summer meroplankton of the northern part of amursky bay (peter
TRANSCRIPT

205
SUMMER MEROPLANKTON OF THE NORTHERN PART OF AMURSKY BAY
(PETER THE GREAT BAY, SEA OF JAPAN)
V.A. Kulikova and V.A. Omelyanenko A.V. Zhirmunsky Institute of Marine Biology, FEB RAS, Vladivostok, 690041
e-mail: [email protected]
In June and August of 2005, meroplankton of the northern part of Amursky Bay under the conditions of reduced salinity and anthropogenic pollution was investigated. A comparative analysis of long-term data (1991, 1996, 1997 and 2005) on taxonomic composition and abundance of meroplankton was conducted. It was shown that distri-bution of pelagic larvae of bottom invertebrates is determined mainly by the location of the adults and the direction of currents. Relatively low salinity (up to 20‰) does not suppress viability of larvae. By the density and species diversity, meroplankton of the northern part of Amursky Bay is very similar to meroplankton of the less desali-nated and relatively clean areas of Peter the Great Bay. All this testifies to a highadaptive capacity of the larvae to unfavorable environmental factors. Reproductive potential of the benthos of Amursky Bay seems to be rather a high to provide success-ful recruitment of benthic communities.
Key words: larvae, meroplankton taxonomic composition, bottom inverteb-rates, salinity, pollution.
Исследовано состояние меропланктона в Амурском заливе в июне и августе 2005 г. в условиях пониженной солености и антропогенного загрязне-ния. Проведен сравнительный анализ многолетних данных (1991, 1996, 1997 и 2005 гг.) таксономического состава и численности меропланктона. Показано, что характер распределения пелагических личинок донных беспозвоночных определяют главным образом местоположение родительских форм и направ-ление течений. Относительно низкая (до 20‰) соленость не подавляет жизне-способности личинок. По величине плотности и видовому разнообразию меро-планктон северной части Амурского залива мало отличается от меропланктона менее опресненных и относительно чистых районов зал. Петра Великого. Все это свидетельствует о высокой адаптационной способности личинок к неблаго-приятным факторам среды. Репродуктивный потенциал бентоса в Амурском заливе достаточно высок для того, чтобы обеспечить сохранение и восстанов-ление бентосных сообществ.
Ключевые слова: личинки, таксономический состав меропланктона, донные беспозвоночные, соленость, загрязнение.
Amursky Bay is one of the most polluted areas in Peter the Great Bay. Indust-rial and domestic effluents of Vladivostok and its suburbs, petroleum productsfrom ships at anchorages, agricultural effluents and non-treated wastewater from

206
V.A. Kulikova and V.A. Omelyanenko
Ussuriysk entering the bay together with the waters of the Razdolnaya River create unfavorable ecological conditions and negatively effect organisms inhabiting the bay (Vaschenko, 2000). In this connection, monitoring studies of organisms of all trophic levels in Amursky Bay become urgent. The persistence of marine benthic communi-ties depends upon the successful recruitment of juveniles into adult habitats, there-fore investigations of the state and long-term dynamics of meroplankton (pelagic larvae of bottom invertebrates) in water areas exposed to anthropogenic pollution are extremely important. The first hydrobiological survey of meroplankton was con-ducted in Amursky Bay in 1991 (Omelyanenko and Kulikova, 2004). In 1996–1997, year-round investigations of meroplankton were conducted at a station located in the shallow northeastern part of the bay (Omelyanenko et al., 2004).
The purpose of the present study was to obtain up-to-date data on taxonomic composition, abundance and distribution of pelagic larvae of bottom invertebrates in the northern part of Amursky Bay, and to analyze long-term changes in the mero-plankton condition.
Material and methodsIn 2005, material was collected on the 11–13 of June at 14 stations, and on the
11–14 of August at 8 stations located in the northern part of Amursky Bay (Fig. 1). Larval plankton was sampled with a Norpak net using 125 μm mesh. At all stations with the depth less than 15 m, vertical net hauls were taken from the bottom to the surface, and at stations with the depth more than 15 m, vertical net hauls were taken from the depth of 15 m to the surface. In areas with the depth less than 3 m, 20-m hori-zontal hauls were performed. At the time of sampling, water temperature and salinity were measured both at the surface and in the lower horizon of the water column.
Samples were treated according to the standard methods using a stamp-dropper, Bogorov chamber, MBC-10 and Ergaval microscopes. Meroplankton was identi-fied to the lowest possible taxon. The data were then normalized to individuals/m3 (ind./m3). Larvae that could not be determined to species level were pooled under the name of a higher taxon. In order to reveal peculiarities of qualitative distribu-tion of meroplankton within the study area, a comparison of meroplankton spe-cies lists was conducted using the method of set theory. In order to reveal interan-nual variations in the abundance and taxonomic composition of meroplankton in Amursky Bay, the data obtained by the authors from 6 stations in June and from 5 stations in August of 1991 were used, as well as year-round collections of 1996 and 1997 at station 6n (see Fig. 1).
ResultsJune. Water surface temperature at different stations varied from 15.3°C to
18.1°C. The most warmed surface layer was registered at stations 1 and 65 located in the mouth of the Razdolnaya River and in the area of the bay adjacent to the river

207
Summer meroplankton
mouth, respectively (hereinafter referred to as «near-mouth area»). On the whole, water temperature near the surface in the coastal eastern and western parts of the study areas was 1–1.5°C higher than in the southwestern and central parts. Bottom water temperature varied from 4.2°C at station 3n to 18.5°C at station 5. Salinity of the surface layer ranged from 2.9‰ in the near-mouth part of the Razdolnaya River to 24.8‰ at station 85, whereas salinity of the bottom layer varied from 21.2‰ at station 24 to 34.6‰ at station 3n, i.e., the maximum salinity was registered in the south and southwestern parts of the study area (Table 1). The warmest sites of the study area displayed the smallest salinity values.
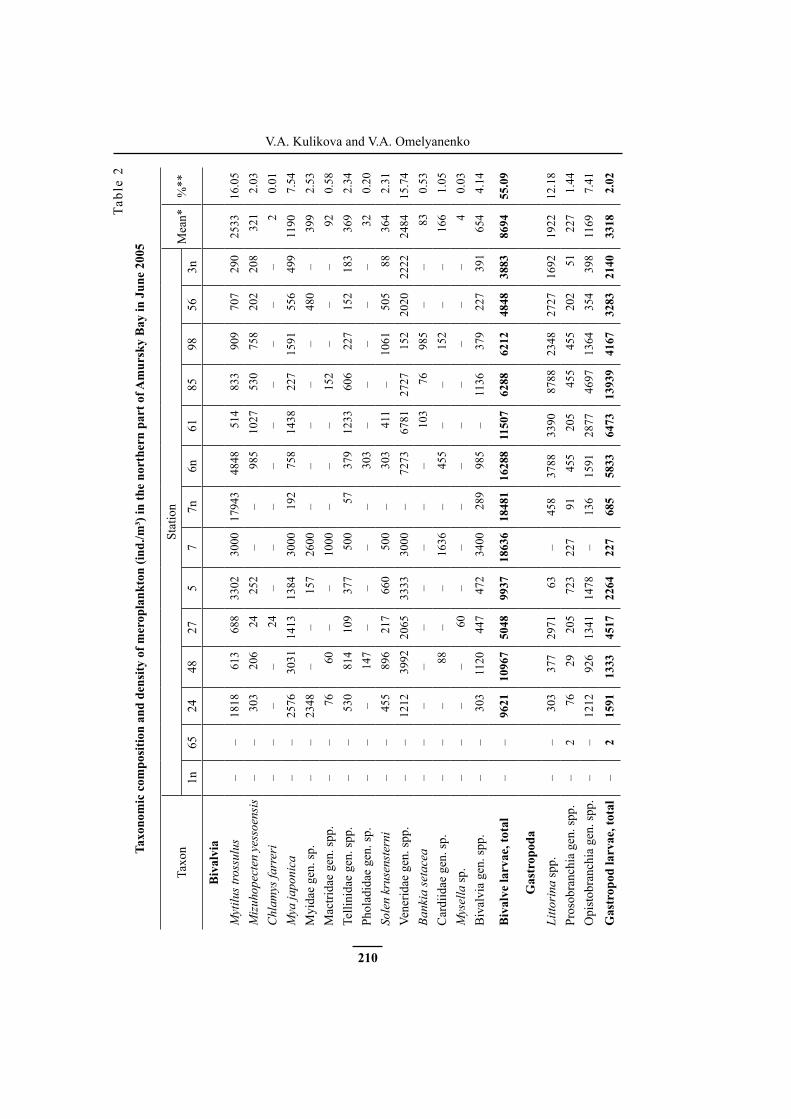
Meroplankton density varied at different stations from 0.003 to 27.8 thousand ind./m³, the mean value was 15.8 thousand ind./m³ (Fig. 2A, Table 2). Minimum number of larvae of bottom invertebrates was found in the mouth and near-mouth area of the Razdolnaya River (stations 1n and 65). Larvae were collected mainly in areas with the depth of 5–6 m (stations 85, 61, 48, 24, 7, 5, 7n and 6n) where their density ranged within 17.5–27.8 thousand ind./m³; at more open and deep-water sites (stations 3n and 56) their density was from 7 to 9 thousand ind./m³.
Bivalvia larvae were present at all stations, except for near-mouth stations 65 and 1n, and dominated practically everywhere; their density varied from 3.9 to 18.6 thousand ind./m³, the mean value was 8.7 thousand ind./m³. Maximum
Fig. 1. A scheme of location of stations in the northern part of Amursky Bay: 1 – June 2005, 2 – August 2005, 3 – June and August 1991, 4 – June and August 1996–1997. Arrows indicate surface currents prevailing in summer (Ecosystems Dynamics…, 2003).

208
V.A. Kulikova and V.A. Omelyanenko
density of bivalve larvae was observed in the coastal areas of the northern and northeastern parts of the study area at stations 6n, 7, 7n, 5, 24 and 48. At the most stations, gastropod larvae were found everywhere except for station 1n; they occupied the second place by abundance. Their density varied within the range of 0.002–13.9 thousand ind./m³, the mean value was 3.3 thousand ind./m³. Maximum
Table 1
Hydrological parameters of the studied area in June and August of 2005
Station Date Depth, m Sampled layer, m
Temperature, °C Salinity, ‰
Surface Lower horizon* Surface Lower
horizon*3n 11.06 18.0 15–0 15.3 4.2 24.7 34.6
56 11.06 16.0 15–0 15.7 6.7 24.5 34.0
61 11.06 6.0 5.5–0 16.4 9.7 21.7 30.6
48 11.06 4.5 4–0 16.9 – 20.4 –
27 11.06 7.0 7–0 16.2 8.6 23.0 30.6
98 12.06 15.0 10–0 15.6 6.7 23.5 30.1
85 12.06 5.5 5–0 15.2 9.4 24.8 30.5
65 12.06 0.8 H.c. 19.5 19.5 2.9 2.9
1n 12.06 0.6 H.c. 19.2 19.2 2.9 2.9
24 12.06 5.0 5–0 17.8 15.4 17.9 21.2
6n 12.06 6.0 5–0 18.1 9.8 20.7 29.8
7 13.06 1.5 H.c. 17.9 17.9 16.4 16.4
7n 13.06 3.5 2–0 17.6 – 18.6 –
5 13.06 5.0 2–0 15.3 18.5 17.7 22.6
3n 11.08 18.0 15–0 – – – –
85 14.08 5.5 5–0 23.6 – 1.2 –
61 11.08 6.0 5.5–0 23.6 – 9.3 –
65 14.08 0.8 H.c. 24.2 – 0.3 –
24 11.08 5.0 5–0 23.4 – 8.5 –
27 11.08 7.0 7–0 24.0 – 13.5 –
7n 11.08 3.5 2–0 23.8 – 17.8 –
6n 11.08 6.0 5–0 24.0 – 18.5 –
*Lower horizon corresponds to the lower layer of the sample.N o t e . H.c. – horizontal catching; «–» not measured.

209
Summer meroplankton
Fig. 2. Meroplankton density in the northern part of Amursky Bay in June (A) and August (B) of 2005. Designations: 1 – other taxonomic groups, 2 – Decapoda, 3 – Echinodermata, 4 – Cirripedia, 5 – Polychaeta, 6 – Gastropoda, 7 – Bivalvia, 8 – surface water salinity.
number of gastropod larvae was recorded in the northwestern shallow area at sta-tion 85. Cirripedia larvae occupied the third place by density. They also were not found in the near-mouth area. At the other stations, their density varied
210
V.A. Kulikova and V.A. Omelyanenko
Tabl
e 2
Taxo
nom
ic c
ompo
sitio
n an
d de
nsity
of m
erop
lank
ton
(ind.
/m³)
in th
e no
rthe
rn p
art o
f Am
ursk
y B
ay in
Jun
e 20
05
Taxo
nSt
atio
nM
ean*
%**
1n65
2448
275
77n
6n61
8598
563n
Biv
alvi
aM
ytilu
s tro
ssul
us–
–18
18
613
688
3302
300
017
943
484
8
514
833
909
707
290
2533
16.0
5M
izuh
opec
ten
yess
oens
is–
– 3
03
206
2
4 2
52–
–
985
102
7 5
30 7
58 2
02 2
08
321
2.0
3C
hlam
ys fa
rrer
i–
––
–
24
––
––
––
––
–
2
0.0
1M
ya ja
poni
ca–
–25
76
3031
1413
1384
300
0
192
7
58 1
438
227
1591
556
499
1190
7.5
4M
yida
e ge
n. sp
.–
–23
48–
– 1
57 2
600
––
––
– 4
80–
399
2.5
3M
actri
dae
gen.
spp.
––
7
6
60
––
100
0–
––
152
––
–
92
0.5
8Te
llini
dae
gen.
spp.
––
530
8
14 1
09 3
77
500
57
379
123
3 6
06 2
27 1
52 1
83 3
69 2
.34
Phol
adid
ae g
en. s
p.–
––
1
47–
––
–
303
––
––
–
32
0.2
0So
len
krus
enst
erni
––
455
8
96 2
17 6
60
500
–
303
4
11–
1061
505
8
8 3
64 2
.31
Vene
ridae
gen
. spp
.–
–12
12
3992
2065
3333
300
0–
727
3 6
781
2727
152
2020
2222
2484
15.7
4Ba
nkia
seta
cea
––
––
––
––
–
103
7
6 9
85–
–
83
0.5
3C
ardi
idae
gen
. sp.
––
–
88
––
163
6–
4
55–
– 1
52–
– 1
66 1
.05
Mys
ella
sp.
––
––
6
0–
––
––
––
––
4 0
.03
Biv
alvi
a ge
n. sp
p.–
– 3
03 1
120
447
472
340
0
289
9
85–
1136
379
227
39
1 6
54 4
.14
Biv
alve
larv
ae, t
otal
––
9621
1096
750
4899
3718
636
1848
116
288
1150
762
8862
1248
4838
8386
9455
.09
Gas
trop
oda
Litto
rina
spp.
––
303
377
2971
6
3–
458
3788
3390
878
823
4827
2716
9219
2212
.18
Pros
obra
nchi
a ge
n. sp
p.–
2
76
2
9 2
05 7
2322
7 9
1 4
55 2
05
455
455
202
5
1 2
27 1
.44
Opi
stob
ranc
hia
gen.
spp.
––
1212
926
1341
1478
–13
615
9128
77 4
697
1364
354
398
1169
7.4
1
Gas
trop
od la
rvae
, tot
al–
215
9113
3345
1722
6422
768
558
3364
7313
939
4167
3283
2140
3318
2.0
2

211
Summer meroplankton
Taxo
nSt
atio
nM
ean*
%**
1n65
2448
275
77n
6n61
8598
563n
Poly
chae
taPr
iono
spio
sp.
––
––
60
409
––
7
6 5
1483
322
7–
404
180
1.14
Poly
dora
spp.
––
7
6–
– 1
2611
4 5
7 6
82–
–15
210
1 4
4
96
0.61
Scol
elep
is sp
.–
––
––
––
–
76
––
––
–
5
0.03
Spio
nida
e ge
n. sp
.–
–
76
– 2
4–
152
––
––
–35
4–
4
30.
27Sp
irorb
is sp
.–
––
165
–
63
––
– 1
03 7
6–
76
13
3
50.
22C
apite
lla c
apita
ta–
––
2
9–
6
3–
–
76
––
76
––
1
70.
11N
erei
dae
gen.
spp.
–7
––
––
38
11
––
––
– 7
6
9
0.06
Tere
belli
dae
gen.
spp.
3–
––
– 5
66–
– 3
79–
––
25
–
70
0.44
Cha
etop
terid
ae g
en. s
pp.
––
––
––
––
– 2
05–
––
–
15
0.09
Phyl
lodo
cida
e ge
n. sp
p.–
––
––
––
––
––
– 2
5 1
9
3
0.02
Scol
oplo
s sp.
––
–37
7421
7 4
09–
68
–13
36–
––
– 4
152.
63O
wen
idae
gen
. spp
.–
– 3
03–
––
––
––
––
–
6
22
0.14
Poly
chae
ta g
en. s
pp.
––
1061
––
––
34
227
––
303
– 6
9
121
0.77
Poly
chae
te la
rvae
, tot
al3
715
1539
6830
216
3530
317
015
1521
5890
975
858
163
110
326.
54
Cir
ripe
dia
Bala
nus c
rena
tus
––
2273
436
–35
53 4
55 4
515
2 2
05 1
5210
61 2
5–
597
3.7
8B.
impr
ovis
us–
– 8
33 4
4216
938
0529
5524
9 7
6–
––
– 2
5 6
11 3
.87
Hes
peri
bala
nus
hesp
eriu
s–
– 5
30 1
42 2
4
94
568
––
––
– 5
1
6 1
01 0
.64
Cht
ham
alus
dal
li–
– 5
30 9
7945
9 5
3511
74 6
8–
2568
606
303
328
120
548
3.4
7
Tabl
e 2
(con
tinu
ed)
212
V.A. Kulikova and V.A. Omelyanenko
Taxo
nSt
atio
nM
ean*
%**
1n65
2448
275
77n
6n61
8598
563n
Cyp
ris la
rvae
––
–
77
––
––
– 1
03 3
79
76
25
19
4
8 0
.31
B. a
mph
itrite
––
––
–
63–
––
––
––
–
4
0.0
3
Bar
nacl
e la
rvae
, tot
al–
–41
6720
7565
280
5051
5236
222
728
7711
3614
3942
917
019
1012
.10
Ech
inod
erm
ata
Oph
iura
sars
i–
–15
216
5–
––
– 9
0914
38 8
3322
7
5119
271
1.72
Stro
ngyl
ocen
trotu
s in
term
ediu
s–
–15
220
6 3
612
6–
– 2
27
411
7
630
3–
611
00.
70
Ast
eroi
dea
gen.
spp.
––
303
360
121
126
––
–
308
152
76
76
3811
10.
71
Ech
inod
erm
larv
ae,
tota
l–
–60
673
115
725
2–
–11
3621
5810
6160
612
663
493
3.12
Phor
onid
aPh
oron
is m
uelle
ri–
––
47
36–
––
––
––
––
6
0.04
Phor
onis
ijim
ai–
–76
112
–12
6–
––
103
7615
225
6 4
80.
31
Dec
apod
a–
––
––
–34
8562
152
––
––
–26
41.
67
Bry
ozoa
––
––
– 3
1–
––
––
––
13
30.
02
Nem
ertin
i–
3–
47
–12
6–
23–
––
––
– 1
40.
09
Mer
opla
nkto
n, to
tal
312
1757
619
280
1071
322
421
2780
319
783
2515
225
274
2340
913
333
9293
6907
1578
310
0
*Mea
n de
nsity
of m
erop
lank
ton.
**Th
e po
rtion
(%) o
f the
tota
l num
ber o
f lar
vae.
No
te. «
–» –
spec
ies w
as n
ot fo
und.
Tabl
e 2
(con
tinu
ed)

213
Summer meroplankton
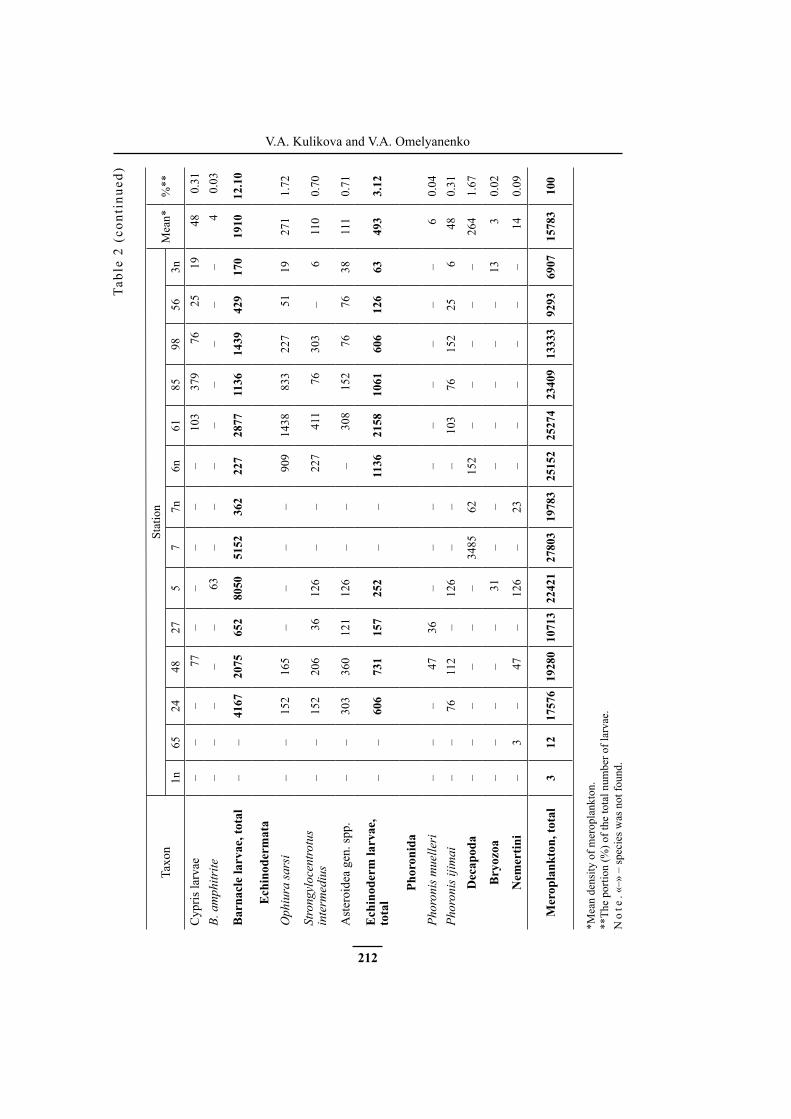
from 0.17 to 8 thousand ind./m³, the mean value was 1.9 thousand ind./m³. Maxi-mum density of barnacle larvae was observed in the northeastern and north-ern areas at stations 5, 7, 24 and 48. Polychaeta larvae were met everywhere, and their density varied from 0.003 to 4.0 thousand ind./m³, the mean value was 1.03 thousand ind./m³. Echinodermata larval density varied from 0.06 to 2.16 thousand ind./m³, the mean value was 0.5 thousand ind./m³. They were not found in the near-mouth and northeastern parts of the study area (stations 65, 1n, 7 and 7n) (Fig. 2A, Table 2).
On the whole, species diversity of meroplankton in the study area was high (Table 2). Larvae of 43 taxa were found, and 23 of them were identified up to a genusor species. At all stations, except near-mouth stations 1n and 65, the number of mero-plankton taxa was more than 20, and maximum diversity (27 taxa) was observed at stations 5 and 48. Species composition of larvae varied slightly from station to sta-tion. Larvae of bivalves and polychaetes were the most diverse everywhere.
Larvae of Mytilus trossulus, Mya japonica and representatives of family Veneri-dae (Bivalvia), Littorina spp. (Gastropoda), Scoloplos sp. (Polychaeta), Balanus improvisus, Balanus crenatus and Chthamalus dalli (Cirripedia), and Ophiura sarsi (Echinodermata) were the most abundant in the study area. However, their quan-titative distribution in the bay was not uniform. Maximum density of M. trossu-lus larvae was observed in the shallow northeastern area at stations 6n, 7n and 7, M. japonica larvae prevailed in the northern part of the study area at stations 24 and 48, and at station 24, high density of larvae of some other unidentified Mya species was found. Representatives of the family Veneridae were abundant almost every-where, and their maximum density was registered at stations 6n and 61. Gastropod larvae of the genus Littorina concentrated mainly in the northwestern shallow part of the study area. High density of Littorina larvae was also observed near the eastern coast at station 6n (Table 2).
Larvae of polychaetes were also distributed very irregularly. Thus, nectochaetes of the most abundant species, Scoloplos sp., were present mainly in the north and northeastern parts of the study area. Their maximum density was registered at stations 48 and 24. Representatives of the family Spionidae were present in plankton almost everywhere. Among them, early nectochaetes of Prionospio sp. and Polydora spp. were the most common, but their density varied greatly from station to station. Lar-vae of the family Terebellidae prevailed in the eastern part of the study area at sta-tions 5 and 6n. Larvae of the other Polychaeta species were rare and not numerous.
Nauplii of the cirripedes B. crenatus and C. dalli were found almost every-where. Their maximum density was registered in the north and northeastern parts of the study area. Most of B. improvisus and Hesperibalanus hesperius larvae were also found there.
Larvae of O. sarsi were concentrated in the western part of the study area at station 61 and in the eastern part at station 6n. They were not found in plankton of the northeastern part of the study area. Plutei of the sea urchin Strongylocentrotus

214
V.A. Kulikova and V.A. Omelyanenko
intermedius were present almost at all stations. Their maximum density was regis-tered in the shallow sites of the study area at stations 61, 98, 48 and 6n. Maximum accumulations of the sea star larvae were recorded at stations 61, 48 and 24.
Decapoda zoea larvae were met only at three stations in the northeastern and northern parts of the study area (stations 6n, 7n and 7); their maximum density was recorded at station 7.
Among larvae of the other groups, actinotrochae of the phoronid Phoronis iji-mai were the most abundant and common. They were found almost everywhere, and their maximum number was recorded at stations 98, 48 and 7 (Table 2).
August. Water surface temperature in August was higher than in June and varied at different stations within a narrow range (from 23.4 to 24.2°C). The most warmed water was in the near-mouth part of the bay, whereas the least warmed water was in the open part of the study area. Salinity in comparison with that of June was highly reduced: its minimum value (0.3‰) was recorded at station 65 near the Razdolnaya River mouth, and its maximum values (17.8 and 18.5‰) were found at stations 7n and 6n in the eastern part of the study area (Table 1).
Total density of meroplankton was lower than in June and varied at different stations from 3.8 to 19.7 thousand ind./m³, the mean value was 9.4 thousand ind./m³. Maximum larval density was recorded at stations 7n and 6n located in the eastern part of the study area; in the central, western and south parts of the study area, at sta-tions 27, 85 and 3n, it was considerably lower, and at station 65 near the Razdolnaya River mouth the larvae were not found (Table 3, Fig. 2B).
Nauplii of the cirripedes were the most abundant group of meroplankton almost everywhere. Their mean density was 3.86 thousand ind./m³ varying in the central part of the study area from 0.1 to 2.0 thousand ind./m³ and in the eastern shallow part from 3.8 to 14.0 thousand ind./m³ (Table 3, Fig. 2B). Bivalve larvae were the second group by abundance. They were distributed rather evenly within the study area; density fluctuations were from 1.45 thousand ind./m³ at station 3n to 3.7 thousand ind./m³ at station 61, the mean value was 2.2 thousand ind./m³. Polychaeta larvae occupied the third place. Their maximum density was recorded at station 7n (5.8 thousand ind./m³), the mean value was 1.4 thousand ind./m³. Abundances of larvae of Nemertini and Gastropoda were almost the same; the mean density values were 0.8 and 0.79 thousand ind./m³, respectively. Gastropod larvae were distributed evenly with some prevalence at stations 6n and 85, whereas the overwhelming majority of nemerteans pilidia was found at station 7n (5.1 thousand ind./m³). Density of larvae of the other groups was lower, and Asteroidea (Echinodermata) dominated among them.
Species diversity of meroplankton was high (Table 3). Larvae of 58 taxa were found. Maximum species diversity was registered in the central and northeastern parts of the study area (37 taxa), minimum species diversity was found at station 85 near the western coast. As well as in June, larvae of Bivalvia, Polychaeta and Cir-ripedia were the most diverse.

215
Summer meroplankton
Table 3
Taxonomic composition and density (ind./m³) of meroplankton in the northern partof Amursky Bay in August 2005
TaxonStation
Mean* %**24 27 7n 6n 61 85 3n
Bivalvia
Mytilus trossulus – – 19 – – – – 3 0.03Modiolus kurilensis 38 133 19 – 38 73 246 78 0.83Crenomutilus grayanus 38 – – – – – – 5 0.06Musculista senhousia – – – 38 38 – – 11 0.11Mytilidae gen. sp. 19 114 19 – 114 – 13 40 0.42Chlamys farreri – 19 – – 152 – – 24 0.26Chlamys swifti – 19 – – – 16 32 9 0.10Mya japonica 38 – – 284 76 – – 57 0.60Myidae gen. spp. 114 133 – – – 31 13 41 0.44Mactridae gen. sp. – 57 76 – 152 – – 41 0.43Crassostrea gigas 1364 739 227 947 1932 760 145 873 9.24Teredo navalis 152 – 38 – 38 42 38 44 0.46Bankia setacea 95 76 19 76 227 42 – 76 0.81Tellinidae gen. spp. 246 189 38 549 265 318 120 247 2.61Barnea sp. – 76 227 – – 99 347 107 1.13Zirfea pilsbryi – – – – – – 38 5 0.06Pholadidae gen. sp. 19 – 492 – – – – 73 0.77Protothaca euglipta – – – – – – 6 1 0.01Ruditapes philippinarum – – – 57 – – – 8 0.09Veneridae gen. spp. 57 – 114 95 227 151 107 107 1.13Mysella sp. 76 – – – – – 25 14 0.15Kellia japonica – 38 – – – – 13 7 0.08Bivalvia gen. spp. 246 398 379 284 455 323 309 342 3.62
Bivalve larvae, total 2500 1989 1667 2330 3712 1854 1452 2215 23.44
Gastropoda
Caecidae gen. spp. – – 38 – – – – 5 0.06Alaba vladivostokensis 76 95 38 701 76 333 – 188 1.99Prosobranchia gen. spp. 530 246 341 758 568 698 101 463 4.90Opistobranchia gen. spp. 189 76 322 – 76 130 133 132 1.40
Gastropod larvae, total 795 417 739 1458 720 1161 234 789 8.35

216
V.A. Kulikova and V.A. Omelyanenko
TaxonStation
Mean* %**24 27 7n 6n 61 85 3n
Polychaeta
Prionospio sp. 19 38 4545 38 38 42 657 768 8.13Polydora spp. – 95 682 417 76 208 19 214 2.26Pseudopolydora kempi 379 133 57 720 38 16 – 192 2.03P. paucibranchiata 19 – – 57 – – – 11 0.11Scolelepis sp. – – 19 38 – – 69 18 0.19Capitella capitata – – – – 76 42 – 17 0.18Nereidae gen. sp. – – 152 57 – – – 30 0.31Chaetopteridae gen. sp. – – – 38 – – 13 7 0.08Phyllodocidae gen. sp. – 57 114 95 189 – 6 66 0.70Pectinariidae gen. sp. – – 57 57 – – 13 18 0.19Pilargidae gen. sp. 19 – 114 – 38 – – 24 0.26Magelonidae gen. sp. – – 57 – – – 6 9 0.10Polychaeta gen. spp. – – 19 – 76 – 13 15 0.16
Polychaete larvae, total 436 322 5814 1515 530 307 795 1389 14.69
Cirripedia
Balanus crenatus 360 57 19 5170 189 42 13 836 8.84B. improvisus 3371 1932 4091 8333 2008 188 6 2847 30.13Hesperibalanus hesperius – – – – – 99 – 14 0.15Cypris larvae 95 19 246 322 – 208 82 139 1.47Rhizocephala gen. spp. – – 19 114 – 16 – 21 0.22
Barnacle larvae, total 3826 2008 4375 13939 2197 552 101 3857 40.81
Echinodermata
Strongylocentrotus nudus – – – – – – 107 15 0.16S. intermedius – – 19 – – – 13 5 0.05E. cordatum – – – – 38 – 69 15 0.16Echinoidea gen. spp. (flat urchins) – – – – – – 57 8 0.09Ophiura sarsi – – 38 – – – 139 25 0.27Amphipholis kochii – – – – – – 13 2 0.02Asteroidea gen. sp. (bipinnaria) – – – – – – 57 8 0.09Asteroidea gen. sp. (brachiolaria) – – 265 – – – 120 55 0.58
Echinoderm larvae, total – – 322 – 38 – 574 133 1.41
Table 3 (cont inued)

217
Summer meroplankton
Larvae of the barnacle B. improvisus (up to 8.3 thousand ind./m³), different nemerteans species (up to 5.1 thousand ind./m³), the polychaete Prionospio sp. (up to 4.5 thousand ind./m³), the oyster Crassostrea gigas (up to 1.9 thousand ind./m³) were the most abundant. Oyster larvae were met everywhere and prevailed at stations 61 and 24; their minimum abundance was registered at station 3n. Larvae of the most polychaete species were more abundant at stations 7n and 6n in the eastern coastal part of the study area, and they were the least abundant at stations 61 and 27. Density of nauplii of the barnacle B. improvisus was high everywhere except for station 3n. Larvae of the barnacle B. crenatus were not numerous. At station 6n, larvae of these cirripedes constituted the most portion of meroplankton. Maximum density of nemer-teans larvae was observed at station 7n. Decapod zoea prevailed with a considerable overbalance at stations 7n, 6n and 3n. Sea star larvae were present only at stations 7n and 3n. Larvae of all the other echinoderms species, among which Strongylocentrotus nudus and O. sarsi dominated, were more numerous at station 3n (Table 3). Actino-trochae of the phoronids were met only at stations 27 and 7n, whereas in June they were found in the other parts of the study area. Evidently, they were larvae of different phoronid species whose adults inhabit at a considerable distance from each other.
DiscussionWhen comparing data on the meroplankton abundance in the northeastern area
of Amursky Bay in June and in August of 2005, it turned out that the total larvae density in the second half of summer was lower both throughout the entire study
TaxonStation
Mean* %**24 27 7n 6n 61 85 3n
Decapoda 133 38 360 322 – 16 366 176 1.87
Phoronida – 38 76 – – – – 16 0.17
Nemertini – 38 5152 57 76 – 278 800 8.47
Anthozoa 76 76 265 38 – – – 65 0.69
Ascidiacea – 38 – – – – – 5 0.06
Polycladida – – 19 – – – – 3 0.03
Tornaria – – – – – – 13 2 0.02
Total meroplankton 7765 4962 18788 19659 7273 3891 3813 9450 100
*Mean meroplankton density.**Portion (%) of the total number of larvae.N o t e . «–» – species was not found.
Table 3 (cont inued)

218
V.A. Kulikova and V.A. Omelyanenko
area and at every station (Fig. 2, Table 2, 3). Meroplankton density was reduced the most considerably at stations in the northwestern and top parts of Amursky Bay, it was lower 6 times at station 85 and 2–2.5 times lower at the other stations. At the same time, near the eastern coast of the study area the meroplankton density was only slightly reduced. Such variations in the meroplankton abundance in August occurred as the result of re-distribution of dominant taxonomic groups. In June, lar-vae of Gastropoda and Bivalvia prevailed everywhere, and in August, their density and portion significantly decreased. Density of polychaete larvae in August alsodecreased at the most stations. Station 7n where abundance of polychaete larvae increased due to Prionospio sp. was the exception. Barnacle larvae density at a number of stations decreased (stations 3n and 85) or remained unchanged (stations 24 and 61). However, significant increase in the cirripedes larvae was registeredat stations 6n, 7n and 27.
In August, the species richness of meroplankton was considerably higher than in June. Among the total 73 meroplankton taxa recorded, 43 taxa (59%) were found in June and 62 (85%) – in August (Tables 2, 3). Moreover, larvae of many species including those that were the most abundant in June dropped out of plankton in August. For example, larvae of the gastropod Littorina spp., the scallop Mizuho-pecten yessoensis, the barnacle C. dalli and the polychaete Scoloplos sp. were absent. The number of the polychaete species remained unchanged while the total species list in August included 40% of the species that were not recorded in June. There was a significant growth in the number of Bivalvia and Echinodermata spe-cies: 12 additional bivalve species and 4 additional echinoderm species appeared in plankton. Instead of larvae of 2–3 gastropod species of the family Littorinidae, larvae of 3 other gastropod species (Alaba vladivostokensis, Brochina derjugini and Fartulum bucerius) were found. Larvae of the C. dalli and Balanus amphitrite bar-nacles were absent in plankton, but larvae of Rhizocephala appeared. Larvae of 5 polychaete species disappeared but almost the same number of larvae of the other polychaete species appeared. Thus, in August the meroplankton species diversity increased: the number of species at different stations varied from 25 to 37 while in June it constituted 16–25. Especially sharp increase in the species number took place at stations 7 (from 16 to 37) and 3n (from 25 to 37).
However, meroplankton density not always positively correlated with the taxo-nomic diversity: even in a case of the great number of taxa, the total larval abun-dance could be very low or, vice versa, it could be high but only due to one or several dominant species. Thus, in June at station 7, the larvae of B. improvisus, M. trossulus, two species of the genus Mya and one unidentified representative ofthe order Decapoda constituted the greatest portion of the total 18 meroplankton taxa. At station 85, gastropod larvae (the family Littorinidae) prevailed absolutely. At station 6n, 23 taxa were recorded but the larvae of M. trossulus and the species from the families Littorinidae and Veneridae dominated. At station 5, 26 taxa were

219
Summer meroplankton
found, and the larvae of M. trossulus, the species from the family Veneridae, and the barnacles B. crenatus and B. improvisus prevailed. In August, rather high abundance of larvae at station 61 was ensured by the oyster C. gigas veligers and the barnacle B. improvisus nauplii, whereas at station 7n larvae of one unidenti-fied nemerteans species created more than 1/3 of the total meroplankton density, 18.8 thousand ind./m³. In August, meroplankton density did not correlate with taxonomic diversity almost at all stations. Thus, at station 3n where the num-ber of taxa was the greatest (37), meroplankton density had the minimum value (3.8 thousand ind./m³).
It is known that in the coastal waters of temperate latitudes, low species diver-sity of zooplankton together with its high density often indicate unfavorable ecologic situation, i.e., a high pollution level and/or water eutrophication. In the northern part of Amursky Bay, the lowest species diversity in June and rather high abundance of meroplankton was recorded at stations 7 and 7n (Fig. 3). At station 7n, larvae of M. trossulus constituted 18 of 19.8 thousand ind./m³ of the total meroplankton den-sity. At station 7, larvae of M. trossulus, M. japonica, Mya sp., Veneridae gen. spp., B. improvisus and one unidentified species of the coastal crab constituted 18 of27.8 thousand ind./m³ of the total meroplankton density. Hydrochemical parameters at these two shallow-water stations were different. Station 7n had lower oxygen concentration, lower salinity and pH, high phosphate and nitrate contents (Fig. 3). Phytoplankton abundance, which is an indicator of eutrophication level, was higher at station 7n than at station 7 (Fig. 4). Thus, station 7n was characterized by less favorable hydrochemical conditions and by higher eutrophication compared to sta-tion 7, and meroplankton density and its taxonomic diversity was lower at station 7n than at station 7. Stations 85, 5 and 6n were characterized by high meroplankton abundance also had rather low values of density and biomass of phytoplankton. Thus, in the northern part of Amursky Bay, a tendency to the inverse dependence between densities of mero- and phytoplankton was observed (Fig. 4).
Analysis of data on salinity and meroplankton abundance at different stations showed that in June in the central part of the study area with relatively high salinity (22–24.8‰), meroplankton density was lower than in the eastern and northeastern parts where salinity was less than 20‰. It might indicate that reduced salinity did not negatively affect pelagic larvae. Such a conclusion conforms to the results of a number of experimental studies on the effect of salinity on the development of mollusks and echinoderms (Table 4). According to these data, larvae of a number of mollusk and echinoderm species (except for the intertidal species of the family Littorinidae) can better adapt to low salinity than adults. In the northern part of Amursky Bay, taxonomic diversity of meroplankton at stations with low salinity turned out to be not lower than at the stations with higher salinity. This indicates that larvae of a number of other taxonomic groups of bottom invertebrates are also tolerant to freshening.

220
V.A. Kulikova and V.A. Omelyanenko
Fig.
3. B
ioge
ns c
once
ntra
tion,
mer
opla
nkto
n de
nsity
and
the
num
ber o
f mer
opla
nkto
n ta
xa in
the
north
ern
part
of A
mur
sky
Bay
in Ju
ne 2
005.
Des
igna
-tio
ns: 1
– N
O3,
μg/l;
2 –
NO
2, μg
/l; 3
– P
O4,
μg/
l; 4
– Si
O3,
μg/l;
5 –
mer
opla
nkto
n de
nsity
, tho
usan
d in
d./m
³; 6
– th
e nu
mbe
r of m
erop
lank
ton
taxa
.

221
Summer meroplankton
In August, salinity was significantly lower than in June, from 0.3 to 9.3‰ inthe northwestern part of the study area and from 13.5 to 18.5‰ in the northeastern part (Table 1). Minimum salinity was registered at stations 85 and 65. The highest density of meroplankton was recorded at stations with the highest salinity. Larvae of Bivalvia, Gastropoda, Polychaeta and Cirripedia were distributed comparatively evenly within the entire area studied while larvae of Echinodermata, Phoronida, Nemertini and Decapoda were present only at sites with higher salinity (Fig. 2). On the whole, meroplankton abundance was higher at stations 61, 24, 7n and 6n where salinity was within the range of 8.5–18‰. At stations 7 and 6n with salinity of 17–18‰, meroplankton density was the highest. At station 65 with almost fresh water (0.3‰), larvae were completely absent, and at station 85 (1.2‰) they were rarely met. Thus, in August almost at all stations salinity was lower than the lowest limits of larvae salinity tolerance for the most species. This could be one of the rea-sons for decrease in larvae abundance in August.
Distribution of larvae of mollusks and echinoderms conforms well to bottom fauna distribution in the northern part of Amursky Bay (Manuilov and Petrenko, 1983; Manuilov, 1990; Selin, 1991; Zvyagintsev, 1991; Lutaenko, 2003). Thus, one of the most common for Amursky Bay species, the oyster C. gigas, creates biogenic reefs in the coastal zone including the near-mouth area with low salinity. These reefs include a great number of other bivalve species (Chlamys farreri, Arca boucardi, Crenomytilus grayanus, M. trossulus and others) as well as the other groups of bot-tom invertebrates. Epifauna of oyster shells consists of mollusks Hiatella arctica, Modiolus kurilensis, Musculista senhousia, Mya japonica, Ruditapes philippina-rum (Zvyagintsev, 1991). In the coastal zone of Skrebtsova Island located in the northestern area of the bay, bottom invertebrates community formed by the mussels C. grayanus and M. kurilensis and included about 40 animal species has been found
Fig. 4. Density of phyto- and meroplankton in the northern part of Amursky Bay in June 2005: 1 – phytoplankton density, 105 cells/m3; 2 – meroplankton density, 103 ind./m3.

222
V.A. Kulikova and V.A. Omelyanenko
(Selin et al., 1991). All these mollusk species as well as many other associated inver-tebrates ensure a high meroplankton density in this area during the reproduction period that coincides with the warmest part of a year.
In June, larvae of three Littorina species prevailed in the northern part of Amursky Bay. They were present at all stations but maximum abundance was regis-tered at stations 6n and 85 near the eastern and western coasts, respectively, i.e., in the area of adult Littorina settlements (Ivanova et al., 2008). Among bivalves, larvae of M. trossulus and M. japonica prevailed in meroplankton of the northeastern and top parts of the bay, whereas larvae of M. yessoensis were numerous in the western part. These sites are just the places where populations of adults are located. In the coastal zone of the top part of the bay, the barnacle-invader B. improvisus inhabits and creates mass colonies with the mean density values of 3.5–4.0 thousand ind./m² at the depth of 0–1.5 m (Zvyagintsev, 2005). Maximum abundance of B. improvisus larvae was also recorded in this area.
Table 4The lower limits of salinity tolerance of bottom invertebrates and their larvae (‰)
Species Larvae Adult Reference
Littorina mandshurica 18–20 8 Yaroslavtseva et al., 2000
Littorina squalida 22 20 Yaroslavtseva et al., 2000
Lottia versicolor, L. radiata, L. dorsuosa, Notoacmea sp. 22–24 – Yaroslavtseva and Sergeeva, 2000
Mytilus trossulus 8 8 Yaroslavtseva et al., 1986
Crassostrea gigas 16–18 24 Yaroslavtseva et al., 1990
Mizuhopecten yessoensis 18–20 26 Yaroslavtseva et al., 1988
Apostihopus japonicus 20 22 Kashenko, 1992, 2002
Styela clava 20–24 24 Kashenko, 1996
Eupentacta fraudatrix 22 22 Kashenko, 2000
Asterias amurensis 18–22 22–24 Kashenko, 2005
Patiria pectinifera 18–20 20 Kashenko, 2003
Strongylocentrotus intermedius 18–20 24 Yaroslavtseva and Sergeeva, 1991
Strongylocentrotus nudus 18–20 22 Yaroslavtseva and Sergeeva, 1991
Ischnochiton hakodadensis 16 24 Kashenko, 1987
Balanus improvisus – Less than 10 Zevina and Strelkov, 1983
Balanus crenatus – 10 Zevina and Strelkov, 1983
N o t e . «–» – no data.
223
Summer meroplankton
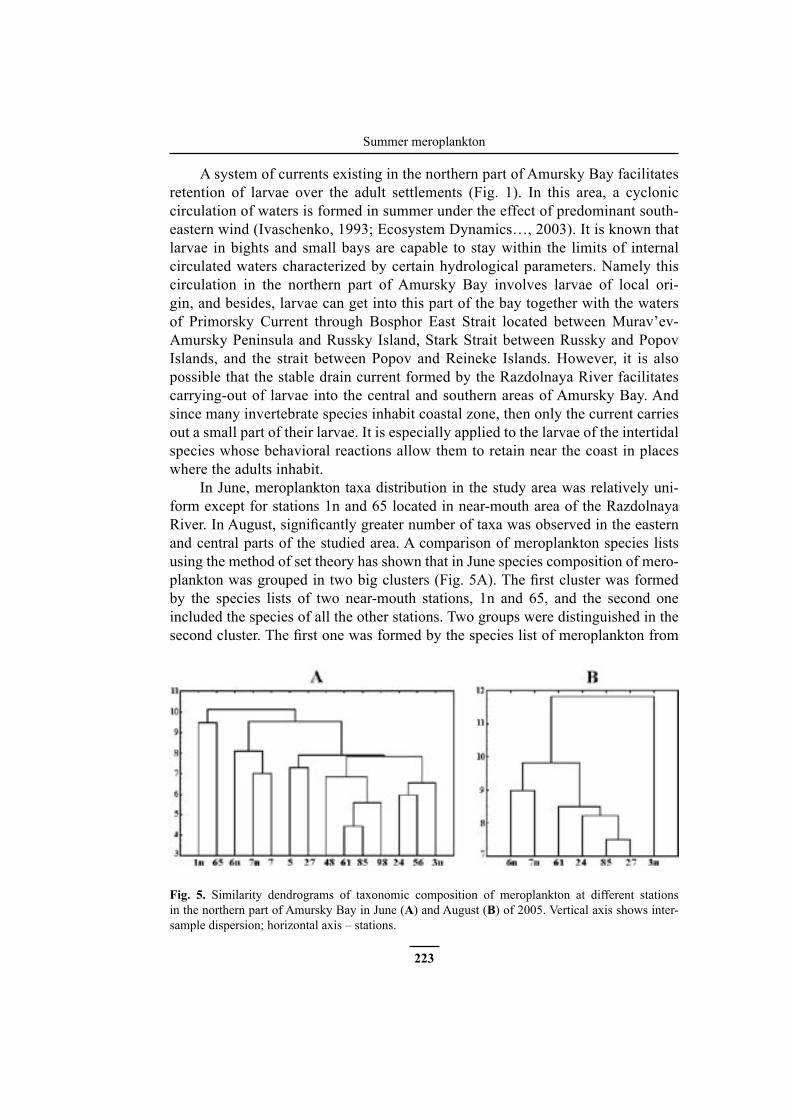
A system of currents existing in the northern part of Amursky Bay facilitates retention of larvae over the adult settlements (Fig. 1). In this area, a cyclonic circulation of waters is formed in summer under the effect of predominant south-eastern wind (Ivaschenko, 1993; Ecosystem Dynamics…, 2003). It is known that larvae in bights and small bays are capable to stay within the limits of internal circulated waters characterized by certain hydrological parameters. Namely this circulation in the northern part of Amursky Bay involves larvae of local ori-gin, and besides, larvae can get into this part of the bay together with the waters of Primorsky Current through Bosphor East Strait located between Murav’ev-Amursky Peninsula and Russky Island, Stark Strait between Russky and Popov Islands, and the strait between Popov and Reineke Islands. However, it is also possible that the stable drain current formed by the Razdolnaya River facilitates carrying-out of larvae into the central and southern areas of Amursky Bay. And since many invertebrate species inhabit coastal zone, then only the current carries out a small part of their larvae. It is especially applied to the larvae of the intertidal species whose behavioral reactions allow them to retain near the coast in places where the adults inhabit.
In June, meroplankton taxa distribution in the study area was relatively uni-form except for stations 1n and 65 located in near-mouth area of the Razdolnaya River. In August, significantly greater number of taxa was observed in the easternand central parts of the studied area. A comparison of meroplankton species lists using the method of set theory has shown that in June species composition of mero-plankton was grouped in two big clusters (Fig. 5A). The first cluster was formedby the species lists of two near-mouth stations, 1n and 65, and the second one included the species of all the other stations. Two groups were distinguished in the second cluster. The first one was formed by the species list of meroplankton from
Fig. 5. Similarity dendrograms of taxonomic composition of meroplankton at different stations in the northern part of Amursky Bay in June (A) and August (B) of 2005. Vertical axis shows inter-sample dispersion; horizontal axis – stations.

224
V.A. Kulikova and V.A. Omelyanenko
the northeastern shallow part of study area. The second group included species from the northwestern, innermost and central parts of the study area. Maximum similarity of species composition was observed in the northwestern shallow part of the bay (stations 61 and 85). In August, meroplankton composition of the central deepwater part of the bay significantly differed from that of all the other stations. Species ofthe northeastern area (stations 6n and 7n), as well as in June, were united in one sub-cluster (Fig. 5B). In August, the level of similarity of larvae species composi-tion was slightly lower than in June (Fig. 5), probably, due to the increased number of species in August.
Fig. 6. Meroplankton density in the northern part of Amursky Bay in June (A) and August (B) in different years.

225
Summer meroplankton
Investigations conducted in 1991 (Omelyanenko and Kulikova, 2004), 1996 and 1997 (Omelyanenko et al., 2004) and in 2005 in the northeastern coastal area of Amursky Bay at station 6n (Fig. 1) have shown considerable interannual vari-ations of abundance of both the total of meroplankton and separate taxonomic groups and species (Fig. 6; Tables 2, 3). In June 2005, meroplankton density at this station exceeded that of 1991, 1996 and 1997 more than 20, 2 and 4 times, respectively. Abundance of Bivalvia, Gastropoda and Polychaeta larvae increased particularly. Thus, in 2005 density of Bivalvia larvae was 13, 2 and 8 times higher than in 1991, 1996 and 1997, respectively, and density of Gastropoda larvae was 7, 2 and 2 times higher, respectively. Abundance of Decapoda and Polychaeta lar-vae increased significantly while Cirripedia abundance, on the contrary, reduced1.5–3 times. In August, when the total abundance of meroplankton was lower than in June, the proportions of meroplankton density of different taxonomic groups in different years remained almost unchanged at station 6n. Thus, in August 2005 density of Bivalvia larvae was 5, 2 and 8 times higher than in 1991, 1996 and 1997, respectively, density of Polychaeta larvae was higher 10, 2 and 32 times, and density of Cirripedia larvae was higher 6 and more times. Such interannual fluctuations of meroplakton density could be connected with fluctuations of tem-perature, salinity, and currents direction. Nevertheless, a tendency to the increase in meroplankton abundance can be evidently traced since 1991, and in 2005 both the total larvae density and larvae density of separate taxonomic groups was signifi-cantly higher than in previous years (Fig. 6, Tables 2, 3). A stable increase in mero-plankton density is typical not only for station 6n but also for the entire study area of Amursky Bay (Fig 6, Tables 2, 3). At all stations, the total abundance of larvae in 2005 was 1.3–20 times higher than in 1991, except for station 24 located in the innermost part of the bay, where meroplankton density in June 1991 and 2005 was similar. Total abundance of meroplankton often raised due to the increase in abun-dance of larvae of certain species. For example, in August 2005 such species were the oyster C. gigas (about 1 thousand ind./m³) and the barnacles B. improvisus and B. crenatus, and in June 2005, larvae of Littorina prevailed.
Increase in abundance of larvae of bottom invertebrate during the recent ten years conforms to the modern data on the state of bottom fauna of Amursky Bay (Belan et al., 2002). Investigations conducted in August 2001 by a joint expedi-tion of the Institute of Marine Biology FEB RAS and the Far Eastern Research Institute of Hydrometeorology showed a decrease in pollution level in Peter the Great Bay and in its negative effect on the bottom fauna. In 2001, bottom fauna of Amursky Bay was characterized by a high species diversity, and pollution-sensitive benthic species became more abundant. At present, Amursky Bay practi-cally does not differ from relatively clean areas of Peter the Great Bay by compo-sition and abundance of meroplankton (Kulikova et al., 2004; Omelyanenko and Kulikova, 2002).

226
V.A. Kulikova and V.A. Omelyanenko
Conclusion
In summer period, pelagic larvae of bottom invertebrates were met everywhere in the northern part of Amursky Bay except for the most freshened area adjacent to the Razdolnaya River mouth. Larvae distribution is determined mainly by location of adults and direction of currents inside the bay. Low salinity does not depress vital capacity of larvae, and only in the area adjacent to the river mouth where salinity is lower than the lowest limits of salinity tolerance of larvae, meroplank-ton is absent or its abundance is extremely low. A dependence of meroplankton abundance upon eutrophication level has been revealed: decrease in meroplankton abundance was observed at a number of stations with a high microalgae density. According to long-term data (1991, 1996, 1997, 2005), an increase in meroplank-ton abundance in the northern part of Amursky Bay took place during the recent decade. The present study testifies to a high adaptive capacity of the larvae to unfa-vorable environmental factors. Reproductive potential of the bottom invertebrates of Amursky Bay seems to be rather a high to provide successful recruitment of benthic communities.
ReferencesBelan, T.A., Oleynik, E.V., Lishavskaya, T.S. and Moshchenko, A.V., Ecological investigations in
the coastal zone of Peter the Great Bay (Sea of Japan), Abstracts of the International Workshop on Global Changes in the Far East, Vladivostok, October 2–3, 2002, Vladivostok: Dalnauka, 2002, pp. 15–18.
Ecosystem Dynamics, Formation of Bioproductivity and Bioresources of the World Ocean: The Current State, Seasonal and Inter-Annual Variability of Hydrometeorological Regime as well as Long-Term Variations of Composition and Structure of Benthic Communities of the Sea of Japan. Research Report of the Far-Eastern Research Institute of Hydrometeorology (State Register no. 01200308225), Vladivostok: FERHRI, 2003, 131 p.
Ivanova, M.B., Belogurova, M.S. and Tsurpalo, A.P., The composition and distribution of intertidal biota in the estuarine zone of Amursky Bay (Peter the Great Bay, Sea of Japan), Ecological Studies and the State of the Ecosystem of Amursky Bay and the Estuarine Zone of the Razdol-naya River (Sea of Japan), vol. 1, Vladivostok: Dalnauka, 2008, pp. 92–142.
Ivashchenko, E.A., Water circulation in Peter the Great Bay, Geographic Research in the Shelf of the Far-Eastern Seas, Vladivostok: Far East State Univ., 1993, pp. 31–61.
Kashenko, S.D., Temperature and Salinity Adaptation of Some Chiton Species in Peter the Great Bay at Different Ontogenetic Stages, Vladivostok: Inst. Mar. Biol., FEB USSR Acad. Sci., 1987, 24 p.
Kashenko, S.D., Effect of desalination on development of the Far-Eastern trepang, Russian Journal of Marine Biology, 1992, no. 3–4, pp. 43–52.
Kashenko, S.D., Effect of dilution on settlement and metamorphosis of larvae of the ascidian Styela clava, Russian Journal of Marine Biology, 1996, vol. 22, no. 3, pp.162–166.
Kashenko, S.D., Combined effect of temperature and salinity on the development of the holo-thurian Eupentacta fraudatrix, Russian Journal of Marine Biology, 2000, vol. 26, no. 3, pp. 188–193.

227
Summer meroplankton
Kashenko, S.D., Reactions of the larvae of the sea cucumber Apostichopus japonicus to sharp desalination of surface water: a laboratory study, SPC Beche-de-mer Inform. Bull., 2002, no. 16, pp. 15–21.
Kashenko, S.D., The reaction of the starfish Asterias amurensis and Patiria pectinifera (Asteroidea) from Vostok Bay (Sea of Japan) to a salinity decrease, Russian Journal of Marine Biology, 2003, vol. 29, no. 2, pp. 110–114.
Kashenko, S.D., Responses of embryos and larvae of the starfish Asterias amurensis to changes in temperature and salinity, Russian Journal of Marine Biology, 2005, vol. 31, no. 5, pp. 294–302.
Kulikova, V.A., Omelyanenko, V.A. and Tarasov, V.G., The impact of pollution on the mero-plankton of Gaidamak Bight (Vostok Bay, Sea of Japan), Russian Journal of Ecology, 2004, no. 2, pp. 91–97.
Lutaenko, K.A., Bivalve molluscan fauna of Amursky Bay (Sea of Japan) and adjacent areas. Part 2. Families Trapezidae – Periplomatidae. Ecological and biogeographic characteristics of the fauna, Bull. Russ. Far East Malacol. Soc., 2003, vol. 7, pp. 5–84.
Manuilov, V.A., Underwater Landscapes of Peter the Great Bay, Vladivostok: Far East State Univer-sity Publ. House, 1990, 168 p.
Manuilov, V.A. and Petrenko, V.S., Reefs of Peter the Great Bay, Paleogeographic Analysis and Stratigraphy of the Far-Eastern Anthropogene, Vladivostok: Far East Science Center, USSR Acad. Sci., 1983, pp. 62–72.
Omelyanenko, V.A. and Kulikova, V.A., Modern state of meroplankton in Vostok Bay (Peter the Great Bay, Sea of Japan) // Abstracts of the International Workshop on Global Changes in the Far East, Vladivostok, October 2–3, 2002, Vladivostok: Dalnauka, 2002, pp. 76–81.
Omelyanenko, V.A. and Kulikova, V.A., Pelagic larvae of Bivalvia, Gastropoda and Echinodermata in Amursky Bay (Peter the Great Bay, Sea of Japan), Proceedings of the Workshop «Climate vari-ability and human activities in relation to Northeast Asian Land-Ocean Interaction and Their Implications for Coastal Zone Management», APN Project 2004-18 NMY, Nanjing, China, Dec. 4–8, 2004, Nanjing: Nanjing Univ., 2004, pp. 125–129.
Omelyanenko, V.A., Kulikova, V.A. and Pogodin, A.G., The Meroplankton of Amursky Bay (Peter the Great Bay, Sea of Japan), Russian Journal of Marine Biology, 2004, vol. 30, no. 3, pp. 159–174.
Selin, N.I., Zhirmunsky, A.V., Levin, V.S., Ponurovsky, S.K. and Matochkin, A.G., Composition and distribution of macroepibenthos in Amursky Bay, Sea of Japan, Russian Journal of Marine Biology, 1991, no. 6, pp. 61–69.
Vashchenko, M.A., Pollution in Peter the Great Bay, Sea of Japan, and its biological consequences, Russian Journal of Marine Biology, 2000, vol. 26, no. 3, pp. 155–166.
Yaroslavtseva, L.M., Naidenko, T.Kh., Sergeeva, E.P. and Yaroslavtsev, P.V., Relationship to freshing of the edible mussel from the Sea of Japan at various stages of development, Soviet Journal of Marine Biology, 1986, no. 4, pp. 40–47.
Yaroslavtseva, L.M., Naidenko, T.Kh., Sergeeva, E.P. and Yaroslavtsev, P.V., Effect of decreased salinity on the Mizuhopecten yessoensis in ontogenesis, Soviet Journal of Marine Biology, 1988, no. 5, pp. 55–60.
Yaroslavtseva, L.M. and Sergeeva, E.P., Effect of decreased salinity of different stages of develop-ment of sea urchins of genus the Strongylocentrotus, Russian Journal of Marine Biology, 1991, no. 6, pp. 47–52.
Yaroslavtseva, L.M. and Sergeeva, E.P., Effect of desalination on the littoral Tecturid Gastropods in early development, Russian Journal of Marine Biology, 2000, vol. 26, no. 1, pp. 32–36.
Yaroslavtseva, L.M., Sergeeva, E.P. and Kashenko, S.D., Variations of sensitivity to desalination in the giant oyster ontogenesis, Russian Journal of Marine Biology, 1990, no. 6, pp. 36–42.

228
V.A. Kulikova and V.A. Omelyanenko
Yaroslavtseva, L.M., Sergeeva, E.P. and Kulikova, V.A., Salinity adaptations of the gastropods Littorina mandshurica and Littorina squalida at different life cycle stages, Russian Journal of Marine Biology, 2000, vol. 26, no. 4, pp. 264–268.
Zevina, G.B. and Strelkov, V.I., Morphological changes of Balanus improvisus plates in different regions of the area, Biological Resources of the Black Sea, Moscow: Moscow Univ. Publ. House, 1983, pp. 136–146.
Zvyagintsev, A.Yu., Seasonal changes of epifauna on valves of the giant oyster in Amursky Bay, Sea of Japan, Russian Journal of Marine Biology, 1991, no. 2, pp. 71–76.
Zvyagintsev, A.Yu., Marine Fouling in the North-West Part of the Pacific Ocean, Vladivostok: Dalnauka, 2005, 431 p.