short-term cold-exposure does not improve insulin sensitivity in rats
TRANSCRIPT

Comp. Biochem. Physiol. Vol. 117A, No. 2, pp. 231–238, 1997 ISSN 0300-9629/97/$17.00Copyright 1997 Elsevier Science Inc. PII S0300-9629(96)00254-X
Short-Term Cold-Exposure Does Not Improve InsulinSensitivity in Rats
Elisabeth Agosto, Maria Cimmino, Yves Minaire, and Alain GeloenUMR 5578-CNRS, Laboratoire de Physiologie des Regulations Energetiques, Cellulaires et Moleculaires,
Lyon, France
ABSTRACT. Effects of noradrenergic activation induced by short-term cold-exposure (7 days at 4°C) on whole-body glucose utilization and tissue glucose uptake were investigated in rats. Measurements were realized onanesthetized normothermic animals at four different levels of insulinemia, within physiological range, allowingcalculation of insulin sensitivity and responsiveness. Whole-body glucose utilization increased as a logarithmicfunction of insulinemias, and was always higher in cold-exposed than in control rats. However, neither insulinsensitivity nor responsiveness, literally, appeared different between the two groups. In the diaphragm, the onlystudied working muscle, glucose uptake was largely higher than in resting muscles. At basal insulin concentration,glucose uptake was higher in cold-exposed than in control rats and increased in the two groups with insulinemia.Among resting muscles, glucose uptake was increased by previous cold exposure in gastrocnemius, soleus, andtibialis. However, insulin sensitivity and responsiveness were found augmented only in the two former. In inter-scapular brown adipose tissue, glucose uptake was largely higher in cold-exposed than in control rats, but nodifference could be evidenced in insulin sensitivity or responsiveness. In white adipose tissues, glucose uptakeincreased with insulinemia. Insulin responsiveness and sensitivity were higher only in the retroperitoneal depot.comp biochem physiol 117A;2:231–238, 1997. 1997 Elsevier Science Inc.
KEY WORDS. BAT, cold exposure, euglycemic-hyperinsulinemic clamp, glucose uptake, insulin respon-siveness, insulin sensitivity, muscles, WAT
INTRODUCTION duced by short-term cold-exposure (4°C, 7 days) on insulinsensitivity. In the present study, whole-body glucose utiliza-
A decreased sympathetic nervous activity in diseases char-tion, as well as tissue glucose uptake and insulin sensitivity,
acterized by insulin resistance, such as obesity and NIDDM,were studied in rats reared either at neutral or cold ambient
has often been noted (7,10). Nevertheless, the hypothesistemperature. Measurements were made under euglycemic-
of a noradrenergic modulation of insulin sensitivity remainshyperinsulinemic clamp, with injection of 2[3H]-deoxyglu-
poorly investigated in vivo for two main reasons. Firstly, thecose and [14C] saccharose, in anesthetized normothermic
difficulty to test the metabolic effects of noradrenaline liesrats at neutral ambient temperature. Graded insulin infu-
in the fact that it is mainly a neurotransmitter. Therefore,sion rates were used to establish the dose-response curvesa great quantity of exogenous noradrenaline is required toneeded to calculate insulin sensitivity in the physiological
mimic physiologic stimulation of the sympathetic system atrange of plasma insulin concentrations. Following Bergman
the synaptic sites. This problem can be solved in using ex-et al. (1), we considered that during euglycemic clamp the
perimental conditions known to enhance sympathetic ner-‘‘initial slope’’ of the relationship between insulinemia and
vous activity, such as cold-exposure (6). Secondly, thean insulin-related effect is the best index of insulin sensi-
complete evaluation of insulin sensitivity requires the estab-tivity.
lishment of a dose-response curve between plasma insulinSince the slopes of the dose-response curves between
concentration and some measurable insulin-dependentplasma insulin concentrations and whole body glucose utili-
metabolic process (11,16).zation were similar, our findings show that short-term cold-
To test the hypothesis of a noradrenergic modulation ofexposure does not improve whole-body insulin sensitivity.
insulin sensitivity we undertook to measure the persistenteffects of the sustained increased noradrenaline release in-
MATERIALS AND METHODSAddress reprint requests to: A. Geloen, PhD, Laboratoire de Physiologie desAnimalsRegulations Energetiques, Cellulaires et Moleculaires, UMR 5578-CNRS,
Faculte de Medecine Lyon-Nord, 8 avenue Rockefeller, 69373 LYONMale Sprague-Dawley rats (Iffa-Credo, L’Arbresle, France)Cedex 08, France. Tel. 010 (33) 78 77 72 83; Fax 010 (33) 78 77 71 75.
Received 30 November 1995; accepted 14 July 1996. weighing 180–220 g, were maintained in animal quarters

232 E. Agosto et al.
under controlled lighting (light on from 7.00 to 19.00 hr) the difference between the two GIR60–120 measured at thelowest and the highest insulinemias (11). Insulin whole-and had free access to laboratory chow (A03; UAR, Epinay/
Orge, France) and water. They were divided into two body sensitivity was calculated as the ‘‘initial slope’’ of theregression line between GIR60–120 and insulinemias in thegroups. One group (controls, n 5 44) was maintained at
thermoneutral temperature (25 6 1°C), whereas the other physiological range, i.e., below 100 mU/l (1).(cold-exposed, n 5 39) was exposed for 7 days to the cold(4 6 1°C).
Glucose Metabolic Index
Glucose uptake by individual tissues was estimated as ‘‘tissueExperimental Protocol glucose metabolic index’’ (13). Briefly, 45 min before the
end of the clamp, the glucose analogue 2-[1,2-3H]-deoxy-d-CONTROL ANIMALS WERE CANNULATED AT 200–220 G.
glucose (32.5 µCi; Isotopchim, Ganagobie-Peyruis, France)Cold-exposed animals were cannulated before cold-expo-and [U-14C]-saccharose (4 µCi; Du Pont de Nemours, Lessure at 180–200 g. Two polyethylene catheters (EO 3403;Ulis, France) were administered i.v. together as a bolus, inBiotrol, Villeron, France) were inserted under halothane250 µl of saline solution containing 0.25% albumin. Plasmaanesthesia, one into the right jugular vein, and the othersamples (50 µl) were collected at 0, 2, 5, 7, 10, 15, 20, 30,into the left carotid. They were exteriorized through an in-and 45 min after the administration of the tracer bolus forcision in the nape of the neck and filled with citrate-salineestimation of a plasma radioactivity disappearance curve.(24.5 mM of tri-Sodium Citrate dihydrate in 0.9% saline).At the end of the clamp, rats were killed by cervical disloca-The carotid catheter was used for blood sampling and thetion and tissues were rapidly removed: diaphragm, musclesjugular for intravenous infusion. All studies were conductedof the hindlimb (extensor digitorum longus, tibialis, soleus,at least 4–5 days after surgery to allow the animals to over-plantaris and gastrocnemius), white adipose tissues (epidid-come the surgical stress and to adapt to the cannulation.ymal and retroperitoneal WAT), and interscapular brownAll animals were deprived of food for 5 hr before the study.adipose tissue (IBAT). These tissues were immediatelyCold-exposed rats were removed from the cold 30 min be-weighed and frozen for storage.fore the beginning of the clamp.
For subsequent analysis, tissues and plasma samples wereAll animals were anesthetized with an intraperitoneal in-solubilized in 2 ml potassium hydroxide (3%) at 65°C in ajection of sodium pentobarbital (30 mg/kg BW). Body tem-shaking water bath for 2 hr. To reduce quenching, 100 µlperature was maintained at 38°C with a heating plate andof hydrogen peroxide (30%) was added to the vials whichmonitored throughout the experiment.were kept at 65°C for one additional hour. Ten milliliters ofscintillant (Insta-Gel, Packard) were then added and double
Euglycemic-hyperinsulinemic Clamp Studies isotope counting performed in a scintillation counter (SL3000 Intertechnic, Kontron). Quench corrections were ap-Euglycemic-hyperinsulinemic clamps were carried out ac-plied on an external standard. The extracellular volume ofcording to Kraegen et al. (12). Rats were infused with hu-each tissue sample was estimated using 14C-saccharose andman neutral insulin (Actrapid HM, 40 U/ml; Novo Indus-was used to calculate the intracellular 2DG-6-phosphatetrie, Denmark) while maintaining blood glucose at the basalcontent (C*m (T) dpm/g at T 5 45 min), according to Homlevel (6 mM). Preliminary measures on awake, fasting unre-et al. (8). Glucose uptake (ng glucose/mg⋅min) was thenstrained animals showed that basal plasma glucose levelscalculated as ‘‘tissue glucose metabolic index’’, R′g (Kraegenwere 6 mM (6.0 6 0.1 mM n 5 12). Insulin was continu-et al., 1985):ously infused at either 2, 4, 6, or 10 mU/kg ⋅ min (in 0.9%
saline and 1% bovine albumin, fatty acid free) at 16.6 µl/R′g 5
C*m (T)
#T
0(C*p /Cp)
dtmin for 2 hr to achieve steady plasma insulin concentra-tions. Blood samples of 25 µl were taken at 5-min intervalsto determine glycemia (Glucose Analyser II, Beckman), and100 µl of blood was taken at 0, 60, and 120 min of the where C*p is plasma 3H-2DG (dpm/ml) and Cp is plasmaclamp for insulin measurements by a double-antibody radio- glucose (µg/ml). R′g insulin responsiveness was calculated asimmunoassay (CIS Biointernational, Gif/Yvette, France). the difference between the two R ′g values measured at theHematocrits were also determined on 25 µl samples at 0 lowest and the highest insulinemias. R′g insulin sensitivityand 120 min of the clamp. To maintain euglycemia, glucose was calculated as the slope of the regression line betweenat different concentrations (5–40%) was infused at variable R′g and logarithm of insulinemias.rates using a peristaltic roller pump, the delivery rate ofwhich was controlled after each clamp.
Hepatic Glucose Output (HGO)This needed glucose infusion rate plateaued between 60and 120 min (GIR60–120 in mg glucose/kg ⋅ min) and was A study was conducted on 19 cannulated rats. Experimental
conditions were exactly the same as in hyperinsulinemic-considered as a measure of the whole-body glucose utiliza-tion. Insulin whole-body responsiveness was calculated as euglycemic clamp procedure. Animals were exposed to

Glucose Uptake in Cold-Adapting Rats 233
warm or cold environment and divided into two groups, Plasma insulin levels reached during the second hour ofthe clamp were not significantly different, for a given insulinsubmitted or not to a 6 mU/kg ⋅ min hyperinsulinemic-
euglycemic clamp. Rats were removed from the cold 30 min infusion rate, in control and cold-exposed animals. Insulininfusion at 2 mU/kg ⋅ min did not significantly change thebefore experiments and anesthetized. Labeled glucose (d-
[6-3H]-glucose; Dositek, Orsay, France) was administrated plasma insulin levels either in control or in cold-exposedrats. In the two groups of rats, further increases in insulinas a 40 µCi bolus at t0, followed by continuous infusion at
a rate of 0.9 µCi/min for 2 hr. Glucose specific activity was infusion rates to 4, 6, and 10 mU/kg ⋅ min resulted, respec-tively, in a 2-, 3-, and 6-fold increase in plasma insulindetermined on 100 µl plasma samples every 15 min during
the second hour. levels.In the two groups of rats, GIR60–120 increased as a nearlyThe specific activity plateau was used to calculate HGO
at the basal state or to measure effect of hyperinsulinemia linear function of insulin infusion rates (Fig. 1A) and as alogarithmic function of insulinemias between 33 and 210on HGO.
Plasma [6-3H]-glucose radioactivity was measured on the mU/l (Fig. 1B), the values being always higher in cold-exposed than in control rats.supernatants of barium hydroxide-zinc sulfate precipitates
of plasma samples after evaporation to dryness. Insta-Gel Insulin responsiveness was calculated in the two groupsof rats as the difference in GIR60–120 obtained at the lowest(Packard) was added and samples were counted for 10 min
in a scintillation counter. To determine concentration of and the highest insulinemias, i.e., 33 and 210 mU/l (con-trols) and 35 and 178 mU/l (cold-exposed rats). The ob-perfused tracer accurately, three aliquots of each infusate
were run through the same procedure and counted together tained values, 34.9 and 33.6 mg/kg ⋅ min for control andcold-exposed rats, respectively, were not significantly dif-with the plasma samples.
The difference between glucose appearance, as deter- ferent.Insulin sensitivity was estimated as the ‘‘initial slope’’ ofmined by the tracer method, and the exogenous glucose in-
fusion rate, represents the residual endogenous glucose pro- the regression lines of GIR60–120 in the range of physiologicalinsulinemias, i.e., lower than 100 mU/l (Fig. 1B). The val-duction.ues obtained 47.42 and 50.29 mg ⋅ L/kg ⋅ min ⋅ mU forcontrol and cold-exposed rats, respectively, were not sig-
Plasma Noradrenaline and Adrenaline Concentrations nificantly different.Blood samples were collected from rats in conditions exactlyidentical as these of rats undergoing hyperinsulinemic-
Glucose Metabolic Index in Individual Tissueseuglycemic clamp. Plasma samples were kept at 280°C un-The glucose metabolic index (R′g) was considered as repre-til determination. Plasma were mixed with ice-cold perchlo-sentative of glucose uptake in different types of muscles. Itsric acid, centrifuged at 4°C, then catecholamines were puri-variations in response to increased plasma insulin concen-fied by chromatography on alumina and assayed by reversed-trations are given in Fig. 2.phase HPLC with electro-chemical detection (22).
Diaphragm, the only working muscle during the presentexperimental conditions, showed the highest R′g. At basal
Statistical Analysis plasma insulin concentrations, R ′g was significantly higherin cold-exposed than in control rats reared at thermoneu-All experimental data are expressed as means 6 SEM. Thetrality. R′g increased with the logarithm of plasma insulinvalues were examined using two-way analysis of varianceconcentration, the slope of the regression line being steeper(ANOVA). A Fisher PLSD post-hoc test was used for groupin control than in cold-exposed rats, suggesting a cold-comparisons. Statistical significance was set at P , 0.05.induced reduction in insulin sensitivity.
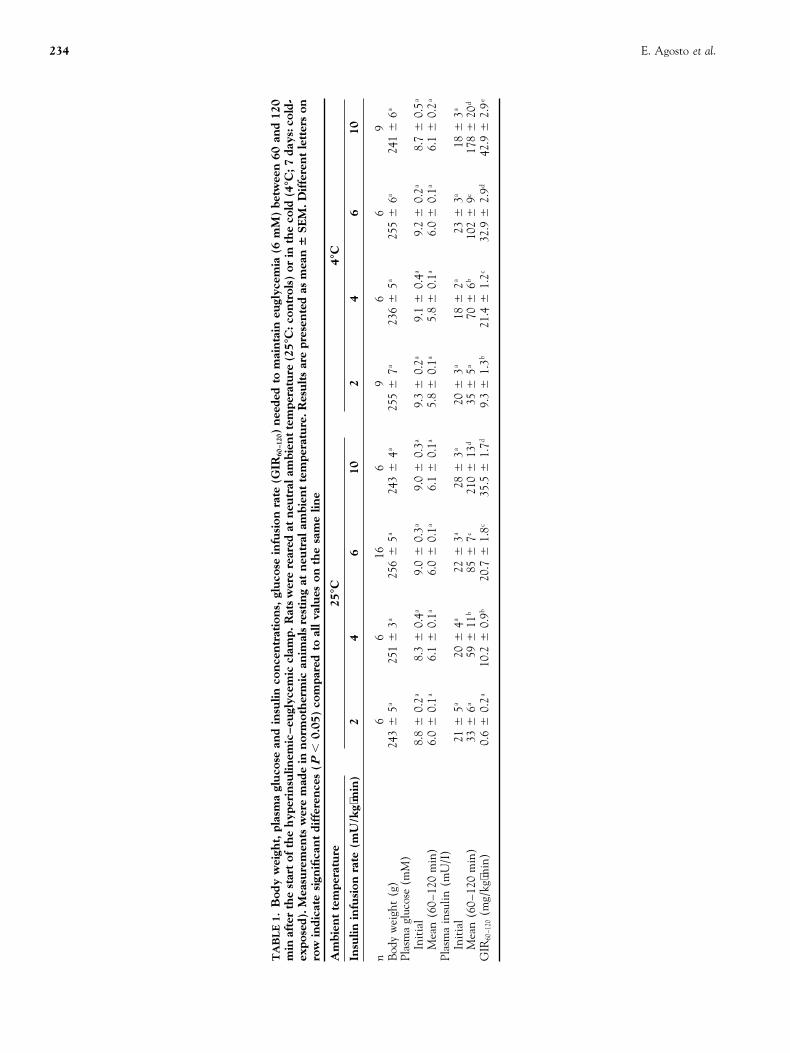
R′g was comparable in all resting muscles (soleus, gastroc-RESULTS nemius, tibialis, EDL, plantaris). At basal plasma insulinBody Weight, Basal Plasma Glucose and Insulin concentrations, R′g was significantly higher in tibialis muscleConcentrations (Table 1) from cold-exposed than from control rats. In all the studied
muscles, values increased with the logarithm of plasma insu-Body weight, basal plasma glucose, and insulin concentra-lin concentration, but the slopes of the regression lines weretions, in 5-hr fasted animals, were not significantly affectedsignificantly steeper (P , 0.05) in cold-exposed than inby previous cold-exposure (4°C, 7 days).control rats only in soleus and gastrocnemius. This suggeststhat insulin sensitivity is increased by previous cold-expo-
Whole-body Glucose Utilization During Euglycemic- sure in these two resting muscles. In most of the studiedhyperinsulinemic Clamp (Table 1) muscles, insulin responsiveness, i.e., the amplitude of the
R′g response to augmented plasma insulin concentrations,Glucose infusion rates (GIR60–120) needed to maintain eugly-cemia (6 mM) at low insulinemias (,40 mU/l) were higher was not significantly different in control and cold-exposed
rats, except for soleus and gastrocnemius. In these two mus-in cold-exposed than in control rats.
234 E. Agosto et al.
TA
BL
E1.
Bod
yw
eigh
t,pl
asm
agl
uco
sean
din
sulin
con
cen
trat
ion
s,gl
uco
sein
fusi
onra
te(G
IR60
–120
)n
eede
dto
mai
nta
ineu
glyc
emia
(6m
M)
betw
een
60an
d12
0m
inaf
ter
the
star
tof
the
hyp
erin
sulin
emic
–eu
glyc
emic
clam
p.R
ats
wer
ere
ared
atn
eutr
alam
bien
tte
mpe
ratu
re(2
5°C
:con
trol
s)or
inth
eco
ld(4
°C;7
days
:col
d-ex
pose
d).M
easu
rem
ents
wer
em
ade
inn
orm
oth
erm
ican
imal
sre
stin
gat
neu
tral
ambi
ent
tem
pera
ture
.Res
ult
sar
epr
esen
ted
asm
ean
6SE
M.
Dif
fere
nt
lett
ers
onro
win
dica
tesi
gnifi
can
tdi
ffer
ence
s(P
,0.
05)
com
pare
dto
all
valu
eson
the
sam
elin
e
Am
bien
tte
mpe
ratu
re25
°C4°
C
Insu
linin
fusi
onra
te(m
U/k
g⋅m
in)
24
610
24
610
n6
616
69
66
9B
ody
wei
ght
(g)
243
65a
251
63a
256
65a
243
64a
255
67a
236
65a
255
66a
241
66a
Plas
ma
gluc
ose
(mM
)In
itia
l8.
86
0.2
a8.
36
0.4a
9.0
60.
3a9.
06
0.3a
9.3
60.
2a9.
16
0.4a
9.2
60.
2a8.
76
0.5
a
Mea
n(6
0–12
0m
in)
6.0
60.
1a
6.1
60.
1a6.
06
0.1a
6.1
60.
1a5.
86
0.1a
5.8
60.
1a6.
06
0.1a
6.1
60.
2a
Plas
ma
insu
lin(m
U/I
)In
itia
l21
65a
206
4a22
63a
286
3a20
63a
186
2a23
63a
186
3a
Mea
n(6
0–12
0m
in)
336
6a59
611
b85
67c
210
613
d35
65a
706
6b10
26
9c17
86
20d
GIR
60–1
20(m
g/kg
⋅min
)0.
66
0.2
a10
.26
0.9b
20.7
61.
8c35
.56
1.7d
9.3
61.
3b21
.46
1.2c
32.9
62.
9d42
.96
2.9
e
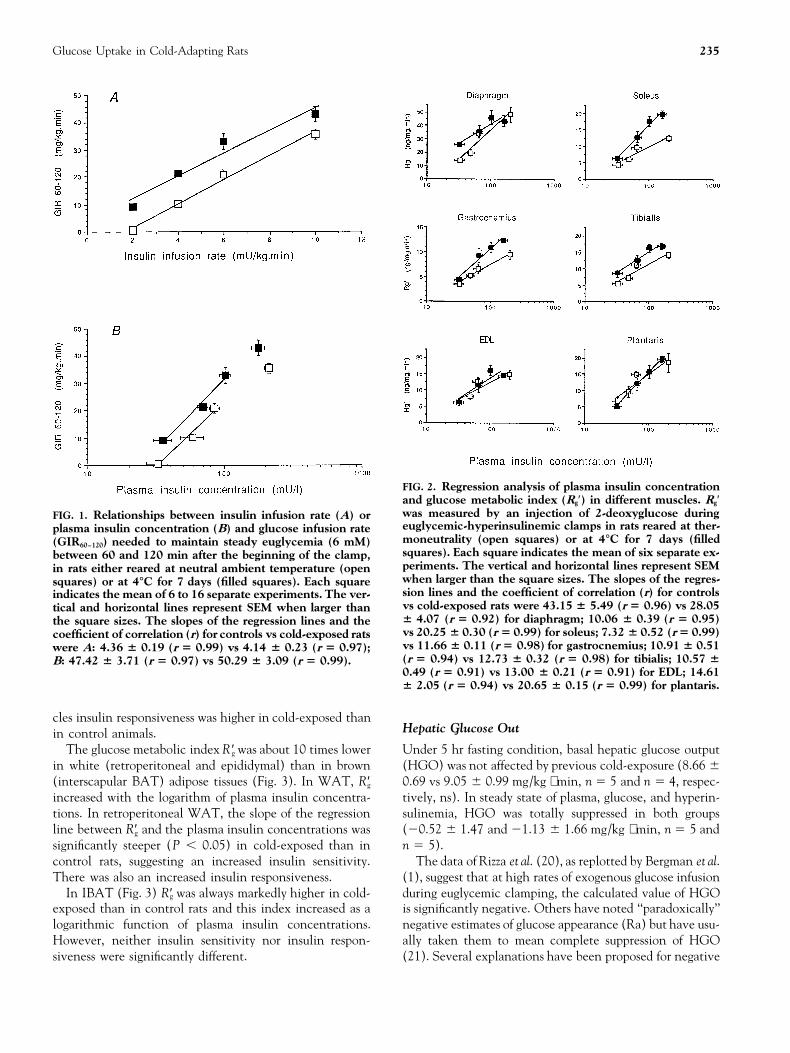
Glucose Uptake in Cold-Adapting Rats 235
FIG. 2. Regression analysis of plasma insulin concentrationand glucose metabolic index (Rg′) in different muscles. Rg′was measured by an injection of 2-deoxyglucose duringFIG. 1. Relationships between insulin infusion rate (A) oreuglycemic-hyperinsulinemic clamps in rats reared at ther-plasma insulin concentration (B) and glucose infusion ratemoneutrality (open squares) or at 4°C for 7 days (filled(GIR60–120) needed to maintain steady euglycemia (6 mM)squares). Each square indicates the mean of six separate ex-between 60 and 120 min after the beginning of the clamp,periments. The vertical and horizontal lines represent SEMin rats either reared at neutral ambient temperature (openwhen larger than the square sizes. The slopes of the regres-squares) or at 4°C for 7 days (filled squares). Each squaresion lines and the coefficient of correlation (r) for controlsindicates the mean of 6 to 16 separate experiments. The ver-vs cold-exposed rats were 43.15 6 5.49 (r 5 0.96) vs 28.05tical and horizontal lines represent SEM when larger than6 4.07 (r 5 0.92) for diaphragm; 10.06 6 0.39 (r 5 0.95)the square sizes. The slopes of the regression lines and thevs 20.25 6 0.30 (r 5 0.99) for soleus; 7.32 6 0.52 (r 5 0.99)coefficient of correlation (r) for controls vs cold-exposed ratsvs 11.66 6 0.11 (r 5 0.98) for gastrocnemius; 10.91 6 0.51were A: 4.36 6 0.19 (r 5 0.99) vs 4.14 6 0.23 (r 5 0.97);(r 5 0.94) vs 12.73 6 0.32 (r 5 0.98) for tibialis; 10.57 6B: 47.42 6 3.71 (r 5 0.97) vs 50.29 6 3.09 (r 5 0.99).0.49 (r 5 0.91) vs 13.00 6 0.21 (r 5 0.91) for EDL; 14.616 2.05 (r 5 0.94) vs 20.65 6 0.15 (r 5 0.99) for plantaris.
cles insulin responsiveness was higher in cold-exposed thanHepatic Glucose Outin control animals.
The glucose metabolic index R ′g was about 10 times lower Under 5 hr fasting condition, basal hepatic glucose output(HGO) was not affected by previous cold-exposure (8.66 6in white (retroperitoneal and epididymal) than in brown
(interscapular BAT) adipose tissues (Fig. 3). In WAT, R′g 0.69 vs 9.05 6 0.99 mg/kg ⋅ min, n 5 5 and n 5 4, respec-tively, ns). In steady state of plasma, glucose, and hyperin-increased with the logarithm of plasma insulin concentra-
tions. In retroperitoneal WAT, the slope of the regression sulinemia, HGO was totally suppressed in both groups(20.52 6 1.47 and 21.13 6 1.66 mg/kg ⋅ min, n 5 5 andline between R′g and the plasma insulin concentrations was
significantly steeper (P , 0.05) in cold-exposed than in n 5 5).The data of Rizza et al. (20), as replotted by Bergman et al.control rats, suggesting an increased insulin sensitivity.
There was also an increased insulin responsiveness. (1), suggest that at high rates of exogenous glucose infusionduring euglycemic clamping, the calculated value of HGOIn IBAT (Fig. 3) R′g was always markedly higher in cold-
exposed than in control rats and this index increased as a is significantly negative. Others have noted ‘‘paradoxically’’negative estimates of glucose appearance (Ra) but have usu-logarithmic function of plasma insulin concentrations.
However, neither insulin sensitivity nor insulin respon- ally taken them to mean complete suppression of HGO(21). Several explanations have been proposed for negativesiveness were significantly different.
236 E. Agosto et al.
values of HGO during euglycemic-hyperinsulinemic clamp:isotope discrimination, inappropriate modeling, large varia-tions of isotopic enrichment.
Plasma Noradrenaline and Adrenaline Content
Plasma noradrenaline contents were 174 6 29 vs 186 6 14pg/ml for control and cold-exposed animals (respectively,ns). Plasma adrenaline contents were 53 6 14 and 37 65.7 pg/ml (respectively, ns).
DISCUSSION
Our findings indicate that increased noradrenergic activa-tion induced by cold-exposure does not improve whole-body insulin sensitivity. Indeed, the slopes of the dose re-sponse curves between plasma insulin concentrations andwhole-body glucose utilization are not different betweencontrol and cold-exposed animals.
The measurement conditions, with control and cold-ex-posed rats, at neutral ambient temperature, were intention-ally chosen so that only persistent metabolic alterationswere taken into account. In our model, sympathetic nervoussystem activity was similar in both control and cold-exposedanimals. Therefore, the effects we observed did not resultfrom stimulatory effect of sympathetic activity on glucoseuptake (14). Since we found that cold-exposure did not en-hance insulin sensitivity when the animals were put backto control conditions, our results suggest that a short-termincreased endogenous noradrenaline release does not persis-tently improve insulin sensitivity. It could be argued thatalterations in glucose metabolism induced by cold disappearvery rapidly when rats return to warm environment, sug-gesting that alterations only occurred under a high sympa-thetic activity. This was shown in conscious cold-accli-mated rats re-exposed at 25°C for 18 h. Indeed, under theseconditions, the net rate of 2-DG uptake returned to controlvalues (28). However, in our study, after 30 min in a warmenvironment, GIR were still significantly increased in cold-exposed animals. Therefore, it cannot be concluded thatthe effects of cold-exposure on glucose tolerance vanishedwhen rats returned to warm conditions. Under these experi-mental conditions, our results suggest that insulin sensitiv-FIG. 3. Regression analysis of plasma insulin concentration
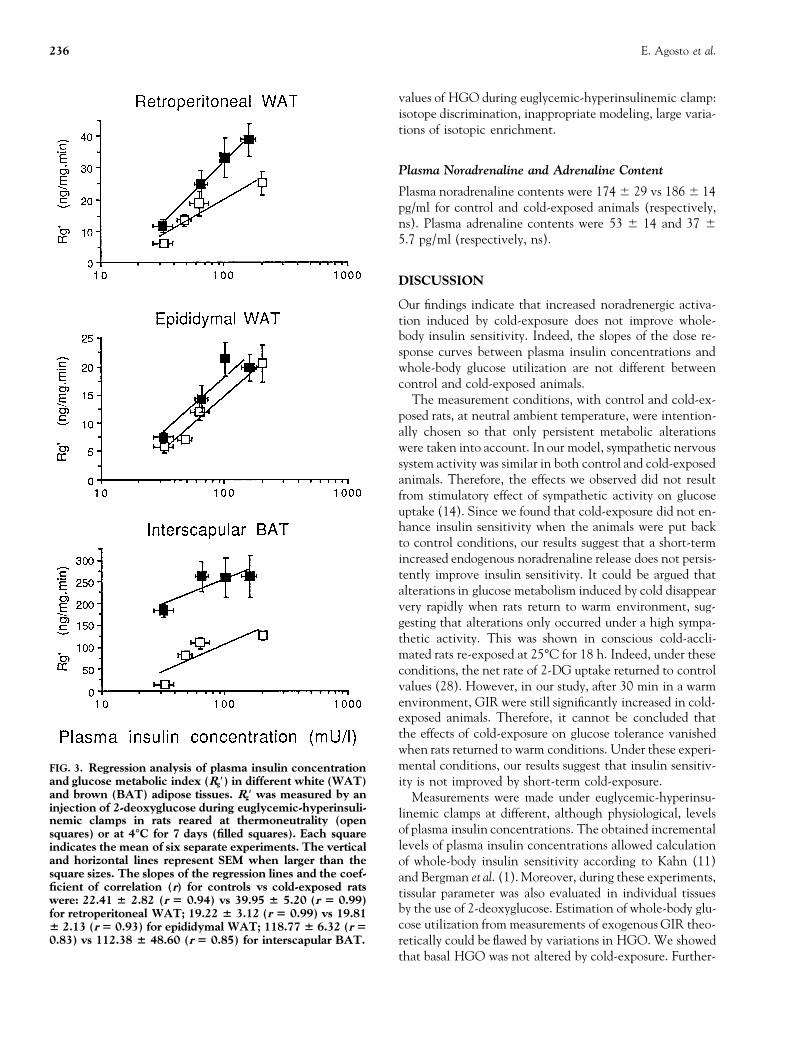
and glucose metabolic index (Rg′) in different white (WAT) ity is not improved by short-term cold-exposure.and brown (BAT) adipose tissues. Rg′ was measured by an Measurements were made under euglycemic-hyperinsu-injection of 2-deoxyglucose during euglycemic-hyperinsuli- linemic clamps at different, although physiological, levelsnemic clamps in rats reared at thermoneutrality (open
of plasma insulin concentrations. The obtained incrementalsquares) or at 4°C for 7 days (filled squares). Each squarelevels of plasma insulin concentrations allowed calculationindicates the mean of six separate experiments. The vertical
and horizontal lines represent SEM when larger than the of whole-body insulin sensitivity according to Kahn (11)square sizes. The slopes of the regression lines and the coef- and Bergman et al. (1). Moreover, during these experiments,ficient of correlation (r) for controls vs cold-exposed rats tissular parameter was also evaluated in individual tissueswere: 22.41 6 2.82 (r 5 0.94) vs 39.95 6 5.20 (r 5 0.99)
by the use of 2-deoxyglucose. Estimation of whole-body glu-for retroperitoneal WAT; 19.22 6 3.12 (r 5 0.99) vs 19.81cose utilization from measurements of exogenous GIR theo-6 2.13 (r 5 0.93) for epididymal WAT; 118.77 6 6.32 (r 5
0.83) vs 112.38 6 48.60 (r 5 0.85) for interscapular BAT. retically could be flawed by variations in HGO. We showedthat basal HGO was not altered by cold-exposure. Further-

Glucose Uptake in Cold-Adapting Rats 237
more, the insulin suppressive effect on HGO, during hyper- tissues are negated by the absence of changes in many oth-ers, resulting in a nearly statu quo at the whole-body level.insulinemic-euglycemic clamp at 6 mU/kg ⋅ min, was iden-
tical between control and short-term cold-exposed rats. In the present study, glucose utilization in the whole animalwas increased at basal insulin concentration, suggesting theThis result is supported by Smith et al. (26) who showed,
both at basal and elevated plasma insulin concentrations, enhancement of a direct non-insulin mediated glucose up-take (NIMGU) (2). The use of 2-DG at basal plasma insulinthat HGO was low in 5-hr fasted rats and not significantly
different in control and cold-acclimated animals. Finally, concentration revealed that the cold effect on NIMGU wasmainly present in brown adipose tissue and to a lesser degreethe rats were purposely kept under anesthesia throughout
the euglycemic-hyperinsulinemic clamp in order to avoid in diaphragm and tibialis muscle. Noradrenaline release bysympathetic nerves is enhanced by cold exposure and it hasany increase in glucose utilization due to muscular tone or
activity. Actually, Penicaud et al. (19) observed that glucose been shown that in the absence of insulin, glucose uptakewas increased in vitro in brown adipocytes by this catechol-utilization by postural muscles was four- to fivefold reduced
by anesthesia. Thus, it was feared that the permanent but amine (15). In addition, noradrenaline increased oxygenconsumption in perfused skeletal muscles from cold-exposedpossibly small increase induced by previous cold exposure
be hidden by the variable and possibly large utilization by rats (24).Insulin mediated glucose uptake exists in addition tophasic or tonic contractions. On another hand, it is known
that pentobarbital anesthesia leads to alteration in glucose NIMGU in insulin-sensitive tissue (heart, skeletal muscles,and adipose tissues) and is activated by cold-exposure orhomeostasis. A transient hyperglycemia and an increased
glucose production were observed by Penicaud et al. (19). exercise (2). In the present study, insulin sensitivity wasincreased in soleus and gastrocnemius muscles and in retro-However, these modifications lasted 40 min, i.e., they were
likely to vanish before the beginning of the GIR measure- peritoneal white adipose tissue from rats previously exposedto cold. Increased insulin sensitivity can be related to manyment (60 min) and the injection of 2-deoxyglucose (75
min). different factors such as an increased tissue blood flow facili-tating the access to insulin receptors (4,9,23).Acute cold-exposure is known to increase glucose turn-
over and clearance rates in rats (5) and in dogs (17,18) in In conclusion, our findings are in agreement with thegenerally accepted increased glucose tolerance in responsewhich little or no change in glucose and insulin concentra-
tions occurs. In eviscerated fasted rats, glucose turnover was to cold-exposure (reviewed in Bukowiecki (2)); neverthe-less, this opinion needs to be further discussed in terms ofthe same in controls and acutely cold-exposed animals in
spite of lower insulinemias in the latter (25). In unanesthe- insulin sensitivity. Using the dose-response methodologyadvocated by Kahn (11), Bergman et al. (1) and more re-tized unrestrained rats a short cold-exposure (5°C, 2 days)
significantly reduced the insulin response to intravenous cently Mikines et al. (16), the present experiment suggeststhat the global cold-induced augmentation in glucose utili-glucose injection while glucose tolerance was improved
(27). Peripheral glucose uptake was increased in anesthe- zation and tolerance is an enduring phenomenon related tothe uneven participation of different tissues. Among them,tized rats previously exposed to cold (5°C, 5 days) while
plasma insulin concentration remained unchanged (3). increased sensitivity and responsiveness can be strictlyclaimed only for soleus and gastrocnemius muscles and ret-Cold acclimation (4°C, 3 weeks), on the other hand, in-
creased substantially (60%) whole-body glucose utilization roperitoneal adipose tissue. Despite the increased insulinsensitivity and responsiveness in some specific tissues, short-in rats under euglycemic-hyperinsulinemic clamp in spite
of comparable plasma insulin concentrations [123 vs 129 term cold-exposure does not significantly improve whole-body insulin sensitivity and responsiveness.mU/l in cold-acclimated and control rats, respectively;
(26)].The transition from whole-body insulin dose-response
This study was supported by the Centre de Recherche en Nutrition Hu-curve to glucose-uptake in individual tissues deserves special maine, Lyon and the Laboratoires Hoechst, Paris. The authors wouldconsideration. In agreement with data previously collected like to thank Dr. T. Pauchard for technical assistance and Dr. J.-M.(3,26), the present study showed that, at any given level of Cottet-Emard for the measures of plasma noradrenaline and adrenaline.plasma insulin concentration within the physiologicalrange, glucose utilization is consistently higher in cold-ex-posed than in control rats. However, no claim for increased Referenceswhole-body insulin sensitivity or responsiveness could be 1. Bergman, R.N.; Finegood, D.T.; Ader, M. Assessment of insu-
lin sensitivity in vivo. Endocr. Rev. 6:45–86;1985.made, at least if these parameters are measured according2. Bukowiecki, L.J. Energy balance and diabetes. The effects ofto Kahn (11) and Bergman (1) through a physiological
cold exposure, exercise training, and diet composition on glu-range of plasma insulin concentrations. Indeed, whole-bodycose tolerance and glucose metabolism in rat peripheral tis-
response to insulin infusions integrates all the individual sues. Can. J. Physiol. Pharmacol. 67:382–393;1989.changes in the different tissues, insulin-sensitive or not. It 3. Cunningham, J.J.; Gulino, M.A.; Meara, P.A.; Bode, H.H.
Enhanced hepatic insulin sensitivity and peripheral glucoseis very possible that the differences which do exist in some

238 E. Agosto et al.
uptake in cold acclimating rats. Endocrinology 117:1585– exercise and detraining on insulin action in trained men. J.Appl. Physiol. 66:704–711;1989.1589;1985.
4. DeFronzo, R.A.; Ferrannini, E.; Sato, Y.; Felig, P.; Wahren, 17. Minaire, Y.; Forichon, J.; Dallevet, G.; Jomain, M.J. Com-bined effects of cold and somatostatin on glucose kinetics inJ. Synergistic interaction between exercise and insulin on
peripheral glucose uptake. J. Clin. Invest. 68:1468–1474; dogs. Eur. J. Appl. Physiol. 46:249–259;1981.18. Minaire, Y.; Forichon, J.; Freminet, A. Effects of endogenous1981.
5. Depocas, F.; Masironi, R. Body glucose as fuel for thermogene- glucagon on glucose kinetics in shivering dogs. J. Appl. Phys-iol. 52:1458–1463;1982.sis in the white rat exposed to cold. Am. J. Physiol. 199:1051–
1055;1960. 19. Penicaud, L.; Ferre, P.; Kande, J.; Leturque, A.; Issad, T.; Gi-rard, J. Effect of anesthesia on glucose production and utiliza-6. Depocas, F.; Behrens, W.A.; Foster, D.O. Noradrenaline-in-
duced calorigenesis in warm- and in cold-acclimated rats: The tion in rats. Am. J. Physiol. 252:E365–E369;1987.20. Rizza, R.A.; Mandarino, L.J.; Gerich, J.E. Dose-response char-interrelation of dose of noradrenaline, its concentration in ar-
terial plasma, and calorigenic response. Can. J. Physiol. Phar- acteristics for effects of insulin on production and utilizationof glucose in man. Am. J. Physiol. 240:E630–E639;1981.macol. 56:168–174;1978.
7. Himms-Hagen, J. Defective thermogenesis in obese animals. 21. Rizza, R.A.; Mandarino, L.J.; Gerich, J.E. Cortisol-induced in-sulin resistance in man: impaired suppression of glucose pro-J. Obes. Weight Regul. 6:179–199;1987.
8. Hom, F.G.; Goodner, C.J.; Berrie, M.A. A (3H)2-deoxyglu- duction and stimulation of glucose utilization due to a postre-ceptor defect of insulin action. J. Clin. Endocrinol. Metab.cose method for comparing rates of glucose metabolism and
insulin responses among rat tissues in vivo. Validation of the 54:131–138;1982.22. Sagnol, M.; Claustre, J.; Cottet-Emard, J.M.; Pequignot, J.M.;model and the absence of an insulin effect on brain. Diabetes
33:141–152;1984. Fellman, N.; Coudert, J.; Peyrin, L. Plasma free and sulphatedcatecholamines after ultra-long exercise recovery. Eur. J.9. James, D.E.; Burleigh, K.M.; Storlien, L.H.; Bennett, S.P.;
Kraegen, E.W. Heterogeneity of insulin action in muscle: In- Appl. Physiol. 60:91–97;1990.23. Schultz, T.A.; Lewis, S.B.; Westbie, D.K.; Wallin, J.D.; Ger-fluence of blood flow. Am. J. Physiol. 251:E422–E430;1986.
10. Jeanrenaud, B.; Halimi, S.; Van de Werve, G. Neuro-endo- ich, J.E. Glucose delivery: A modulator of glucose uptake incontracting skeletal muscle. Am. J. Physiol. 233:E514–E518;crine disorders seen as triggers of the triad: Obesity–insulin
resistance–abnormal glucose tolerance. Diabetes Metab. Rev. 1977.24. Shiota, M.; Masumi, S. Effect of norepinephrine on consump-1:261–291;1985.
11. Kahn, C.R. Insulin resistance, insulin insensitivity and insulin tion of oxygen in perfused skeletal muscle from cold-exposedrats. Am. J. Physiol. 254:E482–E489;1988.unresponsiveness: A necessary distinction. Metabolism
27(Suppl. 2):1893–1902;1978. 25. Smith, O.L.K.; Davidson, S.B. Shivering thermogenesis andglucose uptake by muscles of normal or diabetic rats. Am. J.12. Kraegen, E.W.; James, D.E.; Bennett, S.P.; Chisholm, D.J. In
vivo insulin sensitivity in the rat determined by euglycemic Physiol. 242:R109–R115;1982.26. Smith, S.A.; Young, P.; Cawthorne, M.A. Quantification inclamp. Am. J. Physiol. 245:E1–E7;1983.
13. Kraegen, E.W.; James, D.E.; Jenkins, A.B.; Chisholm, D.J. vivo of the effects of insulin on glucose utilization in individualtissues of warm- and cold-acclimated rats. Biochem. J. 237:Dose-response curves for in vivo insulin sensitivity in individ-
ual tissues in rats. Am. J. Physiol. 248:E353–E362;1985. 789–795;1986.27. Vallerand, A.L.; Lupien, J.; Bukowiecki, L.J. Interactions of14. Liu, X.; Perusse, F.; Bukowiecki, L.J. Chronic norepinephrine
infusion stimulates glucose uptake in white and brown adipose cold exposure and starvation on glucose tolerance and insulinresponse. Am. J. Physiol. 245:E575–E581;1983.tissues. Am. J. Physiol. 266:R914–R920;1994.
15. Marette, A.; Bukowiecki, L.J. Stimulation of glucose transport 28. Vallerand, A.L.; Perusse, F.; Bukowiecki, L.J. Stimulatory ef-fects of cold-exposure and cold-acclimation on glucose uptakeby insulin and norepinephrine in isolated rat brown adipo-
cytes. Am. J. Physiol. 257:C714–C721;1989. in rat peripheral tissues. Am. J. Physiol. 259:R1043–R1049;1990.16. Mikines, K.J.; Sonne, B.; Tronier, B.; Galbo, H. Effect of acute