exist to begin preliminary D. antillarum relocation trials, however, to
support greater urchin retention, efforts to scale up restoration and configure present coral
outplantings so that optimum cluster-biomass is achieved in a shorter amount of time should be
the next steps taken to increase efficacy of coral-urchin restoration programs for enhanced reef
recovery.

62
Figures
Figure 4-1: “Diadema Retention Model” for A. cervicornis restoration sites. The relationship of
cluster-biomass and urchin retention is represented by the black line and the 95%
confidence intervals are represented by the red-dotted lines. The blue dashed line
represents the point at which 75% urchin retention is achieved for cluster-biomass
(3782.609cm TLE/m2).

63
Figure 4-2: A. cervicornis restoration sites identified in the Florida Keys for preliminary D.
antillarum relocations in 2017.
Figure 4-3: A. cervicornis restoration sites identified in the Florida Keys for preliminary D.
antillarum relocations in 2018.

64
Figure 4-4: A. cervicornis restoration sites identified in the Florida Keys for preliminary D.
antillarum relocations in 2019.
Figure 4-5: A. cervicornis restoration sites identified in the Florida Keys for preliminary D.
antillarum relocations in 2020.

65
Table 4-1: Proportion of Clusters Present for Each Category for Years 2017-2020.
Year Good
(75-84.9%)
Better
(85-94.5%)
Best
(>95%)
Total
2017 246 (6.77) 153 (4.21) 565 (15.5) 964
(26.48)
2018 130 (3.58) 172 (4.73) 2,241 (61.68) 2,543
(69.99)
2019 8 (0.22) 51(1.40) 3,573 (98.27) 3,632
(98.89)
2020 0 (0) 0 (0) 3,636 (3636) 3,636
(100)
Table of total clusters present for each year in and the proportion they represent of total clusters
on the reef assigned for good, better, and best D. antillarum retention rates.
Proportions are recorded in parentheses.

66
CHAPTER 5
DISCUSSION
In the 1980s Diadema antillarum were decimated across the Florida Keys and wider
Caribbean reefs. This has been linked to macroalgae overgrowth and suffocation of scleractinian
corals resulting in loss of reef biodiversity (Bak et al., 1984; Lessios et al., 1984; Hughes et al.,
1987; Carpenter, 1990) creating a bleak future for the ecologically vital Florida Keys Reef Tract.
It has been postulated that restoration of both D. antillarum urchin species and Acropora
cervicornis coral species can create a symbiotic partnership between the two and assist with reef
recovery back to a coral dominated state. This study examined various strategies to assist this by
assessing six primary objectives using two experiments to generate a model predicting where
successful urchin relocations might take place to assist enhanced reef recovery. To achieve this,
experiments were conducted to assess the following general hypotheses and objectives for each:
Hypothesis 1: Relocation of D. antillarum to coral restoration sites does not affect health
of the urchins and its objectives (a) determine the parameters necessary to promote D. antillarum
health in an ex situ environment; (b) understand potential habitat usage and behavior of D.
antillarum and T. ventricosus in a controlled environment when compared to expected normal
behavior; (c) understand proper health metrics for monitoring D. antillarum health for in situ
relocations and studies.
Hypothesis 2: With increasing A. cervicornis density, D. antillarum retention within coral
clusters increases and its objective (a) determine the coverage of A. cervicornis coral needed to
promote urchin retention over time when urchins were relocated to coral clusters.
Hypothesis 3: With multiple restoration efforts taking place, suitable habitat currently
exists to support D. antillarum relocation and its objectives (a) predict and identify current
restoration efforts that may facilitate the recovery of urchins by selecting suitable relocation to

67
reefs with appropriate coral infrastructure (coverage); (b) create a tool for managers charged with
implementing coral restoration programs in conjunction with D. antillarum recovery programs.
Diadema antillarum Health and Behavior Parameters Ex Situ: Applications for In Situ
Monitoring and Relocation Efforts
Current ex situ rearing efforts being conducted privately (Mr. Martin Moe, Islamorada,
FL) and in the aquarium industry (Mr. John Than, The Florida Aquarium, Tampa, FL) may soon
achieve standard protocols for mass production of these urchins allowing for a surplus of urchins
to be used for relocation efforts. On-land aquaculture provides many potential benefits for the
recovery of sea urchins such as mass production, control to produce uniform age class and size,
and potential annual restoration efforts versus reliance on seasonal factors to induce spawning
and settling in the wild.
Classification as an invertebrate renders certain ethical husbandry parameters required for
marine vertebrates, unnecessary to consider in design of experiments. D. antillarum is not
currently listed as a protected species under the U.S. Endangered Species Act and has not been
considered for listing on the IUCN Red List (IUCN 2016). In contradiction to the listing status of
D. antillarum, it is widely recognized that the species has been extensively affected from the
1980’s mass mortality event, where 93% of the Caribbean population was decimated (Lessios,
1988). Monitoring the health of urchins used in experiments can be beneficial for understanding
if current practices are detrimental to the health of the urchins, as well as contribute reasons for
good or poor urchin survivorship when relocated to coral restoration sites. Health and behavior is
also a concern for current efforts of D. antillarum propagation ex situ for translocation in situ.
Our first hypothesis- relocation of D. antillarum to coral restoration sites does not affect
health of the urchins- was addressed through the objectives: (1a) determine the parameters
necessary to promote D. antillarum health in an ex situ environment; (1b) understand potential

68
habitat usage and behavior of D. antillarum and T. ventricosus in a controlled environment when
compared to expected normal behavior; and (1c) understand proper health metrics for monitoring
D. antillarum health for in situ relocations and studies. These objectives specifically addressed
concerns with D. antillarum health in regards to moving them from the wild to another reef
habitat, as well as better understanding for behavior of ex situ urchins and parameters that could
potentially lead to poor health of urchins when relocated to reef sites.
Objective 1a: determine the parameters necessary to promote D. antillarum health in an ex
situ environment.
A protocol for parameters to maintain urchin health ex situ was not fully attained for this
objective. Urchins were adversely affected during trials in this experiment. By assessing health
of the urchins in ex situ trials, we highlighted the importance of controlling external abiotic
factors that may possibly cause stress in the urchins and decrease the overall accuracy of any
trials being conducted. Facilities conducting ex situ urchin breeding programs to increase
individuals for use in coral-urchin restoration efforts, should be fully equipped to provide
necessary temperature adjustments, supply of food for routine feeding regimes, and structure that
mimics in situ habitat for urchins to inhabit.
Objective 1b: understand potential habitat usage and behavior of D. antillarum and T.
ventricosus in a controlled environment when compared to expected normal behavior.
Concern also arises when considering altered behavior of ex situ raised urchins for urchin
relocation efforts. It has been observed that urchins raised in ex situ environments do not display
the same shelter-seeking or predator avoidance behaviors when compared to in situ collected D.
antillarum (Sharp and Delgado, pers. comm.). Urchins in ex situ trials showed abnormal shelter-
seeking behaviors before a decline in their health. It is uncertain whether these behaviors were
due to poor conditions or a result of the urchins being held ex situ, however it is clear their
behaviors were not similar to wild D. antillarum used for in situ experiments. In situ relocated

69
urchins immediately sought shelter under shaded habitat or within coral clusters. These urchins
appeared to possess innate knowledge to seek shelter and avoid predation. Selecting for natural
behaviors such as these or training ex situ raised urchins may increase the likelihood of urchin
relocation success where urchins seek shelter immediately, decreasing the likelihood of predation
and increasing their chances for survival.
Ex situ trials may provide necessary insight to urchin biology as we strive to fill in the
numerous gaps in our knowledge, but they need to closely approximate the target environment
where urchins will be relocated- the relatively climatically stable staghorn dominated restoration
sites of the Florida Keys. Further, future mass production through ex situ aquaculture of D.
antillarum as a restoration strategy needs to consider imprinting shelter seeking and predator
avoidance behavior if these behaviors are to be maintained throughout land-based holding prior
to translocation of D. antillarum to an environment that requires foraging and predator
avoidance.
Objective 1c: understand proper health metrics for monitoring D. antillarum health for in
situ relocations and studies.
Criteria used to evaluate the health of urchins in ex situ trials were outlined from a health
evaluation protocol developed by Francis-Floyd et al. (in press) as an evaluation tool for the
release of ex situ raised urchins in relocation efforts. These protocols not only proved to be
sufficient for evaluating the health of urchins ex situ, but also in situ studies for evaluating
stability in urchin health and effects of relocation practices. In contrast to the ex situ trials at
CFC, urchin health during in situ trials showed no significant declines in health throughout the
experiment where urchins exhibited the same healthy characteristics from collection to the end of
the experiment on Day 20. This suggests current relocation protocols from the collection,

70
transportation, and release onto coral clusters are not detrimental to the health of urchins, nor do
they promote predation when ample shelter is provided.
Understanding this provides practitioners with criteria to better interpret results of
relocation efforts. If efforts were unsuccessful and proper relocation protocols were followed,
failure may have resulted in areas outside of urchin health and relocation parameters, factors
including reef complexity, urchin homing behavior, or other unknown variables. Knowing this
can allow researchers to isolate other possibilities of failure, and explore further issues more in
depth. Experimental
Acropora cervicornis Densities to Promote Diadema antillarum Retention
Our second hypothesis- with increasing A. cervicornis density D. antillarum retention within
clusters increases- was addressed with the objective to (2a) determine the coverage of A.
cervicornis coral needed to promote urchin retention over time when urchins were relocated to
coral clusters. Current coral restoration efforts focus on propagation and restoration of A.
cervicornis fragments for outplanting at designated restoration sites (Young et al., 2012). While
not historically associated as D. antillarum habitat, A. cervicornis coral thickets as a result of
coral restoration efforts may provide an interim habitat for juvenile D. antillarum to grow
outside of predation prone test diameters and head start adult populations at restoration sites with
potential to reseed downstream reefs.
Objective 2a: determine the coverage of A. cervicornis coral needed to promote urchin
retention over time when urchins were relocated to coral clusters.
In situ trials identified units for cluster-biomass that may promote 75% or more retention of
juvenile D. antillarum when relocated to A. cervicornis clusters where appropriate coral
coverage was present. This study demonstrated a potential threshold for A. cervicornis coral
cluster density of 25 corals/0.25m2 or more needed for juvenile D. antillarum to have a retention

71
rate >75% of translocated urchins after 20 days. The implications of these findings are important
for managerial decisions, corroborate previous studies, and lay the ground work for additional
urchin relocation experiments on reef habitats. Providing habitat to maintain D. antillarum
populations could enhance both coral and urchin restoration success. Stocking restoration sites
with proper coral cover could provide D. antillarum with the necessary habitat to outgrow
predation vulnerability and form breeding populations to increase recruitment of urchins to the
reef site. In return, increased D. antillarum densities at the restoration site can decrease
overgrowth of macroalgae (Sammarco, 1982; Burdick, 2008; Hernandez et al., 2008) and
increase growth of the surrounding coral (Edmunds and Carpenter, 2001; Idjadi et al., 2010)
providing opportunity for reversal of the currently macroalgae dominated ecosystem.
Applications for Management
Our third hypothesis- With multiple restoration efforts taking place, suitable habitat
currently exists to support D. antillarum relocation- was supported with the objectives to: (3a)
create a tool for managers charged with implementing coral restoration programs in conjunction
with D. antillarum recovery programs, and (3b) predict and identify current restoration efforts
that may facilitate the recovery of urchins by selecting suitable translocation reefs with
appropriate coral infrastructure (coverage) Through these objectives, we were able to prove there
are sites that currently exist to begin coral-urchin restoration efforts and greater abundances of
sites that will become available over the next three years as A. cervicornis clusters continue to
grow.
Objective 3a: predict and identify current restoration efforts that may facilitate the
recovery of urchins by selecting suitable translocation reefs with appropriate coral
infrastructure (coverage).
Acropora cervicornis has been the focus of many restoration groups, however, a diverse
array of other coral species can greatly increase structural complexity on coral reefs (Alvarez-

72
Filip et al., 2011). Current efforts to propagate A. palmata, bouldering corals, and pillar corals
are underway, yet still require ample amount of time and replication to maximize success of
these programs (NOAA Workshop, 2016). In the meantime, the proven success of A. cervicornis
restoration efforts can provide an interim habitat for juvenile D. antillarum to be moved to
restoration sites either from in situ collection sites, or ex situ hatcheries.
A Diadema Retention Model (DRM) was created to predict urchin retention at restoration
sites when relocated to coral clusters that have properties of suitable coral density and ground
cover. The model allowed us to input desired urchin retention rates and output a specific cluster-
biomass needed to promote retention. Using this information, we identified sites where habitat
currently exists to support preliminary urchin relocations as well as which sites would become
available over the next 3 years as coral growth continues within clusters. As coral-urchin
restoration is scaled up, additional data can be incorporated into the DRM for enhanced
predictive results of a successful coral-urchin restoration site, however this tool currently
provides a baseline of sites where combined restoration efforts may begin.
Currently, less than 30% of coral clusters outplanted to reefs have sufficient A.
cervicornis cover to promote successful urchin retention based on our studies. However, in years
2018, 2019, and 2020, this greatly increases to 69.99, 99.89, and 100% respectively under
current outplanting and site restoration regimes. Clusters achieve optimal cluster-biomass in
about three years, however, unknown limitations such as coral disease, coral bleaching, and other
coral stressors may result in a diminished cluster-biomass over time. Recognizing that these
stressors may contribute to a decrease in number of clusters available, it is important to consider
increasing the abundance of corals in a shorter timeframe to incorporate urchin relocations as
soon as will allow. Coral cluster-biomass could be greatly increased if larger coral size and

73
greater coral thicket densities were outplanted to restoration sites in a shorter period of time.
CRF ECO strategies showed great promise in generating sufficient coral cluster-biomass needed
for successful urchin relocations, close to only one year after outplanting. These efforts to
maximize coral cluster density can be beneficial for not only increasing the thickets of corals
placed onto reefs, but also the amount of structure they provide for urchin relocations. If D.
antillarum retention within a restoration site can be maintained, urchins can restore natural
herbivorous benefits to the reef ecosystem, consuming macroalgae, and providing suitable
habitat for coral colonies to grow and thrive (Sammarco et al., 1982; Edmunds and Carpenter,
2001; Nedimyer and Moe, 2011). In return, the increased healthy coral structure can provide
habitat for relocated urchins to increase local populations, and possibly reseed downstream reefs.
Objective 3b: create a tool for managers charged with implementing coral restoration
programs in conjunction with D. antillarum recovery programs.
Resources for management and recovery of reef species can be limited with funding,
biologically available specimens, and resources for monitoring and maintenance of the targeted
objective in reef recovery. These resources are inextricably valuable and conserving resources
used, to generate a large impact, is of the utmost importance. Diadema antillarum populations
are in slow recovery at the majority of reef sites in the Florida Keys and the Caribbean, but have
been cited as essential for the recovery of coral reefs to a coral-dominated stage (Edmunds and
Carpenter, 2001). Utilizing relocation efforts to promote coral cover should be paired with the
best possible knowledge to maximize resources and promote protection of the urchins from
predators to ensure that their persistence at a reef site is met with the objectives of reducing
macroalgae cover and promoting survival into adult spawning sizes. As coral reefs have been
flattened (Alvarez-Filip et al., 2009), there may be less area for relocated D. antillarum to reside,
and reefs may hold carrying capacities for urchins based off of available complexity of the reef

74
(Diadema Workshop, 2017). Added structure to the reef through A. cervicornis restoration
efforts can increase the structural complexity of a reef short to long-term by adding coral cover
that was not previously existent. As shown above, a certain threshold of coral cluster-biomass
can increase the percentage of urchins retained when relocated to a coral cluster. Identifying sites
that currently exist to promote the greatest maximum urchin retention can provide managers and
practitioners with valuable knowledge to ensure that their efforts are successful and a wise use of
resources.
The DRM is a valuable tool for managers looking to incorporate urchins into their current
restoration programs, and holds important implications for restoration protocols at present. The
results of this study show that the goal of restoration programs should be to increase the amount
of coral on the reef, but also to increase the structural complexity of reefs that are restored.
Increasing complexity of the reefs can increase the number of dynamic interactions needed to
promote not only the recovery of D. antillarum, but also bring back dynamic reef interactions
that can promote overall normal function of reefs pre-1970.
Future Work
To effectively stimulate reversal back to coral dominated reefs, management strategies
are highly encouraged to include holistic species recovery together. Some sites currently exist to
begin preliminary D. antillarum relocation trials, however, to support greater urchin retention,
the potential reef sites identified as coming online in 2018-2020 should be the subject of greater
maintenance and care to ensure they can support D. antillarum relocation at this critical time in
the recovery of the Florida Keys Reef Tract.
In addition to future sites that were identified as potential successful urchin relocation
sites, it is important to consider the scaling up of coral restoration efforts so that greater areas of
habitat may return to structural complexity suitable to promote dynamic interactions of other reef

75
organisms. ECO methods to prompt greater coral cover in shorter amounts of time may be
favorable for increasing habitat necessary to promote D. antillarum urchin recovery and should
be tested in initial urchin relocation trials.
Coral density experiments should be replicated on reefs to evaluate urchin retention at
sites with greater fish abundance and variable substrate complexities. Underlying reef rugosity
should be consideration in urchin relocation sites and one that is investigated further, and
possibly added to the DRM. Sites with greater reef rugosity may promote greater urchin
retention rates where urchins can hide in crevices, and use complexity provided from A.
cervicornis to forage at night, where coral can provide greater protection from local predatory
fish and invertebrates.
These implications are important for successful local relocation and reef recovery efforts,
however, to generate an even larger impact, principles of large-scale reef connectivity should
also be considered. Future work to understand the connectivity of reefs and D. antillarum larval
dispersal may offer strategies for selecting reef sites where coral-urchin restoration efforts are
most beneficial across large-scale regions, ultimately contributing to a naturally functioning reef
habitat which may promote natural resiliency giving reef ecosystems a chance to overcome
effects of hysteresis and other global stressors including climate change, ocean acidification, or
increasing storm intensities. Coral reefs are complex and dynamic ecosystems with complex
factors that can affect their health and recovery. As such, greater attention is needed for the
development of recovery strategies that incorporate an equally complex and dynamic restoration
approach. The combination of programs that incorporate recovery of multiple niche groups, such
as coral restoration and urchin relocations, is one solution that may initiate the return of coral to
degraded reef communities and enhance reef recovery rates.
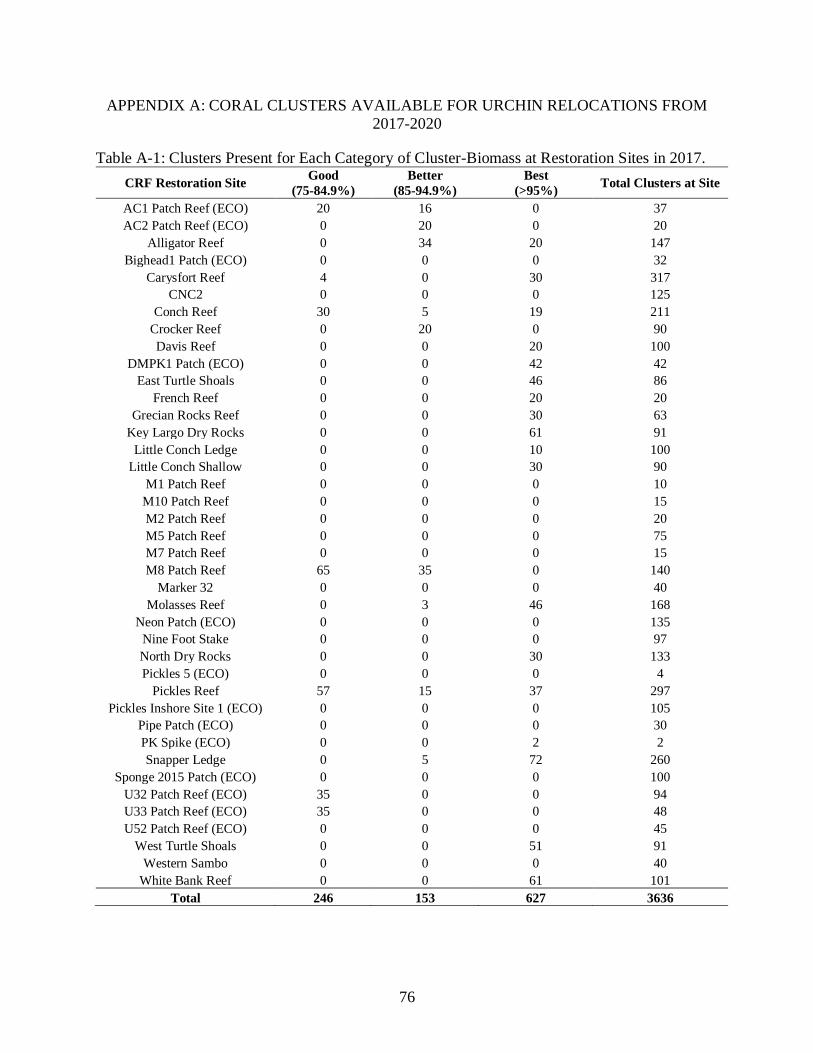
76
APPENDIX A: CORAL CLUSTERS AVAILABLE FOR URCHIN RELOCATIONS FROM
2017-2020
Table A-1: Clusters Present for Each Category of Cluster-Biomass at Restoration Sites in 2017.
CRF Restoration Site Good
(75-84.9%)
Better
(85-94.9%)
Best
(>95%) Total Clusters at Site
AC1 Patch Reef (ECO) 20 16 0 37
AC2 Patch Reef (ECO) 0 20 0 20
Alligator Reef 0 34 20 147
Bighead1 Patch (ECO) 0 0 0 32
Carysfort Reef 4 0 30 317
CNC2 0 0 0 125
Conch Reef 30 5 19 211
Crocker Reef 0 20 0 90
Davis Reef 0 0 20 100
DMPK1 Patch (ECO) 0 0 42 42
East Turtle Shoals 0 0 46 86
French Reef 0 0 20 20
Grecian Rocks Reef 0 0 30 63
Key Largo Dry Rocks 0 0 61 91
Little Conch Ledge 0 0 10 100
Little Conch Shallow 0 0 30 90
M1 Patch Reef 0 0 0 10
M10 Patch Reef 0 0 0 15
M2 Patch Reef 0 0 0 20
M5 Patch Reef 0 0 0 75
M7 Patch Reef 0 0 0 15
M8 Patch Reef 65 35 0 140
Marker 32 0 0 0 40
Molasses Reef 0 3 46 168
Neon Patch (ECO) 0 0 0 135
Nine Foot Stake 0 0 0 97
North Dry Rocks 0 0 30 133
Pickles 5 (ECO) 0 0 0 4
Pickles Reef 57 15 37 297
Pickles Inshore Site 1 (ECO) 0 0 0 105
Pipe Patch (ECO) 0 0 0 30
PK Spike (ECO) 0 0 2 2
Snapper Ledge 0 5 72 260
Sponge 2015 Patch (ECO) 0 0 0 100
U32 Patch Reef (ECO) 35 0 0 94
U33 Patch Reef (ECO) 35 0 0 48
U52 Patch Reef (ECO) 0 0 0 45
West Turtle Shoals 0 0 51 91
Western Sambo 0 0 0 40
White Bank Reef 0 0 61 101
Total 246 153 627 3636

77
Table A-2: Clusters Present for Each Category of Cluster-Biomass at Restoration Sites in 2018.
CRF Restoration Site Good
(75-84.9%)
Better
(85-94.9%)
Best
(>95%) Total Clusters at Site
AC1 Patch Reef (ECO) 0 0 37 37
AC2 Patch Reef (ECO) 0 0 20 20
Alligator Reef 0 0 147 147
Bighead1 Patch (ECO) 1 0 30 32
Carysfort Reef 20 7 79 317
CNC2 0 0 100 125
Conch Reef 26 2 177 211
Crocker Reef 0 0 50 90
Davis Reef 0 0 50 100
DMPK1 Patch (ECO) 0 0 42 42
East Turtle Shoals 0 0 86 86
French Reef 0 0 20 20
Grecian Rocks Reef 0 0 33 63
Key Largo Dry Rocks 0 0 61 91
Little Conch Ledge 0 0 35 100
Little Conch Shallow 0 0 90 90
M1 Patch Reef 0 0 0 10
M10 Patch Reef 0 0 0 15
M2 Patch Reef 0 0 0 20
M5 Patch Reef 0 0 0 75
M7 Patch Reef 0 0 0 15
M8 Patch Reef 0 0 100 140
Marker 32 0 0 0 40
Molasses Reef 0 4 164 168
Neon Patch (ECO) 0 0 135 135
Nine Foot Stake 0 55 0 97
North Dry Rocks 30 20 50 133
Pickles 5 (ECO) 0 0 4 4
Pickles Reef 21 56 190 297
Pickles Inshore Site 1 (ECO) 0 0 105 105
Pipe Patch (ECO) 0 0 30 30
PK Spike (ECO) 0 0 2 2
Snapper Ledge 32 19 189 260
Sponge 2015 Patch (ECO) 0 0 100 100
U32 Patch Reef (ECO) 0 0 94 94
U33 Patch Reef (ECO) 0 0 48 48
U52 Patch Reef (ECO) 0 0 45 45
West Turtle Shoals 0 0 51 91
Western Sambo 0 0 0 40
White Bank Reef 0 0 61 101
Total 130 163 2425 3636
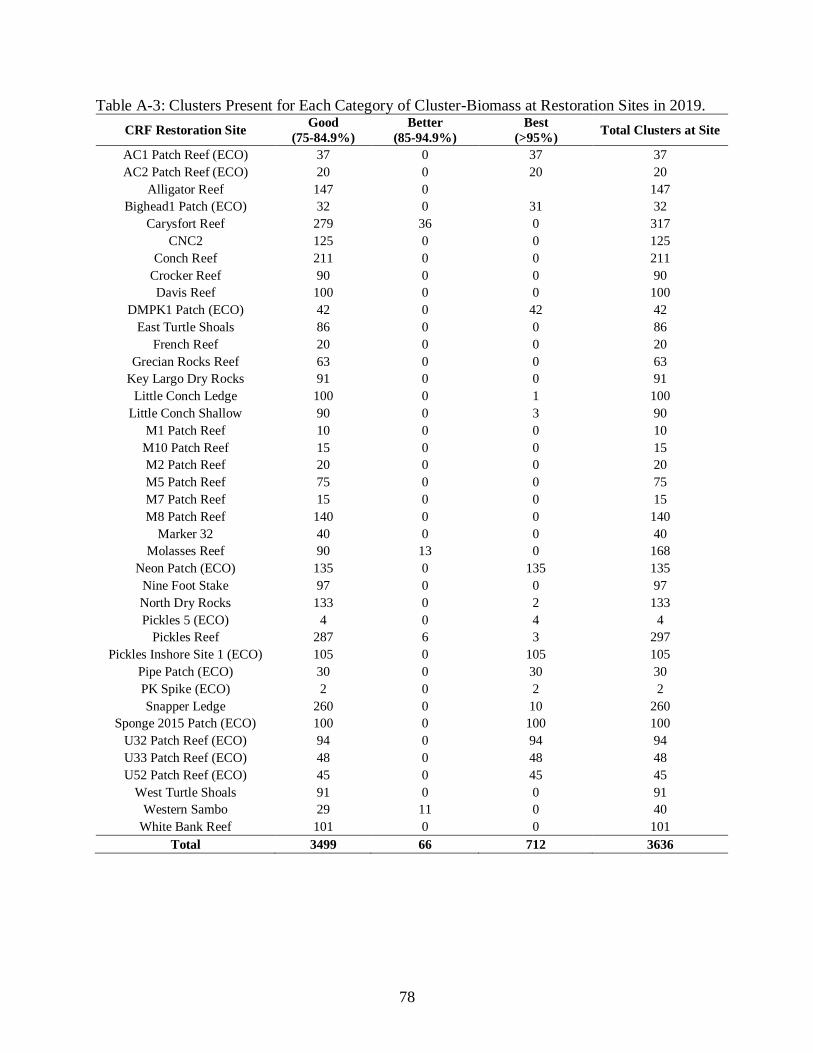
78
Table A-3: Clusters Present for Each Category of Cluster-Biomass at Restoration Sites in 2019.
CRF Restoration Site Good
(75-84.9%)
Better
(85-94.9%)
Best
(>95%) Total Clusters at Site
AC1 Patch Reef (ECO) 37 0 37 37
AC2 Patch Reef (ECO) 20 0 20 20
Alligator Reef 147 0 147
Bighead1 Patch (ECO) 32 0 31 32
Carysfort Reef 279 36 0 317
CNC2 125 0 0 125
Conch Reef 211 0 0 211
Crocker Reef 90 0 0 90
Davis Reef 100 0 0 100
DMPK1 Patch (ECO) 42 0 42 42
East Turtle Shoals 86 0 0 86
French Reef 20 0 0 20
Grecian Rocks Reef 63 0 0 63
Key Largo Dry Rocks 91 0 0 91
Little Conch Ledge 100 0 1 100
Little Conch Shallow 90 0 3 90
M1 Patch Reef 10 0 0 10
M10 Patch Reef 15 0 0 15
M2 Patch Reef 20 0 0 20
M5 Patch Reef 75 0 0 75
M7 Patch Reef 15 0 0 15
M8 Patch Reef 140 0 0 140
Marker 32 40 0 0 40
Molasses Reef 90 13 0 168
Neon Patch (ECO) 135 0 135 135
Nine Foot Stake 97 0 0 97
North Dry Rocks 133 0 2 133
Pickles 5 (ECO) 4 0 4 4
Pickles Reef 287 6 3 297
Pickles Inshore Site 1 (ECO) 105 0 105 105
Pipe Patch (ECO) 30 0 30 30
PK Spike (ECO) 2 0 2 2
Snapper Ledge 260 0 10 260
Sponge 2015 Patch (ECO) 100 0 100 100
U32 Patch Reef (ECO) 94 0 94 94
U33 Patch Reef (ECO) 48 0 48 48
U52 Patch Reef (ECO) 45 0 45 45
West Turtle Shoals 91 0 0 91
Western Sambo 29 11 0 40
White Bank Reef 101 0 0 101
Total 3499 66 712 3636
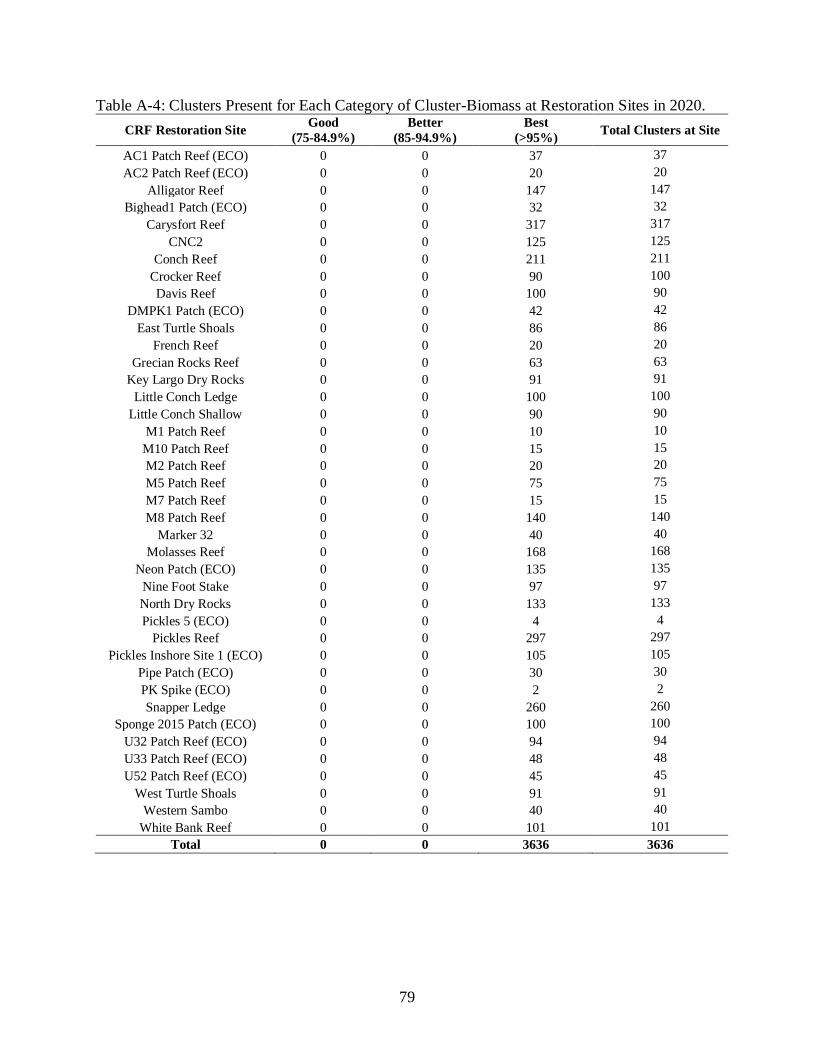
79
Table A-4: Clusters Present for Each Category of Cluster-Biomass at Restoration Sites in 2020.
CRF Restoration Site Good
(75-84.9%)
Better
(85-94.9%)
Best
(>95%) Total Clusters at Site
AC1 Patch Reef (ECO) 0 0 37 37
AC2 Patch Reef (ECO) 0 0 20 20
Alligator Reef 0 0 147 147
Bighead1 Patch (ECO) 0 0 32 32
Carysfort Reef 0 0 317 317
CNC2 0 0 125 125
Conch Reef 0 0 211 211
Crocker Reef 0 0 90 100
Davis Reef 0 0 100 90
DMPK1 Patch (ECO) 0 0 42 42
East Turtle Shoals 0 0 86 86
French Reef 0 0 20 20
Grecian Rocks Reef 0 0 63 63
Key Largo Dry Rocks 0 0 91 91
Little Conch Ledge 0 0 100 100
Little Conch Shallow 0 0 90 90
M1 Patch Reef 0 0 10 10
M10 Patch Reef 0 0 15 15
M2 Patch Reef 0 0 20 20
M5 Patch Reef 0 0 75 75
M7 Patch Reef 0 0 15 15
M8 Patch Reef 0 0 140 140
Marker 32 0 0 40 40
Molasses Reef 0 0 168 168
Neon Patch (ECO) 0 0 135 135
Nine Foot Stake 0 0 97 97
North Dry Rocks 0 0 133 133
Pickles 5 (ECO) 0 0 4 4
Pickles Reef 0 0 297 297
Pickles Inshore Site 1 (ECO) 0 0 105 105
Pipe Patch (ECO) 0 0 30 30
PK Spike (ECO) 0 0 2 2
Snapper Ledge 0 0 260 260
Sponge 2015 Patch (ECO) 0 0 100 100
U32 Patch Reef (ECO) 0 0 94 94
U33 Patch Reef (ECO) 0 0 48 48
U52 Patch Reef (ECO) 0 0 45 45
West Turtle Shoals 0 0 91 91
Western Sambo 0 0 40 40
White Bank Reef 0 0 101 101
Total 0 0 3636 3636

80
LIST OF REFERENCES
Alvarez-Filip L, Dulvy N, Gill J, Côté I, ARW (2009) Flattening of Caribbean coral reefs:
region-wide declines in architectural complexity. Proceedings of the Royal Society B
276: 3019-3025 doi 10.1098/rspb.2009.0339
Anholt B, Werner E (1995) Interaction Between Food Availability and Predation Mortality
Mediated by Adaptive Behavior. Ecology 76: 2230-2234 doi 10.2307/1941696
Anthony, KR, Marshall, PA, Abdulla, A, Beeden, R, Bergh, C, Black, R, Eakin, CM, Game, ET,
Gooch, M, Graham, NA, Green, A, Heron, SF, van Hooidonk, R, Knowland, C,
Mangubhai, S, Marshall, N, Maynard, JA, McGinnity, P, McLeod, E, Mumby, PJ,
Nyström, M, Obura, D, Oliver, J, Possingham, HP, Pressey, RL, Rowlands, GP,
Tamelander, J, Wachenfeld, D, Wear, S (2015) Operationalizing resilience for adaptive
coral reef management under global environmental change. Glob. Change Biol. 21, 48–
61.
Aronson R, Precht W (2001) White-band disease and the changing face of Caribbean coral reefs.
The Ecology and Etiology of Emerging Marine Diseases 159: 25-38 doi 10.1007/978-94-
017-3284-0_2
Bak R, van Eys G (1975) Predation of the sea urchin Diadema antillarum Philippi on living coral
SpringerLink. Oecologia 20: 111-115 doi 10.1007/BF00369023
Bauer J (1976) Growth, Aggregation, and Maturation in the Echinoid, Diadema Antillarum.
Bulletin of Marine Science 26: 273-277
Bauer J (1980) Observations on Geographical Variations in Population Density of the Echinoid
Diadema Antillarum within the Western North Atlantic. Bulletin of Marine Science 30:
509-515
Bauer J, Agerter C (1987) Isolation of Bacteria Pathogenic for the Sea Urchin Diadema
Antillarum (Echinodermata: Echinoidea). Bulletin of Marine Science 40: 161-165
Baums I (2008) A restoration genetics guide for coral reef conservation. Molecular Ecology 17:
2796-2811 doi 10.1111/j.1365-294X.2008.03787.x
Bechtel J, Gayle P, Kaufman L (2006) The return of Diadema antillarum to Discovery Bay:
patterns of distribution and abundance. In: Proceedings of 10th international coral reef
symposium, Okinawa 1:367–375
Beisner B, Haydon D, Cuddington K (2003) Alternative stable states in ecology. Frontiers in
Ecology and the Environment 1: 376-382 doi 10.1890/1540-
9295(2003)001[0376:ASSIE]2.0.CO;2
Bellwood D, Hughes T, Folke C, Nyström M (2004) Confronting the coral reef crisis. Nature
429: 827-833 doi doi:10.1038/nature02691

81
Bielmyer G, Brix K, Capo T, Grosell M (2005) The effects of metals on embryo-larval and adult
life stages of the sea urchin, Diadema antillarum. Aquatic Toxicology 74: 254–263 doi
10.1016/j.aquatox.2005.05.016
Bodmer M, Rogers A, Speight M, Lubbock N, Exton D (2015) Using an isolated population
boom to explore barriers to recovery in the keystone Caribbean coral reef herbivore
Diadema antillarum | SpringerLink. Coral Reefs 34: 1011-1021 doi 10.1007/s00338-015-
1329-4
Brander L, Beukering P, Cesar H (2007) The recreational value of coral reefs: A meta-analysis.
Ecological Economics 63: 209–218 doi 10.1016/j.ecolecon.2006.11.002
Brown C, Saunders M, Possingham H, Richardson A (2013) Interactions between global and
local stressors of ecosystems determine management effectiveness in cumulative impact
mapping. Diversity and Distributions 20: 538-546 doi 10.1111/ddi.12159
Burdick D (2008) The effectiveness of macroalgal reduction and Diadema antillarum addition in
limiting macroalgal growth and facilitating coral recovery 11th International Coral Reef
Symposium, Ft. Lauderdale, FL, pp 1204-1208
Carpenter R (1984) Predator and population density control of homing behavior in the Caribbean
echinoid Diadema antillarum. Marine Biology 82: 101-108 doi 10.1007/BF00392768
Carpenter R (1986) Partitioning Herbivory and Its Effects on Coral Reef Algal Communities.
Ecological Monographs 56: 345-364 doi 10.2307/1942551
Carpenter R, Edmunds P (2006) Local and regional scale recovery of Diadema promotes
recruitment of scleractinian corals. Ecology Letters 9: 271-280
Chandler L, Walters L, Sharp W, Hoffman E (In Press) Genetic structure of natural and
broodstock populations of the long-spined sea urchin, Diadema antillarum, throughout
the Florida Keys. Bulletin of Marine Science 93
Chiappone M, Rutten L, Swanson D, Miller S (2009) Population Status of the urchin Diadema
antillarum in the Florida Keys 25 years after the Caribbean mass mortality. 11th
International Coral Reef Symposium, Ft. Lauderdale, FL, pp 706-710
Chiappone M, Swanson D, Miller S, Smith S (2011) Large-scale surveys on the Florida Reef
Tract indicate poor recovery of the long-spined sea urchin Diadema antillarum. Coral
Reefs 21: 155-159 doi 10.1007/s00338-002-0232-y
Clemente S, Hernández JC, Toledo K, Brito A (2007) Predation upon Diadema aff. antillarum in
barren grounds of the Canary Islands. Scientia Marina 71: 745-754
Clemente S. and Hernandez J.C. (2008) Influence of wave exposure and habitat complexity in
determining spatial variation of the sea urchin Diadema aff. antillarum Echinoidea:

82
Diadematidae) populations and macroalgal cover (Canary Islands–Eastern Atlantic
Ocean). Biologı´a Tropical 56, 229–254.
Como S, Rossi F, Lardicci C (2006) Caging experiment: Relationship between mesh size and
artifacts. Journal of Experimental Marine Biology and Ecology 335: 157–166 doi
10.1016/j.jembe.2006.03.002
Costanza R, de Groot R, Sutton P, van der Ploeg S, Anderson S, Kubiszewski I, Farber S, Turner
R (2014) Changes in the global value of ecosystem services. Global Environmental
Change 26: 152–158 doi 10.1016/j.gloenvcha.2014.04.002
Crabbe M (2016) Comparison of Two Reef Sites on the North Coast of Jamaica over a 15-Year
Period. American Journal of Climate Change 5: 2 doi doi:10.4236/ajcc.2016.51002
Dame E (2008) Assessing the Effect of Artificial Habitat Structure on Translocation of the Long-
Spined Sea Urchin, Diadema antillarum, in Curaçao (Netherlands Antilles). Bulletin of
Marine Science 82: 247-254
Debrot AO, Nagelkerken I (2006) Recovery of the long-spined sea urchin Diadema antillarum in
Curaçao (Netherlands Antilles) linked to lagoonal and wave sheltered shallow rocky
habitats. Bull Mar Sci 79:415–424
Dixson D, Abrego D, Hay M (2014) Chemically mediated behavior of recruiting corals and
fishes: A tipping point that may limit reef recovery. Science 345: 892-897 doi
10.1126/science.1255057
Dizon R, Yap H (2005) Effects of coral transplantation in sites of varying distances and
environmental conditions. Marine Biology 148: 933-943 doi 10.1007/s00227-005-0142-y
Edmunds P, Carpenter R (2001) Recovery of Diadema antillarum reduces macroalgal cover and
increases abundance of juvenile corals on a Caribbean reef. PNAS 98: 5067-5071 doi
10.1073/pnas.071524598
Estes J, Palmisano J (1974) Sea otters: their role in structuring nearshore communities. Science
185: 1058-1060
Feehan C, Brown M, Sharp W, Lauzon-Gauy J, Adams D (2016) Fertilization limitation of
Diadema antillarum on coral reefs in the Florida Keys. Ecology 97: 1897-1904 doi
10.1002/ecy.1461
Filbee-Dexter K, Scheibling RE (2014) Sea urchin barrens as alternative stable states of
collapsed kelp ecosystems. Mar. Ecol. Prog. Ser. 495, 1– 25. (doi:10.3354/meps10573)
Forcucci D (1994) Population Density, recruitment and 1991 mortality event of Diadema
antillarum in the Florida Keys. Bulletin of Marine Science 54: 917-928

83
Fricke, HW (1974) Possible influence of predators on the behaviour of Diadema sea urchins.
Marine Biology 27, 59-62
Fung T, Seymour, RM, Johnson, CR (2011) Alternative stable states and phase shifts in coral
reefs under anthropogenic stress. Ecology 92: 967–982.
Gardner T, Cote I, Gill J, Grant A, Watkinson A (2003) Long-Term Region-Wide Declines in
Caribbean Corals. Science 301: 958-960 doi DOI: 10.1126/science.1086050
Graham N, Bellwood D, Cinner J, Hughes, TP, Norstrom A, Nystrom M (2013) Managing
resilience to reverse phase shifts in coral reefs. Frontiers in Ecology and the Environment
11: 541-548 doi 10.1890/120305
Graham N, Nash K (2013) The importance of structural complexity in coral reef ecosystems.
Coral Reefs 32: 315-326 doi 10.1007/s00338-012-0984-y
Gratwicke B, Speight M (2005) Effects of habitat complexity on Caribbean marine fish
assemblages. Marine Ecology Progress Series 292: 301-310 doi 10.3354/meps292301
Haley MP, Solandt JL (2001) Population fluctuations of the sea urchin Diadema antillarum and
Tripneustes ventricosus at Discovery Bay, Jamaica: a case of biological succession?
Caribb J Sci 37:239–245
Hall S, Raffaelli D, Turrell W (1990) Predator-Caging Experiments in Marine Systems: A
Reexamination of Their Value. The American Naturalist 136: 657-672
Harborne A, Renaud P, Tyler H, Mumby P (2009) Reduced density of the herbivorous urchin
Diadema antillarum inside a Caribbean marine reserve linked to increased predation
pressure by fishes. Coral Reefs 28: 783-791 doi 10.1007/s00338-009-0516-6
Hay M (1984) Patterns of Fish and Urchin Grazing on Caribbean Coral Reefs: Are Previous
Results Typical? Ecology 65: 446-454 doi 10.2307/1941407
Hay M, Taylor P (1985) Competition between herbivorous fishes and urchins on Caribbean
reefs. Oecologia 65: 591-598 doi 10.1007/BF00379678
Hendricks, ML, Hoopes, RL, Arnold, DA, and Kaufman, ML (2002) Homing of hatchery-reared
American shad to the Lehigh River, a tributary to the Delaware River. North American
Journal of Fisheries Management 22: 243-248.
Hereu B, Zabala M, Linares C, Sala E (2005) The effects of predator abundance and habitat
structural complexity on. Marine Biology 146: 293-299 doi 10.1007/s00227-004-1439-y
Hoey A, Bellwood D (2011) Suppression of herbivory by macroalgal density: a critical feedback
on coral reefs? Ecology Letters 14: 267-273 doi 10.1111/j.1461-0248.2010.01581.x

84
Holt R (1977) Predation, apparent competition, and the structure of prey communities.
Theoretical Population Biology 12: 197-229
Hughes T (1994) Catastrophes, Phase Shifts, and Large-Scale Degradation of a Caribbean Coral
Reef. Science 265: 1547-1551
Hughes T, Graham N, Jackson J, Mumby P, Steneck R (2010) Rising to the challenge of
sustaining coral reef resilience. Trends in Ecology and Evolution 25: 633-642 doi
10.1016/j.tree.2010.07.011
Hughes T, Reed D, Boyle M (1987) Herbivory on coral reefs: community structure following
mass mortalities of sea urchins ☆. Journal of Experimental Marine Biology and Ecology
113: 39–59 doi 10.1016/0022-0981(87)90081-5
Hughes T, Rodrigues M, Bellwood D, Ceccarelli D, Hoegh-Goldberg O, McCook L,
Moltschaniwskyj N, Pratchett M, Steneck R, Willis B (2007) Phase Shifts, Herbivory,
and the Resilience of Coral Reefs to Climate Change. Current Biology 17: 360-365 doi
10.1016/j.cub.2006.12.049
Idjadi J, Haring R, Precht W (2010) Recovery of the sea urchin Diadema antillarum promotes
scleractinian coral growth and survivorship on shallow Jamaican reefs. Marine Ecology
Progress Series 403: 91-100 doi 10.3354/meps08463
Idjadi J, Lee S, Bruno J, Precht W, Allen-Requa L, Edmunds P (2006a) Rapid phase-shift
reversal on a Jamaican coral reef. Coral Reefs 25: 209-211 doi 10.1007/s00338-006-
0088-7
Jones GP, McCormick MI, Srinivasan M, Eagle JV (2004) Coral decline threatens fish
biodiversity in marine reserves. PNAS 101: 8251-8253 doi 10.1073/pnas.0401277101
Karlson R, Levitan D (1989) Recruitment-limitation in open populations of Diadema antillarum:
an evaluation | SpringerLink. Oecologia 82: 40-44 doi 10.1007/BF00318531
Kier, PM, Grant, RE (1965) Echinoid distribution and habits, Key Largo Coral Reef Preserve,
Florida. Smithson. Misc. Collect., 149(6): 68 pp.
Kintzing M, Butler M (2014) The Influence of Shelter, Conspecifics, and Threat of Predation on
the Behavior of the Long-Spined Sea Urchin (Diadema antillarum). Journal of Shellfish
Research 33: 781-785 doi 10.2983/035.033.0312
Kissling D, Precht W, Miller S, Chiappone M (2014) Historical reconstruction of population
density of the echinoid Diadema antillarum on Florida Keys shallow bank-barrier reefs.
Bulletin of Marine Science 90: 665-679 doi info:doi/10.5343/bms.2013.1022
Knowlton N (1992) Thresholds and Multiple Stable States in Coral Reef Community Dynamics.
Integrative and Comparative Biology 32: 674-682 doi 10.1093/icb/32.6.674

85
Knowlton N (2001) The future of coral reefs. PNAS 98: 5419–5425 doi
10.1073/pnas.091092998
Kobayashi N (1980) Comparative sensitivity of various developmental stages of sea urchins to
some chemicals. Marine Biology 58: 163-171 doi 10.1007/BF00391872
Lawrence J, Cao X, Chang Y, Wang P, Yu Y, Lawrence A, Watts, SA (2009) Temperature
Effect on Feed Consumption, Absorption, and Assimilation Efficiencies and Production
of the Sea Urchin Strongylocentrotus intermedius. Journal of Shellfish Research 28: 389-
395 doi 10.2983/035.028.0223
Lee SC (2006) Habitat complexity and consumer‐mediated positive feedbacks on a Caribbean
coral reef. Oikos 112: 442-447 doi 10.1111/j.0030-1299.2006.14247.x
Lee T, Williams E (1999) Mean distribution and seasonal variability of coastal currents and
temperature in the Florida Keys with implications for larval recruitment. Bulletin of
Marine Science 64: 25-56
Lessios H (1988a) Mass Mortality of Diadema antillarum in the Caribbean: What Have We
Learned? Annual Review of Ecology and Systematics 19: 371-393
Lessios H (1988b) Population dynamics of Diadema antillarum (Echinodermata: Echinoidea)
following mass mortality in Panamá. Marine Biology 99: 515-526 doi
10.1007/BF00392559
Lessios H, Cubit J, Robertson D, Shulman M, Parker M, Garrity S, Levings, SC (1984) Mass
mortality of Diadema antillarum on the Caribbean coast of Panama. Coral Reefs 3: 173-
182 doi 10.1007/BF00288252
Lessios H, Garrido M, Kessing B (2001) Demographic history of Diadema antillarum, a
keystone herbivore on Caribbean reefs doi 10.1098/rspb.2001.1806
Lessios H, Robertson D, Cubit J (1984) Spread of Diadema mass mortality throughout the
Caribbean. Science 226: 335-337
Lessios HA (2016) The great Diadema antillarum die-off: 30 years later. Annu. Rev. Mar.
Sci. 8:267–83
Levitan DR (1988a) Algal-urchin biomass responses following mass mortality of Diadema
antillarum Philippi at Saint John, U.S. Virgin Islands. Journal of Experimental Marine
Biology and Ecology 119: 167–178 doi 10.1016/0022-0981(88)90231-6
Levitan, DR (1988b) Asynchronous spawning and aggregative behavior in the sea urchin
Diadema antillarum Phillipi. Pp. 18 I- 186 in Echinoderm Biology. Proceedings of the
Sixth International Echinoderm Conference, R. Burke, ed. Balkema, Rotterdam.

86
Levitan DR (1989) Density‐Dependent Size Regulation in Diadema Antillarum: Effects on
Fecundity and Survivorship. Ecology 70: 1414-1424 doi 10.2307/1938200
Levitan, DR, Genovese, SJ (1989) Substratum dependant predator—prey dynamics: patch reefs
as refuges from gastropod predation. Journal of Experimental Marine Biology and
Ecology 130, 111–118.
Lewis J (2011) Feeding and digestion in the tropical sea urchin Diadema antillarum Philippi.
http://dxdoiorg/101139/z64-048 42: 549-557 doi 10.1139/z64-048
Liddell W, Ohlhorst, SL (1986) Changes in benthic community composition following the mass
mortality of Diadema at Jamaica. Journal of Experimental Marine Biology and Ecology
95: 271-278
Lirman D (2001) Competition between macroalgae and corals: effects of herbivore exclus. Coral
Reefs doi 10.1007/s003380000125
Lirman D, Schopmyer S, Galvan V, Drury C, Baker A, Baums I (2014) Growth Dynamics of the
Threatened Caribbean Staghorn Coral Acropora cervicornis: Influence of Host Genotype,
Symbiont Identity, Colony Size, and Environmental Setting. PLOSone. doi
10.1371/journal.pone.0107253
Macia S, Robinson M, Nalevanco A (2007) Experimental dispersal of recovering Diadema
antillarum increases grazing intensity and reduces macroalgal abundance on a coral reef.
Marine Ecology Progress Series 348: 173-182 doi 10.3354/meps06962
Macintyre I, Glynn P, Hinds F (2005) Evidence of the role of Diadema antillarum in the
promotion of coral settlement and survivorship. Coral Reefs 24: 273-273 doi
10.1007/s00338-005-0492-4
Maciá S, Robinson M, Nalevanko A (2007) Experimental dispersal of recovering Diadema
antillarum increases grazing intensity and reduces macroalgal abundance on a coral reef.
Marine Ecology Progress Series 348: 173-182 doi 10.3354/meps06962
Maliao R, Turingan R, Lin J (2008) Phase-shift in coral reef communities in the Florida Keys
National Marine Sanctuary (FKNMS), USA. Marine Biology 154: 841-853 doi
10.1007/s00227-008-0977-0
McBride, SC, Pinnix, WD, Lawrence, JM, Lawrence, AL, Mulligan, TM (1997) The effect of
temperature on production of gonads by the sea urchin Strongylocentrotus franciscanus
fed natural and prepared diets. J. World Aquat. Soc.: 28 :357-365.
McClanahan T (1999) Predation and the Control of the Sea Urchin Echinometra viridisand
Fleshy Algae in the Patch Reefs of Glovers Reef, Belize. Ecosystems 2: 511-523 doi
10.1007/s100219900099

87
McClanahan, TR and Muthiga, NA (1988) Changes in Kenyan coral reef community structure
and function due to exploitation. Hydrobiologia 166, 269–76.
McClanahan T, Mwachireya S (1994) Fish and sea urchin herbivory and competition in Kenyan
coral reef lagoons: the role of reef management - ScienceDirect. Journal of Experimental
Marine Biology and Ecology 184: 237-254
McCook L (1999) Macroalgae, nutrients and phase shifts on coral reefs: scientific issu. Coral
Reefs 18: 357-367 doi 10.1007/s003380050213
McCook L, Jompa J, Diaz-Pulido G (2001) Competition between corals and algae on coral reefs:
a review of evidence and mechanisms. Coral Reefs 19: 400-417 doi
10.1007/s003380000129
Mieog J, Olsen J, Berkelmans R, Bleuler-Martinez S, Willis B, van Oppen M (2009) The Roles
and Interactions of Symbiont, Host and Environment in Defining Coral Fitness. PLOSone
4 doi 10.1371/journal.pone.0006364
Miller M, Bourque A, Bohnsack J (2002) An analysis of the loss of acroporid corals at Looe
Key, Florida, USA: 1983–2000. Coral Reefs 21: 179-182 doi 10.1007/s00338-002-0228-
7
Miller R, Adams A, Ogden N, Ogden J, Ebersole J (2003) Diadema antillarum 17 years after
mass mortality: is recovery beginning on St. Croix? Coral Reefs 22: 181-187 doi
10.1007/s00338-003-0301-x
Miller R, Adams A, Ebersole J, Ruiz E (2007) Evidence for positive density-dependent effects in
recovering Diadema antillarum populations. Journal of Experimental Marine Biology
and Ecology 349: 215–222 doi 10.1016/j.jembe.2007.05.014
Miller L, Gaylord B (2007) Barriers to flow: The effects of experimental cage structures on
water velocities in high-energy subtidal and intertidal environments. Journal of
Experimental Marine Biology and Ecology 344: 215-228
Miller M, AMS (2006) Lessons-learned from the experimental key species restoration. CRC
Press, Boca Raton, FL
Miller M, Kerr K, Williams D (2016) Reef-scale trends in Florida Acropora spp. abundance and
the effects of population enhancement. PeerJ 4 doi 10.7717/peerj.2523
Millot, N (1953) Light emission and light perception in species of Diadema. Nature, Lond. 171,
973–974
Millot, N (1954) Sensitivity to light and the reactions to changes in light intensity of the
echinoid Diadema antillarum Philippi. Phil. Trans. R. Soc. 238, 187–220

88
Moses CS, Bonem RM (2001) Recent population dynamics of Diadema antillarum and
Tripneustes ventricosus along the north coast of Jamaica, W. I. Bulletin of Marine
Science 68: 327-336
Mumby P (2006) The Impact Of Exploiting Grazers (Scaridae) On The Dynamics Of Caribbean
Coral Reefs. Ecological Applications 16: 747-769 doi 10.1890/1051-
0761(2006)016[0747:TIOEGS]2.0.CO;2
Mumby P, Hastings A, Edwards H (2007) Thresholds and the resilience of Caribbean coral reefs.
Nature 450: 98-101 doi 10.1038/nature06252
Mumby P, Hedley J (2006) Revisiting the catastrophic die-off of the urchin Diadema antillarum
on Caribbean coral reefs: Fresh insights on resilience from a simulation model.
Ecological Modelling 196: 131–148 doi 10.1016/j.ecolmodel.2005.11.035
National Marine Fisheries Service. March 2015. Recovery Plan for Elkhorn (Acropora palmata)
and Staghorn (A. cervicornis) Corals. Prepared by the Acropora Recovery Team for the
National Marine Fisheries Service, Silver Spring, MD. 167 p.
National Ocean and Atmospheric Administration (2016) Workshop to Advance the Science and
Practice of Caribbean Coral Restoration, Ft. Lauderdale, FL
Nedimyer K, Moe M (2011) Techniques Development for the Re-establishment of the Long-
spined Sea Urchin, Diadema antillarum, on Two Small Patch Reefs
in the upper Florida Keys. National Oceanic and Atmospheric Administration
Norström, AV, Nyström, M , Lokrantz , J, and Folke, C (2009) Alternative states on coral reefs:
beyond coral macroalgal phase shifts. Marine Ecology Progress Series 376: 295–306.
Ogden J, Brown R, Slaseky N (1973) Grazing by the Echinoid Diadema antillarum Philippi:
Formation of Halos around West Indian Patch Reefs. Science 182: 715-717
Ogden JC, Lobel PS (1978) The role of herbivorous fishes and urchins in coral reef
communities. Environmental Biology of Fishes 3: 49-63 doi 10.1007/BF00006308
Paddack, EA (2009) Recent Region-wide Declines in Caribbean Reef Fish Abundance. Current
Biology 19: 590-595 doi 10.1016/j.cub.2009.02.041
Porter JW, Lewis SK, Porter KG (2015) The effect of multiple stressors on the Florida Keys
coral reef ecosystem: A landscape hypothesis and a physiological test. Limnology and
Oceanography 44: 941-949 doi 10.4319/lo.1999.44.3_part_2.0941
Randall J, Schroeder R, Starck W (1964) Notes on the Biology of the Echinoid Diadema
antillarum. Carib J Sci 4: 421-433

89
Robertson D (1991) Increases in surgeonfish populations after mass mortality of the sea urchin
Diadema antillarum in Panamá indicate food limitation. Marine Biology 111: 437-444
doi 10.1007/BF01319416
Rogers A, Lorenzen K (2016) Does Slow and Variable Recovery of Diadema antillarum on
Caribbean Fore-Reefs Reflect Density-Dependent Habitat Selection? Frontiers in Marine
Science 3 doi doi:10.3389/fmars.2016.00063
Sammarco P (1980) Diadema and its relationship to coral spat mortality: Grazing, competition,
and biological disturbance. Journal of Experimental Marine Biology and Ecology 45:
245-272 doi 10.1016/0022-0981(80)90061-1
Sammarco P (1982) Effects of grazing by Diadema antillarum Philippi (Echinodermata:
Echinoidea) on algal diversity and community structure. Journal of Experimental Marine
Biology and Ecology 65: 83-105 doi doi:10.1016/0022-0981(82)90177-0
Scheffer M, Carpenter S, Foley J, Folke C, Walker B (2001) Catastrophic shifts in ecosystems.
Nature 413: 591-596 doi doi:10.1038/35098000
Shulman MJ (1984) Resource limitation and recruitment patterns in a coral reef fish assemblage.
Journal of Experimental Marine Biology and Ecology 74: 85-109
Shulman M (1990) Aggression among sea urchins on Caribbean coral reefs. Journal of
Experimental Marine Biology and Ecology 140: 239-245
Spirlet C, Grosjean P, Jangoux M (2000) Optimization of gonad growth by manipulation of
temperature and photoperiod in cultivated sea urchins, Paracentrotus lividus (Lamarck)
(Echinodermata)185: 85–99 doi 10.1016/S0044-8486(99)00340-3
Steiner S, Williams S (2006) The density and size distribution of Diadema antillarum in
Dominica (Lesser Antilles): 2001–2004. Marine Biology 149: 1071-1078 doi
10.1007/s00227-006-0279-3
Stocker L (1986) Artifactual effects of caging on the recruitment and survivorship of a subtidal
colonial invertebrate. Marine Ecology Progress Series 34: 305-307
Tanner J (1995) Competition between scleractinian corals and macroalgae: An experimental
investigation of coral growth, survival and reproduction. Journal of Experimental Marine
Biology and Ecology 190: 151–168 doi 10.1016/0022-0981(95)00027-O
The Florida Aquarium Center for Conservation and Disney’s Animals, Science and Environment
(2017) Long-Spined Sea Urchin Workshop: Husbandry and Research Needs to Help
Restore Coral Reefs, Apollo Beach, FL
Tuya F, Martin J, Luque A (2003) Patterns of nocturnal movement of the long-spined sea urchin
Diadema a. Helgoland Marine Research 58: 26-31 doi 10.1007/s10152-003-0164-0

90
Tuya F, Boyra A, Sanchez-Jerez P, Barbera C, Haroun R (2004) Relationships between rocky-
reef fish assemblages, the sea urchin Diadema antillarum and macroalgae throughout the
Canarian Archipelago. Marine Ecology Progress Series 278: 157-169 doi
10.3354/meps278157
Wallace M, Temple S (1987) Releasing Captive-Reared Andean Condors to the Wild. The
Journal of Wildlife Management 51: 541-550 doi 10.2307/3801266
Weil E, Losada F, Bone D (1984) Spatial Variation in density and size of the echinoid Diadema
antillarum Philippi on some Venezuelan coral reefs. Bijdragen tot de Dierkunde 54: 73-
82
Williams D, Miller M, Kramer K (2008) Recruitment failure in Florida Keys Acropora palmata,
a threatened Caribbean coral. Coral Reefs 27: 697-705 doi 10.1007/s00338-008-0386-3
Williams I, Polunin N (2001) Large-scale associations between macroalgal cover and grazer
biomass on mid-depth reefs in the Caribbean. Coral Reefs 19: 358-366 doi
10.1007/s003380000121
Young C, Schopmeyer S, Lirman D (2012) A Review of Reef Restoration and Coral Propagation
Using the Threatened Genus Acropora in the Caribbean and Western Atlantic. Bulletin of
Marine Science 88: 1075-1098 doi/10.5343/bms.2011.1143
Zaneveld J, Burkepile D, Shantz A, Pritchard C, McMinds R, Payet J, Welsh R, Correa A,
Lemoine N, Rosales S, Fuchs C, Maynard J, Thurber R (2016) Overfishing and nutrient pollution
interact with temperature to disrupt coral reefs down to microbial scales. Nature
Communications. doi:101038/ncomms11833 doi doi:10.1038/ncomms11833

91
BIOGRAPHICAL SKETCH
Kayla graduated spring of 2017 with a Master of Science in Fisheries and Aquatic
Sciences. Kayla was a distance student residing in Key Largo, Florida where she was employed
at Coral Restoration Foundation.