sister-chromatid exchange formation at supra-optimal growth temperatures
TRANSCRIPT

Mutation Research, 108 (1983) 293-299 293 Elsevier Biomedical Press
Sister-chromatid exchange formation at supra-optimal growth temperatures
Pablo Hernhndez and Crisanto Guti6rrez * lnstituto de Biologia Celular, Veldzquez-144. Madrid-6 (Spain)
(Received 5 August 1982) (Accepted 29 October 1982)
Summary
The present research was mainly focused on characterizing the formation of sister-chromatid exchanges at both optimal and supra-optimal growth temperatures. Under these conditions (25, 30 and 35°C) meristem cells of Al l ium cepa L. exhibited a roughly constant cell-cycle time, and modifications in other cell-cycle parameters were negligible. Second-division chromosomes of cells incubated at 30 and 35°C showed increased SCE yields as compared with those detected in cells maintained at the optimal temperature (25°C). When cells with unifilarly BrdUrd-substituted DNA were damaged by irradiation with visible light, we obtained almost the same SCE yields at the various temperatures tested. We suggest that this production of SCEs could be the result of a reduced number of lesions produced by light (perhaps as a consequence of reduced intracellular free oxygen at high temperature) a n d / o r of an increased efficiency in the repair capacity of the cells at these temperatures.
The analysis of SCE formation in undamaged cells incubated at different temper- atures during BrdUrd treatment has shown that the optimal growth temperature appears to be the experimental condition in which the cells are able to exhibit the lowest frequency of SCE.
During the last few years, the use of the sister-chromatid exchange (SCE) test has become one of the most widespread and useful techniques for screening DNA- damaging agents with potential mutagenic a n d / o r carcinogenic action (Latt et al., 1981). However, in spite of much work carried out on the SCE phenomenon (Latt,
* To whom correspondence should be addressed.
Abbreviations: BrdUrd, bromodeoxyuridine; dThd, thymidine; FdUrd, fluorodeoxyuridine; SCE, sister- chromatid exchange; Urd, uridine.
0027-5107/83/0000-0000/$03.00 © Elsevier Science Publishers

294
1981; Wolff, 1982; Sandberg, 1982), many questions about the exact nature of SCEs remain open.
A recent approach involves the analysis of the effect of growth temperature on SCE formation by using different cell types: Chinese hamster (Kato, 1980; Speit, 1980), Xenopus laevis (Speit, 1980), Allium cepa (Gutirrrez et al., 1981) and human lymphocytes (Abdel-Fadil et al., 1982). Moreover, the effect of hyperthermia alone or in combination with radiation, usually provoking heat shocks in the cells, has been studied by applying different methods coming from cytogenetics, biochemistry, radiation research, etc. in order to improve techniques useful in cancer therapy (Streffer et al., 1978; Livingstone and Dethlefsen, 1979; Dikomey, 1981).
Root meristem cells of many plants have the ability to grow over a wide range of temperature; e.g., adventitious roots of onion bulbs can grow between 5 and 35°C, 25°C being the optimum as determined by the root growth rate (L6pez-Shez et al., 1969). In a previous work we analyzed the effect of sub-optimal growth temperatures on the incidence of SCE (Gutirrrez et al., 1981). Under those conditions, the lowering of the culture temperature to 5°C led to an 8-fold increase in the total cell-cycle duration, but maintained a constancy in the relative lengths of cell-cycle phases as measured by the steady frequencies of the cell-cycle compartments G 1, S, G 2 and the mitotic phases (Gonzhlez-Fern~ndez et al., 1971). The aim of the present research was to analyze the formation of SCEs under supra-optimal growth tempera- tures, where both root growth and cycle time appear to be roughly constant (L6pez-S&ez et al., 1966, 1969), in an attempt to ascertain factors affecting SCE production other than those directly related to modifications in cell-cycle parame- ters.
Material and methods
Root meristem cells of Allium cepa L. were used. Onion bulbs were grown in filtered tap water at a constant temperature, and were continuously aerated by bubbling at 20 ml air /min. After the roots reached 10-15 mm in length, the tap water was replaced with BrdUrd medium containing 1 0 - a M BrdUrd, 1 0 -9 M FdUrd and 10 -6 M U r d . This treatment lasted for 40 h (two cycles), or 20 h (one cycle under our conditions). When the second cycle occurred in the absence of BrdUrd, the roots were washed with tap water and maintained for 20 h in a BrdUrd-free solution containing l0 -4 M dThd and 10 -6 M Urd.
Visible-light irradiations were carried out by exposing roots, submerged in tap water, to 1.68 x 10 -7 J / / m 2 from a Philips incandescent lamp (light fluence = 700 j /m2/sec , as determined by a Yellow Spring Instr. Model 65A Radiometer). During irradiation, temperature changes were prevented by changing the tap water every 10 min; the onions were exposed to light from the side and were continuously rotated to ensure a uniform illumination of each meristem.
After the 2-cycle treatment, the roots, still attached to the bulbs, were treated with 0.05% colchicine for 2.5 h. Then, the last 2 cm of the roots were cut off and transferred to an ethanol-acetic acid (3: 1) fixative solution overnight. Finally, a
295
modified fluorescent-plus-Giemsa (FPG) staining technique was carried out (Schvartzman and Guti~rrez, 1980). After the scoring of 200-300 chromosomes mean SCE values per second-division chromosome were calculated, and the results were only taken into account when the observed distribution of chromosomes with different numbers of SCEs fitted the theoretical Poisson distribution (calculated from the experimental mean values) according to the X 2 test. Then, 95% confidence limits of the mean were estimated.
Results
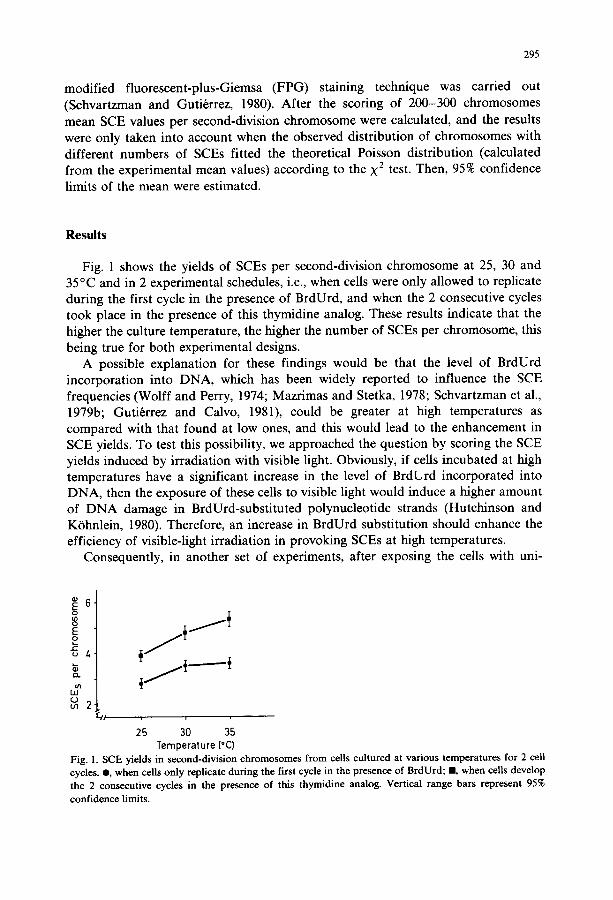
Fig. 1 shows the yields of SCEs per second-division chromosome at 25, 30 and 35°C and in 2 experimental schedules, i.e., when cells were only allowed to replicate during the first cycle in the presence of BrdUrd, and when the 2 consecutive cycles took place in the presence of this thymidine analog. These results indicate that the higher the culture temperature, the higher the number of SCEs per chromosome, this being true for both experimental designs.
A possible explanation for these findings would be that the level of BrdUrd incorporation into DNA, which has been widely reported to influence the SCE frequencies (Wolff and Perry, 1974; Mazrimas and Stetka, 1978; Schvartzman et al., 1979b; Guti6rrez and Calvo, 1981), could be greater at high temperatures as compared with that found at low ones, and this would lead to the enhancement in SCE yields. To test this possibility, we approached the question by scoring the SCE yields induced by irradiation with visible light. Obviously, if cells incubated at high temperatures have a significant increase in the level of BrdUrd incorporated into DNA, then the exposure of these cells to visible light would induce a higher amount of DNA damage in BrdUrd-substituted polynucleotide strands (Hutchinson and K/3hnlein, 1980). Therefore, an increase in BrdUrd substitution should enhance the efficiency of visible-light irradiation in provoking SCEs at high temperatures.
Consequently, in another set of experiments, after exposing the cells with uni-
6. o
E o
JE: u /.
~ 2
25 30 35 Temperature (°C)
Fig. 1. SCE yields in second-division chromosomes from cells cultured at various temperatures for 2 cell cycles. O, when cells only replicate during the first cycle in the presence of BrdUrd; m, when cells develop the 2 consecutive cycles in the presence of this thymidine analog. Vertical range bars represent 95% confidence limits.

296
TABLE 1
SCE YIELDS AT VARIOUS TEMPERATURES AFTER VISIBLE LIGHT (VL) IRRADIATION BETWEEN THE FIRST CYCLE (IN THE PRESENCE OF BrdUrd) AND THE SECOND CYCLE (IN THE ABSENCE OF THIS THYMIDINE ANALOG)
Temperature SCEs per chromosome Increase in SCE (°C) after VL
Non-irradiated Irradiated
25 2.75 + 0.19 a 4.30 + 0.30 1.55 30 3.48 + 0.23 3.70 + 0.25 0.22 35 3.62 + 0.23 3.99 + 0.22 0.37
a Mean+95% confidence limits.
filarly BrdUrd-substituted D N A to an equal dose of visible light (1.68 × 107 j /m2) , we determined the frequencies of SCEs at various temperatures. In every experiment, and based on previous findings (Schvartzman et al., 1979a; Schvartzman and Guti~rrez, 1980), the exposure to light took place in the middle of the 2-cycle treatment, namely, during the non-replicative period located between the first and second D N A replication rounds while the irradiation temperature was the same as that of the culture. As shown in Table 1, we obtained a roughly constant production of SCEs after exposure to visible light. Thus, we conclude that an increase in the level of BrdUrd substitution of DNA, if it exists, does not appear to be the major factor responsible for the SCE formation at high temperatures, which is in agree- ment with previous suggestions (Speit, 1980).
Discussion
Temperature dependence of SCEs in BrdUrd-treated cells The observation that cells incubated at supra-optimal temperatures for 2 consecu-
tive cycles in the presence of BrdUrd elicited increased SCE frequencies is in agreement with previous findings (Speit, 1980; Kato, 1980; Abdel-Fadil et al., 1982). In the present paper, we report a similar SCE response in second-division chro- mosomes of Allium cepa L. meristem cells that have developed only their first cycle
in the presence of BrdUrd (Fig. 1). One explanation put forward to explain the temperature dependence of SCEs
involves the accumulation of DNA damage at high temperature (Kato, 1980). Although the possible involvement of thermal damage in SCE production at high temperatures cannot be presently discarded, the fact that both root growth and cycle parameters remain roughly constant between 25 and 35°C is an indication that the treatments must be considered within a physiological range. Thus, thermal damage, if it actually occurs at these temperatures, is not expected to make an important contribution to this SCE response, as was also previously suggested (Speit, 1980).
Therefore, it seems logical to correlate these SCEs with the D N A replication process, since sister-chromatid exchange formation in damaged cells has recently

297
been related to alterations in the DNA replication pattern (Painter, 1980). In other plant material (Helianthus roots), which alsoshows an ability to grow between 10 and 38°C, it has been well documented that meristem cells exhibited distinct chromosome replication patterns at different temperatures (Van 't Hof et al., 1978). Based on these observations, we postulate that the results shown in Fig. 1 for undamaged cells could be mainly due to a given modification in the chromosome- replication pattern.
Temperature independence of SCEs induced by visible light The SCE response obtained after irradiation with light at various temperatures
has to be explained in terms of the relationship between DNA damage and chromosome replication. We found that the increases in SCE frequencies in irradia- ted cells are significantly lower at 30 and 35°C, with respect to 25°C (Table 1). This situation is different from that previously observed at temperatures below 25°C, where roughly the same increases were detected at all the temperatures tested (Guti6rrez et al., 1981). Therefore, it seems that the ability of an equal dose of visible light to induce SCEs is significantly reduced when cells are incubated under supra-optimal temperatures at which roots can still grow properly.
The failure of light exposure to induce SCEs may be the result of a reduced number of DNA lesions present at the time of chromosome replication (Schvartz- man and Guti6rrez, 1980). Assuming that irradiation leads to the same initial amount of DNA damage, mainly uracilyl radicals (Hutchinson and K6hnlein, 1980), there exist at least 3 possible explanations. Firstly, the light-induced radicals, eventually leading to DNA lesions and SCEs, might be cured rapidly through a non-enzymatic process, since the amount of intracellular free oxygen, known to
~ 6-
E o
~ 4. e~
u 113
2.
/ 0 " , ,% I 0
I
d
-1.00
-0.75 ~:
e-
-0.50 o CB
¢Y
0.25
5 15 25 35
Temperature (*C) Fig. 2. The SCE yields obtained in second-division chromosomes of cells incubated in the presence of BrdUrd for the first cycle only (O) or for the 2 consecutive cycles ( i ) , are compared with the root growth rates (©) obtained at different temperatures. SCE data for 5 -25°C are taken from Guti,~rrez et al. (1981), and those for 25 (overlapped point), 30 and 35°C are from the present paper. Vertical bars represent 95% confidence limits. Root-growth rates at different temperatures are taken from L6pez-Shez et al. (1969).

298
enhance SCE frequencies (Gutirrrez and L6pez-Shez, 1982), could be markedly reduced at high temperatures (Garcia-Herdugo et al., 1974). Moreover, SCE yields shown in Table 1 seemed to mimic those obtained when irradiation took place in the presence of 100% nitrogen, which reduces SCE frequencies in irradiated cells down to the control level (Gutirrrez and Schvartzman, 1981). Secondly, the low enhance- ment in SCE frequencies after irradiation with light at high temperatures (Table 1), could otherwise suggest an increase in the repair efficiency of the cells under the conditions tested in the present study. Thirdly, there could be a combination of the previous alternatives, which perhaps is the more feasible explanation.
Growth rate and SCE formation The relationship between certain characteristics of cell growth and SCE yields has
been analyzed by using different cell types and experimental conditions. Snope and Rary (1979) showed that, in human lymphocytes, the SCE frequency significantly varied with duration of the cell cycle. More recently, Speit (1980) and Kato (1980) in Chinese hamster cells, Gutirrrez et al. (1981) in Allium cepa meristem cells, and Abdel-Fadil et al. (1982) in human lymphocytes, found that the growth temperature has a clear modulating action on the production of SCEs in undamaged, BrdUrd- treated cells. Moreover, when SCE yields and growth rates in root meristem cells were plotted against a wide range of incubation temperatures (5-35°C), the lowest SCE frequency was found in cells incubated at the optimal culture temperature, as determined, in our experiments, by the highest root-growth rate (Fig. 2). From these results, obtained from cells that had completed, in the presence of BrdUrd, either the first cycle or two consecutive cycles, we conclude that optimal growth appears to be a condition in which the cells are able to exhibit the lowest SCE frequency.
Acknowledgements
We are indebted to Prof. Lrpez-S~tez for helpful comments on the manuscript. The technical assistance of M.L. Martinez and M.C. Partearroyo is also much appreciated. The present research was partly supported by a grant from the 'Comisibn Asesora para la Investigacibn Cientifica y Trcnica', and from a research agreement between CSIC (Spain) and the University of Chile (Chile).
References
Abdel-Fadil, M.R., C.G. Palmer and N. Heerema (1982) Effect of temperature variation on sister-chro- matid exchange and cell-cycle duration in cultured human lymphocytes, Mutation Res., 104, 267-273.
Dikomey, E. (1981) Differential cytotoxic effects of hyperthermia below and above 43°C alone or in combination with X irradiation, Radiat. Res., 88, 489-501.
Garcia-Herdugo, G., J. Hidalgo, R. Molina and J.F. L6pez-S/lez (1974) Effect of aeration on the cell cycle in Allium roots, Caryologia, 27, 143-151.
Gonzalez-Fernandez, A., G. Gim6nez-Martin and C. de la Torre (1971) The duration of interphase periods at different temperatures in onion root tips cells, Cytobiologie, 3, 367-371.

299
Guti6rrez, C., and A. Calvo (1981) Approximation of baseline and BrdU-induced SCE frequencies, Chromosoma, 83, 685-695.
Guti6rrez, C., and J.F. L6pez-Shez (1982) Oxygen dependence of sister-chromatid exchanges, Mutation Res., 103, 295-302.
Guti&rez, C., and J.B. Schvartzman (1981) Sister-chromatid exchanges as a measure of the damage produced by visible light on BrdUrd-substituted chromosomes, in: E. Seeberg and K. Kleppe (Eds.), Chromosome Damage and Repair, Plenum, New York, pp. 567-571.
Guti~rrez, C., J.B. Schvartzman and J.F. L6pez-Sb, ez (1981) Effect of growth temperature on the formation of sister-chromatid exchanges in BrdUrd-substituted chromosomes, Exp. Cell Res., 134, 73-79.
Hutchinson, F., and W. K6hnlein (1980) The photochemistry of 5-bromouracil and 5-iodouracil in DNA, in: F.E. Hahn, H. Kersten, W. Kersten and W. Szybalski (Eds.), Progress in Molecular and Subcellular Biology, Vol. 7, Springer, Berlin, pp. 1-42.
Kato, H. (1980) Temperature-dependence of sister chromatid exchange: an implication for its mechanism, Cancer Genet. Cytogenet., 2, 61-67.
Latt, S.A. (1981) Sister chromatid exchange formation, Annu. Rev. Genet., 15, 11-55. Latt, S.A., J. Allen, S.E. Bloom, A. Carrano, E. Falke, D. Kram, E. Schneider, R. Schreck, R. Tice, B.
Whitfield and S. Wolff (1981) Sister chromatid exchanges: a report of the Gene-Tox program, Mutation Res., 87, 17-62.
Livingstone, G.K., and L.A. Dethlefsen (1979) Effects of hyperthermia and X irradiation on sister chromatid exchange (SCE) frequency in Chinese hamster ovary (CHO) cells, Radiat. Res., 77, 512-520.
L6pez-S~iez, J.F., G. Gim6nez-Martin and A. Gonzidez-Fern~ndez (1966) Duration of the cell division cycle and its dependence on temperature, Z. Zellforsch., 75,591-600.
L6pez-Sb.ez, J.F., G. Gonzhlez-Bernhldez, A. Gonzhlez-Fernhndez and G. Garcia-Ferrero (1969) Effect of temperature and oxygen tension on root growth, cell cycle and cell elongation, Protoplasma, 67, 213-221.
Mazrimas, J.A., and D.G. Stetka (1978) Direct evidence for the role of incorporated BudR in the induction of sister-chromatid exchanges, Exp. Cell Res., 117, 23-30.
Painter, R.B. (1980) A replication model for sister-chromatid exchange, Mutation Res., 70, 337-341. Sandberg, A.A. (Ed.) (1982) Sister-Chromatid Exchange, Liss, New York. Schvartzman, J.B., and C. Guti&rez (1980) The relationship between the cell time available for repair and
the effectiveness of a damaging treatment in provoking the formation of sister-chromatid exchanges, Mutation Res., 72, 483-489.
Schvartzman, J.B., R. Postigo and C. Guti~rrez (1979a) Analysis of visible light-induced sister-chromatid exchanges in 5-bromodeoxyuridine-substituted chromosomes, Chromosoma, 74, 317-328.
Schvartzman, J.B., F. Cortes, A. Gonz/dez-Fernhndez, C. Guti~rrez and J.F. L6pez-Shez, (1979b) On the nature of sister-chromatid exchanges in 5-bromodeoxyuridine-substituted chromosomes, Genetics, 92, 1251-1264.
Snope, A.J., and J.M. Rary (1979) Cell-cycle duration and sister-chromatid exchange frequency in cultured human lymphocytes, Mutation Res., 63, 345-349.
Speit, G. (1980) Effects of temperature on sister-chromatid exchanges, Hum. Genet., 55, 333-336. Streffer, C., D. van Beuningen, F. Dietzel, E. R~ttinger, J.E. Robinson, E. Scherer, S. Seeber and K.R.
Trott (Eds.) (1978) Cancer Therapy by Hyperthermia and Radiation, Urban and Schwartzenberg, Munich/Baltimore.
Van't Hof, J., C.A. Bjerknes and J.H. Clinton (1978) Replicon properties of chromosomal DNA fibers and the duration of DNA synthesis of sunflower root-tip meristem cells at different temperatures, Chromosoma, 66, 161-171.
Wolff, S. (Ed.) (1982) Sister-Chromatid Exchange, Wiley, New York. Wolff, S., and P. Perry (1974) Differential Giemsa staining of sister chromatids and the study of
sister-chromatid exchanges without autoradiography, Chromosoma, 48, 341-353.