synaptic reorganization in explanted cultures of rat hippocampus
TRANSCRIPT

Ž .Brain Research 815 1999 304–316
Research report
Synaptic reorganization in explanted cultures of rat hippocampus
Rafael Gutierrez a,b,c,), Uwe Heinemann a´a Institut fur Physiologie der Charite, Humboldt UniÕersitat zu Berlin, Tucholsky-Str. 2, 10117 Berlin, Germany¨ ´ ¨
b Departamento de Fisiologıa, Biofısica y Neurociencias del CINVESTAV, Apdo. Postal 14-740, Mexico, D.F. 07000, Mexico´ ´ ´c Instituto Mexicano de Psiquiatrıa, Laboratory of Neurophysiology, Calzada Mexico-Xochimilco 101, Tlalpan 14370, Mexico´ ´
Accepted 13 October 1998
Abstract
Due to loss of afferent innervation, synaptic reorganization occurs in organotypic hippocampal slice cultures. With extra- andŽ .intracellular recordings, we confirm that the excitatory loop from the dentate gyrus DG to CA3 and further to CA1 is preserved.
However, hilar stimulation evoked antidromic population spikes in the DG which were followed by a population postsynaptic potentialŽ .PPSP ; intracellularly, an antidromic spike with a broad shoulder or EPSPrIPSP sequences were induced. Synaptic responses wereblocked by glutamate receptor antagonists. Stimulation of CA1 induced a PPSP in DG. Dextranamine stained pyramidal cells of CA1
Ž .were shown to project to DG. After removal of area CA3, DG’s and mossy fibers’ MF stimulation still elicited PPSPs and EPSPrIPSPsequences in area CA1 which disappeared when a cut was made through the hippocampal fissure. During bicuculline perfusion, hilarstimulation caused EPSPs in granule cells and spontaneous and evoked repetitive firing appeared even after its isolation from areas CA3and CA1. Collateral excitatory synaptic coupling between granule cells was confirmed by paired recordings. Besides the preservation ofthe trisynaptic pathway in this preparation, new functional synaptic contacts appear, presumably due to MF collateral sprouting andformation of pathways between areas CA1 and DG. q 1999 Elsevier Science B.V. All rights reserved.
Keywords: Hippocampal cultured slice; Mossy fiber; Recurrent excitation; Dentate gyrus
1. Introduction
Ž .In the normal hippocampus, the dentate gyrus DGŽ .granule cells activate via the mossy fiber pathway MF
w xCA3 pyramidal cells 49 , which in turn innervate areaCA1. Direct projections from the DG to area CA1 and
w xvice-versa do not normally exist 24,33 . Mossy fibersoriginate from both leaflets of the DG and project via
w xinfra- and suprapyramidal tracts to area CA3 7 . Unlikew xCA3 pyramidal cells 26 , granule cells do not interact with
each other by recurrent or collateral excitatory synapsesw x12 , unless a chronic form of temporal lobe epilepsy had
w xbeen induced 6,35 . The excitability of granule cells iscontrolled by an elaborated network of feedback and feed-forward inhibition, whereby the DG may serve as a filterfor the convergent input from many sensory systems to the
) Corresponding author. Departamento de Fisiologıa, Biofısica y Neu-´ ´rociencias del CINVESTAV, Apdo. Postal 14-740, Mexico, D.F. 07000.´Fax: q52-5-747-7105; E-mail: [email protected]
hippocampus proper and also serves to prevent spread ofepileptiform activity from the entorhinal cortex to the
w xhippocampus 3,20,27,34 .Although organotypic cultures of the hippocampus
w xmaintain a high degree of tissue organization 15–17 , andthe electrophysiological properties of the MF-pyramidalcell connections have been found to be comparable to the
w xin vivo situation 16 , in vitro culture conditions will favorthe formation of aberrant synapses due to loss of afferentfiber input from the contralateral hippocampus, the sep-tum, the entorhinal cortex and other structures. In slicescultured with the roller tube technique Zimmer and
w x w xGahwiler 51 , and Caeser and Aertsen 5 , have consis-¨tently described MF sprouting to the molecular layer of theDG.
Reorganization of hippocampal synaptic circuitry alsooccurs under pathological conditions, such as temporallobe epilepsy. There, the MF sprout into the inner molecu-lar layer of the DG and this is thought to cause recurrentexcitatory synaptic transmission. Evidence for MF sprout-
0006-8993r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0006-8993 98 01101-9

( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´ 305
ing has been found in several models of chronic epilepsyw x4,6,14,25,48 , in tissue of genetically epileptic-prone ani-
w x w xmals 32 , and tissue of epileptic patients 2,21 , afterw x w xkainate injections 10,12,29,30,35,36 , and kindling 35,36 ,
where giant synaptic boutons develop and make contactwith granule cell dendrites.
Although formation of aberrant synaptic contacts hasw xbeen previously reported 13,51,52 , in anatomical studies
using the roller tube slice culture technique, a functionalcharacterization of newly formed synapses in the culturedhippocampus has not yet been carried out. Here, we pre-sent electrophysiological and morphological evidence forsuch reorganization using the air to medium interface
w xtechnique 46 . Some of these results were reported inw xabstract form 19 .
2. Materials and methods
Cultured hippocampal slices were prepared from 6 to 9w xdays old Wistar rats as described by Stoppini et al. 46 .
Briefly, after decapitation under deep ether anesthesia, thehippocampi were dissected and 400 mm thick slices were
Ž .cut McIlwain tissue chopper under sterile conditions inŽ .gassed 95% O –5% CO ice-cold preparation medium2 2
Žminimum essential medium, MEM, Gibco, Eggenstein,. ŽGermany; pH 7.35 and placed on culture membranes 0.4
mm Millicell culture plate inserts; Millipore, Eschborn,.Germany . Two slices were seeded on each culture mem-
brane without having contact with each other. The mem-Žbranes were placed in 6-chamber incubation wells Falcon,
.New Jersey, USA containing 1.2 ml of the culturingŽmedium 50% MEM, 25% Hank’s balanced salts solution,
HBSS, Sigma, Deisenhofen, Germany; 25% horse serum,Gibco; penicillin and streptomycin 10 000 ur10 000mgrml, Gibco; NaHCO , 7.5%; L-glutamin, 2 mM; pH3
. Ž . Ž7.3 and kept in a CO 5% incubator Heraeus, Hanau,2.Germany at 36.58C. They were fed exchanging 0.6 ml of
the culturing medium with freshly prepared and previouslyprewarmed medium every other day.
For recordings, the cultured slices were transferred onthe membranes to an interface chamber and constantly
Ž . Žperfused with prewarmed 358C and oxygenated 95%. Ž .O –5% CO artificial cerebrospinal fluid ACSF con-2 2
taining—in mM—NaCl, 124; KCl, 3; NaH PO , 1.25;2 4
MgSO , 2; CaCl , 2; NaHCO , 26; glucose 10; pH 7.35.4 2 3
The drugs used were diluted at the desired concentration inŽ .the ACSF, namely: DL -2-amino-5-phosphonovaleric acid
Ž2APV; 30 mM; Research Biochemicals, Natick, MA,. Ž .USA ; 6-nitro-7-sulphamoylbenzo f quinolaxine-2,3-dione
Ž .NBQX; 10 mM; gift from Novo Nordisk, Denmark andŽ .bicuculline methiodide 20 mM; Sigma . Extracellular and
intracellular recordings were performed in different sets ofŽa total of 70 cultured slices at different ages in vitro see
.Section 3 .
ŽBipolar platinum wire electrodes diameter 50 mm,f50–100 mm tip separation; approximate resistance 0.6
.MV were used for stimulation that consisted of squarepulses with a duration of 0.1 ms applied over the hilarformation of the DG or just over the proximal CA3 area tostimulate the MF pathway or within the granule cell or
Žmolecular layers of the DG 200–300 mm apart from the. Žrecording site , and at various sites in the CA1 region see
.below . As controls, extracellular responses of the DG tohilar stimulation were also recorded from slices acutely
Ž . Žprepared from juvenile rats ns4 at an age 10 to 15. Ždays old matching that of the cultured slices 6–9 days
.after birthq4–9 days in vitro . Care was taken to stimu-late the hilar region at about the same distance to therecording electrode as done in the slice cultures.
Ž .Glass microelectrodes 10 MV filled with 150 mMNaCl were used for field potential recordings. For intra-
Ž .cellular recordings, glass microelectrodes 70–100 MV
Žwere pulled with a Brown Flaming horizontal puller Sutter.Instruments and filled with 2 M potassium acetate. The
signals were acquired with conventional extra- and intra-cellular recording techniques and off-line processed with
Ž .the pCLAMP6 program Axon Instruments .Ž .In four cultured slices 7–15 DIV tetramethyl–rhoda-
Ž .mine–dextranamine Molecular Probes and fluorescein–Ž .dextranamine Molecular Probes were used for staining of
granule and CA1 pyramidal cells, respectively. Small crys-tals were placed in the appropriate regions, where some ofthe dye was taken up into the neurons and anterogradelyand retrogradely transported. These slices were re-in-cubated for 12 h before fixation. Proper filters were usedfor taking microphotographs of the fluorescent stainings.
3. Results
The different experiments were carried out in severalsets of slices out of a total of 70 cultured slices at different
Ž .intervals after explantation see below . Some slices wererecorded on two or three consecutive days. The time inculture of specific representative experiments is indicatedin the figure legends. Up to 2 days after explantation, theslice cultures had a whitish appearance. Thereafter, theybecame more transparent and the pyramidal cell layers andthe U-shaped DG were clearly distinguishable although thelower blade of the DG could remain somewhat thinnerthan the superior blade neighboring area CA1. Althoughcare was taken to completely isolate the hippocampus, itcannot be excluded that a portion of the entorhinal cortexmight be present in the explants. Except for a broadeningof the granule and pyramidal cell layers that did not seemto be as extended as that observed with the roller tube
w xtechnique 5,15 , the hippocampal slices cultured with theair to medium interface technique maintained their grosscytoarchitecture.

( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´306
3.1. ConnectiÕity of the slice culture: field potentialrecordings
The normal flow of information in the hippocampusw xfollows a trisynaptic pathway 1 . The input to the DG is
conveyed by the perforant path which synapses onto gran-ule cells which in turn project through the mossy fibers toarea CA3 and this to area CA1 through the Schaffer
Ž .collaterals Fig. 1A . To assess the connectivity of thedifferent hippocampal regions, field potential recordingswere carried out systematically in all areas of the hip-pocampus proper and the DG in 50 cultures. Stimulation ofthe hilus, where the mossy fibers pass on their way fromthe DG to CA3, always evoked a population EPSP or a
spike, depending on stimulus intensity, in the pyramidalŽ .cell layers of area CA3 Fig. 2D and a population EPSP in
Ž .CA1 Fig. 2B . In the granule cell layer of the DG, anantidromic population spike was elicited by hilar stimula-tion. In contrast to the population responses obtained from
Žthe slices acutely prepared from juvenile rats ns4; Fig..3C the antidromic population spike was followed by a
Ž .high amplitude PSP Fig. 1B . Depending on stimulusintensity, field potential responses could reach amplitudes
Ž .of up to 15 mV see Fig. 1 , but usually responses of 3–6mV were elicited for routine analysis. The threshold stimu-lus intensity for eliciting a population spike was usuallyaround 2.5 V, however, field PSP could be elicited beforethe apparition of population spikes. Also, to check whether
Ž . Ž .Fig. 1. A Diagram depicting the normal pattern of information flow within the hippocampus. B Schematic representation of the hippocampus and fieldpotential recordings evoked by hilar stimulation in a slice on the 7th DIV. The numbers accompanying the traces signal the corresponding recordingpositions noted in the schema. Notice the antidromic population spike followed by a population PSP in the granular layer and the shift of polarity in themolecular layer of the DG. All traces are averages of six field potentials evoked at 0.1 Hz. The asterisks show the position of the stimulating bipolarelectrode. Abbreviations: PP—perforant path; DG—dentate gyrus; GC—granular cell; Int—interneuron; MrH-C—mossyrhilar cells; SUBIC—sub-iculum; HF—hippocampal fissure; H—hilus; MF—mossy fibers; SC—Schaffer collaterals.

( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´ 307
Ž . Ž .Fig. 2. A Schematic representation of a cultured hippocampal slice 7 DIV . Numbers in the CA1 region signal different stimulation sites. Lines I and IIŽ . Ž .represent the sites of the cuts see Section 2 . B Field potential recordings in the pyramidal layer of CA1 upon hilar stimulation in control condition andŽ . Ž . Ž .after making the different cuts. C Responses in the granule cell layer of DG B upon stimulation at the sites indicated by the numbers before Control
Ž . Ž . Ž . Ž .and after separation of DG from CA1. D Responses in CA3 I upon stimulation in CA1 Stim 3 and MF Stim 6 after cut I. All traces are averages of6–10 field potentials evoked at 0.1 Hz.
enhanced activity after preparation and culturing of theslices with penicillin present in the medium could beresponsible for synaptic reorganization, eight slices wereincubated during 5 days in culture medium containing
Ž .TTX 1 mM and recorded in the presence and after washout of TTX. After wash-out of TTX, these slice culturesresponded with the same high amplitude PSP in the DG
Ž .upon hilar stimulation not shown .

( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´308
Ž .Fig. 3. A Superimposed field potentials evoked in the DG by hilar stimulation with a paired pulse protocol at 10, 20, 30, 40, 50, 60, 80, 100, 120 and 150Ž .ms in a cultured slice 18 DIV . Notice the interval-dependent inhibition of both the recurrent population PSP and the antidromic population spike.
Ž .Stimulus artifacts were deleted for clarity. B Superimposed field potentials evoked by hilar stimulation in the DG in control condition and afterŽ . Ž .pharmacological treatment. Left panel shows the action of bicuculline arrow on the population PSP; middle panel, effect of NBQX arrow and right
Ž .panel, complete block of the population PSP arrow with simultaneous perfusion of NBQX and 2APV. The recordings were taken from different slices onŽ .the 8th DIV. C Evoked field potential in the DG upon hilar stimulation in an acutely prepared slice of a 15 day-old rat. Notice the lack of the population
PSP after the antidromic spike. All traces are averages of 6–10 field potentials evoked at 0.1 Hz.
Stimulation of the stratum radiatum where the Schaffercollaterals pass on their way from area CA3 to area CA1,evoked a population PSP and spike in CA1 and an an-tidromic spike in CA3. Interestingly, stimulation of areaCA1 at different positions evoked a population PSP, even-tually superimposed by a population spike in the granule
Ž . Ž .cell layer of the DG Fig. 2C ns6 .Fig. 1B illustrates field potential responses in various
layers of the DG induced by hilar stimulation. In thegranule cell layer of the DG, an antidromic populationspike was followed by a positive field potential whichreversed in polarity as the recording electrode was movedtowards the stratum moleculare. Upon hilar stimulation,the population PSP that followed the antidromic spike in
Ž .the DG was present in all the tested cultures ns70 .These field potential responses were present also in cul-
Ž .tures 1 month after explantation ns7 . Interestingly,field potential responses in the DG could be elicited byhilar stimulation without being preceded by an antidromicpopulation spike.
To further assess the flow of excitation within thetrisynaptic pathway DG–CA3–CA1, cuts were carried out
at different levels of this pathway to selectively disrupt theŽinformation transfer between the different regions ns9
. Ž . Ž .cultured slices after 7–8 days in vitro DIV Fig. 2 .After determining the control population responses to stim-ulation of the hilus and of area CA1, cuts were made witha sharp blade under microscope observation leaving theculture membranes intact and the dissected regions wereseparated 2 mm. Recordings were resumed an hour afterthe cuts. Isolated areas were then stimulated with the sameintensity used before the cuts to check for responses thatwould indicate remaining connecting fibers. In no case,connectivity was found between them after the cuts.
The pyramidal cell layer of area CA1 responded with apopulation EPSP after hilar stimulation. After cutting alongthe hippocampal fissure following the perforant path axisand ending close to the pyramidal cell layer in area
Ž .CA2–CA3 Fig. 2A, cut I , the population EPSP in CA1was reduced upon the same stimulation. After cutting area
Ž .CA3 Fig. 2A, cut II , which conveys the information fromthe DG to area CA1, these responses were totally abol-
Ž .ished Fig. 2B . Likewise, in response to stimulation ofarea CA1 in the alveus, the pyramidal cell layer, the

( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´ 309
stratum radiatum and the stratum lacunosum moleculare apositive field potential or a population spike could be
Ž .recorded in the granule cell layer of the DG Fig. 2C . Theamplitude of the field potential and the superimposedpopulation spike grew in amplitude and its latency wasshortened as the stimulating electrode was moved from thealveus towards stratum lacunosum-moleculare of area CA1.Again, carrying out cut I, responses in the DG to stimula-
Ž .tion of area CA1 were completely abolished Fig. 2C . Fig.2D shows that CA3 still responded with a populationantidromic spike and EPSP upon CA1 stimulation after cutI was done and hilar stimulation still evoked a populationEPSP and spike in area CA3.
3.2. Characteristics of responses eÕoked by hilar stimula-tion in the dentate gyrus: field potential recordings
A paired pulse stimulation protocol showed that both,the antidromic spike and the field synaptic potentials inDG evoked by hilar stimulation, over area CA4, presented
Ž .paired pulse inhibition Fig. 3A . A series of field potentialrecordings were also carried out in 10 acutely preparedslices from four juvenile rats of an age matching that of
Ž .the cultured slices 10- to 15-days old . In this case, hilarstimulation evoked antidromic population spikes in the DGthat were not followed by any further field potential
Ž .component Fig. 3C .To assess the nature of the PSP that followed the
antidromic spike in the cultured hippocampal slices, gluta-mate receptor antagonists were perfused. The field poten-tial response was drastically reduced by the AMPArkainate
Ž .receptor antagonist NBQX 10 mM and totally blocked bythe simultaneous perfusion of the NMDA receptor antago-
Ž . Ž . Ž . Žnist 2APV 30 mM and NBQX 10 mM ns10 Fig.. Ž .3B . Bicuculline 20 mM was also able to reduce the
Ž . Ž .amplitude of the field potential response ns6 Fig. 3B .In contrast to studies in acute slices from normal animalsbicuculline often induced repetitive discharges upon hilarstimulation. Although the PSP that followed the antidromicspike was completely blocked by the glutamate receptorantagonists, the partial suppressive effect of bicucullineand the strong inhibition shown by the paired pulse proto-col suggested that GABA mediated inhibition contributesA
to the field potential response.
3.3. Characteristics of responses to hilar stimulation in thedentate gyrus: intracellular recordings
To further explore the nature of the population PSP thatfollowed the antidromic spike in the DG after hilar stimu-lation, intracellular recordings were obtained from granulecells of the upper leaflet close to or in the joint of bothleaflets of the DG in 30 cultured slices. Granule cells wererecorded close to the border with stratum moleculare toavoid impaling hilar neurons or pyramidal-shaped neurons
w xat the border between the DG and the hilar region 39 .
Ž .The resting membrane potential RMP of the recordedŽ .cells was 64"5.8 mV mean"S.D. and the input resis-
Ž .tance was 68.9"34 MV ns45 cells . The mean dura-Žtion of the action potentials measured at half maximal
. Ž .amplitude was 2.5"0.9 ms ns25 cells . Without ex-ception, all cells displayed spontaneous firing of action
Žpotentials and frequent depolarizing PSPs mean frequency. Ž .3.4"3.1rs , which were often grouped bursts of 2 to 6 .
In no case spontaneous hyperpolarizing IPSPs were ob-Ž .served Fig. 4A1 .
Depolarizing current injection caused trains of actionpotentials with spike frequency adaptation as typical for
Ž .granule cells Fig. 4A2 . However, in four neurons a shortburst of action potentials followed by a pronounced hyper-polarization was induced by intracellular depolarizing cur-rent injection. These neurones were not further analyzed inthe present study since they were thought to be interneu-
w xrons 37 . Upon hilar stimulation and while observing withan extracellular microelectrode the presence of antidromicfield action potentials in the vicinity of the intracellularlyrecorded cells, all cells responded with an antidromic spikewith a hump in its repolarization phase or a hyperpolariza-
Ž .tion after the spike Fig. 4B1,B2 . After lowering thestimulus intensity, a compound EPSPrIPSP was observed,i.e., the threshold intensity for evoking an antidromicaction potential was higher than the intensity needed to
Žobtain a compound EPSPrIPSP for each cell Fig..4B1,B2 . The mean amplitudes of the EPSP and IPSP
Ž .were 2.8"1.6 and 4.0"2.5 mV, respectively ns15 .The latencies to peak were 3.8"1.4 ms for the EPSPs and
Ž .12.4"2.3 ms for the IPSPs ns15 . In seven cells, onlyEPSPs were observed upon subthreshold stimulation foreliciting antidromic spikes with a mean amplitude of 8.4"
Ž .4.7 mV Fig. 4B1 . The latency to the peak of these EPSPswas 5.8"2 ms.
During hyperpolarization of the cell, the hump of therepolarization phase of the action potential became morepronounced. Under control conditions hilar stimulation didnot elicit repetitive firing originating from the broadened
Ž .antidromic spike Fig. 4B . However, we noted in threecells that after an evoked EPSPrIPSP sequence, severalEPSPs and depending on stimulus intensity also EPSPs
Žsuperimposed by action potentials appeared Fig. 5C, Con-.trol .Hilar stimulation at threshold intensity for inducing
postsynaptic potentials during injection of hyper- and de-polarizing currents was used to determine the reversalpotential for the EPSP. The depolarizing component re-
Žversed polarity at y57.8"10.3 mV at RMPsy66"7.1.mV; ns12 .
Stimulation within the granule or molecular layers ofŽ . Žthe DG also evoked a compound EPSPrIPSP ns6 Fig.
.4B3 . Stimulation of the pyramidal cell layer of CA1 alsoelicited postsynaptic potentials alone or with spikes, de-
Ž .pending on the stimulus intensity, in granule cells ns5Ž .Fig. 4C1 . Conversely, hilar stimulation evoked EPSPs in

( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´310
Ž .Fig. 4. Intracellular recordings in the DG and CA1 in hippocampal cultured slices and synaptic responses to hilar stimulation. A1 Spontaneous activityŽ . Ž . Ž .presents grouped EPSPs and spikes in granule cells. A2 I–V curve of the same cell with an input resistance of 66 MV 13 DIV . B Characteristic
Ž . Ž . Ž .responses of granule cells to hilar and DG stimulation. B1 superimposed traces of the initial subthreshold EPSPs 5 V and orthodromic 7, 8 V andŽ . Ž .antidromic 9 V spikes evoked by increasing stimulation intensities in the hilar region in a slice at 6 DIV. B2 cell in which an IPSP was more evident
Ž . Ž . Ž . Ž . Ž .upon subthreshold 5 V stimulation.With higher intensities, a compound EPSPrIPSP 6.5 V and antidromic spike were evoked 7 V 13 DIV . B3Ž . Ž .compound EPSPrIPSP evoked by stimulation given within the DG, 250 mM apart from the recording electrode 14 DIV . C1 Typical intracellular
Ž .response in a granule cell upon stimulation of the pyramidal layer of CA1 under bicuculline perfusion; onset latency of the EPSP was 3.3 ms. C2 EPSPŽ .recorded in a pyramidal cell in CA1 upon hilar stimulation; onset latency of the EPSP was 3.2 ms 6 DIV . Vertical scale—20 mV—applies to all traces.
pyramidal cells of area CA1 at low intensity stimulationŽ .ns3; Fig. 4C2 .
3.4. Effects of bicuculline on postsynaptic potentials indentate gyrus granule cells induced by hilar stimulation.
Since GABA dependent inhibition could obscure theA
presence of EPSPs, we carried out recordings of granuleŽ .cells during perfusion of bicuculline 20 mM . Bicuculline
partially blocked the spontaneous synaptic activity andŽ .spontaneous paroxysmal activity appeared Fig. 5A . This
activity was generally rhythmic at 0.3"0.4rs with depo-Ž .larization shifts of long duration 2.1"1.8 s; ns7 super-
imposed by high frequency neuronal discharges. Sub-threshold hilar stimulation for evoking an antidromic spike
Ž .induced an EPSP Fig. 5B,C . The determination of the
reversal potential of the pharmacologically isolated EPSPwas not possible due to firing of the cells; however, it was
Žnoticed that it was shifted in depolarizing direction Fig..5A, Bicuculline . When threshold stimulation for evoking
antidromic spikes was provided, repetitive spiking wasŽ .always induced Fig. 5C .
The pharmacologically isolated EPSP was slightly re-duced and shortened in the presence of the NMDA recep-tor antagonist 2APV and completely blocked by perfusion
Žof the AMPArkainate receptor antagonist NBQX Fig..5C . In the presence of the glutamate receptor antagonists,
the antidromic spikes could still be evoked while both theevoked IPSPs and EPSPs and spontaneous synaptic poten-
Ž . Ž .tials were completely blocked Fig. 5C,D ns8 .Since the stimulation of CA1 was able to produce
postsynaptic potentials in granule cells, the spontaneous
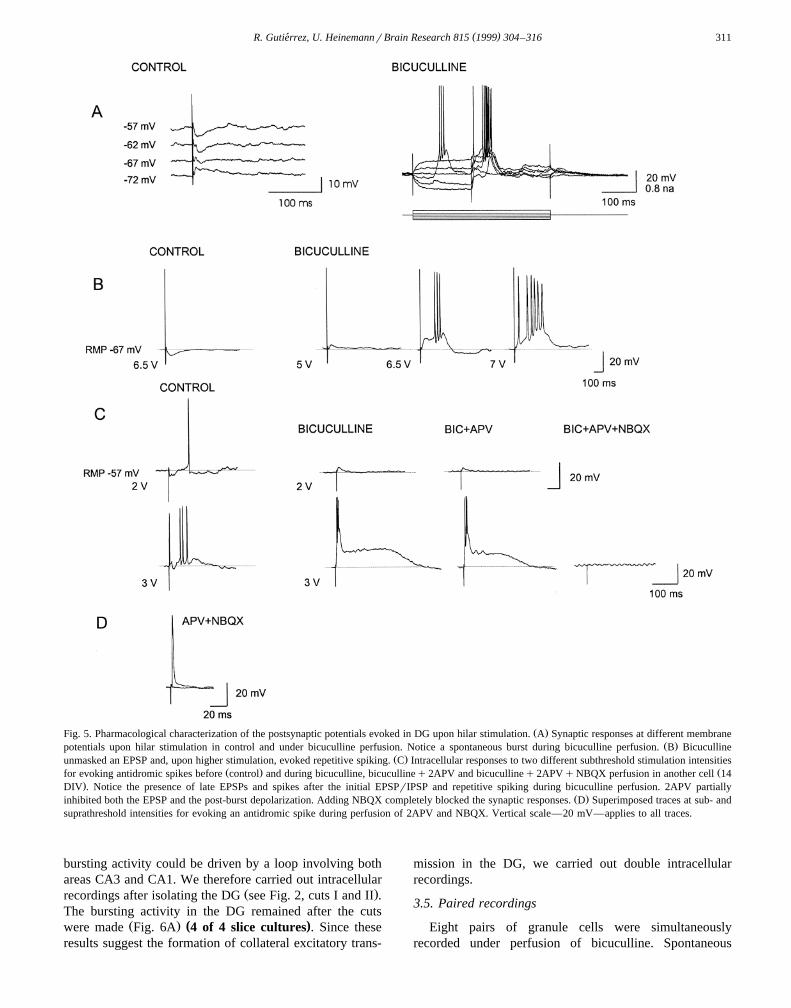
( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´ 311
Ž .Fig. 5. Pharmacological characterization of the postsynaptic potentials evoked in DG upon hilar stimulation. A Synaptic responses at different membraneŽ .potentials upon hilar stimulation in control and under bicuculline perfusion. Notice a spontaneous burst during bicuculline perfusion. B Bicuculline
Ž .unmasked an EPSP and, upon higher stimulation, evoked repetitive spiking. C Intracellular responses to two different subthreshold stimulation intensitiesŽ . Žfor evoking antidromic spikes before control and during bicuculline, bicucullineq2APV and bicucullineq2APVqNBQX perfusion in another cell 14
.DIV . Notice the presence of late EPSPs and spikes after the initial EPSPrIPSP and repetitive spiking during bicuculline perfusion. 2APV partiallyŽ .inhibited both the EPSP and the post-burst depolarization. Adding NBQX completely blocked the synaptic responses. D Superimposed traces at sub- and
suprathreshold intensities for evoking an antidromic spike during perfusion of 2APV and NBQX. Vertical scale—20 mV—applies to all traces.
bursting activity could be driven by a loop involving bothareas CA3 and CA1. We therefore carried out intracellular
Ž .recordings after isolating the DG see Fig. 2, cuts I and II .The bursting activity in the DG remained after the cuts
Ž . ( )were made Fig. 6A 4 of 4 slice cultures . Since theseresults suggest the formation of collateral excitatory trans-
mission in the DG, we carried out double intracellularrecordings.
3.5. Paired recordings
Eight pairs of granule cells were simultaneouslyrecorded under perfusion of bicuculline. Spontaneous

( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´312
Ž . Ž .Fig. 6. A Spontaneous activity of a granule cell in an isolated DG under bicuculline perfusion. Notice the presence of depolarization shifts. BSimultaneous recordings of two pairs of granule cells under bicuculline perfusion. Notice synchronized spontaneous bursting activity. Left panel depicts a
Ž . Ž .long-lasting burst. C Spontaneous activity of a pair of cells simultaneously recorded showing the presence of EPSPs in a cell upper trace as the otherŽ . Ž .cell fires action potentials lower trace . D Paired recordings in DG showed that intracellularly evoked spiking of a cell evokes EPSPs in the follower cell
during perfusion of bicuculline. A single presynaptic action potential is depicted, whereas three superimposed traces of the follower cell were acquiredusing the spike as trigger for the analysis.
bursting activity was present in all the recordings andŽ .appeared to be synchronized in all the pairs Fig. 6B .
Evidence for direct excitatory synaptic transmission be-tween granule cells was suggested by the presence of
ŽEPSPs in one of the cells while the other fired ns3.pairs . Fig. 6B and C shows examples of intracellular
recordings of two different pairs of granule cells, in whichspontaneous activity under bicuculline perfusion shows astrong synchronization and EPSPs could be seen after
spiking of one cell. In Fig. 6D, another pair is depicted, inwhich intracellular stimulation to make a cell fire consis-tently evoked EPSPs in the other cell.
3.6. Dextranamine staining
Four slices at 7–15 DIV were reincubated for 12 h afterplacing under microscopical observation tetramethyl–
Ž .rhodamine–dextranamine Molecular Probes and fluores-

( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´ 313
Ž .Fig. 7. Tetramethyl–rhodamine–dextranamine stained fibers originated from CA1 pyramidal cells project to the granule cell layer of the DG 7–8 DIV .Granule cells, stained with fluorescein–dextranamine, are depicted in the upper blade of the DG. The arrow on the right panel shows the same granule cellpointed on the left panel at a higher magnification.
Ž .cein–dextranamine Molecular Probes over the granulecell layer of the DG and over the pyramidal cell layer of
Ž .CA1, respectively Fig. 7 . Slices were then fixed andphotographed under a fluorescence microscope with properfilters. This confirmed the presence of fibers extendingfrom area CA1 over the upper blade of the DG to granulecells.
4. Discussion
Our data show that explanted cultures of hippocampalslices preserve the intrinsic connectivity from the DG toarea CA3 and from area CA3 to CA1. However, a strongsynaptic reorganization is suggested by the following find-
Ž .ings: i stimulation experiments and dextranamine stain-ing revealed mutual interactions between area CA1 and the
Ž .DG. ii Hilar stimulation induced PSPs below and abovethreshold for activation of antidromic potentials in DG.Ž .iii Intracellular stimulation of granule cells induced EP-SPs in a neighboring neuron. Interestingly, under bicu-culline, spontaneous and evoked epileptiform dischargescould be observed in these cells.
In cultured slices, the DG is totally deprived fromafferent input, while areas CA3 and CA1 are just partiallydeprived since they loose their normal interaction partners
w xin other areas 51 . Therefore, only synaptic contacts ofintrinsic origin are present under culture conditions. It wastherefore rather suggestive that the PSPs following an-
tidromic stimulation represent EPSPs mediated by recur-rent excitatory collaterals formed by MF. Two findingssuggest that on top of such reorganization recurrent excita-tory connections are formed between granule cells. Afterisolating the DG from areas CA3 and CA1 spontaneousepileptiform discharges were still seen in granule cellsafter application of the GABA receptor antagonist bicu-A
culline. We should bear in mind, however, that in theisolated DG, part of area CA3–4 and the hilus wereattached. Moreover, paired recordings in such conditionsrevealed a high synchrony between the synaptic events inthe recorded granule cells and intracellular induction ofaction potentials in a cell caused postsynaptic potentials ina second cell. This confirms previous suggestions on mossy
w xfibre spouting causing excitatory synaptic inputs 13,51,52 .The possibility, however, that this high synchrony mightbe also driven by an enhanced number of electrical synapsesis open to future investigations.
Reciprocal projections between the DG and CA1 do notw xnormally exist 24,33 . However, morphological studies
have shown projections from CA1 towards the DG andw xvice versa after denervation 23 , and after partial destruc-
w xtion of area CA3 8,9 . In culture conditions, fibers fromgranule cells have been shown to extend to the granular
w xlayer of the DG and, although rarely, to CA1 5 . Wereport here, projections from area CA1 extending to themolecular and even to the granular layer of the DG.Indeed, our electrophysiological characterization of theseconnections shows that functional synapses are formedbetween these two areas since stimulation of the MF and

( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´314
the DG produces synaptic responses in CA1 and viceversa, even after disruption of area CA3 which normallyconveys this information.
The formation of these aberrant connections does notdepend on the presence of sustained hyperexcitability thatcould be evoked by the presence of penicillin in the culturemedium, since avoiding all electrical activity by culturingthe slices under TTX for 5 days from the day of explanta-tion did not prevent their generation.
Indeed, these apparent direct connections between CA1and DG explain in part why stimulation of the molecularlayer of the DG induces EPSPrIPSP sequences in granulecells. A second putative source for the induction of EPSPsin DG after hilar stimulation is suggested by experiments
w xof Muller and Misgeld 28 . These authors observed EPSPs¨in DG interneurons after stimulation of CA3 and sponta-neous EPSPs after application of 4AP. Similar observa-
w xtions have been also made by Scharfman 38 . This sug-gests that an excitatory projection from area CA3 to theDG exists and this may have proliferated into the deaffer-ented DG. This may explain why stimulation with lowintensities of the hilus induced EPSPrIPSP sequences inthe cultured DG. The last input source to DG could be the
w xdirect inhibitory CA1–CA3-hilar connections 41 . How-ever, under bicuculline perfusion, the possible input fromthese interneurons is blocked. Our recordings of juvenilehippocampi were done without any drug treatment and wefound no evidence for this sort of activity. This mightsuggest that, if these connections also exist in the juvenileanimal, they do not go further from the hilar region to theDG.
That an extended collateral reinnervation of granulecells of the DG is formed in culture is also suggested bythe finding that stimulation of the hilus produces in fieldpotential recordings an antidromic population spike andsynaptic field potentials in the DG. These responses are
Žnot likely to be due to direct stimulation by current.spread of afferent fibers in the stratum moleculare of DG,
since they were evoked with rather low current intensitiesŽ .0.1–0.3 mA at the most during 0.1 ms applied in thehilar region at least 100 mm apart. Therefore, filtering ofthe current is expected by neuronal and glial capacitances.Moreover, hilar stimulation evoked spikes with a latencyto the stimulation artifact as well as EPSPs upon sub-threshold stimulation. Our recordings in acutely preparedjuvenile hippocampi show no evidence of synapticallyevoked population PSPs, where only antidromic populationspikes were seen. Intracellular recordings in the culturedDG revealed excitatory and inhibitory synaptic potentialsevoked by hilar stimulation. When GABA -mediated inhi-A
bition was suppressed the EPSP increased in amplitudeand the reversal potential of the EPSP was shifted indepolarizing direction. This suggests that synaptic inhibi-tion is clearly expressed as also suggested by our pairedpulse recordings. The isolated EPSP or repetitive spikinginduced by hilar stimulation after application of bicu-
culline was completely blocked by the application of 2APVand NBQX suggesting that these are caused by glutamate.
Our findings compare with previous studies in whichMF sprouting was induced by lesioning area CA3 by
w xkainate. Sloviter 43 , showed that paired pulse inhibitionw xin the DG is impaired and, Cronin et al. 10 showed that
hilar stimulation was able to produce EPSPs in the pres-ence of bicuculline also in the DG. These observationssuggested the presence of recurrent excitatory synapses,but they were revealed only after GABA -dependent inhi-A
bition was blocked. Moreover, denervation of granule cellsin vivo induces MF sprouting into the molecular layerw x3,14,23 .
Although the cultured slices seem to retain guidancew xcues for the growth of the MF 11 , they form however
new synapses with granule cells, with interneurons in theDG and possibly also with cells in area CA1. It is knownthat granule cells are still undergoing massive proliferationat P7 and continue forming synapses primarily during the
w x w xpostnatal period 40 . Gaiarsa and Heimrich 18 , reportedthat DG-hilus explants develop MF axons projecting toarea CA3 in cocultured dislocated hippocampal slices.However, in addition to the normal giant MF terminals,they found numerous small fibers which did not maintainthe restricted distribution of the MF to CA3.
Most features present in the recordings from granulecells under culture conditions are similar to those obtainedfrom acutely prepared slices. The intrinsic membrane prop-erties of the cells, i.e., spike duration, spike frequencyadaptation, input membrane resistance and pharmacology
w xare well in agreement with previous reports 22,37,38,45 .We observed in the cultured granule cells spontaneousspiking and depolarizing synaptic activity which are rare inrecordings with sharp microlectrodes in acute slices; alsono spontaneous hyperpolarizing IPSPs were observed. Mostof the spontaneous PSPs however were blocked by perfu-sion of bicuculline suggesting that these represent depolar-
w xizing IPSPs, as has often been noted in granule cells 31 .The small EPSP together with a reversal potential ratherclose to resting membrane potential suggest that in normalconditions, this EPSP is strongly controlled by the shunt-
w xing action of a GABA mediated superimposed IPSP 44 .A
4.1. Pathophysiological implications
Our experiments with bicuculline show that in slicecultures there are epileptiform discharges induced not onlyin areas CA3 and CA1 which is rather normal in slices
w xfrom young animals 47 , but also in the DG. Interestingly,in intracellular recordings such spontaneous depolarizingshifts remained after isolation of the DG from areas CA1and CA3, suggesting that these connections are not essen-tial for the formation of spontaneous epileptiform dis-charges in the present preparation. This puts emphasis onthe formation of recurrent excitatory collaterals from MF
Ž w x.to neighboring granule cells see also Ref. 50 .

( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´ 315
An aberrant MF rearrangement has been associated withw xmedically intractable temporal lobe epilepsy 2,21 , and it
is proposed that the affected circuitry could contribute tofocal seizure onset. However, these functional recurrent
w xexcitatory connections are tonically inhibited 10 . This isin accordance with the hypothesis that MF sprouting in-volves new granule cell synapses to basket cells as well as
Ž w x.to other granule cells see Ref. 12,42 . Moreover, ourresults also show that a possible loop comprising CA1–DGinteractions might also be involved in the generation of ahyperexcitable state of granule cells. Whether formation ofrecurrent axon collaterals is a necessary prerequisite forthe generation of drug resistant temporal lobe epilepsy isyet unknown. Our findings suggest that the formation ofsuch abnormal synapses facilitates the development ofepileptiform discharges in the DG and that it may be animportant step in generation of drug resistant temporallobe epilepsy.
Acknowledgements
The authors thank Richard Kovacs for providing the´figure of fluorescence staining and Drs. W. Muller, T.¨Gloveli and A. Draguhn for their comments on earlierversions of the manuscript. Dr. Rafael Gutierrez was a´recipient of an Alexander von Humboldt Research Fellow-ship. This research was supported by Grant SFB 515, TPB1.
References
w x1 P. Andersen, T.V.P. Bliss, K.K. Skrede, Lamellar organization ofŽ .the hippocampal excitatory pathways, Exp. Brain Res. 13 1971
222–238.w x2 T.L. Babb, W.R. Kupfer, J.K. Pretorius, P.H. Crandall, M.F.
Levesque, Synaptic reorganization by mossy fibers in human epilep-Ž .tic fascia dentata, Neuroscience 42 1991 351–363.
w x3 J.W. Bekenstein, E.W. Lothman, The role of the perforant pathwayas a trophic factor for neurotransmission in the rat dentate gyrus,
Ž .Hippocampus 1 1991 405–413.w x4 Y. Ben-Ari, E. Tremblay, M. Berger, L. Nitecka, Kainic acid seizure
Ž .syndrome and binding sites in developing rats, Brain Res. 14 1984284–288.
w x5 M. Caeser, A. Aertsen, Morphological organization of rat hippocam-Ž .pal slice cultures, J. Comp. Neurol. 307 1991 87–106.
w x6 J.E. Cavazos, T.P. Sutula, Progressive neuronal loss induced bykindling: a possible mechanism for mossy fiber synaptic reorganiza-
Ž .tion and hippocampal sclerosis, Brain Res. 527 1990 1–6.w x7 B.J. Clairborne, D.G. Amaral, W.M. Cowan, A light and electron
microscopic analysis of the mossy fibers of the rat dentate gyrus, J.Ž .Comp. Neurol. 246 1986 435–458.
w x8 T.M. Cook, K.A. Crutcher, Extensive target loss during developmentŽ .results in mossy fibers in the region superior CA1 of the rat
Ž .hippocampal formation, Dev. Brain Res. 21 1985 19–30.w x9 T.M. Cook, K.A. Crutcher, Lesion-induced CA1 mossy fibers in the
Ž .rat represent a neoinnervation, Exp. Brain Res. 70 1988 433–436.w x10 J. Cronin, A. Obenaus, C.R. Houser, F.E. Dudek, Electrophysiology
of dentate granule cells after kainate-induced synaptic reorganizationŽ .of the mossy fibers, Brain Res. 573 1992 305–310.
w x11 M.E. Dailey, J. Buchanan, D.E. Bergles, S.J. Smith, Mossy fibergrowth and synaptogenesis in rat hippocampal slices in vitro, J.
Ž .Neurosci. 14 1994 1060–1078.w x12 F.E. Dudek, A. Obenaus, J.S. Schweitzer, J.P. Wuarin, Functional
significance of hippocampal plasticity in epileptic brain: electrophys-iological changes of the dentate granule cells associated with mossy
Ž .fiber sprouting, Hippocampus 4 1994 259–265.w x13 M. Frotscher, B.H. Gahwiler, Synaptic organization of intra-¨
cellularly stained CA3 pyramidal neurons in slice cultures of ratŽ .hippocampus, Neuroscience 24 1988 541–551.
w x14 M. Frotscher, J. Zimmer, Lesion-induced mossy fibers to the molec-ular layer of the rat fascia dentata: identification of postsynapticgranule cells by the Golgi-EM technique, J. Comp. Neurol. 215Ž .1983 298–311.
w x15 B.H. Gahwiler, Organotypic monolayer cultures of nervous tissue, J.¨Ž .Neurosci. Methods 4 1981 329–342.
w x16 B.H. Gahwiler, Development of the hippocampus in vitro: cell types,¨Ž .synapses, and receptors, Neuroscience 11 1984 751–760.
w x17 B.H. Gahwiler, Slice cultures of cerebellar, hippocampal and hy-¨Ž .pothalamic tissue, Experientia 40 1984 235–243.
w x18 J.L. Gaiarsa, B. Heimrich, Restoration of mossy fiber projection inslice co-cultures of dislocated dentate gyrus and degranulated hip-
Ž .pocampus, Dev. Brain Rev. 86 1995 250–258.w x19 R. Gutierrez, U. Heinemann, Synaptic reorganization of explanted´
Ž .cultures of the rat hippocampus, Soc. Neurosci. 22 1996 Abstr.823.19.
w x20 U. Heinemann, H. Beck, J.P. Dreier, E. Ficker, J. Stabel, C.L.Zhang, The dentate gyrus as a regulated gate for the propagation of
Ž .epileptiform activity, in: C.E. Riback, C.M. Gall, I. Mody Eds. ,The Dentate Gyrus and its Role in Seizures, Elsevier, Amsterdam,1992, pp. 273–280.
w x21 M. Isokawa, M.F. Levesque, T.L. Babb, J. Engel, Single mossy fiberaxonal systems of human dentate granule cells studied in hippocam-pal slices from patients with temporal lobe epilepsy, J. Neurosci. 13Ž .1993 1511–1522.
w x22 J.D.C. Lambert, R.S.G. Jones, A reevaluation of excitatory aminoacid-mediated synaptic transmission in rat dentate gyrus, J. Neuro-
Ž .physiol. 64 1990 119–132.w x23 S. Laurberg, J. Zimmer, Lesion-induced sprouting of hippocampal
mossy fiber collaterals to the fascia dentata in developing and adultŽ .rats, J. Comp. Neurol. 200 1981 433–459.
w x24 R. Lorente de No, Studies on the structure of the cerebral cortex: II.´Continuation of the study of the Ammonic system, J. Psychol.
Ž .Neurol. 46 1934 113–177.w x25 L. Mello, E.A. Cavalheiro, T.L. Babb, W.R. Kupfer, J.K. Pretorius,
A.M. Tan, D.M. Finch, Pilocarpine-induced status epilepticus leadsto chronic seizures and mossy fiber sprouting in the hippocampus,
Ž .Epilepsia 31 1990 675.w x26 R. Miles, R.K.S. Wong, Excitatory synaptic interactions between
Ž .CA3 neurons in the guinea-pig hippocampus, J. Physiol. LondonŽ .373 1986 371–373.
w x27 U. Misgeld, M. Frotscher, Postsynaptic GABAergic inhibition ofnon-pyramidal neurons in the guinea-pig hippocampus, Neuro-
Ž .science 19 1986 193–206.w x28 W. Muller, U. Misgeld, Picrotoxin- and 4-aminopyridine-induced¨
activity in hilar neurons in the guinea pig hippocampal slice, J.Ž .Neurophysiol. 65 1991 141–147.
w x29 J.V. Nadler, B.C. Perry, C.W. Cotman, Selective reinnervation ofhippocampal area CA1 and the fascia dentata after destruction of
Ž .CA3–CA4 afferents with kainic acid, Brain Res. 182 1980 1–9.w x30 M.M. Okasaki, D.A. Evenson, J.V. Nadler, Hippocampal mossy
fiber sprouting and synapse formation after status epilepticus in rats:visualization after retrograde transport of biocytin, J. Comp. Neurol.
Ž .352 1995 515–534.w x31 T.S. Otis, K.J. Staley, I. Mody, Perpetual inhibitory activity in

( )R. Gutierrez, U. HeinemannrBrain Research 815 1999 304–316´316
mammalian brain slices generated by spontaneous GABA release,Ž .Brain Res. 545 1991 142–150.
w x32 X. Qiao, J.L. Noebels, Developmental analysis of hippocampalmossy fiber outgrowth in a mutant mouse with inherited spike-wave
Ž .seizures, J. Neurosci. 13 1993 4622–4635.w x33 S. Ramon y Cajal, Histologie du Systeme Nerveux de l’Homme et´ `
des Vertebres, Maloine, Paris, 1911.´ ´w x34 G. Rausche, J. Dreier, C.L. Zhang, U. Heinemann, Prevention of
spread of seizure-like activity from the entorhinal cortex to theŽ .hippocampus, in: E.J. Speckmann, M.J. Gutnick Eds. , Epilepsy
and Inhibition, Urban and Schwarzenberg, Munster, 1990, pp. 297–¨312.
w x35 A. Represa, I. Jorquera, G. Le Gal La Salle, Y. Ben-Ari, Epilepsyinduced collateral sprouting of hippocampal mossy fibers: does itinduce the development of ectopic synapses with granule cell den-
Ž .drites?, Hippocampus 3 1993 257–268.w x36 A. Represa, H. Pollard, J. Moreau, G. Ghilini, M. Krestchatisky, Y.
Ben-Ari, Mossy fiber sprouting in epileptic rats is associated with atransient increased expression of a-tubulin, Neurosci. Lett. 156Ž .1993 149–152.
w x37 H.E. Scharfman, Differentiation of the rat dentate neurons by mor-phology and electrophysiology in hippocampal slices: granule cells,spiny hilar cells and aspiny ‘fast-spiking’ cells, in: C.E. Riback,
Ž .C.M. Gall, I. Mody Eds. , The Dentate Gyrus and its Role inSeizures, Elsevier, Amsterdam, 1992, pp. 93–112.
w x38 H.E. Scharfman, EPSPs of dentate gyrus granule cells during epilep-tiform bursts of dentate hilar ‘mossy’ cells and area CA3 pyramidal
Ž .cells in disinhibited rat hippocampal slices, J. Neurosci. 14 19946041–6057.
w x39 H.E. Scharfman, Electrophysiological diversity of pyramidal-shapedneurons at the granule cell layerrhilus border of the rat dentate
Ž .gyrus recorded in vitro, Hippocampus 5 1995 287–305.w x40 A.R. Schlessinger, W.M. Cowan, D.I. Gottlieb, An autoradiographic
study of the time of origin and the pattern of granule cell migrationŽ .in the dentate gyrus of the rat, J. Comp. Neurol. 159 1975
149–176.w x41 A. Sik, A. Ylinen, M. Penttonen, G. Buzsaki, Inhibitory CA1–´
Ž .CA3-hilar region feedback in the hippocampus, Science 265 19941722–1724.
w x42 R.S. Sloviter, Permanently altered hippocampal structure, excitabil-ity, and inhibition after experimental status epilepticus in the rat: the‘dormant basket cell’ hypothesis and its possible relevance to tempo-
Ž .ral lobe epilepsy, Hippocampus 1 1991 41–66.w x43 R.S. Sloviter, Possible functional consequences of synaptic reorgani-
zation in the dentate gyrus of kainate-treated rats, Neurosci. Lett.Ž .137 1992 91–96.
w x44 K.J. Staley, I. Mody, Shunting of excitatory input to dentate gyrusgranule cells by a depolarizing GABA receptor-mediated post-A
Ž .synaptic conductance, J. Neurophysiol. 68 1992 197–212.w x45 K.J. Staley, T. Otis, I. Mody, Membrane properties of dentate gyrus
granule cells: comparison of sharp microelectrode and whole-cellŽ .recordings, J. Neurophysiol. 67 1992 1346–1358.
w x46 L. Stoppini, P.A. Buch, D. Muller, A simple method for organotypic¨Ž .cultures of nervous tissue, J. Neurosci. Methods 37 1991 173–182.
w x47 J.W. Swann, K.L. Smith, R.J. Brady, Localized excitatory synapticinteractions mediate the sustained depolarization of electrographic
Ž .seizures in developing hippocampus, J. Neurosci. 13 1993 4680–4689.
w x48 D.L. Tauck, J.V. Nadler, Evidence of functional mossy fiber sprout-ing in hippocampal formation of kainic acid-treated rats, J. Neurosci.Ž .5 1985 1016–1022.
w x49 D.M. Terrian, R.L. Gannon, M.A. Rea, Glutamate is endogenousamino acid selectively released by rat hippocampal mossy fibersynaptosomes concomitantly with prodynorphin-derived peptides,
Ž .Neurochem. Res. 15 1990 1–5.w x50 J.-P. Wuarin, F.E. Dudek, Electrographic seizures and new recurrent
excitatory circuits in the dentate gyrus of hippocampal slices fromŽ .kainate-treated epileptic rats, J. Neurosci. 14 1996 4438–4448.
w x51 J. Zimmer, B.H. Gahwiler, Cellular and connective organization of¨slice cultures of the rat hippocampus and fascia dentata, J. Comp.
Ž .Neurol. 228 1984 432–446.w x52 J. Zimmer, B.H. Gahwiler, Growth of hippocampal mossy fibers: a¨
lesion and coculture study of organotypic slice cultures, J. Comp.Ž .Neurol. 264 1987 1–313.