supporting online material for -...
TRANSCRIPT

www.sciencemag.org/cgi/content/full/333/6044/883/DC1
Supporting Online Material for
The Structure of the Kinesin-1 Motor-Tail Complex Reveals the Mechanism of Autoinhibition
Hung Yi Kristal Kaan, David D. Hackney, Frank Kozielski*
*To whom correspondence should be addressed. E-mail: [email protected]
Published 12 August 2011, Science 333, 883 (2011) DOI: 10.1126/science.1204824
This PDF file includes
Materials and Methods SOM Text Figs. S1 to S6 Tables S1 and S2 References

Materials and Methods
Protein expression and purification
The motor domain of Drosophila Melanogaster kinesin (residues 1-381) was initially cloned into the vector pET21(16). Polymerase chain reaction (PCR) was carried out to amplify residues 1-365 and to include restriction sites BamHI and XhoI. The product was inserted into a modified pET28a expression vector (Invitrogen), yielding a Ulp-1 protease-cleavable N-terminal SUMO fusion and hexahistidine-tagged protein. It was then expressed and purified as previously described(17), apart from the following changes: the plasmid was transformed into E. coli BL21-CodonPlus (Stratagene), Buffer A contained 20 mM Tris pH 8.8, 1 mM MgCl2, 200 mM NaCl, 20 mM Imidazole, 1 mM EGTA, protease buffer contained 20 mM Tris pH 8.8 and 250 mM NaCl, and gel filtration buffer contained 20 mM Tris pH 8.8, 300 mM NaCl, 1 mM DTT, 1 mM EGTA, and 2 mM MgCl2.
The non-fusion tail peptide (NF937-952) was obtained by initial expression as a
fusion with thioredoxin followed by removal of the thioredoxin by cleavage with Tev protease. NF937-952 also contains an additional GlySer and ThrSer appended to the N- and C-termini respectively. Ser 181 was replaced with cysteine in DKH392(CLM) that is a cysteine-lite mutant (CLM) of Drosophila DKH392(18) with the two surface exposed cysteines 45 and 338 replaced by serines and containing a C-terminal His6-tag. The H136E mutation was introduced into wild type DKH392 with a C-terminal His6-tag. The D185R mutation was introduced into DKH392(CLM) with an additional S156C substitution. All constructs were confirmed by DNA sequencing and were purified by Ni-affinity chromatography, essentially as previously described(19).
Crystallization of dimer and dimer-tail complex
Crystals of the motor domain dimer appeared after 2 days in hanging drops by mixing 1 µl of protein with 1 µl of reservoir solution containing 24 % polyethylene glycol-3350, 0.15 M Ammonium sulfate, and 0.1 M HEPES sodium pH 7.8 in VDX plates (Hampton Research) at 19°C. Micro streak seeding was carried out to produce single and larger crystals. A block-shaped crystal with dimensions of approximately 0.1 x 0.1 x 0.1 mm was immersed in cryoprotectant solution (28.8 % polyethylene glycol-3350, 0.18 M Ammonium sulfate, 0.12 M HEPES sodium pH 7.8, 0.36 M NaCl, and 20% erythritol) and flash frozen in liquid nitrogen.
Intially, we crystallized the motor domain dimer in complex with tail peptide 910-
952. However, the crystals diffracted no better than 3 Å. We were able to solve the structure and saw electron density for only ten residues of the peptide. Hence, we decided to use a shorter tail peptide for crystallization. Purified motor domain (10 mg/ml) was

2
incubated with tail peptide 937-952 (6.1 mg/ml) in a ratio of (1 monomeric motor : 2 tails) for 2 hours on ice before setting up of crystal trays. Crystals of the motor domain dimer-tail complex appeared after 2 days in hanging drops by mixing 1 µl of protein-inhibitor complex with 1 µl of reservoir solution containing 18 % polyethylene glycol-3350, 0.2 M Potassium chloride, and 0.1 M HEPES sodium pH 7.4 in VDX plates (Hampton Research) at 19°C. Micro streak seeding was carried out to produce single and larger crystals. A block-shaped crystal with dimensions of approximately 0.3 x 0.1 x 0.1 mm was immersed in cryoprotectant solution (21.6 % polyethylene glycol-3350, 0.24 M Potassium chloride, 0.12 M HEPES sodium pH 7.4, 0.36 M NaCl, and 20% polyethylene glycol-200) and flash frozen in liquid nitrogen.
Data collection and processing
Diffraction data for the dimer and dimer-tail complex were recorded at the European Synchrotron Radiation Facility (ESRF) ID23-2 and Swiss Light Source (SLS) PXI respectively. Data were processed using iMosflm(20) and scaled using Scala(21) from the CCP4 suite of programs(22). The dimer and dimer-tail structures were solved by molecular replacement (Phaser(23)) using one molecule of human kinesin-1 motor domain(24) (PDB code: 1BG2) and one molecule of the refined dimer structure as search models respectively. Refinement was carried out with Refmac5(25). The calculation of Rfree used 5% of data. Electron-density and difference density maps all σA-weighted, were inspected and the model was improved using Coot(26). Model geometry was analysed using Molprobity(27). For the dimer structure, 90.31 % (587) of the residues are in the preferred regions, 6.77 % (44) are in the allowed regions, and 2.92 % (19) are outliers as shown by the Ramachandran Plot. For the dimer-tail complex, 94.14 % (1270) of the residues are in the preferred regions, 4.97 % (67) are in the allowed regions, and 0.89 % (12) are outliers. Protein domain interfaces were analyzed using PDBePISA(28). Figures are prepared using PyMOL(29).
Cross-linking
Disulfide cross-linked DKH392(CLM)-S181C was prepared by oxidation using copper and phenanthroline(30). DKH392(CLM)-S181C was pre-incubated with 4 mM DTT to keep Cys181 reduced. The DTT was then removed by precipitation with 60 % ammonium sulfate followed by two washes of the pellet by suspension in 60 % ammonium sulfate and centrifugation. The washed pellet was dissolved in 20 mM MOPS/KOH pH 7.0, 200 mM NaCl, 2 mM MgCl2 and 0.1 mM ATP. Oxidation was initiated by addition of 20 µM CuCl2 and 40 µM phenanthroline. After 1 hour at 25°C, the reaction was stopped by addition of excess EDTA. Control experiments indicated that MT-stimulated ATPase activity and exposed SH groups (detected with 5,5’-dithiobis-2-nitrobenzoic acid (DTNB)) were lost approximately in parallel. The extent of cross linking was monitored by SDS PAGE using SDS sample buffer that did not contain

3
sulphydryl compounds and that contained N-ethylmaleimide to block the buried SH groups on the protein and prevent disulfide exchange after denaturation. Typically 10-15 % of the protein was resistant to cross linking even with higher concentrations of copper and longer incubations. This likely represents protein in which one or both Cys181 side chains of a dimer have been chemically modified to a form that cannot be oxidized to a disulfide.
Kinetics of MT-stimulated mant-ADP dissociation
The bound ADP of DKH392(CLM)-S181C was first exchanged for mant-ADP by equilibration with a 5-fold excess of mant-ATP and 4 mM DTT and then subjected to centrifuge gel filtration on a column of Sephadex G25 equilibrated with 15 % sucrose in 200 mM NaCl, 2 mM MgCl2 and 20 µM mant-ATP before oxidation as described above. Cross-linking was reversed by treatment with 10 mM DTT for 24 hours at 21°C . Release of mant-ADP was determined as previously described(31) in A25 buffer with 50 mM KCl and 7.5 % sucrose (after mixing). Excitation was at 285 nm and fluorescence was detected using a 417 nm long pass filter. Reactions containing MTs were supplemented with 10 µM paclitaxel. Offsets were applied to the fluorescence data with microtubules, in order to correct for the fluorescence of the microtubule tryptophans, so that the traces with and without microtubules start at the same values.

4
SOM Text
Structure of kinesin motor domain dimer
The crystal structure of D. melanogaster kinesin motor domain dimer was solved at a resolution of 2.7 Å. Data collection and refinement statistics are shown in table S1. The final model comprises of residues Glu 10 to Lys 356 (excluding residues 246 to 258) and Glu 10 to Lys 354 (excluding residues 244 to 259) for molecules A and B respectively. The excluded region is Loop L11, which is disordered and not modelled. Residues 10 to 13, which are implicated in forming the cover-neck bundle, is observed stabilizing the docking of the neck linker via the formation of a ß-sheet(32, 33). The asymmetric unit consists of a dimer, where the monomers are not related by crystallographic symmetry. Each monomer has one molecule of Mg2+ADP bound in the nucleotide-binding site.
The motor domains of the dimer have the same fold as other kinesin homologues:
eight major β-strands in the core, surrounded by three major α-helices on each side. Overall, the two monomers are related by a non-crystallographic two-fold symmetry (about 180°) and interact mainly at the coiled-coil domain via the neck-linker (Fig. S1a). The two monomers of our kinesin dimer overlay very well in the catalytic core up to the neck linker, but differ slightly in the position of the coiled-coil domain (r.m.s.d. = 0.84 Å) (Fig. S1c). The kinesin dimer has only four residues on the catalytic core that contribute to the interfacial surface with the coiled-coil domain: Asn 167 and Arg 168 of molecule B and Lys 166 of molecules A and B (Fig. S1a).

5
Fig. S1. Structure of motor domain dimer. (A) D. melanogaster kinesin motor domain dimer with Mg2+ADP (red, sticks) bound in the nucleotide binding pocket. Ser 181 (yellow, spheres), which was modified to cysteine for cross-linking experiments, and catalytic core residues (green, spheres) interacting with the coiled-coil domain are shown. (B) Structure of R. norvegicus kinesin motor domain dimer, where monomers exhibit a 120° rotational symmetry (PDB ID: 3KIN). Several structures of the minus-end directed kinesin Ncd dimer(34-36) reveal different angles of rotation between the monomers (ranging from 75° to 180°); thus, alluding to the fact that the kinesin dimers have inherent flexibility and variability to adopt various conformations for motility. (C) Overlay of monomers (chain A: orange, B: blue) of dimer structure revealing differing position of the α-helical regions forming the coiled-coil domain.

6
Fig. S2. Comparison of the free motor domain dimer and the dimer-tail complex. Overlay of monomers of the dimer structure (blue) with that of the dimer-tail complex (magenta) reveals virtually similar conformations of the catalytic cores but differing positions of the α-helical region forming the coiled-coil domain. In addition, the positions of residues 342-344 of the neck linker are slightly different in the free dimer versus the dimer-tail structure. At this position, the neck linker of the dimer-tail complex is in closer contact to the motor domain core than that of the free dimer.

7
Fig. S3. Interactions at dimer-tail interface. A closer look at the hydrogen bond interactions (black dotted lines), characteristic of a ß-sheet, between tail domain (green) and the N-terminal of the ß4 strand of each motor domain.

8
Fig. S4. Conservation of residues in the head-tail interface. Multiple sequence alignment by ClustalW of metazoan kinesin-1s (Dm, Drosophila melanogaster, GI:125414; Hs-A,B,C, Homo sapiens isoforms, GI:143811412, GI:417216, GI:3043586; Sp, Strongylocentrotus purpuratus – sea urchin, GI:10270; and Lp, Loligo pealii – squid, GI:125415). The alignment for two regions from the head and one region from the tail are shown. Phe 123, Ile 126, Phe 135 and Phe 179 have hydrophobic interactions with the tail domain and are completely conserved (red boxes). Asp 185 interacts with the central Lys of the tail peptide and it has the conservative change to Glu in the other kinesin-1s. Ser 181 that was used for cross-linking after conversion to Cys is also conserved except for the natural substitution to Cys in human isoform B. His 136 is in the new interface between heads that is formed by tail binding and it is also conserved. Fungal kinesin-1s as indicated for Neurospora (Nc, Neurospora crassa, GI:6016437) are more highly diverged, but still show at most conservative changes in the key residues indicated above. The tail residues observed in the structure are indicated by the green box. They are highly conserved, even for the fungal example. However, the fungal

9
sequence completely lacks the positively charged auxiliary binding site (ABS) region (conserved positive charges indicated by the blue box) on the N-terminal side of the IAK region. This region in fungal kinesin-1s is actually an insert that is not present in metazoan kinesin-1s. In metazoan kinesins, the ABS region on the N-terminal side of the IAK region binds MTs in an ATP independent manner(37-39) and increases the affinity of tail domains for motor domains. However, the ABS region alone is not sufficient for inhibition of metazoan kinesin(19), nor required in fungal kinesins that lack the ABS region(40). This suggests a more secondary role for the ABS region of providing additional stabilizing interactions in metazoan kinesins and as a site for binding of other proteins such as light chains(41) that fungi lack. Interestingly, fungi also lack the negatively charged cluster in metazoan kinesins near helix α3 that is a potential interaction site for the positively charged ABS.
10
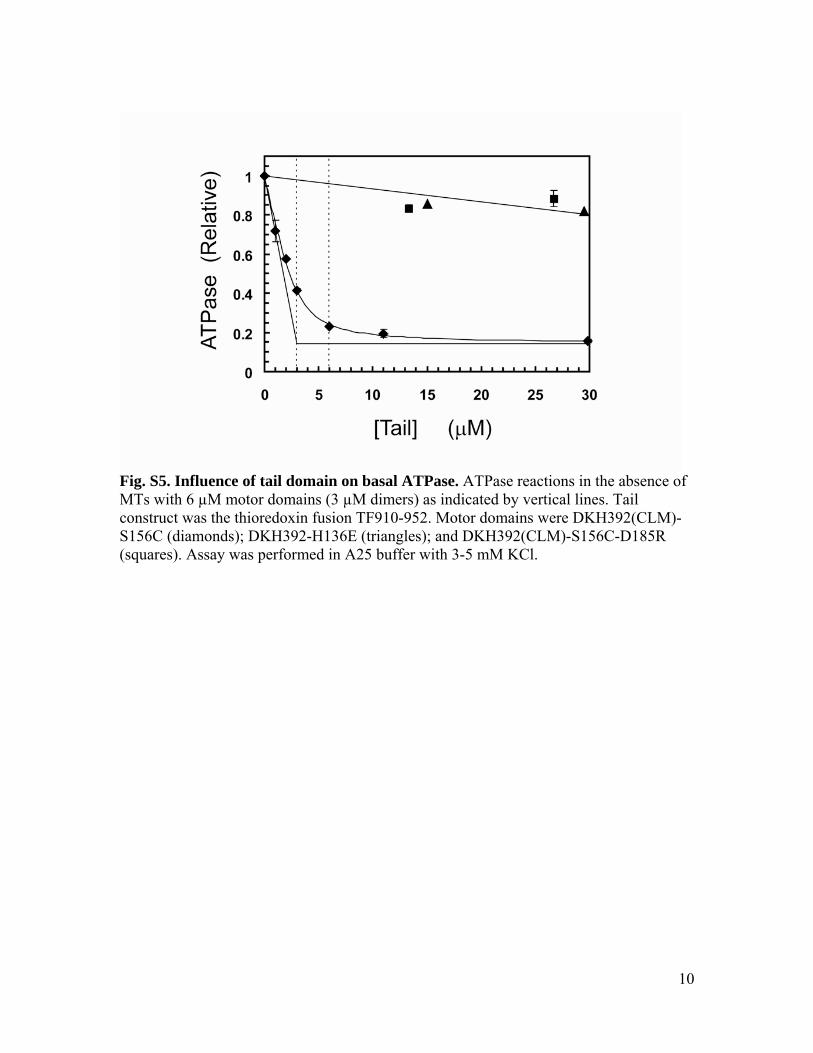
Fig. S5. Influence of tail domain on basal ATPase. ATPase reactions in the absence of MTs with 6 µM motor domains (3 µM dimers) as indicated by vertical lines. Tail construct was the thioredoxin fusion TF910-952. Motor domains were DKH392(CLM)-S156C (diamonds); DKH392-H136E (triangles); and DKH392(CLM)-S156C-D185R (squares). Assay was performed in A25 buffer with 3-5 mM KCl.

11
Fig. S6. Influence of cross-linking on MT-ATPase. MT-ATPase measurements were performed as previously described(19) in A25 buffer (25 mM ACES/KOH (pH 6.9), 2 mM magnesium acetate, 2 mM potassium EGTA, 0.1 mM potassium EDTA) with 50 mM KCl, 0.25 μM MTs and 10 µM paclitaxel. The 100 % value for the MT-ATPase of the unmodified enzyme was 19 s-1. The amount of monomer remaining in the oxidized and DTT-treated samples was determined by densitometry of stained SDS gel samples relative to a serial dilution of the unmodified sample. Assay was performed at 25°C in duplicate or greater with the standard deviation indicated.

12
Table S1. Data collection and refinement statistics. Dimer Dimer-tail Data collection Space group P212121 C2221
Cell dimensions a, b, c (Å) 76.9, 89.2, 138.4 113.8, 190.7, 146.0 α, β, γ (°) 90.0, 90.0, 90.0 90.0, 90.0, 90.0 Resolution (Å) 30.0 - 2.7 (2.84 - 2.70)* 30.0 - 2.2 (2.32 - 2.20)* Rmerge (%) 8.3 (39.5) 5.7 (36.6) I/σI 12.0 (3.6) 21.7 (5.5) Completeness (%) 99.9 (100.0) 99.9 (99.6) Redundancy 4.1 (4.1) 8.8 (8.3) Refinement Resolution (Å) 2.7 2.2 No. reflections 35890 80517 Rwork/ Rfree 21.0 / 28.9 19.5 / 25.2 No. atoms Protein 5152 10982 Ligand/ion 2 Mg2+ADP 4 Mg2+ADP Water 181 656 B-factors Protein 41.3 43.1 Ligand/ion 30.0 28.9 Water 35.5 43.0 R.m.s deviations Bond lengths (Å) 0.012 0.012 Bond angles (º) 1.82 1.79
One crystal was used for data collection for each structure. *Highest resolution shell is shown in parenthesis.

13
Table S2. MT-stimulated mant-ADP release rates of wild type and mutant motor domains. Construct Rate (µM-1s-1) Wild type 11.6 ± 0.9 H136E 11.8 ± 0.5 CLM-S156C 10.4 ± 0.7 CLM-S156C-D185R 11.6 ± 0.9 All constructs are DKH392 with substitutions as indicated. (CLM is 'Cys-lite mutant'.) Head dimers at 0.075 µM (dimer concentration) were equilibrated with excess mant-ADP in A25 buffer containing 50 mM KCl and 15% sucrose and then mixed at 30°C with an equal volume of the same buffer without sucrose and with 0.4 mM ATP and 0.6 µM MTs.

References and Notes 1. R. D. Vale, T. S. Reese, M. P. Sheetz, Identification of a novel force-generating
protein, kinesin, involved in microtubule-based motility. Cell 42, 39 (1985). doi:10.1016/S0092-8674(85)80099-4 Medline
2. N. Hirokawa, Kinesin and dynein superfamily proteins and the mechanism of organelle transport. Science 279, 519 (1998). doi:10.1126/science.279.5350.519 Medline
3. S. A. Endow, F. J. Kull, H. Liu, Kinesins at a glance. J. Cell Sci. 123, 3420 (2010). doi:10.1242/jcs.064113 Medline
4. D. D. Hackney, M. F. Stock, Kinesin’s IAK tail domain inhibits initial microtubule-stimulated ADP release. Nat. Cell Biol. 2, 257 (2000). doi:10.1038/35010525 Medline
5. M. F. Stock et al., Formation of the compact confomer of kinesin requires a COOH-terminal heavy chain domain and inhibits microtubule-stimulated ATPase activity. J. Biol. Chem. 274, 14617 (1999). doi:10.1074/jbc.274.21.14617 Medline
6. K. J. Verhey, J. W. Hammond, Traffic control: Regulation of kinesin motors. Nat. Rev. Mol. Cell Biol. 10, 765 (2009). doi:10.1038/nrm2782 Medline
7. K. A. Dietrich et al., The kinesin-1 motor protein is regulated by a direct interaction of its head and tail. Proc. Natl. Acad. Sci. U.S.A. 105, 8938 (2008). doi:10.1073/pnas.0803575105 Medline
8. D. D. Hackney, N. Baek, A. C. Snyder, Half-site inhibition of dimeric kinesin head domains by monomeric tail domains. Biochemistry 48, 3448 (2009). doi:10.1021/bi8022575 Medline
9. F. J. Kull, S. A. Endow, Kinesin: Switch I & II and the motor mechanism. J. Cell Sci. 115, 15 (2002). Medline
10. F. Kozielski et al., The crystal structure of dimeric kinesin and implications for microtubule-dependent motility. Cell 91, 985 (1997). doi:10.1016/S0092-8674(00)80489-4 Medline
11. W. Hwang, M. J. Lang, M. Karplus, Force generation in kinesin hinges on cover-neck bundle formation. Structure 16, 62 (2008). doi:10.1016/j.str.2007.11.008 Medline
12. S. Rice et al., A structural change in the kinesin motor protein that drives motility. Nature 402, 778 (1999). doi:10.1038/45483 Medline
13. J. Q. Cheng, W. Jiang, D. D. Hackney, Interaction of mant-adenosine nucleotides and magnesium with kinesin. Biochemistry 37, 5288 (1998). doi:10.1021/bi972742j Medline
14. C. V. Sindelar, K. H. Downing, An atomic-level mechanism for activation of the kinesin molecular motors. Proc. Natl. Acad. Sci. U.S.A. 107, 4111 (2010). doi:10.1073/pnas.0911208107 Medline
15. K. Hahlen et al., Feedback of the kinesin-1 neck-linker position on the catalytic site. J. Biol. Chem. 281, 18868 (2006). doi:10.1074/jbc.M508019200 Medline

16. W. Jiang, M. F. Stock, X. Li, D. D. Hackney, Influence of the kinesin neck domain on dimerization and ATPase kinetics. J. Biol. Chem. 272, 7626 (1997). doi:10.1074/jbc.272.12.7626 Medline
17. H. Y. Kaan, V. Ulaganathan, D. D. Hackney, F. Kozielski, An allosteric transition trapped in an intermediate state of a new kinesin–inhibitor complex. Biochem. J. 425, 55 (2010). doi:10.1042/BJ20091207
18. T. G. Huang, J. Suhan, D. D. Hackney, Drosophila kinesin motor domain extending to amino acid position 392 is dimeric when expressed in Escherichia coli. J. Biol. Chem. 269, 32708 (1994). Medline
19. A. G. W. Leslie, in Joint CCP4 and ESF-EACMB Newsletter on Protein Crystallography, no. 26 (Daresbury Laboratory, Warrington, UK, 1992), pp. xx–xx.
20. P. R. Evans, Scaling and assessment of data quality. Acta Crystallogr. D Biol. Crystallogr. 62, 72 (2006). doi:10.1107/S0907444905036693 Medline
21. Collaborative Computational Project No. 4, Acta Crystallogr. D50, 760 (1994).
22. A. J. McCoy et al., Phaser crystallographic software. J. Appl. Cryst. 40, 658 (2007). doi:10.1107/S0021889807021206
23. F. J. Kull, E. P. Sablin, R. Lau, R. J. Fletterick, R. D. Vale, Crystal structure of the kinesin motor domain reveals a structural similarity to myosin. Nature 380, 550 (1996). doi:10.1038/380550a0 Medline
24. G. N. Murshudov, A. A. Vagin, E. J. Dodson, Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. D Biol. Crystallogr. 53, 240 (1997). doi:10.1107/S0907444996012255 Medline
25. P. Emsley, K. Cowtan, Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126 (2004). doi:10.1107/S0907444904019158 Medline
26. V. B. Chen et al., MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 66, 12 (2010). doi:10.1107/S0907444909042073 Medline
27. E. Krissinel, K. Henrick, Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 372, 774 (2007). doi:10.1016/j.jmb.2007.05.022 Medline
28. L. L. C. Schrodinger, Pymol (2010); www.schrodinger.com/.
29. K. Kobashi, Catalytic oxidation of sulfhydryl groups by o-phenanthroline copper complex. Biochim. Biophys. Acta 158, 239 (1968). Medline
30. D. D. Hackney, M. F. Stock, Kinesin tail domains and Mg2+ directly inhibit release of ADP from head domains in the absence of microtubules. Biochemistry 47, 7770 (2008). doi:10.1021/bi8006687 Medline
31. A. S. Khalil et al., Kinesin’s cover-neck bundle folds forward to generate force. Proc. Natl. Acad. Sci. U.S.A. 105, 19247 (2008). doi:10.1073/pnas.0805147105 Medline

32. E. P. Sablin et al., Direction determination in the minus-end-directed kinesin motor ncd. Nature 395, 813 (1998). doi:10.1038/27463 Medline
33. F. Kozielski, S. De Bonis, W. P. Burmeister, C. Cohen-Addad, R. H. Wade, The crystal structure of the minus-end-directed microtubule motor protein ncd reveals variable dimer conformations. Structure 7, 1407 (1999). doi:10.1016/S0969-2126(00)80030-1 Medline
34. M. Yun et al., Rotation of the stalk/neck and one head in a new crystal structure of the kinesin motor protein, Ncd. EMBO J. 22, 5382 (2003). doi:10.1093/emboj/cdg531 Medline
35. H. Yonekura et al., Mechanism of tail-mediated inhibition of kinesin activities studied using synthetic peptides. Biochem. Biophys. Res. Commun. 343, 420 (2006). doi:10.1016/j.bbrc.2006.02.169 Medline
36. M. A. Seeger, S. E. Rice, Microtubule-associated protein-like binding of the kinesin-1 tail to microtubules. J. Biol. Chem. 285, 8155 (2010). doi:10.1074/jbc.M109.068247 Medline
37. S. Seiler et al., Cargo binding and regulatory sites in the tail of fungal conventional kinesin. Nat. Cell Biol. 2, 333 (2000). doi:10.1038/35014022 Medline
38. Y. L. Wong, S. E. Rice, Kinesin’s light chains inhibit the head- and microtubule-binding activity of its tail. Proc. Natl. Acad. Sci. U.S.A. 107, 11781 (2010). doi:10.1073/pnas.1005854107 Medline