spytag/spycatcher functionalization of e2 nanocages with...
TRANSCRIPT

SpyTag/SpyCatcher Functionalization of E2 Nanocages with Stimuli-Responsive Z‑ELP Affinity Domains for Tunable MonoclonalAntibody Binding and Precipitation PropertiesAndrew R. Swartz and Wilfred Chen*
Department of Chemical and Biomolecular Engineering, University of Delaware, Colburn Laboratory, 150 Academy Street, Newark,Delaware 19716, United States
*S Supporting Information
ABSTRACT: E2 nanocages functionalized with Z-domain-elastin-like polypeptide affinity ligands (Z-ELP40) usingSortase A (SrtA) ligation have been shown to be a promisingscaffold for purifying monoclonal antibodies (mAbs) based onaffinity precipitation. However, the reversible nature of SrtAreaction has been attributed to the low ligation efficiency(<25%) and has significantly limited the practical utility of thetechnology. Here, we reported an improved conjugationplatform using the SpyTag/SpyCatcher pair to form aspontaneous isopeptide bond between SpyTag-E2 and Z-ELP-SpyCatcher fusion proteins of two different ELP chain-lengths. Using this system, E2 ligation efficiencies exceeding90% were obtained with both 40- and 80-repeat Z-ELP-SpyCatcher fusions. This enabled the production of nanocages fully functionalized with Z-ELP for improved aggregation andmAb binding. Compared to the 50% decorated Z-ELP40-E2 nanocages produced by SrtA ligation, the fully decorated Z-ELP80-Spy-E2 nanocages exhibited a 10 °C lower transition temperature and a 2-fold higher mAb binding capacity. The improvedtransition property of the longer Z-ELP80 backbone allowed for >90% recovery of Z-ELP80-Spy-E2 nanocages at roomtemperature using 0.1 M ammonium sulfate after mAb elution. The flexibility of customizing different affinity domains onto theSpyTag-E2 scaffold should expand our ability to purify other non-mAb target proteins based on affinity precipitation.
■ INTRODUCTION
Affinity precipitation is a bioseparation method that involvesthe capture and precipitation of a target protein in bulksolution1 and has been proposed as a more cost-effective andscalable alternative to conventional Protein A affinitychromatography for the purification of therapeutic monoclonalantibodies (mAbs).2,3 The two main strategies for affinityprecipitation utilize either multivalent affinity ligand cross-linking (primary effect) or stimuli-responsive affinity ligands(secondary effect) to trigger the formation of larger aggregatesfor precipitation.4 Primary effect affinity ligands such asmultivalent haptens have been used to cross-link with dimericantibodies into oligomeric aggregates;5,6 however, this strategyis limited by low precipitation yields, slow dissolution kinetics,and the requirement of an additional separation step afterelution to remove the dissociated affinity ligands.7 mAbpurification using stimuli-responsive affinity ligands has alsobeen implemented using either pH8 or temperature9
responsive synthetic polymers and biopolymers such aselastin-like polypeptides (ELP).10,11 However, utility of thetechnique is hindered by the harsh environmental stimuli suchas low pH, high temperature, and/or high salt concentrationnecessary to induce phase transition of the affinity ligand,
resulting in an increase in mAb aggregation12,13 or thecoprecipitation of other cell culture impurities.14
Our group has developed a new protein-based affinityscaffold that combines the benefits of both primary andsecondary effect affinity precipitation. We functionalized thesurface of a self-assembled 60-mer E2 nanocage with an affinityligand consisting of a Protein A derived Z-domain-40-repeatELP (Z-ELP40) fusion and demonstrated binding-inducedmultivalent cross-linking as the initial capture mechanism ofmAbs from cell culture, followed by a selective precipitation ofregenerated nanocages after mAb elution for easy separation.15
The 25 nm E2 nanocage from Bacillus stearothermophilus wasselected as an ideal scaffold due to its high structural stability(Tm > 80 °C) and the ability to genetically modify the exteriorN-terminus for protein conjugation.16 Because large geneticfusions have been shown to disrupt proper E2 self-assembly,we modified a truncated E2 subunit with a small N-terminaltriglycine tag to allow for post-translational, site-specificconjugation of Z-ELP40 using Staphylococcus aureus Sortase A(SrtA) ligation.17,18 Using partially functionalized Z-ELP-E2
Received: June 26, 2018Revised: August 7, 2018Published: August 10, 2018
Article
pubs.acs.org/bcCite This: Bioconjugate Chem. 2018, 29, 3113−3120
© 2018 American Chemical Society 3113 DOI: 10.1021/acs.bioconjchem.8b00458Bioconjugate Chem. 2018, 29, 3113−3120
Dow
nloa
ded
via
UN
IV O
F D
EL
AW
AR
E o
n Se
ptem
ber
19, 2
018
at 1
1:11
:24
(UT
C).
Se
e ht
tps:
//pub
s.ac
s.or
g/sh
arin
ggui
delin
es f
or o
ptio
ns o
n ho
w to
legi
timat
ely
shar
e pu
blis
hed
artic
les.
nanocages, an affinity precipitation process capable of highyield and efficient impurity clearance equivalent to Protein Achromatography was developed for the purification ofindustrial mAbs from cell culture harvests.19,20
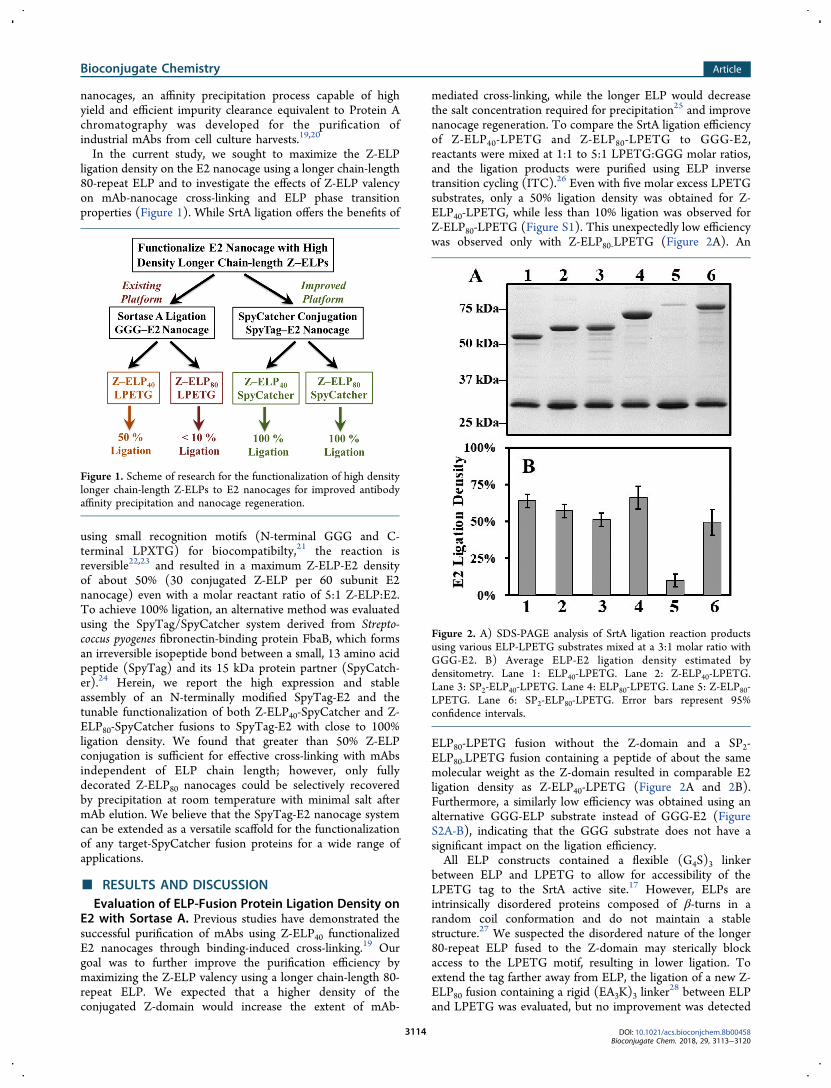
In the current study, we sought to maximize the Z-ELPligation density on the E2 nanocage using a longer chain-length80-repeat ELP and to investigate the effects of Z-ELP valencyon mAb-nanocage cross-linking and ELP phase transitionproperties (Figure 1). While SrtA ligation offers the benefits of
using small recognition motifs (N-terminal GGG and C-terminal LPXTG) for biocompatibilty,21 the reaction isreversible22,23 and resulted in a maximum Z-ELP-E2 densityof about 50% (30 conjugated Z-ELP per 60 subunit E2nanocage) even with a molar reactant ratio of 5:1 Z-ELP:E2.To achieve 100% ligation, an alternative method was evaluatedusing the SpyTag/SpyCatcher system derived from Strepto-coccus pyogenes fibronectin-binding protein FbaB, which formsan irreversible isopeptide bond between a small, 13 amino acidpeptide (SpyTag) and its 15 kDa protein partner (SpyCatch-er).24 Herein, we report the high expression and stableassembly of an N-terminally modified SpyTag-E2 and thetunable functionalization of both Z-ELP40-SpyCatcher and Z-ELP80-SpyCatcher fusions to SpyTag-E2 with close to 100%ligation density. We found that greater than 50% Z-ELPconjugation is sufficient for effective cross-linking with mAbsindependent of ELP chain length; however, only fullydecorated Z-ELP80 nanocages could be selectively recoveredby precipitation at room temperature with minimal salt aftermAb elution. We believe that the SpyTag-E2 nanocage systemcan be extended as a versatile scaffold for the functionalizationof any target-SpyCatcher fusion proteins for a wide range ofapplications.
■ RESULTS AND DISCUSSIONEvaluation of ELP-Fusion Protein Ligation Density on
E2 with Sortase A. Previous studies have demonstrated thesuccessful purification of mAbs using Z-ELP40 functionalizedE2 nanocages through binding-induced cross-linking.19 Ourgoal was to further improve the purification efficiency bymaximizing the Z-ELP valency using a longer chain-length 80-repeat ELP. We expected that a higher density of theconjugated Z-domain would increase the extent of mAb-
mediated cross-linking, while the longer ELP would decreasethe salt concentration required for precipitation25 and improvenanocage regeneration. To compare the SrtA ligation efficiencyof Z-ELP40-LPETG and Z-ELP80-LPETG to GGG-E2,reactants were mixed at 1:1 to 5:1 LPETG:GGG molar ratios,and the ligation products were purified using ELP inversetransition cycling (ITC).26 Even with five molar excess LPETGsubstrates, only a 50% ligation density was obtained for Z-ELP40-LPETG, while less than 10% ligation was observed forZ-ELP80-LPETG (Figure S1). This unexpectedly low efficiencywas observed only with Z-ELP80‑LPETG (Figure 2A). An
ELP80-LPETG fusion without the Z-domain and a SP2-ELP80‑LPETG fusion containing a peptide of about the samemolecular weight as the Z-domain resulted in comparable E2ligation density as Z-ELP40-LPETG (Figure 2A and 2B).Furthermore, a similarly low efficiency was obtained using analternative GGG-ELP substrate instead of GGG-E2 (FigureS2A-B), indicating that the GGG substrate does not have asignificant impact on the ligation efficiency.All ELP constructs contained a flexible (G4S)3 linker
between ELP and LPETG to allow for accessibility of theLPETG tag to the SrtA active site.17 However, ELPs areintrinsically disordered proteins composed of β-turns in arandom coil conformation and do not maintain a stablestructure.27 We suspected the disordered nature of the longer80-repeat ELP fused to the Z-domain may sterically blockaccess to the LPETG motif, resulting in lower ligation. Toextend the tag farther away from ELP, the ligation of a new Z-ELP80 fusion containing a rigid (EA3K)3 linker
28 between ELPand LPETG was evaluated, but no improvement was detected
Figure 1. Scheme of research for the functionalization of high densitylonger chain-length Z-ELPs to E2 nanocages for improved antibodyaffinity precipitation and nanocage regeneration.
Figure 2. A) SDS-PAGE analysis of SrtA ligation reaction productsusing various ELP-LPETG substrates mixed at a 3:1 molar ratio withGGG-E2. B) Average ELP-E2 ligation density estimated bydensitometry. Lane 1: ELP40-LPETG. Lane 2: Z-ELP40-LPETG.Lane 3: SP2-ELP40-LPETG. Lane 4: ELP80-LPETG. Lane 5: Z-ELP80-LPETG. Lane 6: SP2-ELP80-LPETG. Error bars represent 95%confidence intervals.
Bioconjugate Chemistry Article
DOI: 10.1021/acs.bioconjchem.8b00458Bioconjugate Chem. 2018, 29, 3113−3120
3114

(Figure S3). Because of the reversibility of SrtA ligation andthe limited ligation for Z-ELP80-LPETG,
29,30 we decided topursue an alternative conjugation strategy utilizing the SpyTag-SpyCatcher pair.24
Expression and Conjugation of Spytag-E2 and Z-ELP-SpyCatcher Constructs. To overcome the limitations of theSrtA reaction, SpyTag/SpyCatcher conjugation was selected asan alternative because of its irreversible and rapid isopeptidebond formation.31 Although we were successful in displayingthe GGG tag on the N-terminus of the E2 monomer, there arevery few reports on the genetic fusion of other peptides ontoE2.32 To identify the best insertion site, the 13-amino acidSpyTag was attached to different locations on the native N-terminal linker of the E2 monomer and expressed in E. coli.Various levels of soluble expression were obtained for allSpyTag-E2 fusion constructs (Figure S4A). The soluble E. colilysate was heated to 70 °C for the partial purification of the E2nanocages and to check for thermostability (Figure S4B). AllSpyTag-E2 constructs were at least partially soluble after 70 °Cincubation, and dynamic light scattering confirmed assemblednanoparticles of the expected size (Figure S5A). Z-ELP40-SpyCatcher and Z-ELP80-SpyCatcher were expressed andpurified using ITC (Figure S6). SpyTag-E2(158) andSpyTag-E2(173) were selected to test for conjugation withthe Z-ELP-SpyCatcher fusions because they exhibited thehighest culture productivity and low polydispersity (FiguresS4C and S5B). Samples were mixed for 1 h in PBS followed byone ITC cycle to remove any unreacted E2. The reactionresulted in high conjugation of both Z-ELP40- and Z-ELP80-SpyCatcher fusion on either SpyTag-E2(158) or SpyTag-E2(173) (Figure 3). SpyTag-E2(158) was selected for future
investigation because it contained the same N-terminalextension as the well-characterized GGG-E2(158) from ourprevious studies17 for better comparison.To compare the two conjugation methods, all reactants were
mixed at a 0.75:1 ELP:E2 molar ratio for 4 h followed by oneITC cycle (Figure S7). In contrast to the low conversion ofSrtA ligation, SpyTag/SpyCatcher conjugation resulted inclose to 100% efficiency for both Z-ELP40- and Z-ELP80-SpyCatcher, as no unreacted Z-ELP was detected by SDS-PAGE in the purified products. The simplicity of the 2-component reaction and the high efficiency afforded bySpyTag/SpyCatcher significantly advance the capabilities ofthe nanocage affinity precipitation technology and enableaccess to all 60 E2 subunits for a highly tunable platform.
Effect of Ligation Density on Nanocage Aggregationand IgG Binding Capacity. To examine the versatility of theSpyTag/SpyCatcher system to generate a wide range of Z-ELP-functionalized E2 nanocages, reactions were prepared at0.25:1, 0.5:1, 0.75:1, and 1:1 Z-ELP:E2 molar ratios for the 40-and 80-repeat ELP. For comparison, SrtA ligation reactionswere used to prepare different levels of functionalized E2nanocages using 0.5:1, 1:1, 2:1, and 4:1 Z-ELP40:E2 molarratios (Figure 4A). The E2 ligation density was proportional to
the reactant ratio for the SpyTag/SpyCatcher system, while asignificant excess amount of Z-ELP was required to drive theconversion to 50% for the SrtA ligation (Figure 4B). Theaverage conjugation efficiency for the SpyTag/SpyCatcherproducts exceeded 90% (Figure 4C), indicating that any targetZ-ELP conjugation density may be achieved by simply mixingat the target Z-ELP:E2 reactant ratio. Because the SpyTag/SpyCatcher system eliminates the requirement for SrtA andthe need to remove excess Z-ELP reactants, this system offers asubstantially more streamlined and cost-efficient method togenerate Z-ELP functionalized E2 nanocages.The potential benefits of higher Z-ELP ligation densities on
the nanocage transition temperature (Tt) and IgG bindingcapacity were further investigated. Increasing the ligationdensity from 50% to 100% resulted in a 5 °C lower Tt, andconjugation with the longer 80-repeat ELP further decreasedthe Tt value by 6−7 °C across all samples (Figure 5A). Theseresults again highlight the benefit of the SpyTag/SpyCatchersystem as only the fully decorated Z-ELP80-E2 nanocageprovides a Tt value close to room temperature, a conditionhighly desirable for nanocage recovery using only a smallincrease in ionic strength after mAb elution.To ensure that the longer ELP and the presence of a larger
SpyCatcher fusion have no impact on IgG binding, the bindingcapacity per nanocage was evaluated using an industrial mAb(human IgG1) in the presence of excess IgG. As expected, ahigher Z-ELP ligation density resulted in a higher bindingcapacity, and the ELP chain-length had no influence in mAb
Figure 3. Conjugation of SpyTag-E2(158) (Lane 1) and SpyTag-E2(173) (Lane 2) with Z-ELP40-SpyCatcher (Lane 3) and Z-ELP80-SpyCatcher (Lane 4) using either a 2:1 or 5:1 molar ratio ofSpyCatcher:SpyTag substrates. Conjugation products were analyzedby SDS-PAGE for Z-ELP40-SpyCatcher and SpyTag-E2(158) (Lanes5−6), Z-ELP80-SpyCatcher and SpyTag-E2(158) (Lanes 7−8), Z-ELP40-SpyCatcher and SpyTag-E2(173) (Lanes 9−10), and Z-ELP80-SpyCatcher and SpyTag-E2(173) (Lanes 11−12).
Figure 4. A) SDS-PAGE analysis of the conjugation of Z-ELP40-SpyCatcher (Lanes 1−4) or Z-ELP80-SpyCatcher (Lanes 5−8) ontoSpyTag-E2 and the SrtA-mediated ligation of Z-ELP40-LPETG (Lanes9−12) onto GGG-E2. Different ratios of Z-ELP:E2 were used for theSpyTag-SpyCatcher reactions (0.25:1, 0.5:1, 0.75:1, and 1:1) and theSrtA ligation (0.5:1, 1:1, 2:1, and 4:1). B) The resulting ELP-E2ligation densities (determined by densitometry) as a function of theinitial Z-ELP:E2 reactant ratios for Z-ELP40-Spy-E2 (blue square), Z-ELP80-Spy-E2 (red circle), and Z-ELP40-SrtA-E2 (green triangle). C)Comparison of average ligation efficiencies of the conjugationreaction. Error bars represent 95% confidence intervals.
Bioconjugate Chemistry Article
DOI: 10.1021/acs.bioconjchem.8b00458Bioconjugate Chem. 2018, 29, 3113−3120
3115
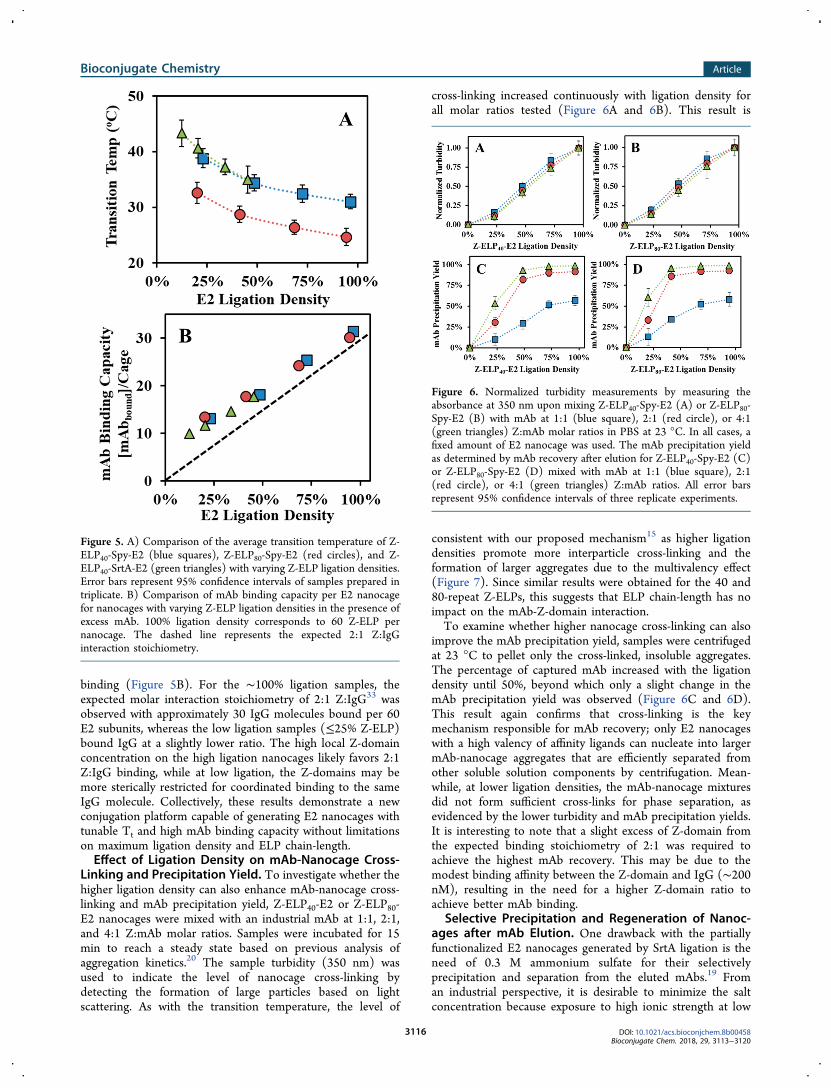
binding (Figure 5B). For the ∼100% ligation samples, theexpected molar interaction stoichiometry of 2:1 Z:IgG33 wasobserved with approximately 30 IgG molecules bound per 60E2 subunits, whereas the low ligation samples (≤25% Z-ELP)bound IgG at a slightly lower ratio. The high local Z-domainconcentration on the high ligation nanocages likely favors 2:1Z:IgG binding, while at low ligation, the Z-domains may bemore sterically restricted for coordinated binding to the sameIgG molecule. Collectively, these results demonstrate a newconjugation platform capable of generating E2 nanocages withtunable Tt and high mAb binding capacity without limitationson maximum ligation density and ELP chain-length.Effect of Ligation Density on mAb-Nanocage Cross-
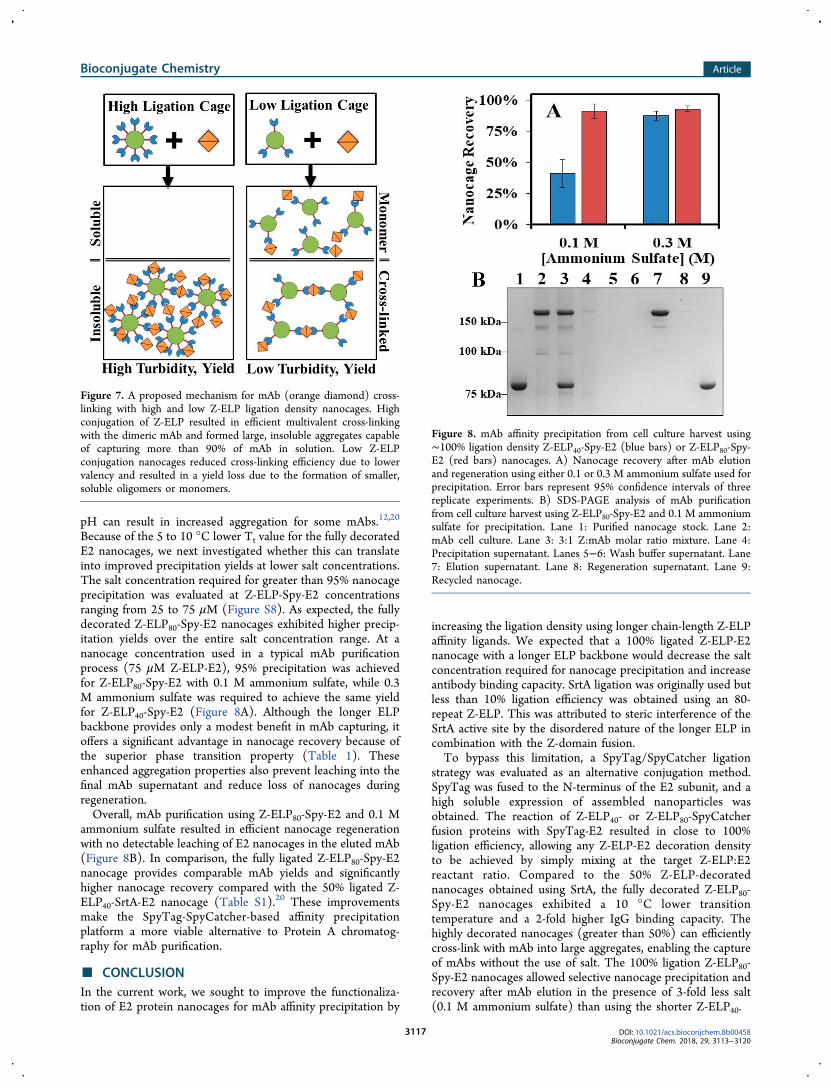
Linking and Precipitation Yield. To investigate whether thehigher ligation density can also enhance mAb-nanocage cross-linking and mAb precipitation yield, Z-ELP40-E2 or Z-ELP80-E2 nanocages were mixed with an industrial mAb at 1:1, 2:1,and 4:1 Z:mAb molar ratios. Samples were incubated for 15min to reach a steady state based on previous analysis ofaggregation kinetics.20 The sample turbidity (350 nm) wasused to indicate the level of nanocage cross-linking bydetecting the formation of large particles based on lightscattering. As with the transition temperature, the level of
cross-linking increased continuously with ligation density forall molar ratios tested (Figure 6A and 6B). This result is
consistent with our proposed mechanism15 as higher ligationdensities promote more interparticle cross-linking and theformation of larger aggregates due to the multivalency effect(Figure 7). Since similar results were obtained for the 40 and80-repeat Z-ELPs, this suggests that ELP chain-length has noimpact on the mAb-Z-domain interaction.To examine whether higher nanocage cross-linking can also
improve the mAb precipitation yield, samples were centrifugedat 23 °C to pellet only the cross-linked, insoluble aggregates.The percentage of captured mAb increased with the ligationdensity until 50%, beyond which only a slight change in themAb precipitation yield was observed (Figure 6C and 6D).This result again confirms that cross-linking is the keymechanism responsible for mAb recovery; only E2 nanocageswith a high valency of affinity ligands can nucleate into largermAb-nanocage aggregates that are efficiently separated fromother soluble solution components by centrifugation. Mean-while, at lower ligation densities, the mAb-nanocage mixturesdid not form sufficient cross-links for phase separation, asevidenced by the lower turbidity and mAb precipitation yields.It is interesting to note that a slight excess of Z-domain fromthe expected binding stoichiometry of 2:1 was required toachieve the highest mAb recovery. This may be due to themodest binding affinity between the Z-domain and IgG (∼200nM), resulting in the need for a higher Z-domain ratio toachieve better mAb binding.
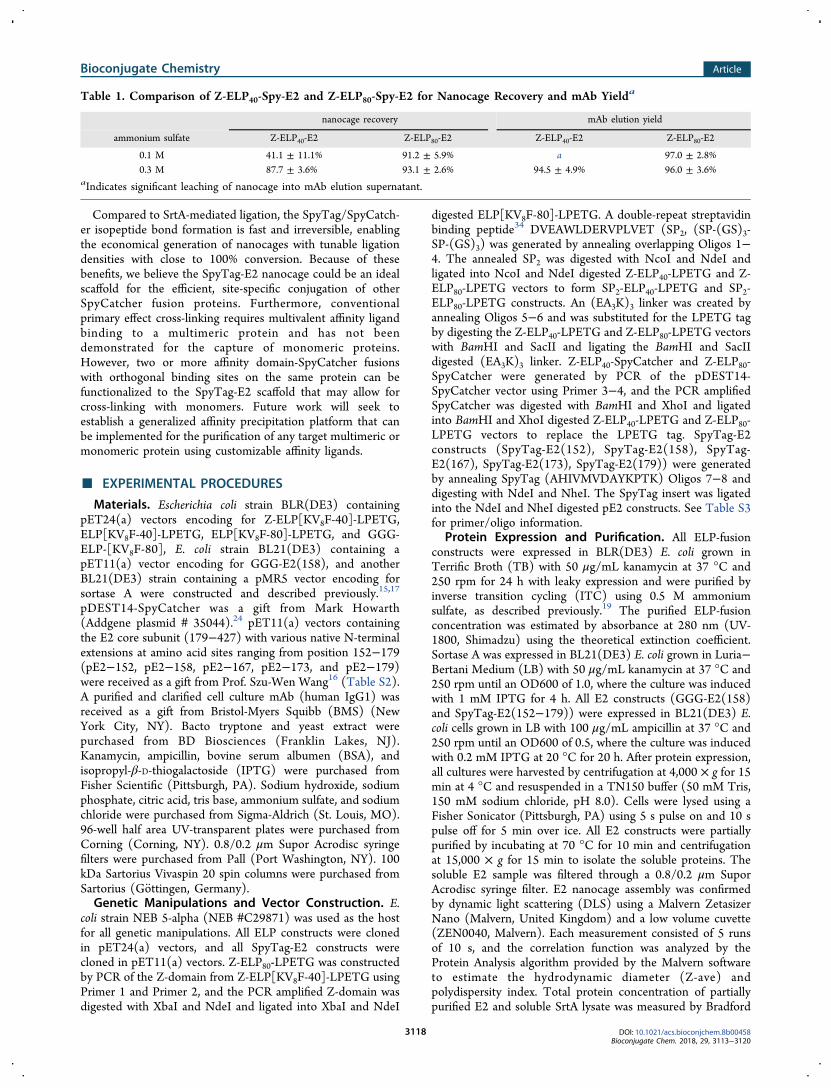
Selective Precipitation and Regeneration of Nanoc-ages after mAb Elution. One drawback with the partiallyfunctionalized E2 nanocages generated by SrtA ligation is theneed of 0.3 M ammonium sulfate for their selectivelyprecipitation and separation from the eluted mAbs.19 Froman industrial perspective, it is desirable to minimize the saltconcentration because exposure to high ionic strength at low
Figure 5. A) Comparison of the average transition temperature of Z-ELP40-Spy-E2 (blue squares), Z-ELP80-Spy-E2 (red circles), and Z-ELP40-SrtA-E2 (green triangles) with varying Z-ELP ligation densities.Error bars represent 95% confidence intervals of samples prepared intriplicate. B) Comparison of mAb binding capacity per E2 nanocagefor nanocages with varying Z-ELP ligation densities in the presence ofexcess mAb. 100% ligation density corresponds to 60 Z-ELP pernanocage. The dashed line represents the expected 2:1 Z:IgGinteraction stoichiometry.
Figure 6. Normalized turbidity measurements by measuring theabsorbance at 350 nm upon mixing Z-ELP40-Spy-E2 (A) or Z-ELP80-Spy-E2 (B) with mAb at 1:1 (blue square), 2:1 (red circle), or 4:1(green triangles) Z:mAb molar ratios in PBS at 23 °C. In all cases, afixed amount of E2 nanocage was used. The mAb precipitation yieldas determined by mAb recovery after elution for Z-ELP40-Spy-E2 (C)or Z-ELP80-Spy-E2 (D) mixed with mAb at 1:1 (blue square), 2:1(red circle), or 4:1 (green triangles) Z:mAb ratios. All error barsrepresent 95% confidence intervals of three replicate experiments.
Bioconjugate Chemistry Article
DOI: 10.1021/acs.bioconjchem.8b00458Bioconjugate Chem. 2018, 29, 3113−3120
3116
pH can result in increased aggregation for some mAbs.12,20
Because of the 5 to 10 °C lower Tt value for the fully decoratedE2 nanocages, we next investigated whether this can translateinto improved precipitation yields at lower salt concentrations.The salt concentration required for greater than 95% nanocageprecipitation was evaluated at Z-ELP-Spy-E2 concentrationsranging from 25 to 75 μM (Figure S8). As expected, the fullydecorated Z-ELP80-Spy-E2 nanocages exhibited higher precip-itation yields over the entire salt concentration range. At ananocage concentration used in a typical mAb purificationprocess (75 μM Z-ELP-E2), 95% precipitation was achievedfor Z-ELP80-Spy-E2 with 0.1 M ammonium sulfate, while 0.3M ammonium sulfate was required to achieve the same yieldfor Z-ELP40-Spy-E2 (Figure 8A). Although the longer ELPbackbone provides only a modest benefit in mAb capturing, itoffers a significant advantage in nanocage recovery because ofthe superior phase transition property (Table 1). Theseenhanced aggregation properties also prevent leaching into thefinal mAb supernatant and reduce loss of nanocages duringregeneration.Overall, mAb purification using Z-ELP80-Spy-E2 and 0.1 M
ammonium sulfate resulted in efficient nanocage regenerationwith no detectable leaching of E2 nanocages in the eluted mAb(Figure 8B). In comparison, the fully ligated Z-ELP80-Spy-E2nanocage provides comparable mAb yields and significantlyhigher nanocage recovery compared with the 50% ligated Z-ELP40-SrtA-E2 nanocage (Table S1).20 These improvementsmake the SpyTag-SpyCatcher-based affinity precipitationplatform a more viable alternative to Protein A chromatog-raphy for mAb purification.
■ CONCLUSIONIn the current work, we sought to improve the functionaliza-tion of E2 protein nanocages for mAb affinity precipitation by
increasing the ligation density using longer chain-length Z-ELPaffinity ligands. We expected that a 100% ligated Z-ELP-E2nanocage with a longer ELP backbone would decrease the saltconcentration required for nanocage precipitation and increaseantibody binding capacity. SrtA ligation was originally used butless than 10% ligation efficiency was obtained using an 80-repeat Z-ELP. This was attributed to steric interference of theSrtA active site by the disordered nature of the longer ELP incombination with the Z-domain fusion.To bypass this limitation, a SpyTag/SpyCatcher ligation
strategy was evaluated as an alternative conjugation method.SpyTag was fused to the N-terminus of the E2 subunit, and ahigh soluble expression of assembled nanoparticles wasobtained. The reaction of Z-ELP40- or Z-ELP80-SpyCatcherfusion proteins with SpyTag-E2 resulted in close to 100%ligation efficiency, allowing any Z-ELP-E2 decoration densityto be achieved by simply mixing at the target Z-ELP:E2reactant ratio. Compared to the 50% Z-ELP-decoratednanocages obtained using SrtA, the fully decorated Z-ELP80-Spy-E2 nanocages exhibited a 10 °C lower transitiontemperature and a 2-fold higher IgG binding capacity. Thehighly decorated nanocages (greater than 50%) can efficientlycross-link with mAb into large aggregates, enabling the captureof mAbs without the use of salt. The 100% ligation Z-ELP80-Spy-E2 nanocages allowed selective nanocage precipitation andrecovery after mAb elution in the presence of 3-fold less salt(0.1 M ammonium sulfate) than using the shorter Z-ELP40.
Figure 7. A proposed mechanism for mAb (orange diamond) cross-linking with high and low Z-ELP ligation density nanocages. Highconjugation of Z-ELP resulted in efficient multivalent cross-linkingwith the dimeric mAb and formed large, insoluble aggregates capableof capturing more than 90% of mAb in solution. Low Z-ELPconjugation nanocages reduced cross-linking efficiency due to lowervalency and resulted in a yield loss due to the formation of smaller,soluble oligomers or monomers.
Figure 8. mAb affinity precipitation from cell culture harvest using∼100% ligation density Z-ELP40-Spy-E2 (blue bars) or Z-ELP80-Spy-E2 (red bars) nanocages. A) Nanocage recovery after mAb elutionand regeneration using either 0.1 or 0.3 M ammonium sulfate used forprecipitation. Error bars represent 95% confidence intervals of threereplicate experiments. B) SDS-PAGE analysis of mAb purificationfrom cell culture harvest using Z-ELP80-Spy-E2 and 0.1 M ammoniumsulfate for precipitation. Lane 1: Purified nanocage stock. Lane 2:mAb cell culture. Lane 3: 3:1 Z:mAb molar ratio mixture. Lane 4:Precipitation supernatant. Lanes 5−6: Wash buffer supernatant. Lane7: Elution supernatant. Lane 8: Regeneration supernatant. Lane 9:Recycled nanocage.
Bioconjugate Chemistry Article
DOI: 10.1021/acs.bioconjchem.8b00458Bioconjugate Chem. 2018, 29, 3113−3120
3117
Compared to SrtA-mediated ligation, the SpyTag/SpyCatch-er isopeptide bond formation is fast and irreversible, enablingthe economical generation of nanocages with tunable ligationdensities with close to 100% conversion. Because of thesebenefits, we believe the SpyTag-E2 nanocage could be an idealscaffold for the efficient, site-specific conjugation of otherSpyCatcher fusion proteins. Furthermore, conventionalprimary effect cross-linking requires multivalent affinity ligandbinding to a multimeric protein and has not beendemonstrated for the capture of monomeric proteins.However, two or more affinity domain-SpyCatcher fusionswith orthogonal binding sites on the same protein can befunctionalized to the SpyTag-E2 scaffold that may allow forcross-linking with monomers. Future work will seek toestablish a generalized affinity precipitation platform that canbe implemented for the purification of any target multimeric ormonomeric protein using customizable affinity ligands.
■ EXPERIMENTAL PROCEDURES
Materials. Escherichia coli strain BLR(DE3) containingpET24(a) vectors encoding for Z-ELP[KV8F-40]-LPETG,ELP[KV8F-40]-LPETG, ELP[KV8F-80]-LPETG, and GGG-ELP-[KV8F-80], E. coli strain BL21(DE3) containing apET11(a) vector encoding for GGG-E2(158), and anotherBL21(DE3) strain containing a pMR5 vector encoding forsortase A were constructed and described previously.15,17
pDEST14-SpyCatcher was a gift from Mark Howarth(Addgene plasmid # 35044).24 pET11(a) vectors containingthe E2 core subunit (179−427) with various native N-terminalextensions at amino acid sites ranging from position 152−179(pE2−152, pE2−158, pE2−167, pE2−173, and pE2−179)were received as a gift from Prof. Szu-Wen Wang16 (Table S2).A purified and clarified cell culture mAb (human IgG1) wasreceived as a gift from Bristol-Myers Squibb (BMS) (NewYork City, NY). Bacto tryptone and yeast extract werepurchased from BD Biosciences (Franklin Lakes, NJ).Kanamycin, ampicillin, bovine serum albumen (BSA), andisopropyl-β-D-thiogalactoside (IPTG) were purchased fromFisher Scientific (Pittsburgh, PA). Sodium hydroxide, sodiumphosphate, citric acid, tris base, ammonium sulfate, and sodiumchloride were purchased from Sigma-Aldrich (St. Louis, MO).96-well half area UV-transparent plates were purchased fromCorning (Corning, NY). 0.8/0.2 μm Supor Acrodisc syringefilters were purchased from Pall (Port Washington, NY). 100kDa Sartorius Vivaspin 20 spin columns were purchased fromSartorius (Gottingen, Germany).Genetic Manipulations and Vector Construction. E.
coli strain NEB 5-alpha (NEB #C29871) was used as the hostfor all genetic manipulations. All ELP constructs were clonedin pET24(a) vectors, and all SpyTag-E2 constructs werecloned in pET11(a) vectors. Z-ELP80-LPETG was constructedby PCR of the Z-domain from Z-ELP[KV8F-40]-LPETG usingPrimer 1 and Primer 2, and the PCR amplified Z-domain wasdigested with XbaI and NdeI and ligated into XbaI and NdeI
digested ELP[KV8F-80]-LPETG. A double-repeat streptavidinbinding peptide34 DVEAWLDERVPLVET (SP2, (SP-(GS)3-SP-(GS)3) was generated by annealing overlapping Oligos 1−4. The annealed SP2 was digested with NcoI and NdeI andligated into NcoI and NdeI digested Z-ELP40-LPETG and Z-ELP80-LPETG vectors to form SP2-ELP40-LPETG and SP2-ELP80-LPETG constructs. An (EA3K)3 linker was created byannealing Oligos 5−6 and was substituted for the LPETG tagby digesting the Z-ELP40-LPETG and Z-ELP80-LPETG vectorswith BamHI and SacII and ligating the BamHI and SacIIdigested (EA3K)3 linker. Z-ELP40-SpyCatcher and Z-ELP80-SpyCatcher were generated by PCR of the pDEST14-SpyCatcher vector using Primer 3−4, and the PCR amplifiedSpyCatcher was digested with BamHI and XhoI and ligatedinto BamHI and XhoI digested Z-ELP40-LPETG and Z-ELP80-LPETG vectors to replace the LPETG tag. SpyTag-E2constructs (SpyTag-E2(152), SpyTag-E2(158), SpyTag-E2(167), SpyTag-E2(173), SpyTag-E2(179)) were generatedby annealing SpyTag (AHIVMVDAYKPTK) Oligos 7−8 anddigesting with NdeI and NheI. The SpyTag insert was ligatedinto the NdeI and NheI digested pE2 constructs. See Table S3for primer/oligo information.
Protein Expression and Purification. All ELP-fusionconstructs were expressed in BLR(DE3) E. coli grown inTerrific Broth (TB) with 50 μg/mL kanamycin at 37 °C and250 rpm for 24 h with leaky expression and were purified byinverse transition cycling (ITC) using 0.5 M ammoniumsulfate, as described previously.19 The purified ELP-fusionconcentration was estimated by absorbance at 280 nm (UV-1800, Shimadzu) using the theoretical extinction coefficient.Sortase A was expressed in BL21(DE3) E. coli grown in Luria−Bertani Medium (LB) with 50 μg/mL kanamycin at 37 °C and250 rpm until an OD600 of 1.0, where the culture was inducedwith 1 mM IPTG for 4 h. All E2 constructs (GGG-E2(158)and SpyTag-E2(152−179)) were expressed in BL21(DE3) E.coli cells grown in LB with 100 μg/mL ampicillin at 37 °C and250 rpm until an OD600 of 0.5, where the culture was inducedwith 0.2 mM IPTG at 20 °C for 20 h. After protein expression,all cultures were harvested by centrifugation at 4,000 × g for 15min at 4 °C and resuspended in a TN150 buffer (50 mM Tris,150 mM sodium chloride, pH 8.0). Cells were lysed using aFisher Sonicator (Pittsburgh, PA) using 5 s pulse on and 10 spulse off for 5 min over ice. All E2 constructs were partiallypurified by incubating at 70 °C for 10 min and centrifugationat 15,000 × g for 15 min to isolate the soluble proteins. Thesoluble E2 sample was filtered through a 0.8/0.2 μm SuporAcrodisc syringe filter. E2 nanocage assembly was confirmedby dynamic light scattering (DLS) using a Malvern ZetasizerNano (Malvern, United Kingdom) and a low volume cuvette(ZEN0040, Malvern). Each measurement consisted of 5 runsof 10 s, and the correlation function was analyzed by theProtein Analysis algorithm provided by the Malvern softwareto estimate the hydrodynamic diameter (Z-ave) andpolydispersity index. Total protein concentration of partiallypurified E2 and soluble SrtA lysate was measured by Bradford
Table 1. Comparison of Z-ELP40-Spy-E2 and Z-ELP80-Spy-E2 for Nanocage Recovery and mAb Yielda
nanocage recovery mAb elution yield
ammonium sulfate Z-ELP40-E2 Z-ELP80-E2 Z-ELP40-E2 Z-ELP80-E2
0.1 M 41.1 ± 11.1% 91.2 ± 5.9% a 97.0 ± 2.8%0.3 M 87.7 ± 3.6% 93.1 ± 2.6% 94.5 ± 4.9% 96.0 ± 3.6%
aIndicates significant leaching of nanocage into mAb elution supernatant.
Bioconjugate Chemistry Article
DOI: 10.1021/acs.bioconjchem.8b00458Bioconjugate Chem. 2018, 29, 3113−3120
3118

protein assay purchased from Bio-Rad (Hercules, CA) usingBSA as a standard. Protein expression was confirmed byCoomassie stained, 10% acrylamide SDS-PAGE using a Bio-Rad Mini-PROTEAN electrophoresis system (Hercules, CA).Protein purity was estimated using densitometry analysis ofSDS-PAGE gels using Thermo MyImage software (Waltham,MA).Sortase A Ligation of ELP-LPETG Fusions to GGG-E2.
Sortase A reactions were prepared with 70 °C partially purified10 μM GGG substrate, 10 μM SrtA in soluble lysate, and avariable concentration of ITC purified LPETG substrate (1:1to 5:1 LPETG:GGG molar ratio) in a reaction buffer (TN150,6 mM CaCl2) and mixed for 4 h at 37 °C. The ligation productwas purified by ITC using 0.5 M ammonium sulfate andresuspended in phosphate buffered saline (PBS, 25 mMsodium phosphate, 150 mM sodium chloride, pH 7.2). Theunreacted ELP-LPETG fusion protein was removed by 100kDa diafiltration into PBS using Sartorius Vivaspin 20 spincolumns. The purified ligation products were confirmed byCoomassie stained 10% acrylamide SDS-PAGE, and the ELPfusion-E2 ligation density was estimated by densitometryanalysis.Z-ELP-SpyCatcher Conjugation to SpyTag-E2. Purified
Z-ELP40-SpyCatcher or Z-ELP80-SpyCatcher was mixed witheither 10 μM or 25 μM E2 SpyTag-E2 at target molar ratios inPBS for 1−4 h at room temperature followed by one ITC cycleinto PBS. The conjugation products were confirmed byCoomassie stained 10% acrylamide SDS-PAGE, and the Z-ELP-E2 ligation density was estimated by densitometryanalysis. 100% ligation density assumes 60 Z-ELP conjugatedper 60-subunit E2 nanocage. Ligation efficiency was calculatedby (Z-ELP-E2 ligation density)/([Z-ELP]:[E2] reactant ratio).Transition Temperature Measurement. Transition
temperature (Tt) measurements were made using a ShimadzhuUV-1800 spectrophotometer. 100 μL samples were added intriplicate to an 8-well multicell microcuvette (Shimadzhu) andequilibrated for 10 min at 20 °C, and the absorbance at 350nm was measured every 0.5 degrees from 20 to 60 °C at aramp rate of 0.5 degree per min with a 30 s equilibration beforeeach measurement. The absorbance curves were analyzed bythe provided Tm Analysis Software (Shimadzhu), and the Tt,defined as the maximum slope of the transition profile, wasevaluated.Absorbance Measurement. Absorbance measurements
were performed using a Synergy plate reader from BioTek(Winooski, VT) run at 23 °C. 100 μL of experimental samplesand buffer controls were well mixed and added to a half-area96-well UV transparent plate. For all measurements, theabsorbance signal was buffer subtracted and corrected for pathlength (1 cm). For mAb or nanocage concentration, theabsorbance was measured at 280 nm, and the theoreticalextinction coefficient was used to calculate concentration. Forturbidity evaluation, the absorbance was measured at 350 nm.Z-ELP-E2 Nanocage mAb Binding Capacity. To
evaluate the nanocage binding capacity for an industrialmAb, nanocages with varying Z-ELP ligation density wereprepared at 25 μM E2 concentration in microcentrifuge tubesand mixed with excess mAb at room temperature for 2 h inPBS at 23 °C. Samples were then adjusted to 0.5 Mammonium sulfate, mixed for 10 min at 23 °C, and centrifugedat 15,000 × g for 10 min to pellet the precipitated nanocagesand bound mAb. The supernatant was removed, and theabsorbance was measured to evaluate unbound mAb
concentration. The bound mAb concentration was calculatedby mass balance. The mAb binding capacity per 60-subunit E2nanocage was estimated by [mAb]bound/(60*[E2]).
mAb-Nanocage Cross-Linking and PrecipitationYield. An industrial mAb was prepared in microcentrifugetubes with the Z-ELP40-Spy-E2 or Z-ELP80-Spy-E2 nanocagesat 1:1, 2:1, 4:1 Z:mAb molar ratios in triplicate. The sampleswere mixed for 30 min at 23 °C, and the turbidity wasmeasured using absorbance at 350 nm. The samples were thencentrifuged at 15,000 × g for 10 min at 23 °C to pellet theinsoluble components, and the supernatant was removed. Thepellet was resuspended in 1x volume elution buffer (50 mMsodium citrate, pH 3.5) and mixed for 15 min at 23 °C. Thesamples were adjusted to 0.5 M ammonium sulfate andcentrifuged at 15,000 × g for 10 min at 23 °C to selectivelyprecipitate the dissociated nanocage. The elution supernatantwas removed, and the absorbance was measured at 280 nm toevaluate mAb precipitation yield by mass balance. The purityof the mAb elution samples was confirmed by SDS-PAGE toensure no residual nanocage.
Nanocage Regeneration and mAb Affinity Precip-itation. ∼100% ligation Z-ELP40-Spy-E2 or Z-ELP80-Spy-E2precipitation yield in elution buffer was evaluated using acentral composite DOE design (25, 50, and 75 μM E2concentration and 0.05, 0.15, and 0.25 M ammonium sulfate)with three replicates of the centerpoint (50 μM and 0.15 M).After adding ammonium sulfate at 23 °C, the samples werecentrifuged at 15,000 × g for 5 min at 23 °C, the supernatantabsorbance at 280 nm was measured to evaluate nanocageconcentration, and the nanocage precipitation yield wascalculated by mass balance. The effect of ammonium sulfateon nanocage regeneration during mAb affinity precipitationwas studied. The nanocage was mixed with an industrial mAbcell culture harvest sample at a 3:1 Z:mAb molar ratio in cellculture media in triplicate for 5 min at 23 °C and centrifuged at15,000 × g for 5 min at 23 °C. The pelleted mAb-nanocagecomplex was washed with PBS followed by 25 mM sodiumcitrate pH 5.0. After another centrifugation, the pellet wasresuspended in elution buffer and mixed for 15 min. Theelution samples were adjusted to either 0.1 or 0.3 Mammonium sulfate for selective nanocage precipitation. Thepurified mAb was removed in the supernatant, and mAbelution yield was calculated by measuring absorbance at 280nm. The pelleted nanocage was regenerated in elution bufferand then buffer exchanged back into PBS using ITC. Thenanocage recovery was evaluated after one purification cycle bymeasuring absorbance at 280 nm.
■ ASSOCIATED CONTENT*S Supporting InformationThe Supporting Information is available free of charge on theACS Publications website at DOI: 10.1021/acs.bioconj-chem.8b00458.
Supporting figures and tables on the Sortase A ligationreactions, SpyTag-E2 expression and characterization,comparison of conjugation methods, and geneticmanipulations (PDF)
■ AUTHOR INFORMATIONCorresponding Author*E-mail: [email protected]. Phone: +1 302 831 6327.
Bioconjugate Chemistry Article
DOI: 10.1021/acs.bioconjchem.8b00458Bioconjugate Chem. 2018, 29, 3113−3120
3119

ORCIDWilfred Chen: 0000-0002-6386-6958NotesThe authors declare no competing financial interest.
■ ACKNOWLEDGMENTS
We would like to thank BMS Biologics Process Development(Devens, MA) for providing the mAb and for their technicaland analytical support. This work was supported by a grantfrom NSF (CBET1403724 and DMR1609621).
■ REFERENCES(1) Gupta, M. N., Kaul, R., Guoqiang, D., Dissing, U., andMattiasson, B. (1996) Affinity Precipitation of Proteins. J. Mol.Recognit. 9, 356−359.(2) Thommes, J., and Etzel, M. (2007) Alternatives to chromato-graphic separations. Biotechnol. Prog. 23, 42−45.(3) Hammerschmidt, N., Tscheliessnig, A., Sommer, R., Helk, B.,and Jungbauer, A. (2014) Economics of recombinant antibodyproduction processes at various scales: Industry-standard compared tocontinuous precipitation. Biotechnol. J. 9, 766−775.(4) Hilbrig, F., and Freitag, R. (2003) Protein purification by affinityprecipitation. J. Chromatogr. B: Anal. Technol. Biomed. Life Sci. 790,79−90.(5) Handlogten, M. W., Stefanick, J. F., Deak, P. E., and Bilgicer, B.(2014) Affinity-based precipitation via a bivalent peptidic hapten forthe purification of monoclonal antibodies. Analyst 139, 4247−4255.(6) Bilgicer, B., Thomas, S. W., III, Shaw, B. F., Kaufman, G. K.,Krishnamurthy, V. M., Estroff, L. A., Yang, J., and Whitesides, G. M.(2009) A non-chromatographic method for the purification of abivalently active monoclonal IgG antibody from biological fluids. J.Am. Chem. Soc. 131, 9361−9367.(7) Pearson, J. C., Clonis, Y. D., and Lowe, C. R. (1989) Preparativeaffinity precipitation of L-lactate dehydrogenase. J. Biotechnol. 11,267−274.(8) Taipa, M. A., Kaul, R. H., Mattiasson, B., and Cabral, J. M.(2000) Recovery of a monoclonal antibody from hybridoma culturesupernatant by affinity precipitation with Eudragit S-100. Biosepara-tion 9, 291−298.(9) Anastase-Ravion, S., Ding, Z., Pelle, A., Hoffman, A. S., andLetourneur, D. (2001) New antibody purification procedure using athermally responsive poly(N-isopropylacrylamide)-dextran derivativeconjugate. J. Chromatogr., Biomed. Appl. 761, 247−254.(10) Madan, B., Chaudhary, G., Cramer, S. M., and Chen, W.(2013) ELP-z and ELP-zz capturing scaffolds for the purification ofimmunoglobulins by affinity precipitation. J. Biotechnol. 163, 10−16.(11) Sheth, R. D., Jin, M., Bhut, B. V., Li, Z., Chen, W., and Cramer,S. M. (2014) Affinity precipitation of a monoclonal antibody from anindustrial harvest feedstock using an ELP-Z stimuli responsivebiopolymer. Biotechnol. Bioeng. 111, 1595−1603.(12) Sheth, R. D., Madan, B., Chen, W., and Cramer, S. M. (2013)High-throughput screening for the development of a monoclonalantibody affinity preicpitation step using ELP-Z stimuli responsivebiopolymers. Biotechnol. Bioeng. 110, 2664−2676.(13) Arosio, P., Jaquet, B., Wu, M., and Morbidelli, M. (2012) Onthe role of salt type and concentration on the stability behavior of amonoclonal antibody solution. Biophys. Chem. 168−169, 19−27.(14) Gagnon, P. (2012) Technology trends in antibody purification.Journal of Chromatography A 1221, 57−70.(15) Swartz, A. R., Sun, Q., and Chen, W. (2017) Ligand-InducedCross-Linking of Z-Elastin-like Polypeptide-Functionalized E2Protein Nanoparticles for Enhanced Affinity Precipitation of Anti-bodies. Biomacromolecules 18, 1654−1659.(16) Dalmau, M., Lim, S., Chen, H. C., Ruiz, C., and Wang, S.-W.(2008) Thermostability and molecular encapsulation within anengineered caged protein scaffold. Biotechnol. Bioeng. 101, 654−664.
(17) Chen, Q., Sun, Q., Molino, N. M., Wang, S.-W., Boder, E. T.,and Chen, W. (2015) Sortase A-mediated multi-functionalization ofprotein nanoparticles. Chem. Commun. 51, 12107−12110.(18) Sun, Q., Blackstock, D., and Chen, W. (2015) Post-translationalmodification of bionanoparticles as a modular platform for biosensorassembly. ACS Nano 9, 8554−8561.(19) Swartz, A. R., Xu, X., Traylor, S. J., Li, Z. J., and Chen, W.(2018) One-step affinity capture and precipitation for improvedpurification of an industrial monoclonal antibody using Z-ELPfunctionalized nanocages. Biotechnol. Bioeng. 115, 423−432.(20) Swartz, A. R., Xu, X., Traylor, S. J., Li, Z. J., and Chen, W.(2018) High-efficiency affinity precipitation of multiple industrialmAbs and Fc-fusion proteins from cell culture harvests using Z-ELP-E2 nanocages. Biotechnol. Bioeng. 115, 2039−2047.(21) Levary, D. A., Parthasarathy, R., Boder, E. T., and Ackerman,M. E. (2011) Protein-protein fusion catalyzed by Sortase A. PLoS One6, e18342.(22) Antos, J. M., Truttmann, M. C., and Ploegh, H. L. (2016)Recent advances in sortase-catalyzed ligation methodology. Curr.Opin. Struct. Biol. 38, 111−118.(23) Row, R. D., Roark, T. J., Philip, M. C., Perkins, L. L., and Antos,J. M. (2015) Enhancing the efficiency of sortase−mediated ligationsthrough nickel−peptide complex formation. Chem. Commun. 51,12548−12551.(24) Zakeri, B., Fierer, J. O., Celik, E., Chittock, E. C., Schwarz-Linek, U., Moy, V. T., and Howarth, M. (2012) Peptide tag forming arapid covalent bond to a protein, through engineering a bacterialadhesin. Proc. Natl. Acad. Sci. U. S. A. 109, E690−E697.(25) Meyer, D. E., and Chilkoti, A. (2004) Quantification of theeffects of chain length and concentration on the thermal behavior ofelastin-like polypeptides. Biomacromolecules 5, 846−851.(26) Hassouneh, W., Christensen, T., and Chilkoti, A. (2010)Elastin-like polypeptides as purification tag for recombinant proteins.Current Protocols Protein Science 61, 6.11.1−6.11.16.(27) Reichheld, S. E., Muiznieks, L. D., Keeley, F. W., and Sharpe, S.(2017) Direct observation of structure and dynamics during phaseseparation of an elastomeric protein. Proc. Natl. Acad. Sci. U. S. A. 114,E4408−E4415.(28) Chen, X., Zaro, J. L., and Shen, W. C. (2013) Fusion proteinlinkers: property, design and functionality. Adv. Drug Delivery Rev. 65,1357−69.(29) Liu, F., Luo, E. Y., Flora, D. B., and Mezo, A. R. (2014)Irreversible sortase A-mediated ligation driven by diketopiperazineformation. J. Org. Chem. 79, 487−492.(30) Guimaraes, C. P., Witte, M. W., Theile, C. S., Bozkurt, G.,Kundrat, L., Blom, A. E. M., and Ploegh, H. L. (2013) Site-specific C-terminal internal loop labeling of proteins using sortase-mediatedreactions. Nat. Protoc. 8, 1787−1799.(31) Reddington, S. C., and Howarth, M. (2015) Secrets of acovalent interaction for biomaterials and biotechnology: SpyTag andSpyCatcher. Curr. Opin. Chem. Biol. 29, 94−99.(32) Domingo, G. J., Orru, S., and Perham, R. N. (2001) Multipledisplay of peptides and proteins on a macromolecular scaffold derivedfrom a multienzyme complex. J. Mol. Biol. 305, 259−267.(33) Jendeberg, L., Persson, B., Andersson, R., Karlsson, R., Uhlen,M., and Nilsson, B. (1995) Kinetic analysis of the interaction betweenprotein a domain variants and human Fc using plasmon resonancedetection. J. Mol. Recognit. 8, 270−278.(34) Lamla, T., and Erdmann, V. A. (2004) The nano-tag, astreptavidin-binding peptide for the purification and detection ofrecombinant proteins. Protein Expression Purif. 33, 39−47.
Bioconjugate Chemistry Article
DOI: 10.1021/acs.bioconjchem.8b00458Bioconjugate Chem. 2018, 29, 3113−3120
3120