substrate-specific kinetics of dicer-catalyzed rna processing
TRANSCRIPT

doi:10.1016/j.jmb.2010.09.030 J. Mol. Biol. (2010) 404, 392–402
Contents lists available at www.sciencedirect.com
Journal of Molecular Biologyj ourna l homepage: ht tp : / /ees .e lsev ie r.com. jmb
Substrate-Specific Kinetics of Dicer-CatalyzedRNA Processing
Srinivas Chakravarthy1,2, Samuel H. Sternberg3,Colleen A. Kellenberger3 and Jennifer A. Doudna1,2,3,4⁎1Howard Hughes Medical Institute, Berkeley, CA 94720, USA2Department of Molecular and Cell Biology, Berkeley, CA 94720, USA3Department of Chemistry, University of California, Berkeley, CA 94720, USA4Physical Biosciences Division, Lawrence Berkeley National Laboratory, Berkeley, CA 94720, USA
Received 5 August 2010;received in revised form7 September 2010;accepted 14 September 2010Available online13 October 2010
Edited by A. Pyle
Keywords:Dicer;siRNA;miRNA;TRBP;kinetics
*Corresponding author. E-mail [email protected] used: miRNA, mic
interfering RNA; dsRNA, double-sttrans-activation response (TAR) RNdsRBD, dsRNA binding domain; RIsilencing complex; Ago-2, Argonautelectrophoretic mobility shift assay.
0022-2836/$ - see front matter © 2010 E
The specialized ribonuclease Dicer plays a central role in eukaryotic geneexpression by producing small regulatory RNAs—microRNAs (miRNAs)and short interfering RNAs (siRNAs)—from larger double-stranded RNA(dsRNA) substrates. Although Dicer will cleave both imperfectly base-paired hairpin structures (pre-miRNAs) and perfect duplexes (pre-siRNAs)in vitro, it has not been clear whether these are mechanistically equivalentsubstrates and how dsRNA binding proteins such as trans-activationresponse (TAR) RNA binding protein (TRBP) influence substrate selectionand RNA processing efficiency. We show here that human Dicer is muchfaster at processing a pre-miRNA substrate compared to a pre-siRNAsubstrate under both single and multiple turnover conditions. Maximalcleavage rates (Vmax) calculated by Michaelis–Menten analysis differed bymore than 100-fold under multiple turnover conditions. TRBP was found toenhance dicing of both substrates to similar extents, and this stimulationrequired the two N-terminal dsRNA binding domains of TRBP. Theseresults demonstrate that multiple factors influence dicing kinetics. WhileTRBP stimulates dicing by enhancing the stability of Dicer–substratecomplexes, Dicer itself generates product RNAs at rates determined at leastin part by the structural properties of the substrate.
© 2010 Elsevier Ltd. All rights reserved.
Introduction
MicroRNAs (miRNAs) and short interferingRNAs (siRNAs) play central roles in controllingeukaryotic gene expression and have attracted
ress:
roRNA; siRNA, shortranded RNA; TBRP,A binding protein;SC, RNA-inducede 2; EMSA,
lsevier Ltd. All rights reserve
increasing attention as potential therapeutic agents.In the cytoplasm, many of these RNAs are generatedfrom cleavage of longer double-stranded RNA(dsRNA) substrates by the enzyme Dicer, a largemultidomain, ribonuclease III family protein.1,2However, the precursors to these RNAs are structu-rally distinct: pre-miRNAs are predicted to beimperfectly base-paired hairpins with a variable-size loop, whereas pre-siRNAs are usually perfectlybase-paired duplexes of variable length with noloop. Some species have functionally distinct Dicersthat process different kinds of substrates. Forexample, in Drosophila melanogaster, Dcr-1 andDcr-2 are responsible for the biogenesis of miRNAsand siRNAs, respectively.3 However, there is onlyone known Dicer in the human system that
d.

393Substrate-specific Dicing Kinetics
putatively processes both pre-miRNAs and pre-siRNAs. Understanding how processing rates forhuman Dicer are affected by substrate differencesand by Dicer's protein-binding partners is para-mount to identifying the molecular mechanismsthat underlie regulatory RNA production.The catalytic core of Dicer, represented by the
structure of the enzyme from Giardia intestinalis,comprises a PAZ domain N-terminal to two ribo-nuclease III domains.4 This structure, likely to becommon to all Dicer enzymes, led to a model fordsRNA recognition in which substrates are posi-tioned for cleavage to produce dsRNA products of aspecific 21- to 27 -bp length.1 However, it remainsunclear how Dicer accommodates substrate RNAswith the different helical structures and thermody-namic stabilities that characterize pre-miRNAs andpre-siRNAs.In most eukaryotes, Dicer contains additional
domains beyond those found in the Giardiaenzyme, including an N-terminal DExH/D-boxhelicase, a domain of unknown function (DUF 283)and a C-terminal dsRNA binding domain (dsRBD)(Fig. 1a). These domains function at least in part torecruit dsRNA binding proteins that are centralplayers in RNA-mediated gene silencing.6–9
Studies of RNA samples isolated frommammaliancells having or lacking Dicer show that a majority ofDicer products are miRNAs.10,11 Some examples ofendogenous siRNAs have been identified, but theiroverall occurrence is relatively uncommon in thesecell types.12,13 Nevertheless, siRNA precursors suchas short hairpin RNAs are thought to be processedby human Dicer, and some studies suggest that suchprocessing has a vital role in enhancing siRNAefficiency by facilitating optimal RNA-inducedsilencing complex (RISC) loading.14,15
In addition to Dicer's potential for differentialsubstrate recognition, double-stranded RNA bind-ing proteins that directly interact with Dicer couldinfluence its activity.16 These proteins, typicallycomprising two to three consecutive dsRBDs con-nected by variable-length linkers, include RDE-4 inCaenorhabditis elegans,17 R2D2 and Loquacious/R3D1 in D. melanogaster18,19 and trans-activationresponse (TAR) RNA binding protein (TRBP) andPACT in Homo sapiens.6,7 In C. elegans, RDE-4 isthought to be exclusively involved in the siRNApathway.20 In D. melanogaster, the substrate prefer-ences detected for R2D2 and Loquacious/R3D1suggest roles in selecting pre-siRNA and pre-miRNA substrates, respectively, and in specifyingguide strand selection during RISC assembly.19,21–23In humans, TRBP ensures efficient recruitment ofmiRNA onto Argonaute 2 (Ago-2) in RISC andenhances the stability and efficiency of both Dicerand RISC.6,14,24–26 Perhaps as a result, changes insteady-state levels of certain miRNA families cor-relate with TRBP truncation mutations that charac-
terize some colorectal cancers.27 PACT has also beenshown to bind to human Dicer,7,26 although its effecton dsRNA binding or processing remains unclear.To investigate the effects of substrate variants and
TRBP on dicing kinetics, we compared the proces-sing rates of two distinct dsRNA substrates contain-ing two-nucleotide 3′ overhangs: a 73-nucleotidehairpin with an imperfect stem (human pre-let-7a)and a 35-bp perfect duplex (Fig. 1a). Dicer showed amarked preference for processing pre-let-7a overthe 35-bp duplex, an effect most pronounced undermultiple turnover conditions in which substrate isin excess over enzyme. TRBP increased the rate ofendonucleolytic cleavage of both substrates by ∼4-to 5-fold under multiple turnover conditions. Thisstimulatory effect required the two N-terminaldsRBDs of TRBP. These results reveal substantialvariability in Dicer-catalyzed small RNA proces-sing depending on substrate sequence and second-ary structure. Although TRBP stimulates dicingactivity, substrate-dependent differences in dicingrates are inherent to human Dicer itself. Ourfindings imply that levels of miRNAs and siRNAsare influenced at least in part by precursorstructure and/or sequence. Furthermore, levels ofregulatory RNA in the cell could change as aconsequence of mutations that alter substrate RNAstructure or stability.
Results and Discussion
Dicing rates vary with different substrates
In vitro, human Dicer will cleave various dsRNAsubstrates including pre-miRNAs and short or longperfect duplexes to generate miRNAs and siRNAs,respectively. These substrates are structurally dis-tinct, leading to potential differences in dicing ratesthat could contribute to variability in miRNA and/or siRNA levels in vivo. To directly test dicing ratesof these two substrate types, we designed a 73-nucleotide hairpin with the sequence of the humanpre-miRNA pre-let-7a and a 35-bp perfect duplexrepresenting a pre-siRNA. Both substrates con-tained 3′ two-nucleotide overhangs for optimalrecognition by Dicer (Fig. 1a).We performed dicing assays under single turn-
over conditions, in which Dicer is present in largemolar excess over the substrate. Under thesereaction conditions, the rate of product formationis not limited by product release, since each enzymereacts at most with one substrate molecule. Cleav-age of pre-let-7a was considerably faster than thatobserved for the 35-bp duplex. The t1/2 (time tocleave half of the starting material) for pre-let-7a wasb5 min, whereas for the 35-bp duplex it was∼80 min (Fig. 1b).
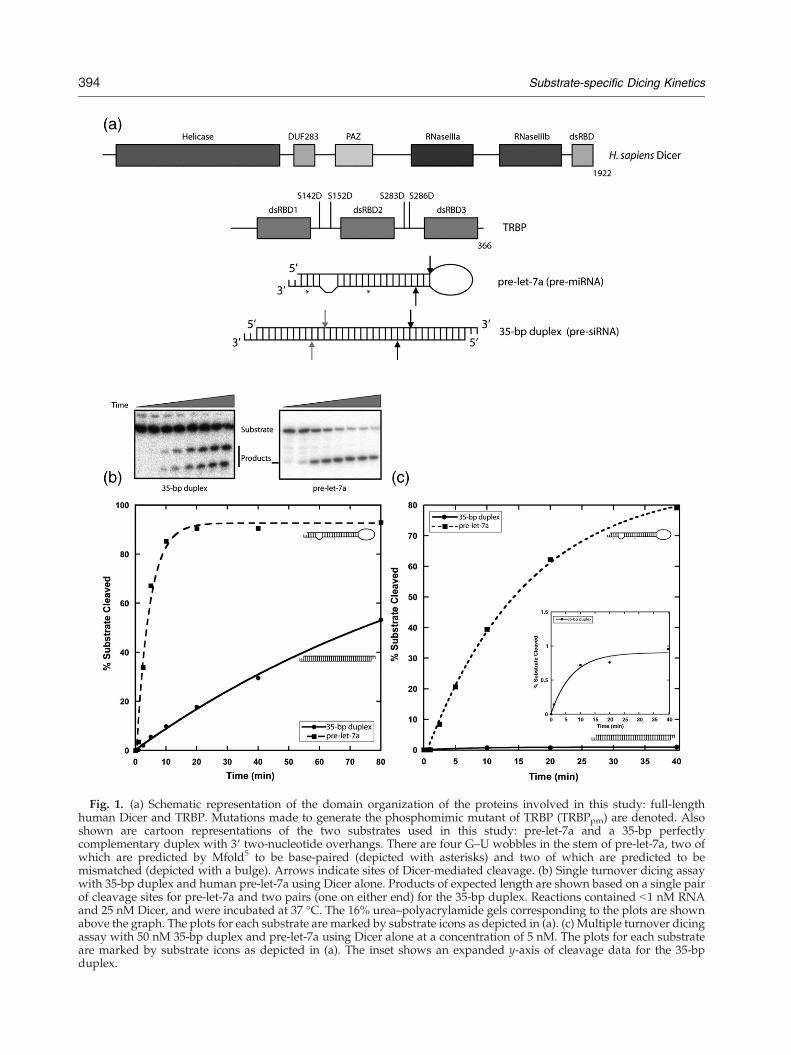
Fig. 1. (a) Schematic representation of the domain organization of the proteins involved in this study: full-lengthhuman Dicer and TRBP. Mutations made to generate the phosphomimic mutant of TRBP (TRBPpm) are denoted. Alsoshown are cartoon representations of the two substrates used in this study: pre-let-7a and a 35-bp perfectlycomplementary duplex with 3′ two-nucleotide overhangs. There are four G–U wobbles in the stem of pre-let-7a, two ofwhich are predicted by Mfold5 to be base-paired (depicted with asterisks) and two of which are predicted to bemismatched (depicted with a bulge). Arrows indicate sites of Dicer-mediated cleavage. (b) Single turnover dicing assaywith 35-bp duplex and human pre-let-7a using Dicer alone. Products of expected length are shown based on a single pairof cleavage sites for pre-let-7a and two pairs (one on either end) for the 35-bp duplex. Reactions contained b1 nM RNAand 25 nM Dicer, and were incubated at 37 °C. The 16% urea–polyacrylamide gels corresponding to the plots are shownabove the graph. The plots for each substrate are marked by substrate icons as depicted in (a). (c) Multiple turnover dicingassay with 50 nM 35-bp duplex and pre-let-7a using Dicer alone at a concentration of 5 nM. The plots for each substrateare marked by substrate icons as depicted in (a). The inset shows an expanded y-axis of cleavage data for the 35-bpduplex.
394 Substrate-specific Dicing Kinetics

395Substrate-specific Dicing Kinetics
Under multiple turnover conditions, in whichsubstrate concentration is in molar excess over thatof the enzyme, an even more striking difference insubstrate processing rates was observed. With a 10-fold excess of substrate over Dicer, the t1/2 for pre-let-7a was ∼15 min, whereas b1% of the 35-bp duplexwas cleaved in the same amount of time (Fig. 1c).Thus, Dicer catalyzes cleavage of multiple pre-let-7asubstrates per enzyme molecule while being unableto process the 35-bp substrate in multiple turnoverfashion under identical reaction conditions.Equilibrium binding affinities of each substrate for
Dicer are similar (Table 1), which suggests that thedifferences in rate and extent of Dicer-mediatedcleavage with these substrates are not simply aconsequence of substrate binding affinity. Otherpossible explanations include inefficient productrelease or suboptimal substrate loading, resultingin catalytic defects and/or the formation of nonpro-ductive enzyme–substrate complexes. Electropho-retic mobility shift assays (EMSAs) performed inprior studies, as well as our own filter binding data,showed that Dicer has a very low affinity forduplexes of the length that represents dicingproducts (Table 1).24 Additionally, the results from
Table 1. Equilibrium dissociation constants (Kds) forprotein:RNA complexes
Dicing substrates Dicing product
pre-let-7a 35-bp duplex 19-bp duplex
TRBP 1.3±0.2 nM b90 pMa 0.29±0.05 nMb
TRBPpm 2.9±0.6 nM b100 pMa 1.04±0.07 nMb
Dicer 1.8±0.4 nM 3.4±0.5 nM 700±100 nMc
Dicer–TRBP b50 pMa b51 pMa 0.24±0.04 nMb
Dicer–TRBPpm b40 pMa b52 pMa NDDicer–TRBPRBD3 1.3±0.1 nM 3.0±0.4 nM ND
Equilibrium dissociation constants were determined by filterbinding as described in Materials and Methods. Data wereanalyzed with a standard binding isotherm unless otherwisestated, according to the equation: fraction bound=A×[protein]/(Kd+[protein]), where A is the amplitude of the binding curve.ND, not determined. Errors, where included, represent thestandard fitting error.
a Because of the extremely low equilibrium dissociationconstants for these complexes, it was not possible to lower theRNA concentration sufficiently below the Kd. Therefore, bindingisotherms were fit with the solution of a quadratic equationdescribing a bimolecular dissociation reaction, as describedpreviously.38 Kds are reported as an upper bound because theRNA concentration in the reactions could not be preciselydetermined, thereby limiting the accuracy of the linearregression fit.
b Binding curves displayed apparent negative cooperativityand were best fit by allowing a variable Hill coefficient (n=0.4–0.6). Reported Kds are K0.5 values, that is, the proteinconcentration at which the fraction RNA bound is equal to 50%.
c The low affinity of this complex prevented completedefinition of the saturation regime in the binding curve. Toavoid underestimating the Kd as a result, the amplitude was fixed(A=1) when fitting the data. The reported Kd represents an upperbound estimate.
our single turnover experiments demonstrated thatcleavage rates already differ by N10-fold underconditions in which product release should notinfluence the accumulation of product. We thereforespeculate that the 35-bp duplex is “misloaded” ontoDicer, thereby retarding the progress of the reaction.This might not occur with pre-let-7a if its structuralfeatures preclude nonproductive binding to Dicer.This possibility is consistent with the observationthat in vivo, human Dicer is predominantly respon-sible for the production of miRNAs.10 It is notewor-thy that even among pre-miRNAs, there naturallyexist many variations in structure such as the size ofthe loop and the number of mismatches in the stem,which may lead to considerable differences in theway distinct substrates are recognized and pro-cessed by Dicer. Recent studies have revealed pre-miRNAs with structural properties that make themunsuitable substrates for Dicer and are insteadprocessed by Ago-2.28,29 It is therefore conceivablethat Dicer has a specific set of substrate structuralrequirements that lead to considerable variability incatalytic efficacy.
TRBP has a stimulatory effect on dicing kinetics
TRBP binds and stabilizes Dicer, modifies Dicer'sendonucleolytic activity and has been suggested tohelp load product dsRNAs onto Ago-2 duringformation of RISCs.6,24 In addition, TRBP trunca-tions correlate with changes in the populationdistributions of certain endogenous miRNAs invivo.27 Studies using chimeric constructs of RDE-4and TRBP showed that the two N-terminal dsRBDscontribute largely to the dsRNA binding affinity ofTRBP.30 Whether and how TRBP influences Dicer'sselection and speed of dsRNA substrate processinghas not been well characterized.To test the influence of TRBP on dicing kinetics,
recombinantly expressed Dicer and TRBP wereincubated together to form a heterodimeric com-plex, which was then purified by size-exclusionchromatography to remove any free TRBP.8 Controlexperiments showed that excess TRBP binds tightlyto dsRNA and sequesters it from the Dicer–TRBPcomplex, inhibiting dicing without necessarilyreflecting the behavior of the Dicer–TRBP complexitself (data not shown).Equilibrium binding analysis of Dicer–dsRNA
complexes in the presence and absence of TRBP,by means of an EMSA, suggested that TRBPenhances complex stability with both the pre-let-7aand the 35-bp duplex (Fig. 2a). The Dicer–TRBPcomplex gave rise to a slowly migrating ternarycomplex with either dsRNA substrate, whereasDicer alone did not. To obtain quantitative informa-tion about dsRNA binding affinities, we performedfilter binding assays to measure equilibrium disso-ciation constants (Kds) of Dicer alone and the Dicer–
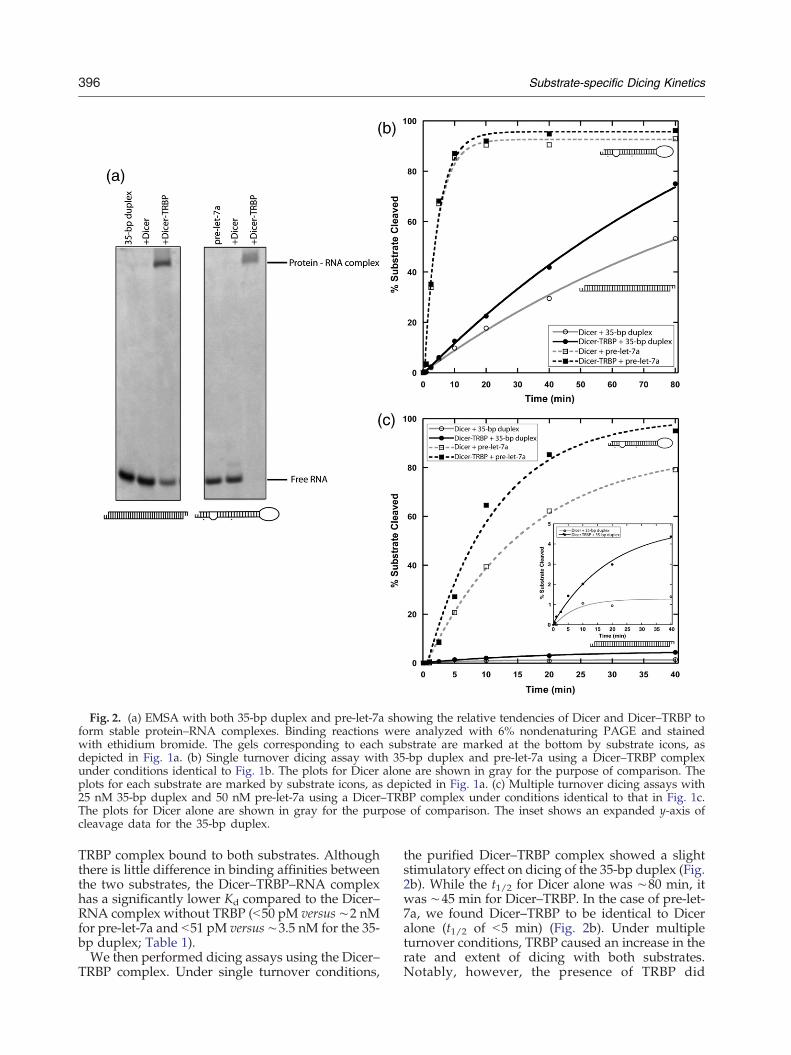
Fig. 2. (a) EMSA with both 35-bp duplex and pre-let-7a showing the relative tendencies of Dicer and Dicer–TRBP toform stable protein–RNA complexes. Binding reactions were analyzed with 6% nondenaturing PAGE and stainedwith ethidium bromide. The gels corresponding to each substrate are marked at the bottom by substrate icons, asdepicted in Fig. 1a. (b) Single turnover dicing assay with 35-bp duplex and pre-let-7a using a Dicer–TRBP complexunder conditions identical to Fig. 1b. The plots for Dicer alone are shown in gray for the purpose of comparison. Theplots for each substrate are marked by substrate icons, as depicted in Fig. 1a. (c) Multiple turnover dicing assays with25 nM 35-bp duplex and 50 nM pre-let-7a using a Dicer–TRBP complex under conditions identical to that in Fig. 1c.The plots for Dicer alone are shown in gray for the purpose of comparison. The inset shows an expanded y-axis ofcleavage data for the 35-bp duplex.
396 Substrate-specific Dicing Kinetics
TRBP complex bound to both substrates. Althoughthere is little difference in binding affinities betweenthe two substrates, the Dicer–TRBP–RNA complexhas a significantly lower Kd compared to the Dicer–RNA complex without TRBP (b50 pM versus ∼2 nMfor pre-let-7a and b51 pM versus ∼3.5 nM for the 35-bp duplex; Table 1).We then performed dicing assays using the Dicer–
TRBP complex. Under single turnover conditions,
the purified Dicer–TRBP complex showed a slightstimulatory effect on dicing of the 35-bp duplex (Fig.2b). While the t1/2 for Dicer alone was ∼80 min, itwas ∼45 min for Dicer–TRBP. In the case of pre-let-7a, we found Dicer–TRBP to be identical to Diceralone (t1/2 of b5 min) (Fig. 2b). Under multipleturnover conditions, TRBP caused an increase in therate and extent of dicing with both substrates.Notably, however, the presence of TRBP did

397Substrate-specific Dicing Kinetics
nothing to ameliorate the relatively inefficientprocessing of the 35-bp duplex compared to pre-let-7a (Fig. 2c). With pre-let-7a, the t1/2 for Diceralone was ∼15 min, compared to ∼8 min for theDicer–TRBP complex (Fig. 2c).TRBP-mediated stimulation of dicing could occur
at multiple stages of the dicing reaction trajectory.The much higher affinity of the Dicer–TRBPcomplex for RNA substrates, as compared to Diceralone (Table 1), strongly suggests that TRBP accel-erates dicing in part by assisting in substraterecruitment. It is also possible that substrate bindingexerts some amount of conformational strain onDicer, which TRBP may alleviate via its interactionwith Dicer and by properly orienting the substratein Dicer's active site. Furthermore, we cannotexclude the possibility that TRBP also plays a rolein facilitating product release and enzyme turnover.Future experiments will be needed to fully explainthe mechanistic basis for TRBP's stimulatory effecton dicing.
TRBP requires high RNA binding affinity tostimulate dicing
In order to obtain a more detailed picture of theeffect of substrate variation and the presence ofTRBP on Dicer function, we performed dicing assaysunder multiple turnover conditions using a series ofsubstrate concentrations. Michaelis–Menten plotsobtained from these data revealed that Dicer'smaximum cleavage rate (Vmax) for pre-let-7a is100-fold higher than that for the 35-bp perfectduplex. We also find that the Dicer–TRBP complexshows a 4- to 5-fold increase in Vmax over thatmeasured for Dicer alone, irrespective of substratetype (Fig. 3a and b). Thus, the specific sequence andstructure of RNA substrates have substantial effectson catalysis rates, resulting in Vmax differences span-ning multiple orders of magnitude. Furthermore,TRBP leads to an overall enhancement in cleavagerate, regardless of the specific substrate. Surpri-singly, Km values extracted from the Michaelis–Menten curves are in the tens of nanomolar range(Fig. 3a and b), both in the presence and in theabsence of TRBP, whereas the Kds for substratebinding are up to three orders of magnitude lower(Table 1). Because Km does not equal Kd for thesecomplexes, product formation may not be the rate-limiting step. For example, substrate molecules mayneed to undergo conformational changes after initialbinding in order to form productive enzyme–substrate complexes.The stimulation of dicing activity by TRBP could
be due either to its dsRNA binding affinity, to aconformational change brought about by protein–protein interactions between TRBP and Dicer, or to acombination of both. Previous RNA-binding studiesusing wild-type or mutated forms of TRBP impli-
cated the first two dsRBDs in RNA binding; whiledsRBD2 was clearly shown to have the highestaffinity for dsRNA,31 dsRBD1 also enhanced theRNA binding affinity of a TRBP–RDE4 chimera.30 Inagreement, studies of truncation mutants showedthat the C-terminal dsRBD3 is dispensable for RNAbinding30,32 and contributes primarily to protein–protein interactions and Dicer binding.6,33 Further-more, it has also been shown in Drosophila that atruncation mutant of Loquacious containingdsRBD2 and dsRBD3 is sufficient both for bindingDcr-1 (via dsRBD3) and for enhancing Dcr-1-mediated miRNA production (via dsRBD2).34
In order to investigate the roles of the dsRNA-binding and Dicer-binding dsRBDs of TRBP instimulating dicing, we designed a truncation con-struct (TRBPRBD3) that lacked dsRBD1 and dsRBD2but retained its ability to interact with Dicer. Wewere able to purify the Dicer–TRBPRBD3 complex,which failed to produce an electrophoretic mobilityshift with either the pre-let-7a or the 35-bp dsRNAsubstrate, similar to the behavior of Dicer alone(data not shown). Additionally, the measuredbinding affinity of Dicer–TRBPRBD3 for either sub-strate is within error of the affinity measured forDicer alone (Table 1). These observations stronglysuggest that TRBP confers high dsRNA bindingaffinity to the Dicer–TRBP complex via its two N-terminal dsRBDs.The Dicer–TRBPRBD3 complex was tested in dicing
assays with both the pre-let-7a and the 35-bp duplexunder multiple turnover conditions. In both cases,this enzyme complex produced little or no stimula-tion of dicing relative to the rate observed for Diceralone (Fig. 3c). However, we have not completelyeliminated the possibility that TRBPRBD3 has a lowerbinding affinity for Dicer compared to that of full-length TRBP and therefore dissociates appreciablyduring the reaction. Nevertheless, these data indi-cate that dsRBD2—and perhaps dsRBD1 to a lesserextent—is required for TRBP's stimulatory affect ondicing. In agreement with studies of Loquacious andDcr-1 from Drosophila,34 dsRBD3 is required forDicer binding. Future studies of TRBP containingpoint mutations in dsRBD1 and dsRBD2 that inhibitRNA binding will be needed to investigate whetherthese domains are also involved in protein–proteininteractions that affect dicing kinetics.
A TRBP phosphorylation mimic does notsignificantly affect dicing kinetics
The posttranslational phosphorylation of TRBP invivo has been shown to increase the apparent half-life of Dicer and affect cellular miRNA levels.25
TRBP expressed in Escherichia coli is devoid ofposttranslational modifications, whereas TRBPexpressed in a baculovirus system exhibits hetero-geneous phosphorylation25 (E. Ma and J.A.D.,
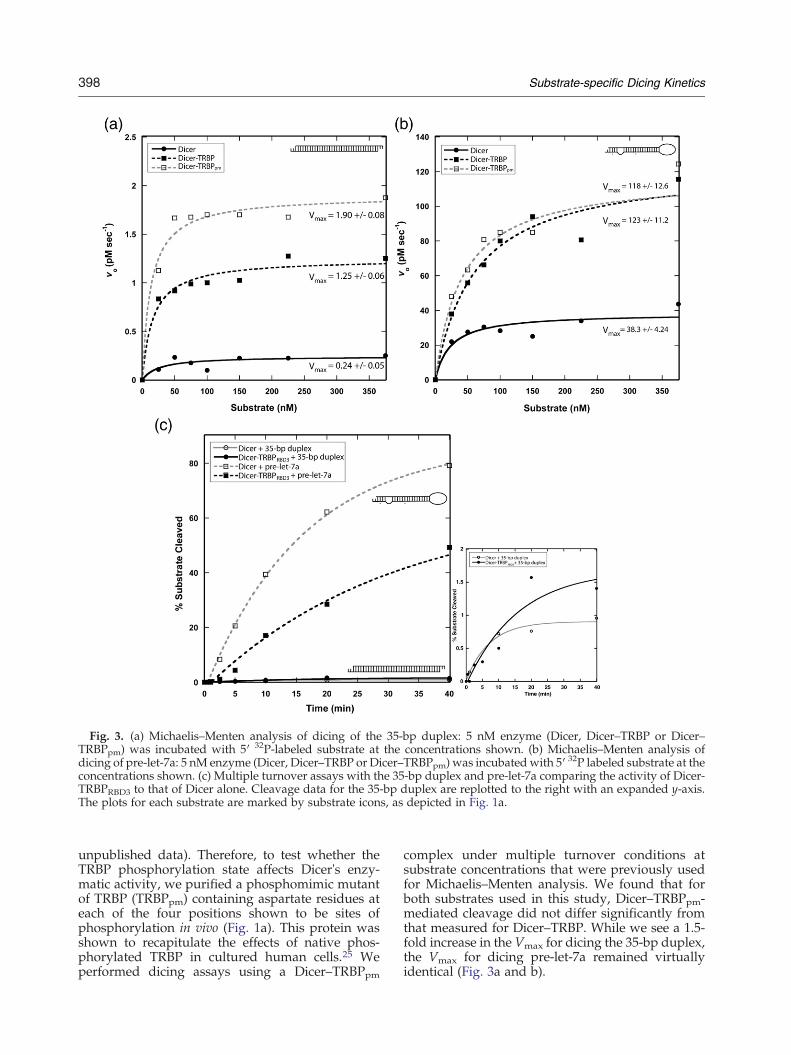
Fig. 3. (a) Michaelis–Menten analysis of dicing of the 35-bp duplex: 5 nM enzyme (Dicer, Dicer–TRBP or Dicer–TRBPpm) was incubated with 5′ 32P-labeled substrate at the concentrations shown. (b) Michaelis–Menten analysis ofdicing of pre-let-7a: 5 nM enzyme (Dicer, Dicer–TRBP or Dicer–TRBPpm) was incubated with 5′ 32P labeled substrate at theconcentrations shown. (c) Multiple turnover assays with the 35-bp duplex and pre-let-7a comparing the activity of Dicer-TRBPRBD3 to that of Dicer alone. Cleavage data for the 35-bp duplex are replotted to the right with an expanded y-axis.The plots for each substrate are marked by substrate icons, as depicted in Fig. 1a.
398 Substrate-specific Dicing Kinetics
unpublished data). Therefore, to test whether theTRBP phosphorylation state affects Dicer's enzy-matic activity, we purified a phosphomimic mutantof TRBP (TRBPpm) containing aspartate residues ateach of the four positions shown to be sites ofphosphorylation in vivo (Fig. 1a). This protein wasshown to recapitulate the effects of native phos-phorylated TRBP in cultured human cells.25 Weperformed dicing assays using a Dicer–TRBPpm
complex under multiple turnover conditions atsubstrate concentrations that were previously usedfor Michaelis–Menten analysis. We found that forboth substrates used in this study, Dicer–TRBPpm-mediated cleavage did not differ significantly fromthat measured for Dicer–TRBP. While we see a 1.5-fold increase in the Vmax for dicing the 35-bp duplex,the Vmax for dicing pre-let-7a remained virtuallyidentical (Fig. 3a and b).
399Substrate-specific Dicing Kinetics
Paroo et al. showed that the relative abundance ofcertain miRNAs varied between cells expressing thephosphomimic TRBP mutant and those expressing anonphosphorylatable TRBP mutant.25 Notably, inthis study, let-7a was among the miRNAs whoseabundance decreased in the TRBPpm-containingcells. The absence of an effect of TRBPpm on dicingrates in this study suggests that instead of alteringdicing kinetics, TRBP phosphorylation may influencemiRNA levels by an indirect mechanism. A moredetailed analysis with pre-miRNAs representative ofboth the up-regulated and the down-regulatedpopulations would nevertheless be desirable toevaluate the possibility that TRBP phosphorylationinfluences dicing kinetics in other contexts. We notethat the dissociation constants for TRBPpm–RNAcomplexes are comparable to those measured forwild-type TRBP (Table 1), consistent with the ideathat TRBP stimulates dicing by a mechanism thatgenerally depends on its RNA binding affinity.
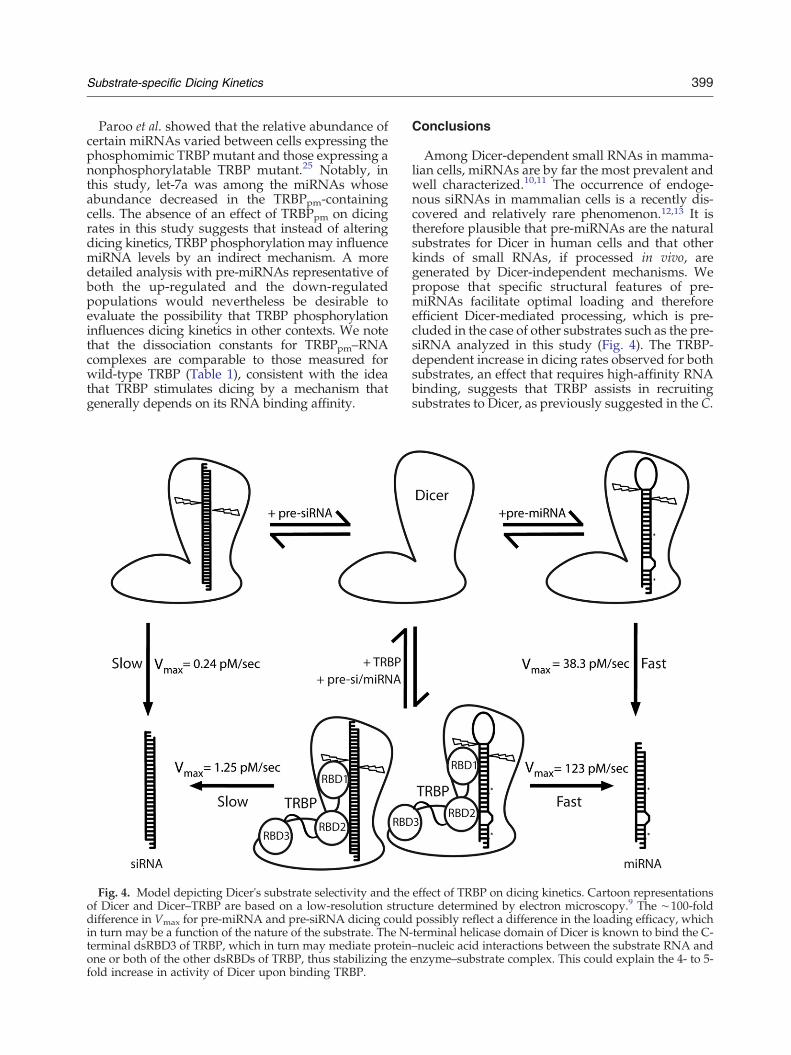
Fig. 4. Model depicting Dicer's substrate selectivity and theof Dicer and Dicer–TRBP are based on a low-resolution strucdifference in Vmax for pre-miRNA and pre-siRNA dicing couldin turn may be a function of the nature of the substrate. The Nterminal dsRBD3 of TRBP, which in turn may mediate proteinone or both of the other dsRBDs of TRBP, thus stabilizing thefold increase in activity of Dicer upon binding TRBP.
Conclusions
Among Dicer-dependent small RNAs in mamma-lian cells, miRNAs are by far the most prevalent andwell characterized.10,11 The occurrence of endoge-nous siRNAs in mammalian cells is a recently dis-covered and relatively rare phenomenon.12,13 It istherefore plausible that pre-miRNAs are the naturalsubstrates for Dicer in human cells and that otherkinds of small RNAs, if processed in vivo, aregenerated by Dicer-independent mechanisms. Wepropose that specific structural features of pre-miRNAs facilitate optimal loading and thereforeefficient Dicer-mediated processing, which is pre-cluded in the case of other substrates such as the pre-siRNA analyzed in this study (Fig. 4). The TRBP-dependent increase in dicing rates observed for bothsubstrates, an effect that requires high-affinity RNAbinding, suggests that TRBP assists in recruitingsubstrates to Dicer, as previously suggested in the C.
effect of TRBP on dicing kinetics. Cartoon representationsture determined by electron microscopy.9 The ∼100-foldpossibly reflect a difference in the loading efficacy, which
-terminal helicase domain of Dicer is known to bind the C-–nucleic acid interactions between the substrate RNA andenzyme–substrate complex. This could explain the 4- to 5-

400 Substrate-specific Dicing Kinetics
elegans and D. melanogaster systems,30,34,35 and/orstabilizes the interaction between Dicer and its RNAsubstrates (Fig. 4). However, Dicer itself possesses anintrinsic ability to distinguish between substrateswith different sequences and structural character-istics. Together, these results show that the specificnature of the dicing substrate heavily influences thecatalytic production of regulatory RNAs and couldtherefore be an important aspect of small RNAbiogenesis in vivo and for the development of RNAi-based therapeutics.
Materials and Methods
Sample preparation
We purified Dicer, TRBP and Dicer–TRBP complexes bymeans of published protocols.36 We used site-directedmutagenesis to incorporate the TRBP point mutantsS142D, S152D, S283D and S286D, and purified TRBPpm
25
with the same procedure as for wild-type TRBP. Humanpre-let-7a was synthesized by in vitro transcription with T7RNApolymerase fromaDNA template containing a doubleribozyme system to ensure homogeneous 5′ and 3′ ends.37
The two strands of the 35-bp duplex (a and b) and those ofthe 19-bp duplex (a and b) were synthesized by IDT(Integrated DNA Technologies, Coralville, IA). All sub-strates were purified with 16% urea-PAGE, and purifiedpre-let-7a, 35-bp duplex b and 19-bp duplex a were then 5′end-labeled with T4 polynucleotide kinase (New EnglandBiolabs Inc., Beverly, MA) and [γ-32P]ATP. The substrateswere heated at 65 °C for 10 min in annealing buffercontaining 3 mM MgCl2, 30 mM NaCl and 100 mM Tris–HCl (pH 7.5), followed by flash cooling for refolding pre-let-7a and slow cooling to anneal the 35- and 19-bp duplexes.The sequences of the substrates are as follows:
pre-let-7a: 5′-UGAGGUAGUAGGUUGUAUA-GUUUUAGGGUCACACCCACCACUGGGA-GAUAACUAUACAAUCUACUGUCUUACC-3′
35-bp duplex a: 5′-UGAGGUAGUAGGUUGUAUA-GUUUGAAAGUUCACGAUU-3′
35-bp duplex b: 5′-UCGUGAACUUUCAAACUAUA-CAACCUACUACCUCAAA-3′
19-bp duplex a: 5′-GUCACAUUGCCCAAGUCUCTT-3′
19-bpduplex b: 5′-GAGACUUGGGCAAUGUGACTT-3′
Kinetic assays
Single turnover assays were performed in 45 -μLmixtures, which consisted of 25 nM enzyme and b1 nM5′ 32P-labeled substrate. Upon incubation at 37 °C, 5 -μLaliquots were removed and mixed with 6 μL of loadingbuffer at time points of 0.5, 1, 2.5, 5, 10, 20, 40 and 80 min.These samples were heated at 70 °C for 10 min prior tobeing run on a 16% urea–polyacrylamide gel and quantifiedwith a phosphorimager (GE Healthcare). The data were
plotted with Kaleidagraph (Synergy software). Multipleturnover dicing assays were also performed in 45 -μLmixtures containing 5 nM enzyme and 5′ 33P-labeledsubstrate at concentrations of 25, 50, 75, 100, 150, 225 and375 nM. Reactions were incubated at 37 °C, and 5 -μLaliquots were taken after 0.5, 1, 2.5, 5, 10, 20, 40 and 80 min.Variations in the time points were required in order toadapt to varying levels of dicing activity with differentsubstrates and different proteins, as indicated in the figureaxes. After quantification, initial velocities (vo) at eachsubstrate concentration (S) were determined by linearregression, and Vmax along with standard errors werecalculated by fitting to theMichaelis–Menten equation, vo=(Vmax×S)/(Km+S), with Kaleidagraph.
Electrophoretic mobility shift assay
The 35-bp duplex and pre-let-7a RNAswere annealed in100 mM Tris–HCl (pH 7.5), 3 mMMgCl2 and 30 mMNaClby heating at 65 °C for 10 min and either slow cooled (35-bp duplex) or flash cooled (pre-let-7a). Each of theseRNAs (∼10.5 pmol) were then incubated with equimolarDicer, Dicer–TRBP or Dicer–TRBPRBD3 for 1 h at 4 °C in20 mM Tris–HCl (pH 7.5), 25 mM NaCl, 5 mM EDTA(ethylenediaminetetraacetic acid), 1 mM DTT and 1%glycerol. Reactions were analyzed on a 6% nondenaturingpolyacrylamide gel by staining with ethidium bromide.
Filter binding assays
Serial dilutions of Dicer, TRBP or Dicer–TRBP constructswere incubated with b1 nM of 5′ 32P-radiolabeled 35-bpduplex, pre-let-7a or 19-bp duplex in binding buffercontaining 20 mM Tris–HCl (pH 7.5), 25 mM NaCl,1 mM DTT, 1% glycerol and 0.01% Igepal CA-630.Reactions with Dicer were supplemented with 5 mMEDTA to prevent RNA cleavage by Mg2+ chelation. Afterreactions were incubated (25- or 50 -μL volume) on ice for1 h, samples were applied by vacuum to a dot-blot appa-ratus containing two pieces ofWhatman filter paper belowthree membranes: 0.2 -μmpore size Tuffryn (Pall Co.), 0.1 -μm pore size Protran (Whatman) and Hybond-N (Amer-sham). Themembraneswere equilibrated in binding bufferfor at least 20min prior to use. After applying reactions andwashing with 50 μL of binding buffer, membranes weredried and exposed overnight on a phosphor screen. Theamounts of free RNA (retained on Hybond-N membrane)and protein-bound RNA (retained on Protran membrane)were quantified with a phosphorimager (GE Healthcare).The extent of complex aggregation, as detected byradioactivity on the Tuffryn membrane, was negligible.The fraction of RNA bound, calculated as the ratio ofradioactivity on the Protran membrane to the sum ofradioactivity on the Protran and Hybond-N membranes,was plotted as a function of protein concentration. Kdswere determined by fitting the data to binding isothermswith KaleidaGraph (see Table 1 for details).
References
1. MacRae, I. J., Zhou, K. & Doudna, J. A. (2007).Structural determinants of RNA recognition andcleavage by Dicer. Nat. Struct. Mol. Biol. 14, 934–940.

401Substrate-specific Dicing Kinetics
2. Tomari, Y. & Zamore, P. D. (2005). Perspective:machines for RNAi. Genes Dev. 19, 517–529.
3. Carthew, R. W. & Sontheimer, E. J. (2009). Origins andmechanisms of miRNAs and siRNAs. Cell, 136,642–655.
4. Macrae, I. J., Zhou, K., Li, F., Repic, A., Brooks, A. N.,Cande, W. Z. et al. (2006). Structural basis for double-stranded RNA processing by Dicer. Science, 311,195–198.
5. Zuker, M. (2003). Mfold web server for nucleic acidfolding and hybridization prediction. Nucleic AcidsRes. 31, 3406–3415.
6. Haase, A. D., Jaskiewicz, L., Zhang, H., Laine, S., Sack,R., Gatignol, A. & Filipowicz, W. (2005). TRBP, aregulator of cellular PKR and HIV-1 virus expression,interacts with Dicer and functions in RNA silencing.EMBO Rep. 6, 961–967.
7. Lee, Y., Hur, I., Park, S. Y., Kim, Y. K., Suh, M. R. &Kim, V. N. (2006). The role of PACT in the RNAsilencing pathway. EMBO J. 25, 522–532.
8. Ma, E., MacRae, I. J., Kirsch, J. F. & Doudna, J. A.(2008). Autoinhibition of human dicer by its internalhelicase domain. J. Mol. Biol. 380, 237–243.
9. Wang, H. W., Noland, C., Siridechadilok, B., Taylor,D. W., Ma, E., Felderer, K. et al. (2009). Structuralinsights into RNA processing by the human RISC-loading complex. Nat. Struct. Mol. Biol. 16, 1148–1153.
10. Calabrese, J. M., Seila, A. C., Yeo, G. W. & Sharp, P. A.(2007). RNA sequence analysis defines Dicer's role inmouse embryonic stem cells. Proc. Natl Acad. Sci. USA,104, 18097–18102.
11. Chiang, H. R., Schoenfeld, L. W., Ruby, J. G., Auyeung,V. C., Spies, N., Baek, D. et al. (2010). MammalianmicroRNAs: experimental evaluation of novel andpreviously annotated genes. Genes Dev. 24, 992–1009.
12. Watanabe, T., Totoki, Y., Toyoda, A., Kaneda, M.,Kuramochi-Miyagawa, S., Obata, Y. et al. (2008).Endogenous siRNAs from naturally formed dsRNAsregulate transcripts in mouse oocytes. Nature, 453,539–543.
13. Meiri, E., Levy, A., Benjamin, H., Ben-David, M.,Cohen, L., Dov, A. et al. (2010). Discovery ofmicroRNAs and other small RNAs in solid tumors.Nucleic Acids Res. 38, 6234–6246.
14. Gregory, R. I., Chendrimada, T. P., Cooch, N. &Shiekhattar, R. (2005). HumanRISC couplesmicroRNAbiogenesis and posttranscriptional gene silencing. Cell,123, 631–640.
15. Maniataki, E. &Mourelatos, Z. (2005). A human, ATP-independent, RISC assembly machine fueled by pre-miRNA. Genes Dev. 19, 2979–2990.
16. Paroo, Z., Liu, Q. & Wang, X. (2007). Biochemicalmechanisms of the RNA-induced silencing complex.Cell Res. 17, 187–194.
17. Parrish, S. & Fire, A. (2001). Distinct roles for RDE-1and RDE-4 during RNA interference in Caenorhabditiselegans. RNA, 7, 1397–1402.
18. Forstemann, K., Tomari, Y., Du, T., Vagin, V. V.,Denli, A. M., Bratu, D. P. et al. (2005). NormalmicroRNA maturation and germ-line stem cell main-tenance requires Loquacious, a double-strandedRNA-binding domain protein. PLoS Biol. 3, e236.
19. Liu, Q., Rand, T. A., Kalidas, S., Du, F., Kim, H. E.,Smith, D. P. & Wang, X. (2003). R2D2, a bridge
between the initiation and effector steps of theDrosophila RNAi pathway. Science, 301, 1921–1925.
20. Tabara, H., Sarkissian, M., Kelly, W. G., Fleenor, J.,Grishok, A., Timmons, L. et al. (1999). The rde-1 gene,RNA interference, and transposon silencing in C.elegans. Cell, 99, 123–132.
21. Jiang, F., Ye, X., Liu, X., Fincher, L., McKearin, D. &Liu, Q. (2005). Dicer-1 and R3D1-L catalyze micro-RNA maturation in Drosophila. Genes Dev. 19,1674–1679.
22. Tomari, Y., Du, T. & Zamore, P. D. (2007). Sorting ofDrosophila small silencing RNAs. Cell, 130, 299–308.
23. Tomari, Y., Matranga, C., Haley, B., Martinez, N. &Zamore, P. D. (2004). A protein sensor for siRNAasymmetry. Science, 306, 1377–1380.
24. Chendrimada, T. P., Gregory, R. I., Kumaraswamy, E.,Norman, J., Cooch, N., Nishikura, K. & Shiekhattar, R.(2005). TRBP recruits the Dicer complex to Ago2 formicroRNA processing and gene silencing.Nature, 436,740–744.
25. Paroo, Z., Ye, X., Chen, S. & Liu, Q. (2009).Phosphorylation of the human microRNA-generatingcomplex mediates MAPK/Erk signaling. Cell, 139,112–122.
26. Kok, K. H., Ng, M. H., Ching, Y. P. & Jin, D. Y. (2007).Human TRBP and PACT directly interact with eachother and associate with Dicer to facilitate theproduction of small interfering RNA. J. Biol. Chem.282, 17649–17657.
27. Melo, S. A., Ropero, S., Moutinho, C., Aaltonen, L. A.,Yamamoto, H., Calin, G. A. et al. (2009). A TARBP2mutation in human cancer impairs microRNAprocessing and DICER1 function. Nat. Genet. 41,365–370.
28. Cifuentes, D., Xue, H., Taylor, D. W., Patnode, H.,Mishima, Y., Cheloufi, S. et al. (2010). A novelmiRNA processing pathway independent of Dicerrequires Argonaute2 catalytic activity. Science, 328,1694–1698.
29. Cheloufi, S., Dos Santos, C. O., Chong, M. M. &Hannon, G. J. (2010). A dicer-independent miRNAbiogenesis pathway that requires Ago catalysis.Nature, 465, 584–589.
30. Parker, G. S., Maity, T. S. & Bass, B. L. (2008). dsRNAbinding properties of RDE-4 and TRBP reflect theirdistinct roles in RNAi. J. Mol. Biol. 384, 967–979.
31. Daviet, L., Erard, M., Dorin, D., Duarte, M., Vaquero,C. & Gatignol, A. (2000). Analysis of a bindingdifference between the two dsRNA-binding domainsin TRBP reveals the modular function of a KR-helixmotif. Eur. J. Biochem. 267, 2419–2431.
32. Gatignol, A., Buckler, C. & Jeang, K. T. (1993).Relatedness of an RNA-binding motif in humanimmunodeficiency virus type 1 TAR RNA-bindingprotein TRBP to human P1/dsI kinase and DrosophilaStaufen. Mol. Cell Biol. 13, 2193–2202.
33. Daniels, S. M., Melendez-Pena, C. E., Scarborough,R. J., Daher, A., Christensen, H. S., El Far, M. et al.(2009). Characterization of the TRBP domain requiredfor dicer interaction and function in RNA interference.BMC Mol. Biol. 10, 38.
34. Ye, X., Paroo, Z. & Liu, Q. (2007). Functional anatomyof the DrosophilamicroRNA-generating enzyme. J. Biol.Chem. 282, 28373–28378.

402 Substrate-specific Dicing Kinetics
35. Parker, G. S., Eckert, D. M. & Bass, B. L. (2006). RDE-4preferentially binds long dsRNA and its dimerizationis necessary for cleavage of dsRNA to siRNA. RNA,12, 807–818.
36. MacRae, I. J., Ma, E., Zhou, M., Robinson, C. V. &Doudna, J. A. (2008). In vitro reconstitution of the
human RISC-loading complex. Proc. Natl Acad. Sci.USA, 105, 512–517.
37. Ferre-D'Amare, A. R. & Doudna, J. A. (1996). Use ofcis- and trans-ribozymes to remove 5′ and 3′ hetero-geneities frommilligrams of in vitro transcribed RNA.Nucleic Acids Res. 24, 977–978.