structural basis for antibody-mediated neutralization of ... · west africa every year.the...
TRANSCRIPT

RESEARCH ARTICLE◥
STRUCTURAL BIOLOGY
Structural basis forantibody-mediated neutralizationof Lassa virusKathryn M. Hastie,1 Michelle A. Zandonatti,1 Lara M. Kleinfelter,2 Megan L. Heinrich,3
Megan M. Rowland,3 Kartik Chandran,2 Luis M. Branco,3 James E. Robinson,4
Robert F. Garry,3,5 Erica Ollmann Saphire1,6*
The arenavirus Lassa causes severe hemorrhagic fever and a significant disease burden inWest Africa every year. The glycoprotein, GPC, is the sole antigen expressed on the viralsurface and the critical target for antibody-mediated neutralization. Here we present thecrystal structure of the trimeric, prefusion ectodomain of Lassa GP bound to a neutralizingantibody from a human survivor at 3.2-angstrom resolution. The antibody extensivelyanchors two monomers together at the base of the trimer, and biochemical analysissuggests that it neutralizes by inhibiting conformational changes required for entry. Thiswork illuminates pH-driven conformational changes in both receptor-binding and fusionsubunits of Lassa virus, illustrates the unique assembly of the arenavirus glycoproteinspike, and provides a much-needed template for vaccine design against these threats toglobal health.
Lassa virus (LASV) is the etiologic agent ofLassa fever, an often-fatal viral hemorrhagicfever that is endemic in West Africa, with anestimated 20 to 70% lethality (1, 2). The vi-rus appears to have extended its geographic
spread (3), and outbreaks in 2016 were accom-panied by demonstrated human-to-human trans-mission in Africa and Germany (4). There is noapproved Lassa fever vaccine, and the nucleosideanalog ribavirin and supportive therapy are theonly treatment options currently in use for LASVinfection. Attempts to use convalescent plasmaagainst LASV have met with variable results (5).LASV is a member of the arenavirus family, whichincludes more than 30 known pathogens thatexist on all populated continents on Earth. TheOld World family of arenaviruses contains LASV;lymphocytic choriomeningitis virus (LCMV), whichcauses febrile illness, neurological disease, and birthdefects with a 2 to 5% seroprevalence in NorthAmerica and Europe; and the 80% lethal hem-orrhagic fever virus Lujo (LUJV), which emergedin Southern Africa in 2008 (6). Arenaviruses inthe New World category include Machupo virus(MACV) and Junín virus (JUNV), the causative
agents of Bolivian and Argentinian hemorrhagicfever, respectively, as well as numerous other agentssuch as Sabía and Guanarito.As the sole antigen on the viral surface, the
arenavirus glycoprotein complex (GPC) is theprimary target of protective humoral immuneresponses (7, 8) and a focus for vaccine designefforts. The virion form of GPC is a trimer ofheterodimers, each containing the receptor-binding subunit GP1 and the transmembrane,fusion-mediating subunit GP2 (9). GPC also en-codes an unusual stable signal peptide (SSP) thathas many critical functions and is retained in thevirion as part of the complex. Specifically, it isrequired for proper processing of GPC into theGP1 and GP2 subunits and modulation of the pHof infectivity, and it may have additional chap-erone functions during GPC maturation (10–17).The GPC precursor is trafficked from the endo-plasmic reticulum to the Golgi, where it is heavilyN-glycosylated and processed by cellular proteases[signal peptidase (SPase) SKI1/SP1] into its matureform, which comprises noncovalently linked GP1,GP2, and SSP (18). GPC must also interact withERGIC-53 in the exocytic pathway in order toform infectious virions (19).The GPC trimer on the resulting virion must
engage several host receptors to mediate entryof target cells. GPC of LASV binds to a xylose–glucaronic acid sugar, called matriglycan (20);on a-dystroglycan (a-DG) (21–23); or to alter-native receptors at the cell surface (24). LASVthen enters the endocytic pathway, where itbinds to lysosome-associated membrane pro-tein 1 (LAMP1) before membrane fusion (9, 25, 26).The GPC of pathogenic New World arenaviruses
binds to transferrin receptor 1 (27) as its cel-lular receptor.Multiple structures of individual GP1 and GP2
subunits of various arenaviruses have been deter-mined (28–33), along with a GP1-GP2 complex ofLCMV (34). That structure, however, did not re-veal the organization of GPC on the arenavirussurface that is relevant for antibody neutraliza-tion and vaccine design.A study of more than 100 antibodies from hu-
man survivors of Lassa fever found that the major-ity of the neutralizing response to LASV targetedthe quaternary assembly of the prefusion GPCtrimer, rather than either subunit alone (7). Fur-ther, the majority of the neutralizing response wascontained in a single competition group, termedGPC-B. Here, we present the 3.2-Å crystal struc-ture of the prefusion GPC trimer of LASV, in com-plex with the human neutralizing antibody 37.7H,which is directed against the quaternary GPC-Bepitope. This structure reveals the first look atthe prefusion arenavirus GP trimer; suggests thatconformational changes occur in the GP1 subunit,as well as the GP2 subunit upon exposure to lowpH; and illuminates reasons why GPC must beenzymatically processed to oligomerize and bindone of its extracellular receptors. It also illuminateswhat appears to be the most vulnerable region onthe LASV trimer targeted for antibody-mediatedneutralization and suggests that such antibodiesfunction by blocking conformational changes re-quired for binding an intracellular receptor andfor fusion.
Structure determination
Structure determination of an arenavirus GPCtrimer has been previously hindered by meta-stability of the protein: the propensity of GP1and GP2 to separate and for GP2 to spring into itspostfusion, six-helix bundle conformation. Mindfulof the success of cysteine-linkage strategies for HIVand respiratory syncytial virus (35–40), we genet-ically modified the LASV glycoprotein ectodomainby (i) making the point mutations R207C andG360C (41) to covalently link GP1 and GP2 to-gether, (ii) introduction of a proline via an E329Pmutation in the metastable region of HR1 of GP2,and (iii) by replacing the native S1P GP1-GP2 cleav-age site with a furin site (RRLL to RRRR) to en-able efficient processing of the GP in DrosophilaS2 cells (fig. S1A). Among hundreds of modifi-cations screened over 10 years, these three, incombination, provided stable, crystallizable pro-tein. Size-exclusion chromatography coupled withmultiangle light scattering (SEC-MALS) and SDS–polyacrylamide gel electrophoresis analysis of theresulting protein (termed GPCysR4) demonstratesthat the GP elutes as a monomer and that theprotein is efficiently processed into GP1 and GP2subunits but that the two subunits remain asso-ciated (fig. S1, B and C). Further, enzyme-linkedimmunosorbent assay analysis with a panel ofhuman antibodies demonstrates that GPCysR4is recognized by neutralizing antibodies that re-quire native association between the GP1 and GP2subunits (7) and is not recognized by antibodiesagainst post-fusion GP2 (fig. S1D). Together, these
RESEARCH
Hastie et al., Science 356, 923–928 (2017) 2 June 2017 1 of 6
1Department of Immunology and Microbial Science, TheScripps Research Institute, La Jolla, CA 92037, USA.2Department of Microbiology and Immunology, AlbertEinstein College of Medicine, Bronx, NY, USA. 3Zalgen Labs,Germantown, MD, USA. 4Department of Pediatrics, School ofMedicine, Tulane University, New Orleans, LA, USA.5Department of Microbiology and Immunology, TulaneUniversity, New Orleans, LA, USA. 6Skaggs Institute forChemical Biology, The Scripps Research Institute, La Jolla,CA, USA.*Corresponding author. Email: [email protected]
Corrected 26 June 2017. See full text. on D
ecember 26, 2019
http://science.sciencem
ag.org/D
ownloaded from

results suggest that GPCysR4 is in its native,prefusion state.Monomeric GPCysR4 was incubated with
excess Fab 37.7H and subjected to SEC-MALSanalysis. SEC-MALS indicated the formation oftrimeric GP-Fab complexes in addition to mono-meric GP-Fab complexes (fig. S1B).Crystals of both the monomeric and trimeric
fractions of the GPCysR4-Fab 37.7H complexformed in space group P6122 and diffract to3.2 Å with a trimer of GP bound to three Fabsin the asymmetric unit (table S1 and fig. S1E).Phases were determined with an iterative ap-proach by using molecular replacement with a
related Fab structure and the LCMV GP crystalstructure (34).
Architecture of the trimer
In the crystal structure, the soluble LASV GPCysR4trimer adopts a compact tripod shape that closelymatches the tomographic reconstruction of thetrimeric GPC spike from authentic Lassa virions(9) (Fig. 1, A and B). Notably, the arenavirus GPlacks a central three-helix fusion subunit core evi-dent in other class I glycoproteins such as Ebolavirus GP (42), HIV-1 Env (36, 39), and influenzahemagglutinin (HA) (43) (fig. S2). Instead, the1775-Å2 surface area buried at the trimeric in-
terface arises from interactions between both theGP1 and GP2 subunits between monomers, par-ticularly a helices 1, 2, and 3; the C-terminal tailof GP1; and heptad repeat 1 (HR1) of GP2 (Fig. 1Cand fig. S4). Our recent work demonstrated thatmutation of either S138 or L143 (Lassa number-ing) to arginine prevented rescue of recombinantLCMV virions (34). In LASV, these residues lieat the trimeric interface on a2 and contact theGP1 C terminus of the neighboring monomer(Fig. 1C and fig. S4). Hence, contacts observed toform the trimer interface in the crystal structureare essential for virus viability.GPs of the arenavirus family are heavily gly-
cosylated. Lassa GP has 11 potential N-linkedglycosylation sites on each monomer, which to-gether comprise ~25% of the total mass of theprotein. In the trimeric structure presented here,we can now visualize the location of the majorityof these glycans (fig. S5). The glycans shield theside and lower portions of the trimer, leaving onlya few regions vulnerable to antibody binding—(i) the b-sheet face of GP1 where LAMP-1 andthe New World arenavirus receptor transferrinreceptor 1 (TfR1) bind (28, 30), (ii) three regionsof GP2, the fusion peptide, fusion loop, and HR2,and (iii) the GP1- and GP2-involving trimericinterfaces (Fig. 1D and fig. S6A). Modeling of thelarger, oligo-mannose glycans that might bepresent on the viral surface (44) reinforces thesignificant effect that glycosylation has on theneutralizing antibody evasion for the arenavirusfamily (8, 45) (fig. S6A). Further, analysis of glycan-protein interactions in this structure reveals themolecular basis for the specific and strong in-fluences of glycosylation on viral fitness (46). Forexample, Bonhomme and co-workers found thatrecombinant LCMV bearing a mutation to re-move the N79 glycan site (Lassa numbering) wasunable to be rescued (46). We find here that thisglycan packs against the fusion peptide (fig. S6B)and may shield and provide stability to this highlyhydrophobic region.
Receptor binding in the context ofthe trimer
The overall structure of LASV GPCysR4 aligns wellwith the previously determined structure of theLCMV GP monomer with a 2-Å RMSD over theentire structure and a 1-Å RMSD over the coreelements (b sheets and a helices of GP1 andall of GP2) (Fig. 2A). Differences between thetwo structures outside the core can be mapped tothree main regions: (i) the flexible loops con-necting the upper, b-sheet face and the lowera-helical face of GP1, (ii) the ~20 C-terminal res-idues of GP1, and (iii) the fusion peptide (Fig. 2A).In LCMV, the C terminus of GP1 lies in close ap-position to the N terminus of GP2. In the LASVtrimer, the C terminus is translated 30 Å, pointsaway from GP2 into the apex of the trimer, andpacks against a1 and a2 of the neighboring mono-mer (Figs. 1C and 2B). Enzymatic cleavage of GP1from GP2 is required to achieve this separationof the termini and formation of this trimeric as-sembly. Last, the fusion peptide adopts a confor-mation in the LASV trimer different from that in
Hastie et al., Science 356, 923–928 (2017) 2 June 2017 2 of 6
Fig. 1. Organization and glycosylation of the LASV trimer. (A) Cartoon representation of thetrimer from the front, side, and top. The GP1 subunit of each monomer is in a light shade and the GP2subunit in a dark shade. In the top view, spheres indicate positions of the C terminus of GP1 and theN terminus of GP2 at the trimeric interface. (B) The crystal structure of the LASV GP trimer(cartoon) docked into the tomographic reconstruction of the LASV GPC spike from fixed virions(surface, EMD-3290), in the same orientations as shown in (A). (C) GP monomer A is shownin cartoon representation and is colored by domain as in fig. S3. Structural elements involved intrimerization are indicated. GP monomers B and C are shown as surfaces. (D) The glycans visiblein the crystal structure are illustrated as atomic spheres, with those glycans attached to GP1illustrated in light shades and those attached to GP2 illustrated in dark shades.
RESEARCH | RESEARCH ARTICLE
Corrected 26 June 2017. See full text. on D
ecember 26, 2019
http://science.sciencem
ag.org/D
ownloaded from
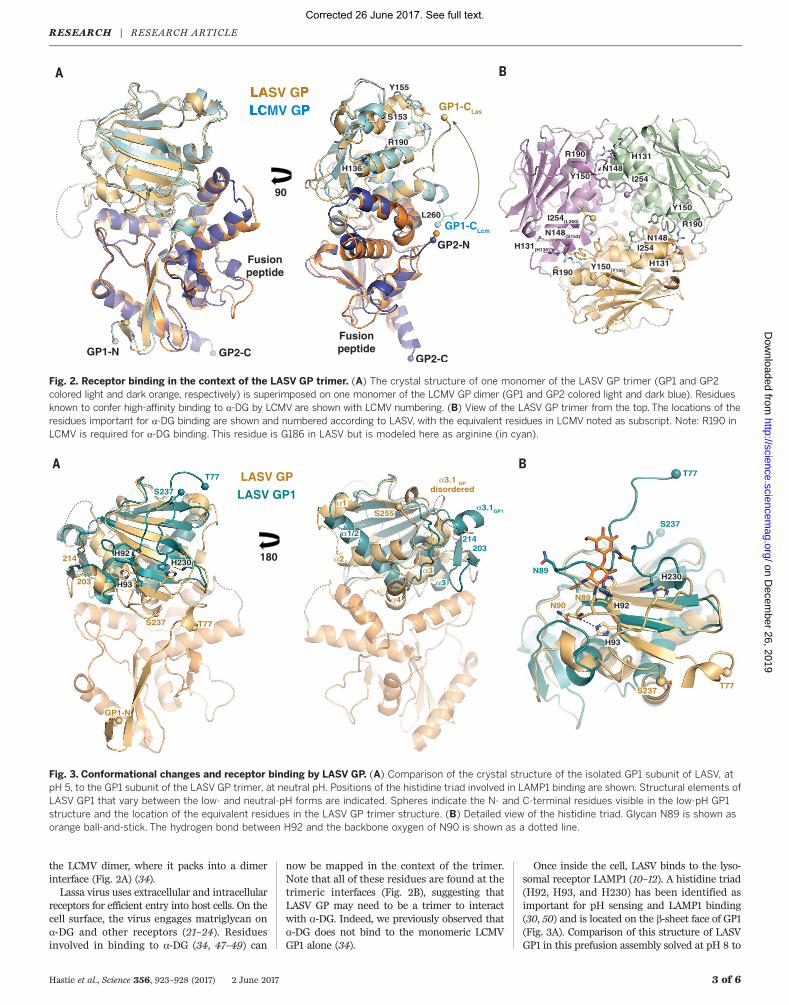
the LCMV dimer, where it packs into a dimerinterface (Fig. 2A) (34).Lassa virus uses extracellular and intracellular
receptors for efficient entry into host cells. On thecell surface, the virus engages matriglycan ona-DG and other receptors (21–24). Residuesinvolved in binding to a-DG (34, 47–49) can
now be mapped in the context of the trimer.Note that all of these residues are found at thetrimeric interfaces (Fig. 2B), suggesting thatLASV GP may need to be a trimer to interactwith a-DG. Indeed, we previously observed thata-DG does not bind to the monomeric LCMVGP1 alone (34).
Once inside the cell, LASV binds to the lyso-somal receptor LAMP1 (10–12). A histidine triad(H92, H93, and H230) has been identified asimportant for pH sensing and LAMP1 binding(30, 50) and is located on the b-sheet face of GP1(Fig. 3A). Comparison of this structure of LASVGP1 in this prefusion assembly solved at pH 8 to
Hastie et al., Science 356, 923–928 (2017) 2 June 2017 3 of 6
Fig. 2. Receptor binding in the context of the LASV GP trimer. (A) The crystal structure of one monomer of the LASV GP trimer (GP1 and GP2colored light and dark orange, respectively) is superimposed on one monomer of the LCMV GP dimer (GP1 and GP2 colored light and dark blue). Residuesknown to confer high-affinity binding to a-DG by LCMV are shown with LCMV numbering. (B) View of the LASV GP trimer from the top. The locations of theresidues important for a-DG binding are shown and numbered according to LASV, with the equivalent residues in LCMV noted as subscript. Note: R190 inLCMV is required for a-DG binding. This residue is G186 in LASV but is modeled here as arginine (in cyan).
Fig. 3. Conformational changes and receptor binding by LASV GP. (A) Comparison of the crystal structure of the isolated GP1 subunit of LASV, atpH 5, to the GP1 subunit of the LASV GP trimer, at neutral pH. Positions of the histidine triad involved in LAMP1 binding are shown. Structural elements ofLASV GP1 that vary between the low- and neutral-pH forms are indicated. Spheres indicate the N- and C-terminal residues visible in the low-pH GP1structure and the location of the equivalent residues in the LASV GP trimer structure. (B) Detailed view of the histidine triad. Glycan N89 is shown asorange ball-and-stick. The hydrogen bond between H92 and the backbone oxygen of N90 is shown as a dotted line.
RESEARCH | RESEARCH ARTICLE
Corrected 26 June 2017. See full text. on D
ecember 26, 2019
http://science.sciencem
ag.org/D
ownloaded from

the structure of LASV GP1 solved as an isolatedsubunit at pH 5 shows differences in the posi-tions and interactions made by the three histi-dines. In the neutral-pH, prefusion complex, H92packs against N89, another glycan identified byBonhomme and coauthors (46) as essential forLCMV rescue (Fig. 3B and fig. S6B). In the low-pH structure of GP1 alone, H92 is in essentiallythe same location, but its interaction partner, N89,is shifted ~10 Å away. Similarly, in the neutral-pH,prefusion complex, H93 is located on the under-side of a b sheet and makes a hydrogen bond withthe main-chain oxygen of N90. In the low-pHstructure of GP1 alone, H93 is instead oriented180 degrees away from its location in the GPtrimer and is unable to make the same hydrogenbond. Presumably, the local environment expe-rienced by these residues at neutral and low pHmay alter the structural stability of the entire GP1subunit to release its hold on GP2.Other significant structural differences be-
tween the neutral-pH complex and the low-pHGP1 include elongation and rotation of a1 anda2 from two antiparallel helices in GP1 of theneutral-pH prefusion complex to a single helixin the low-pH GP1 monomer, as well as rear-rangement of residues 200 to 214 from a helix-loop structure in the prefusion complex to atwo-stranded b sheet in the low-pH monomer
(Fig. 3A). Further, in the low-pH GP1, the visibletermini are T77 and S237, and both are orientedupward, away from where GP2 would be. In theprefusion complex, residues 59 to 75 in the N ter-minus form an extended b strand, which assembleswith strand b11 of the GP2 T loop; residue S237continues on to form a4 in the GP1 C terminus.Hence, the helices, loops, and C terminus of GP1are flexible and adopt alternate positions whennot bound by GP2.Conformational changes in GP2 are well known
during fusion. Based on these structural data, itis tempting to speculate that important confor-mational changes also occur in GP1 during low-pH–mediated LASV entry.Old and New World arenaviruses share a sim-
ilar organization of the GP1 core and are ~60%identical in GP2 by sequence, which allowsmodeling of other arenavirus receptor and anti-body interactions in the context of a GPC trimer.Arenaviruses in the New World category bindTfR1 as their cellular receptor, and a structure ofMACV GP1 in complex with a TfR1 monomer isavailable (28). MACV GP1 aligns to the prefusionLASV GP1 with a 2.5-Å RMSD for the core, andmodeling illustrates the relative positions of theTfR1-binding sites relative to each other in thearenavirus trimer (fig. S7A). Neutralizing anti-bodies are known against this TfR1-binding site
for JUNV (32), and modeling of the JUNV-Fabcomplex into the LASV trimer assembly suggeststhat three such Fab fragments could bind in thecontext of the JUNV GPC trimer (fig. S7B).
Structural definition of the anti-LASV37.7H epitope
The antibody 37.7H against LASV was isolatedfrom a Sierra Leonean survivor of Lassa fever.This antibody neutralizes viruses representing allfour known lineages of LASV in vitro (7) and offersprotection from lethal LASV challenge in guineapig (51) and potentially nonhuman primates. Theantibody simultaneously binds two GPmonomersat the base of the GP trimer, where it engages fourdiscontinuous regions of LASV GP, two in “site A”and two in “site B” (Fig. 4, A and B). Site A containsresidues 62 to 63 of the N-terminal loop of GP1and residues 387 to 408 in the T-loop and HR2of GP2. Site B contains residues 269 to 275 of thefusion peptide and residues 324 to 325 of HR1of GP2 (Fig. 4B). In total, 37.7H buries 1620 Å2
of GP: ~1000 Å2 of GP at site A and ~620 Å2 ofGP at site B. Although nearly the entire surfaceburied on GP belongs to GP2, the presence of bothGP1 and GP2 is critical for 37.7H recognition (7),likely because GP1 is required to maintain theproper prefusion conformation of GP2 for 37.7Hbinding.
Hastie et al., Science 356, 923–928 (2017) 2 June 2017 4 of 6
Fig. 4. Structural definition of the 37.7H epitope. (A) The GP trimer bound to three 37.7H Fabs from the top (left) and side (right). Cartoon representationsof the 37.7H Fab heavy and light chains are shown in dark and light gray, respectively. Each Fab binds to two GP monomers, shown as surface represen-tations, near the base of the trimer. (B) Monomers A and C are shown as surface representations, with the heavy- and light-chain CDRs shown as darkand light gray tubes, respectively. The footprint of the antibody is colored yellow, with the binding site indicated. Side-chain interactions at the GP-37.7Hinterface are magnified in the inset boxes. GP elements are colored as indicated in fig. S3. Note that only selected residues are shown for clarity.
RESEARCH | RESEARCH ARTICLE
Corrected 26 June 2017. See full text. on D
ecember 26, 2019
http://science.sciencem
ag.org/D
ownloaded from
The antibody 37.7H also recognizes the GPCof LCMV (7) but does not recognize the GPC ofthe more distantly related Old World arenavirusLUJV nor the GPC of New World arenaviruses(7). Sequence comparison among these arenavi-ruses demonstrates nearly complete sequence con-servation throughout the 37.7H epitope for allLASV lineages and LCMV (fig. S8). However, thesequences of LUJO, JUNV, and MACV GPCs arefar more divergent, particularly in HR2 of GP2,which is heavily involved in binding to 37.7H.
The 37.7H antibody neutralizes bystabilizing the prefusion GP
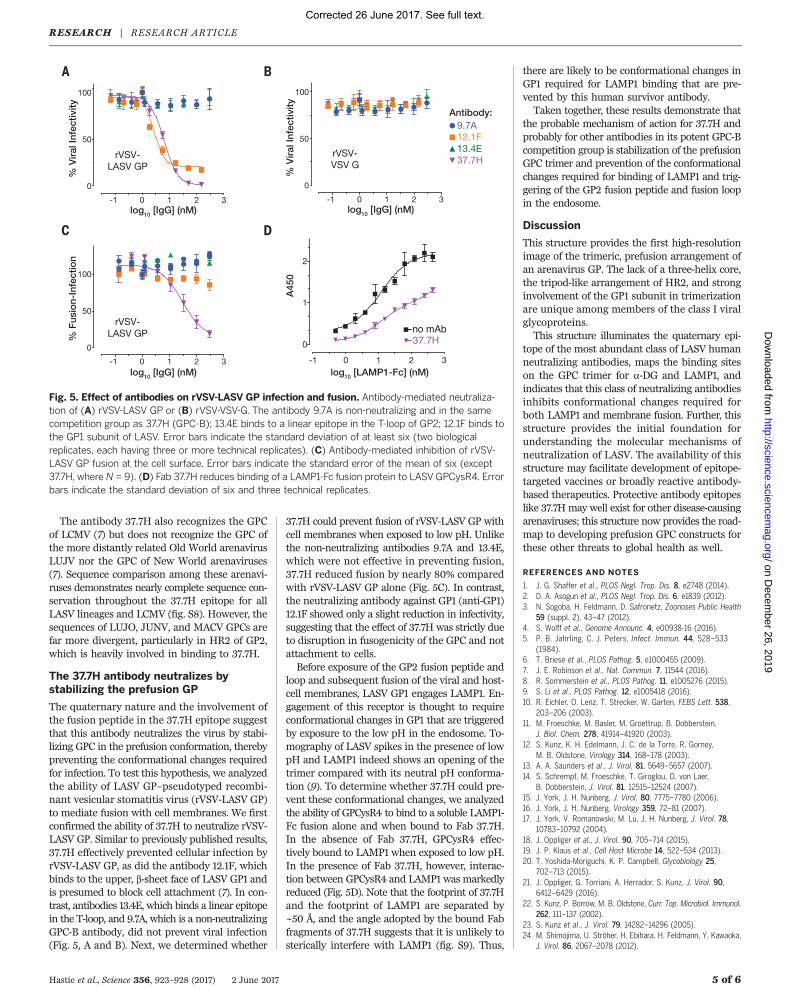
The quaternary nature and the involvement ofthe fusion peptide in the 37.7H epitope suggestthat this antibody neutralizes the virus by stabi-lizing GPC in the prefusion conformation, therebypreventing the conformational changes requiredfor infection. To test this hypothesis, we analyzedthe ability of LASV GP–pseudotyped recombi-nant vesicular stomatitis virus (rVSV-LASV GP)to mediate fusion with cell membranes. We firstconfirmed the ability of 37.7H to neutralize rVSV-LASV GP. Similar to previously published results,37.7H effectively prevented cellular infection byrVSV-LASV GP, as did the antibody 12.1F, whichbinds to the upper, b-sheet face of LASV GP1 andis presumed to block cell attachment (7). In con-trast, antibodies 13.4E, which binds a linear epitopein the T-loop, and 9.7A, which is a non-neutralizingGPC-B antibody, did not prevent viral infection(Fig. 5, A and B). Next, we determined whether
37.7H could prevent fusion of rVSV-LASV GP withcell membranes when exposed to low pH. Unlikethe non-neutralizing antibodies 9.7A and 13.4E,which were not effective in preventing fusion,37.7H reduced fusion by nearly 80% comparedwith rVSV-LASV GP alone (Fig. 5C). In contrast,the neutralizing antibody against GP1 (anti-GP1)12.1F showed only a slight reduction in infectivity,suggesting that the effect of 37.7H was strictly dueto disruption in fusogenicity of the GPC and notattachment to cells.Before exposure of the GP2 fusion peptide and
loop and subsequent fusion of the viral and host-cell membranes, LASV GP1 engages LAMP1. En-gagement of this receptor is thought to requireconformational changes in GP1 that are triggeredby exposure to the low pH in the endosome. To-mography of LASV spikes in the presence of lowpH and LAMP1 indeed shows an opening of thetrimer compared with its neutral pH conforma-tion (9). To determine whether 37.7H could pre-vent these conformational changes, we analyzedthe ability of GPCysR4 to bind to a soluble LAMP1-Fc fusion alone and when bound to Fab 37.7H.In the absence of Fab 37.7H, GPCysR4 effec-tively bound to LAMP1 when exposed to low pH.In the presence of Fab 37.7H, however, interac-tion between GPCysR4 and LAMP1 was markedlyreduced (Fig. 5D). Note that the footprint of 37.7Hand the footprint of LAMP1 are separated by~50 Å, and the angle adopted by the bound Fabfragments of 37.7H suggests that it is unlikely tosterically interfere with LAMP1 (fig. S9). Thus,
there are likely to be conformational changes inGP1 required for LAMP1 binding that are pre-vented by this human survivor antibody.Taken together, these results demonstrate that
the probable mechanism of action for 37.7H andprobably for other antibodies in its potent GPC-Bcompetition group is stabilization of the prefusionGPC trimer and prevention of the conformationalchanges required for binding of LAMP1 and trig-gering of the GP2 fusion peptide and fusion loopin the endosome.
Discussion
This structure provides the first high-resolutionimage of the trimeric, prefusion arrangement ofan arenavirus GP. The lack of a three-helix core,the tripod-like arrangement of HR2, and stronginvolvement of the GP1 subunit in trimerizationare unique among members of the class I viralglycoproteins.This structure illuminates the quaternary epi-
tope of the most abundant class of LASV humanneutralizing antibodies, maps the binding siteson the GPC trimer for a-DG and LAMP1, andindicates that this class of neutralizing antibodiesinhibits conformational changes required forboth LAMP1 and membrane fusion. Further, thisstructure provides the initial foundation forunderstanding the molecular mechanisms ofneutralization of LASV. The availability of thisstructure may facilitate development of epitope-targeted vaccines or broadly reactive antibody-based therapeutics. Protective antibody epitopeslike 37.7Hmay well exist for other disease-causingarenaviruses; this structure now provides the road-map to developing prefusion GPC constructs forthese other threats to global health as well.
REFERENCES AND NOTES
1. J. G. Shaffer et al., PLOS Negl. Trop. Dis. 8, e2748 (2014).2. D. A. Asogun et al., PLOS Negl. Trop. Dis. 6, e1839 (2012).3. N. Sogoba, H. Feldmann, D. Safronetz, Zoonoses Public Health
59 (suppl. 2), 43–47 (2012).4. S. Wolff et al., Genome Announc. 4, e00938-16 (2016).5. P. B. Jahrling, C. J. Peters, Infect. Immun. 44, 528–533
(1984).6. T. Briese et al., PLOS Pathog. 5, e1000455 (2009).7. J. E. Robinson et al., Nat. Commun. 7, 11544 (2016).8. R. Sommerstein et al., PLOS Pathog. 11, e1005276 (2015).9. S. Li et al., PLOS Pathog. 12, e1005418 (2016).10. R. Eichler, O. Lenz, T. Strecker, W. Garten, FEBS Lett. 538,
203–206 (2003).11. M. Froeschke, M. Basler, M. Groettrup, B. Dobberstein,
J. Biol. Chem. 278, 41914–41920 (2003).12. S. Kunz, K. H. Edelmann, J. C. de la Torre, R. Gorney,
M. B. Oldstone, Virology 314, 168–178 (2003).13. A. A. Saunders et al., J. Virol. 81, 5649–5657 (2007).14. S. Schrempf, M. Froeschke, T. Giroglou, D. von Laer,
B. Dobberstein, J. Virol. 81, 12515–12524 (2007).15. J. York, J. H. Nunberg, J. Virol. 80, 7775–7780 (2006).16. J. York, J. H. Nunberg, Virology 359, 72–81 (2007).17. J. York, V. Romanowski, M. Lu, J. H. Nunberg, J. Virol. 78,
10783–10792 (2004).18. J. Oppliger et al., J. Virol. 90, 705–714 (2015).19. J. P. Klaus et al., Cell Host Microbe 14, 522–534 (2013).20. T. Yoshida-Moriguchi, K. P. Campbell, Glycobiology 25,
702–713 (2015).21. J. Oppliger, G. Torriani, A. Herrador, S. Kunz, J. Virol. 90,
6412–6429 (2016).22. S. Kunz, P. Borrow, M. B. Oldstone, Curr. Top. Microbiol. Immunol.
262, 111–137 (2002).23. S. Kunz et al., J. Virol. 79, 14282–14296 (2005).24. M. Shimojima, U. Ströher, H. Ebihara, H. Feldmann, Y. Kawaoka,
J. Virol. 86, 2067–2078 (2012).
Hastie et al., Science 356, 923–928 (2017) 2 June 2017 5 of 6
Fig. 5. Effect of antibodies on rVSV-LASV GP infection and fusion. Antibody-mediated neutraliza-tion of (A) rVSV-LASV GP or (B) rVSV-VSV-G. The antibody 9.7A is non-neutralizing and in the samecompetition group as 37.7H (GPC-B); 13.4E binds to a linear epitope in the T-loop of GP2; 12.1F binds tothe GP1 subunit of LASV. Error bars indicate the standard deviation of at least six (two biologicalreplicates, each having three or more technical replicates). (C) Antibody-mediated inhibition of rVSV-LASV GP fusion at the cell surface. Error bars indicate the standard error of the mean of six (except37.7H, where N = 9). (D) Fab 37.7H reduces binding of a LAMP1-Fc fusion protein to LASVGPCysR4. Errorbars indicate the standard deviation of six and three technical replicates.
RESEARCH | RESEARCH ARTICLE
Corrected 26 June 2017. See full text. on D
ecember 26, 2019
http://science.sciencem
ag.org/D
ownloaded from

25. L. T. Jae et al., Science 344, 1506–1510 (2014).26. G. Pasqual, J. M. Rojek, M. Masin, J. Y. Chatton, S. Kunz,
PLOS Pathog. 7, e1002232 (2011).27. S. R. Radoshitzky et al., Nature 446, 92–96 (2007).28. J. Abraham, K. D. Corbett, M. Farzan, H. Choe, S. C. Harrison,
Nat. Struct. Mol. Biol. 17, 438–444 (2010).29. T. A. Bowden et al., J. Virol. 83, 8259–8265 (2009).30. H. Cohen-Dvashi, N. Cohen, H. Israeli, R. Diskin, J. Virol. 89,
7584–7592 (2015).31. S. Igonet et al., Proc. Natl. Acad. Sci. U.S.A. 108, 19967–19972
(2011).32. S. Mahmutovic et al., Cell Host Microbe 18, 705–713 (2015).33. M. L. Parsy, K. Harlos, J. T. Huiskonen, T. A. Bowden, J. Virol.
87, 13070–13075 (2013).34. K. M. Hastie et al., Nat. Struct. Mol. Biol. 23, 513–521 (2016).35. J. S. McLellan et al., Science 342, 592–598 (2013).36. J. P. Julien et al., Science 342, 1477–1483 (2013).37. A. Krarup et al., Nat. Commun. 6, 8143 (2015).38. Y. D. Kwon et al., Nat. Struct. Mol. Biol. 22, 522–531 (2015).39. D. Lyumkis et al., Science 342, 1484–1490 (2013).40. R. W. Sanders et al., PLOS Pathog. 9, e1003618 (2013).41. Single-letter abbreviations for the amino acid residues are
as follows: A, Ala; C, Cys; D, Asp; E, Glu; F, Phe; G, Gly; H, His;I, Ile; K, Lys; L, Leu; M, Met; N, Asn; P, Pro; Q, Gln; R, Arg; S, Ser;T, Thr; V, Val; W, Trp; and Y, Tyr. The R207C mutation is Cysreplacing Arg207.
42. J. E. Lee et al., Nature 454, 177–182 (2008).43. S. J. Gamblin et al., Science 303, 1838–1842 (2004).44. A. R. Goncalves et al., J. Virol. 87, 11504–11515 (2013).45. J. B. McCormick, S. W. Mitchell, M. P. Kiley, S. Ruo,
S. P. Fisher-Hoch, J. Med. Virol. 37, 1–7 (1992).
46. C. J. Bonhomme, K. A. Knopp, L. H. Bederka, M. M. Angelini,M. J. Buchmeier, PLOS ONE 8, e53273 (2013).
47. S. C. Smelt et al., J. Virol. 75, 448–457 (2001).48. B. M. Sullivan et al., Proc. Natl. Acad. Sci. U.S.A. 108, 2969–2974
(2011).49. M. N. Teng, P. Borrow, M. B. Oldstone, J. C. de la Torre, J. Virol.
70, 8438–8443 (1996).50. H. Cohen-Dvashi, H. Israeli, O. Shani, A. Katz, R. Diskin, J. Virol.
90, 10329–10338 (2016).51. R. W. Cross et al., Antiviral Res. 133, 218–222
(2016).
ACKNOWLEDGMENTS
The authors acknowledge the Viral Hemorrhagic Fever ResearchConsortium, the Viral Hemorrhagic Fever ImmunotherapeuticConsortium (VIC), and NIH grant 1U19AI109762-01 (E.O.S.,K.C., J.E.R., L.M.B., and R.F.G); NIH grant R21 AI116112 (E.O.S.);NIH Training Program grant T32 GM007491 (L.M.K.); NIH contractHHSN272200900049C (J.E.R., E.O.S., R.F.G., and L.M.B.); anInvestigators in Pathogenesis of Infectious Diseases award fromthe Burroughs Wellcome Fund (E.O.S.); and beamline 12-2 of theStanford Synchrotron Radiation Lightsource (Palo Alto, CA), aswell as beamlines 19-ID, 23-ID-B, and 23-ID-D of the AdvancedPhoton Source (Argonne, IL) for data collection. We are alsograteful to S. Whelan, Harvard Medical School, for providing theLAMP1-Fc expression plasmid used in these studies. K.M.H.designed and cloned the constructs, produced recombinantprotein, crystallized the complex, built and refined the model,performed the receptor-binding experiments, analyzed the data,and wrote the manuscript; M.A.Z. cloned the constructs andproduced recombinant protein; L.M.K. and K.C. performed the VSV
neutralization and fusion assays; J.E.R., L.M.B., M.L.H., M.M.R., andR.F.G generated and produced antibodies used throughout thestudies; and E.O.S. analyzed the data and wrote the manuscript. Allauthors commented on the manuscript. Crystallographic structurefactors and coordinates are deposited into the Protein Data Bankwith accession number 5VK2. Use of the Stanford SynchrotronRadiation Lightsource (SSRL), SLAC National AcceleratorLaboratory, is supported by the U.S. Department of Energy (DOE),Office of Science, Office of Basic Energy Sciences, under contractDE-AC02-76SF00515. The SSRL Structural Molecular BiologyProgram is supported by the DOE Office of Biological andEnvironmental Research and by the National Institute of GeneralMedical Sciences (NIGMS), NIH (including P41GM103393). Thisresearch used resources of the Advanced Photon Source, a DOEOffice of Science User Facility operated for the DOE Office ofScience by Argonne National Laboratory under contract DE-AC02-06CH11357. The contents of this publication are solely theresponsibility of the authors and do not necessarily represent theofficial views of NIGMS or NIH. Correspondence and requests formaterials should be addressed to E.O.S.. This is manuscript 29458from The Scripps Research Institute.
SUPPLEMENTARY MATERIALS
www.sciencemag.org/content/356/6341/923/suppl/DC1Materials and MethodsFigs. S1 to S9Table S1References (52–69)
9 January 2017; accepted 28 April 201710.1126/science.aam7260
Hastie et al., Science 356, 923–928 (2017) 2 June 2017 6 of 6
RESEARCH | RESEARCH ARTICLE
Corrected 26 June 2017. See full text. on D
ecember 26, 2019
http://science.sciencem
ag.org/D
ownloaded from

Structural basis for antibody-mediated neutralization of Lassa virus
M. Branco, James E. Robinson, Robert F. Garry and Erica Ollmann SaphireKathryn M. Hastie, Michelle A. Zandonatti, Lara M. Kleinfelter, Megan L. Heinrich, Megan M. Rowland, Kartik Chandran, Luis
DOI: 10.1126/science.aam7260 (6341), 923-928.356Science
, this issue p. 923Sciencemechanisms and antibody neutralization and provides a template for vaccine design.GPC bound to a neutralizing antibody from a human survivor of the disease. The structure gives insight into viral entry
report the high-resolution structure of the trimeric ectodomain ofet al.GPC that engages host cell receptors. Hastie glycoproteinRNA virus in the arenavirus family. There is no available vaccine. The only antigen on the viral surface is the
Lassa fever is a severe, sometimes fatal, hemorrhagic disease. It is caused by the Lassa virus, a single-strandedA step on the path to a Lassa vaccine
ARTICLE TOOLS http://science.sciencemag.org/content/356/6341/923
MATERIALSSUPPLEMENTARY http://science.sciencemag.org/content/suppl/2017/05/31/356.6341.923.DC1
CONTENTRELATED http://stm.sciencemag.org/content/scitransmed/6/253/253ps10.full
REFERENCES
http://science.sciencemag.org/content/356/6341/923#BIBLThis article cites 66 articles, 27 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.ScienceScience, 1200 New York Avenue NW, Washington, DC 20005. The title (print ISSN 0036-8075; online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Copyright © 2017, American Association for the Advancement of Science
on Decem
ber 26, 2019
http://science.sciencemag.org/
Dow
nloaded from