structure of the integral membrane protein caax …people.virginia.edu/~jz3x/journal_club/pryor et...
TRANSCRIPT

DOI: 10.1126/science.1232048, 1600 (2013);339 Science
et al.Edward E. Pryor Jr.Structure of the Integral Membrane Protein CAAX Protease Ste24p
This copy is for your personal, non-commercial use only.
clicking here.colleagues, clients, or customers by , you can order high-quality copies for yourIf you wish to distribute this article to others
here.following the guidelines
can be obtained byPermission to republish or repurpose articles or portions of articles
): April 9, 2013 www.sciencemag.org (this information is current as of
The following resources related to this article are available online at
http://www.sciencemag.org/content/339/6127/1600.full.htmlversion of this article at:
including high-resolution figures, can be found in the onlineUpdated information and services,
http://www.sciencemag.org/content/suppl/2013/03/28/339.6127.1600.DC1.html can be found at: Supporting Online Material
http://www.sciencemag.org/content/339/6127/1600.full.html#relatedfound at:
can berelated to this article A list of selected additional articles on the Science Web sites
http://www.sciencemag.org/content/339/6127/1600.full.html#ref-list-1, 22 of which can be accessed free:cites 57 articlesThis article
http://www.sciencemag.org/content/339/6127/1600.full.html#related-urls1 articles hosted by HighWire Press; see:cited by This article has been
http://www.sciencemag.org/cgi/collection/biochemBiochemistry
subject collections:This article appears in the following
registered trademark of AAAS. is aScience2013 by the American Association for the Advancement of Science; all rights reserved. The title
CopyrightAmerican Association for the Advancement of Science, 1200 New York Avenue NW, Washington, DC 20005. (print ISSN 0036-8075; online ISSN 1095-9203) is published weekly, except the last week in December, by theScience
on
Apr
il 9,
201
3w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

Dungey cycle during substorms are given by the ECPCmodel (18).
10. K. Oksavik, V. L. Barth, J. Moen, M. Lester, J. Geophys.Res. 115, A12308 (2010).
11. K. Hosokawa et al., J. Geophys. Res. 114, A04318 (2009).12. A. G. Wood, S. E. Pryse, J. Moen, Ann. Geophys. 27, 3923
(2009).13. M. Lockwood et al., Ann. Geophys. 23, 3513 (2005).14. M. Lockwood et al., J. Geophys. Res. 111, A02306
(2006).15. J. C. Foster et al., J. Geophys. Res. 110, A09S31 (2005).16. J. J. Sojka et al., Geophys. Res. Lett. 20, 1783 (1993).17. G. Crowley et al., J. Geophys. Res. 105, 5215 (2000).18. S. W. H. Cowley, M. Lockwood, Ann. Geophys. 10,
103 (1992).19. G. S. Bust, G. Crowley, J. Geophys. Res. 112, A05307
(2007).20. TEC is the number of electrons present along a
transionospheric path between two points, with units ofelectrons per square meter, where 1016 electrons/m2 = 1TEC unit (TECU) (21). TEC is important in determiningthe scintillation and group delay of radio waves passingthrough the ionosphere. It is determined by observing
carrier phase delays of received radio signals transmittedfrom satellites above the ionosphere; here, we use GPSsatellites. TEC is strongly affected by solar activity.
21. A. J. Coster, J. C. Foster, P. J. Erickson, GPS World 14,42 (2003).
22. C. Stolle et al., Ann. Geophys. 24, 107 (2006).23. J. M. Ruohoniemi, K. B. Baker, J. Geophys. Res. 103,
(A9), 20797 (1998).24. E. G. Thomas et al., J. Geophys. Res., (2013);
10.1002/jgra.5011625. S. K. Morley, M. Lockwood, Ann. Geophys. 24, 961 (2006).26. M. Lockwood et al., Ann. Geophys. 23, 3495 (2005).27. M. Lockwood, M. Hairston, I. D. Finch, A. P. Rouillard,
J. Geophys. Res. 114, A01210 (2009).
Acknowledgments: This work in China was supported by theNational Basic Research Program (grant 2012CB825603),the National Natural Science Foundation (grants 41274149,41104091, 41031064, and 40890164), and the OceanPublic Welfare Scientific Research Project, State OceanicAdministration (201005017). The Norwegian contribution wassupported by the Research Council of Norway. SuperDARNis a collection of radars funded by national scientific funding
agencies in Australia, Canada, China, France, Japan, SouthAfrica, the United Kingdom, and the United States. TheVirginia Tech authors acknowledge the support of NSF awardsAGS-0946900 and AGS-0838219 and a graduate researchfellowship from the Virginia Space Grant Consortium. The GPSTEC acquisition effort is led by A. J. Coster at MIT HaystackObservatory. The GPS TEC and SuperDARN data are availableon the public database http://sd-software.ece.vt.edu/tiki/tiki-index.php?page=DaViT TEC. We also thank the NASA OMNIWeb formaking available the solar wind/IMF data (http://omniweb.gsfc.nasa.gov/html/sc_merge_data1.html) and World Data CenterC1, Kyoto, for the AE indices (http://wdc.kugi.kyoto-u.ac.jp/wdc/Sec3.html).
Supplementary Materialswww.sciencemag.org/cgi/content/full/339/6127/1597/DC1Supplementary TextFigs. S1 to S4Movie S1References
15 October 2012; accepted 31 January 201310.1126/science.1231487
Structure of the Integral MembraneProtein CAAX Protease Ste24pEdward E. Pryor Jr.,1,2* Peter S. Horanyi,1,2* Kathleen M. Clark,1,3* Nadia Fedoriw,1,3*Sara M. Connelly,1,3 Mary Koszelak-Rosenblum,1,4 Guangyu Zhu,1,4 Michael G. Malkowski,1,4,5
Michael C. Wiener,1,2† Mark E. Dumont1,3,6†
Posttranslational lipidation provides critical modulation of the functions of some proteins.Isoprenoids (i.e., farnesyl or geranylgeranyl groups) are attached to cysteine residues in proteinscontaining C-terminal CAAX sequence motifs (where A is an aliphatic residue and X is any residue).Isoprenylation is followed by cleavage of the AAX amino acid residues and, in some cases, byadditional proteolytic cuts. We determined the crystal structure of the CAAX protease Ste24p, a zincmetalloprotease catalyzing two proteolytic steps in the maturation of yeast mating pheromonea-factor. The Ste24p core structure is a ring of seven transmembrane helices enclosing avoluminous cavity containing the active site and substrate-binding groove. The cavity is accessibleto the external milieu by means of gaps between splayed transmembrane helices. Wehypothesize that cleavage proceeds by means of a processive mechanism of substrate insertion,translocation, and ejection.
Isoprenoid groups are conjugated to proteinsby means of cysteine residues of CAAX ac-ceptor sequences in which the cysteine attach-
ment site is followed by two aliphatic amino acidresidues and one unspecified residue at the pro-tein C terminus. Isoprenylation is generally ac-companied by two subsequent processing steps,proteolytic cleavage of the AAX residues andcarboxymethylation of the newly exposed car-bonyl group of the modified cysteine residue
(fig. S1). Some isoprenylated proteins also un-dergo additional proteolytic processing, includ-ing an additional cleavage by the same proteasethat initially removes the AAX residues. At leasttwo classes of enzymes are responsible for thecleavage of isoprenylated proteins and peptides.One of these is the Ras-converting enzyme (Rce)family of type II prenyl proteases, responsiblefor proteolytic processing of signal-transducingproteins including Ras (1, 2) and the Gg subunitsof heterotrimeric GTP-binding protein (G protein)complexes (3). The other is the Ste24p family oftype I prenyl proteases, first identified in yeast onthe basis of its role in maturation of the matingpheromone a-factor (4–6). Extensive characteri-zation of the role of Ste24p in a-factor processinghas been conducted in the yeast system (7). Ste24pis localized to the endoplasmic reticulum mem-brane. Its proteolytic activity requires zinc, con-sistent with the fact that Ste24p contains the zincmetalloprotease signature motif HEXXH (8, 9).
A human ortholog of Ste24p, zinc metallo-protease STE24 (ZMPSTE24), can complement
the full function of yeast Ste24p (6). The onlyknown substrate for ZMPSTE24 is prelamin A,the precursor to the nuclear intermediate filamentprotein lamin A. Lamins provide mechanical sta-bility to the nuclear envelope, function as scaf-folds for localization of other proteins and forcytoskeletal attachment, regulate chromatin, andare implicated in transcription and DNA repairand replication (10).Mutations in either ZMPSTE24or the processing site of prelaminA are associatedwith a spectrum of premature-aging diseases re-ferred to as progeria (11). The severity of differ-ent forms of progeria is reported to be correlatedwith extent of loss of ZMPSTE24 activity (12).Also, ZMPSTE24 (and Saccharomyces cerevisiaeSte24p) are inhibited by antiviral drugs designedto target the HIV aspartyl protease, and this off-target interaction may give rise to some of thesevere side effects of these drugs (13, 14).
Ste24p from Saccharomyces cerevisiae(ScSte24p) has been overexpressed previouslyin S. cerevisiae cells and purified (9, 15). To iden-tify forms of the protein with enhanced stabilityand suitability for crystallization, we cloned andpurified orthologs from nine yeast species close-ly related to S. cerevisiae. S. mikatae Ste24p(SmSte24p) is 96% identical to ScSte24p andis 37% identical to H. sapiens ZMPSTE24(fig. S2). Purified SmSte24p is enzymatically ac-tive (fig. S3), and we obtained crystals of thisprotein that diffracted anisotropically to 3.1 Åresolution (and isotropically to 3.9 Å resolution).After obtaining a native data set and proceedingto make selenomethionine-containing SmSte24p(16), we discovered that the Structural GenomicsConsortium (SGC) had solved the structure ofhuman ZMPSTE24. Because of SGC’s openaccess policy, the coordinates were deposited inthe Protein Data Bank (PDB) before publication[PDB: 4AW6 (17)]. Thus, we solved the structureof SmSte24p by a combination of molecular re-placement (MR) and single-wavelength anoma-lous diffraction (SAD) of the bound catalytic zincatoms.
1Membrane Protein Structural Biology Consortium, USA. 2De-partment of Molecular Physiology and Biological Physics, Box800886, University of Virginia, Charlottesville, VA 22908–0886, USA. 3Department of Pediatrics, Box 703, University ofRochester School of Medicine and Dentistry, Rochester, NY14642, USA. 4Hauptman-Woodward Institute, 700 EllicottStreet, Buffalo, NY 14203, USA. 5Department of StructuralBiology, State University of New York at Buffalo, 700 EllicottStreet, Buffalo, NY 14203, USA. 6Department of Biochemistryand Biophysics, Box 712, University of Rochester School ofMedicine and Dentistry, Rochester, NY 14642, USA.
*These authors contributed equally to this work.†Corresponding author. E-mail: [email protected](M.C.W.); [email protected] (M.E.D.)
29 MARCH 2013 VOL 339 SCIENCE www.sciencemag.org1600
REPORTS
on
Apr
il 9,
201
3w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from
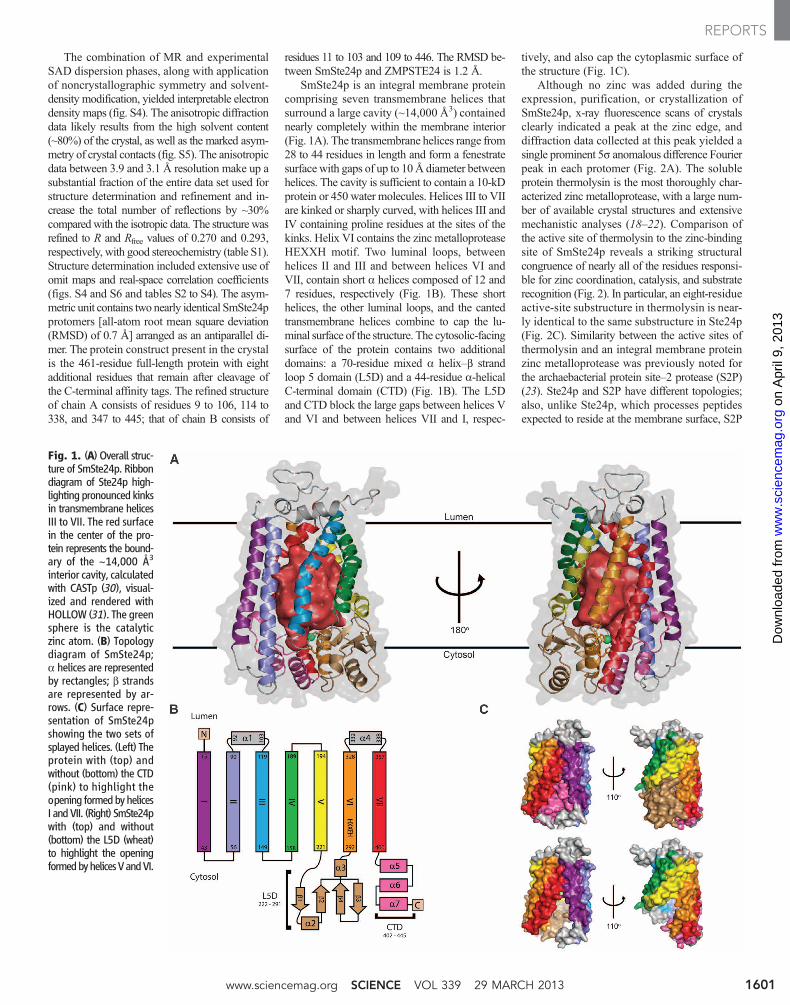
The combination of MR and experimentalSAD dispersion phases, along with applicationof noncrystallographic symmetry and solvent-density modification, yielded interpretable electrondensity maps (fig. S4). The anisotropic diffractiondata likely results from the high solvent content(~80%) of the crystal, as well as the marked asym-metry of crystal contacts (fig. S5). The anisotropicdata between 3.9 and 3.1 Å resolution make up asubstantial fraction of the entire data set used forstructure determination and refinement and in-crease the total number of reflections by ~30%comparedwith the isotropic data. The structure wasrefined to R and Rfree values of 0.270 and 0.293,respectively, with good stereochemistry (table S1).Structure determination included extensive use ofomit maps and real-space correlation coefficients(figs. S4 and S6 and tables S2 to S4). The asym-metric unit contains two nearly identical SmSte24pprotomers [all-atom root mean square deviation(RMSD) of 0.7 Å] arranged as an antiparallel di-mer. The protein construct present in the crystalis the 461-residue full-length protein with eightadditional residues that remain after cleavage ofthe C-terminal affinity tags. The refined structureof chain A consists of residues 9 to 106, 114 to338, and 347 to 445; that of chain B consists of
residues 11 to 103 and 109 to 446. The RMSD be-tween SmSte24p and ZMPSTE24 is 1.2 Å.
SmSte24p is an integral membrane proteincomprising seven transmembrane helices thatsurround a large cavity (~14,000 Å3) containednearly completely within the membrane interior(Fig. 1A). The transmembrane helices range from28 to 44 residues in length and form a fenestratesurface with gaps of up to 10Å diameter betweenhelices. The cavity is sufficient to contain a 10-kDprotein or 450 water molecules. Helices III to VIIare kinked or sharply curved, with helices III andIV containing proline residues at the sites of thekinks. Helix VI contains the zinc metalloproteaseHEXXH motif. Two luminal loops, betweenhelices II and III and between helices VI andVII, contain short a helices composed of 12 and7 residues, respectively (Fig. 1B). These shorthelices, the other luminal loops, and the cantedtransmembrane helices combine to cap the lu-minal surface of the structure. The cytosolic-facingsurface of the protein contains two additionaldomains: a 70-residue mixed a helix–b strandloop 5 domain (L5D) and a 44-residue a-helicalC-terminal domain (CTD) (Fig. 1B). The L5Dand CTD block the large gaps between helices Vand VI and between helices VII and I, respec-
tively, and also cap the cytoplasmic surface ofthe structure (Fig. 1C).
Although no zinc was added during theexpression, purification, or crystallization ofSmSte24p, x-ray fluorescence scans of crystalsclearly indicated a peak at the zinc edge, anddiffraction data collected at this peak yielded asingle prominent 5s anomalous difference Fourierpeak in each protomer (Fig. 2A). The solubleprotein thermolysin is the most thoroughly char-acterized zinc metalloprotease, with a large num-ber of available crystal structures and extensivemechanistic analyses (18–22). Comparison ofthe active site of thermolysin to the zinc-bindingsite of SmSte24p reveals a striking structuralcongruence of nearly all of the residues responsi-ble for zinc coordination, catalysis, and substraterecognition (Fig. 2). In particular, an eight-residueactive-site substructure in thermolysin is near-ly identical to the same substructure in Ste24p(Fig. 2C). Similarity between the active sites ofthermolysin and an integral membrane proteinzinc metalloprotease was previously noted forthe archaebacterial protein site–2 protease (S2P)(23). Ste24p and S2P have different topologies;also, unlike Ste24p, which processes peptidesexpected to reside at the membrane surface, S2P
Fig. 1. (A) Overall struc-ture of SmSte24p. Ribbondiagram of Ste24p high-lighting pronounced kinksin transmembrane helicesIII to VII. The red surfacein the center of the pro-tein represents the bound-ary of the ~14,000 Å3
interior cavity, calculatedwith CASTp (30), visual-ized and rendered withHOLLOW (31). The greensphere is the catalyticzinc atom. (B) Topologydiagram of SmSte24p;a helices are representedby rectangles; b strandsare represented by ar-rows. (C) Surface repre-sentation of SmSte24pshowing the two sets ofsplayed helices. (Left) Theprotein with (top) andwithout (bottom) the CTD(pink) to highlight theopening formedbyhelicesI andVII. (Right) SmSte24pwith (top) and without(bottom) the L5D (wheat)to highlight the openingformedbyhelicesVandVI.
www.sciencemag.org SCIENCE VOL 339 29 MARCH 2013 1601
REPORTS
on
Apr
il 9,
201
3w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

catalyzes intramembrane proteolysis of trans-membrane proteins (24–26).
The interactions between Ste24p and itssubstrates that determine the specificity of theinteraction remain poorly experimentally char-acterized; to date, only mutations immediatelyC-terminal to each of the cleavage sites in thea-factor precursor have been shown to affectcleavage efficiencies in in vivo studies (27, 28).To characterize a likely substrate-binding sitein SmSte24p, we first produced a compositemodel of thermolysin with an occupied activesite based on three structures of thermolysin incomplex with substrates and inhibitors (Fig. 3A).We then generated an electron density envelopecorresponding to the composite contents of thethermolysin active site (Fig. 3B) and mapped thisonto the structure of SmSte24p by means of thesame transformation matrix used to calculate theRMSD between the two enzymes’ active-site resi-dues. Finally, we fitted a farnesylated peptide frag-ment of the a-factor precursor (including its scissilebond) to this density (Fig. 3, C and D). The mod-eled position of the substrate places the farnesylattachment site at a position adjacent to the inter-nal cavity that affords ample volume for it to adopta variety of conformations (Fig. 3, C and D). How-ever, the modest resolution of our current structureprecludes a detailed interpretation of this modeling.
The substrate-binding groove, formed by L5Dand CTD, is contained at the cytoplasmic face ofthe internal transmembrane cavity of SmSte24p.The groove, nearly coplanar with the membranesurface and ~40 Å in length, traverses the cavity(Fig. 4, A and B). Each end of the groove is lo-cated between one of the pairs of splayed trans-membrane helices (Fig. 1C). The high degree ofsimilarity between the active sites of SmSte24pand thermolysin strongly suggests that the po-larity (N-to-C direction) of peptide-protein sub-strates with respect to the active site is identicalin the two enzymes. Thus, SmSte24p substrates
can be expected to be oriented in the groove withtheir C termini pointing toward the portal betweenhelices VII and I of Ste24p and their N termini
pointing toward the portal between helices VandVI (Fig. 4C). The localization of residues involvedin enzymatic activity to the cytosolic-proximal
Fig. 2. Analysis of the SmSte24p active site. (A) The active site of SmSte24pcontains the HEXXH motif in helix VI (H297–H301), the catalytic residue(E390), and substrate-coordinating residues N263, A264, H434, and R440.Mutation of H297, E298, or H301 diminishes Ste24p function (4, 32). Thecatalytic zinc atom is represented by anomalous electron difference den-
sity contoured at 5s. (B) The active site of thermolysin (PDB:1LND)showing the HEXXH-motif (H142–H146), the catalytic residue (E166),and the substrate-coordinating residues N112, A113, R203, and H231.(C) Overlay of thermolysin and SmSte24p active sites showing their sim-ilarity (RMSD of overlaid residues: 0.5 Å).
Fig. 3. Modeling of a-factor precursor binding to SmSte24. (A) Surface representation of the substrate-binding pocket of thermolysin with three different substrate/inhibitor molecules overlaid. (B) Electrondensity envelope calculated from (A). (C) Electron density envelope from (B) mapped onto the SmSte24pstructure with an a-factor precursor peptide fragment (ACVIA) positioned within the density. (D) Alternateview of the modeled a-factor precursor peptide with attached farnesyl chain. The white area indicated bythe asterisk identifies fenestration between helices V and VI.
29 MARCH 2013 VOL 339 SCIENCE www.sciencemag.org1602
REPORTS
on
Apr
il 9,
201
3w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

portion of the cavity is reflected in higher sequenceidentity and conservation between SmSte24pand ZMPSTE24 in this part of the cavity (fig. S7).
These observations, together with previous-ly reported functional similarity between Ste24pand ZMPSTE24, particularly the ability ofZMPSTE24 to fully complement a deletion ofSTE24 in processing yeast a-factor (6), suggesta possible pathway for substrate processing byCAAX proteases. In considering a small sub-strate such as the a-factor precursor (36 resi-dues in length), substrate binding could occurby entry of either its C or N terminus into thebinding groove. However, prelamin A contains afolded domain of 647 residues N-terminal to bothZMPSTE24 cleavage sites (fig. S1A). Because itis unlikely that such a large domain could gain
access to the active site, the most likely path-way for entry of this substrate (and, by inference,all the substrates of this type of enzyme) into theactive site is by means of insertion of its C ter-minus (Fig. 4D). The second proteolytic cleav-ages of the a-factor precursor and prelamin Acould occur through processive further exten-sion of the substrate C terminus into the substrategroove or cavity after the first cleavage (Fig. 4E).However, the presence of the large folded do-main of ZMPSTE24 dictates that withdrawal ofthe final N-terminal cleavage products wouldhave to occur by means of reversal of the initialroute of entry. Because L5D and CTD block thegaps between helices V and VI and helices VIIand I, respectively (Fig. 1C), movement of oneor both domains may be required for substrate
processing. However, relatively modest confor-mational changes or movement may suffice toenlarge the fenestrations that already exist inthese structural regions (Fig. 3D). Note that mostof the missense mutations in human ZMPSTE24that result in progeria map to the putative en-trance and exit portals (fig. S8).
A possible role for the voluminous internalcavity is suggested by the consumption of awater molecule in each turnover of proteolyticenzymes. Although biological membranes arewater-permeable, the dwell-time of water in themembrane interior is low, and maximizing theeffective concentration of water may promotethe proteolytic cleavage reaction by mass action.Thus, the cavity may serve as a water reservoir tofacilitate CAAX protease substrate processing.
Fig. 4. Proposed Ste24p mechanism. (A) Cutaway view of the SmSte24pactive site from the luminal surface. The modeled a-factor precursor peptide(from Fig. 3, C and D) is shown in blue. (B) Surface representation of SmSte24ppositioned into a lipid bilayer model obtained from a published moleculardynamics simulation (33). Blue spheres denote gaps between helices V and VIand between helices VII and I that lead to the substrate-binding groove andinternal active site contained within the internal transmembrane cavity (Fig. 1A).
(C) Schematic representation, to scale, of the continuous groove that traversesthe cavity [determined with MOLE 2.0 (34)]. The ~40 Å tunnel can accom-modate ~13 residues of substrate. (D) Insertion of the a-factor precursor Cterminus into the entrance portal formed by helices V and VI. Cleavage bySmSte24p occurs at the CAAXmotif indicated by the red star. (E) Subsequentsubstrate translocation, egress through the VII to I portal, and positioning ofthe second cleavage site.
www.sciencemag.org SCIENCE VOL 339 29 MARCH 2013 1603
REPORTS
on
Apr
il 9,
201
3w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from

Other integral membrane proteases appear torely on different mechanisms for obtaining ac-cess to water. The catalytic serine of rhomboidproteases is positioned at the base of a funnel-shaped periplasmic-accessible cavity that maybe gated (29). The zinc atom of S2P is accessibleto the cytoplasm by means of a narrow water-permeable channel present in the closed (to sub-strate) conformation of the enzyme (23). The roleof the large cavity, the mechanism of specific rec-ognition of cleavage sites with divergent sequences,the unusual dual-cleavage process, and the pos-sible role of the farnesyl group in recognition aresome of the questions raised by this provocativestructure.
References and Notes1. J. M. Dolence, L. E. Steward, E. K. Dolence, D. H. Wong,
C. D. Poulter, Biochemistry 39, 4096 (2000).2. E. Kim et al., J. Biol. Chem. 274, 8383 (1999).3. J. C. Otto, E. Kim, S. G. Young, P. J. Casey, J. Biol. Chem.
274, 8379 (1999).4. V. L. Boyartchuk, M. N. Ashby, J. Rine, Science 275, 1796
(1997).5. K. Fujimura-Kamada, F. J. Nouvet, S. Michaelis,
J. Cell Biol. 136, 271 (1997).6. A. Tam et al., J. Cell Biol. 142, 635 (1998).7. S. Michaelis, J. Barrowman, Microbiol. Mol. Biol. Rev.
76, 626 (2012).8. C. V. Jongeneel, J. Bouvier, A. Bairoch, FEBS Lett. 242,
211 (1989).9. A. Tam, W. K. Schmidt, S. Michaelis, J. Biol. Chem.
276, 46798 (2001).10. T. A. Dittmer, T. Misteli, Genome Biol. 12, 222 (2011).
11. H. J. Worman, J. Pathol. 226, 316 (2012).12. J. Barrowman, P. A. Wiley, S. E. Hudon-Miller,
C. A. Hrycyna, S. Michaelis, Hum. Mol. Genet. 21, 4084(2012).
13. C. Coffinier et al., Proc. Natl. Acad. Sci. U.S.A. 104,13432 (2007).
14. S. E. Hudon et al., Biochem. Biophys. Res. Commun.374, 365 (2008).
15. K. M. Clark et al., Protein Expr. Purif. 71, 207 (2010).16. M. G. Malkowski et al., Proc. Natl. Acad. Sci. U.S.A.
104, 6678 (2007).17. A. Quigley et al., Science 339, 1604 (2013).18. D. G. Hangauer, A. F. Monzingo, B. W. Matthews,
Biochemistry 23, 5730 (1984).19. W. R. Kester, B. W. Matthews, Biochemistry 16, 2506
(1977).20. B. W. Matthews, Acc. Chem. Res. 21, 333 (1988).21. B. W. Matthews et al., Nat. New Biol. 238, 41 (1972).22. B. W. Matthews, J. N. Jansonius, P. M. Colman,
B. P. Schoenborn, D. Dupourque, Nat. New Biol. 238,37 (1972).
23. L. Feng et al., Science 318, 1608 (2007).24. M. S. Brown, J. Ye, R. B. Rawson, J. L. Goldstein, Cell
100, 391 (2000).25. T. E. Golde, C. B. Eckman, Sci. STKE 2003, RE4
(2003).26. M. S. Wolfe, R. Kopan, Science 305, 1119 (2004).27. G. Huyer et al., Eukaryot. Cell 5, 1560 (2006).28. C. E. Trueblood et al., Mol. Cell. Biol. 20, 4381 (2000).29. S. Urban, Biochem. J. 425, 501 (2010).30. J. Dundas et al., Nucleic Acids Res. 34(Web server issue),
W116 (2006).31. B. K. Ho, F. Gruswitz, BMC Struct. Biol. 8, 49 (2008).32. W. K. Schmidt, A. Tam, S. Michaelis, J. Biol. Chem. 275,
6227 (2000).33. J. B. Klauda et al., J. Phys. Chem. B 114, 7830 (2010).34. K. Berka et al., Nucleic Acids Res. 40 (Web server issue),
W222 (2012).
Acknowledgments: Author contributions (authorship ofrelevant supplementary materials: Materials and Methodssections denoted by *): target selection and yeast homologyscreening (M.E.D.*); cloning (S.M.C); expression, solubilization,purification, and functional characterization with accompanyingfig. S3 (K.M.C.*, N.F.*, M.E.D.*); preliminary crystallization anddiffraction screening (M.K.-R., G.Z., M.G.M.*); SmSte24pcrystallization (E.E.P.*), SmSte24p crystallography (E.E.P.*,P.S.H.*, M.C.W.). M.C.W. conceptualized and wrote the mainpaper, with the following exceptions: introduction (M.E.D.),figures and captions (E.E.P, P.S.H., M.C.W.). All authorscritically read the manuscript before submission. In particular,M.E.D. contributed substantially to the editing, revision, andimprovement of the manuscript. This work was supported byProtein Structure Initiative: Biology grant U54 GM094611(NIH) for membrane protein structural genomics. Data werecollected at Southeast Regional Collaborative Access Team(SER-CAT) 22-ID beamline at the Advanced Photon Source,Argonne National Laboratory. Use of the Advanced PhotonSource was supported by the U.S. Department of Energy, Officeof Science, Office of Basic Energy Sciences, under ContractNo. W-31-109-Eng-38. We thank P. Loll, R. Nakamoto, andJ. Zimmer for useful discussion. Atomic coordinates andstructure factors have been deposited in the Protein Data Bankwith accession number 4IL3. Vectors and clones are availablefrom the NIH Protein Structure Initiative Materials Repository,under a Materials Transfer Agreement. The authors declare nocompeting financial interests.
Supplementary Materialswww.sciencemag.org/cgi/content/full/339/6127/1600/DC1Materials and MethodsFigs. S1 to S8Tables S1 to S4References (35–58)
26 October 2012; accepted 8 February 201310.1126/science.1232048
The Structural Basis ofZMPSTE24-Dependent LaminopathiesAndrew Quigley,1* Yin Yao Dong,1* Ashley C. W. Pike,1* Liang Dong,1 Leela Shrestha,1
Georgina Berridge,1 Phillip J. Stansfeld,2 Mark S. P. Sansom,2 Aled M. Edwards,3 Chas Bountra,1
Frank von Delft,1 Alex N. Bullock,1 Nicola A. Burgess-Brown,1 Elisabeth P. Carpenter1†
Mutations in the nuclear membrane zinc metalloprotease ZMPSTE24 lead to diseases of lamin processing(laminopathies), such as the premature aging disease progeria and metabolic disorders. ZMPSTE24processes prelamin A, a component of the nuclear lamina intermediate filaments, by cleaving it attwo sites. Failure of this processing results in accumulation of farnesylated, membrane-associatedprelamin A. The 3.4 angstrom crystal structure of human ZMPSTE24 has a seven transmembranea-helical barrel structure, surrounding a large, water-filled, intramembrane chamber, capped by azinc metalloprotease domain with the catalytic site facing into the chamber. The 3.8 angstromstructure of a complex with a CSIM tetrapeptide showed that the mode of binding of the substrateresembles that of an insect metalloprotease inhibitor in thermolysin. Laminopathy-associatedmutations predicted to reduce ZMPSTE24 activity map to the zinc metalloprotease peptide–bindingsite and to the bottom of the chamber.
The nuclear lamina is composed of inter-mediate filaments that give structure tothe nuclei of cells in many multicellular
organisms (1, 2). ZMPSTE24 (also known as
farnesylated-protein converting enzyme 1, FACE-1,Hs Ste24) is an inner nuclear membrane zincmetalloprotease that is essential for the matura-tion of laminA, one of three lamin proteins foundin the lamina of differentiated cells (3–5). LaminA and B precursor proteins undergo a cascadeof C-terminal, posttranslational modificationsinitiated by farnesylation of the cysteine on theC-terminal CAAX motif (where A is any ali-phatic residue and X is any residue). ZMPSTE24or the Rce1 protease (6) subsequently cleave
theAAX residues from theC terminus of prelaminA (the CAAX cleavage reaction), followed bycarboxymethylation of the farnesyl cysteine.ZMPSTE24 then cleaves an additional 15 resi-dues from the C terminus of prelamin A. Thisremoves the membrane-embedded farnesyl cys-teine (fCys) and releases mature lamin A into thenucleoplasm (5). The ZMPSTE24 yeast homo-log, Ste24, performs two similar cleavage reac-tions on the yeast mating pheromone a-factor (7).
Mutations in either prelamin A or ZMPSTE24cause a family of diseases called laminopathies,which result from accumulation of farnesylated,unprocessedprelaminA in thenucleus. ZMPSTE24-null mice have spontaneous bone fractures, cardio-myopathy, severe muscle weakness, and hair loss(3, 8). In humans, the severity of ZMPSTE24-dependent laminopathies correlates with the levelof residual protease activity (9). The most severeZMPSTE24-dependent laminopathies are neo-natal lethal restrictive dermopathy (RD) (4, 10–12)and the premature aging (progeria) disease atypicalHutchinson-Gilford progeria syndrome (HGPS) (13).Milder laminopathies—such as mandibuloacraldysplasia type B (MAD-B) (14) and, potentially,metabolic syndrome (MS) (15)—have a range ofsymptoms including lipodystrophy (LD) and in-sulin resistance (16), bone and skin abnormalities,and cardiomyopathy (14, 15, 17). During normalaging, ZMPSTE24 expression decreases in vas-cular smooth muscle cells, which leads to accu-mulation of unprocessed prelamin A, disrupting
1Structural Genomics Consortium, University of Oxford, OldRoad Campus Research Building, Roosevelt Drive, Oxford,OX3 7DQ, UK. 2Department of Biochemistry, University ofOxford, Oxford, OX1 3QU, UK. 3Structural Genomics Consortium,University of Toronto, Toronto, Ontario M5G 1L7, Canada.
*These authors contributed equally to this work.†Corresponding author. E-mail: [email protected]
29 MARCH 2013 VOL 339 SCIENCE www.sciencemag.org1604
REPORTS
on
Apr
il 9,
201
3w
ww
.sci
ence
mag
.org
Dow
nloa
ded
from