storage of root reserves in rhodes grass introduction aldous
TRANSCRIPT

STORAGE OF ROOT RESERVES IN RHODES GRASS
H. WEINMANN
(WITH FIVE FIGURES)
Introduction
It is a well-known fact that the roots of many perennial herbaceous plantsfunction as organs of storage, even where there are no special morphologicalmodifications of the root system. Root-storage phenomena in herbaceousplants have been studied in detail by a number of workers. GRABER andassociates (7) found that in lucerne, polysaccharides and nitrogen increasedin percentage and amount in the roots during the mature stages whereasspring growth was associated with a decrease in the percentage of nitrogenand soluble sugars in both tops and roots. The authors conclude that "stor-age is greatly accelerated during the periods of blossoming and seed forma-tion. At such periods, the plant produces reserve foods in excess of theamounts used in vegetative extension and a surplus is stored in the roots.. . .
WILLARD (20), likewise working on lucerne, made similar observations.ALDOUS (1) found that in the roots of Vernonia baldwinii, Andropogon sco-parius, Symphoricarpus vulgaris, Rhus glabra and Verbena stricta the per-centage of nitrogen, total sugars and starch in general decreased to aboutthe time of flowering, after which considerable increases of these constituentswere recorded. ALDOUS thinks that after the time of flowering "the plantsbegin to restore the reserves which must have been drawn on to produce thetop growth." Similarly, GRANDFIELD (8) reports a 23.8 per cent. increasein the amount of total carbohydrates and a 16.8 per cent. increase in theamount of total nitrogen in the roots of Verbena stricta during the periodbetween budding and maturity, and approximately the same increases inthe roots of Solidago rigida and Vernonia baldwinii. According to BEWSand VANDERPLANK (5), the percenitage of total carbohydrates increaseddurinig the entire season and that of total soluble sugars during wiinter only,in the corms of Hypoxis rooperi, while the percentages of the same compoundsdecreased in the leaves between spring and autumn. The authors think,however, that in this plant "the large carbohydrate reserves are drawn uponito only a very slight extent when new leaves are produced in spring.
More recently data have been collected with regard to the storage andutilization of root reserves in various grasses. STURKIE (16) found that therootstocks of Sorghurn halepense were developed after the maturing of aerialplant parts; this was confirmed by SAMPSON and MCCARTY (14) in Stipapulchra, where an accumulation of carbohydrate reserves in the roots wasobserved during maturation. In the roots of Digitaria eriantha and Themeda
467
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
triandra, BEWS and BAYER (4) found only somewhat irregular fluctuationsof sugars anid polysaccharides, whereas in Elyonurus ambiguus and Muehlen-bergia gracilis, MCCARTY (9) recorded an accumulation of sugars and starchin the stem bases, rhizomes and roots at the end of the season with the cessa-tion of growth. That mineral elements may also be included in the substancesstored in the roots, is shown by the work of RICHARDSON, TRUMBLE andSHAPTER (12, 13) on Lolium subulatum and Phalaris tuberosa. In bothgrasses, considerable losses of nitrogen, phosphorus, and potassium occurredfrom the herbage portions during maturity; amounting in Phalaris tuberosato 36, 22, and 12 per cent. of the total intake of N, K20, and P20,5 respectively.In the case of Phalaris tuberosa, the losses of nitrogen and phosphorus couldbe accounted for by translocation to the basal internodes and to the root sys-tem; potassium was actually lost from all portions of the plant, amountingto 27.8 per cent. of the amount found in the entire plant shortly beforeflowering.
The writer, who has been engaged for several years in the study of thestorage of root reserves in some South African Highveld grasses, found thatin the roots of Trachypogon plumosus, Tristachya hispida and Digitaria tri-cholaenoides, the percentage of nitrogen and total sugars (17) as well as ofphosphorus (unpublished data) decreased during the growing period andincreased in autumn and winter. In connection with a study of the growthrhythm of the same three species and also of Cynodon dactylon, Brachiariaserrata and Eragrostis chalcantha, ALTONA (2) was recently able largely toconfirm the results hitherto obtained by examining the percentages of nitro-gen and total sugars in the roots of these grasses at their various stages ofdevelopment. In general, the curves of nitrogen and total sugar content ofthe roots show a definite seasonal trend reaching a maximum in mid-winterand dropping at the time of shooting to reach a minimum at the time offlowering.
Seasonal chemical changes in the roots coincide largely with correspond-ilng changes in the chemical composition of the herbage in the opposite direc-tion. Decreases in the percentage of nitrogen, phosphorus, potassium andother constituents in the herbage during the growing season have beenreported by many authors in various countries. When the total amountsof these constituents in the herbage per unit area are calculated, it is shown,however, that absorption continues for a considerable part of the growingseason. While the decrease of the percentage of nitrogen, phosphorus andpotassium during the greater part of the growing season is merely due to theprogressive accumulation of carbohydrates, actual losses from the herbageof total amounts of these constituents occur during autumn when the per-centages of the same constituents in the roots increase (17).
468
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.

WEINMANN: STORAGE OF ROOT RESERVES
Naturally, mere pereentage figures are not fully conclusive for the char-acterization of such processes as translocation and storage where variousopposite processes or processes having opposite effects may go on at the sametime. Thus, prolonged absorption of mineral elements by the roots from thesoil may mask translocation of the same elements from shoots to roots,whereas an exceedingly high inerease of one group of constituents may undercertain circumstances result in a decrease of the percentage of another group,though that group may actually inerease in amount. Only a determinationof the total amounts of the various constituents present in shoots and rootsat different stages of growth and maturation will indicate to what- extentconstituents have been traiislocated from shoot to root and vice versa. This,however, is rarely possible with plants growing in the field, sinee a collectingof the whole root bulk is mostly impracticable. For a more detailed investi-gation of the processes of absorption, translocation, and storage, pot experi-ments conducted under controlled conditions are more suitable.
The present contribution deals with the storage of root reserves in Chlorisgayana KUNTH, Rhodes Grass, which was grown under conitrolled conditionsin pots and was harvested at different stages of maturation.
Materials and methodsFor this experiment cylindrical metal tins approximately 2 feet in height
and with an open surface of 1250 cm2 were used. These tins were firstbrought to equal weight by placing a few quartz stones in the bottom, andwere then filled with ordinary veld soil collected at Frankenwald, the Botani-cal Research Station of the Witwatersrand University, niear Johannesburg,South Africa, where the experimelnt was conducted.
The lower half of the tins was filled with 29.2 lb. of sub-soil consistingof coarse decomposed granite, the upper half with 32.2 lb. of top-soil (thefigures expressinig absolute dry weight). Top-soil as well as sub-soil werethoroughly mixed before being distributed into the single pots; the top-soilwas also sieved through a wide-meshed sieve in order to exclude coarserstones. The top-soil used was an acid loamy sand, poor in organic matterand nutrients, particularly in nitrogen and phosphorus. The dry top-soil(as used) contained 29.85 per cent. stones (determined by using a sieve of 20mesh-per-inch) and 2.35 per cent. organic matter (loss on ignition). Theresults of chemical and mechanical anialyses of the top-soil fine earth aregiven in table I.
For the mechanical analysis the Boyoucos hydrometer method was used(6), nitrogen was determined by the ordinary Kjeldahl method, phosphorusand potassium were extracted by treating 100 gui. of air-dry soil with 200ml. of 10 per cent. HCl on a boiling water bath for 3 hours. The maximumwater retaining capacity of the top-soil (as used) was 34.0 per cent. and that
469
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
TABLE ICHEMICAL AND MECHANICAL ANALYSES OF TOP-SOIL FINE EARTH. RESULTS
GIVEN IN PERCENTAGE OF DRY MATTER
MECHANICAL ANALYSES
PARTICLE SIZE IN MILLIMETERS
SAND SILT CLAY TOTAL COLLOIDS1.0-0.05 0.05-0.005 < 0.005 < 0.008
76.5 8.1 15.4 16.0
CHEMICAL ANALYSES
NITROGEN PHOSPHORIC POTASHOXIDE
0.107 0.015 0.087
of the sub-soil was 29.7 per cent., equalling approximately 20 lb. of waterper pot.
Thirty six tins were filled with soil as described above; one series of 12 tinsreceived no fertilizer treatment (0) ; another series of 12 tins received a ferti-lizer treatment in the form of sulphate of ammonia, phosphate, and potashsalt (PNK) ; and the third series of 12 tins were given the same treatment asthe second with the difference that a threefold quantity of ammonium sul-phate was given (PN3K). The actual amounts of nutrients applied per potwere as follows:
P = 6 gm. raw rock and superphosphate mixture (27 per cent.)= 1.44 gm. P205;
K = 1.2 gm. muriate of potash (60 per cent.)= 0.72 gm. K20;
N =3 gm. sulphate of ammonia (21.1 per cent.)= 0.63 gm. N;
N3=9 gmi. t t "= 1.89 gm. N.
The phosphate was distributed and mixed into the soil surface about one inchdeep on Dec. 15, 1936; sulphate of ammonia and potassium chloride wereapplied in the form of a solution on Dec. 17; on the 18th a number of seedsof Rhodes Grass were sown into each pot. Germination was satisfactory andafter about one month the seedlings were singled out so that finally ten plantsremained in each pot.
In contrast to the plants of the PNK and PN3K series the plants of the 0series were very poor from the beginning, and it was feared that they woulddie on account of a marked deficiency of nutrients. In order to prevent this,
470
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.

WEINMANN: STORAGE OF ROOT RESERVES
to ten pots of this series a small amount of PNK equalling 1/20 of the properPNK treatment was given on January 25, 1937; two pots of this series werekept entirely unfertilized as an absolute control. The whole experiment thusconsisted altogether of four treatments:
0 pnk PNK PN3KNumber of pots ............ 2 10 12 12
It later became evident, however, that the plants of the 0 series overcametheir first set back and did not die; on the other hand, the small pnk treat-ment was not able to produce any significant difference as compared with the0 plants. Practically, both of these series can be regarded as control series;the plants of both series remained true "deficiency" plants.
The plants were kept in the open air but were, in the early stages ofgrowth, sheltered against heavy storms by means of a movable roof of corru-gated iron which was removed as soon as rain had ceased. For watering,collected rain water was used and the water content of the pots was kept moreor less constant by frequent weighing. The water supply was controlledaccording to the requirements of the growing plants. Thus, when the seedswere sown into the pots the soil possessed a water content equalling 33 percent. of the maximum water retaining capacity of the pots; the water con-tent was raised to 50 per cent. of the M.W.R.C. on Dec. 30, to 60 per cent.on Feb. 13, to 70 per cent. on March 2, and to 75 per cent. on April 3. OnApril 17, when the plants approached maturity, all watering was stopped andthe pots were allowed to dry out gradually.
The plants were harvested at three different stages of maturity; namely,three to four pots of each series on March 23, April 17, and May 18, 1937.The two pots of the absolute control series (0) were harvested on the lastdate as well. The total herbage of each pot was first cut off directly at thebase including also the subaerial parts of the stems, and the whole root systemwas collected by washing it free from the surrounding soil. Care was takento avoid losses of roots, and at the same time to shorten the procedure ofwashing as much as possible, in order to avoid any risk of loss of constituentsfrom the tissue by leaching. Roots and shoots were then air dried in a glasshouse. When air-dry, their weight was determined; hereafter the materialof one treatment and harvest-roots and shoots separately-was united andfinely ground. The ground material was stored in airtight bottles and keptfor the various determiniations.
The following constituents were determined in shoots and roots: Totalsugars, total hydrolyzable carbohydrates, nitrogen, phosphorus, potassiumand soluble ash.
Total sitgars.-5 gm. of the airdry substance were (after the addition of1 gem. CaCO3) extracted twice with 200 ml. of 95 per cent. alcohol for a
471
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
period of two hours under a reflux condenser. The residue was then re-moved by filtering and the alcohol distilled off under reduced pressure; theremaining syrup was taken up with distilled water, cleared with neutral leadacetate, and deleaded with potassium oxalate. The extract was then hydro-lyzed by boiling with HCl under a reflux condenser for 11 hours (2.5 ml.conc. HCl were added to 100 ml. solution). Hereafter the solution wasneutralized and made up to 500 ml. volume. The reducing power of thisextract was then estimated by the MUNSON and WALKER method (10) fol-lowed by the BERTRAND method for the estimation of cuprous oxide (3).From the amount of copper precipitate the glucose equivalent was found byreference to the tables of MUNSON and WALKER.
Total hydrolyzable carbohydrates.-As recommended by BEWS and VAN-DERPLANK (5), 5 gm. of airdry substance were hydrolyzed with 200 ml. of1.1 per cent. HCl in a beaker, covered with a watch glass, on a boiling waterbath for 41 hours. The hydrolyzed and filtered extract was then neutralized,cleared and deleaded, made up to 500 ml. volume, and its reducing powerdetermined as described above. The results were also expressed as glucose.
Nitrogen.-Nitrogen was estimated by the ordinary Kjeldahl method,using a mixture of K2SO4 and CuSO4 (9: 1 parts by weight) as a catalystin the digestion.
Phosphorus.-A sample of the plant material was incinerated after theaddition of saturated lime water in order to avoid losses of organic phos-phorus in the ignition. After removal of the silica, phosphates were pre-cipitated in the nitric acid extract of the plant ash as ammonium phospho-molybdate according to LORENZ (18).
Potassium.-Potassium was determined as potassium perchlorate in theHCl extract of the plant ash after removal of silica, sulphates, phosphates,calcium, iron, aluminium, and magnesium (18).
Soluble ash.-A weighed ash sample of the plant material was treatedwith hot HCl. The liquid was then filtered off and the remaining insolubleresidue (SiO2) was washed, ignited, and weighed. The difference total plantash minus SiO2 was reported as soluble ash.
Dry matter.-The dry matter content was determined by drying a sepa-rate sample in an electric oven for 12 hours at a temperature of 1050 C. Allresults of the chemical determinations were expressed as percentages of drymatter.
Results
YIELDS AND GENERAL DEVELOPMENT OF THE PLANTS
In their general development and growth rhythm, the plants of the PNKand PN3K series were soon far in advance of the plants of the 0 and pnkseries. The leaves of the PN3K plants had a darker green color than the
472
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.

WEINMANN: STORAGE OF ROOT RESERVES 473
leaves of the other plants, owing to their high nitrogen supply; the plants of0 and pnk series, on the other hand, were poor and yellowish, and were notable to form stems and infloresceneces. Both, PNK and PN3K plants, pro-duced stems and inflorescences, but the PN3K plants did not only form morebulk than the PNK plants but produced also a larger number of infloreseencesper plant (see fig. 1). The final average number of infloreseences formed by
I-
FIG. 1. Infloreseences produced under varying nutrients. From left to right:pnk series: received 1/20 of proper PNK treatment;PNK series: received proper PNK treatment;PN,K series: three-fold quantity of ammonium sulphate.
(Photographed April 17, 1937).
ten plants (as counted on April 17 after the flowering period was finished)was 10.5 and 27.2 for PNK and PN3K planits respectively. Maturation wasaccelerated by both these fertilizer treatments: the plants of PNK and PN3Ktreatment possessed relatively early (middle of March) a fairly large propor-tion of dead, yellowish leaves-PN3K plants again more than PNK plants-whereas 0 and pnk plants remained in the green, vegetative stage through-out the growing season, though yellowing occurred here to a certain extent,probably owing to deficiency of nutrients.
Three to four pots of each series were harvested at three different stagesof maturation, namely on March 23, April 17, and May 18, 1937, respec-tively. At the first date, flowering was just commencing, while at the seconddate the flowering period was definitely finished. The first frosts wererecorded on April 22, 24, and 29, followed by frosts on every day during Maywith the exception of a warm spell from the 6th to the 14th. The thirdharvesting date (May 18), was, therefore, well after the growing season had
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.
PLANT PHYSIOLOGY
ceased. At this last date, the PN3K plants possessed only very few greenleaves, the PNK plants a few more, whereas the plants of 0 and pnk serieswere still almost entirely green. While it is frequently observed thatnitrogen in excess retards maturation, the earlier maturation of the PN3Kplants in this case may be partly due to the greater daily fluctuations in thewater content of the pots of these better developed plants owing to theirhigher total transpiration. Seeding was in full swing near the last harvest-ing date and many seeds had fallen before this day. Further losses of seedsin handling the plants were unavoidable to a certain extent but, as far aspossible, seeds were collected and included in the samples.
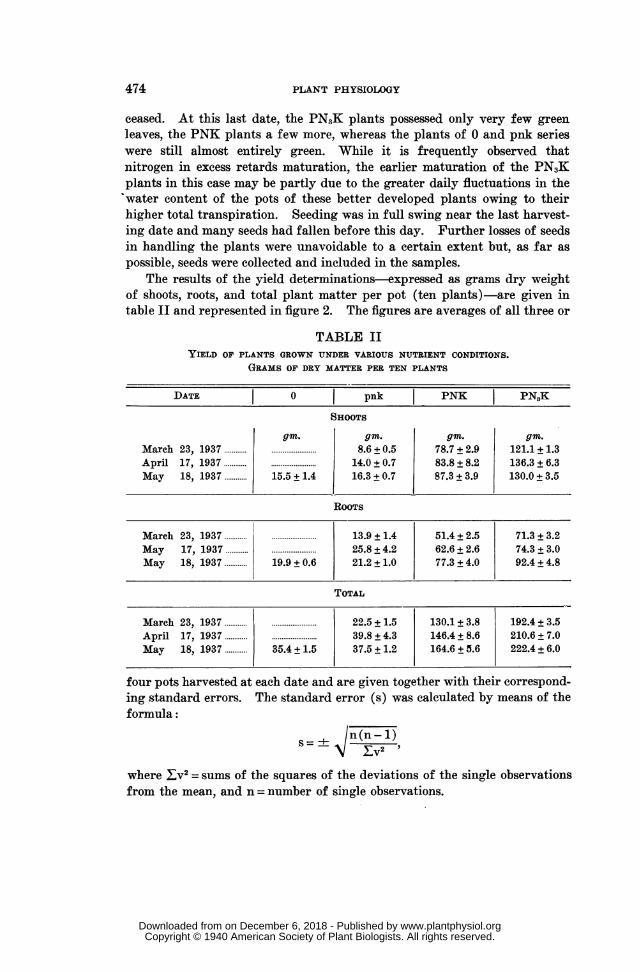
The results of the yield determinations-expressed as grams dry weightof shoots, roots, and total plant matter per pot (ten plants)-are given intable II and represented in figure 2. The figures are averages of all three or
TABLE IIYIELD OP PLANTS GROWN UNDER VARIOUS NUTRIENT CONDITIONS.
GRAMS OF DRY MATTER PER TEN PLANTS
DATE 0 pnk PNK PN3K
SHOOTS
gin. gm. gm. gm.March 23, 1937 ......................... 8.6 + 0.5 78.7 + 2.9 121.1 + 1.3April 17, 1937 14.0 ± 0.7 83.8 + 8.2 136.3 + 6.3May 18, 1937 ....... 15.5 + 1.4 16.3 + 0.7 87.3 + 3.9 130.0 + 3.5
ROOTS
March 23, 1937......... ..... .......
13.9+
1.4 51.4+
2.5 71.3 + 3.2
May 17, 1937 25.8 + 4.2 62.6 + 2.6 74.3 + 3.0May 18, 1937 ......... 19.9 + 0.6 21.2 + 1.0 77.3 + 4.0 92.4 + 4.8
TOTAL
March 23, 1937 22.5 + 1.5 130.1 + 3.8 192.4 + 3.5April 17, 1937 ......... ....................... 39.8 + 4.3 146.4 + 8.6 210.6 + 7.0May 18, 1937 35.4 + 1.5 37.5 ± 1.2 164.6 + 5.6 222.4 + 6.0
four pots harvested at each date and are given together with their correspond-ing standard errors. The standard error (s) was calculated by means of theformula:
s-+ Jn(n-1)
where Zv2 = sums of the squares of the deviations of the single observationsfrom the mean, and n = number of single observations.
474
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.

WEINMANN: STORAGE OF ROOT RESERVES
7 nilJbllx
FIG. 2. Yields; grams of dry matter per ten plants.
A comlparison of the yields of the three sampling dates shows that withregard to the shoots, growth was practically finished on April 17, no signifi-cant difference being betweenl the yields of April 17, 1937 and May 18, 1937with the possible exception of the pnk plants which show also during thisperiod a slight and perhaps significant increase of 2.3 gm.+1 0.99. Thegrowth rhythm was, however, different in the roots-at least in PNK andPN,K plants-which also increased in the very last part of the growingseason, i.e., after flowering and durinlg formation of seeds. The changes inthe root system between April 17 and May 18 were as follows:
in pnk a decrease of 4.6 gin. ± 4.3 (=- 17.8 per cent.);in PNK an increase of 14.7 gm. -+- 4.8 (=+23.5 per cent.);in PN,K an increase of 18.1 gin. ± 5.7 (=+±24.4 per cent.).
The decrease is small and uncertain for its relatively high error in thepnk plants. At any rate, the pnk plants were, on account of a generaldeficiency of nutrients, hinldered in their normal development. But signifi-cant increases of the root system occurred in the well-fertilized plants duringthe late stages of maturation, when aerial growth had already ceased. Thisis in agreemenlt with observations of several other mvorkers already mentioned.
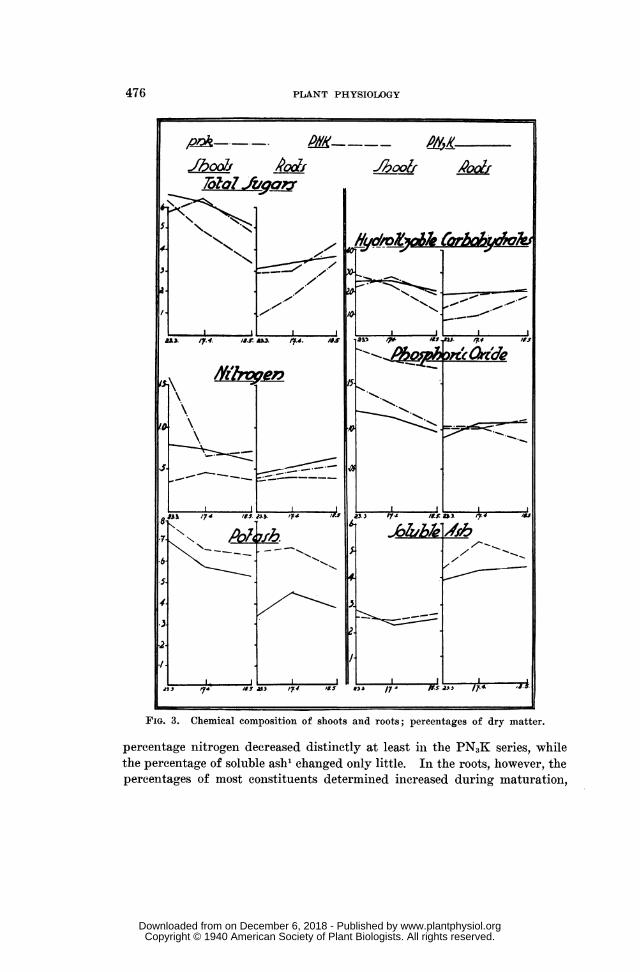
CHEMICAL CHANGES IN THE PLANTS DURING MATURATION
The results of the chemical determinations are given in table III and figure3. The figures show that in the shoots the percentage of most constituentsdetermined decreased during maturation. This is particularly distinct withreg,ard to sugars, hydrolyzable carbohydrates, phosphorus and potassium1;
1 Potassium and soluble ash could not he determined in the pnk series owing to lackof sufficient material.
W-..
475
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.
PLANT PHYSIOLOGY
FIG. 3. Chemical composition of shoots and roots; percentages of dry matter.
percentage nitrogen decreased distinctly at least in the PN3K series, whilethe percentage of soluble ash1 changed only little. In the roots, however, thepercentages of most constituents determined increased during maturation,
476
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.
WEINMANN: STORAGE OF ROOT RESERVES
TABLE IIICHEMICAL COMPOSITION' OF SHOOTS AND ROOTS.EXPRESSED AS PERCENTAGE OF DRY MATTER
pnk PNK PN3KCONSTITUENT DATE
SHOOTS ROOTS SHOOTS ROOTS SHOOTS ROOTS
Total 3/23/37 5.80 0.97 6.52 2.90 6.55 3.04SUTarS 4/17/37 6.34 1.87 4.72 2.97 6.25 3.31
Sugars 5/18/37 4.77 3.38 3.32 4.37 5.06 3.73
Hydrolyz. 3/23/37 23.25 8.06 29.50 14.00 25.50 20.65Carbohy- 4/17/37 27.20 10.17 23.75 20.27 26.50 21.00drates 5/18/37 20.75 19.20 11.95 21.90 21.25 21.75
3/23/37 1.55 0.41 0.35 0.36 0.81 0.47Nitrogen 4/17/37 0.65 0.48 0.48 0.40 0.72 0.52
5/18/37 0.73 0.53 0.37 0.39 0.59 0.62
Phosphoric 3/23/37 0.149 0.104 0.192 0.096 0.122 0.087Oxide 4/17/37 0.127 0.097 0.178 0.096 0.113 0.099OXide 5/18/37 0.097 0.083 0.167 0.108 0.093 0.105
3/23/37 .. 0.77 0.63 0.69 0.33Potash 4/17/37 0.65 0.64 0.56 0.44
5/18/37 .. 0.62 0.55 0.52 0.36
Soluble 3/23/37 2.52 4.28 2.74 3.90Ash 4/17/37 2.38 5.27 2.20 4.19
5/18/37 2.53 4.69 2.45 4.35
though in some cases the changes were considerably smaller than in theshoots. Thus, in the roots of the PN3K plants the percentages of sugars andhydrolyzable carbohydrates increased only slightly. The same holds true fornitrogen in all series and for phosphorus in the PNK and PN3K series, whilethe percentage of potassium decreased in the roots of both well fertilizedseries towards the end of maturation and that of soluble ash in the PNKseries only.
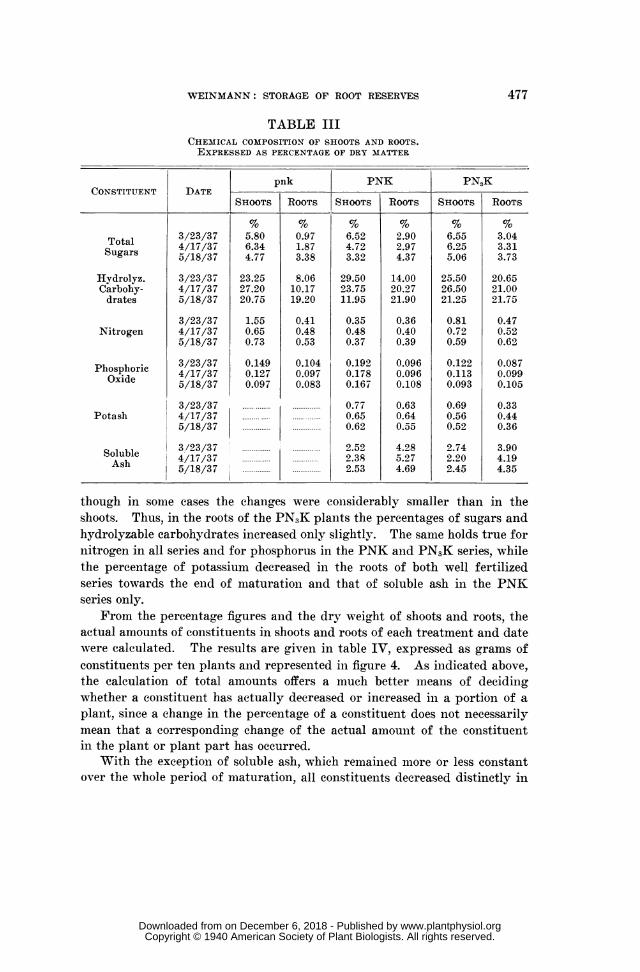
From the percentage figures and the dry weight of shoots and roots, theactual amounts of constituents in shoots and roots of each treatment and datewere calculated. The results are given in table IV, expressed as grams ofconstituents per ten plants and represented in figure 4. As indicated above,the calculation of total amounts offers a much better means of decidingwhether a constituent has actually decreased or increased in a portion of aplant, since a change in the pereentage of a constituent does not necessarilymean that a corresponding change of the actual amount of the constituentin the plant or plant part has occurred.
With the exception of soluble ash, which remained more or less constantover the whole period of maturation, all constituents decreased distinctly in
477
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.
PLANT PHYSIOLOGY
* CA Om 01
rz1i:~=~~ 000Z'-- '-~ r-
r--l - 0)000C4 CA C"C] C] CN000mr-o) o> o,4 * *
(= C CJt6 t6 t-:
iitInC(000 000 000iCA
-~1.4 r-l-i(
* 0 t-
lz ) t-.:00 ti
*qCAtC]4 IR Qs C'i
0 t0 t-C: e CA
,- 0t0 .1 L..O oi-
c 00 -m m t-*~ *; C
0O C000
CAC]: C; C;010101
000
t-
000Ll
cy "I t-
C) C>
010m i!CYi0~
CO-1 in In
0- c0~ 1t-O ,-0C)1Y 0- o00 'qC 0CIC)
o 449 t-oi~ &55 00
*tcOCO-! O:~ll
ZZ l~ma
01 cm "licCOOCCA01q
00 0 CJ01
C5 C;- O COr--q
- CO- 1C4 1C4000
Ll
oE 0'--0 0 0
liM 0 t GC>C> 00
0 --O- 0$ 00 m0 00 .
o P I0 i4to 000 000
20>000C1ie 000 000
cC~-
-n 4.c-4c
0
EH o
CC+9 O ~ +
0z2 P-0CT
478
0H
E-400
EzGo
E-40H.-
z4
z
ME-400m
tz
E-4 Z~
0.
M pgED P4
4 ¢pM O
o M
E¢
E-4
z00
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.
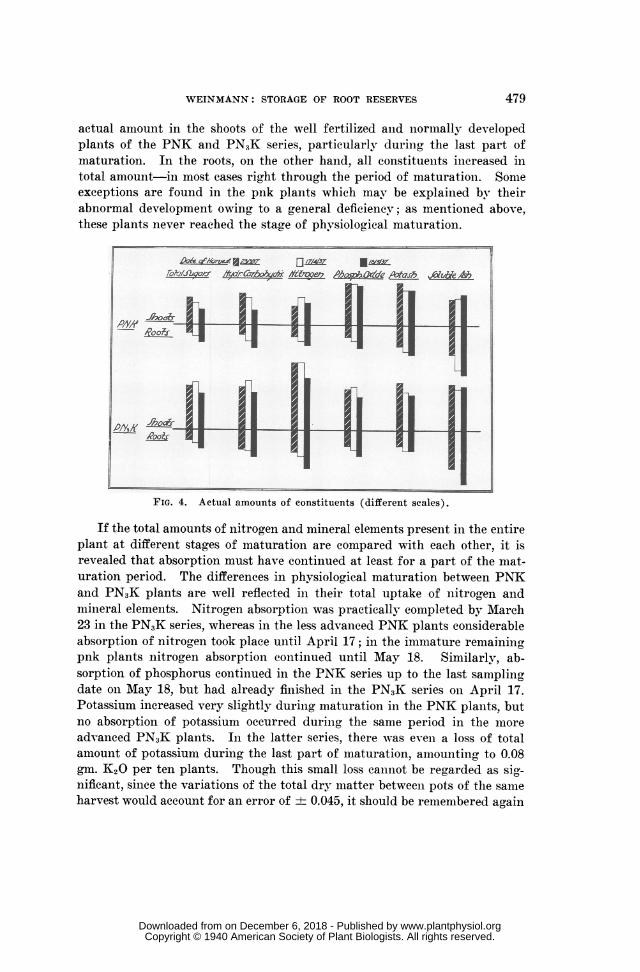
WEINMANN: STORAGE OF ROOT RESERVES
actual amount in the shoots of the well fertilized and lnormally developedplants of the PNK and PNIK series, particularly during the last part ofmaturation. In the roots, on the other hand, all constituelnts iniereased intotal amount-in most cases right through the period of maturation. Someexceptions are found in the pnk plants which may be explained by theirabnormal development owing to a general deficiency; as mentioned above,these plants never reached the stage of phvsiological maturation.
! Eo~AS>a* >dS Lqe DZEa X e
FIG. 4. Actual amounts of constituents (different scales).
If the total amounts of nitrogen and mineral elements present in the entireplant at different stages of maturation are compared with each other, it isrevealed that absorption must have continued at least for a part of the mat-uration period. The differences in physiological maturation between PNKand PN3K plants are well reflected in their total uptake of nlitrogen andmineral elements. Nitrogen absorption was practically completed by March23 in the PN3K series, whereas in the less advanced PNK plants considerableabsorptionl of nitrogen took place until April 17; in the immature remainingpnk plants nitrogen absorption conitinued until May 18. Similarly, ab-sorption of phosphorus continued in the PNK series up to the last samplingdate on May 18, but had already finished in the PN3K series oni April 17.Potassium increased very slightly during maturation in the PNK plants, butno absorption of potassium occurred during the same period in the moreadvanced PN3K plants. In the latter series, there was even a loss of totalamount of potassium during the last part of maturation, amounting to 0.08gm. K2O per ten plants. Though this small loss caninot be regarded as sig-nificant, sinee the variations of the total dry matter between pots of the sameharvest would account for an error of ± 0.045, it should be remembered again
479
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
that substantial losses of potash from all portions of the plant during thelate stages of maturation have been recorded by RICHARDSON, TRUMBLE andSHAPTER (13) in Phalaris tuberosa. Similar losses of nitrogen and mineralelements by remigration into the soil during maturation are also reported byWILFARTH, ROMER and WIMMER (19), SEKERA (15) and PENSTON (11).
The figures of soluble ash are only of limited physiological value, sincesome mineral elements (such as phosphorus and sulphur) are partly lost inthe ignition and are therefore not quantitatively included. It would appear,however, from the total amounts of soluble ash that absorption continuedright through the period of maturation, again more pronounced in the PNKseries than in the PN3K series. In both series, the bulk of mineral elementsabsorbed during this period accumulated only in the roots, not appreciablyaffecting percentages and amounts of soluble ash in the shoots.
It should be remembered that in the interpretation of the data presenteda number of actual and possible errors must be taken into account. Thestandard error of the dry matter (as given in table II) would naturally appearproportionally as standard error of each single constituent of the dry matter,limiting the conclusions as to the significance of the differences recorded.Possible variations in the chemical composition of plants of the same treat-ment and harvest but from different pots are not even included here, sinceonly combined samples were analyzed. In addition, some inherent errors inmethods would probably influence the results at least slightly: small amountsof plant matter were lost at the end of maturation by the loss of seeds andrain may have leached out constituents from dead leaves.
Furthermore, as far as carbohydrates are concerned, one would not expectthat the constituents disappearing from the shoots should be recovered inexactly the same amounts in the roots. Sugars translocated from shoots toroots may there be polymerized to various types of polysaccharides, part ofwhich (such as pentosans, hemi-celluloses and celluloses) do not appear as"hydrolyzable carbohydrates" under the conditions of hydrolysis employed.On the other hand, photosynthetic activity followed by immediate transloca-tion of the photosynthetic products to the roots, would result in an increaseof the amounts of carbohydrates in the roots without a corresponding de-crease in the shoots. Appreciable losses of carbohydrates may possibly occurin the respiration of living plant parts, particularly also of actively growingroots, or in the decomposition of dead parts.
While for all these reasons too far-reaching conclusions should be avoided,the results should be conclusive proof that in Chloris gayana during theperiod of maturation appreciable amounts of organic reserves as well as ofmineral elements are translocated from shoots to roots, in part accompaniedby continued contemporaneous absorption by the roots of nitrogen and min-eral nutrients from the soil.
480
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.
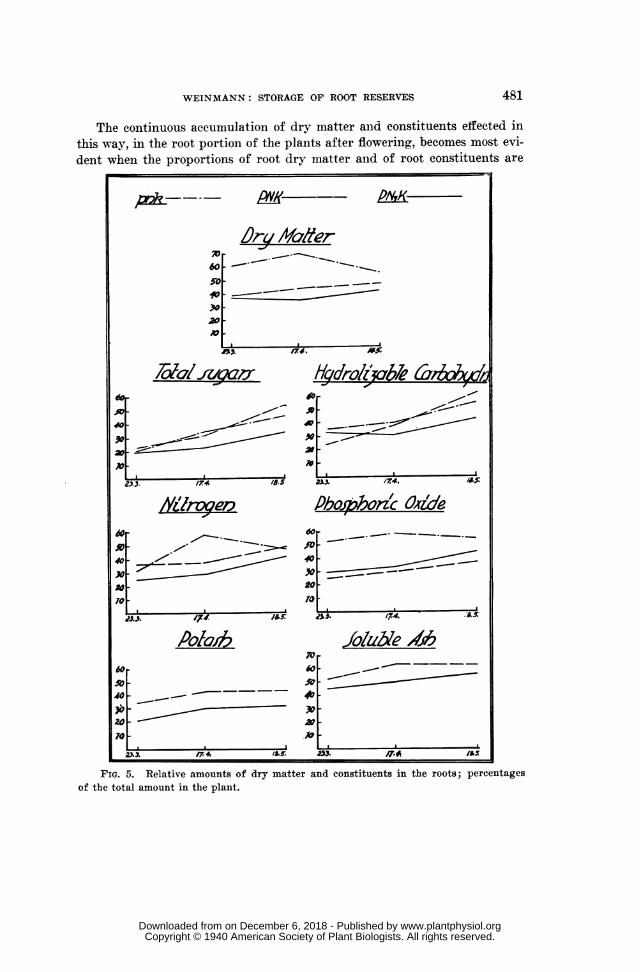
WEINMANN: STORAGE OF ROOT RESERVES
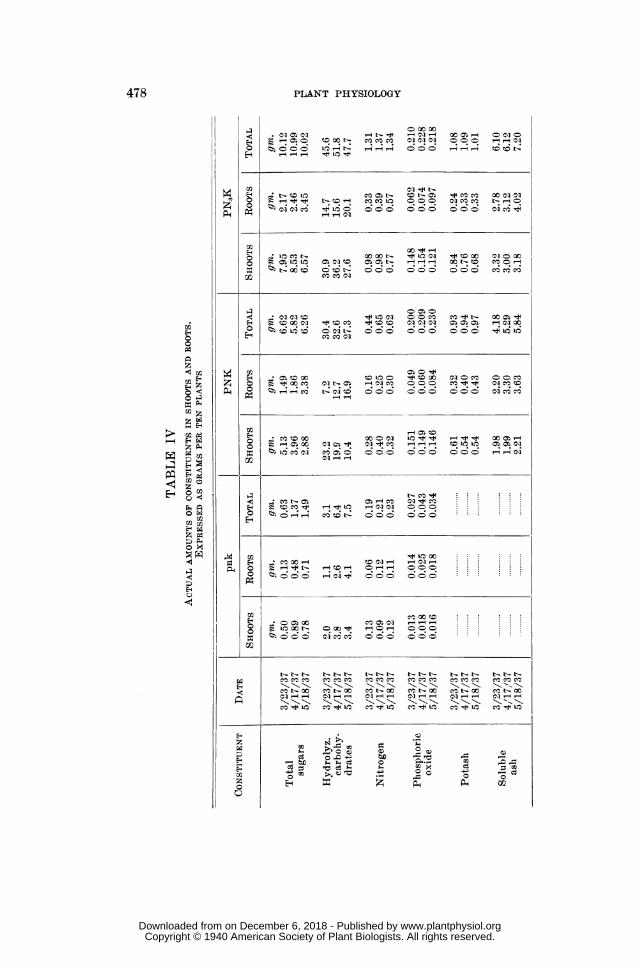
The continuous accumulation of dry matter anid constituents effected inthis way, in the root portion of the plants after flowering, becomes most evi-dent when the proportions of root dry matter and of root constituents are
Y --
D9ray /alter_-
60
'03020
b&iodro/zabk&3469
40
60
480
Ad78
60r
40
20
40
IV
D-3. /174 /8.5
/~-----,
:w0 ===-M.1. /)Y
0Polob
3L. 174.
A boornzc OLrzde60iv
40
'0
70
60
.0
aL!. .
%,. 17.4.A-/
jolZie ih=_-
/Al
FIG. 5. Relative amounts of dry matter and constituents in the roots; percentagesof the total amount in the plant.
481
f
.-Il
.--- .
--l-
1-
Ar.
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
expressed as percentages of the corresponding amounts present in the enitire( amount in roots x 100\
plant amount in wole plat) As can be seen from table V and figure 5,amount in whole plant/TABLE V
RELATIVE AMOUNTS OF DRY MATTER AND CONSTITUENTS IN THE ROOTS.EXPRESSED AS PERCENTAGES OF THE TOTAL AMOUNT IN THE PLANT
CONSTITUENT DATE pnk PNK PN3K
3/23/37 61.8 39.6 37.1Dry matter 4/17/37 64.8 42.8 35.3
5/18/37 56.6 47.0 41.5
3/23/37 20.7 22.5 19.8Total sugars 4/17/37 35.0 32.0 22.4
5/18/37 47.7 54.0 34.4
3/23/37 35.5 23.7 32.2Hydrolyz. 4/17/37 40.7 38.9 30.1carbohydrates 5/18/37 54.7 61.9 42.2
3/23/37 31.6 36.4 25.2Nitrogen 4/17/37 57.2 38.5 28.5
5/18/37 47.8 48.4 42.5
Phosphoric 3/123/37 51.9 24.5 29.5oxide ~4/17/37 58.2 28.7 32.5oxide 5/18/37 52.9 36.5 44.5
3/23/37 34.4 22.2Potash 4/17/37 42.6 30.3
5/18/37 44.3 32.7
3/23/37 52.6 45.6Soluble ash 4/17/37 62.4 51.0
5/18/37 .I.62.2 55.8
these relative amounts of dry matter and constituents in the roots increaseddistinctly during maturation, the only exceptions occurring in the pnkseries, probably owing to the retarded development of these deficiency plants.
Summary
Chloris gayana KUNTH, Rhodes Grass, was grown under controlled con-ditions in pots receiving different fertilizer treatments and was harvested atthree different stages of maturity.
In the well fertilized plants significant increases of the root system oc-curred after the time of flowering and during the formation of seeds, whengrowth of the aerial parts had already ceased.
During this period considerable amounts of sugars, hydrolyzable carbo-hydrates, nitrogen, phosphorus and potassium were lost from the shoots andwere largely recovered in the roots.
482
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.

WEINMANN: STORAGE OF ROOT RESERVES
In addition to translocation from the shoots, nitrogen and mineral ele-ments also accumulated in the roots to a certain extent by simultaneous ab-sorption from the soil. The duration of the absorption period varied, how-ever, for different elements; it also varied for plants grown under differentnutritional conditions and hence reaching different stages of physiologicalmaturation.
The relative amounts of dry matter and constituents present in the roots(expressed as percentages of the total amounts present in the whole plant)increased, in most cases, considerably during autumn, indicating a con-tinuous storage of these substances in the roots with the approach of maturity.
Abnormally developed and immature remaining deficiency plants showedin part a correspondingly abnormal behavior with regard to root storage andabsorption.
The writer is greatly indebted to PROF. J. PHILLIPS, Head of the Depart-ment of Botany of the University of the Witwatersrand, for his interest andthe facilities provided in his Department; to PROF. H. STEPHEN, Head ofthe Department of Chemistry, for placing a laboratory and apparatus at hisdisposal; to DR. B. SEGAL of the same Department for frequent advice andto MR. A. H. BUNTING for his assistance in the analytical work. Sincerethanks are also accorded to the National Research Council and Board ofSouth Africa for having assisted the writer with a grant in connection withthis research.
UNIVERSITY OF THE WITWATERSRANDJOHANNESBURG, SOUTH AFRICA
LITERATURE CITED1. ALDOUS, A. E. Relation of organic food reserves to the growth of soilie
Kansas pasture plants. Jour. Amer. Soc. Agron. 22: 385-392.1930.
2. ALTONA, R. E. A report of an investigation of the seasonal movemelntsof root reserves in six highveld grasses. Unpublished B.Se.Hons.Thesis, Univ. of the Witwatersrand. 1939.
3. BERTRAND, G. Le dosaoe des sucres reducteurs. Bull. Soc. Chimi., ser.3, 35: 1285-1299. 1906.
4. BEWS, J. W., and BAYER, A. W. Researches on the vegetation of Natal-Series III, Section I. On the reaction of Digitaria eriantha var.stolonifera (Stapf) and Themeda triandra (Forsk) to the winterseason. South African Jour. Sci. 28: 158-168. 1931.
5. , and VANDERPLANK J. E. Storaoe anid other carbohy-drates in a Natal succulent and a Natal geophyte and their behaviorbefore, during, and after the winter resting season. Ann. Bot. 44:689-719. 1930.
483
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.

PLANT PHYSIOLOGY
6. Bouyoucos, G. J. A comparison of the hydrometer method and thepipette method for making mechanical analysis of soils, with newdirections. Jour. Amer. Soc. Agron. 22: 747-751. 1930.
7. GRABER, L. F., NELSON, N. T., LEUKEL, W. A., and ALBERT, W. B.Organic food reserves in relation to the growth of alfalfa and otherperennial herbaceous plants. Wisconsin Agr. Exp. Sta. Res. Bull.80. 1927.
8. GRANDFIELD, C. 0. The relation of organic food reserves to the effect ofcutting pasture weeds at different stages of growth. Jour. Amer.Soc. Agron. 22: 709-718. 1930.
9. MCCARTY, E. C. Seasonal march of carbohydrates in Elyonurus am-biguus and Muehlenbergia gracilis andc their reaction under mod-erate grazing use. Plant Physiol. 10: 727-738. 1935.
10. MUNSON, L. S., and WALKER, P. H. The unification of reducing sugarmethods. Jour. Amer. Chem. Soc. 28: 663-686. 1906.
11. PENSTON, N. L. Return of mineral elements to the soil by plants.Nature 136: 268-269. 1935.
12. RICHARDSON, A. E. V., TRUMBLE, H. C., and SHAPTER, R. F. Factorsaffecting mineral content of pastures. Council Sci. & Ind. Res.,Australia, Bull. 49. 1931.
13. , , and . The influence ofgrowth stage and frequency of cutting on the yield and compositionof a perennial grass-Phalaris tuberosa. Council Sci. & Ind. Res..Australia, Bull. 66. 1932.
14. SAMPSON, A. W., and MCCARTY, E. C. The carbohydrate metabolism ofStipa pulchra. Hilgardia 5: 61-100. 1930.
15. SEKERA. Der Mineralstoffwechsel der Gerste. Zeitschr. Pflanzener-nahr., Diingung & Bodenk. B, 7: 535. 1928.
16. STURKIE, D. G. The influence of various top-cutting treatments on rootstocks of Johnson grass (Sorghum halepense). Jour. Amer. Soc.Agron. 22: 82-93. 1930.
17. WEINMANN, H. Investigations on the storage of root reserves in someSouth African grasses. Unpublished M.Sc. Thesis, Univ. of theWitwatersrand. 1938.
18. WIESSMANN, H. Agrikulturchemisches Praktikum. Paul Parey, Berlin.1926.
19. WILFARTH, R., ROMER, H., and WIMMER, G. Vber die Niihrstoffauf-nahme der Pflanzen in verschiedenen Zeiten ihres Wachstums.Landw. Vers.-Sta. 63: 1-70. 1906.
20. WILLARD, C. J. Root reserves of alfalfa with special reference to timeof cutting and yield. Jour. Amer. Soc. Agron. 22: 595-602. 1930.
484
www.plantphysiol.orgon December 6, 2018 - Published by Downloaded from Copyright © 1940 American Society of Plant Biologists. All rights reserved.