stem cells & cancer three tumor biology puzzles: 1.most tumors are of a clonal origin but tumor...
TRANSCRIPT

Stem Cells & Cancer
Three tumor biology puzzles:1. Most tumors are of a clonal origin but tumor cells are heterogeneous.2. It is very difficult to establish stable tumor cell lines from tumors.3. Large numbers of established tumor cells have to be injected to re-initiate
an orthotopic tumor in mice.
Recent key reviews:1. Reya T et al. Stem cells, cancer, and cancer stem cells. Nature 414, 105-111, 2001.2. Visvader JE, and Lindeman GJ. Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nat Rev Cancer 8: 755-768, 2008.3. Dick JE. Stem cell concepts renew cancer research. Blood 112: 4793-4807, 2008.4. Shackleton M, Quintana E, Fearon ER, and Morrison SJ. (2009). Heterogeneity in cancer: cancer stem cells versus clonal evolution. Cell 138, 822-829, 2009.
(Dean Tang, Basic Concepts of Tumor Biology, Sept. 22, 2010)

1. Characteristics & Definition
2. Types & (Adult) SC Identification
3. SC Niche & Plasticity
4. SCs & Cancer
5. Cancer Stem Cells (CSCs)
Stem Cells & Cancer

Flatworm(planarian)
Newt MRL mice

Stem cell development
Terminal differentiation
Death (PCD)
Senescence
Stem cells
Progenitors/Precursor cells

-Rare -Generally small- Normally localized in a ‘protected’ environment called NICHE, where they only occasionally divide. - But they possess HIGH PROLIFERATIVE POTENTIAL and can give rise to large clones of progeny in vitro or in vivo following injury or appropriate stimulation.- Possess the ability to SELF-RENEW (i.e., asymmetric or symmetric cell division)- Can generate (i.e., DIFFERENTIATE into) one or multiple or all cell types (uni-, oligo-, multi-, pluri-, or toti-potent).
Stem Cells

SC
Committed cells
SC Self-renewal, Proliferation, and Differentiation

SC Development: Self-renewal, proliferation, differentiation
LT-SC ST-SC Lateprogenitors
Differen-tiated cells
Differen-tiating cells
Earlyprogenitors
?
Pro
life
rati
onDifferentiation
Transformationtargets
Self-renewal
Niche Commitment Differentiation

•Mouse ESCs were generated early 1980s by Evans and Martin. •mES cells are cultured on mouse fibroblast feeders (irradiated or mitomycin C-treated) together with LIF..mES cells are widely used in gene targeting.•Human ES (hES) cells were first created by Jim Thomson (Uni. Wisconsin) in 1998. •hES cells were initially cultured also on mouse fibroblast feeders but without LIF. Now they can be maintained in defined medium with high bFGF (100 ng/ml), activin, and some other factors.
Embryonic Stem Cells (ESCs)

• Lef
tove
r or
dea
d-e
nd
IV
F e
mb
ryos
(P
GD
) How can hES cells be derived?
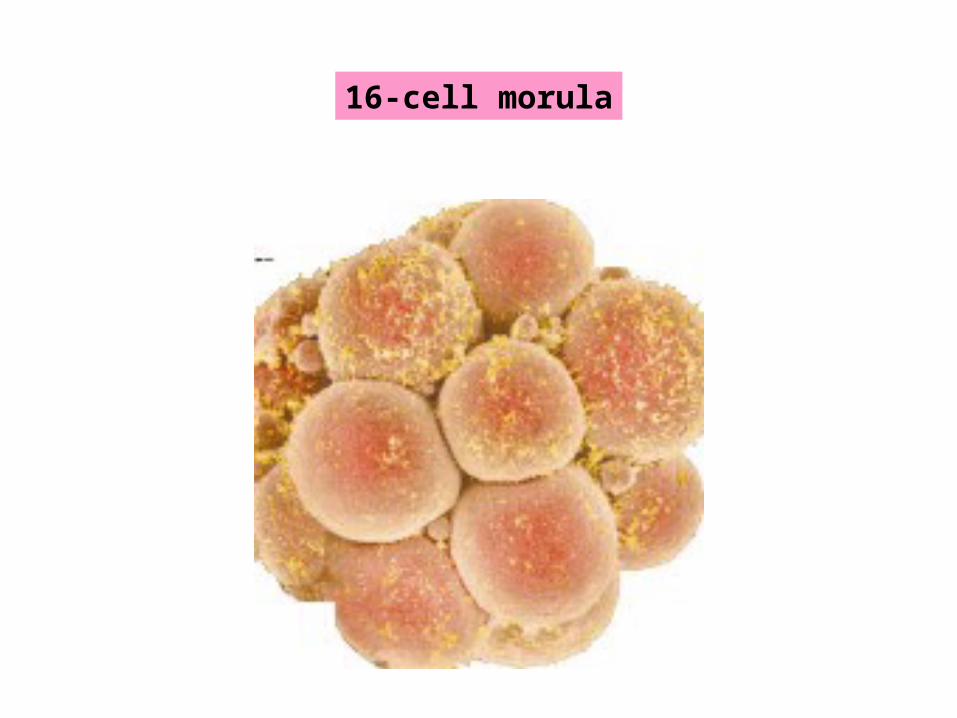
16-cell morula

Primitive ectoderm Trophectoderm
Primitive Endoderm
A. Nagy
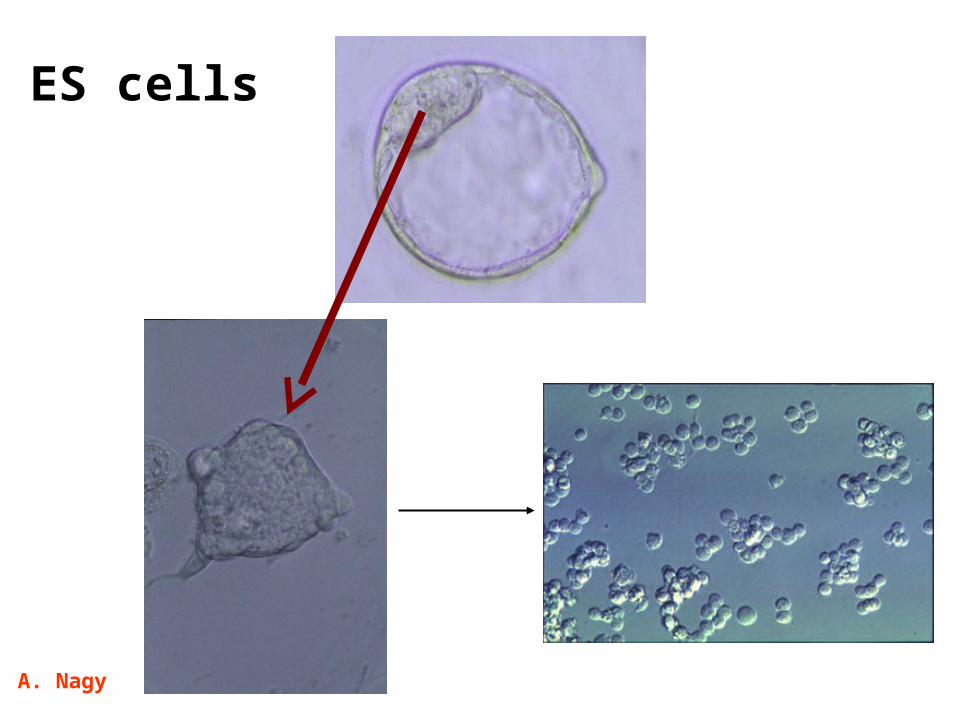
ES cells
A. Nagy

TS cells A. Nagy

A. Nagy

A. Nagy
A. Nagy

heart pancreas testis
liver brain kidney
A. Nagy


•Derived from umbilical cord •Primarily blood stem cells•Also contain mesenchymal stem cells that can differentiate into bone, cartilage, heart muscle, brain, liver tissue etc.*CB-SC could be stimulated to differentiate into neuron, endothelial cell, and insulin-producing cells
Cord Blood Stem Cells (CB-SC)
Germline Stem Cells (GSC)
Other ‘embryonic’ SCs

How to identify and characterize (adult) stem cells?
1. Marker analysis2. Clonal/clonogenic assays3. Side population (SP): BCRP or ABCG24. Label-retaining cells (LRC)5. Aldefluor assay (Aldh1 expression)6. Cell size-based enrichment7. Genetic marking

Passegué, Emmanuelle et al. (2003) Proc. Natl. Acad. Sci. USA 100, 11842-11849
Hematopoietic and progenitor cell lineages
(~1:5,000 or 0.02%;lifetime self-renewal)
(~1:1,000 or 0.1%;self-renewal for 8 wks)
(No self-renewal)

(Nestin)
(GFAP)
(Pax6)(A2B5)
(NG2)(MBP)
(NeuM)
(Mash-1)
(PDGFR)

Sue Fischer

How to identify and characterize (adult) stem cells?
1. Marker analysis2. Clonal/clonogenic assays3. Side population (SP): BCRP or ABCG24. Label-retaining cells (LRC)5. Aldefluor assay (Aldh1 expression)6. Cell size-based enrichment7. Genetic marking

CLONAL vs CLONOGENIC ASSAYS
Clonal
*Plate cells at clonal density(50-100 cells/wellin 6-well plateor 10-cm dishor T25 flask)
*Plate single cellsinto 96-well plates(or using flow sorting)- limiting dilution
Holoclone Mero- or paraclone
a. Cloning efficiency (CE; %)b. Clonal size (cell number/clone)c. Clonal development (tracking)d. Clone types
A clone: a two-dimensional structure
Plating efficiency
Prolif. potential
Clonogenic
‘In-gel’ assays(plate cells at low density)
‘On-gel’ assays(plate at low density)
a. Efficiency (%)b. Colony/sphere size (cell number)c. Colony/sphere development (tracking)d. Immunostaining/tumor exp.
A colony/sphere: a 3-D structure
Colonies(colony-formationassays)
Anchorage-independ.survival
Prolif.
Spheres(sphere-formation assays)
Gels: Agar Agarose Methylcellulose Matrigel Poly-HEMA fibroblasts

Mixing Experiments to Demonstrate the Clonality of Clones/Spheres
DU145 RFP:DU145 GFP (1:1) MC
DU145:DU145 GFP (1:1) MC
phas
eG
FP
DU145:DU145 GFP (1:1) Clonal Assay

How to identify and characterize (adult) stem cells?
1. Marker analysis2. Clonal/clonogenic assays3. Side population (SP): BCRP or ABCG24. Label-retaining cells (LRC)5. Aldefluor assay (Aldh1 expression)6. Cell size-based enrichment7. Genetic marking

Identification of HSC by SP
Zhou et al., Nature Med 7, 1028, 2001

How to identify and characterize (adult) stem cells?
1. Marker analysis2. Clonal/clonogenic assays3. Side population (SP): BCRP or ABCG24. Label-retaining cells (LRC)5. Aldefluor assay (Aldh1 expression)6. Cell size-based enrichment7. Genetic marking

LRCs in the Bulge ARE Stem Cells
Tumbar et al., Science 303, 359-363, 2004; Fuchs et al., Cell 116, 769, 2004Fuchs E: The tortoise and the hair: Slow-cycling cells in the stem cell race.
Cell 137, 811-819, 2009.

How to identify and characterize (adult) stem cells?
1. Marker analysis2. Clonal/clonogenic assays3. Side population (SP): BCRP or ABCG24. Label-retaining cells (LRC)5. Aldefluor assay (Aldh1 expression)6. Cell size-based enrichment7. Genetic marking

Stem Cell Niche
The most important function of a stem cell niche is to keep the stem cells quiescent and from differentiating and simultaneously maintain their “stemness” (i.e., the repertoire of gene expression profiles characteristic of stem cells).

Stem Cell Niche in Hair Follicles: The Bulge
Moore KA & Lemischka IR. Science 311, 1880-1885, 2006
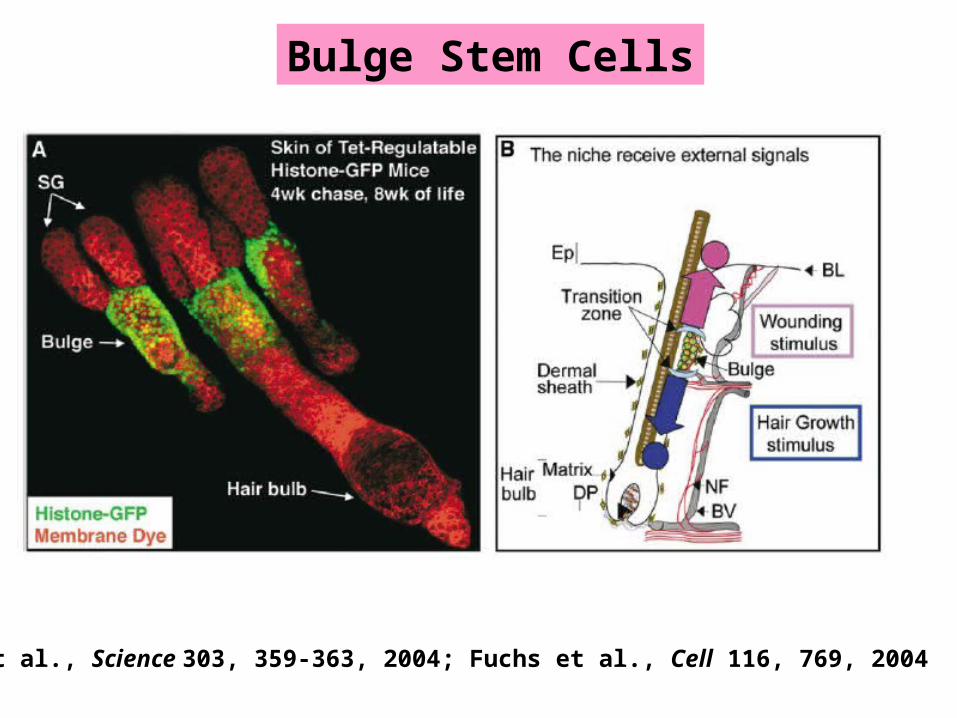
Bulge Stem Cells
Tumbar et al., Science 303, 359-363, 2004; Fuchs et al., Cell 116, 769, 2004

Stem Cell Niche in Small Intestine: The Crypt
Moore KA & Lemischka IR. Science 311, 1880-1885, 2006

Stem Cell Niches in BM
Moore KA & Lemischka IR. Science 311, 1880-1885, 2006
Naveiras O et al., Bone-marrow adipocytes as negative regulators of the hematopoietic microenvironment. Nature 460, 259, 2009.
Mendez-Ferrer, S et al., Mesenchymal and hematopoietic stem cells form a unique bone marrow niche. Nature 466, 829-834, 2010.
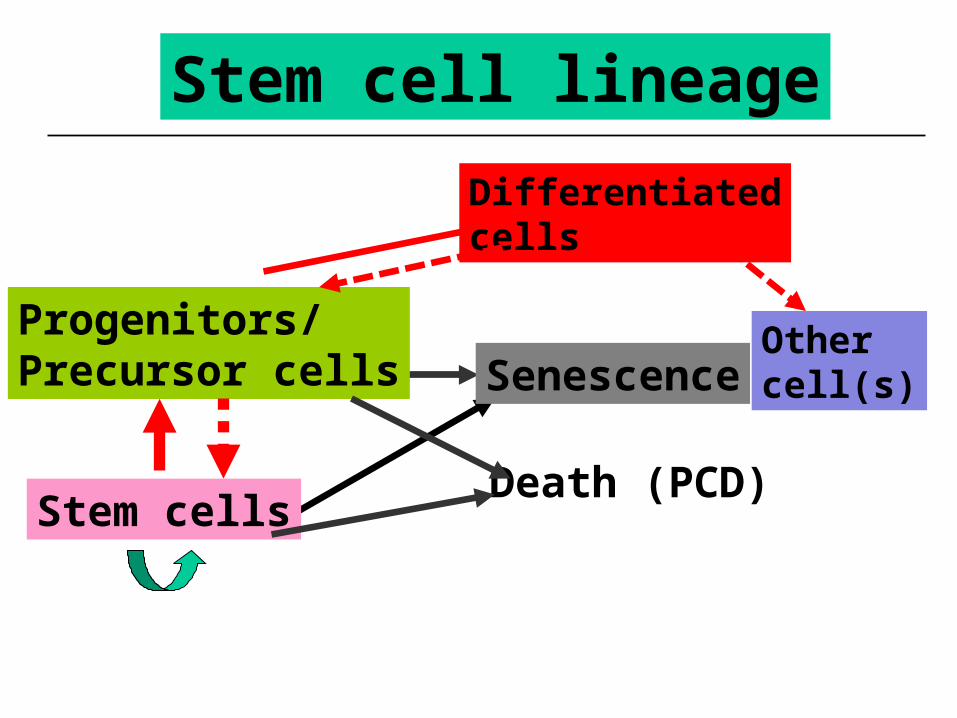
Stem cell lineage
Differentiatedcells
Death (PCD)
Senescence
Stem cells
Progenitors/Precursor cells
Other cell(s)

Adult Stem Cell Plasticity
Plasticity: the ability of SCs to regenerate and trans-differentiate into (many) other cell types (the cell type-specific programming of apparently committed primary progenitors is not irrevocably fixed, but may be radically re-specified in response to a single
transcriptional regulator. Heyworth C et al., EMBO J. 21, 3770-3781, 2002).
**Transdifferentiation vs Dedifferenitation: Transdifferentiation refers to adult stem cells directly differentiating into other cell lineages of cells; de-differentiation refers to somatic stem/progenitor cells first reverting back to a more primitive state then differentiating into a specific cell type.

Blelloch R. Nature 455, 604-605, 2008

*First report: Long-term cultured adult brain (stem) cells can reconstitute the whole blood in lethally irradiated mice (Bjornson et al., Science 283, 534-537, 1999).
*Cells from skeletal muscle have hematopoietic potential (Jackson et al., PNAS 96, 14482-14486, 1999) and can also “differentiate” into many other cell types (Qu-Petersen, Z, et al., JCB 157, 851-864, 2002).
*CNS “SCs” can “differentiate” into muscle cells (Clarke et al., Science 288, 1660-1663, 2000; Galli et al., Nat. Neurosci 3, 986-991, 2000; Tsai and McKay, J. Neurosci 20, 3725-3735, 2000).
*Vice versa, “SCs” from blood or bone marrow can “transdifferentiate” into muscle (Ferrari et al., Science 279, 1528-1530, 1998; Gussoni et al., Nature 401, 390-394, 1999), hepatocytes (Petersen et al., Science 284, 1168-1170, 1999; Lagasse et al., Nat Med 6, 1229-1234, 2000), cardiac myocytes (Orlic et al., Nature 410, 701-705, 2001), or neural cells (Mezey et al., Science 290, 1779-1782, 2000; Brazelton et al., Science 290, 1775-1779, 2000).
*Bone marrow appears to contain two distinct SCs: the HSC and MSC. A single HSC could contribute to epithelia of multiple organs of endodermal and ectodermal origin (Krause et al., Cell 105 369-377, 2001). MSC, on the other hand, can adopt a wider range of fates (endothelial, liver, neural cells, and perhaps all cell types) (Pittenger et al., Science 284, 143-146, 1999;
Schwartz et al., JCI 109, 1291-1302, 2002; Jiang et al., Nature 418, 41-49, 2002).*Pluripotent “SCs” have also been isolated from skin that can “differentiate” into neural cells, epithelial
cells, and blood cells (Toma et al., Nat Cell Biol. 3, 778-784, 2001)*Highly purified adult rat hepatic oval “stem’ cells, which are capable of differentiating into hepatocytes
and bile duct epithelium, can “trans-differentiate” into pancreatic endocrine hormone-producing cells when cultured in a high glucose environment (Yang et al., PNAS 99, 8078-8083, 2002)
“Transdifferentiation” of Stem Cells: Exciting!!!

Cell fusion as one mechanism of trans-differentiation: Not only stem/progenitor cells but also terminally differentiated cells can fuse with other cells
Cardiomyocytes fuse with surrounding noncardiomyocytes and reenter the cell cycle(Matsuura K et al., JCB 167, 351, 2004)

De-differentiation: Cell-cycle re-entry*Many ‘post-mitotic’ cells such as hepatocytes, endothelial cells, and Schwann cells have long
been known to retain proliferative (progenitor) potential.*Dedifferentiation is a genetically regulated process that may ensure a return path to the undifferentiated state when necessary (Katoh et al., PNAS 101, 7005, 2004).*Regeneration of male GSC by spermatogonial dedifferentiation in vivo (Brawley and Matunis,
Science 304, 1331, 2004).*Conversion of mature B cells into T cells by dedifferentiation to uncommitted progenitors
(Nature 449, 473-477, 2007).*During Salamander limb regeneration, complete de-differentiation to a pluripotent state is not required – Progenitor cells in the blastema keep a memory of their tissue origin (Nature 460, 60-65, 2009).*Epigenetic reversion of post-implantation epiblast to pluripotent embryonic cells (Nature,
461, 1292-1295, 2009).*Evidence for cardiomyocyte renewal in humans (Bergmann O et al., Science 324, 98-102, 2009). (Cardiomyocytes turn over at an estimated rate of ~1% per year at age 20, declining to 0.4% per year at age 75. At age 50, 55% of human cardiomyocytes remain from birth while 45% were generated afterward. Over the first decade of life, cardiomyocytes often undergo a final round of DNA synthesis and nuclear division without cell division, resulting in ~25% of human cardiomyocytes being binucleated.)*Neuregulin 1/ErbB4 signaling induces cardiomyocyte proliferation and repair of heart injury
(Bersell et al., Cell, 138, 257-270, 2009).*MafB/c-Maf deficiency enables self-renewal of differentiated functional macrophages (Aziz A,
et al., Science 326, 867-871, 2009).

Pancreatic -cells: Interesting insulin-producing cells
*Insulin-producing -cells in adult mouse pancreas can self-duplicate during normal homeostasis as well as during injury (Dor et al., Nature 429, 41, 2004).
*In vivo reprogramming of adult pancreatic exocrine cells to cells using 3 TFs (Ngn3, Pdx1, and Mafa), suggesting aparadign for directing cell reprogramming without reversion to a pluripotent cell state (Zhou et al., Nature455, 627-632, 2008).
*In response to injury, a population of pancreatic progenitorscan generate glucagon-expressing alpha cells that thentransdifferentiate (with ectopic expression of Pax4) intobeta cells (Collmbat et al, Cell 138, 449-462, 2009).
*Conversion of adult pancreatic -cells to -cells after extreme -cell loss (Nature 464, 1149-1154, 2010).

NU
CL
EA
R R
EP
RO
GR
AM
MIN
G
CE
LL
126
, 652
-655
, 200
6

Induced Pluripotent Cells (iPS cells)(Infection of somatic cells with 2-4 factors: Sox2, Oct4, Klf-4, Myc)
Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006 Aug 25;126(4):663-76. Maherali N, ……., Hochedlinger K. Directly reprogrammed fibroblasts show global epigenetic remodeling and widespread tissue contribution. Cell Stem Cell. 2007 Jun 7;1(1):55-70.Okita K, Ichisaka T, Yamanaka S. Generation of germline-competent induced pluripotent stem cells. Nature. 2007 Jul 19;448(7151):313-7. Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from
adult human fibroblasts by defined factors. Cell 131, 861-872, 2007.Yu J, ……, Thomson JA. Induced pluripotent stem cell lines derived from human somatic cells. Science. 2007 Dec
21;318(5858):1917-20. Hanna J, …. Jaenisch R. Treatment of sickle cell anemia mouse model with iPS cells generated from autologous skin (Science,
318, 1920-1923, 2007).Nakagawa M, …… Yamanaka S. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat Biotechnol. 2008 Jan;26(1):101-6.Park IH, …….. Daley GQ. Reprogramming of human somatic cells to pluripotency with defined factors. Nature. 2008 Jan 10;451(7175):141-6.
Kobayashi T et al., Generation of rat pancreas in mouse by interspecific blastocyst injection of pluripotent stem cells. Cell 142,
787-799, 2010.
*Up to now: >2000 PubMed publications on iPS cells!*Yamanaka S: Elite and stochastic models for induced pluripotent stem cell generation. Nature 460: 49-52, 2009.*Yamanaka S & Blau HM. Nuclear reprogramming to a pluripotent state by three
approaches. Nature 465, 704, 2010

Development and epigenetic (re)programming
Transcription factors and chromatin remodelers and key modulators of ‘potency’
Hochedlinger 2009, Development 136, 509-523

Nicholas CR & Kriegstein AR. Cell reprogramming gets direct. Nature 463, 1031-1032, 2010.Vierbuchen T et al., Direct conversion of fibroblasts to functional neurons by defined factors.
Nature 463, 1035-1041, 2010 (Ascl1, Brn2, and Myt1l). Ieda M, et al., Direct reprogramming of fibroblasts into functional cardiomyocytes by defined
factors. Cell 142, 375-386, 2010 (Gata4, Mef2c, and Tbx5).Bonfanti P, et al., Microenvironmental reprogramming of thymic epithelial cells to skin multi-
potent stem cells. Nature 466, 978-982, 2010.Bussard KM, et al., Reprogramming human cancer cells in the mouse mammary gland. Cancer
Res 70, 6336-6343, 2010.

Perinatal stem/progenitors
functional maturation
PCD
Stem Cells & Tumorigenesis
(Terminal) differentiation
Adult Stem/Progenitor Cells
Death
Senescence
ESCs
Tumor cells

Several fundamental tumor biology questions
1. Why is it so difficult to establish tumor cell lines from established tumors or even metastases?
2. Why tens or hundreds of thousands of establishedtumor cells have to be injected to initiate anorthotopic tumor?
3. Tumors are clonal (i.e., all tumors were initially derived from ‘going bad’ of one cell) but why is thetumor itself heterogeneous?

Cell type Cell# injected Incidence Latency (days)
Du145 1,000 0/4 10,000 0/4 100,000 1/4 103 500,000 3/5 53, 53, 59
LAPC4 100 0/4 1,000 0/4 10,000 0/4 100,000 0/4 500,000 3/6 43, 43, 48
LAPC9 100 0/3 1,000 0/910,000 4/8 46, 53, 75, 75100,000 6/9 32, 42, 42, 45, 62, 691,000,000 4/4 48, 56, 56, 69
Tumorigenecities of Orthotopically Implanted Prostate Cancer Cells

PSA
AR
CD57
CK5
Nanog

Several fundamental tumor biology questions
1. Why is it so difficult to establish tumor cell lines from tumors or even metastases?
2. Why tens or hundreds of thousands of establishedtumor cells have to be injected to initiate anorthotopic tumor?
3. Tumors are clonal (i.e., all tumors were initially derived from ‘going bad’ of one cell) but why is thetumor itself heterogeneous (i.e., comprisingmultiple cell types)?
These questions can be potentially explained by the presence of stem-like cells in the tumor, i.e., tumor (orcancer) stem cells

Cancer stem cells (CSC): Tumorigenic cells
Hewitt, HB. Studies of the quantitative transplantation of mouse sarcoma. Brit. J. Cancer. 7, 367-383, 1953 (~0.01% of the tumor cells are tumor stem cells; limiting dilution method).
Bruce, W.R & van der Gaag, H. A quantitative assay for the number of murine lymphoma cells capable of proliferation in vivo. Nature 199, 79-80, 1963 (~0.8% total cells forming spleen colonies).
Wodinsky, I., Swiniarski, J., and Kensler, CJ. Spleen colony studies of leukemia L1210. I. Growth kinetics of lymphocytic L1210 cells in vivo as determined by spleen colony assay. Cancer Chemother. Rep. 51, 415-421, 1967 (1-3% total cells forming spleen colonies).Bergsahel, DE & Valeriote FA. Growth characteristics of a mouse plasma cell tumor. Cancer Res. 28, 2187-2196, 1968 (<4.4% cells are tumor stem cells). Park CH, Bergsagel DE, and McCulloch, EA. Mouse myeloma tumor stem cells: a primary cell culture assay. JNCI 46, 411-422, 1971 (0.7 - 1.2% clonogenic, tumor stem cells).Hamburger, A.W. and Salmon SE. Primary bioassay of human tumor stem cells. Science 197, 461-463, 1977
0.001 - 0.1% myeloma cells forming colonies in soft agar).Fidler, IJ and Kripke, ML. Metastasis results from preexisting variant cells within a malignant tumor. Science 197, 893-895, 1977.Salmon, SE, Hamburger, A.W., Soehnlen, B., Durie, B.G.M., Alberts, D.S., and Moon, T.E.. Quantitation of
differential sensitivity of human -tumor stem cells to anticancer drugs. N. Eng. J. Med. 298, 1321-1327, 1978.
Sell, S., and Pierce, G.B. Maturation arrest of stem cell cell differentiation is a common pathway for the cellular origin of teratocarcinomas and epithelial cancers. Lab. Invest. 70, 6-22, 1994.Trott, KR. Tumor stem cells: the biological concept and its application in cancer treatment. Radiother. Oncol. 30, 1-5, 1994.
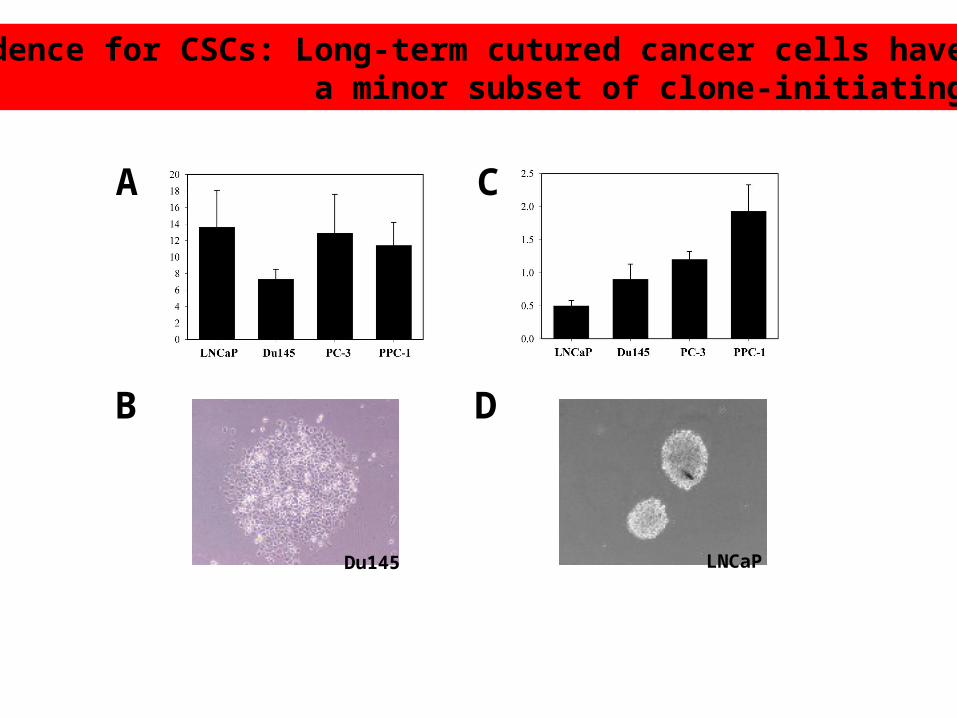
Evidence for CSCs: Long-term cutured cancer cells have only a minor subset of clone-initiating cells
A C
B D
Du145 LNCaP

Only 0.01 -0.1% of the actutely purified tumor cells have the sphere-forming abilities

Leukemic stem cells (LSC): Classic examples of CSC
Lapidot T, et al. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 367, 645-648, 1994.
Blair et al., Blood 89, 3104-3112, 1997Bonnet, D., & Dick, J.E. Nature Med. 3, 730-737, 1997
1. Most of the leukemic cells are unable to proliferate extensively and only a small, defined subset of cells was consistently clonogenic.
2. LSCs for human AML were identified prospectively and purified as [Thy1-, CD34+, CD38-] cells from various patient samples and they represent 0.2 - 1% of the total.
3. The LSCs are the only cells capable of transferring AML from human patient to NOD/SCID mice and are referred to as SCID leukemia-initiating cells (SL-IC).

CSC studies in human solid tumors (2003 - 2008)CSC studies in human solid tumors (2003 - 2008)Tumor type Samples Marker Mice Transplantation Results Ref.Breast cancer 9 (1 primary; 8 met.) CD44+CD24-/loESA+ NOD/SCID mice mammary fat pad >50 fold enrichment 1
FACS pretreated with VP16 in tumorigenicityBreast cancer 4 xenotransplants ALDH+ NOD/SCID mice humanized mammary 500 ALDH+ cells generate T; 2
(from 2 primary; 2 met.) FACS fat pad 20 ALDH+CD44+CD24-Lin- cells generate T
Brain tumors 7 primary tumors CD133+ (MACS) 6-8 wk NOD/SCID intracranial injection CD133+ more tumorigenic 3
Prostate cancer 7 (4 primary, CD44+21hiCD133+ (MACS) no tumor experiments marker+ cells more clonogenic 4 1 benign, 2 LN mets) purified from long-term cultured cells
Colon cancer 17 (6 primary, 10 liver CD133+ (double MACS) 8 wk NOD/SCID renal capsule 1 CSC/57,000 T. cells 5 & 1 retroperitoneal met.) irradiated 1 CSC/262 CD133+ cells
Colon cancer 19 primary (5 Dukes A) CD133+ (FACS or MACS) SCID subcutaneous 3,000 CD133+ cells generate T 6 Colon cancer 21 primary CRC CD133+ 5-6 wk nude mice subcutaneous 2,500 CD133+ cells generate T 7
25 CD133+-derived spheres generate T
Colon cancer 2 primary, 6 xenografts EpCAMCD166+CD44+ 6-8 wk NOD/SCID subcutaneous 150 EpCAMCD166+CD44+ 8(FACS) cells generate T
Pancreatic cancer 10 (2 primary; 2 met.) CD44+CD24+ESA+ NOD/SCID subcutaneous+pancreas >100 fold enrichment 9Pancreatic cancer 11 (6 met.); sorting for 7 CD133+ (MACS) 8-12 wk nude mice pancreas 500 CD133+ cells generate T 10
L3.6pl metastatic line CD133+CXCR4+ (FACS) the CD133+CXCR4+ pop. mediates met. Head & Neck 25 primary (3 recurrences) CD44+Lin- (FACS) NOD/SCID & Rag2-/- subcutaneous 5,000 CD44+Lin- cells generate T 11
9 for sorting (4 primary+5 xenografts) only 13/25 HNSCC samples gave tumorsMelanoma 7 (1 primary; 4 LN & ABCB5+ (MACS) NOD/SCID subcutaneous 1 MMIC/1 million bulk T cells 12
2 visceral met.) 1ary xeno: 1 MMIC/160,000 ABCB5+ cells 2ary xeno: 1 MMIC/120,000 ABCB5+ cells
Lung cancer 19 (18 primary; 1 met.) CD133+ (FACS) 4 wk SCID or nude subcutaneous 104 CD133+ cells generate T. 13
Liver cancer 28 primary (only 13 used) CD45-CD90+ (MACS) SCID intrahepatic CD45-CD90+ more tumorigenic 14
1. Al-Hajj M, et al. PNAS 2003;100:3983-8. 2. Ginestier C et al., Cell Stem Cell 2007;1:555-67.3. Singh SK, et al. Nature 2004;432:396-401. 4. Collins AT, et al. Cancer Res 2005; 65: 10946-51.5. O'Brien CA, et al. Nature 2007; 445:106-10. 6. Ricci-Vitiani L, et al. Nature 2007; 445:111-5.7. Todaro M et al., Cell Stem Cell 2007; 1: 389-402. 8. Dalerba P et al., PNAS 2007; 104: 10158-63.9. Li C et al., Cancer Res. 2007; 67:1030-7 10. Hermann PC et al., Cell Stem Cell 2007;1:313-3211. Prince ME et al., PNAS 2007; 104: 973-8. 12. Schatton T, et al., Nature 2008; 451:345-9.13. Eramo A et al., Cell Death Differ. 2008; 15: 504-14. 14. Yang ZF et al., Cancer Cell 2008; 13: 153-66.

Identification of CSC in Solid Tumors by Markers
Al-Hajj, M et al., Prospective identification of tumorigenic breast cancer cells. PNAS 100, 3983-3988, 2003.
Hemmati, H.D et al., Cancerous stem cells can arise from pediatric brain tumors. PNAS 100, 15178-15183, 2003 (using neural SC markers).
Galli R et al., Isolation and characterization of tumorigenic, stem-likeneural precursors from human glioma. Cancer Res. 64, 7011, 2004.
Singh SK et al. Identification of human brain tumour initiating cells.
Nature 432: 396-401, 2004 (using CD133 as a marker).Patrawala L, et al. Highly purified CD44+ prostate cancer cells from
xenograft human tumors are enriched in tumorigenic and metastatic progenitor cells. Oncogene 25, 1696-1708, 2006.

Identification of CSCs by SP
Kondo T, et al. Persistence of a small population of cancer stem-like cells in the C6 rat glioma cell line. PNAS 101: 781-786, 2004.Hirschmann-Jax C, et al. A distinct "side population" of cells with high
drug efflux capacity in human tumor cells. PNAS 101: 14228, 2004.Patrawala, L., et al. Side population (SP) is enriched in tumorigenic,
stem-like cancer cells whereas ABCG2+ and ABCG2- cancer cells are similarly tumorigenic. Cancer Res. 65, 6207, 2005.
Haraguchi N, et al. Characterization of a side population of cancer cells from human gastrointestinal system. Stem Cells 24, 506-13, 2006.
Chiba T, et al. Side population purified from hepatocellular carcinoma cells harbors cancer stem cell-like properties. Hepatology 44:240-51, 2006.
Szotek PP, et al., Ovarian cancer side population defines cells with stem cell-like characteristics and Mullerian Inhibiting Substance responsiveness. PNAS 103:11154-9, 2006.

The SP in LAPC-9 tumor is more tumorigenic
LAPC9-SP 100 2/8 (52 and 108 d)1,000 3/4 (108 d)1,500 3/3 (60 d)
LAPC9-NSP 1,500 0/8150,000 0/7 (terminated in 9 mo.)300,000 1/6 (92 d)
e

Identification of putative CSCs by quiescence
Pece S, et al., Biological and molecular heterogeneity of breast cancer correlates with their cancerStem cell content. Cell 140, 62-73, 2010.

Identification of putative CSCs by size selection
Small(<10 µm)
Large(>30 µm)
small
large
100 1000

-1187Exon 1
TSS (+1)
ATG (164)K5 promoter
116
718Exon 2
Identification of putative CSCs by promoter tracking

Where did tumor cells come from?
LT-SC ST-SC Lateprogenitors
Differen-tiated cells
Differen-tiating cells
Earlyprogenitors
?
Pro
life
rati
onDifferentiation
Transformationtargets
Self-renewal
Niche Commitment Differentiation

Progenitor or differentiated cells as transformation targets or CSCs
Passegue, E., et al. Normal and leukemic hematopoiesis: are leukemias a stem cell disorder or a reacquisition of stem cell characteristics? Proc. Natl. Acad. Sci. USA. 100 (Suppl 1): 11842-11849, 2003.
Huntly, B.J., et al. MOZ-TIF2, but not BCR-ABL, confers properties of leukemic stem cells to committed murine hematopoietic progenitors. Cancer Cell. 6: 587-596, 2004.
Jamieson, C.H., et al. Granulocyte-macrophage progenitors as candidate leukemic stem cells in blast-crisis CML. NEJM. 351: 657-667, 2004.
Krivtsov, AV et al., Transformation from committed progenitor to leukemia stem cells initiated by MLL-AF-9. Nature 442, 818-822, 2006.
McCormack MP, et al. The Lmo2 oncogene initiates leukemia in mice byinducing thymocyte self-renewal. Science 327, 879-883, 2010.

Where do tumors REALLY come from?Houghton J et al., Gastric cancer originating from bone
marrow-derived cells. Science. 2004, 306: 1568-71. Direkze NC, et al., Bone marrow contribution to tumor-
associated myofibroblasts and fibroblasts. Cancer Res. 2004, 64: 8492-5.
Aractingi S et al., Skin carcinoma arising from donor cells in a kidney transplant recipient. Cancer Res. 2005, 65:1755-60.
Kaplan RN et al., VEGFR1-positive hematopoietic bone marrowprogenitors initiate the pre-metastatic niche. Nature. 2005, 438, 820-827.
Riggi N et al., Development of Ewing’s sarcoma from primary bone marrow-derived mesenchymal progenitor cells. Cancer Res. 2005, 65:11459-11468.
Palapattu GS et al., Epithelial architectural destruction is necessary for bone marrow derived cell contribution to regenerating prostate epithelium. J. Urol, 2006, 176:813-18.

Cancer Stem Cells & Treatment
Weissman, Nature 414, 105-111, 2001

How to specifically target CSCs?
*Identify functional CSC molecules (e.g., CD44, c-KIT, Bmi-1, Nanog) Knock downthese molecules to inhibit CSC properties (e.g., Jeter et al., 2009; Levina Vet al., Elimination of human lung CSCs through targeting of the stem cellfactor-c-kit autocrine signaling loop. Cancer Res. 70, 338-346, 2010).
*Identify CSC-specific cell-surface markers Develop antibody or prodrug basedtherapeutics
*Take advantage of differential signaling requirement between normal and cancer SC(Yilmaz OH et al., Pten dependence distinguishes haematopoietic stem cells
from leukaemia-initiating cells. Nature. 2006 441:475-82)

Using stem/progenitor cells to treat tumors:
*Using neural progenitor cells alone: these cells produce large amounts of TGF(Staflin et al., Cancer Res. 64, 5347-5354, 2004)
*Using neural progenitor cells to deliver cytokines or cytotoxic genes or products--- Benedetti et al., Nat. Med. 6, 447-450, 2000--- Aboody et al., PNAS 97, 12846-12851, 2000--- Herrlinger et al., Mol. Ther. 1, 347-357, 2000--- Ehtesham et al., Cancer Res. 62, 5657-5663, 2002--- Ehtesham et al., Cancer Res. 62, 7170-7174, 2002--- Barresi et al., Cancer Gene Ther. 10, 396-402, 2003
*Using IL23-expressing BM-derived neural stem-like cells to attack glioma cells--- Yuan X et al., Cancer Res. 66, 2630-2638, 2006
*Using hMSC to attack Kaposi’s sarcoma--- Khakoo AY et al., JEM. 203, 1235-1247, 2006

‘Reprogramming’ the microenvironment of CSC to treat tumors:
Kulesa P et al. Reprogramming metastatic melanoma cells to assume a neural crest cell-like phenotype in an embryonic microenvironment. PNAS103, 3752-3757, 2006.
Topczewska JM., et al., Embryonic and tumorigenic pathways converge via Nodalsignaling: role in melanoma aggressiveness. Nature Med. 12, 925-932,2006.

PSA
AR
CD57
CK5
Nanog
Rx

CSC

PSA
AR
CD57
CK5
Nanog

PSA
AR
CD57
CK5
Nanog
Rx
Rx
