social organization of woodchucks (marmota monax)
TRANSCRIPT

Behav Ecol Sociobiol (1992) 31:393-400 Behavioral Ecology and Sociobiology © Springer-Verlag 1992
Social organization of woodchucks (Marmota monax) Paul T. Meier *
Department of Zoological and Biomedical Sciences, Ohio University, Athens, OH 45701, USA
Received September 26, 1990 / Accepted July 18, 1992
Summary. The social organization of woodchucks (Mar- mota monax) in southeast Ohio was studied at two sites, at one for two 2 years (1979-1980) and the other for 3 years (1981 1983). Spatial organization was deter- mined by trapping and radio tracking. The home ranges of adult females did not overlap in the early spring but during late spring and summer there was some overlap (< 10%) as females expanded their home ranges. Adult females tended to occupy the same home range in con- secutive years. Some adult males occupied well-defined home ranges that did not overlap the home ranges of other males but did overlap extensively the home range of one to three adult females. These males tended to occupy the same home range in consecutive years. In- fants used the same home range of their dam until about 2-3 months of age when most males and females appar- ently dispersed. About 35% of the juvenile females did not disperse until their second spring, just before their mother's new litter first emerged from their burrow. The average social group consisted of an adult male with two female kin groups comprising an adult female, an offspring (usually female) of the previous year, and the young of the year. Interactions within the kin group and with the adult male were relatively frequent and generally amicable. Interactions between kin groups both within and between different social groups were relatively rare and agonistic. The social organization of woodchucks in Ohio differs from that described in pre- vious studies of woodchucks elsewhere and from that predicted by current models proposed by others on the evolution of social organization of marmots.
Introduction
The evolution of social behavior in species within the genus Marmota has received considerable attention (Bar- ash 1974a, 1989; Elliot 1975; Armitage 1981, 1988; Mi-
* Present address: Biology Department, Cedar Crest College, 100 College Drive, Allentown, PA 18104, USA
chener 1983; Arnold 1990a, b). Olympic marmots (Mar- mota olympus) and hoary marmots (M. caligata) are highly social and live in small colonies typically compris- ing an adult male, several adult females, 2-year-olds, yearlings, and infants (Barash 1973, 1974b). The area inhabited by the colony is available equally to all colony members. Yellow-bellied marmots (M. flaviventris) are less social. Although they live in groups similar in com- position to those of Olympic marmots and hoary mar- mots, resources are usually not shared equally among all colony members (Armitage 1962, 1986, 1988). Re- sources may be shared among members of a matriline, but members of different matrilines do not share re- sources even though they may be closely related (Armi- tage 1984). Woodchucks (M. monax) have been consid- ered to be asocial, with no prolonged associations be- tween individuals outside of the breeding season (Griz- zell 1955; de Vos and Gellespie 1960; Anthony 1962; Bronson 1964; Ferron and Quellet 1989). However, Ha- milton (1934) stated that adult woodchucks occasionally shared burrows at times other than the breeding season.
Barash (1974a) suggested that the difference in social organization among marmot species is the result of dif- ferences in the age of natal dispersal. Natal dispersal is delayed longest in Olympic marmots because they ex- perience the shortest vegetative growing season. The high degree of sociality exhibited by Olympic marmots is due to the need to inhibit dispersal of undersized young. Yellow-bellied marmots and woodchucks have progressively longer vegetative growing seasons, earlier age of natal dispersal, and are less social.
Armitage (1981) also noted a correlation between de- gree of sociality and age of dispersal in North American ground-dwelling sciurids. He proposed that sociality was a life-history tactic that allowed continuation of repro- ductive investment beyond weaning in large species liv- ing where the growing season is too short to enable off- spring to become reproductively mature in their first year of life.
Michener (1983) also examined the relationships be- tween sociality and life history traits in ground-dwelling sciurids. She noted that although the most social ground

394
squirrels are large, their act ive season ranges f rom short , to long, to con t inuous . She sugges ted tha t increased so- c ia l i ty was co r re l a t ed wi th increased over lap in the act ive seasons be tween y o u n g and adults . In asocia l species, adul t s en ter h i b e r n a t i o n weeks before y o u n g enter h iber- na t ion . Social species have increased co inc idence o f ac- tive season o f adul t s and y o u n g t h r o u g h p r o l o n g e d ac- t ive seasons or shor t ened act ive seasons assoc ia ted wi th de layed dispersal .
In this paper , I f irst descr ibe the social o rgan i za t i on o f w o o d c h u c k s in sou theas t Ohio. I then c o m p a r e their social o rgan i za t i on with tha t o f w o o d c h u c k s in Pennsyl- van ia and o the r species o f m a r m o t s to test the mode l s p r o p o s e d by Ba ra sh (1974a), A r m i t a g e (1981), and Mi- chener (1983).
Materials and methods
Two study sites were used. Study Site 1 was a 41-ha site 10 km southeast of Athens, Ohio and was studied from spring of 1979 through the summer of 1980. Study Site 2 was a 60-ha site 12 km northeast of Athens, Ohio and was studied from spring of 1981 through fall 1983. Both sites contained woodland and field habitats. The woodland habitat was second-growth woodlands and the field habitat was grazed by farm animals.
Woodchucks were captured with live-traps placed at burrow entrances. Woodchucks were ear-tagged, weighed, sexed, classified as infant, yearling, or adult according to the criteria of Snyder et al. (1961), and their reproductive condition noted. Infants were also marked with non-toxic for dye for visual identification.
Most of the captured adults and yearlings and some infants were tagged with radio-transmitters. Transmitter collars were made of nylon webbing of various colors, which aided in visual identifica- tion. An attempt was made to locate each radio-tagged individual at least twice daily, once between sunrise and 1000 hours (EST) and again between 1600 hours and sunset, during the period from March through October.
An individual's home range was determined by connecting the outermost location points for that individual. Sites where an indi- vidual was found less than 2% of the total times that it was located were not used in determining the individual's home range.
Results
Spacing behavior
The fo l lowing desc r ip t ion o f spac ing behav io r is der ived f rom the m o v e m e n t s o f 31 r a d i o - t a g g e d w o o d c h u c k s consis t ing o f B adu l t males , 14 adu l t females, 2 year l ing males , 5 year l ing females, 1 in fan t male , a n d 1 infant female (Table 1). A d d i t i o n a l i n f o r m a t i o n on the move- men t o f infants was o b t a i n e d f rom obse rva t ions and t r app ing .
Adult females. Afte r emergence f rom h i b e r n a t i o n in ear ly M a r c h unt i l m i d - M a y , adu l t females used on ly a smal l a rea a r o u n d the b u r r o w tha t they h a d used as a h ibe rna - culum. The average size o f the h o m e range o f 6 r ad io - t agged adu l t females was 2512 m 2 (SD = 761). The aver- age d is tance be tween neares t ne ighbor adu l t females was 104 m. A t tha t t ime, spac ing was p r o b a b l y no t ma in - t a ined by in te rac t ions a m o n g the females because the home ranges o f females were n o t con t iguous (Fig. 1 A).
Table 1. Dates and the number of times located for the radio-tagged woodchucks used in the analysis of spacing patterns
ID Sex Age Dates Year No. of times located
274 M Adult 25 June-12 July 1979 72 262 M Adult 25 March-28 April 1980 36 419 M Adult 17 April-23 May 1980 48 440 M Adult 7 May-23 June 1980 56 472 M Adult 27 March-] 7 Aug 1981 152 472 M Adult 23 Apri l16 July 1982 72 472 M Adult 22 Feb- 4 Aug 1983 109 401 M Adult 4 May- 8 July 1982 67 401 M Adult 2 March- 4 April 1983 38 24 M Adult 20 May-10 July 1982 34
420 M Adult 15 Aug- 1 Sept 1982 14 254 F Adult 5 July-17 Aug 1979 81 418 F Adult 17 April-23 May 1980 44 417 F Adult 18 April-17 May 1980 39 426 F Adult 25 April-17 May 1980 33 414 F Adult 24 April-24 May 1980 42 431 F Adult 16 May- 6 June 1980 26 464 F Adult 16 May-20 June 1980 41 469 F Adult 3 April-24 July 1981 119 459 F Adult 29 April-30 June 1981 62 452 F Adult 29 April-27 Oct 1981 172 452 F Adult 31 May-10 July 1982 18 436 F Adult 8 May- 5 Oct 1981 151 436 F Adult 2 July-11 Sept 1982 40 436 F Adult 24 Feb-17 March 1983 33 491 F Adult 22 May18 July 1981 57 491 F Adult 31 May-16 July 1982 26 491 F Adult 27 April-26 May 1983 20 483 F Adult 8 July-27 Aug 1981 53
8 F Adult 20 April-16 July 1982 116 412 M Yrtng 28 April-26 July 1983 33
7 M Yrlng 18 May-20 July 1982 18 463 F Yrlng 14 April- 6 May 1981 21 451 F Yrlng 14 April- 3 June 1981 44 478 F Yrlng 4 May- 7 May 1982 5 482 F Yrlng 14 May-23 May 1982 14
5 F Yrlng 14 July-- 2 Sept 1982 22 288 a F Yrlng 12 March-26 June 1980 12
10" F Yrlng 6 April- 6 Oct 1983 54 412 b M Infant 14 Aug-10 Sept 1982 18 417 M Infant 20 Aug- 8 Oct 1982 19
a Data on the movements of these two yearling females were ob- tained by observation, not by radio-tracking. Both wore collars which aided in identification b This infant male was also radio-tagged as a yearling
Af te r the y o u n g were b o r n in mid -Apr i l , females g r adua l l y increased the size o f thei r home range to for- age in a reas where vege ta t ion was m o r e plent iful . O f 19 females 17 m o v e d thei r y o u n g when they were 3 -4 weeks o ld f rom the na ta l b u r r o w to a n o t h e r b u r r o w tha t was closer to fo rag ing areas. The adu l t females con- t inued to e x p a n d their home range unt i l mid-June , when the size o f the home range s tabi l ized for the r e m a i n d e r o f the year . The average size o f the s u m m e r home range o f 13 a d u l t females was 13485 m 2 ( S D = 2 4 0 8 ) . D u r i n g the summer , the home ranges o f a d u l t females were in con tac t bu t wi th lit t le ove r l ap ( < 5%, Fig. 1 A). Al l adu l t females con t inued to use the h i b e r n a c u l u m / n a t a l b u r r o w

395
. . . . . . . . iiiii!iiiiiiii;!ii i > :: :: !;ii!iiiii!i
':: iliiiiii iiiiiii I
• :iiii!:: .:!:i:i:i:i:i:!:{:i:
~:i:!:: . . . . . . . :.::i!!~i![!i~i!i~i!ii~!!!. !i!i!i!!!ii!!ili!i!i::i::!:!~!!!#!!!!!::::" i!!!i!#i!!!i!i!!!i!i!!{
ii}iiiiiiBii!iiiii!iiiii!iiiii::: ~ ...:.:.: . . . . . . . . . ~iili]
iiii<::,:!iiiiiiii , ? • iiiii!iiiiiiiiii!i!i%iiiiiii;i i iiiiiiiii iiiil i 1 . . . . ':: ' iiiiiiiiiiiU":iiiiiiiii iiiiiiiiiiiiiiii ::
iii!iii = woodland
Q = burrows I00 m
9[ - hibernacula
Fig. 1 A, B. Home ranges of adult female and adult male wood- chucks. A Partial home ranges of 5 adult females showing areas of overlap. Shaded areas are the home ranges of the females during early spring (before 15 May). Note that the home ranges of females may overlap with each other. B Home ranges of two adult maies. Note that the home ranges of males are not contiguous with each other but do overlap the home ranges of adult females
during the summer but the frequency of their visits to that burrow varied considerably (X= 18% of times lo- cated, range 4~25%). In the fall the trend observed in the spring was reversed; the home range contracted to a small area around the hibernaculum.
Adult males. Adult males can be classified into two types based upon their space utilization behavior: (1) resident males whose activity was confined to a well defined area; and (2) transient males, which spent only short periods of time in any one area. The behavior of transient males is discussed elsewhere (Meier 1985).
The home range of radio-tagged resident adult males (n = 8) in the spring included the hibernacula/natal bur- rows of 1-3 (x = 2.0) adult females (Fig. 1 B). The males moved frequently between these burrows, making at least daily visits to each of them. The home ranges of resident adult males were not contiguous with each other in the spring or in the summer (Fig. 1 B). The home range of males in the spring was 85% of the size of the summer home range (16075m 2, SD=2941) . One resident adult male was radio-tagged for 2 consecutive years and another male for 3 years; these males used
the same home range each year. Three other adult males were recaptured but not radio-tagged within their home ranges of the previous year, when they had been radio- tagged.
Yearlings. Less information is available on the home ranges of yearlings. Five yearling females and two year- ling males were radio-tagged (Table 1). Four of the fe- males and one of the males had also been captured as infants and had remained in their dams' home range.
One of the yearling males wandered over a large area, covering a distance of about 1 km during the 2 months that his movements were monitored. The second yearling male had a home range that overlapped that of his dam by about 30% and was completely within the home range of his putative sire. The yearling's home range was similar to that used by an adult female that had died in March of that year.
Four of the five radio-tagged yearling females appar- ently dispersed out of the study area in May when they were about 1 year old. One of these was the only yearling female known to have produced a litter. She was lactat- ing when captured on 14 April but her space utilization behavior in late April and early May was not typical of adult females with young; she ranged over a wider area and relocated more frequently. After 7 May I was unable to locate or recapture her.
Only one radio-tagged yearling female is known to have not dispersed out of the study area. However, two additional yearling females were observed often enough (one 12 times from 12 March to 20 June, the other 54 times from 6 April to 6 October) to allow estimation of their home ranges. Two of these yearling females used portions of their dams' home range. During April, their dams shifted their home range towards one end, result- ing in little overlap between the yearlings' and adults' home ranges. The home range of the third yearling bor- dered the home range of her dam. This area had been used the previous year by an adult female that had moved to another area. This yearling female was recap- tured as an adult in the same area.
Infants. The following description of spacing behavior of infants is based on 28 individuals (17 females and 11 males) that I captured between 5 May and 2 June at burrows occupied by other infants (putative siblings) and an adult female (putative mother). Information on the movements of infants is from two radio-tagged indi- viduals and observations of marked individuals.
Infants first appeared above ground in early May. During the first 2-3 weeks above ground, the young foraged and played among themselves and with their mother in close proximity to the burrow from which they had emerged. At 3 weeks after emergence, the in- fants began to follow their mother to foraging areas further away from the burrow and also began to use other burrow systems within their dam's home range.
Five (3 males and 2 females) of the 28 infants cap- tured were later (earliest date was 17 June) captured outside of their mother 's home range. The remaining 23 infants were last observed within their mother 's home
396
range. Nine infants were last seen between 7 May and 12 June, and 3 others disappeared during July. Eleven other infants (9 females and 2 males) remained within the home range of their mother until at least September (range of date of last observation: 2 Sep tembe~l 8 No- vember). These infants used the same areas used by their mother during the summer months, but they were usual- ly (102 of 132 observations) observed at burrows other than the one occupied by their mother. All infants recap- tured or observed as yearlings (n = 7) were within their dams' home range. All seven of these individuals (six females and one male) probably used the same hiberna- culum as their dam as they were last seen before hiberna- tion and first seen after hibernation at the hibernaculum.
Social organization
Females form social groups consisting of an adult female and her offspring of the current year and sometimes (at least 6 out of 19 litters produced) one of her female offspring from the previous year. Males attach to one or more female kin groups to form harems. Within a harem comprising more than one female kin group, the adult male was the only member that regularly inter- acted with members of more than one female kin group. Interactions between female kin groups both within and among harems were rarely observed (n = 4 interactions during the entire study). There was very little overlap ( x < 5 % of home range) between the areas occupied by female kin groups regardless of whether or not they be- longed to the same harem.
Within a female kin group, interactions were more frequent. During the spring, from the breeding season until the young were 2-5 weeks of age, the adult male made daily visits to the burrows occupied by the adult females and yearlings. Adult females and yearlings shared the same burrow in early spring but occupied different burrows beginning about the time the adult female gave birth. Interactions between adult females and yearlings virtually ceased at that time.
Social behavior and interactions
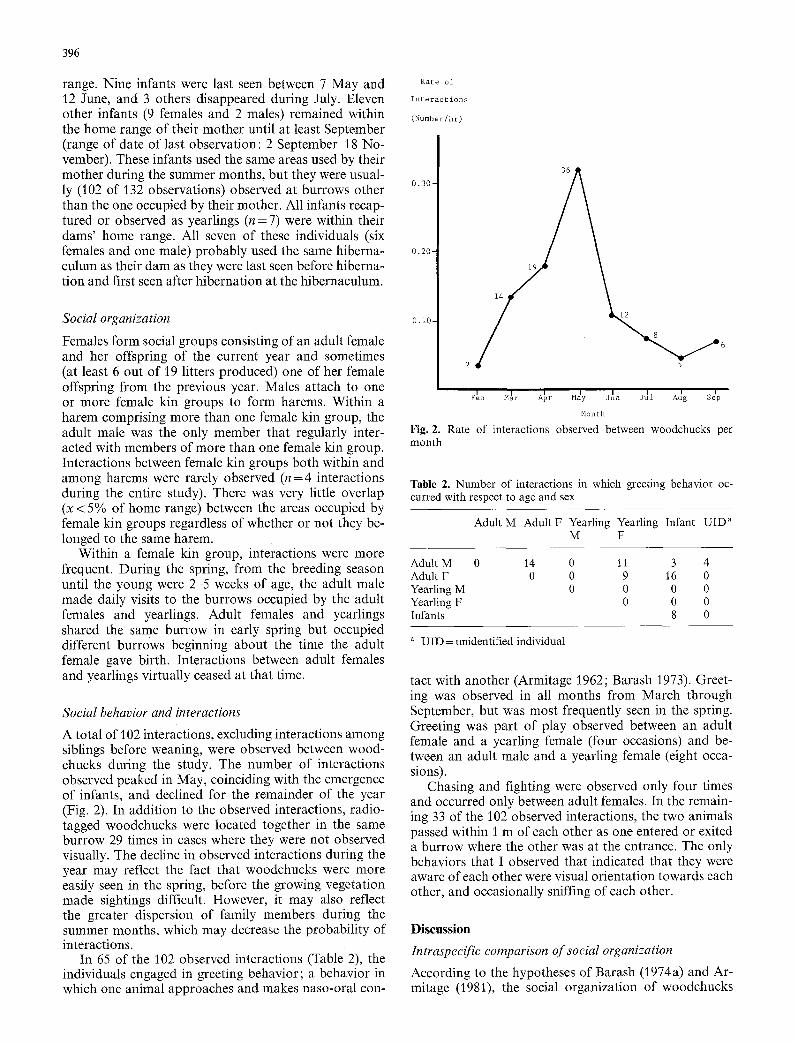
A total of 102 interactions, excluding interactions among siblings before weaning, were observed between wood- chucks during the study. The number of interactions observed peaked in May, coinciding with the emergence of infants, and declined for the remainder of the year (Fig. 2). In addition to the observed interactions, radio- tagged woodchucks were located together in the same burrow 29 times in cases where they were not observed visually. The decline in observed interactions during the year may reflect the fact that woodchucks were more easily seen in the spring, before the growing vegetation made sightings difficult. However, it may also reflect the greater dispersion of family members during the summer months, which may decrease the probability of interactions.
In 65 of the 102 observed interactions (Table 2), the individuals engaged in greeting behavior; a behavior in which one animal approaches and makes naso-oral con-
Rate of
Interactions
(Number/hr)
0.30-
O. 20..~
0 .~ (
36
19
14
/ 2 5
F~b M~r A~r M~y J~n J~l A~g Sgp
Month
Fig. 2. Rate of interactions observed between woodchucks per month
Table 2. Number of interactions in which greeting behavior oc- curred with respect to age and sex
Adult M Adult F Yearling Yearling Infant UID a M F
0 14 0 11 3 4 0 0 9 16 0
0 0 0 0 0 0 0
8 0
Adult M Adult F Yearling M Yearling F Infants
a UID = unidentified individual
tact with another (Armitage 1962; Barash 1973). Greet- ing was observed in all months from March through September, but was most frequently seen in the spring. Greeting was part of play observed between an adult female and a yearling female (four occasions) and be- tween an adult male and a yearling female (eight occa- sions).
Chasing and fighting were observed only four times and occurred only between adult females. In the remain- ing 33 of the 102 observed interactions, the two animals passed within I m of each other as one entered or exited a burrow where the other was at the entrance. The only behaviors that I observed that indicated that they were aware of each other were visual orientation towards each other, and occasionally sniffing of each other.
Discussion
Intraspecific comparison of social organization
According to the hypotheses of Barash (1974a) and Ar- mitage (1981), the social organization of woodchucks

Table 3. Comparison of four populations of marmots (Marmota) with respect to ab- solute and relative age of female natal dis- persal and overlap in the active seasons of juveniles and adults
397
M. M. M. M. monax" monax b flaviventris ° olympus d
Rate of weight gain 18.8 19.9 14 16.6 infants (g/day) Active season Adults 7 7 4.5 3.5 (months) Infants 5 5 2.5 2.0 Age of female 2 dispersal (months) e 13 (65%) 2 5 (100%) 13-25 (79%) 25-27 (75%) No. of months active 2-5 infant before dispersal 7.5 yrlng 2-5 4-8 7.5 Age of female first 12% yrlng 53% yrlng 25% 2-yr. 3-yr. reproduction 88% 2-yr. 47% 2-yr. 75% 3-yr. No. of months active before first 5 yrlng 5 yrlng 7 2-yr reproduction 12 2-yr. 12 2-yr. 11 3-yr 12.5 % Overlap of adult and infant active seasons 90-95 9~95 95 100 90 100 (adult/infant)
a Marmota monax in Ohio, data from this study, Meier (1985) b M. monax in Pennsylvania, data from Snyder et al. (1961) c M.flaviventris, data from Armitage (1981); Armitage and Downhower (1974); Armitage et al. (1976) a M. olympus, data from Barash (1973) ° The number in parentheses is the percentage of female young that dispersed or died before the oldest age indicated for each population
in Ohio and Pennsylvania should be similar because they experience vegetative growing seasons of similar length (Table 3). However, there are major differences in social organization between these populations. Three of these differences will be discussed: spacing behavior, age of juvenile dispersal, and relative frequency of amicable and agonistic behaviors.
S p a c i n g behav ior . All adult female and some adult male woodchucks in Ohio occupy distinct, well-defined home ranges throughout the year and from one year to the next. There is little intrasexual overlap in home ranges. Intrasexual spacing of adults is probably maintained through aggressive interactions and scent marking with- in the home range. In this regard, woodchucks can be considered to be territorial, that is, adults defend exclu- sive (with regard to sex) areas (Brown and Orians 1970).
In contrast, Bronson (1964) found that the overlap in home ranges of woodchucks in Pennsylvania averaged 36% (range 4-77%). The use of space within the areas of overlap was determined through simple dominance hierarchies among neighbors. Bronson did not distin- guish between woodchucks of different age or sex when determining overlap in home ranges. He did provide an example of overlap in home ranges of five neighboring woodchucks consisting of two adult females, one adult male, one yearling male and an unidentified yearling (Bronson 1964). There was little overlap in the home ranges of the adult females. Most of the overlap was between the adult male with one of the adult females and between the adult females with the yearlings. Thus, the apparent difference in spacing behavior between the Ohio and Pennsylvania populations may only reflect dif-
ferences in interpretation rather than real differences in behavior.
Ferron and Quellet (1989) described the spatial orga- nization of a northern population of woodchucks as ter- ritorial. Unlike the Ohio population, there was virtually no overlap in the home ranges of adult males and adult females after the dispersal of young. They suggested that this difference in spacing behavior between the Ohio and northern population was related to a lower population density in the northern population. Bronson (1964) also found that the degree of overlap in home ranges de- creased within the Pennsylvania population with a de- crease in density. Davis (1981) suggested that density of woodchucks in Pennsylvania was related to food abundance.
Disper sa l . All infant woodchucks in Pennsylvania and most infants in Ohio apparently disperse during their first summer, at about two months of age. However, in Ohio, 25% (7/28) of the infants that were captured did not disperse until their second spring, when they were about 13 months old. These infants were usually females, but one male was also known to have dispersed as a yearling.
Young should delay dispersal if the benefits they gain outweigh the costs of sharing resources that contribute to their mother 's future reproductive success. One poten- tial benefit to young is increased protection from preda- tion by group vigilance. However, young woodchucks that delay dispersal probably gain little increase in pro- tection from predation because they tend not to forage in groups and alarm calling was rarely heard (n= 17 during the study).

398
Young that delay dispersal may benefit by gaining access to some resource important for survival and re- production. For woodchucks, these resources are most likely food and burrows.
I did not measure food supply for my population of woodchucks, but it appeared that food was in excess. Foraging areas of woodchucks were recognizable be- cause of paths through the area but there was no obvious depletion of the vegetation in those areas during the summer. In addition, the home ranges of individuals monitored for successive years did not change in size although farming activities (grazing by farm animals, cutting of hay) in the study area did change during that time.
Burrows also appeared to be in excess, as there were more burrow systems than woodchucks. However, only certain burrows appeared to be suitable for use as hiber- nacula. Although I do not have data to test whether some burrows were more suitable (i.e. increased the probability of survival during the winter) for use as hi- bernacula, indirect evidence indicates that it is likely. Hibernacula were usually located in areas of rock out- crops and differed from other burrows in a number of features (Meier 1985). Also, woodchucks used the same burrows for hibernacula each winter. If hibernacula were limited, then it may benefit a juvenile to share its mother's hibernaculum until it reaches a size at which it can successfully compete for one of its own.
In Pennsylvania, where all young woodchucks dis- perse at an earlier age, there is less information on the availability of hibernacula. Bronson's (1963, 1964) study area was located in the ammunition storage area of Let- terkeny Army Ordnance Depot. The majority of wood- chuck burrows, including at least some hibernacula, were located in earthen bunkers that occurred in parallel lines throughout the study area. The home ranges of adults included 2-3 bunkers. If burrows located in these bunkers were suitable for use as hibernacula, then hiber- nacula were probably not limiting. Thus the difference in the age of dispersal of some young woodchucks in Ohio relative to Pennsylvania most likely reflects a dif- ference in the availability of hibernacula.
Social behavior. Bronson (1963, 1964) described the so- cial behavior of woodchucks in Pennsylvania as essen- tially agonistic. He observed 66 agonistic interactions in the field, consisting of 27 chases, 13 threats, and 26 passive interactions where only one individual displayed an overt response. He observed greeting behavior only twice in the field, and these apparently occurred in agon- istic situations.
Bronson's (1964) data do not include interactions among siblings and their mother before weaning, and 29 of the 102 interactions that I observed in the Ohio population were of that type. Of the remaining 79 inter- actions, that I observed, 75 of them were classified as nonagonistic. All of the nonagonistic interactions were among siblings and their mother or between adult males with adult females or subadults. The generally nonag- gressive nature of interactions between mothers and off- spring most likely reflect the delay in dispersal of young.
In the Pennsylvania population of woodchucks, there were no prolonged associations between adult males and adult females outside of the breeding season (Bronson 1964). In contrast, in the Ohio population adult males and adult females associated with each other throughout the year and often from one year to the next. This in- creased length of association and the accompaning ami- cable interactions between the male and female most likely reflects the mating strategy of some males (Meier 1985).
Interspecific comparison of social organization
The typical composition of social groups within Olympic marmots (Barash 1973) and yellow-bellied marmots (Ar- mitage 1962, 1984, 1986, 1988) is the same as that found for woodchucks in Ohio : a male with one to three adult females and the immature offspring of the females from their previous two litters. However, the frequency and nature of interactions among individuals within the so- cial group differ among the three species.
When reproductively mature, female woodchucks in Ohio have either dispersed or gained access to portions of their mother's home range. No interactions between a mother and her reproductively mature female offspring were observed. In contrast, some female yellow-bellied marmots remain with their mother when reproductively mature and continue to interact amicably with her and their sisters (Armitage 1984, 1986, 1988). However, inter- actions between unrelated females within the social unit are agonistic (Armitage and Johns 1982; Armitage 1986, 1988). Interactions between unrelated females are usual- ly much less frequent than between related females and decline over time.
In Olympic marmots, interactions between all resi- dent females are frequent and most often amicable (Bar- ash 1973). Unfortunately, the exact genealogical rela- tionships of the individuals within the social group are unknown. If the social organization of Olympic mar- mots follows the pattern observed in yellow-bellied mar- mots, then the social groups are most likely composed of highly related females.
There are two competing hypotheses that have been proposed to explain the difference in degree of sociality among species of marmots. The first hypothesis, pro- posed by Michener (1983) is that greater social tolerance is correlated with greater coincidence of the active peri- ods of young and adults via prolonged active seasons or shortened active seasons associated with delayed dis- persal. Her hypothesis does not explain the difference in social organization between the Ohio and Pennsylva- nia populations of woodchucks, because the two popula- tions have very similar circannual cycles (Table 3). Her hypothesis also does not explain the difference in social organization among Olympic marmots, yellow-bellied marmots and woodchucks in Ohio. Among these popu- lations there is no correlation between degree of sociality with either coincidence of the active seasons of young and adults nor with the length of time young are active before dispersal (Table 3).

399
The second hypothesis, proposed by Barash (1974a) and modified by Armitage (1981), is that increased socia- lity occurs in species of marmots when they experience relatively short growing seasons resulting in reproduc- tive maturity being delayed to age two or later. Increased sociality in these species is due to the necessity to inhibit for longer periods of time the dispersal of undersized young. This hypothesis is rejected as an explanation for the difference in social organization between wood- chucks in Ohio and Pennsylvania because these popula- tions experience active seasons of equal length (Table 3). Nor does their hypothesis explain the difference in degree of sociality among woodchucks in Ohio, yellow- bellied marmots, and Olympic marmots.
Natal dispersal of female woodchucks in Ohio is bi- modal. Approximately 65% of the captured female in- fants were not observed in their natal area after their first summer. The remaining females remained in their natal area until they were yearlings. Most female wood- chucks first reproduce as two year olds [7 of 8 in this study; 47% in Pennsylvania (Snyder and Christian 1960), 74% in New York (Hamilton 1934), 75% in Mar- yland (Grizzell 1955)]. In yellow-bellied marmots and Olympic marmots, natal dispersal of about 75% of the females also occurs at least one year before most become reproductively mature (Table 3).
Although the absolute age of female natal dispersal increases from woodchucks to yellow-bellied marmots to Olympic marmots, the difference is not due to differ- ent growth rates of the young among those populations (Table 3). Rather, the difference in age of dispersal re- flects the amount of time that young are active, and thus able to gain weight, each year. The number of months that female young are active before dispersal is approximately equal in all three species (Table 3). Hence there is no difference in the length of time that dispersal of undersized young must be inhibited through social interactions among the three species of marmots. Female yearling yellow-bellied marmots are apparently of sufficient size for dispersal as evidenced by the equal survivorship of nondispersing and dispersing yearlings (Downhower and Armitage 1981).
Because dispersal does not occur and social interac- tions are much less frequent during hibernation, it is the period of time that young are active before dispersal that should be considered when testing the hypotheses of Barash (1974a) and Armitage (1981). After females have been active for about 7 months after birth, 65% of M. monax, 79% of M. flaviventris (Armitage and Downhower 1974), and 75% of M. olympus (Barash 1973) have either dispersed or died. Because the number of months young are active before dispersal of about 25% of the female young is similar in all three species, other factors besides length of growing season and de- layed dispersal of undersized young must be important in determining the degree of sociality in species of mar- mots. Increased sociality in yellow-bellied marmots and Olympic marmots is correlated with retention of two- year old female offspring and three-year old female off- spring respectively. Retention is not because those off- spring are of insufficient size for dispersal. Also, the
offspring that are retained within their natal colonies do not have more opportunities for social interaction with their mother than l-year-old female woodchucks that disperse as yearlings.
The alpine marmot M. marmota has a similar degree of sociality to the Olympic marmot (Barash 1976; Ar- nold 1990a, b). Arnold found that the high degree of sociality in alpine marmots was not due to the inhibition of dispersal of undersized young. Instead, he suggests that the sociality is due to the benefits of joint hiberna- tion. The variation in degree of sociality among the North American marmots may reflect differences in the benefits of joint hibernation. The benefits of joint hiber- nation is likely to vary with elevation, thus correlating with the pattern observed by Barash (1974a, 1989) and Armitage (1981, 1988) between length of growing season and degree of sociality.
I am not arguing that delayed dispersal is unimpor- tant in the evolution of social behavior in marmots and ground squirrels in general. The trend remains that spe- cies of ground-dwelling sciurids that have delayed dis- persal tend to be more social than those species that do not have delayed dispersal (Armitage 1981). How- ever, the problem that also needs to be addressed is why in those species that have an equal amount of time in which mothers interact with at least some of their immature female offspring, some of those species (e.g. M. flaviventris and M. olympus) extend social behavior to among at least some adult females whereas other spe- cies (e.g.M. monax) do not continue social behavior into adulthood.
Acknowledgements. I am grateful to Vere Smith and A. Peter and Virginia Smith for allowing me to use their property as study sites. [ thank the members of my dissertation committee, T. Koertve- lyessy, S. Moody, E. Peterson, J. Rovner and especially my advisor G. Svendsen for their guidance during the study. Scott Moody, Carolyn Renzulli and my wife Deborah provided encouragement. I also wish to thank the three anonymous reviewers for their helpful comments on an earlier draft of the manuscript. The research was supported in part by grants to the author from the Society of Sigma Xi, the Theodore Roosevelt Memorial Fund of the American Museum of Natural History, the John Houk Memorial Grant of the Ohio University Graduate Student Senate, and the Department of Zoological and Biomedical Sciences, Ohio University.
References
Anthony M (1962) Activity and behavior of the woodchuck in southern Illinois. Occas Pap CC Adams Center Ecol Studies 6 : 1-25
Armitage KB (1962) Social behavior of a colony of the yellow- bellied marmot (Marmota flaviventris). Anita Behav 10:319- 331
Armitage KB (1981) Sociality as a life-history tactic of ground squirrels. Oecologia 48 : 36-49
Armitage KB (1984) Recruitment in yellow-bellied marmot popula- tions: kinship, philopatry and individual variability. In: Murie JO, Michener GR (eds) Biology of ground-dwelling squirrels. Univ of Nebraska Press, Lincoln, pp 375-403
Armitage KB (1986) Marmot polygyny revisited: determinants of male and female reproductive strategies. In: Rubenstein DS, Wrangham RW (eds) Ecology of social evolution: field studies of birds and mammals. Princeton Univ Press, Princeton, pp 303-331

400
Armitage KB (1988) Resources and social organization of ground- dwelling squirrels. In: Slobodchikoff CN (ed) The ecology of social behavior. Academic Press, New York, pp 131-155
Armitage KB, Downhower JF (1974) Demography of yellow-bel- lied marmot populations. Ecology 55 : 1233-1245
Armitage KB, Johns DW (1982) Kinship, reproductive strategies and social dynamics of yellow-bellied marmots. Behav Ecol Sociobiol 11:55-63
Armitage KB,Downhower JF, Svendsen GE (1976) Seasonal changes in the weights of marmots. Am Midl Nat 96:36-51
Arnold W (1990a) The evolution of marmot sociality: I. Why dis- perse late? Behav Ecol Sociobiol 27:229-237
Arnold W (1990b) The evolution of marmot soeiality: II. Costs and benefits of joint hibernation. Behav Ecol Sociobio127 : 239- 246
Barash DP (1973) The social behavior of the Olympic marmot. Anita Behav Monogr 6:171-249
Barash DP (1974a) The evolution of marmot societies: a general theory. Science 85 : 415-420
Barash DP (1974b) The social behavior of the ho'ary marmot (Mar- mota caligata). Anita Behav 22:256 261
Barash DP (1989) Marmots, social behavior and ecology. Stanford Univ Press, California
Bronson FH (1963) Some correlates of interaction rates in natural populations of woodchucks. Ecol 44:637-644
Bronson FH (1964) Agonistic behavior of woodchucks. Anita Be- hay 12:470-478
Brown JL, Orians GH (1970) Spacing behavior in mobile animals. Annu Rev Ecol Syst 1:239-262
Davis DE (1981) Mechanism for decline in a woodchuck popula- tion. J Wildl Manage 45:658-668
Downhower JF, Armitage KB (1981) Dispersal of yearling yellow- bellied marmots (Marmotaflaviventris). Anita Behav 29:1064- 1069
Elliot PF (1975) Longevity and the evolution of polygamy. Am Nat 109:281-287
Ferron J, Quellet JP (1989) Temporal and intersexual variations in use of space in regard to social organization in the wood- chuck (Marmota monax), Can J Zool 67:1642-1649
Grizzell RA (1955) A study of the southern woodchuck Marmota monax monax. Am Midl Nat 53:257-293
Hamilton WJ (1934) The life history of the rufescent woodchuck. Ann Carnegie Mus 23:85-178
Meier PT (1985) Behavioral ecology, social organization and mat- ing system of woodchucks (Marmota monax) in southeast Ohio. Ph D Diss, Ohio Univ, Athens, Ohio
Michener GR (1983) Kin identification, matriarchies, and the evo- lution of sociality in ground dwelling sciurids. In: Eisenberg J, Kleiman D (eds) Recent advances in mammalian behavior. (Spec Publ Amer Soc Mammal 7) pp 528-571
Snyder RL, Christian JJ (1960) Reproductive cycle and litter size in the woodchuck. Ecology 41 : 647-656
Snyder RL, Davis DE, Christian JJ (1961) Seasonal changes in the weights of woodchucks. J Mammal 42:297-312
Vos A de, Gellespie DI (1960) A study of woodchucks on an Ontar- io farm. Can Field Nat 74:140-145