structural resiliency of an egf-like subdomain bound to its target
TRANSCRIPT

Protein Science (1996), 5:195-203. Cambridge University Press. Printed in the USA. Copyright 0 1996 The Protein Society
Structural resiliency of an EGF-like subdomain bound to its target protein, thrombin
RICHARD HRABAL,' ELIZABETH A. KOMIVES,* AND FENG NI'
the Montreal Joint Centre for Structural Biology, Montreal, Quebec, Canada H4P 2R2 ' Biotechnology Research Institute, National Research Council of Canada, and
*Department of Chemistry and Biochemistry, University of California at San Diego, La Jolla, California 92093-0601
(RECEIVED August 28, 1995; ACCEPTED November 15, 1995)
Abstract
The thrombin-bound structures of native peptide fragments from the fifth EGF-like domain of thrombomodulin were determined by use of NMR and transferred NOE spectroscopy. The bound peptides assume an EGF-like structure of an antiparallel @-sheet, a novel structural motif observed for a bound peptide in protein-peptide complexes. There is a remarkable structural resiliency of this structure motif manifested in its ability to accom- modate a different number of residues within the disulfide loop. Docking experiments revealed that the key contacts with thrombin are hydrophobic interactions between the side chains of residues Ile 414 and Ile 424 of thrombo- modulin and a hydrophobic pocket on the thrombin surface. Residues Leu 415, Phe 419, and Ile 420, which would have been buried in intact EGF-like domains, are unfavorably exposed in the complex of thrombin with the EGF-like thrombomodulin fragment, thus providing a rationale for the enhancement of binding affinity upon the deletion of Ile 420. The unique @-sheet structures of the bound peptides are specified by the presence of disulfide bridges in the peptides because a corresponding linear thrombomodulin fragment folds into a sheet structure with a dif- ferent backbone topology. The different bound conformations for the linear and the cyclized peptides indicate that side-chain interactions within a specific environment may dictate the folding of bound peptides in protein- peptide complexes.
Keywords: EGF-like domain; protein-peptide interaction; thrombomodulin; thrombin; transferred NOE
EGF-like repeats are autonomous folding units of -50 amino acid residues occurring in many multidomain proteins (Appella et al., 1988; Engel, 1989). The regulatory roles of the EGF-like domains in protein interactions have promoted an enormous amount of research progress into the structure-function relation- ships of these protein modules (Campbell & Bork, 1993; Groe- nen et al., 1994). In solution, EGF and most EGF-like modules fold into a compact structure with an N-terminal subdomain or- ganized by two disulfide bonds, followed by a relatively inde- pendent C-terminal subdomain of 15-20 amino acid residues (Groenen et al., 1994). The apparently conserved structural scaf- fold of EGF-like domains may allow the tailoring of binding specificity by the replacement of surface residues, as in EGF- like modules found in different proteins (Appella et al., 1988; Campbell & Bork, 1993). Once the essential binding residues are identified, it may become possible to design selective peptido- mimetic ligands to interfere with the specific interactions be- tween EGF or EGF-like proteins and their molecular targets.
Reprint requests to: Feng Ni, Biotechnology Research Institute, Na- tional Research Council Canada, 6100 Royalmount Avenue, Montreal, Quebec, Canada H4P 2R2; e-mail: [email protected].
Advances in structure-function relationships of EGF and EGF-like protein modules culminated in a recent attempt to de- sign thrombin-specific ligands (Lougheed et al., 1995) to mimic part of the functions of thrombomodulin, a membrane-bound glycoprotein with six EGF-like repeats in its extracellular struc- ture (Esmon, 1989). An analog peptide with a smaller cyclic loop was constructed by deleting residues Glua8 and Ile420 from the native sequence Ea,CPEGYILDDGF1420CTDIDE in the C-terminal subdomain of the fifth EGF-like module of human thrombomodulin (Hayashi et al., 1990; Tsiang et al., 1990, 1992). The design was conducted in the absence of the three-dimensional structure of the native peptide and was based on the rationale (Lougheed et al., 1995) that the five residues C409PEGY413 are required to maintain the sequence and struc- ture conservation in almost all EGF-like domains (Appella et al., 1988; Campbell & Bork, 1993), whereas residues Y4131LDD417 and D4231DE426 are important for binding to thrombin (Naga- shima et al., 1993). The cyclized molecule Ca9PEGYILDD GF4&TDIDE has an enhanced thrombin-binding affinity (Lougheed et al., 1995) and binds specifically to the fibrinogen recognition exosite of thrombin with a bound structure identi- cal to the C-terminal subdomain of all EGF-like molecules
195

196
(Srinivasan et al., 1994). A parallel X-ray structure analysis re- vealed that the native thrombomodulin fragment ECP410E41 GYILDDGFICTDIDE folds into a similar molecular shape in its complex with thrombin (Mathews et al., 1994), but with a chain reversal centered around residues P r ~ , , ~ - G l u ~ ~ instead of residues G 1 ~ ~ ~ ~ - G 1 y ~ ~ ~ observed in the analog peptide CPE411G412YILDDGFCTDIDE (Srinivasan et al., 1994). The displacement by a single residue in the crystal structure allows the side chain of the conserved aromatic residue Tyr413 to make intimate intermolecular contacts with its target protein, throm- bin (Mathews et al., 1994). The sanle Tyr residue in the analog peptide is buried in the intrapeptide and three-stranded &sheet, making it apparently inaccessible for thrombin binding (Sriniva- san et al., 1994).
In this paper, transferred NOE spectroscopy was used to de- termine the thrombin-bound structure of a cyclized EGF-like fragment C4wPEGY1414LDDGF1420CTD1424DE, corresponding to the native sequence of the C-terminal subdomain of the fifth EGF-like repeat in human thrombomodulin. The new bound structure shows how the disulfide-bonded native peptide, while still adopting an EGF-like backbone conformation, can inter- act with thrombin in a similar fashion as captured in the crystal structure of the thrombin-thrombomodulin peptide com- plex. The same binding mode was also observed for a peptide, C40,EAPEGY1414LDDGFIC421TD1424DE, containing a non- native disulfide bond between the fourth cysteine (Cys 407) and the sixth cysteine (Cys 421) residues in the EGF-like protein do- main. The critical residues, 11e414 and Ile424, for thrombin bind- ing are brought into spatial proximity by a 0-sheet structure, a hitherto novel structural motif observed for a bound peptide in protein-peptide complexes (Siligardi & Drake, 1995; Stanfield & Wilson, 1995).
Results
Transferred nuclear Overhauser effects
In the presence of thrombin, the disulfide-cyclized native throm- bomodulin peptide C40,PEGY1414LDDGF1420CTD1424DE (fur- ther referred to as TM52+5C, Table l ) experienced changes of the chemical shifts and line widths of some proton resonances with respect to its free form. The largest chemical shift changes are from the aliphatic side-chain resonances of Ile 414 (0.05 ppm) and Ile 424 (0.04 ppm), the amide proton resonance of Ile 414 (0.05 ppm), and one of the PCH, protons of Asp 416 (0.07 ppm).
R. Hrabal et al.
For the peptide with a shifted disulfide bond, C407EAPEGY 14,4LDDGFIC421TD1424DE (referred to as TM52+2+5C, Ta- ble l), the changes upon binding are of a similar nature, but less pronounced, except for both of the Asp 416 0CH2 protons (0.05 ppm). These spectral changes were also observed for the des-Ile 420 analog peptide, C409PEGY1414LDDGFC421TD1424DE (further referred to as TM52-1+5C, Table I ) , which was estab- lished to interact specifically with the fibrinogen recognition exo- site of thrombin (Srinivasan et al., 1994; Lougheed et al., 1995). Therefore, the native peptides, TM52+5C and TM52+2+5C, must have identical, or at least very similar, binding contacts with the thrombin exosite compared with the Ile 420-deleted thrombomodulin peptide TM52-1+5C studied previously (Srini- vasan et al., 1994).
The spectral changes in the thrombomodulin peptides were a function of the concentration ratios of the peptides and throm- bin, indicating reversibility of the thrombin-peptide complexes and dynamic spectral averaging for the peptides between the free and thrombin-bound states (Ni, 1994). The free peptides gave rise only to a few intraresidue and sequential NOE crosspeaks with very low intensities (spectra not shown), suggesting that these peptides themselves do not have predominant structural preferences in solution. However, upon binding with thrombin, the NOESY spectra exhibited a large number of medium- and long-range NOE crosspeaks (Figs. 1,2), indicating that they are thrombin-induced transferred NOES (Albrand et al., 1979; Ni, 1994; Ni & Scheraga, 1994) unique to the bound peptides. The observation of these transferred NOEs again indicates that the thrombomodulin fragments interact specifically with thrombin, as established previously (Srinivasan et al., 1994).
Figure 3 presents an overview of important transferred NOEs observed for the thrombomodulin peptides. The transferred NOE connectivities were characteristic of the secondary struc- ture types for the C-terminal subdomains within all EGF-like protein modules (Campbell & Bork, 1993; Srinivasan et al., 1994). For peptide TM52+5C, for example, there were strong NOE contacts between the NH protons of residues Gly 412 and Tyr 413, and between the C,H proton of Glu 41 1 and the NH proton of Gly 412 (Fig. 3A), characterizing a type-I1 6-turn in- volving residues Glu 411 and Gly 412, as observed previously for TM52-1+5C (Srinivasan et al., 1994). Long-range NOEs be- tween the side chains of Tyr 413 and Asp 423/11e 424, between the backbone and the side-chain protons of Asp 416 and Ile 420, and between the a C H protons of Leu 415 and Cys 421, were characteristic of a two-stranded antiparallel P-sheet involving residues Tyr 413-Asp 416 and Phe 419-Ile 424. Most of the
Table 1. Sequences of EGF-like fragments from human thrombomodulina
Abbreviation Sequence DCT ( PM) IC50 (PM)
.~
TM52-1+5C: C4mPEGYI414LDDGF-CTDI4zdDE 24 f 2 23 f 3 TM52+5C: C,~~PEGYI~I~LDDGFICTDI~~~DE 35 * 5 65 f 10 TM52+2+5C: CEA,o9PEGYIql4LDDGFICTDI424DE 100 f 10 300 f 30
a DCT, concentration of peptides required to double the clotting time of fibrinogen catalyzed by thrombin; IC50, concen- tration of peptides required to achieve 50% of inhibition of protein C activation catalyzed by the thrombin-thrombomodulin complex. Inhibition constants are from Lougheed et al. (1995) for TM52-1+5C and from Blackmar et al. (1995) for TM52+5C and TM52+2+5C.

Thrombin-bound structures of EGF-like domains
A
B
G 4 1 2
G412 E41 I
G 4 1 8
E408
C412
0 0 Q
C418 ,
8 .'5 8 .'O
A
B
Fig. 1. Fingerprint (NH-aH) regions of the NOESY spectra of (A) TM52+5C and (B) TM52+2+5C in complex with thrombin. Residues are marked by a one-letter code and their positions in the sequence of thrombomodulin. Roman numerals in B indicate nonsequential NOE COntaCtS (I, D416,~-G418,~; 11, 1414,~-D416~~; 111, E411,H- Y413NH; Iv, L ~ ~ ~ N H - F ~ I ~ , H ; v, 1414~~-D423,~). Corresponding peptides exhibit very weak NOE intensities in the absence of thrombin.
transferred NOEs characteristic of the bound structure of TM52+5C were also found with peptide TM52+2+5C. Two ex- tra residues, Cys 407 and Glu 408, together with Ala 409 (replac- ing Cys 409) and Pro 410, had many more NOE contacts with the rest of the molecule (Fig. 3B). For example, the aromatic ring of Phe 419 makes numerous NOE interactions with Cys 407, Glu 408, and Ala 409 (Fig. 2B). The side-chain protons of Leu 415 also participate in NOE interactions with the aromatic rings of both Tyr 413 and Phe 419.
4 7 .'2 7 .'o 6 .'8 6 . 6
__P410 - Y 4 1 3 a
-P410 -Y413& 82
8 2
7 .'4 7 .'2 7 .'o 6 .'8 6
Fig. 2. Region of the NOESY spectra of (A) TM52+5C and (B) TM52+2+5C, showing NOE connectivities involving the side-chain pro- tons of residues Tyr 413 and Phe 419. None of these NOES were observed in the spectra of the free peptides in the absence of thrombin.
Thrombin-bound structures of the EGF-like subdomains
The bound structure of TM52+5C was calculated using a total of 120 distance constraints derived from the analysis of trans- ferred NOEs. After an initial refinement step, four hydrogen bonds were incorporated, one between Pro 410 CO and Tyr 413 NH, expected for a type I1 P-turn, and another linking Asp 416 CO and Phe 419 NH. The remaining two hydrogen bonds, be- tween Ile 414 CO and Thr 422 NH and between Ile 420 CO and Asp 416 NH, helped to regularize both strands of the antipar- allel &sheet. All of the hydrogen bond constraints were satis- fied in the subsequent refinement steps, producing a group of

198 R . Hrabal et al.
P-E
C-E-A-P-E-G-Y-I-L-D-D-G-F-I-C-T-D-I-D-E
Fig. 3. Summary of significant transferred NOES for (A) TM52+5C and (B) TM52+2+5C. The description indicates the types of protons involved in NOE contacts between the two amino acid residues.
structures with low distance violations and low interaction en- ergies. These structures also produced calculated transferred NOE spectra (not shown) closely matching the experimental data.
Figure 4A shows the superposition of a family of 11 best struc- tures of TM52+5C with excellent convergence properties (Ta- ble 2). The average RMS deviation (RMSD) between a particular structure and the mean structure is -0.5 A for the backbone at- oms and -0.9 A for all heavy atoms, considering well-defined residues Cys 409-Ile 424. The two C-terminal residues, Asp 425 and Glu 426, are not so well-defined, as seen in Figure 4A and from the increase of RMSD to 0.9 and 1.4 A for the backbone and all heavy atoms, respectively, calculated for all 18 residues. The poor definition of the C-terminus may be a result of the lack of long-range NOES and/or a higher flexibility of this part of the bound molecule. On the other hand, the N-terminal residues Pro 410-Tyr 413 form the conserved type I1 0-turn at the be- ginning of a two-stranded antiparallel @-sheet, comprising res- idues Tyr 413-Asp 416 and Phe 419-Ile 424. The position of both strands of the 0-sheet was determined uniquely by NOE contacts between the side-chain protons of Asp 416 and Thr 422 (Fig. 3A). The right-handed twist within the 0-sheet was dictated by the NOE interaction between Ile 414 methyl protons and the PCH, protons of Asp 416.
For TM52+2+5C, a total of 186 NOE constraints were uti- lized for the structure refinement, supplemented by only two pairs of hydrogen bonds, both of which were of the 1-4 type within the two 0-turns. Figure 4B shows a family of 14 struc- tures of TM52+2+5C with the structure statistics given in Ta- ble 2. The average RMSD for residues Cys 407-Ile 424 is 0.3 A
for the backbone atoms and 0.7 A for all heavy atoms. Similar to TM52+5C, the last two residues are poorly defined because of the lack of distance constraints.
The main structural motif of TM52+2+5C clearly consists of a three-stranded antiparallel 0-sheet. The first strand, com- posed of residues Cys 407-Pro 410, can be divided into two parts. The N-terminal part, i.e., residues Cys 407 and Glu 408, is aligned with the third strand, whereas residues Ala 409 and Pro 410 are closer to the second strand, making numerous NOE contacts with it. The first and second strands are connected by the first type I1 0-turn, which is stabilized by a hydrogen bond between Pro 410 CO and Tyr 413 NH. The second strand, span- ning residues Tyr 413-Asp 416, can also be divided into two parts. The first two residues, Tyr 413 and Ile 414, make NOE contacts with the residues of the first strand. The remaining two residues, Leu 415 and Asp 416, contact the third strand. The sec- ond 0-turn is also type 11, stabilized by a hydrogen bond between Leu 415 CO and Gly 418 NH. In comparison with the second 0-turn of TM52+5C, this one in TM52+2+5C is shifted by one residue toward the N-terminus. The third strand contains a bulge extending from Phe 419 through Asp 423, which is important for the molecule to accommodate the non-native and shifted di- sulfide bond between residues Cys 407 and Cys 421.
Discussion
Structural comparison with related EGF-like subdomains
Recent structural studies revealed that the C-terminal sub- domains of the EGF-like modules either form intimate molec-
Table 2. Refinement statistics of the TM52+5C and TM52+2+5C structures bound to thrombina
TM52+5Cb
XPLOR energies (kcal mol") Et,,
ENOE Eelec
Ebond
Eangle
Eirnpr
EVDW
NOE violations >0.5 A >0.1 A
Cartesian coordinate RMSD (A) [409(407)d-426] HAe [409(407)d-426] BAf [409(407)d-424] HAe [409(407)d-424] BAf
-24 k 5 -39 k 7
13 * 1 -134 k 4
40 k 2 87 f 3 7 f l
0 8
1.38 * 0.35 0.89 k 0.17 0.91 k 0.17 0.46 + 0.05
TM52+2+5CC
-38 k 6 -57 * 1
1 6 k 1 -134 k 10
39 k 1 90 k 2
8 f 1
0 10
0.87 k 0.21 0.44 k 0.1 1 0.69 k 0.19 0.29 + 0.04
dues Cys 409-Leu 415 and Ile 420-Ile 424 for TM52+5C and Cys 407- a Structures were superimposed using the backbone atoms of resi-
Leu 415 and Ile 420-Ile 424 for TM52+2+5C, respectively. RMSDs were calculated with respect to the mean structure.
Calculated as an average over 1 1 structures. Calculated as an average over 14 structures. TM52+2+5C.
e All heavy atoms. Backbone atoms.

Thrombin-bound structures of EGF-like domains 199
I
Fig. 4. Backbone structures of (A) TM52+5C and (B) TM52+2+5C in complex with thrombin. The cluster of conformations indicates that the bound structures of residues Cys 409 (or Ala 409) to Ile 424 are well-defined by NOE distance constraints (see Table 2). Atomic coordinates of the structures shown have been deposited with the Brookhaven Protein Data Bank.
ular contacts with other protein domains (Padmanabhan et al., 1993; Mathews et al., 1994; Srinivasan et al., 1994) or are freely accessible in multidomain proteins, leaving room for potential interactions with target proteins (Graves et al., 1994; Picot et al., 1994). Figure 5 shows a comparison of the EGF-like subdomains with a differing number of residues between the disulfide-bonded cysteine pairs. All the structures have the same canonical fold observed in EGF-like protein modules (Camp- bell & Bork, 1993). The first seven residues, CXXGYXX or CXXGFXX, assume a high degree of structural similarity, with
a type I1 /3-turn at the center of the XXGY/F motif and a con- served Gly residue. Furthermore, the side chains of the aromatic residues Tyr (or Phe), are all embedded inside the interstrand space within the P-sheet, making close contacts with the disul- fide bond between the two cysteine residues (Srinivasan et al., 1 994).
The 12-residue disulfide loop in peptide TM52-1+5C allows the formation of an optimized antiparallel &sheet, in compar- ison with the 10-residue loop analogues, where there is always a bulge after the last cysteine residue (Fig. 5) . On the other hand,
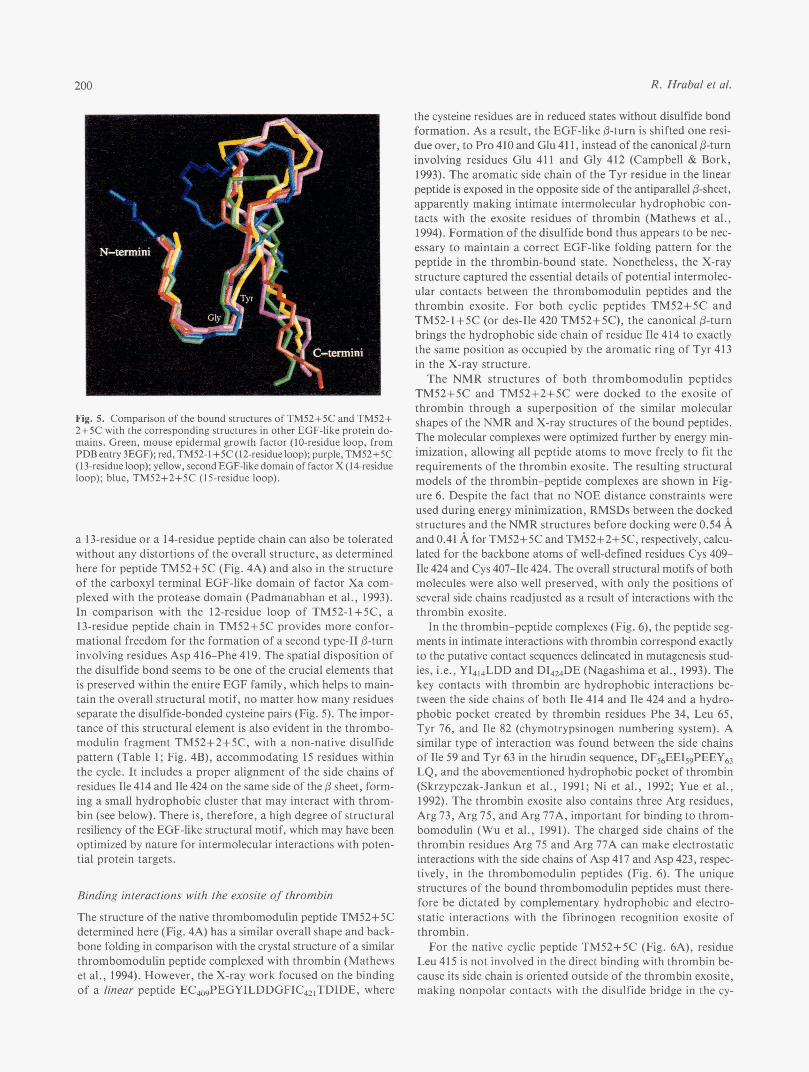
R. Hrabal et al. 200
Fig. 5. Comparison of the bound structures of TM52+5C and TM52+ 2+5C with the corresponding structures in other EGF-like protein do- mains. Green, mouse epidermal growth factor (IO-residue loop, from PDB entry 3EGF); red, TM52-l+5C (12-residue loop); purple, TM52+5C (13-residue loop); yellow, secondEGF-like domain of factor X (14-residue loop); blue, TM52+2+5C (15-residue loop).
a 13-residue or a 14-residue peptide chain can also be tolerated without any distortions of the overall structure, as determined here for peptide TM52+5C (Fig. 4A) and also in the structure of the carboxyl terminal EGF-like domain of factor Xa com- plexed with the protease domain (Padmanabhan et al., 1993). In comparison with the 12-residue loop of TM52-1+5C, a 13-residue peptide chain in TM52+5C provides more confor- mational freedom for the formation of a second type-I1 &turn involving residues Asp 416-Phe 419. The spatial disposition of the disulfide bond seems to be one of the crucial elements that is preserved within the entire EGF family, which helps to main- tain the overall structural motif, no matter how many residues separate the disulfide-bonded cysteine pairs (Fig. 5) . The impor- tance of this structural element is also evident in the thrombo- modulin fragment TM52+2+5C, with a non-native disulfide pattern (Table 1; Fig. 4B), accommodating 15 residues within the cycle. It includes a proper alignment of the side chains of residues Ile 414 and Ile 424 on the same side of the 0 sheet, form- ing a small hydrophobic cluster that may interact with throm- bin (see below). There is, therefore, a high degree of structural resiliency of the EGF-like structural motif, which may have been optimized by nature for intermolecular interactions with poten- tial protein targets.
Binding interactions with the exosite of thrombin
The structure of the native thrombomodulin peptide TM52+5C determined here (Fig. 4A) has a similar overall shape and back- bone folding in comparison with the crystal structure of a similar thrombomodulin peptide complexed with thrombin (Mathews et al., 1994). However, the X-ray work focused on the binding of a linear peptide ECm$EGYILDDGFIC421TDIDE, where
the cysteine residues are in reduced states without disulfide bond formation. As a result, the EGF-like P-turn is shifted one resi- due over, to Pro 410 and Glu 41 1, instead of the canonical &turn involving residues Glu 411 and Gly 412 (Campbell & Bork, 1993). The aromatic side chain of the Tyr residue in the linear peptide is exposed in the opposite side of the antiparallel &sheet, apparently making intimate intermolecular hydrophobic con- tacts with the exosite residues of thrombin (Mathews et al., 1994). Formation of the disulfide bond thus appears to be nec- essary to maintain a correct EGF-like folding pattern for the peptide in the thrombin-bound state. Nonetheless, the X-ray structure captured the essential details of potential intermolec- ular contacts between the thrombomodulin peptides and the thrombin exosite. For both cyclic peptides TM52+5C and TM52-1+5C (or des-Ile 420 TM52+5C), the canonical &turn brings the hydrophobic side chain of residue Ile 414 to exactly the same position as occupied by the aromatic ring of Tyr 413 in the X-ray structure.
The NMR structures of both thrombomodulin peptides TM52+5C and TM52+2+5C were docked to the exosite of thrombin through a superposition of the similar molecular shapes of the NMR and X-ray structures of the bound peptides. The molecular complexes were optimized further by energy min- imization, allowing all peptide atoms to move freely to fit the requirements of the thrombin exosite. The resulting structural models of the thrombin-peptide complexes are shown in Fig- ure 6. Despite the fact that no NOE distance constraints were used during energy minimization, RMSDs between the docked structures and the NMR structures before docking were 0.54 A and 0.41 A for TM52+5C and TM52+2+5C, respectively, calcu- lated for the backbone atoms of well-defined residues Cys 409- Ile 424 and Cys 407-Ile 424. The overall structural motifs of both molecules were also well preserved, with only the positions of several side chains readjusted as a result of interactions with the thrombin exosite.
In the thrombin-peptide complexes (Fig. 6), the peptide seg- ments in intimate interactions with thrombin correspond exactly to the putative contact sequences delineated in mutagenesis stud- ies, i.e., Y1414LDD and D14,DE (Nagashima et al., 1993). The key contacts with thrombin are hydrophobic interactions be- tween the side chains of both Ile 414 and Ile 424 and a hydro- phobic pocket created by thrombin residues Phe 34, Leu 65, Tyr 76, and Ile 82 (chymotrypsinogen numbering system). A similar type of interaction was found between the side chains of Ile 59 and Tyr 63 in the hirudin sequence, DF&EI,$EEY,, LQ, and the abovementioned hydrophobic pocket of thrombin (Skrzypczak-Jankun et al., 1991; Ni et al., 1992; Yue et al., 1992). The thrombin exosite also contains three Arg residues, Arg 73, Arg 75, and Arg 77A, important for binding to throm- bomodulin (Wu et al., 1991). The charged side chains of the thrombin residues Arg 75 and Arg 77A can make electrostatic interactions with the side chains of Asp 417 and Asp 423, respec- tively, in the thrombomodulin peptides (Fig. 6). The unique structures of the bound thrombomodulin peptides must there- fore be dictated by complementary hydrophobic and electro- static interactions with the fibrinogen recognition exosite of thrombin.
For the native cyclic peptide TM52+5C (Fig. 6A), residue Leu 415 is not involved in the direct binding with thrombin be- cause its side chain is oriented outside of the thrombin exosite, making nonpolar contacts with the disulfide bridge in the cy-

Thrombin-bound structures of EGF-like domains 20 1
d f'
Fig. 6. Structures of (A) TM52+5C and (B) TM52+2+5C docked to the fibrinogen recognition exosite of thrombin. Thrombin exosite is rep- resented by a molecular surface presented by residues Met 32-Met 84 (chymotrypsinogen numbering scheme). Light red on the molecular sur- face indicates a hydrophobic patch created by Phe 34, Leu 65, Tyr 76. and Ile 82. Light blue represents the positive charges from residues Arg 67, Arg 75, and Arg 77A. Side chains of the peptide residues in- volved in complex formation are indicated in red (Ile 414 and Ile 424) and blue (Asp 416, Asp 417, and Asp 423), respectively. Side chains of residues Tyr 413, Phe 419, and Ile 420 are colored in brown and the disulfide bond in yellow.
clized peptide. The side chains of residues Phe 419 and Ile 420 may also cluster into hydrophobic patches, but they are exposed unfavorably outside of the thrombin binding surface into the solvent. Phe 419 and Ile 420 therefore may not contribute to the binding of the thrombomodulin peptides with the exosite of
thrombin, as supported by the substitution of these correspond- ing two residues to Ser 419 and Thr 420 in bovine thrombo- modulin (Suzuki et al., 1987). Furthermore, residues Leu 415, Phe 419, and Ile 420 would have lain at the interface between the N-terminal (the A- and B-loops) and the C-terminal (the C-loop) subdomains of the intact fifth EGF-like domain in thrombomodulin, with their hydrophobic side chains masked by residues in the N-terminal subdomain (Campbell & Bork, 1993). The lack of interdomain contacts in peptide TM52+2+5C may be partially compensated by hydrophobic interactions among residues Cys 407, Leu 415, Phe 419, Ile 420, and Cys 421, revealed by extensive NOE contacts among these residues (Fig. 2B). The smaller disulfide loop in the native peptide TM52+5C ap- parently does not allow these extensive hydrophobic contacts seen in the larger peptide TM52+2+5C, resulting in a reduced number of NOES (Fig. 2A). These observations suggest that re- moval of an exposed and unpaired hydrophobic residue may be the reason for the slight enhancement of affinity upon the de- letion of Ile420 in peptide TM52-1+5C in comparison with the native peptide TM52+5C (Table I) .
The study of peptide-protein interactions is of fundamental importance both for an understanding of the structure-function relationships of peptides and proteins and for the rational de- sign of peptidomimetic ligands. Recent progress indicates that the binding of peptides to protein targets seems to require vari- ous conformational motifs, including local turns, or-helices, and extended structures (Siligardi & Drake, 1995; Stanfield & Wil- son, 1995). The current work reveals that sophisticated /3-sheet structures can also be used by peptides to define specific inter- actions with their protein targets. The unique &sheet structures of the bound peptides may be specified by the presence of di- sulfide bridges in the peptides, because a corresponding linear peptide can fold into a sheet structure with a different backbone topology (Mathews et al., 1994). The different bound confor- mations for the linear and cyclic thrombomodulin peptides sup- port the observed context dependencies of &sheet propensities of amino acid residues in proteins (Minor & Kim. 1994; Otzen & Fersht, 1995) and that side-chain interactions within a specific environment may dictate the folding of bound peptides in protein-peptide complexes (Siligardi & Drake, 1995). Because the bound structure is not a predominant one for the free pep- tides in solution, it will be a great challenge to develop strategies to further improve the binding affinities of the EGF-like throm- bomodulin peptides toward thrombin.
Materials and methods
NMR sample preparation
Disulfide-cyclized peptides TM52+5C, with a sequence of CaPEGYILDDGFICTDIDE426, and TM52+2+5C, with a se- quence of C407EAPEGYILDDGFICTDIDE42a (Table l), were synthesized and assayed following procedures described previ- ously (Lougheed et al., 1995). Bovine a-thrombin was purified from bovine plasma prothrombin after a controlled activation by snake venom proteases (Ni et al., 1990). The active site of thrombin was blocked by D-Phe-Pro-Arg-CH,CI (PPACK) to prevent thrombin autolysis. Alternatively, purified bovine pro- thrombin was used directly without activation because the exo- site is available in prothrombin for peptide binding (Ni et al., 1993).

202 R. Hrabal et al.
All NMR samples were prepared as aqueous solutions that were 50 mM in sodium phosphate and 0.2 mM in EDTA. Sam- ples of the free peptides were prepared as 2-3 mM solutions in a mixture of 90% H 2 0 and 10% D20. The pH values of the peptide samples were adjusted to 5.5 by adding a small amount of either HCI or NaOH. Samples of the peptide ligands in com- plexes with thrombin (or prothrombin) were prepared by add- ing the stock solutions of the particular protein with a proper pH value to a pH-adjusted solution of the peptides. In the case of thrombin complexes, the concentration of the peptide was 0.7 mM and the thrombidpeptide ratio 10: 1, whereas the pro- thrombin sample contained 2.5 mM peptide and 15: 1 ratio of peptide to prothrombin.
NMR spectroscopy
All NMR experiments were conducted on a Bruker AMX-500 spectrometer equipped with a BGU unit producing pulsed-field gradients in a 5-mm inverse gradient probe. All NMR data were acquired at 25 "C. Water suppression was achieved by a gradient- echo method WATERGATE (Piotto et al., 1992). Both clean- TOCSY (TOWNY-16) and NOESY experiments contained an extra purging gradient pulse at the end of the mixing period and were acquired as 350 t,-increments, 2,048 data points in each FID. Sine modulation along the t,-dimension was applied by adjusting the initial delay to IN-4Pm/?r (Ni, 1992b) to eliminate t,-ridges in the 2D spectra. NOESY spectra typically had mix- ing times between 100 and 200 ms. NMR data were processed with the FELIX 2.0 software program (Hare Research, Inc.) by multiplication with sine-bell window function shifted by 30" in both dimensions and zero filled to matrices of 2,048 x 1,024 data points each. A polynomial baseline correction was used to fur- ther improve the quality of 2D NMR spectra (Ni, 1992b).
Structure calculations
Structure calculations were carried out with XPLOR (version 3.1, Briinger, 1992). The Sybyl software package (version 6.0, TRIPOS Associates) was employed for the visualization of mo- lecular structures. The intensities of cross peaks in NOESY spec- tra were divided into strong, medium, and weak, respectively, and converted to the corresponding distance constraints of 1.8-2.7 A, 1.8-3.5 A, and 1.8-5.OA, respectively. The accuracy of distance constraints were improved and spin-diffusion in NOESY spectra was corrected through an iterative procedure of distance calibration (Ni, 1994). Hydrogen bonds were treated as additional distance constraints of 1.7-2.2 A without any re- strictions of dihedral angles. Starting structures were generated through distance geometry followed by a few steps of simu- lated annealing for the regularization of covalent geometries (Kuszewski et al., 1992). The full simulated annealing procedure for structure refinement consisted of 12-ps high temperature dy- namics (1,000 K) followed by a cooling period of 27 ps, during which the temperature dropped to 300 K .
The NOE penalty function was set to 50 kcal mol" A-2 with the square-well potential used throughout the calculations. Non- stereospecifically assigned groups of protons were treated uti- lizing [r,$"6 as an effective distance (Clore et al., 1986). Other force constants were uniformly set to 1,000 kcal mol" A - 2 for bond lengths and to 500 kcal mol" radp2 for both bond angles and impropers. The van der Waals energy term was represented
by the Lenard-Jones 12-6 function with a gradual increase of its weight from 0.005 at the beginning of each refinement to 4.0 at the end. During the initial stages of the structural refinement, no electrostatic interactions were incorporated, however, they were added during the final stages with a weight of 0.125 of the VDW term, which helped to further regularize the resulting structures. Correctness of the computed structures were checked by the calculation of NOESY spectra accomplished by in-house programs PDBZNOE, producing matrices of NOE intensities, and GFIDSJ, converting the NOE intensities into regular NMR data (Ni, 1992a, 1994; Ni & Zhu, 1994).
Computation of the thrombin-peptide complexes
The structures of TM52+5C and TM52+2+5C were super- imposed onto the X-ray structure of a linear thrombomodulin peptide bound to thrombin (Mathews et al., 1994). Steric clashes and unfavorable van der Waals interactions were released by 2,000 steps of Powell energy minimization. No NOE constraints were applied during the minimization procedure. Nonbonded interactions were calculated only within the ligand and between the ligand and the enzyme. The weight of the van der Waals en- ergy term was increased to 4.0 and the electrostatic interactions were calculated with the weight of 1 .O. Positions of the enzyme atoms were fixed and only the ligand atoms were allowed to move during the minimization.
Structure coordinates
The atomic coordinates of the structures have been deposited with the Brookhaven Protein Data Bank, accession numbers IFGD and IFGE.
Acknowledgments
We thank Song Hu for providing bovine a-thrombin and for conduct- ing preliminary binding studies with TM52+5C, Betty Zhu for help with NOE simulations, and Bruce Fulton for help with pulse-field-gradient experiments. Professor AI Tulinsky is acknowledged for providing the X-ray coordinates of the second EGF-like domain in complex with fac- tor X and the complex of thrombin with a linear thrombornodulin pep- tide. This work was supported in part by a research grant (to. EN.) from the Medical Research Council of Canada (MRC MT-12566). This is NRCC publication no. 38565.
References
Albrand JP, Birdsall B, Feeney J, Roberts GCK, Burgen ASV. 1979. The use of transferred nuclear Overhauser effects in the study of the confor- mations of small molecules hound to proteins. Int J Biol Macromol
Appella E, Weber IT, Blasi F. 1988. Structure and function of epidermal 1:37-41.
Blackmar C, Healy VL, Hrabal R, Ni F, Komives EA. 1995. Structure/ growth factor-like regions in proteins. FEES Lett 231: 1-4.
activity of the region of thrombomodulin that binds thrombin. Bioorganic Chemistry. In press.
Briinger AT. 1992. XPLOR manual, version 3.1. New Haven, Connecticut: Yale University.
Campbell ID, Bork P. 1993. Epidermal growth factor-like modules. Curr Opin Struct Biol3:385-392.
Clore GM, Gronenborn AM, Carlson G , Meyer EF. 1986. Stereochemistry of binding of the tetrapeptide acetyl-Pro-Ala-Pro-Tyr-NH2 to porcine pancreatic elastase. JMol Biol 190:259-267.
Engel J. 1989. EGF-like domains in extracellular matrix proteins: Localized signals for growth and differentiation? FEES Lett 2Sl:l-7.

Thrombin-bound structures of EGF-like domains 203
Esmon CT. 1989. The roles of protein C and thrombomodulin in the regu- lation of blood coagulation. J Eiol Chem 264:4743-4746.
Graves BJ, Crowther RL, Chandran C, Rumberger JM, Li S, Huang KS, Presky DH, Familletti PC, Wolitzky BA, Burns DK. 1994. Insight into E-selectin/ligand interaction from the crystal structure and mutagenesis of the Lec/EGF domains. Nature 367532-538.
Groenen LC, Nice EC, Burgess AW. 1994. Structure-function relationships for the EGF/TGF-a family of mitogens. Growth Factors 11:235-257.
Hayashi T, Zushi M, Yamamoto S, Suzuki K. 1990. Further localization of binding sites for thrombin and protein C in human thrombomodulin. J Biol Chem 265:20156-20159,
Kuszewski J, Nilges M, Brunger AT. 1992. Sampling and efficiency of met- ric matrix distance geometry: A novel partial metrization algorithm. J
Lougheed JC, Bowman CL, Meininger DP, Komives EA. 1995. Thrombin inhibition by cyclic peptides from thrombomodulin. Protein Sci 4 : 773-780.
Mathews 11, Padmanabhan KP, Tulinsky A, Sadler JE. 1994. Structure of a nonadecapeptide of the fifth EGF domain of thrombomodulin com- plexed with thrombin. Biochemistry 33:13547-13552.
Minor DL Jr, Kim PS. 1994. Context is a major determinant of &sheet pro- pensity. Nature 371 :264-267.
Nagashima M, Lundh E, Leonard JC, Morser J, Parkinson JF. 1993. Alanine-scanning mutagenesis of the epidermal growth factor-like do- mains of human thrombomodulin identifies critical residues for its co- factor activity. J Biol Chem 268:2888-2892.
Ni F. 1992a. Complete relaxation matrix analysis of transferred nuclear Over- hauser effects. JMagn Reson 96:651-656.
Ni F. 1992b. Optimised acquisition and processing of homonuclear 3D NMR spectra. Applications to transferred-NOE experiments. J Magn Reson
Ni F. 1994. Recent developments in transferred NOE methods. Prog NMR Specrrosc 26517-606.
Ni F, Konishi Y, Scheraga HA. 1990. Thrombin-bound conformation of the C-terminal fragments determined by transferred nuclear Overhauser ef- fects. Biochemistry 29:4479-4489.
Ni F, Ning Q, Jackson CM, Fenton JW. 1993. Thrombin exosite for fibrin- ogen recognition is partially accessible in prothrombin. J Biol Chem 268:16899-16902.
Ni F, Ripoll DR, Purisima EO. 1992. Conformational stability of a thrombin- binding peptide derived from the hirudin C-terminus. Biochemistry 31: 2545-2554.
Ni F, Scheraga HA. 1994. Use of transferred NOE to determine the confor- mations of ligands bound to proteins. Acc Chem Res. Submitted.
Biol NMR 2~33-56.
99:391-400.
Ni F, Zhu Y. 1994. Accounting for ligand-protein interactions in the relaxation-matrix analysis of transferred nuclear Overhauser effects. J Magn Reson B 102:180-184.
Otzen DE, Fersht AR. 1995. Side-chain determinants of &sheet stability.
Padmanabhan K, Padmanabhan KP, Tulinsky A, Park CH, Bode W, Huber Biochemistry 345718-5724.
R, Blankenship DT, Cardin AD, Kisiel W. 1993. Structure of human Des(1-45) factor Xa at 2.2 A resolution. JMol Biol232:947-966.
Picot D, Loll PJ, Garavito RM. 1994. The X-ray crystal structure of the mem- brane protein prostaglandin H2 synthase-1. Nuture 367:243-249.
Piotto M, Saudek V, Sklenar V. 1992. Gradient-tailored excitation for single- quantum NMR spectroscopy of aqueous solutions. J Eiomol NMR 2:
Siligardi G, Drake AF. 1995. The importance of extended conformations and,
tides. Biopolymers (Peptide Science) 37:28 1-292. in particular, the PI, conformation for the molecular recognition of pep-
Skrzypczak-Jankun E, Carperos VE, Ravichandran KG, Tulinsky A, West- brook M, Maraganore JM. 1991. Structure of the hirugen and hirulog 1 complexes of alfa-thrombin. JMol Biol221:1379-1393.
Srinivasan J, Hu S, Hrabal R, Zhu Y, Komives EA, Ni F. 1994. Thrombin- bound structure of an ECF subdomain from human thrombomodulin determined by transferred nuclear Overhauser effects. Biochemistry 33: 13553-13560.
Stanfield RL, Wilson IA. 1995. Protein-peptide interactions. Curr Opin Struct Biol5:103-113.
Suzuki K, Kusumoto H, Deyashiki Y, Nishioka J, Maruyama I, Zushi M, Kawahara S, Honda G , Yamamoto S, Horiguchi S . 1987. Structure and
thelium acting as a cofactor for protein C. EMEO J 6:1891-1897. expression of human thrombomodulin, a thrombin receptor on endo-
Tsiang M, Lentz SR, Dittman WA, Duanzhi w, Scarpati EM, Sadler EJ. 1990. Equilibrium binding of thrombin to recombinant human throm- bomodulin: Effect of hirudin, fibrinogen, factor Va, and peptide ana- logues. Biochemistry 29:10602-10612.
Tsiang M, Lentz SR, Sadler EJ. 1992. Functional domains of membrane- bound human thrombomodulin. J Biol Chem 267:6164-6170.
Wu Q , Sheehan JP, Tsiang M, Lentz SR, Birktoft JJ, Sadler EJ. 1991. Sin- gle amino acid substitutions dissociate fibrinogen-clotting and thrombin- binding activities of human thrombin. Proc Nut1 Acad Sci USA 88: 6775-6779.
Yue SY, DiMaio J, Szewczuk Z, Purisima EO, Ni F, Konishi Y. 1992. Characterization of the interactions of a bifunctional inhibitor with or-thrombin by molecular modelling and peptidesynthesis. Protein Eng
661-665.
5:77-85.