specialization of the right hemisphere for visuomotor control csi
TRANSCRIPT

,~rurop,~‘i,,,loy,u. vol 2X. ho 8. pp 763-715. 1990 Pnnted ,n Greal Bnlam
002R-3932 90 53 oO+” 00 t 1990 Perpamon Prey plc
SPECIALIZATION OF THE RIGHT HEMISPHERE FOR VISUOMOTOR CONTROL
R. M. BRACEWELL*, MASUD HUSAIN~ and J. F. STEIN
University Laboratory of Physiology, Parks Road, Oxford OX1 3PT. U.K
(Received 11 Ju/J~ 1989; accepted 9 January 1990)
Abstract-The accuracy of saccades directed towards the remembered positions of targets in left (LVF) or right (RVF) visual hemifield was measured. The majority of right-handed subjects were found to be more accurate at directing their gaze to locations in the LVF than in the RVF. suggesting that the right hemisphere is superior to the left in oculomotor control. Even after completion of a corrective saccade following the primary saccade, subjects systematically undershot target direction and overshot target depth, suggesting that visual feedback normally plays an important role in the fine guidance of gaze after the completion of a primary saccade.
INTRODUCTION
KIMURA [27] first reported that subjects localize small visual stimuli more accurately in the left visual hemifield (LVF) than they do in the right (RVF), suggesting that the right hemisphere may have a special role in perceptual localization. Although Kimura’s results have been confirmed by some investigators [28,29,44] others have either failed to show any differences in accuracy between the hemifields [4,7,8] or have even demonstrated a RVF
advantage [41]. In these perceptual experiments, subjects were required to judge the remembered position
of a visual stimulus with respect to another visual cue, e.g. a grid pattern. However. by far the largest percentage difference in performance between the hemifields has been found on a task which requires subjects to point to targets without visual feedback of the hand. GUIARD et al. [20] showed that pointing, with either left or right hand, can be 25% more accurate in the LVF than it is in the RVF (cf. 7.5% in Kimura’s Experiment 5). This finding may reflect differences in visuomotor localization between the hemispheres. Alternatively, it may be due to a superiority of the right hemisphere for manipulo-spatial ability [14].
In the study presented here, we investigated whether differences in accuracy between hemifields can be demonstrated in another type of visuomotor behaviour, the directing of saccadic eye movements. We show that a LVF advantage can be demonstrated for most right-handed subjects when they are asked to make saccades to the remembered position of visual targets. i.e. without the aid of visual feedback. This result, in conjunction with the results of the pointing experiment of GUIARD et a/. [20], suggests that the right hemisphere is specialized for visuomotor control. A preliminary report has been published in abstract form
CSI.
*Present address: Department of Brain and Cognitive Science. Massachusetts Institute of Technology. Cambridge. MA 02139, U.S.A.
tAuthor for correspondence.
763

764 R. M. BRACEWELL. MAS~X HUSAIN and J. F. STEIN
METHODS Saccades of amplitude > 7.5 are usually executed in two steps-a primary movement. followed by a corrective
saccade [24]. The question arises as to whether saccades are normally pre-programmed in two steps or whether the corrective movement results from the regtstration ofan error by the oculomotor system. There is evidence that visual information presented during a saccade may be used to direct a corrective saccade after the primary mov’ement [Zl, 221. Therefore. measurement of the total angular excursion of the eyes (i.e. primary + corrective saccade) with the target continuously lit is not a good indicator of where the target IS nntially locahzed by the oculomotor system.
But it has also been found that subjects make corrective saccades when directmg their gaze to the remembered position ofvisual targets [3, 491. i.e. in a situation where there is no visual feedback either during or after the primary saccade. This may be because the saccadic system preprogrammes movements in two steps. If this is so. the amplitude of the primary saccade alone is also unlikely to be a good measure ofspatial localization. Therefore. m this study. we have measured the accuracy of the total excursion of gaze tie. primary + any correctivjc saccade) when no visual feedback was available during or after the saccade.
Twenty right-handed subjects. 10 males and 10 females. aged between 20 and 23. were studied (mean lateralit! quotient =78 on Edinburgh Inventory; [37]). All had normal vision. SubJects sat at a viewing distance of 50 cm from a monitor screen. Their head movements were restricted by means ofa bite-bar. They were required to fixate a central spot (subtending 0.04. at the eye) which wsas presented at eye level for’two seconds. after which it was extinguished.
Subjects continued to fixate centrally in the dark for a further second until a small target light (also 0.04 ) appeared for 100 msec at positions which were randomly varied between 2.5 and 10 horizontally to the left or right of the midline. Subjects were required to move their eyes to the point at which the target had flashed. They returned fixation to the central spot when it reappeared 2 set after the target had been extinguished. Since mean saccadic reaction time in this paradigm was 280 msec, subjects were making saccades to the remembered positions of targets. We find no evidence of express saccades of latency < 150 msec when naive subjects perform this task.
We used this particular delayed saccade protocol with a 1 set wait period tn order to eliminate any visual cue (left by persistence) of the central fixation light. Although small microdrifts occur durmg the wait period. there are no systematicdrifts to the left or right within individuals. Targets vvithin the central region ofvision ( < k 2.5 ) were not used because this area of the visual field, or part of it. may be represented in the primary visual cortices of each hemisphere through either callosal or direct geniculostriate connections [9. 251. It should be noted. however. that callosal transfer may be responsible for much larger representations of the ipsilateral visual hemifield in extrastriate cortical areas [Z. 19, 521.
The subjects were given ten practice trials to famiharize themselves with the task. They then performed three blocks of 30 trials. The same pseudorandom sequence of target positions was presented to each SubJect. There were 44 targets in the LVF and 46 in the RVF. The distribution of targets in each hemifield was not significantly different.
The position of each eye was recorded throughout each block oftrials usrng an infrared photoreflectometer. which we calibrated to possess a resolution ofO.06 degree of arc (see also [50] ). Tvvo frequency modulated infrared beams were projected at the high contrast border betvveen the sclera and iris. The radiation refIccted back was measured by two small sensors. The differential of the phase-locked. amplified transducer signals provsides. within k 1 I . 3 sensitive and linear measure of horizontal eye position [50]. Calibrations were performed before each block of trials.
Eye position was measured 200 msec after the completion ofeach saccade. This procedure allowed us to take into account any corrective saccade that might follow the primary movement. Post-saccadic eye position was compared with the eccentricity of the target in each trial in order to determine the accuracy of localization.
All SubJects made disjunctive saccades, i.e. the movements ofeach eye were not ofexactly the same amplitude. so that the point of fixation lay either slightly in front of. or behind, the plane of the display. DitTerences between the visual hemifields for both version (direction) and vergence (depth) accuracy were thcrcfore nnalysed. Details of computations are given with Fig. I.
RESULTS
Since there were no significant differences in the distribution of targets between the hemifields, an estimate of overall accuracy in the LVF or RVF was obtained simply by collapsing the data for each hemifield across eccentricities. These results were further analysed in terms of the constant (mean) error. i.e. systematic under- or overshoot, and the variable error, i.e. the stndard deviation, of version and vergence measures.
Constnnt err01
Overall. subjects tended to undershoot the direction of the target; the mean constant error was 0.21 version (referred to as -0.21 ) which corresponds to an underestimate of target

KItiHT HEMISPHERIC SPECIALIZATION FOR VISUOMOTOR CONTROL 165
plane of target posltlon
dosplay -3 i
‘.. \ ‘\ \
T
I- -l
Fig. I. The amplitude of the saccade (+corrective movement. if present) made by each eye was computed. These are denoted angles L and R. for left and right eye respectively. L and R give deviations from straight ahead of a line of sight drawn from each eye to the plane of the display. The angle subtended between the target and a perpendicular drawn from the mid-pupillary position is denoted T. The accuracy of localization in the plane of the display was determrned by computing ((L- 7)+ (R- 7))/2. This gives the directional error in degrees of version. The angular vergence error, I(;-/I) was computed by solving for r. smce r=;-/I By simple geometry Z= tan(50 i)- tan(d,:i). where i is half the interpupillary distance and d is the distance in cm from the interocular plane to the plane oflocalization. Using similar triangles (shaded)and solving for tan R. it can readily
be shown that d=X(tan R-tan L(X’50)).
position of 3% over all target locations. In depth, subjects tended to overshoot the target by 0.10 vergence (Table 1). Analysis of variance (hemifield x sex) with constant error as the dependent variable revealed no significant differences between hemifields or sexes for either version or vergence: nor was there a significant interaction between these variables
[F(l. 36)< 1.4. P>O.2].
Significant differences did emerge, however, upon analysis of variable errors (standard
deviations) both in version and vergence. For version. the mean variable error in the LVF was 0.57 . whereas in the RVF it was 0.72“
(Table 2). Analysis of variance (hemifield x sex) revealed that the difference between hemitields was significant [F( I, 36)=8.3, P<O.Ol] but, again. there was no effect of sex [F (I. 36)=O.I ). Nor was there any significant interaction between these variables
[F(I. 36)=0.6]. For vergence. the mean variable error was 0.79 in the LVF and 0.95 in the RVF. Analysis
of variance once again demonstrated that the LVF advantage was significant [F (1. 36) = 4.7, P<O.OS] and no effect of sex [F (I. 26)=0.6]. But there was a trend towards a significant sex x hemifield interaction [F (I. 36)= 2.2. P=O. I]. Close inspection of the vergence variable error data (Table 2) shows that whilst males were more accurate in the LVF than in the RVF (0.76 vs I .03 ) females tended to perform equally well in both (0.81. and 0.86’ in LVF and RVF respectively). Moreover. their performance was at about the level of

lhh R M. BKACEM.ELL. MASUII HLSUV and J. F. STEI\
Table I. Constant errors m dlrectlon nnd depth pooled xros~ txget eccentrlcitles for each subJect
(21 Constant error m directIon tiersion degrees)
FcIlXlle~ M3k\ LVF R\‘F LVF RVF
(b) Constant error in depth (\erpencc degrechl
Femalea MAC\ LVF RVF L\‘F R\‘I
Mean SE
0.11 0.93 0.0s 0.24 0.22 - 0.03 ~0.0’ - 0 4’)
0 49 0.h2 PO.1 I -0.1 I
- 0.08 PO.3 0.1-l 0.36
-02x 0.39 0.00 PO.12
-0.20 - 0.72 0.57 0.25
O.hh - 0.75 po.o: 0.1 i po.3: 0.3R ~0.72 0.72
0.20 po.5x 0.:2 o.ii 0.21 - 0.0.3 - 0.1 I I.16
0.1 I - 0.02 0.01 0.21 0.10 0.1’) 0.1 I 0 I5
accuraq of males in the LVF. In short. the trend towards ;I significant scs x hcmilield interaction for vergence errors was due to ;I difference bctwcen the scxcs in pcrformancc in the RVF: in this hemitield males were less accurate than kmales.
Only tkvo subjects. both females (female subjects 7 and X in Table 2). sho\ved lower variable error for version and vergence in the RVl-’ than in the LVl-
Although overall accuracy in each hemifield rcvcaled dill’ercnccs in variable error bctwccn LVF and RVF, it is of some interest to know if this asymmctr~ changed with eccentricity.
I trritrhle errors in version and vergence incrcascd with target occcntricity in both hemifields (Fig. 3). Moreover. the gradients of the error-eccentricity functions for the RVF were significantly larger than those for the LVF (for version. I = - 3.56. I’< O.OOI: for \ergence. t = -6.75. P<O.OOl ). That is. dif~crcnccs bctb+ccn the hcmificlds in variable error increased \vith target eccentricity. Details of regression arc gven with Fig. 2.
Plots of co1~~f~u7~ error against target eccentricity also re\eal interesting relationships. For \,ersion. the magnitude of the u~I~/~~~~s/u~~~ incrca~cd with ccccntricity. Furthcrmorc, it

KIGHT HtZIISPHEKI(’ SPE(‘IALIZATIOI; FOK \‘ISUOMOTOR COZTKOL 76’
Table 2. Variable errors m dIrectIon and depth pooled ucroa target eccentricities for each subject.
The order of subjects is retained rrom Table 1
(a) Varlahle error In direction 1 version degrees j Females Male5
LVF RVF LVF RVF
0.36 0.51 0.69 0.80 0.54 1.1x 0.32 0.4 I 0.58 0.85 0.79 0.9 I 0.57 0.90 0.43 0.76 0.54 0.6X 0.56 0.69 0.56 0.84 0.35 0.76 0.54 0.45 0.76 0.X? 0.74 0.45 0.75 0.65 0.53 0.97 0.56 0.55 0.59 0.71 0.58 0.57
Mean 0.56 0.75 0.58 0.69 SE 0.03 0.08 0.05 0.05
(b) Variable error in depth (vergence degrees)
Females Males LVF RVF LVF RVF
0.53 0.9 I 0.x4 1.47 0.85 1.13 0.50 0.66 0.86 0.93 0.84 0.62 0.9 1 0.70 0.6X 1.1 I 0.84 0.90 0.53 0.x7 0.46 0.89 0.63 0.78 0.74 0.50 0.76 0.95 1.19 0.72 0.63 1.13 0.92 I.21 1.17 1.26 0.82 0.74 0.99 I .47
Mean 0.81 0.86 0.76 I .O! SE 0.07 O.O? 0.07 0.10
increased significantly more in the RVF than in the LVF (top panels of Fig. 3; r=4.80, P<O.OOl ). For vergence, the magnitude of the OIYYS~ICN~~ increased with target eccentricity. However. for this measure. there was no significant difference between the error-eccentricity functions in each hemifield (lower panels of Fig. 3; f = 1.62, P>O. 1).
We considered the possibility that the differences between the error-eccentricity functions in each hemifield were due to the performance recorded for the larger eccentricities in the RVF. However, removal from the data-base of the eight largest target eccentricities in the RVF makes little difference to the results.
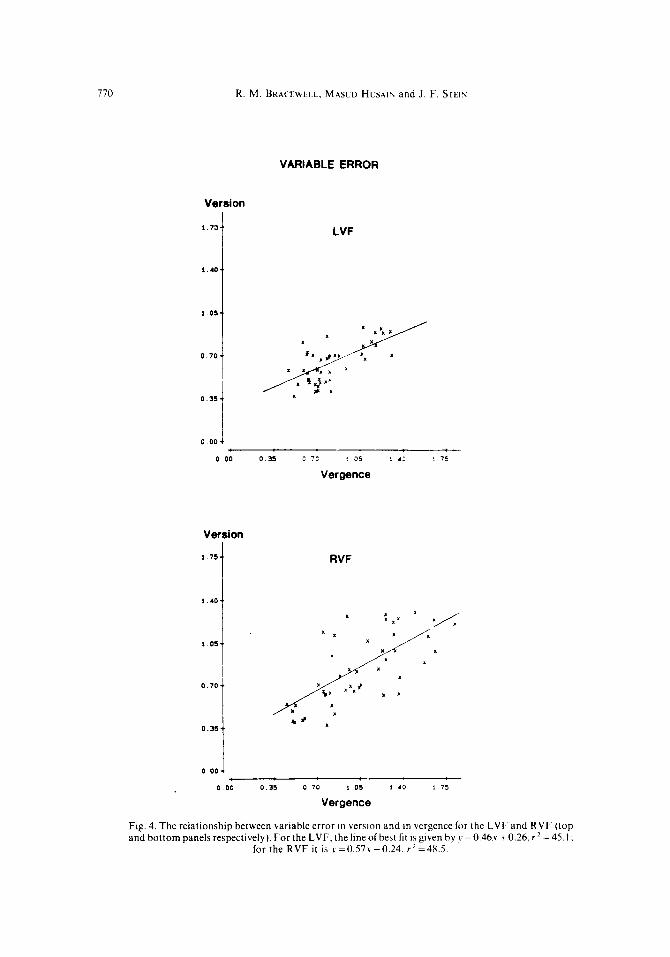
Tlw ~.~I~~fio~~.sl~ip hrt\twrl c~‘~‘o~‘s in wrsioiz cud rqpwc
The above analysis suggests that variable error in version and vergence may be related since both increased with target eccentricity. Exactly how were theerrors in version related to those in vergence’?
Figure 4 illustrates the version-vergence relations for variable error. These are shown separately for each hemifield because the gradients differ significantly (r = - 5.63, P<O.OOl).

768
VE 1 LVF
R. M. BRACEWELL. MASUD HUSAIK and J. F. STEIN
VERSION
VE / RVF
1 .o.
00 75 50 ‘5 100
eccentricity
VERGENCE
VE IRVF
00 z, 50 ‘5 100 00 25 50 ,I 100
eccentricity eccentricity
Fig. 2. Variable error (VE) plotted against target eccentricity in LVF and RVF. The top panels show the relationship for errors in direction (in angles version); the bottom panels for errors in depth (in angles vergence). In both cases,error increases significantly more with eccentricity in the RVF than in the LVF. I.e. asymmetry increases with target eccentricity. For version, the lines of best fit are given by ~~=0.07~+0.27,r2=55.9(inLVF)and~=0.10x+0.l9,r2=72.9(inRVF).Forvergence.thelinesof best fit are ~=0.08x+O.37, r2=38.1 (in LVF) and ~=0.13\-+0.27. rZ=71.0 (in RVF). All of these
regressions are significant at Pi 0.001.
But the important point is that in both hemifields the slope of the versionvergence function is about 0.5, i.e. variable error in version increased by approx 0.5 for every degree of error in vergence. Unlike variable errors, constant errors in version and vergence were not related. Furthermore, there were no significant differences between the hemifields (t =0.66, P~0.4).

CE 1 LVF
KIGHT HEMISPHERIC SPECIALIZATION FOR VISUOMOTOR CONTROL
VERSION
-0.901
DO 25 50 75 :oo
eccentricity
-0 90 J 00 2, ,o -5 :oo
eccentricity
769
VERGENCE
CE LVF
i
CE RVF 0 10 0 I0 I
* X I I . .
-0 M 1. -0 SO I_ PO *, ID 71 100 00 21 50 -5 100
eccentricity eccentricity
Fig. 3. Constant error (CE) plotted against target eccentricity in LVF and RVF. The top panels show the relationship for errors in direction (in angles version): the bottom panels for errors in depth (in angles vergence). Positive error values represent overshoots; negative values undershoots. For version. error increases significantly more with eccentricity in the RVF than in the LVF. For version. thclinesofbest lit aregiven by!= -O.Ol.v-0.12.rz=3.6(in LVF)andj’= -0.08.~+0.26.r’=56.0 (III RVF). For vergence. the lines of best lit are J=O.OXY-0.38. ~‘=38.2 (in LVF) and \.=0.06~-0.2S.r2=24.6(in RVF).All oftheseregressionsaresignificant a1 P<O.OOl.except thefirst
where P=O.l.
Corrective saccades were defined as saccades occurring within 200 msec of the completion of the primary movement (cf. [3. 24.493). Most (91%) such corrective movements occurred when target eccentricities were > 7.5‘. Their frequency varied from
770 R. M. BRACEWELL. MAXD HUSAIN and J. F. STEIY
VARIABLE ERROR
Version
1.0s..
0.70..
0.35 I 0 004
LVF
0 00 0 3s c 7c : 05 I .L : 75
Vergence
Version
1.75 1 RVF
1 01
I 0 70 I ’
0.004 l
0 00 0.35 0 70 1 05 1 .O * 75
Vergence
Fig. 4. The relationship between variable error in version and in vergence for the LVF and RVF (top and bottom panels respectively). For the LVF.thelmeofbest fit ~sgiven byy=0.46.~+0.26,rZ =45.1;
for the RVF it is ~=0.57.~+0.24. r*=48.5.

RIGHT HEMISPHERIC SPECIALIZATION FOR VISUOMOTOR COKTROL 771
subject to subject (from 2-16% of saccades). Seventy-four per cent of corrective saccades
were 10% or less of the amplitude of the primary saccade. Only 2% of corrective movements
were dysconjugate.
DISCUSSION
The experiment reported here was designed to determine whether there are differences in accuracy between saccades directed to targets in the left and right hemifields. We found that overall uariable error (i.e. across all target eccentricities) in both version (direction) and vergence (depth) was significantly smaller in the LVF than in the RVF. Overall comrmt error, in direction or depth, did not differ significantly between the hemifields. However. the increase in constant error in version with eccentricity was significantly larger in the RVF than in the LVF. These results suggest that the right hemisphere is superior to the left in directing saccades to remembered visual targets.
Our results are consistent with several sets of findings. First, it has been shown that conjugate paralysis is more common following lesions of the right hemisphere than of the left [ 1 I, 361. Second, in subjects with left cerebral language lateralization, gaze deviation is more common and severe following injections of sodium amylobarbitone to the right carotid artery than the left [33]. Third, tasks requiring eye movement produce greater interference (increase in reaction time) in the LVF than the RVF [47]. Finally. eye movement reaction times of patients with lesions to the right parietal lobe are longer than those with damage to the corresponding area in the left hemisphere [ 151.
On the face of it our data are not consistent with studies of saccadic reaction times in normal subjects which have demonstrated shorter latencies to targets presented in the RVF than in the LVF [26,40]. But it would be interesting to know whether there was a trade-off between spatial accuracy and reaction time in those particular experiments.
Constunt and variable errors
It has been demonstrated in right-handed individuals that, using either hand, constant error in pointing without visual feedback is much smaller towards targets in the LVF than in the RVF [20]. Constant error has previously been identified with central motor programs [IS, 383, so this result suggests that the right hemisphere is superior to the left in proyrumrniry arm movements to visual targets. Our findings regarding constant error for eye movements are not so strong since differences between the hemifields were not apparent at small eccentricities. However. as target eccentricity increased so the LVF advantage of constant error in version became more apparent. Thus. our constant error data also support the contention that the right hemisphere is superior to the left in programming movements to visual targets.
Differences between the hemificlds for variable error were stronger than for constant error in our experiments. Overall. LVF advantages of 12 and 9% for version and vergence respectively were recorded (where advantage is computed from RVF,,,,, - LVF,,,,,/ RVF,,,,, + LVF,,,,,). These values are much larger than LVF advantages (computed in terms of percentage correct performance in each hemifield) reported in perceptual experiments, in which 7.59;) was the largest value to be recorded [27].
Variable error is considered by some to be solely an indicator of the accuracy of morewent c~.xc~mtior~ processes. e.g. inherent “noise” in the muscles, since, unlike constant error. it

771 R. M. BRACEWELL. MASLD HUSAIN and J. F. STEIY
appears (in investigations of the skeletomotor system) to be more sensitive to movement velocity and amplitude than to experimental conditions such as manipulations of target duration [43, 481. Thus. variable error has been associated with movement performance, not programming.
It is probable. however. that the process of localization is itself not entirely accurate; and that this variability may add to that in the motor system. GNADT et al. have recently found that the variability of saccades to remembered positions increases as the delay between seeing the target and movement onset is lengthened [16]. It was also shown that there is a concomitant fall in peak saccadic velocity. This reduction in velocity is in the direction opposite to that predicted by those models of motor control which consider variability to be a consequence of “noise” in the execution process [48, 511. It is unlikely, therefore. that characteristics of the oculomotor plant alone are responsible for the variability in saccadic end-position. Rather, a variance associated with the memorized location of the target influences the observed variable error.
Thus, variable error, as well as constant error. may be regarded as a measure of oculomotor programming. Therefore, if a LVF advantage is taken to be indicative of a right hemispheric advantage, our data (for both constant and variable errors) suggest that in most right-handed individuals the right hemisphere is superior to the left in programming saccades to remembered positions.
We suggest that one aspect of programming in which the right hemisphere is superior to the left is in localizing visual targets. This would be consistent with the results of several experiments which show a LVF advantage for perceptual localization [27-29, 441. In these previous experiments, subjects were required to indicate where they judged a visual stimulus to have been in relation to other visual cues. i.e. without moving their arm or eyes to the remembered position of the stimulus. Whilst these results suggest that the right hemisphere is specialized for perceptual localization, our findings suggest that the right hemisphere is also superior to the left in risuomotor localization. This hypothesis is consistent with the findings of GUARD er ul. [20] who demonstrated that reaching with either hand is more accurate to targets presented in the LVF. Certainly, there is little evidence to support the suggestion that it is the right kand which is superior to the left in visuomotor control [45].
One region within the right hemisphere which may be responsible for visual localization is the posterior parietal cortex. There is much evidence to suggest that this area plays an important role in both perceptual and visuomotor localization [IO]. It has also been demonstrated that one cytoarchitectoniczone within the posterior parietal cortex is larger in the right hemisphere than on the left [13] and this area may have an important role in directing saccades to visual targets [39]. *
A slightly different explanation to that offered above is that the difference we have observed reflects a superiority of the right hemisphere in rc>r~lcn~hrrir~c/ the spatial location of visual targets. There is certainly considerable evidence to suggest that spatial aspects of memory may be preferentially lateralized to the right hemisphere [IO]. However. it must be appreciated that the latency ofsaccades on this paradigm is very short (mean: 280 msec) and comparable to that observed with saccades to continually lit visual targets. A rcmembercd saccade paradigm was employed simply to ensure that there was no visual feedback of the target during the movement or after the completion of the primary saccade. We have no evidence to suppose that of the many visuomotor control mechanisms involved, the process of remembering spatial location alone is responsible for the hemifield differences that WC have
observed.

RIGHT HEMISPHERIC SPECIALIZATION FOR VISUOMOTOR CONTROL 773
Sex diferences
It has previously been reported that LVF advantages for visual localization are more pronounced in males than females [27-291. In this investigation, for variable error in vergence, there was a similar trend. Females appeared to be just as accurate in the RVF as they were in the LVF; males were as accurate as females in the LVF but they were worse in the RVF (cf. [32]).
Localization in depth
The present investigation is the first divided visual field study in which localization has been assessed in depth as well as in direction, although a number of other studies have examined depth perception in each visual field using random-dot stereograms [6, 12, 171. The demonstration of a LVF advantage for variable error in vergence as well as in version on a task in which essentially only the direction of the target was varied may seem rather surprising. But there is a simple explanation.
The hypothesis has been advanced that perceptual representations of space are encoded in version-vergence co-ordinates [30]. In other words, eye position signals from both eyes are required to localize a target in depth as well as direction. Let s,Z,, be equal to the variance associated with retinal localization by each eye (this obviously scales with eccentricity). Let s & equal the variance associated with the eye position signal, again from each eye. If both these variance terms are the same for each eye, the overall variance ofdirectional localization using binocular vision is given simply by the expression s:~, +s&~ degrees version.
For binocular depth localization, however, the situation is dilferent. In order to compute error in degrees vergence, the variance terms for each eye must be added to give 2(s;‘,, + s&). This is because a given error in estimating direction in each eye is associated with double that error in degrees vergence (refer to Fig. 1). Variable error in vergence is therefore inevitably double that in version if target position is constructed using eye position signals. Exactly how retinal and eye position signals are combined is a matter of current neurophysiological investigation. Interestingly, there is evidence to suggest that neurones within the posterior parietal cortex of monkey are modulated by both version [l] and vergence angle [46].
Implications for the control of saccadic eye movements
Various workers have reported that corrective saccades are sometimes made when subjects are required to make saccades in the dark [3,49]. The results of these studies, as well as our own, argue against the idea that retinal error signals are necessarJ9 to generate small corrective saccades [3 1,421. They do not, however, exclude a role in oculomotor control for visual feedback at the end of a saccade.
Clearly, our data show that er’eri rvlrelt the!. rmtlc corrective movements, subjects made errors in localization in three-dimensional space: In terms ofdirection, the mean error was an undershoot of 0.21’ (variability: 0.65”) which corresponds to a mean underestimate of 3% over all target positions. This agrees well with another finding of direction underestimation of 4% [34]. Saccadic localization in depth. however. has not previously been measured. We found that constanr errors corresponded to disparities (differences in angular vergence) which are just within Panum’s fusional limit (approximately 0.2‘ at the fovea [35]), but the mwiahle errors were so large that it would not have been possible consistently to fuse the target had it been illuminated at the end of the saccade. HALLETT and LIGHTSTONE [21, 221 showed that visual information presented during a

774 R. M. BKACEWELL. MASULI HUSAIV and J. F. STEIN
saccade may be used to direct a corrective saccade after the primary movement. Our results suggest that the oculomotor system may also use visual information at the end of a primary saccade to direct the fine guidance of gaze onto a target. Therefore, saccades appear not to be entirely preprogrammed. Furthermore, a comparison of desired eye position with that achieved after the primary saccade [49] is not sufficient to generate the correct error signal for the corrective saccade: visual inspection appears to be necessary as well [31.42].
In summary, the findings presented in this study demonstrate that saccades are directed more accurately to the remembered positions oftargets in the LVF than to those in the RVF. Furthermore. it is shown that although visual feedback is not necessary to generate corrective saccades, it is likely to have a role in the fine guidance of gaze after the completion of the primary saccade. We propose that the left hemifield advantage reported here relates to the superiority of the right hemisphere in localizing visual targets.
A~~rlo~c/edyemr,Its-We are grateful to Dr. F. H. C. Marriott for his advice on statistical analyslb.
I.
2.
3.
4. 5.
6.
7.
8.
1).
IO. II.
12.
1.
IJ. Ii.
16.
17. IX. 19.
20
?I.
23
REFERENCES
ANU~KSEN. R. A., Ess~cr. G. K. and SIEGEL, R. M. Encodmg of spatial location by postertor parietal neurons. Scrrncc~ 230, 456-458. 1985. ANTONINI. A., BEKLUCCHI, G. and LEPORE. F. Physiological organization of callosal connections of a visual lateral suprasylvian cortical area in the cat. .I. Nruroph~siol. 49, 902 ~921. 1983. BECKER. W. and FUCHS, A. F. Further properties of the human saccadlc system: eye movements and correctton saccades with and without visual fixation points. i’ision Rrs. 19, 967 983. 1969. BIKKETT. P. Measures of lateraiity and theories of hemispheric processes. ~~~,u~~,ps~t,ho/oc/itr 15.693 696. 1977 BRACEWTLL. R. M.. HUSAIN. M. and STEIN. J. F. Differences in the accuracy of human saccadcs In left and right visual fields. J. Physiol. 382. 98~. 1987. BKEITMEYEK. B.. JULESZ. B. and KROPFT. W. Dynamic random-dot stereograms reveal up iloun anIsotropy and left-right isotropy between cortical hemifields. S&we 187. 369-270. 1975. BRYIIEN. M. P. Perceptual asymmetry in vision: relation to handedness. eycdness and speech lateralization. C‘0rre.y 9, 419435. 1973. BKYDE~. M. P. Response bias and hemispheric differences in dot locahzation. Perccpr. I’.s~~c~hop/~~.~. 19, 23 2X. 1976. BUNT. A. H.. MIYCKLEK. D. S. and JOHANSON. G. W. Demonstration of the bilateral proJection of the central retina of the monkey with horseradish peroxidase neurograph). J. ~‘ornp. ~Vclrrol. 171. hlY 630. 1077 Dt. RESZI. E. Disordrrs o/ Spurial E.~p/orurw~~ ud C‘oyrlirim. John Wiley. Chlchcstcr. 19X?. DI Rrxz~. E.. COLOhlBO. A.. FAGLIONI. P. and GILHEKTOP;I. N. ConJugate eazc parcscs 111 strohc potlents with unilateral damage: an unexpected instance of hemtspheric asymmetry. rl~~/z.\ Acwro/. 39, 4x2 4X6. 19X?. DUKUFOKU. M. and KIMURA. D. Right hemisphere specialization for depth perception rcllected in \ibual tield differences. Nurrcrc~. Lord. 231, 394-395. 1971. EIUFLHFRC;. D. and GALABURI)A. A. M. Inferior parletal lobule. Diverecnt cytoarchltectonlcasymmetrlcs in the human brain. Arc~hs Nrurol. 41, 843 -852, 1984. G,\\%~.%%Ic;A. M. S. and Lt DOUX, J. E. 7%~ Ime<qrarct/ .I/rm/. Plenum Prcsb. NC\\ 1’orh. 107X. GIIW~~I. F.. CASAZZA. M.. Mus~c~o. M. and A\A%zIvI.(;. Oculomotor dlaordcrs in cortical Icsmns in man: the role of unilateral neglect. :V~urops!rkolt~~/~u 21, 543 553. 1983 GZAIIT. J. n’.. BKACEW~LL, R. M. and ANIIIIKSLN. R. A. Saccadlc eye movementa ulthout visual lccdhach: memory-linked sensorlmotor spatial transformation. Swq /or ~~rlcrrj.\~,c,rl(,c .Ah.srruc~l 13. 1090. IYX7. GKABO\VS~C,\. A. Lateral differences in the detection ofstercoscopic depth. iVelcr~‘p.~~(,II”/l,(/irr 21.24’) 257. 19x3. GKAVI~. R. Constant errors In the execution and appreclatmn of movement. Hrcrrt~ 95, 640 660. 1Y7?. GKOSS. C. G.. BI.NI)EK. D. B. and MISHKI~. M. Contributronh of the corpus callosum and the anterwr commlssurc lo visual activation of inferior temporal neurons. Brtrljl Rr,.\. 131, ‘27 23Y. lY77. GIIAKII. Y.. DIAI. G. and BI.A~;HATON. D. Left-hand advantapc m right-handcr\ for apat~al constant error: prellmmary e\,idence in a unimanual ballistic aimed movement. ~c’to~,p.\?.~,/,,,lljyrcr 21, I I I I t5. lYX3. HALL~TI. P. E. and LIC~HTSTOW:. A. D. Saccadic eye movement\ toward\ \ttmu11 triggered by prior saccades. I.I\/ou Rr\. 16, 99 106. 1976. HAI.L~TT. P. E. and LICIHTSTOUL. A. D. Saccadic eye movement\ to tlashcd tarpcts. I’r\!o?l Kc.\. 16. 107 114. I976

KIC;HT HFIIISPHEKIC SPECIALIZATIO’; FOK VISUOMOTOK COSTKOL 77;
23. HASSLE. R. M. and !k%\‘ENSKi. A. A Accuracy ofspat~al localizations near the time ofsaccadtceye mo\emcnts. C7.\1on Re\. 25. 1077-1082. 1985.
24. HEN%)%. D. B. lnvesttgation mto corrective saccadic eye movements for retixation amphtudes of IO degrees and belou. 17sion Rrs. 19. 57. 61. 1979.
25. HUBCL. D. H. and WIESEL. T N. Cortical and callosal connections concerned with the \erttcal mertdian of\isual fields m the cat. J. ,Yruropl~~.srol. 30. IS61 -1573. 1967.
26. HUTTOV. J. T. and PALET. J. Lateral saccadic latenctes and handedness. .~ru~op.s~c~/~(~Ir,yiu 24. 449 151. IYXh. 27. KIS~I.KA. D. Spatial localizatton in the left and right visual fields. Curl. J. Pcrciwl. 23. 445 45X. IYhY. 1X. LLVY. J. and REII). M. Variattons m Hriting posture and cerebral orpamsation. St rrnw 194. 337 33Y. 1976 29. LEVI’. J. and REID, M. Variations in cerebral orgamsation as a functton of handedness. hand posture III wrttng
and sex. J. rup. Psrckol. 107, 119-144. 1978. 30. LL~BFK~. R. K. The Metrtc of Btnocular Visual Space. J. Opr. Sot,. .-lrn. 40. 627-642. 1930 31, MATHEK. J. A. Some aspects or the motor organization of the oculomotor system. J. .‘Lfor. &/I[II. 17, 373 387.
19x5. 3’. M~GLoNE.. J. Sex differences in human bratn asymmetry: a critical sur\cy. T/w &/WI I Brc~ru SC. 3, 215 263.
1980. 33. Mtaoo~. K. J.. LOKISG. D. W.. LLE. G. P.. Btwoss. B. S.. NICHOLS. F T.. THOhlPSOx. E. E.. THOUPSOX. W. 0.
and HElLhlAS. K. M. Hemisphere asymmetry for eye gaze mechanisms. Brarn 112. 103-l I I. 19X9. 34. MILLER. J. M. Information used by the perceptual and oculomotor systems regarding the amplitude ofsaccadic
and pursutt eye movements. I ‘isron Rr.5. 20. 59 68. 1980. 35. MITCHELL. D. E. Retinal disparity and diplopia. I’~.sron Rrs. 6, 441 -351. 1966. 36. MOHK. J. P.. GAUTIEK. J. C.. HIEK. D. B. and STEIX. R. W. In Strokr Vol. I, H. J. M. BAKUETT. J. P. MOHK. B. M
STFIX and F. M. YATX (Edttors). pp. 377450. Churchill Ltwn~stone. New 1’orL. 1986. 37. OLU~IELU. R. C. The assessment and analysis of handedness: The Edinburgh Inventory .~~~~r~r~ps~c~lf~~lo~~~~~ 9.
97 113.1971. 3X. PAILLAKU. J. and BROLCHON. M. A. A proprioceptt\e contrtbution to the spatial encoding of posttion cues for
ballistic movements. Brain Rr.5. 71. 273-284. 1974. 39. PIEHROT-DESE~LLIC;N~. CH.. GRAY. F. and BKC~ET. P. Infarcts of both Inferior parietal lobules v.ith impatrment
of vtsually guided eye movements. peripheral wsual inattention and optic atasia. Brnirr 109, XI 97. 19X6. 40. PIHOZZOLO. F J. and RAYNER. K. Handedness. hemisphertc spectalization and saccadic eye movement
latencies. .Yc,~trop.~!c/~o/oclra 18, 2’5 129. 1980. 41. POHL. W.. BUTTEKS. N. and G~OIX;LASS. H. Spatial dtscrtmmation systems and cerebral latcraltsation. Cwrc\-
8, 305 314. 1972. 42. PK~HLAL;(.. C. and JEAVUEROIJ. M. Corrective saccades: dependence on retinal reaRerent stgnals. I ‘I.WW Rc.r. 15.
465 469. 1975. 43. PKAHLA\(.. C.. PELLISON. D. and GOOLIALI-. M A. Visual control of reaching movements wthout \tsion of the
limb. E\-p. Brarn Rrs. 62. 293 302. 1986. 44. ROH~KTSHA\~. S. and SHELUO~. M. Lateralit! effects tn Judfement of the identity and position of letters: a signal
detectton analysir. Q. J. r\-p P.YJx~/I~. 28. I I5 121. 1976. 35. Roy. E. A. and ELLIOTT. D. Manual asymmetrtes In visually directed atming. c‘un. J. P.\~~chr/.40. 109 121. 19x6. 46. SAK(AT,I. H.. SHIHCTA~I. H. and KAI\ AZO. K. Spattal properttes of \tsual tixation neurons m posterior partetal
assowtton cortex of the monl\cy J .~l,lr~~)pl~~..\~~,/. 43. 1654 1672. 19X0. 47. SAVA. D.. LIOTTI. M and RIZZOLATTI. G. Right hemisphere supermrity for programming oculomotion:
wtdence from simple reaction time eiperimcnts. .\c~t~~~p.\~‘[./~r,i~,~/~[r 26. 201 21 I. 198X. 4X. SC-Hhlll)l. R. A.. ZI:LAZVIK. H.. HA~~~IU. B.. FK..\u. J. S. and Gt IY\. J. T. Motor-output \ariabihty : il theory
for the accurac\ of rapid motor acts. P,\I.~/I~I/. Rc-. 86. 41 5 -151. 1970. 4Y. Wrtrl K. R B. and D~&)bt. R. B. The metncs of hortrontal saccadtc eye movements in normal humans. I?.wvr
Kc\. 12. 467 375. 1971. 50. WHI:I I FSS. L. L.. BONITO\. R. M. and COHN \. G. H Eye-movement responses to step and pulse-step stimuli. J.
O/I/. SIU. .-lr,1. 52. 210 213. iY6h. 51. WKKHT. C. E and MIXI K. D. E. Condrtmns for a lrnear speed accuracy trade-off tn aimed movomcnts. Q. J.
I’\P. P.\l<~/I~~/. 35.A. 279 296. IYX? 52. VA\ ESSI 1. D. C.. NI.\VSOLII. W. T. and BI\H\. J. L. The pattern of Interhemispheric connections and its
relattonship to extrastrtate \IW;II arcas 111 the mac:tqu~. .I. \c,wr~\c+ 2. 265 33. 19X2.