studies on isolated rat adrenal cells metabolism of hydroxylated sterols
TRANSCRIPT

Molecular and Cellular Endocrinology 3 (1975) 375-383. 0 North-Holland Pub]. Comp.
STUDIES ON ISOLATED RAT ADRENAL CELLS
METABOLISM OF HYDROXYLATED STEROLS*
H. E. FALKE, H. J. DEGENHART, G. J. A. ABELN and H. K. A. VISSER
Department of Pediatrics, Erasmus University and Academic Hospital Rotterdam and Sophia
Children’s Hospital and Neonatal Unit, Rotterdam, Netherlands
Received 9 April 1975 Accepted 20 June 1975
Corticosterone production by isolated rat adrenal cells in the absence of ACTH is stimulated by 20a-hydroxycholesterol, 22R-hydroxycholesterol and 25hydroxycholesterol. This effect is also seen at sub-maximal ACTH concentrations. Aminoglutethimide (AGJ) inhibits the stimulation caused by the three hydroxylated cholesterols. In the presence of ACTH, AGI also inhibits steroid production both under control conditions and in the presence of the sterols. The stimulation by 25hydroxycholesterol is dose-dependent, both in the presence and absence of a sub-maximal ACTH concentration. At maximal ACTH concentrations 25-hydroxycholesterol does not give additional stimulation. Cycloheximide has no effect on corticosterone production from 25-hydroxycholesterol whether ACTH is present or not. Our results indicate that 25-hydroxycholesterol is a good substrate for the study of the cholesterol side-chain cleaving system and the mechanism of action of ACTH at the level of the intact cell.
Keywords: isolated rat adrenal cells; cholesterol side-chain cleavage; ACTH mechanism of action; 25-hydroxycholesterol; 20a-hydroxycholesterol; 22R-hydroxy- cholesterol.
The adrenal cholesterol side-chain cleaving system is able to convert a wide
variety of sterols, including 5-cholesten-3P,20a-diol (20a-hydroxycholesterol),
5-cholesten-3b,22R-diol (22R-hydroxycholesterol) and 5-cholesten-3P,25-diol
(25-hydroxycholesterol), into steroids (Burstein and Gut, 1971). Usually the
metabolism of these sterols has been studied in vitro using isolated mito-
chondria or crude preparations of the cholesterol side-chain cleaving system
(Burstein and Gut, 1971; Jefcoate et al., 1974). Little is known about the
metabolism of these sterols in the intact adrenal cell. Hall and Young (1968)
*This work was supported by a grant from the Foundation for Medical Research, Sophia Children’s Hospital and Neonatal Unit, Rotterdam.

316 H. E. Falke et al.
observed a conversion of [7r~-~H]20a-hydroxycholesterol into cortisol in
bovine adrenal slices. Sharma (1973) found a production of deoxycorticosterone
and corticosterone from [7a-3H]20a-hydroxycholesterol in isolated rat adrenal
cells. In each case steroid production from 20a-hydroxycholesterol occurred
both in the absence and presence of ACTH.
This paper described the corticosterone production by isolated rat adrenal
cells incubated with 20a-hydroxycholesterol, 22R-hydroxycholesterol or 25
hydroxycholesterol and the effects of ACTH, aminoglutethimide (AGI) and
cycloheximide. Emphasis is put on 25-hydroxycholesterol in view of the fact
that side-chain cleavage of this sterol involves all the steps required for chol-
esterol side-chain cleavage. In the existing models for cholesterol side-chain
cleavage (Burstein and Gut, 1971; Kraaipoel et al., 1975) metabolism of 20a- hydroxycholesterol or 22R-hydroxycholesterol involves only part of the side-
chain cleaving system.
MATERIALS AND METHODS
Materials Incubations were carried out in a Krebs-Ringer bicarbonate buffer (pH 7.4)
containing 0.2% (w/v) glucose and 3 ‘A (w/v) albumin (KRBAG) (Bovine
serum albumin, Cohn Fraction V, Sigma Chemical Co.) or in a modified
Krebs-Ringer buffer with sodium bicarbonate replaced by 21.0 mM HEPES
(KRAHG) (Calbiochem). The use of HEPES buffers (Good et al., 1966) makes
it possible to do experiments in air and produces better pH control as compared
with bicarbonate buffers. Buffers were sterilized by filtration through Millipore
filters (HAWP 02500; pore size 0.45 uM).
Corticotrophin (ACTH) was a purified preparation of porcine ACTH
obtained from Organon Oss (CortrophineB). Aminoglutethimide phosphate
(AGI) was a generous gift from Ciba Co., Arnhem. Cycloheximide was obtained
from Boehringer. Cholesterol and 20a-hydroxycholesterol were obtained from
Sigma Chemical Co., 22R-hydroxycholesterol from Ikapharm and 25-hydroxy-
cholesterol from Steraloids. Solutions of these sterols were made up in ethanol.
All glass surfaces in contact with the cells or solutions containing ACTH
were siliconized (Siliclad, Clay Adams) and sterilized.
Incubations Incubations were carried out in round-bottomed glass centrifuge tubes
under 02-COZ (95-5 ‘A) for KRBAG or in air for KRHAG. The total incubation
volume was 1 ml. ACTH, AGI and cycloheximide were added in 0.1 ml buffer,

Adrenal metabolism of hydroxylated sterols 371
the sterols in 10 ~1 ethanol. After 2-h incubation at 37 “C. in a Dubnoff in-
cubator (100 rpm) the tubes were tightly capped, frozen rapidly and stored at
-20 “C. The number of cells per incubation was about 2 x 105.
Preparation of cell suspensions Suspensions of isolated rat adrenal cells were prepared as described pre-
viously (Falke et al., 1975). Adrenals of male Sprague-Dawley rats (200-250 g)
were used. No difference was observed between cells prepared in KRBAG or
KRHAG.
Analytic procedures Corticosterone was measured by fluorimetry as described previously (Falke
et al., 1975). The values obtained had to be corrected for the small amount of
fluorescence caused by 20a-hydroxycholesterol, 22R-hydroxycholesterol and
25hydroxycholesterol. The fluorescence of 1 ug of each of these sterols is
equivalent to 5.5 ng corticosterone at the maximal fluorescence wavelength of
the latter. The extent of the correction was based on the assumption that only a
small fraction of the sterols present was converted into steroids. That this was
a valid assumption was supported by measurement of the sterols by gas
chromatography. The method described by Kraaipoel et al. (1975) was used
with epicholesterol as an internal standard. Overall recovery of the sterols after
a 2-h incubation with isolated rat adrenal cells was 85-90x.
RESULTS
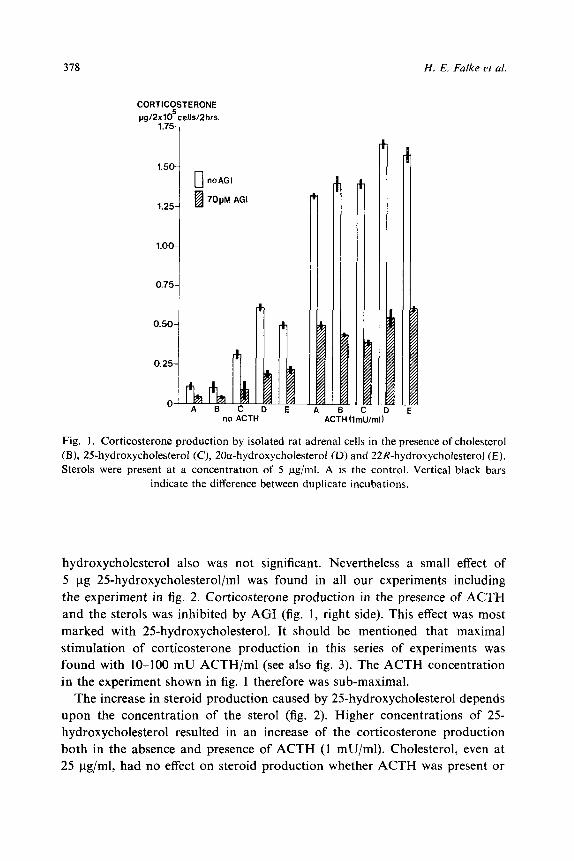
The production of corticosterone by isolated rat adrenal cells under several
conditions is shown in fig. 1. In the absence of ACTH, addition of 5ug/ml of
20a-hydroxycholesterol, 22R-hydroxycholesterol or 25-hydroxycholesterol caus-
ed a marked increase in corticosterone production (fig. 1, left side). 20a-
Hydroxycholesterol proved to be the best substrate, followed by 22R-hydroxy-
cholesterol and 25-hydroxycholesterol, respectively. Addition of cholesterol
failed to increase steroid production. AGI (70pM) inhibited corticosterone
production both in the control incubation and in the presence of each of the
sterols. Fig. 1 (right side) also shows the effect of the sterols on corticosterone
production in the presence of 1 mU ACTH/ml. Significant stimulation under
this condition was found with 5 ug/ml of 20a-hydroxycholesterol and 22R-
hydroxycholesterol. Very small effects were seen with cholesterol and 25-hydro-
xycholesterol. The effect of cholesterol surely is not significant and was not
found in other experiments, like the one shown in fig. 2. The effect of 25-
378 H. E. Falke et al.
CORTICOSTERONE
(1g/2x105cells/2hrs. 1.75,
no ACTH
0 noAGl
70pM AGI
‘H (lmU/n
Fig. 1. Corticosterone production by isolated rat adrenal cells in the presence of cholesterol (B), 2ShydroxycholesteroI (C), 20a-hydroxycholesterol (D) and 22R-hydroxycholesterol (E). Sterols were present at a concentration of 5 pg/ml. A is the control. Vertical black bars
indicate the difference between duplicate incubations.
hydroxycholesterol also was not significant. Nevertheless a small effect of
5 ug 2Shydroxycholesterol/ml was found in all our experiments including
the experiment in fig. 2. Corticosterone production in the presence of ACTH
and the sterols was inhibited by AGI (fig. 1, right side). This effect was most
marked with 2Shydroxycholesterol. It should be mentioned that maximal
stimulation of corticosterone production in this series of experiments was
found with lo-100 mU ACTH/ml (see also fig. 3). The ACTH concentration
in the experiment shown in fig. 1 therefore was sub-maximal.
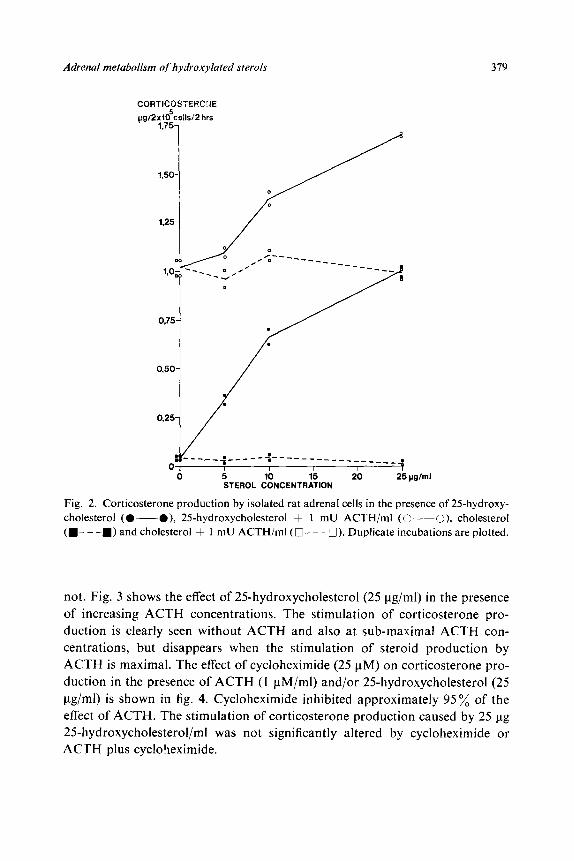
The increase in steroid production caused by 25-hydroxycholesterol depends
upon the concentration of the sterol (fig. 2). Higher concentrations of 25-
hydroxycholesterol resulted in an increase of the corticosterone production
both in the absence and presence of ACTH (1 mu/ml). Cholesterol, even at
25 ug/ml, had no effect on steroid production whether ACTH was present or
Adrenal metabolism of hydroxylated sterols 379
1,25
l&l I-
0,75 ,-
0,x l-
0,25 b-
.I _
0 -r 0
I_____ .-____:__--- # -----es_____
I I I 1 5 STEROL d:NCENTR::ION
20 25 pglml
Fig. 2. Corticosterone production by isolated rat adrenal cells in the presence of 25hydroxy- cholesterol (O---O), 2Shydroxycholesterol + 1 mU ACTH/ml (C--O), cholesterol (m- - -E) and cholesterol + 1 mU ACTH/ml ( q - - -3). Duplicate incubations are plotted.
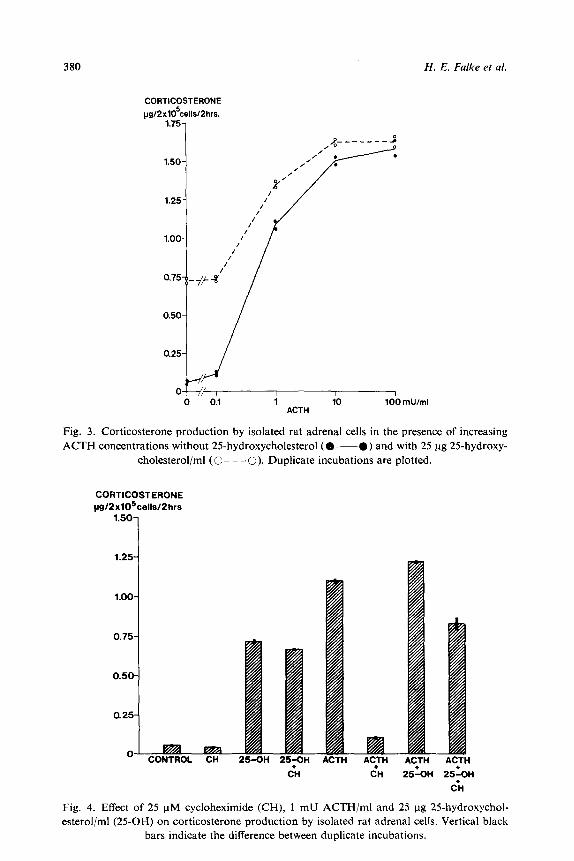
not. Fig. 3 shows the effect of 25-hydroxycholesterol (25 pg/ml) in the presence
of increasing ACTH concentrations. The stimulation of corticosterone pro-
duction is clearly seen without ACTH and also at sub-maximal ACTH con-
centrations, but disappears when the stimulation of steroid production by
ACTH is maximal. The effect of cycloheximide (25 PM) on corticosterone pro-
duction in the presence of ACTH (1 FM/ml) and/or 25-hydroxycholesterol (25
pg/ml) is shown in fig. 4. Cycloheximide inhibited approximately 95% of the
effect of ACTH. The stimulation of corticosterone production caused by 25 ug
25-hydroxycholesterol/ml was not significantly altered by cycloheximide or
ACTH plus cycloheximide.
380 H. E. Falke et al.
CORTICOSTERONE
~g/2x105cells12hrs. 1.751
Fig. 3. Corticosterone production by isolated rat adrenal cells in the presence of increasing ACTH concentrations without 25hydroxycholesterol (O--O) and with 25 ug 25-hydroxy-
cholesterol/ml (0- - -0). Duplicate incubations are plotted.
CORTICOSTERONE ~9/2x105cells/2hrs
ISO-
1.25-
lsm-
0.75-
0.50-
0.25-
0 CONTROL CH 25-OH 25PH ACTH ACTH ACTH ACTH
CH EH 25+OH 25+OH
CH
Fig. 4. Effect of 25 uM cycloheximide (CH), 1 mU ACTH/ml and 25 ug 25-hydroxychol- esterol/ml (25OH) on corticosterone production by isolated rat adrenal cells. Vertical black
bars indicate the difference between duplicate incubations.

Adrenal metabolism of hydroxylated sterols 381
DISCUSSION
Corticosterone production by isolated rat adrenal cells is stimulated by
20a-hydroxycholesterol, 22R-hydroxycholesterol and 25-hydroxycholesterol.
In the presence of a sub-maximal ACTH concentration a similar stimulation
was found. This indicates that these sterols are good substrates for the cholester-
ol side-chain cleaving system in the intact adrenal cell. Hall and Young (1968)
and Sharma (1973), working with bovine adrenal slices and isolated rat adrenal
cells, respectively, previously recovered 3H-labeled C, i-steroids starting
with [7a- 3H]20a-hydroxycholesterol as a substrate. The synthesis of corti-
costerone from these sterols by isolated rat adrenal cells in the absence of
ACTH is compatible with the current hypothesis of the mechanism of action
of ACTH. According to this hypothesis ACTH exerts its effect prior to the
cholesterol side-chain cleaving system (Garren et al., 1971; Schulster, 1974).
The reason for the absence of a stimulating effect of cholesterol, a phenomenon
also reported by Sharma (1973), is not clear. Maybe, under our conditions,
exogeneous cholesterol cannot reach the side-chain cleaving system fast
enough to stimulate steroid synthesis. In addition Sharma (1973) found that
cholesterol inhibited the effect of ACTH on corticosterone production. The
absence of such an effect in our experiments may be due to the difference
in the cholesterol concentration used. The highest cholesterol concentration
in our experiments was 25 ug/ml (= 0.065 mM) while Sharma used 192 ug/ml
(= 0.5 mM). Formation of mono-molecular layers of cholesterol on the cell
surface might have been responsible for the inhibition caused by the high
cholesterol concentration used by Sharma.
In the classical scheme of cholesterol side-chain cleavage 20a-hydroxy-
cholesterol and 22R-hydroxycholesterol are considered as intermediates. In the
recent model proposed by Kraaipoel et al. (1975) however, these two sterols
are merely by-products. The calculations of Burstein and Gut (1971) also
indicate that the largest part of the steroid production from cholesterol does
not involve either sterol as an intermediate. Both in the model of Kraaipoel
et al. (1975) and in the model of Burstein and Gut (1971) the metabolism of
these two sterols involves only part of the cholesterol side-chain cleaving
system. Therefore they are of limited use for the study of the cholesterol side-
chain cleaving system. This is the reason why most of our work was carried out
only with 25-hydroxycholesterol. In our opinion 25-hydroxycholesterol is a
good substrate for the study of the cholesterol side-chain cleaving system.
The occurence of 25-hydroxycholesterol in mammals has been repeatedly
described. It is present among other sterols in human cord blood (Eberlein,
1965) human aorta (Van Lier and Smith, 1967) and brain (Van Lier and Smith,

382 H. E. Falke et al.
1969). Formation of 25-hydroxycholesterol from cholesterol by a 25-hydroxy-
lase was found in rat liver (Bjiirkhem and Gustafsson, 1974). A 25-hydroxylase
for cholecalciferol is also present in mammalian liver (Norman and Henry,
1974). Conversion of 25-hydroxycholesterol to pregnenolone by the cholesterol
side-chain cleaving system was previously found by Burstein and Gut (1971)
in acetone powders of bovine adrenal mitochondria and by Degenhart et al.
(1974) and Jefcoate et al. (1974) in bovine and rat adrenal mitochondria.
Evidence exists that side-chain cleaving of cholesterol and 25-hydroxycholesterol
takes place by the same enzyme system. Metabolism of 25-hydroxycholesterol
is inhibited by AGI an inhibitor of the first step of cholesterol side-chain cleavage
(cholesterol+ 5,2@(22)-cholestadien-3P-01) (Degenhart et al., 1974). This also
indicates that all the steps of cholesterol side-chain cleaving are involved in the
metabolism of 25hydroxycholesterol. The competition between cholesterol
and 25-hydroxycholesterol (Jefccate et al., 1974) is further evidence for con-
version of both sterols by the same enzyme system. The inhibition of the meta-
bolism of 25-hydroxycholesterol by AGI (fig. 1) indicates that also in the
intact cell this sterol is converted by the cholesterol side-chain cleaving system.
The absence of additive stimulation by 25-hydroxycholesterol at maximal
ACTH concentrations points in the same direction. It seems that in isolated
rat adrenal cells at maximal ACTH concentrations the substrate flux to the
side-chain cleaving system matches the full capacity of this system; therefore
no extra effect of 25-hydroxy-cholesterol can be seen. The results on which
Jefcoate et al. (1974) base their assumption that the cytochrome P-450 for
side-chain cleavage of cholesterol is present in three different forms may just
reflect different binding properties of cholesterol and 25-hydroxycholesterol.
ACTH pretreatment in vivo has no effect on the total cholesterol side-chain
cleaving activity (Koritz and Kumar, 1970) nor on the K,, or I’,,,,, of the
enzyme system (Bell and Harding, 1974) in rat adrenal mitochondria. ACTH
therefore appears to have no direct effect on the cholesterol side-chain cleaving
system. Hcwever, Sharma (1973) found that ACTH, but not N6,02-dibutyryl-
adenosine 3’,5’-monophosphate stimulated the conversion of [7a-3H]20a-
hydroxycholesterol in isolated rat adrenal cells. This is in contrast with the
results of Hall and YotTng (1968) who found that ACTH s!ightly inhibited the
conversion of [7a-3H]20a-hydroxycholesterol in bovine adrenal slices. Neither
Hall and Young (1968) nor Sharma (1973) measured specific activities of the
compounds involved which makes exact interpretation of their results difficult.
Our results are consistent with the absence of a direct effect of ACTH on the
side-chain cleaving system. The metabolism of 25-hydroxycholesterol in the
absence of ACTH indicates that the side-chain cleaving system is invariably
present in an active form. At sub-maximal ACTH concentrations only part of

Adrenal metabolism of hydroxylated sterols 383
the activity is needed for side-chain cleavage of endogeneous cholesterol and
an additive effect of 25-hydroxycholesterol can be observed. Only at maximal
ACTH concentrations the full capacity of the side-chain cleaving system is
used for endogeneous cholesterol and 25-hydroxycholesterol cannot give
extra steroid production. Evidence that ACTH has no direct effect on the side-
chain cleaving system comes from our results with cycloheximide. Both in the
absence and in the presence of ACTH cycloheximide had no effect on the
amount of the stimulation caused by 25-hydroxycholesterol (fig. 4). The
failure of cycloheximide to influence the effect of 25-hydroxycholesterol is
further proof for the localization of the effect of ACTH on events prior to the
side-chain cleaving system.
ACKNOWLEDGEMENTS
Mrs. Mary Amesz-Heinrich, Mr. A. A. M. Kempers and Dr. D. A. Price are
gratefully acknowledged for their help with the preparation of the manuscript.
REFERENCES
Bell, J. J. and Harding, B. W. (1974) Biochim. Biophys. Acta 348,285. Bjorkhem, I. and Gustafsson, J. A. (1974) J. Biol. Chem. 249, 2528. Burstein, S. and Gut, M. (1971) Recent Prog. Horm. Res. 27, 303. Degenhart, H. J., Kraaipoel, R. J., Falke, H. E., Leferink, J. G., van Beek, V., de Leeuw-Boon,
H., Abeln, G. and Visser, H. K. A. (1974) J. Steroid Biochem. 5, 308. Eberlein, W. R. (1965) J. Clin. Endocrinol. Metab. 25, 1101. Falke, H. E., Degenhart, H. J., Abeln, G. J. A., Visser, H. K. A. and Croughs, R. J. M. (1975)
Acta Endocrinol. 78, 110. Garren, L. D., Gill, G. N., Masui, H. and Walton, M. (1971) Recent Prog. Horm. Res. 27,433. Good, N. E., Winget, G. D., Winter, W., Connolly, T. N., Tzawa, S. and Singh, R. M. M.
(1966) Biochemistry 5, 467. Hall, P. F. and Young, D. G. (1968) Endocrinology 82, 559. Jefcoate, C. R., Simpson, E. R. and Boyd, G. S. (1974) Eur. J. Biochem. 42, 539. Kraaipoel, R. J., Degenhart, H. J., Leferink, J. G., van Beek, V., de Leeuw-Boon, H. and
Visser, H. K. A. (1975) FEBS Lett. 50, 204. Koritz, S. B. and Kumar, A. M. (1970) J. Biol. Chem. 245, 152. Norman, A. W. and Henry, H. (1974) Recent Prog. Horm. Res. 30,431. Sharma, R. K. (1973) J. Biol. Chern. 248, 5473. Schulster, D. (1974) In: Advances in Steroid Biochemistry and Pharmacology, Vol. 4, Eds.:
M. H. Briggs and G. A. Christie (Academic Press, London, New York) p. 233. Van Lier, J. E. and Smith, L. L. (1967) Biochemistry 6, 3269. Van Lier, J. E. and Smith, L. L. (1969) Tex. Rep. Biol. Med. 27, 167.