sox2 expression associates with stem cell state in...
TRANSCRIPT

1
SOX2 expression associates with stem cell state in human ovarian carcinoma
Running title: SOX2 in ovarian carcinoma stem cells
Petra M. Bareiss*1, Anna Paczulla*1, Hui Wang1, Rebekka Schairer1, Stefan Wiehr2, Ursula
Kohlhofer3, Oliver C. Rothfuss4, Anna Fischer3, Sven Perner5, Annette Staebler3; Diethelm
Wallwiener6, Falko Fend3, Tanja Fehm6, Bernd Pichler2, Lothar Kanz1, Leticia Quintanilla-Martinez3,
Klaus Schulze-Osthoff4,7, Frank Essmann4, Claudia Lengerke1
1Department of Internal Medicine II, University of Tuebingen, Tuebingen, Germany; 2Department of Preclinical
Imaging and Radiopharmacy, Laboratory for Preclinical Imaging and Imaging Technology of the Werner Siemens-
Foundation, University of Tuebingen, Tuebingen, Germany; 3Institute of Pathology, University of Tuebingen,
Tuebingen, Germany; 4Interfaculty Institute for Biochemistry, University of Tuebingen, Tuebingen, Germany; 5Institute of Pathology, University of Bonn, Bonn, Germany; 6Women´s Hospital, University of Tuebingen,
Tuebingen, Germany, 7German Cancer Consortium (DKTK) and German Cancer Research Center, Heidelberg,
Germany * equal contribution (Petra Bareiss and Anna Paczulla have contributed equally and share first authorship)
Author contributions:
1. Conception and design: P.M.B. and C.L.
2. Development of methodology: P.M.B, A.P., H.W., S.W., U.K., O.C.R., F.E., M.K:
3. Acquisition of data: P.M.B, A.P., R.S., H.W., S.W., U.K., A.F., F.E., L.Q.M.
4. Analysis and interpretation of data: P.M.B., A.P., R.S., S.W., O.C.R., S.P., A.S., B.P., L.Q.M.,
K.S.O., F.E., C.L.
5. Writing, review and/or revision of the manuscript: P.M.B., A.P., S.W., S.P., L.Q.M., K.S.O., F.E.,
C.L.
6. Administrative, technical, or material support: D.W., F.F., T.F., B.P., L.Q.M., L.K., K.S.O., F.E., C.L.
7. Study supervision: C.L. Correspondence
Claudia Lengerke, MD
University of Tuebingen
Department of Internal Medicine II
Otfried-Mueller-Strasse 10
72076 Tuebingen
Email: [email protected]
Tel.: +49 7071 29 82899; Fax: +49 7071 29 4524
Keywords: SOX2, ovarian, cancer stem cells, apoptosis
Conflicts of interest: none
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

2
Word count: 4853
Number of figures and tables: 7
Number of references: 46
Supplemental Material: 7 Supplemental Figures, Supplemental Material & Methods
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

3
Abstract
The SRY-related HMG-box family of transcription factors member SOX2 regulates stemness and
pluripotency in embryonic stem cells and plays important roles during early embryogenesis. More
recently, SOX2 expression was documented in several tumor types including ovarian carcinoma,
suggesting an involvement of SOX2 in regulation of cancer stem cells. Intriguingly, however, studies
exploring the predictive value of SOX2 expression with respect to histopathological and clinical
parameters report contradictory results in individual tumors, indicating that SOX2 may play tumor-
specific roles. In this report, we analyze the functional relevance of SOX2 expression in human
ovarian carcinoma. We report that in human serous ovarian carcinoma (SOC) cells SOX2 expression
increases the expression of cancer stem cell markers, the potential to form tumor spheres and the in
vivo tumor-initiating capacity, while leaving cellular proliferation unaltered. Moreover, SOX2-
expressing cells display enhanced apoptosis resistance in response to conventional chemotherapies
and TRAIL. Hence, our data demonstrate that SOX2 associates with stem cell state in ovarian
carcinoma and induction of SOX2 imposes cancer stem cell properties on SOC cells. We propose the
existence of SOX2-expressing ovarian cancer stem cells as a mechanism of tumor aggressiveness
and therapy resistance in human SOC.
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

4
Introduction
Pluripotency-associated stem cell factors such as OCT4 and SOX2 regulate cellular identity in
embryonic stem cells and facilitate the reprogramming of terminally differentiated somatic cells back to
a pluripotent stem cell state (1). SOX proteins are also important regulators of early development in
different tissues, such as the foregut and lung, where for example SOX2 expression controls
bronchogenesis by inhibiting airway branching (2, 3). In adult mice, SOX2 is expressed in different
epithelial compartments marking cells with self-renewal properties (4), and targeted ablation lethally
disrupts epithelial tissue homeostasis (4). SOX2 expression is also found in neural stem cells, where it
promotes stemness by preventing default differentiation into neurons (5).
More recently, SOX2 expression has been demonstrated in several tumor types such as lung (6-10),
breast (11-14), skin (15, 16), prostate (17), ovarian (18, 19), sinonasal (20) as well as different types of
squamous carcinomas (21). However, the SOX2 expression pattern and the correlation with
histopathological status and clinical outcome are highly variable among tumors, suggesting distinct
roles of SOX2 in individual tumors. In breast carcinoma, SOX2 expression is mostly detected in a
minor subset of tumor cells and appears to be an early event in tumor development (13), indicating
potential roles in cancer stem cells (CSCs) biology and involvement in reprogramming processes
generating CSCs. In support of this notion, induction of SOX2 expression in breast carcinoma cell
lines was shown to enhance CSC properties such as tumor sphere potential and in vivo tumorigenicity
(12). Moreover, SOX2 expression was associated with positive lymph-nodal status in early-stage
breast carcinoma (13). In contrast, in human squamous cell lung cancer SOX2 protein overexpression
was associated with smaller tumor size, lower probability of metastasis and improved clinical outcome.
Other than breast carcinoma, squamous cell lung cancer samples displayed homogenous SOX2
expression, arguing against specific roles of SOX2 in lung CSCs.
Ovarian carcinoma has the seventh highest morbidity rate of cancer in women (22). Due to the lack of
early specific symptoms, ovarian carcinoma is mostly diagnosed at advanced metastatic stages that
cannot be cured by surgical resection alone. In spite of initially good response rates to platin-based
chemotherapies, relapse is a common event during the clinical course of the disease (22). An
explanation for ovarian carcinoma relapse is provided by the tumor stem cells hypothesis, proposing
that conventional chemotherapeutic approaches target the fast proliferating bulk of the ovarian cancer
cells while sparing the tumor-initiating CSCs (23). The isolation and molecular characterization of
ovarian CSCs are thus subjects of intense research. Previous studies have suggested ALDH1,
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

5
CD133, CD44, and CD117 as ovarian CSC markers, but plasticity and transition between stem and
non-stem cell states complicates their efficient isolation (24-32).
In this study, we hypothesized that SOX2 expression associates with stem cell state in ovarian
carcinoma. SOX2 expression can be detected in 15% to 60.5% of ovarian carcinomas (18, 19),
depending on the staining methodology. Supporting our hypothesis, we observed that the majority of
SOX2-positive samples displayed SOX2 expression in less than 10% of tumor cells. Moreover, SOX2
expression was enhanced by culture conditions enriching for tumor stem cells. Detailed analyses
performed on SOX2-modified human serous ovarian carcinoma (SOC) cell lines and primary cells
demonstrate that indeed SOX2 expression induces CSC properties, such as expression of stemness
markers, tumor sphere formation and in vivo tumor initiating capacity as well as apoptosis resistance,
thereby strongly promoting in vivo tumorigenicity and enabling selective resistance to conventional
anti-cancer therapies.
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

6
Materials and Methods
Cell culture, tumor spheres and cell growth assays
Ovarian cancer cell lines (Caov-3, OVCAR-3, OVCAR-5; DSMZ, Braunschweig, Germany; last
authentification on January 31st 2013 at DSMZ) and primary cells obtained through dissociation of
tissue samples derived from four patients with SOC were cultured under standard conditions. For the
tumor sphere culture assay, cells were grown in ultra-low attachment plates (Corning, Tewksbury, MA)
with sphere medium and daily added growth factors (20 ng/ml FGF, 20 ng/ml EGF; Sigma-Aldrich,
Steinheim, Germany) as described (33). Spheres were counted between day 5 and 9. To investigate
serial sphere formation, spheres were washed with PBS and dissociated to single cells by
trypsinization. To assess cell growth, 50.000 cells were plated under adherent conditions and counted
on day 3, 6 and 9. For sphere cultures performed to enrich for CSC activity, OVCAR-3 and Caov-3
cells were maintained under sphere culture conditions for 21 days and primary SOC cells for 10 days
before undergoing assessment.
Analysis of a tissue microarray (TMA) of primary human ovarian carcinomas
SOX2 and Ki67 positivity was investigated by immunohistochemistry using polyclonal goat anti-human
SOX2 (AF2018, R&D Systems, Wiesbaden, Germany) and monoclonal mouse anti-human Ki67 (clone
MiB-1, M7240, DakoCytomation, Glostrup, Denmark) on a TMA including 215 human primary ovarian
carcinomas from patients treated at the Women’s Hospital of the University of Tübingen. Detailed
information about the TMA construction and analysis are provided in the Supplemental Material. The
study was approved by the Ethics Committee of the University of Tuebingen.
Lentiviral transduction
Lentiviruses carrying SOX2 shRNA, SOX2 overexpression, corresponding empty GFP- and SOX2-
enhancer reporter constructs (12) were designed, produced and used for transduction as previously
reported (34-36). Details on lentiviral constructs and protocols are provided in the Supplemental
Material.
Flow cytometry analysis of stem cell markers, cell cycle and BrdU assays
To detect ALDH activity, the Aldefluor® assay was used according to the manufacturer’s guidelines
(Stem Cell Technologies, Grenoble, France). Cells were incubated in Aldefluor® assay buffer for 30
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

7
min. Cells from each sample additionally treated with the ALDH inhibitor diethylaminobenzaldehyde
(DEAB) served as negative controls. For flow cytometric analyses anti-CD133 (Miltenyi Biotec,
Bergisch-Gladbach, Germany), anti-TRAIL receptor 1 and 2 antibodies (BioLegend, San Diego, CA)
were used. Dead cells were detected by DAPI staining (100 ng/ml). Cell cycle analysis using
propidium iodide (PI; Sigma) staining and BrdU incorporation assays using mouse anti-BrdU V450
antibody (Roche, Mannheim, Germany) were performed as previously described (35). Flow cytometric
analyses were performed using a FACS Canto II and data analysed using the FACSDiva™ software
(BD Biosciences, Heidelberg, Germany).
Apoptosis assays
Cells seeded at 50.000 cells/cm² were incubated overnight and then treated for 24 hours with
staurosporine (2.5 µM; Sigma-Aldrich) or SuperKiller TRAIL™ (25 ng/ml; Enzo Life Sciences,
Farmingdale, NY), or for 96 hours with cisplatin (5 µM, Medac, Wedel, Germany), carboplatin (100 µM,
Medac) or paclitaxel (5 nM, Bristol-Myers Squibb, New York, USA). Cells were harvested by
trypsinization, fixed in 70% ice-cold ethanol and incubated in PBS containing 50 µg/ml PI and 100
µg/ml RNase A. Cells with subdiploid DNA content (subG1) were assessed by flow cytometry.
Caspase-3/7 activity was assessed by the Caspase-Glo® 3/7 assay (Promega, Madison, WI) and
normalized to protein content following treatment with staurosporine (4 hours) or TRAIL (6 hours).
Immunoblot and immunocytochemistry analyses
Immunoblot and immunocytochemistry analyses were performed using mouse anti-actin (Li-COR
Biosciences, Lincoln, NE), rabbit anti-caspase-3 (Cell Signaling, Danvers, MA), rabbit anti-cleaved
caspase-3 (Asp175) (Cell Signaling), and rabbit anti-SOX2 (D6D9) XP (Cell Signaling) antibodies.
Detection was performed using IRDye® 800CW-conjugated goat anti-rabbit IgG or IRDye® 680 anti-
mouse IgG antibodies (Li-COR Biosciences) and an Odyssey Imager (Li-COR Biosciences). For
detection of the SOX2 knockdown HRP-linked anti-rabbit IgG antibody (Cell Signaling) and ECLTM
Prime Western Blotting Detection Reagent (GE Healthcare, Buckinghamshire, UK) were used.
Gene expression analyses
RNA isolation, cDNA preparation and real-time gene expression analyses were performed as
described (13, 37) using a LightCycler® 480 instrument and LightCycler® probes master mix (for
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

8
SOX2, ALDH1, LIN28, NANOG, OCT4, GAPDH; Roche) or SYBR Green assay (for BBC3, PMAIP1,
BCL2; Eurogentec, Cologne, Germany). Primers and probes are listed in the Supplemental Material.
Relative expression levels were calculated after normalization to the reference gene GAPDH (Probe)
or PBGD (SYBR Green) by using the ΔΔCT method.
Xenotransplantation model
NOD.Cg-Prkdcscid IL2rgtmWjl/Sz (also termed NOD/SCID/IL2Rγnull, abbreviated as NSG) mice (38) were
purchased from Jackson Laboratory (Bar Harbor, ME, USA) and maintained under pathogen-free
conditions. Control and SOX2-overexpressing Caov-3 cells mixed with Matrigel (1:2, BD Biosciences)
were implanted subcutaneously in individual flanks of the same mouse. Tumor growth was monitored
by palpation of the injection site and PET/MR analysis performed using i.v. administered 11-15 MBq of
[18F]FDG as described (39, 40) and summarized in the Supplemental Material. Mice were euthanized
7 to 15 weeks after implantation. For histological analysis, mouse tissues were fixed in 4%
formaldehyde, paraffin-embedded cut in 3-5 µm-sections and stained with H&E. Immunohistochemical
analysis was performed as described (41) on an automated immunostainer (Ventana Medical
Systems, Frankfurt, Germany) according to the company’s protocol for open procedures with slight
modifications. The antibody panel used included SOX2 (SP76; Cell Marque, Rocklin, CA), cleaved
caspase-3 (Asp 175, Cell Signaling), Ki67 (SP6, DCS Innovative Diagnostik Systeme, Hamburg,
Germany), and EpCAM (BerEp4, Dako, Hamburg, Germany).
Statistical analyses
For all experiments, mean values are presented and error bars represent the standard error if not
otherwise indicated. P-values are derived via the application of a 2-tailed, unpaired Student t-test.
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

9
Results
SOX2 expression is enhanced in CSC-enriched SOC cell cultures
SOX2 mRNA expression was investigated by real-time PCR in SOC patient samples and cell lines
including the lines Caov-3 and OVCAR-3 harboring amplifications on the chromosome 3q (Fig. 1A).
Heterogeneous expression of SOX2 was noted (Fig. 1A and Suppl. Fig. 1) mirroring the results
documented in a TMA of human SOC samples (18, 19). Previous data on ovarian and breast cancer
cells reported that sphere cultures increase CSC frequency as compared to two-dimensional (2D)
adherent cultures (27, 33). Indeed, ovarian cancer cell lines grown as spheres for 21 days showed a
higher frequency of ALDHhighCD133+ putative CSCs (OVCAR-3 cells, Fig. 1B) and enhanced
expression of putative stem cell markers in comparison to 2D-cultures (Caov-3 cells, Fig. 1C).
Consistent with a role of SOX2 as a stem cell marker in SOC, SOX2 expression was also enhanced in
sphere cultures of Caov-3 as well as primary ovarian carcinoma cells (Fig. 1C).
SOX2 modulates CSC properties in human SOC cells
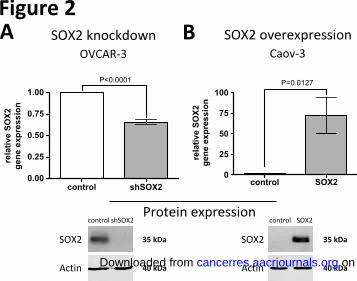
To explore the functional role of SOX2 in ovarian carcinoma, we stably suppressed SOX2 expression
in OVCAR-3, the SOC line with the highest basal SOX2 expression, using two different lentiviruses
containing SOX2-inhibitory shRNAs (Fig. 2A and Suppl. Fig. 2A). Furthermore, Caov-3 cells displaying
low basal SOX2 expression as well as primary SOC cells derived from patients were treated with
SOX2 lentiviruses to study the effects of SOX2 overexpression (Fig. 2A and Suppl. Fig. 3A). Cells
transduced with empty GFP-lentiviruses were used as controls.
Induction of SOX2 expression in both Caov-3 and primary SOC cells was able to enhance the
expression of other putative stem cell markers (LIN28, NANOG, OCT4 and ALDH1; Fig. 3A,B),
suggesting that activation of SOX2 expression is sufficient to facilitate the transition to a stem cell like-
state. Consistent with this notion, enhanced tumor spheres formation was observed in SOX2-
overexpressing cells, while SOX2 knockdown induced the opposite effect (Fig. 4A). Notably, the effect
on tumor sphere formation was also documented in primary SOC cells and was even more
pronounced upon serial replating (Fig. 4B). Single cell tumor sphere assays further confirmed the
higher frequency of sphere-initiating cells in SOX2-expressing versus control cells (Fig. 4C).
To further explore the role of SOX2 as a CSC marker in human SOC, we treated OVCAR-3 cells with
a lentiviral SOX2-reporter construct, previously described to recognize cells with high SOX2 promoter
activity (SOX2+) in breast carcinoma and neural stem cells (12, 42) (Suppl. Fig. 4A). Puromycin
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

10
selection was applied to select for efficiently transduced cells and SOX2+ cells visualized by
fluorescence. Supporting our previous data, SOX2+ cells were enriched in OVCAR-3 cells cultured as
spheres, as compared to 2D cultures (Suppl. Fig. 4B). In primary sphere assays, nearly every SOX2+
cell isolated by FACS gave rise to an individual tumor sphere (SOX2+ sphere), and SOX2+ spheres
were larger than those derived from reporter-negative cells (SOX2- spheres) (Fig. 4D and Suppl. Fig.
4C). Importantly, flow cytometry of SOX2+ primary spheres revealed a mixture of SOX2+ and SOX2-
cells, suggesting that SOX2+ cells undergo both self-renewal and differentiation processes giving rise
to both populations of cells. In contrast, primary SOX2- spheres remained SOX2-negative (Suppl. Fig.
4C). Consistent with these data, cells derived from SOX2- spheres exhausted their sphere generation
potential upon serial replating, while cells derived from SOX2+ spheres maintained sphere formation
(Fig. 4D).
Together, these data indicate that cells with self-renewal capacity segregate to the SOX2+
compartment and suggest that SOX2 induction can activate CSC molecular pathways and functional
properties in human SOC cells.
Induction of SOX2 strongly enhances in vivo tumorigenicity in a NSG mouse model
To explore the relevance of SOX2 expression in SOC cells in vivo, we performed xenotransplantation
experiments of SOX2-overexpressing and control Caov-3 cells in immunopermissive NOD.Cg-
Prkdcscid IL2rgtmWjl/Sz (NSG) mice (38). The same numbers of SOX2-overexpressing and control Caov-
3 cells were implanted subcutaneously in the flanks of 8 weeks old female mice as indicated, and
tumor induction was monitored every second week by palpation of the injection sites. To avoid bias
through different animal hosts, SOX2-overexpressing and control cells were injected in the right and
left flank of the same mouse. When 500.000 cells were injected per flank, control Caov-3 cells
generated tumors at 4 weeks post-injection in 1 out of 4 animals (Fig. 5A). Lowering the number of
transplanted cells to 100.000 or 50.000 cells per animal delayed tumor formation from control Caov-3
cells (Fig. 5A). In contrast, SOX2-overexpressing cells robustly induced tumors in all transplanted
animals and accelerated the appearance of palpable tumor masses (Fig. 5A).
To further consolidate these observations, we performed in vivo PET/MRI analyses and ex vivo
immunohistological analyses at the end of the experiment. At week 15 post-inoculation (p.i.) of
100.000 cells, tumors were detected on both sites in all mice by the sensitive PET/MRI method (Fig.
5D). However, quantitative image analysis of tumor volumes revealed that SOX2-overexpressing cells
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

11
induced much larger tumors than control cells (118.2±19.0 mm3 versus 40.4 mm3 at 15 weeks
following injection of 100.000 cells, and respectively 618.6±244.6 mm3 versus 50.98±21.8 mm3 when
500.000 cells were injected, Fig. 5B), which was also confirmed by immunohistological analysis (Fig.
5C). Interestingly, PET-quantification of [18F]FDG uptake (39) revealed similar metabolic activity in
SOX2-overexpressing and control tumors at both measured time-points (8 and 15 weeks post
injection, p.i., Fig. 5B), indicating that the inductive effects of SOX2 on tumor formation were not
mediated by modulation of metabolic activity. However, due to their larger mass, tumors derived from
SOX2-overexpressing cells partially displayed necrotic areas revealing a heterogeneous uptake of
[18F]FDG at the measured time points.
Interestingly, the pronounced difference of in vivo tumorigenicity between SOX2-overexpressing and
control cells was not due to enhanced cellular proliferation, as revealed by the similar results of the
Ki67 staining performed on explanted tumors (Fig. 5C). SOX2-overexpressing tumors displayed more
necrotic areas and a higher apoptotic activity, as shown by the active caspase-3 staining (Fig. 5C).
These findings are in line with the in vivo PET/MR imaging results where necrotic areas were detected
in all SOX2-overexpressing tumors (Fig. 5B).
Overall, these data strongly suggest that SOX2 mediates tumorigenicity in SOC cells by facilitating
transition to a CSC state with enhanced tumor-initiating properties. To further explore this hypothesis,
we performed a limiting dilution in vivo transplantation assay: 10.000, 1.000, 100 and 10 SOX2-
overexpressing and control Caov-3 cells were transplanted as described above in the contralateral
flanks of n=5 mice per group. In contrast to the results observed with higher numbers of cells (Fig. 5 A
and C), no palpable tumors were documented at seven weeks post-transplantation. However,
immunohistological analysis of the injection sites revealed microscopic human tumor cell clusters in
animals injected with 10.000 or 1.000 cells, but not 100 or 10 cells (H&E staining, Fig. 5E). Staining
with antibodies against human EpCAM (Suppl. Fig. 5) and CA125 (not shown) confirmed correct
detection of human SOC cells. Notably, microscopical tumors were detected more frequently from
SOX2-overexpressing as compared to control cells, which was especially evident with the lowest
number of injected cells (Fig. 5E and F).
SOX2 expression does not affect cell proliferation but enhances the apoptosis resistance of
SOC cells
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

12
Modulation of SOX2 expression did not alter cell cycle progression, BrdU incorporation (Fig. 6A) or
cell growth of in vitro 2D-cultured OVCAR-3 and Caov-3 cells (Fig. 6B). To further explore whether
SOX2 regulates SOC cell proliferation, we analyzed SOX2 expression and Ki67 staining on a tissue
microarray (TMA) of 215 human primary ovarian carcinomas comprising 143 high-grade SOC (see
Supplemental Material for details on TMA construction). In 136 out of 143 high-grade SOC samples,
both SOX2 and Ki67 staining was available. Although SOX2 expression was found in 64.6% of high-
grade SOC samples (Fig. 6C, Suppl. Fig 1 and data not shown), Ki67 positivity was not dependent on
the SOX2 expression level, which is in contrast to findings in other tumor entities (11, 17).
As mentioned above, the histologic analysis of the murine tumors revealed higher levels of active
caspase-3 in SOX2-overexpressing tumors (Fig. 5C). However, enhanced caspase-3 activation is
most likely a secondary effect in response to restrictive in vivo environmental factors (e.g. insufficient
blood supply due to disproportionate tumor growth overriding tumor´s capacity of vessel recruitment),
since in vitro 2D-culture experiments showed reduced levels of spontaneous apoptosis in SOX2-
overexpressing Caov-3 as well as OVCAR-3 cells (Fig. 6D). To test whether SOX2 expression
modulates apoptosis, we incubated the cells with staurosporine and the death ligand TRAIL to activate
the intrinsic and extrinsic pathways of apoptosis. Flow cytometric quantitation of subG1 cell
populations revealed enhanced apoptosis in response to staurosporine and TRAIL in the SOX2
knockdown cells, whereas SOX2 overexpression conferred enhanced resistance (Fig. 6A-B, Suppl.
Fig. 2 and 6). We also assayed activity of caspase-3/7 in substrate cleavage assays (Fig. 7A-B) and
processing of caspase-3 by immunoblot analysis for cleaved caspase-3 (Suppl. Fig. 7), confirming the
resistance-mediating potential of SOX2 expression. Importantly, SOX2 expression in Caov-3 cells also
mediated resistance to carboplatin, cisplatin and paclitaxel (Fig. 7C, Suppl. Fig. 7), indicating SOX2
expression as a molecular driver of chemotherapy resistance in ovarian carcinoma. Notably, the
enhanced sensitivity of OVCAR-3 cells due to SOX2 knockdown was reverted by lentiviral re-
expression of ectopic SOX2 (Fig. 7D). These data demonstrate that the observed phenotype
specifically depends on SOX2 expression levels. Analogue experiments performed in a third cell line
(OVCAR-5) and using an alternative SOX2 shRNA sequence furthermore confirmed these results
(Suppl. Fig. 2 and data not shown).
In an attempt to elucidate the molecular basis for SOX2-induced resistance to TRAIL-mediated
apoptosis, we initially analyzed surface expression of the TRAIL receptors 1 and 2 by flow cytometry.
However, no significant difference in the expression level of TRAIL-R1 or TRAIL-R2 was detected in
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

13
OVCAR-3 and Caov-3 cells in response to SOX2 knockdown and overexpression, respectively (Fig.
5E). Therefore, SOX2 expression modulates apoptosis sensitivity downstream of these death
receptors, as usually seen in so-called type II cells that depend on amplification of death receptor
signaling via the intrinsic apoptotic pathway. As the intrinsic pathway is controlled by the BCL2 protein
family, we analyzed the expression of pro-apoptotic (PUMA/BBC3 and NOXA/PMAIP1) and anti-
apoptotic genes (BCL2) by real-time PCR. In line with the observed apoptosis resistance,
overexpression of SOX2 induced enhanced expression of anti-apoptotic BCL2 while reducing
expression of the pro-apoptotic proteins PUMA/BBC3 and NOXA/PMAIP1 (Fig. 7F).
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

14
Discussion
SOX2 is a key regulator for maintaining the pluripotency and self-renewal of embryonic stem cells and
contributes to the reprogramming of differentiated somatic cells back to a pluripotent stem cell state.
More recently, enhanced SOX2 expression has been detected in several epithelial tumors (6, 7, 9-19)
suggesting that SOX2 also regulates tumorigenesis. Based on its prominent role in pluripotent stem
cell stemness, SOX2 expression has been proposed as a general feature of CSCs (12, 27, 29, 43).
Emerging data, however, demonstrate divergent SOX2 expression patterns and functions across
tumors, suggesting that SOX2 adopts specific roles in individual tumor types. In breast cancer cells,
for instance, SOX2 was shown to promote CSC characteristics such as in vitro tumor sphere formation
and in vivo tumorigenicity (12). When cultured under non-adherent sphere conditions that enrich for
CSCs, breast cancer cells up-regulated SOX2 expression, indicating a tight link between SOX2
expression and functional stem cell state. Furthermore, immunohistochemical analysis of primary
breast carcinomas revealed a heterogeneous SOX2 expression in only a minority of tumor cells (13),
consistent with the putative role of SOX2 as a breast CSC marker. In contrast, squamous cell lung
cancers (9) mostly display homogenous distribution of SOX2 among tumor cells, suggesting that in
this tumor entity SOX2 might also influence non-CSCs. The difference in upstream regulatory
mechanisms reported for SOX2 in individual tumor types further support this hypothesis. In squamous
cell lung cancers SOX2 overexpression is mostly linked to SOX2 gene amplification on the
chromosome 3q26 (9). This in line with the observed homogenous SOX2 expression in all tumor cells,
since genetic amplification events likely persist upon CSC differentiation. In contrast, in breast
carcinomas elevated SOX2 expression has been largely detected in the absence of chromosomal
amplifications and relies on yet unknown upstream regulatory mechanisms (13). Since epigenetic
mechanisms essentially participate in stem cell reprogramming, it is possible that SOX2 expression in
breast CSCs is triggered by epigenetic events, such as altered SOX2 promoter methylation as
previously reported in glioblastoma (44).
In serous ovarian carcinoma, high SOX2 protein expression is associated with histopathologically and
clinically aggressive disease (18, 19). Similar as in breast carcinoma (13), we found that SOX2-
positive ovarian carcinomas display a heterogeneous expression pattern with mostly less than 10% of
SOX2-positive tumor cells, indicating that SOX2 might preferentially regulate the ovarian CSC
compartment. Indeed, SOC sphere cultures enriched for putative ovarian CSCs induced increased
SOX2 enhancer activity and SOX2 expression as compared to 2D-cultures. Consistently, SOX2+ cells
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

15
enriched by detection of the SOX2 enhancer reporter generated tumor spheres from nearly every cell
and demonstrated self-renewal and differentiation properties in serial replating assays. SOX2- cells, in
contrast, gave rise to significantly less primary spheres, and most importantly, could not preserve
sphere initiation properties beyond tertiary spheres.
To explore whether SOX2 expression is sufficient to mediate stemness in ovarian carcinoma cells, we
modulated SOX2 expression in human SOC cell lines and primary cells by lentiviral SOX2 knockdown
and overexpression. Ectopic SOX2 expression enhanced the in vitro tumor sphere potential and
expression of stemness genes such as OCT4, LIN28, NANOG and ALDH1, whereas the SOX2
knockdown showed opposite effects. However, although the frequency of sphere-initiating cells was
greatly enhanced by SOX2 overexpression in Caov3 and patient-derived SOC cells, primary tumor
spheres were initiated by only a fraction of SOX2-overexpressing cells. In contrast, SOX2+ cells
isolated via positivity for the SOX2 enhancer reporter generated spheres from nearly every cell. A
possible explanation for this finding is that, even though SOX2 can facilitate transition to a stem cell
state, this transition occurs only in a subset of tumor cells. Alternatively, particularly high SOX2
expression levels, as detected by the SOX2 enhancer reporter, are needed to accomplish the
transition to a CSC state, which might not be uniformly induced in all cells by lentiviral SOX2
expression.
Upon xenotransplantation in NSG mice, SOX2-overexpressing cells induced tumors earlier and more
frequent than control SOC cells. In vivo PET/MRI analyses as well as histological analyses of
xenotransplanted mice confirmed larger tumor volumes from SOX2-overexpressing than control SOC
cells. Since data in prostate as well as breast cancer suggested that SOX2 promotes tumorigenesis by
inducing cell proliferation (11, 17), we tested whether cell growth and proliferation were affected by
SOX2. Surprisingly, mouse tumors derived from SOX2-overexpressing and control cells showed
similar Ki67 staining. In addition, no differences in cell cycle distribution, BrdU incorporation or cell
growth were observed in 2D cultures of SOX2-modified and control cells. Tumors generated from
SOX2-expressing and control cells showed also similar metabolic activity in the PET assay. These
results were further corroborated by tissue microarray analyses of primary ovarian carcinomas, which
revealed no correlation between the SOX2 expression level and Ki67. Furthermore, limiting dilution
experiments suggest that SOX2 overexpression enhanced the frequency of tumor-initiating cells, since
increased tumor cell clusters could microscopically be detected in animals transplanted with low
numbers of SOX2-overexpressing as compared to control cells. Thus, our data suggest that the
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

16
enhanced tumorigenicity of SOX2-overexpressing ovarian carcinoma cells does not rely on enhanced
cell proliferation, but is rather due to the induction of a CSC state.
Another feature regulating tumor formation is the apoptosis sensitivity of tumor cells. CSCs are
assumed to possess enhanced apoptosis resistance, facilitating tumor generation and escape from
conventional chemotherapies. Intriguingly, SOX2-expressing tumors from transplanted Caov-3 cancer
cells showed enhanced caspase-3 activity as compared to tumors from control cells. Nevertheless,
analyses of SOX2-modified Caov-3, OVCAR-3 and OVCAR-5 cells consistently demonstrated that
higher SOX2 levels prevented apoptosis in response to both intrinsic (e.g. staurosporine,
chemotherapies) and extrinsic (e.g. TRAIL) stimuli, indicating that SOX2 confers apoptosis resistance,
a property classically attributed to CSCs. The obviously discrepant results between caspase-3
activation observed in vitro and in vivo might be explained by several reasons, including a potentially
increased hypoxia of the larger SOX2-expressing tumors, which might result in secondary necrosis.
In ovarian cancer cells SOX2 appears to regulate stemness, tumor-initiating capacity and apoptosis
resistance, which are main features characterizing CSCs, while not modulating proliferation. The
molecular mechanisms of SOX2-mediated stemness remain largely unexplored. In this study, we
observed robust induction of OCT4, LIN28, NANOG and partly ALDH1 upon SOX2 activation. This
could be a direct SOX2-induced transcriptional effect or mediated by the fact that SOX2 activation
induces a stem cell state characterized by expression of these markers. In embryonic stem cells,
SOX2 interacts with the pluripotency proteins OCT4, NANOG and Lin28. In line, suppression of OCT4
and Lin28 by RNA interference was recently shown to inhibit ovarian cancer cell growth and survival
(29). To elucidate the pathways underlying SOX2-mediated apoptosis resistance, we first studied the
surface expression of TRAIL receptors, which was not affected by SOX2 expression and, hence,
indicated an involvement of receptor downstream events. Indeed, expression analysis of apoptosis-
regulatory genes revealed that SOX2 modulated the expression of certain BCL2 members. In SOX2-
overexpressing Caov-3 cells, expression of the anti-apoptotic gene BCL2 was enhanced, while the
expression of the pro-apoptotic genes PUMA/BBC3 and NOXA/PMAIP1 was reduced. These data
indicate that SOX2 modulates the balance of central apoptosis regulators, thereby changing apoptosis
sensitivity. New therapeutic approaches that target BCL2 proteins to enhance apoptosis may therefore
be a valuable tool for targeting SOX2-positive putative ovarian CSCs. Further studies are needed to
explore in detail the mechanisms of apoptosis protection governed by SOX2 and to investigate
whether BCL2, PUMA/BBC3 and NOXA/PMAIP1 or related genes are direct transcriptional targets of
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

17
SOX2 in ovarian carcinoma. In support of this assumption is the recent identification of SOX2-binding
regions in the BCL2 and NOXA/PMAIP1 genes using a ChiP-Seq approach of glioblastoma cells (45).
In summary, our data in ovarian carcinoma cell lines and patient-derived tumor samples suggest that
in this tumor entity SOX2 expression is a CSC marker and can induce cancer stem cell properties
such as stemness, tumor-initiating capacity and apoptosis resistance. SOX2 expression in putative
ovarian CSCs enables their selective survival to conventional chemotherapies and promotes their in
vivo tumorigenicity. We propose that SOX2-expressing CSCs contribute to therapy resistance and
disease relapse in ovarian carcinoma patients and that targeting SOX2 will improve clinical treatment
of ovarian carcinoma by enhancing apoptotic responses to conventional chemotherapies and
exhausting the cancer stem cell fraction.
Acknowledgments This study was supported by the Deutsche Forschungsgemeinschaft (SFB773) and the Baden-
Württemberg-Stiftung (“Adult Stem Cells II” Program). We thank Jana Ihring, Caroline Herrmann and
Sabrina Grimm (University of Tuebingen, Department of Internal Medicine II) for help with apoptosis
assays and FACS analysis, Maren Koenig (University of Tübingen, Department of Preclinical Imaging
and Radiopharmacy, Laboratory for Preclinical Imaging and Imaging Technology of the Werner
Siemens-Foundation), for excellent technical PET/MR imaging support, Claudia Kloss and Dennis
Thiele (University of Tübingen, Institute of Pathology) for help with murine histopathological analyses,
Holm Zaehres (Max-Planck Institute, Münster, Germany) for providing the human SOX2 cDNA, Olga
Kustikova, Axel Schmabach and Christopher Baum from the Hannover Medical School (Hannover,
Germany) for help with lentiviral constructs.
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

18
References
1. Park IH, Zhao R, West JA, Yabuuchi A, Huo H, Ince TA, et al. Reprogramming of human somatic cells to pluripotency with defined factors. Nature. 2008;451:141-6. 2. Dong C, Wilhelm D, Koopman P. Sox genes and cancer. Cytogenet Genome Res. 2004;105:442-7. 3. Gontan C, de Munck A, Vermeij M, Grosveld F, Tibboel D, Rottier R. Sox2 is important for two crucial processes in lung development: branching morphogenesis and epithelial cell differentiation. Dev Biol. 2008;317:296-309. 4. Arnold K, Sarkar A, Yram MA, Polo JM, Bronson R, Sengupta S, et al. Sox2(+) adult stem and progenitor cells are important for tissue regeneration and survival of mice. Cell Stem Cell. 2011;9:317-29. 5. Graham V, Khudyakov J, Ellis P, Pevny L. SOX2 functions to maintain neural progenitor identity. Neuron. 2003;39:749-65. 6. Bass AJ, Watanabe H, Mermel CH, Yu S, Perner S, Verhaak RG, et al. SOX2 is an amplified lineage-survival oncogene in lung and esophageal squamous cell carcinomas. Nat Genet. 2009;41:1238-42. 7. Hussenet T, Dali S, Exinger J, Monga B, Jost B, Dembele D, et al. SOX2 is an oncogene activated by recurrent 3q26.3 amplifications in human lung squamous cell carcinomas. PLoS One. 2010;5:e8960. 8. Rudin CM, Durinck S, Stawiski EW, Poirier JT, Modrusan Z, Shames DS, et al. Comprehensive genomic analysis identifies SOX2 as a frequently amplified gene in small-cell lung cancer. Nat Genet. 2012;44:1111-6. 9. Wilbertz T, Wagner P, Petersen K, Stiedl AC, Scheble VJ, Maier S, et al. SOX2 gene amplification and protein overexpression are associated with better outcome in squamous cell lung cancer. Mod Pathol. 2011;24:944-53. 10. Xiang R, Liao D, Cheng T, Zhou H, Shi Q, Chuang TS, et al. Downregulation of transcription factor SOX2 in cancer stem cells suppresses growth and metastasis of lung cancer. Br J Cancer. 2011;104:1410-7. 11. Chen Y, Shi L, Zhang L, Li R, Liang J, Yu W, et al. The molecular mechanism governing the oncogenic potential of SOX2 in breast cancer. J Biol Chem. 2008;283:17969-78. 12. Leis O, Eguiara A, Lopez-Arribillaga E, Alberdi MJ, Hernandez-Garcia S, Elorriaga K, et al. Sox2 expression in breast tumours and activation in breast cancer stem cells. Oncogene. 2012;31:1354-65. 13. Lengerke C, Fehm T, Kurth R, Neubauer H, Scheble V, Muller F, et al. Expression of the embryonic stem cell marker SOX2 in early-stage breast carcinoma. BMC Cancer. 2011;11:42. 14. Rodriguez-Pinilla SM, Sarrio D, Moreno-Bueno G, Rodriguez-Gil Y, Martinez MA, Hernandez L, et al. Sox2: a possible driver of the basal-like phenotype in sporadic breast cancer. Mod Pathol. 2007;20:474-81. 15. Girouard SD, Laga AC, Mihm MC, Scolyer RA, Thompson JF, Zhan Q, et al. SOX2 contributes to melanoma cell invasion. Lab Invest. 2012;92:362-70. 16. Laga AC, Zhan Q, Weishaupt C, Ma J, Frank MH, Murphy GF. SOX2 and nestin expression in human melanoma: an immunohistochemical and experimental study. Exp Dermatol. 2011;20:339-45. 17. Lin F, Lin P, Zhao D, Chen Y, Xiao L, Qin W, et al. Sox2 targets cyclinE, p27 and survivin to regulate androgen-independent human prostate cancer cell proliferation and apoptosis. Cell Prolif. 2012;45:207-16. 18. Ye F, Li Y, Hu Y, Zhou C, Chen H. Expression of Sox2 in human ovarian epithelial carcinoma. J Cancer Res Clin Oncol. 2011;137:131-7. 19. Zhang J, Chang DY, Mercado-Uribe I, Liu J. Sex-determining region Y-box 2 expression predicts poor prognosis in human ovarian carcinoma. Hum Pathol. 2012;43:1405-12. 20. Schrock A, Goke F, Wagner P, Bode M, Franzen A, Braun M, et al. Sex Determining Region Y-Box 2 (SOX2) Amplification Is an Independent Indicator of Disease Recurrence in Sinonasal Cancer. PLoS One. 2013;8:e59201. 21. Maier S, Wilbertz T, Braun M, Scheble V, Reischl M, Mikut R, et al. SOX2 amplification is a common event in squamous cell carcinomas of different organ sites. Hum Pathol. 2011;42:1078-88. 22. van der Burg ME. Advanced ovarian cancer. Curr Treat Options Oncol. 2001;2:109-18. 23. Steg AD, Bevis KS, Katre AA, Ziebarth A, Dobbin ZC, Alvarez RD, et al. Stem cell pathways contribute to clinical chemoresistance in ovarian cancer. Clin Cancer Res. 2012;18:869-81. 24. Baba T, Convery PA, Matsumura N, Whitaker RS, Kondoh E, Perry T, et al. Epigenetic regulation of CD133 and tumorigenicity of CD133+ ovarian cancer cells. Oncogene. 2009;28:209-18.
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

19
25. Curley MD, Therrien VA, Cummings CL, Sergent PA, Koulouris CR, Friel AM, et al. CD133 expression defines a tumor initiating cell population in primary human ovarian cancer. Stem Cells. 2009;27:2875-83. 26. Keysar SB, Jimeno A. More than markers: biological significance of cancer stem cell-defining molecules. Mol Cancer Ther. 2010;9:2450-7. 27. Kryczek I, Liu S, Roh M, Vatan L, Szeliga W, Wei S, et al. Expression of aldehyde dehydrogenase and CD133 defines ovarian cancer stem cells. Int J Cancer. 2012;130:29-39. 28. Ma I, Allan AL. The role of human aldehyde dehydrogenase in normal and cancer stem cells. Stem Cell Rev. 2011;7:292-306. 29. Peng S, Maihle NJ, Huang Y. Pluripotency factors Lin28 and Oct4 identify a sub-population of stem cell-like cells in ovarian cancer. Oncogene. 2010;29:2153-9. 30. Silva IA, Bai S, McLean K, Yang K, Griffith K, Thomas D, et al. Aldehyde dehydrogenase in combination with CD133 defines angiogenic ovarian cancer stem cells that portend poor patient survival. Cancer Res. 2011;71:3991-4001. 31. Wu Y, Wu PY. CD133 as a marker for cancer stem cells: progresses and concerns. Stem Cells Dev. 2009;18:1127-34. 32. Zhang S, Balch C, Chan MW, Lai HC, Matei D, Schilder JM, et al. Identification and characterization of ovarian cancer-initiating cells from primary human tumors. Cancer Res. 2008;68:4311-20. 33. Dontu G, Abdallah WM, Foley JM, Jackson KW, Clarke MF, Kawamura MJ, et al. In vitro propagation and transcriptional profiling of human mammary stem/progenitor cells. Genes Dev. 2003;17:1253-70. 34. Anastasov N, Klier M, Koch I, Angermeier D, Hofler H, Fend F, et al. Efficient shRNA delivery into B and T lymphoma cells using lentiviral vector-mediated transfer. J Hematop. 2009;2:9-19. 35. Konantz M, Andre MC, Ebinger M, Grauer M, Wang H, Grzywna S, et al. EVI-1 modulates leukemogenic potential and apoptosis sensitivity in human acute lymphoblastic leukemia. Leukemia. 2012. 36. Maetzig T, Brugman MH, Bartels S, Heinz N, Kustikova OS, Modlich U, et al. Polyclonal fluctuation of lentiviral vector-transduced and expanded murine hematopoietic stem cells. Blood. 2011;117:3053-64. 37. Riedt T, Ebinger M, Salih HR, Tomiuk J, Handgretinger R, Kanz L, et al. Aberrant expression of the homeobox gene CDX2 in pediatric acute lymphoblastic leukemia. Blood. 2009;113:4049-51. 38. Shultz LD, Lyons BL, Burzenski LM, Gott B, Chen X, Chaleff S, et al. Human lymphoid and myeloid cell development in NOD/LtSz-scid IL2R gamma null mice engrafted with mobilized human hemopoietic stem cells. J Immunol. 2005;174:6477-89. 39. Judenhofer MS, Wehrl HF, Newport DF, Catana C, Siegel SB, Becker M, et al. Simultaneous PET-MRI: a new approach for functional and morphological imaging. Nat Med. 2008;14:459-65. 40. Wiehr S, von Ahsen O, Rose L, Mueller A, Mannheim JG, Honndorf V, et al. Preclinical Evaluation of a Novel c-Met Inhibitor in a Gastric Cancer Xenograft Model Using Small Animal PET. Mol Imaging Biol. 2012. 41. Kunder S, Calzada-Wack J, Holzlwimmer G, Muller J, Kloss C, Howat W, et al. A comprehensive antibody panel for immunohistochemical analysis of formalin-fixed, paraffin-embedded hematopoietic neoplasms of mice: analysis of mouse specific and human antibodies cross-reactive with murine tissue. Toxicol Pathol. 2007;35:366-75. 42. Kondo T, Raff M. Chromatin remodeling and histone modification in the conversion of oligodendrocyte precursors to neural stem cells. Genes Dev. 2004;18:2963-72. 43. Wong DJ, Liu H, Ridky TW, Cassarino D, Segal E, Chang HY. Module map of stem cell genes guides creation of epithelial cancer stem cells. Cell Stem Cell. 2008;2:333-44. 44. Alonso MM, Diez-Valle R, Manterola L, Rubio A, Liu D, Cortes-Santiago N, et al. Genetic and epigenetic modifications of Sox2 contribute to the invasive phenotype of malignant gliomas. PLoS One. 2011;6:e26740. 45. Fang X, Yoon JG, Li L, Yu W, Shao J, Hua D, et al. The SOX2 response program in glioblastoma multiforme: an integrated ChIP-seq, expression microarray, and microRNA analysis. BMC Genomics. 2011;12:11. 46. Scheble VJ, Braun M, Beroukhim R, Mermel CH, Ruiz C, Wilbertz T, et al. ERG rearrangement is specific to prostate cancer and does not occur in any other common tumor. Mod Pathol. 2010;23:1061-7.
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

20
Figure legends Figure 1: Enhancement of SOX2 expression in CSC-enriched spheres cultures. (A) SOX2 shows
heterogeneous expression in ovarian cancer cell lines and patient samples. (B) OVCAR-3 cells
cultured as spheres for 21 days are enriched for ALDHhighCD133+ putative ovarian cancer stem cells
as measured by flow cytometry. (C) Enhanced expression of SOX2 and other putative stem cell genes
Caov-3 cells cultured for 21 days and patient samples cultured for 10 days under sphere conditions,
as compared to corresponding 2D-cultured cells. Relative gene expression was analyzed by real-time
PCR after normalization to GAPDH. Shown are the fold changes in relative gene expression of cells
cultured in spheres (grey box) relative to cells cultured in 2D (dashed line). Data represent the mean
values ± SEM from three or more independent biological experiments performed in triplicates for the
Caov-3 cell line, and respectively from technical triplicates in the patient samples (* p<0.05, ** p<0.01,
*** p<0.005, n.s. not significant)
Figure 2: Modulation of SOX2 expression. Modulation of SOX2 mRNA and protein expression in
ovarian cell lines after lentiviral SOX2 knockdown (A) or SOX2 overexpression (B) in comparison to
control lentiviruses. The upper panels show relative gene expression levels normalized to GAPDH, as
measured by real-time PCR analysis of three representative independent biological experiments each
performed in triplicates. The lower panels show immunoblot analyses representative for three
independent biological replicates.
Figure 3: SOX2 expression enhances expression of putative stem cell genes. Caov-3 (A) and
primary SOC patient-derived cells (B) were analyzed by real-time PCR and normalized for GAPDH.
Shown are the fold changes in relative gene expression in cells cultured in spheres (grey box) versus
cells cultured in 2D (dashed line). Shown are data from one representative out of three independent
biological experiments performed in triplicates for Caov-3 cells and from technical triplicates in primary
cells (* p<0.05, ** p<0.01, *** p<0.005, n.s. not significant).
Figure 4: SOX2 increases the sphere-forming potential of ovarian cell lines and primary tumor
cells. (A) SOX2-modified and control Caov-3 and OVCAR-3 cells (plated 1250 cells/well) were scored
for primary sphere formation after 9 and 5 days, respectively. Shown are data from three independent
biological experiments performed in triplicates. (B) Primary SOC cells were transduced with
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

21
lentiviruses for GFP-tagged SOX2 or a GFP control. Cells were then directly plated at a density of 625
cells/well without prior FACS. After 5 days sphere formation of transduced (GFP+) and non-
transduced (GFP-) cells was microscopically scored. Primary spheres were subsequently dissociated
to single cells and used for the replating experiments to investigate secondary sphere formation.
Shown are data from technical triplicates. (C) Single SOX2-overexpressing and control Caov-3 cells
were assessed for their sphere-forming potential in 96-well plates. Shown are data from three
independent biological experiments. Secondary spheres were generated by replating of cells
dissociated from one individual primary sphere in each well. For tertiary and quaternary spheres,
pooled spheres dissociated to single cells were replated at a density of 20 cells/well. (D) SOX2+ and
SOX2- OVCAR-3 cells isolated by FACS were plated in sphere conditions 100 cells/well and spheres
counted after 7 days. For all replating assays, spheres were pooled, dissociated to single cells and
replated as indicated or at a density of 100 cells/well. Primary and secondary sphere formation was
analyzed in three or more, tertiary and quaternary sphere formation in two independent biological
experiments performed in triplicates (C, D).
Figure 5: SOX2 expression confers tumorigenic potential in vivo. SOX2-overexpressing and
control Caov3 cells were injected as indicated at same numbers contralaterally in the flanks of NSG
mice (n=5 mice per group). (A) As compared to controls, SOX2-overexpressing tumors were detected
faster and at higher frequency by palpation. (B) SOX2 expression results in increased tumor size but
unaltered metabolic activity, as revealed by [18F]FDG PET/MRI imaging (left panel: representative
image of a tumor-bearing mouse scanned 15 weeks after cell inoculation; right panel: tumor volumetry
and quantification of tracer accumulation). (C) SOX2-overexpressing tumors show similar Ki67 and
enhanced active caspase-3 staining by immunohistochemistry. (D) Limiting dilution assays reveal
increased tumor cell cluster formation of SOX2-overexpressing versus control cells, as shown by
immunohistochemical analysis of the injection site 7 weeks after inoculation. Tumor clusters were
detected in animals injected with 10.000 or 1.000 cells, but not with 100 or 10 cells. (E) H&E staining
of representative samples derived from mice inoculated with 1.000 cells. Pictures were taken at the
indicated magnification.
Figure 6: SOX2 effects on cell proliferation and basal apoptosis. SOX2 expression does not
influence cell cycle distribution, BrdU uptake (A), or cell growth (B) of 2D-cultured cells. (C) SOX2
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

22
protein expression and Ki67 staining in primary high-grade SOC samples. Analysis was performed on
a tissue microarray comprising 143 high-grade SOCs. (D) SOX2 expression modulates basal
apoptosis as measured by percentages of subG1 cells. Shown are data from three or more
independent biological experiments performed in triplicates in Caov-3 and OVCAR-3 cells (A, B, D).
Figure 7: SOX2 expression induces apoptosis resistance. Elevated SOX2 expression reduces
apoptotic responses after treatment with (A) staurosporine (STS), (B) TRAIL and (C)
chemotherapeutic drugs (cisplatin, carboplatin, paclitaxel). (D) SOX2 overexpression restores
apoptosis resistance in SOX2-knockdown cells. (E, F) SOX2 modulation does not change TRAIL-R1
and TRAIL-R2 surface expression, but influences mRNA expression of BCL2, BBC3 and PMAIP1.
Shown are percentages of subG1 cells and caspase-3/7-activities performed in Caov-3 and OVCAR-3
cells (A-D) and differences of median fluorescence intensities (E). Relative gene expression levels
were determined by real-time PCR and normalized to PBGD (D, F). The results of each panel
represent mean values ± SEM from three independent biological experiments performed in triplicates.
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

A
2D-culture
Spheres
ALDH+ CD133+ ALDH+/CD133+0
10
20
30
40
50
60
P=0.0481
P<0.0001
% p
os
itiv
e c
ells
Patient #1
C
Cao
v-3
OVCAR-3
OVCAR-5
OVCAR-8
SK-O
V-3
0
1
2150
250
350
450
e-04
rela
tiv
e S
OX
2 g
en
e e
xp
res
sio
n
#1 #2 #3 #40
0.1
0.20.5
2.5
4.5
e-03
cell lines patient samples
B
SOX2 ALDH1 Lin28 Oct4
0
1
2
35
15
25
rela
tiv
e g
en
e e
xp
res
sio
n
SOX2 ALDH1 Lin28 Nanog Oct401
3
5
7
9
11
rela
tiv
e g
en
e e
xp
res
sio
n i
n
sp
he
res
ve
rsu
s 2
D
Patient #2
***
******
***
***
SOX2 ALDH1 Lin28 Nanog Oct401
3
5
7
9
11
n.s.
** * n.s.
***
OVCAR-3 Caov-3
Figure 1
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177
A B
Protein expression
Actin
SOX2
control SOX2
40 kDa
35 kDa
SOX2 overexpression
Caov-3
control SOX20
25
50
75
100
P=0.0127
rela
tiv
e S
OX
2g
en
e e
xp
res
sio
n
SOX2 knockdown
OVCAR-3
control shSOX20.00
0.25
0.50
0.75
1.00
P<0.0001
rela
tiv
e S
OX
2g
en
e e
xp
res
sio
n
Actin
SOX2
40 kDa
35 kDa
control shSOX2
Figure 2
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

A BCaov-3
0
0.5
1.05.0
205220
600
SOX2 ALDH1 Lin28 Nanog Oct4
***
***
***
***
**control
SOX2
0
1
2
3
4
SOX2 ALDH1 Lin28 Nanog Oct4
Patient #3
n.s.
***
* *
***control
SOX2
0
1
2
15
25
SOX2 ALDH1 Lin28 Nanog Oct4
Patient #4
n.s.n.s.
*** ***
***
control
SOX2
Patient #1
SOX2 ALDH1 Lin28 Nanog Oct4
0
1
3
5
7 ***
******
***
***
control
SOX2
fold
re
lative g
ene
exp
ressio
n
in S
OX
2 o
ve
rexp
ressin
g v
ers
us c
ontr
ol ce
llsFigure 3
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

C
SOX2 overexpressionCaov-3
A
0.0
2.5
5.0
7.5
10.0
12.5
P=0.0161
nu
mb
er
of
sp
he
res
pe
r 1
00
ce
lls
control SOX2
SOX2 overexpression - single spheresCaov-3
control SOX2
0
5
10
15
nu
mb
er
of
sp
he
res
fro
m o
ne
dis
so
cia
ted
sin
gle
sp
he
re
secondary spheres
P=0.0021
control SOX2
0
5
10
15
nu
mb
er
of
sp
he
res
pe
r
20
pla
ted
ce
lls
tertiary spheres
P<0.001
control SOX2
0
5
10
15
20
25
30
nu
mb
er
of
sp
he
res
pe
r 1
00
pla
ted
sin
gle
ce
lls
P=0.0372
primary spheres
control SOX2 0
5
10
15
quaternary spheres
P=0.0085
nu
mb
er
of
sp
he
res
pe
r
20
pla
ted
ce
lls
0
5
10
15
control shSOX2
nu
mb
er
of
sp
he
res
pe
r 1
00
ce
lls
SOX2 knockdownOVCAR-3
B
D
SOX2 positive
0
20
40
60
80
100
SOX2 negative
P<0.001
0
5
10
15
20
25
30
SOX2 positive
SOX2 negative
P<0.001
0
5
10
15
20
25
30
SOX2 positive
SOX2 negative
P=0.0202
0
5
10
15
20
25
30
SOX2 positive
SOX2 negative
P=0.0167
secondary spheres tertiary spheres quaternary spheresprimary spheres
SOX2 reporter cellsOVCAR-3
SOX2 overexpression
control SOX2
0
5
10
15
20
25
30
35
GFP+ GFP-
secondary spheres
P=0.0016 (GFP+)
primary spheres
control SOX2
0
5
10
15P=0.0165 (all)
P<0.001 (GFP+)P<0.001 (all)
nu
mb
er
of
sp
he
res
pe
r 1
00
ce
lls
control SOX2
0
5
10
15
control SOX2
0
5
10
15
20
25
30
35
P<0.001 (GFP+)P<0.001 (all)
P=0.0166 (GFP+)P=0.0292 (all)
secondary spheresprimary spheres
Patient #1 Patient #2
nu
mb
er
of
sp
he
res
pe
r 1
00
ce
lls
nu
mb
er
of
sp
he
res
pe
r 1
00
ce
lls
nu
mb
er
of
sp
he
res
pe
r 1
00
ce
lls
nu
mb
er
of
sp
he
res
pe
r 1
00
ce
lls
P=0.0185
Figure 4
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

C H&E 12.5x SOX2 12.5x Ki-67 12.5x
Co
ntr
ol
SO
X2
B
500.000 cellsA 100.000 cells
ControlSO
X2
0 2 4 60
1
2
3
4
5Control
SOX2
weeks
nu
mb
er
of
pa
lpa
ble
tu
mo
rs
0 2 4 6 8 10 12 140
1
2
3
4
5Control
SOX2
weeksn
um
be
r o
f
pa
lpa
ble
tu
mo
rs
p.i. 15 weeks p.i.V
olu
metr
y [
mm
3]
0
250
500
750
1000
1250
%ID
/cc [
18F
]FD
G
0
1
2
3
4
n=1 n=4 n=4n=4
8 weeks
p.i. 15 weeks p.i.
n=1 n=4 n=4n=4
8 weeks
500.000
cells
100.000
cells
500.000
cells
100.000
cells
Control SOX2
Caspase-3 12.5x
0 1 3 5 7 9 110
1
2
3
4
5Control
SOX2
weeks
nu
mb
er
of
pa
lpab
le t
um
ors
50.000 cells
E SOX2 Control SOX2 Control
H&E 50x H&E 630x
D
Cells injected
Immunohistological tumor
detection after 7 weeks
10.000
SOX2 control
1.000
100
10
4/5 3/5
4/5 1/5
0/5
0/50/5
0/5
Figure 5
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

SOX2 overexpression SOX2 knockdownA
D SOX2 knockdown
B SOX2 overexpression
C Primary SOC samples
Caov-3 OVCAR-3 Caov-3
OVCAR-3
SOX2 overexpression
Caov-3
OVCAR-3
SOX2 knockdown
Control SOX20
10
20
30
40
% B
rdU
po
sit
ive
cells
Control shSOX20
5
10
15
20
25
30
35
% B
rdU
po
sit
ive
ce
lls
Control SOX20
2
4
6
8
10
12
14
P=0.004
Su
b G
1 f
racti
on
%
Control shSOX20
2
4
6
8
10
12
Su
b G
1 f
racti
on
%
SOX2 positivity
negative low medium high0
10
20
30
40
50
60
70
80
90
% K
i-6
7 p
os
itiv
ity
G0-phase S/M/G2-phase
0
10
20
30
40
50
60
70
SOX2
Control
% P
I p
os
itiv
e c
ells
G0-phase S/M/G2-phase
0
10
20
30
40
50
60 Control
shSOX2
% P
I p
os
itiv
e c
ells
days
0 1 2 3 4 5 6 7 8 9 100
2.5
5.0
7.5Control
SOX2
nu
mb
er
of
via
ble
ce
lls
days
0
2.5
5.0
7.5
10.0
12.5
Control
shSOX2
P=0.0053
0 1 2 3 4 5 6 7 8 9 10
(x1
05)
nu
mb
er
of
via
ble
ce
lls
(x1
05)
Figure 6
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

A Staurosporine treatment
Chemotherapytreatment
C
B TRAIL treatment
SOX2 knockdownSOX2 overexpression SOX2 overexpression SOX2 knockdown
SOX2 overexpression
D Rescue of SOX2 expression in SOX2 knockdown OVCAR-3 cells
Staurosporine treatment TRAIL treatment
E SOX2 knockdownSOX2 overexpression F SOX2 overexpressionCaov-3 OVCAR-3
Caov-3 OVCAR-3 Caov-3 OVCAR-3
Caov-3
Caov-3
Control SOX2
0
50
100
150
200
250
anti-apoptotic
P=0.0042
rela
tiv
e g
en
e e
xp
res
sio
no
f B
CL
2
BBC3 PMAIP1
0
20
40
60
80
100
120
Control
SOX2P=0.0001
pro-apoptotic
rela
tiv
e g
en
e e
xp
res
sio
n
TRAILR1 TRAILR2
0
1000
2000
3000Control
shSOX2
Dif
fere
nce
of
med
ian
flu
ore
sce
nc
e in
ten
sit
y
TRAILR1 TRAILR2
0
200
400
600
800
1000
1200Control
SOX2
Dif
fere
nc
e o
f m
ed
ian
flu
ore
sc
en
ce
in
ten
sit
y
Cisplatin Carboplatin Paclitaxel
0
10
20
30
40
50
60
70
Control
SOX2
P=0.0073
P=0.0040
P=0.0232
Su
b G
1 f
rac
tio
n %
(sp
ec
ific
ap
op
tos
is)
Control shSOX2 shSOX20
10
20
30
40
50
60
P=0.0014 P<0.0001
+ SOX2
Su
b G
1 f
rac
tio
n %
(sp
ec
ific
ap
op
tos
is)
Control shSOX2 shSOX20.00
0.25
0.50
0.75
1.00
P<0.0001 P=0.0078
+ SOX2
rela
tiv
e S
OX
2g
en
e e
xp
ress
ion
Control shSOX2 shSOX20
10
20
30
40
50
60P=0.0120
+ SOX2S
ub
G1
fra
cti
on
%(s
pe
cif
ic a
po
pto
sis
)
Control SOX20
25
50
75
100
P<0.0001
rela
tiv
e R
LU
(%
)
Control shSOX20
25
50
75
100
125
150
175
rela
tiv
e R
LU
(%
)
Control shSOX20
100
200
300
400
500
600
rela
tiv
e R
LU
(%
)
Control SOX20
25
50
75
100
P=0.0001
rela
tiv
e R
LU
(%
)
Control SOX20
10
20
30
40
50
60P<0.0001
Su
b G
1 f
rac
tio
n %
(sp
ec
ific
ap
op
tos
is)
Control SOX20
10
20
30
40
50
60
P=0.0073
Su
b G
1 f
rac
tio
n %
(sp
ec
ific
ap
op
tos
is)
Control shSOX20
10
20
30
40
50
60
P=0.0036
Su
b G
1 f
rac
tio
n %
(sp
ec
ific
ap
op
tos
is)
Control shSOX20
10
20
30
40
P=0.0060
Su
b G
1 f
rac
tio
n %
(sp
ec
ific
ap
op
tos
is)
Figure 7
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177

Published OnlineFirst July 18, 2013.Cancer Res Petra M Bareiss, Anna Paczulla, Hui Wang, et al. ovarian carcinomaSOX2 expression associates with stem cell state in human
Updated version
10.1158/0008-5472.CAN-12-4177doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2013/07/18/0008-5472.CAN-12-4177.DC1
Access the most recent supplemental material at:
Manuscript
Authoredited. Author manuscripts have been peer reviewed and accepted for publication but have not yet been
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/early/2013/07/18/0008-5472.CAN-12-4177To request permission to re-use all or part of this article, use this link
on May 26, 2018. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Author manuscripts have been peer reviewed and accepted for publication but have not yet been edited. Author Manuscript Published OnlineFirst on July 18, 2013; DOI: 10.1158/0008-5472.CAN-12-4177