sexual dimorphism in the song system of the carolina wrenthryothorus ludovicianus
TRANSCRIPT

Sexual Dimorphism in the SongSystem of the Carolina Wren
Thryothorus ludovicianus
PAUL M. NEALEN1* AND DAVID J. PERKEL2
1Biology Department, University of Pennsylvania, Philadelphia, Pennsylvania 19104-60182Department of Neuroscience, University of Pennsylvania, Philadelphia, Pennsylvania
19104-6074
ABSTRACTSexual and interspecific differences in the size of passerine bird song repertoires are
related to differences in the size of song-control regions (SCR) within the brain. Most speciesof Thryothorus wrens (family Certhiidae) are known to duet, and, in both sexes, songrepertoire sizes are related to the size of the SCR. However, one member of this genus, theCarolina wren T. ludovicianus, is very sexually dimorphic in its singing behavior: Malesdevelop large song repertoires, whereas females do not sing. In this study, Nissl staining wasused to investigate whether the marked gender difference in the behavior of this species isrelated to sexual dimorphism of the SCR. Carolina wren males, as predicted, possess thelargest premotor song nuclei within the genus; these nuclei could not be identified withinNissl-stained female tissue. The cellular bases for gender differences in SCR morphology alsowere examined: Males and females differed strongly in the size and density of neuronsmaking up the regions in which SCRs exist in the male forebrain. Interspecific comparisonprovided no evidence for a decoupling of behavioral and neural evolution within this clade.Male Carolina wrens possess the largest song repertoires and SCRs within the genus,whereas females of this species represent the opposite behavioral and neural extremes of thissongbird group. These results are consistent with the hypothesis that the size of the passerinesong repertoire is limited by the amount of neural tissue devoted to singing. J. Comp. Neurol.418:346–360, 2000. © 2000 Wiley-Liss, Inc.
Indexing terms: songbird; bird song; duetting repertoire size; HVc; robust nucleus of the
archistriatum
Comparative analysis of complex systems can offer in-sights into evolution and function that cannot be obtainedfrom study of any one species. One complex system that iswell suited for multifaceted study is that of bird song: ahighly patterned, often repetitive vocal signaling used incommunication (Smith, 1991). In most passerines, song isa predominantly male behavior, with females appearing touse song little or not at all; exceptions include duettingspecies, such as the Northern cardinal Cardinalis cardi-nalis (Ritchison, 1986; personal observations) and mostThryothorus wrens (Farabaugh, 1982).
Several neural regions involved in song learning andproduction have been identified (Nottebohm et al., 1976).These are discrete nuclei connected in two principal path-ways (Fig. 1; for review, see Brenowitz et al., 1997) and arereferred to here as song-control regions (SCRs). The ante-rior forebrain pathway (AFP) is crucial for the learningand refinement of song elements, because disruption of
this pathway in juveniles leads to abnormal adult song(Bottjer et al., 1984; Sohrabji et al., 1990; Scharff andNottebohm, 1991). Lesions of AFP nuclei in adults, how-ever, have little influence on song structure and delivery(Bottjer et al., 1984; Scharff and Nottebohm, 1991). Neu-ral activity in the motor pathway (MP) encodes the precisetiming and arrangement of song elements during singing(Yu and Margoliash, 1996), and lesions of the MP nuclei inadult birds cause profound loss of singing behavior (Not-
Grant sponsor: University of Pennsylvania Research Foundation; Grantsponsor: National Institutes of Health; Grant number: MH56646; Grantsponsor: National Science Foundation.
*Correspondence to: Paul M. Nealen, Biology Department, University ofPennsylvania, Philadelphia, PA 19104-6018.E-mail: [email protected]
Received 12 August 1999; Revised 19 November 1999; Accepted 19November 1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 418:346–360 (2000)
© 2000 WILEY-LISS, INC.

tebohm et al., 1976). Passerine SCRs, thus, are crucial forsong learning, production, and perception (see, e.g., Bre-nowitz, 1991) but are not suggested to be involved in anynon-song related behaviors. This provides an exceptionalopportunity to examine the neural basis for a complex yetneurally isolated, learned behavior.
Anatomic investigations of SCR in relationto behavior
Two brain regions within the MP have been common ob-jects of study: the nucleus HVc, used as the proper name(Margoliash et al., 1994) and the premotor nucleus to whichit projects, the robust nucleus of the archistriatum (RA; Fig.1). Examination of these two nuclei revealed strong neuralsexual dimorphism (NSD) in some species. For example,singing is a male-only behavior in the zebra finch (Taeniopy-gia guttata), and males have much larger SCRs than females(Nottebohm and Arnold, 1976). Intersexual study of speciespossessing male and female song (see, e.g., Brenowitz et al.,1985) as well as interspecific comparisons among male song-birds (Arnold et al., 1986; DeVoogd et al., 1993) have re-vealed a general correlation: Larger adult SCR volumes arecorrelated with overall repertoire size (measured as the num-ber of different syllable or song types performed; Brenowitzand Arnold, 1986). These findings suggested that theamount of neural material in these SCRs directly contrib-utes to behavioral function by influencing the variety of songelements that are learned or performed (Ward et al., 1998).However, causality is lacking: Do SCR volumes determinesong output, or vice versa, or does some unknown factorcontrol them both (Brenowitz and Arnold, 1986; Brenowitz,1997)? Other problems of interpretation exist. Thus far, sta-tistical assessment of the relationship between anatomic andbehavioral variables has been limited (however, see Bernardet al., 1996). The structural correlation between these vari-ables is relatively weak: Specifically, the amount of variation
in repertoire size explained by variation in the overall vol-umes of nuclei HVc and RA is typically low (see, e.g., Bre-nowitz et al., 1995). One possible explanation for this weakrelationship is that aspects other than overall SCR volumeare more critical in the determination of repertoire size. It ispossible that neuron number, volume, and density; the ratioof neurons to glia; the degree of neuronal branching; or thedegree of vascularization determine more directly or limitbehavioral versatility.
Sexual dimorphism of brain and behavior inThryothorus wrens
The correlation between song repertoire size and theelaboration of SCRs has been investigated in both sexes ofseveral species of duetting wrens (genus Thryothorus).Male and female bay wrens (T. nigricapillus) have equallysized song repertoires, and NSD of the SCRs is not appar-ent (Brenowitz and Arnold, 1986). Similarly, male andfemale buff-breasted wrens (T. leucotis) exhibit no sexualdimorphism of song repertoire size and only modest NSDof SCRs (Brenowitz et al., 1985). Finally, male rufous-and-white wrens (T. rufalbus) have both larger song reper-toires and larger SCRs than females of the species (Bre-nowitz and Arnold, 1986). In general, SCR volumes andsong repertoire size consistently are related both withinand among species in this clade (see, e.g., Brenowitz andArnold, 1986).
Focus of current study
In terms of singing behavior, one species of Thryothoruswren differs dramatically from its congeners. The Caro-lina wren (T. ludovicianus) is very sexually dimorphic inits singing behavior (Haggerty and Morton, 1995). Malesof this species develop the largest song repertoires re-ported for the genus, averaging 32 songs per male (Mor-ton, 1987). Female Carolina wrens, in contrast to bothsexes of all other members of the genus, do not sing (Hag-gerty and Morton, 1995; personal observations). Duringaggressive male singing (such as territorial defense), fe-males occasionally will respond to male song with a harshchatter (Haggerty and Morton, 1995), which may overlapthe end of male song (personal observations). There is noevidence that this chatter call is learned, because its formvaries little throughout the Carolina wren range (E.S.Morton, personal communication), and it has been heardfrom captive, hand-reared birds (Haggerty and Morton,1995). Therefore, Carolina wren males and females repre-sent the behavioral extremes of the genus in terms of thesize of their learned song repertoires.
We have examined the cytoarchitecture of nuclei HVcand RA (the primary SCR in the motor pathway for songperformance; Fig. 1) in both sexes of the Carolina wren.Our primary goal was to determine whether the markedsexual dimorphism in behavior is matched by sexual di-morphism of SCR morphology in this species. We alsoexamined in detail the basis of SCR overall volumethrough measurements of neuronal size and density. Inaddition, we present a broad examination of the relation-ship between singing behavior and SCR design within thissingle genus by direct comparison of neuroanatomic pa-rameters among these closely related but behaviorallyvery different Thryothorus wrens. In this manner, wewere able to assess the degree of matching in behavioral-neural evolution within this clade.
Fig. 1. Representation of the principal neural pathways related tosong learning and production in passerines. Physiologic and anatomicdata suggest that nucleus HVc and robust nucleus of the archistria-tum organize output of the motor pathway (solid lines). These nucleialso are connected to the anterior forebrain pathway (dashed lines),which is involved in auditory processing and song learning. DLM,medial portion of the dorsolateral nucleus of the thalamus; L-MAN,lateral magnocellular nucleus of the anterior neostriatum; nXIIts,tracheosyringeal portion of the hypoglossal nucleus; X, area X.
347SEXUAL DIMORPHISM OF CAROLINA WREN SONG NUCLEI

MATERIALS AND METHODS
Field methods
During the periods from April to June in 1996–1998,Carolina wrens were located in natural habitats in south-eastern Pennsylvania and central North Carolina. Maleswere located by their singing, which is persistent through-out the day during the breeding cycle (Haggerty and Mor-ton, 1995). The behavior of both sexes was similar betweenlocalities. Animals were mist-netted for collection in ac-cord with state and federal permits. A total of 11 malesand 3 females were collected, and animal sex was verifiedby gonadal inspection. All animals were adult in appear-ance, size, and behavior. All research methods were ap-proved by the University of Pennsylvania Animal Careand Use Committee.
Tissue processing
Collected animals were killed with sodium pentobarbi-tal (Nembutal 0.375 mg/g administered intramuscularly)and perfused transcardially with avian saline and then4% phosphate-buffered paraformaldehyde, pH 7.4. Brainsgenerally were removed from the skulls, postfixed over-night at 4°C (several brains were postfixed in situ andthen removed), and stored at 4°C. Whole brains were thendivided sagittally into halves, cryoprotected in 30% su-crose, and frozen sectioned in the sagittal plane at 25 mmon a sliding microtome (tissue from 1 male was sectionedat 20 mm). Sections were collected into 0.1 M phosphate-buffered saline, pH 7.4, and stored at 4°C. One or twoseries of sections (100 mm spacing) from each hemispherewere collected onto gelatin-subbed microscope slides,stained with cresyl violet (Sigma, St. Louis, MO), andcoverslipped in Eukitt (Calibrated Instruments, Inc.). Leftand right hemispheres were examined separately.
Light microscopy
Forebrain volume. For each series of sections, black-and-white digital images of the forebrain were capturedwith a computer-based imaging system (Hitachi KP-M1CCD camera; Data Translation DT3152 image-capturecard). Total forebrain area was determined for each sec-tion (Scion Image, Scion Corporation). Whole forebrainvolume was computed as the sum of section areas multi-plied by the sampling interval.
HVc volume. Criteria for establishing HVc boundariesfollowed those of Johnson and Bottjer (1995). The dorsalboundary of HVc was the overlying ventricle, and theventral boundary was identified by rows of small, orthog-onally arranged cells (Fig. 2, top). Cell clustering is aknown characteristic of the passerine HVc (Bottjer et al.,1986), and the first instances of cell clustering set thelateral boundary. The medial boundary of HVc was diffi-cult to establish. In practice, this was determined bysharply decreased cell clustering and a transition to auniformly sized population of cells of greatly narroweddorsoventral extent (Johnson and Bottjer, 1995). By usingthese criteria, the boundaries describing nucleus HVcwere traced with a camera lucida from those sections thatcontained the nucleus. Traces were scanned digitally, andarea and volume measures were determined as describedabove.
RA volume. Known characteristics of passerine nu-cleus RA include large, widely spaced neurons (Gurney,1981). Their occurrence, coupled with surrounding rows of
orthogonally arranged, small cells, was used to set theboundaries of nucleus RA in the sagittal plane (Fig. 2,bottom). Typically, it was possible to delimit nucleus RAover a lateral-medial extent of 500–600 mm. For thesesections, nucleus borders were traced and then digitized,as described above. Estimates of overall nucleus volumewere made by summing the traced areas and multiplyingthis sum by the sampling interval, as described above.However, in the sagittal sections used here, the lateraland medial boundaries could not be established clearly inall birds. Thus, we present a minimum estimate of overallnucleus RA volume based on those sections through thelargest portion of the nucleus in which nucleus borderscould be identified clearly.
Soma size distribution. In three males, the size dis-tribution of all cell types within HVc was sampled. Allsections containing a portion of the nucleus were dividedevenly into three groups: those covering the lateral, inter-mediate, or medial portions of the nucleus. One sectionwas chosen from each group, and the somata of cellswithin the section were traced without regard for cell formor size. A minimum of 50 cells (mean, 132 cells) was tracedper sampling location; sampled sections was a minimumof 400 mm apart. Cell soma sizes were sampled similarlyat lateral and medial locations within nucleus RA. Sam-pled sections in RA were a minimum of 300 mm apart; aminimum of 125 cells (mean, 186 cells) were traced persampling location.
Location of presumptive nuclei in females. It wasplanned originally to sample cell sizes within female nu-clei HVc and RA in the same manner. However, theboundaries of neither HVc nor RA could be determined inNissl-stained female tissue; thus, these boundaries aresexually dimorphic (see Results). However, it was still ofinterest to sample female tissue to determine whetherforebrain cytoarchitecture in these same locations alsowas sexually dimorphic. The locations of areas to be sam-pled in the female neostriatum and archistriatum thatcorresponded to the locations of nuclei HVc and RA inmales were chosen as follows.
Sections were chosen from male tissue that corre-sponded to the lateral extreme, center, and medial ex-treme of each nucleus. For each nucleus, the lateral-medial position of the section that contained the middleextent of each nucleus was noted; this section was thentransilluminated and examined under a dissecting micro-scope. Sections representing similar positions in thelateral-medial extent of a female hemisphere were thencompared with the male reference sections with regard tothe size, shape, and placement of the overlying ventricleand hippocampus (for nucleus HVc) and the lamina arch-istriatalis dorsalis and archistriatum (for nucleus RA) toidentify those sections of female tissue that best matchedthe male reference sections for lateral-medial position.Sections representing the lateral-medial extremes of eachmale nucleus were then examined to insure that the fe-male section chosen as the presumptive nucleus centerwas not outside the lateral-medial nucleus boundaries inmale tissue. Sections equidistant from the presumptivenuclei center in female tissue were chosen as lateral andmedial sampling points. Sampling areas within femalesections were then determined by matching dorsal-ventraland anterior-posterior characteristics to those in corre-sponding male tissue.
348 P.M. NEALEN AND D.J. PERKEL

Fig. 2. Nuclei HVc and RA in Nissl-stained tissue from a male Carolina wren. The boundariesdefining each nucleus are indicated by arrows. Dorsal is up; anterior is to the right. Scale bars 5 200 mm.

Thus, the size distribution of cell types within threefemales was sampled in a manner similar to the samplingin males, except that sampling locations were constrainedto two locations (lateral and medial) in both presumptivenuclei HVc and RA. For both sexes, traces were digitized.Soma area and diameter (major axis diameter) were de-termined by imaging software (Scion Image).
Neuron size distribution. Following the same sam-pling scheme described above, the size distribution of neu-rons (only) within HVc and RA also was sampled at threelocations along the lateral-medial extent of each nucleus.Neurons are recognized in cresyl violet-stained tissue by adarkly stained cytoplasm, weakly stained nucleus, anddarkly stained nucleolus (see Bottjer et al., 1986). A min-imum of 25 neurons (mean, 38 neurons) was sampled persection in HVc. A minimum of 25 neurons (mean, 39neurons) was sampled per section in nucleus RA.
Neuron density
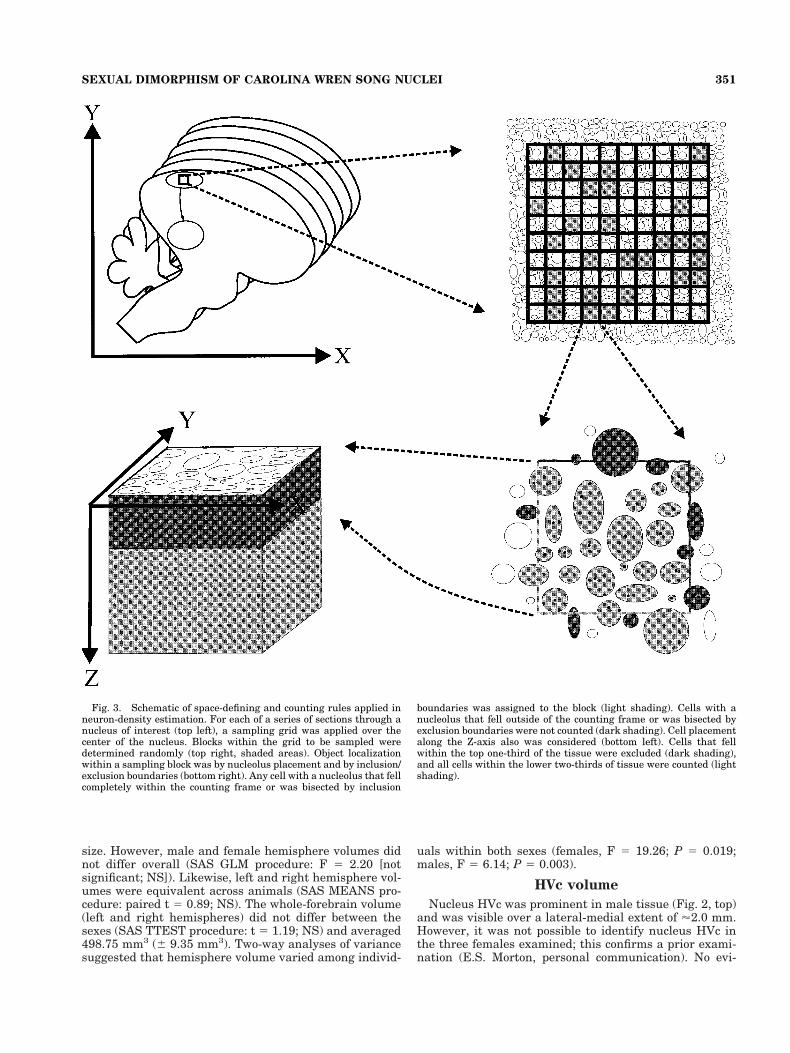
Definition of counting space. By using the same neu-ron identification criteria, the absolute density of neuronswithin both nuclei HVc and RA was examined with ste-reologically correct methods. Assessment of density re-quires the definition of a three-dimensional countingspace, with strict rules for object localization within andoutside of that space (see Coggeshall and Lekan, 1996).The definition of counting space was as follows: First, a10 3 10 reticule grid was centered over the nucleus undermoderate magnification (320), and grid placement wasleft intact when sampling magnification (340) was estab-lished (Fig. 3, top left). Individual blocks within the gridcovered 412.09 mm2 at 340 magnification and defined thecounting frames in the X-Y plane. Within the grid, blocksto be sampled were determined from a random numbertable, whose values represented row/column combinations(e.g., 07 5 seventh block of first row; 21 5 first block ofthird row). Because the sampling grid did not cover com-pletely either nucleus at its maximum cross-sectionalarea, overall placement of the grid was biased toward thecenter of the nucleus, which precluded making estimatesof total neuron numbers within these nuclei (by multipli-cation of density and volume estimates; West, 1999). How-ever, individual sampling units were spaced at randomwithin the grid (Fig. 3, top right), which permitted acomparison of neuron density estimates for the center ofthese nuclei among animals that were sampled uniformly inthis way.
Stereologic counting also requires definition of Z-axislimits to counting frames, because counts through theextent of tissue thickness are biased toward overcounting(Coggeshall and Lekan, 1996). Earlier sampling had re-vealed that the ratio of tissue thickness to depth of focuson our equipment (Olympus BH microscope, 340 lens)was approximately 3:1; thus, the Z-axis extent of thecounting space was set at two-thirds of the section thick-ness (Fig. 3, bottom left). This was the extent of the tissuedepth that could be excluded confidently from the top-of-section focal plane, and its accuracy was tested empiri-cally (see below).
Counting rules. Rules for inclusion of neurons withinthe counting space also were established. The nucleoluswas used as the counting unit, because it is small relativeto section thickness and is less likely than whole cells to besubdivided into two or more sections. Its identification incresyl violet-stained tissue also is relatively certain. Nu-
cleolus placement wholly within the counting frame rep-resented assignment of the neuron within the countingframe. When the nucleolus was bisected by a frame bor-der, its assignment to a counting frame was based oninclusion/exclusion boundaries (Fig. 3, bottom right). Asmall percentage of cells have two nucleoli; their assign-ment was based on dual inclusion within the countingframe or on the frame border that divided them (see Fig. 3,bottom right, top left of frame).
Extent of sampling. Stereologic density estimatesshould be based on a sample of 100–200 counted objects(see West, 1999). Sections sampled here were spaced uni-formly over the full lateral-medial extent of the nucleus asobserved, with a minimum spacing of 100 mm betweensampled sections. On average, 10.5 sections (range, 5–16sections) were sampled within HVc, and 6.8 sections weresampled within RA (range, 5–9 sections) because of themuch greater lateral-medial extent of HVc than RA. Be-cause of the relatively smaller numbers of sections repre-senting RA compared with HVc and because of the less-dense nature of RA (see Results), it was necessary to countmuch greater numbers of blocks within each samplingfield of nucleus RA. On average, 16.1 blocks (range, 10–50blocks) within the sampling grid were sampled for eachsection within HVc, whereas 50 blocks were sampled ineach section of nucleus RA.
Test of counting frame accuracy. The placement ofindividual counting frame limits in the X-Y plane wasrelatively simple, whereas accurate determination ofZ-axis extent was more difficult in practice. Within nu-cleus RA (only), the depth rule established was tested asfollows: After sampling individual blocks for neuron den-sity within each section, a count of all neurons covered bythe 10 3 10 grid was performed without regard to tissuedepth. If the counting depth rule was maintained accu-rately, then, on average, the following relationship wouldobtain
No. of neurons in blocks sampled for depth
5no. of blocks sampled for depth
total no. of blocks in grid ~n 5 100!
3 depth fraction ~ 5 2/3! 3total no. of neuronsunder whole grid . (1)
Whole grid counts were made on a total of 87 sections (7birds 3 2 hemispheres 3 6.2 sections per hemisphere).Analysis of these counts for nucleus RA suggests that thedepth of tissue counted, on average, was 0.836 (6 0.017),slightly higher than the 0.67 target value. More impor-tantly, this empirically determined depth value allowed usto calibrate all of the density counts by the actual amountof tissue sampled. Thus, in all analyses of neuron densitycounts for both nuclei HVc and RA, 0.836 was used torepresent the tissue depth fraction sampled. All statisticalanalyses were performed using SAS software (version6.12; SAS Institute, 1996). Data are reported as themean 6 standard error.
RESULTS
Forebrain volume
Male-female differences in body size (see Haggerty andMorton, 1995) may give rise to sex differences in brain
350 P.M. NEALEN AND D.J. PERKEL
size. However, male and female hemisphere volumes didnot differ overall (SAS GLM procedure: F 5 2.20 [notsignificant; NS]). Likewise, left and right hemisphere vol-umes were equivalent across animals (SAS MEANS pro-cedure: paired t 5 0.89; NS). The whole-forebrain volume(left and right hemispheres) did not differ between thesexes (SAS TTEST procedure: t 5 1.19; NS) and averaged498.75 mm3 (6 9.35 mm3). Two-way analyses of variancesuggested that hemisphere volume varied among individ-
uals within both sexes (females, F 5 19.26; P 5 0.019;males, F 5 6.14; P 5 0.003).
HVc volume
Nucleus HVc was prominent in male tissue (Fig. 2, top)and was visible over a lateral-medial extent of '2.0 mm.However, it was not possible to identify nucleus HVc inthe three females examined; this confirms a prior exami-nation (E.S. Morton, personal communication). No evi-
Fig. 3. Schematic of space-defining and counting rules applied inneuron-density estimation. For each of a series of sections through anucleus of interest (top left), a sampling grid was applied over thecenter of the nucleus. Blocks within the grid to be sampled weredetermined randomly (top right, shaded areas). Object localizationwithin a sampling block was by nucleolus placement and by inclusion/exclusion boundaries (bottom right). Any cell with a nucleolus that fellcompletely within the counting frame or was bisected by inclusion
boundaries was assigned to the block (light shading). Cells with anucleolus that fell outside of the counting frame or was bisected byexclusion boundaries were not counted (dark shading). Cell placementalong the Z-axis also was considered (bottom left). Cells that fellwithin the top one-third of the tissue were excluded (dark shading),and all cells within the lower two-thirds of tissue were counted (lightshading).
351SEXUAL DIMORPHISM OF CAROLINA WREN SONG NUCLEI

dence of cell clustering or stratification of cells indicativeof a ventral or caudal boundary was seen in female tissue.This contrasts with data from other Thryothorus wrens(bay wren, buff-breasted wren: Brenowitz et al., 1985;rufous-and-white wren: Brenowitz and Arnold, 1986). Inthese tropical, duetting species, nucleus HVc is apparentin both male and female tissue, and the size of HVc isrelated to the repertoire size of each sex (Brenowitz et al.,1985).
In species in which song is dominated by componentsproduced by one side of the syrinx (e.g., canaries: Notte-bohm and Nottebohm, 1976), hemispheric differences inthe functionality of nucleus HVc may exist, although in-terhemispheric size differences have not been reported.Similarly, among Carolina wren males, left and right nu-clei HVc did not differ in volume (paired t 5 0.10; NS).However, strong differences in HVc volume exist amongindividual males (F 5 7.39; P 5 0.0014). These volumetricdifferences may be indicative of functional and behavioraldifferences among males of this species. Across all malessampled (n 5 11 birds), total (left and right) HVc volumeaveraged 1.85 mm3 (6 0.11 mm3). This value is greaterthan that reported for three species of tropical, duettingThryothorus wrens in which the HVc total volumes rangefrom '1.0–1.3 mm3 (values estimated from Brenowitz etal., 1985; Brenowitz and Arnold, 1986).
RA volume
In male Carolina wrens, nucleus RA was prominent(Fig. 2, bottom) over a lateral-medial extent of 500–600mm. No hemispheric differences in RA volume were ob-served (paired t 5 0.39; NS). In addition, no differencesamong individual males were found (F 5 1.82; NS). As fornucleus HVc, it was not possible to identify the nucleus RAin Nissl-stained female tissue. No obvious differences inneuron size or density (which could indicate nucleus RA)existed within the dorsal archistriatum. Likewise, no cleararrangement of small cells around a nucleus border wasevident. This again is in contrast with data for tropical,duetting Thryothorus wrens (Brenowitz et al., 1985, Bre-nowitz and Arnold, 1986), in which nucleus RA was iden-tified in female tissue, and its volume was correlated withthe repertoire size performed. By this model, the lack ofsong performance in female Carolina wrens would lead tothe prediction of, at most, a very small nucleus RA (how-ever, see also Gahr et al., 1998).
Interspecific comparison of nucleus RA volume againsuggests that Carolina wren males are extreme in thegenus. In the 11 males sampled here, our minimum esti-mate for nucleus RA overall volume (left and right) aver-aged 0.55 mm3 (6 0.02 mm3). As found for nucleus HVc,this SCR volume was larger than that reported for thetropical, duetting Thryothorus species ('0.25–0.35 mm3;Brenowitz et al., 1985; Brenowitz and Arnold, 1986).
HVc cell size
To test for potential sex differences in the cellularmake-up of these forebrain regions, cell soma area anddiameter were determined from digitized traces of cellsomata. All cell types present were sampled (both neuronsand nonneuronal cells) at three locations (lateral, inter-mediate, and medial) within HVc for males and at twolocations within the neostriatum for females. Samplinglocations in female tissue were matched to those corre-sponding to lateral and medial nucleus HVc in male tis-
sue. A total of 1,900 cells were sampled in three males,and 1,794 cells were sampled in three females. The distri-butions of cell sizes within HVc differed very stronglybetween the sexes (SAS NPAR1WAY procedure;Kolmogorov-Smirnov statistic [Ka] 5 13.56; P , 0.0001;Fig. 4A). The average cell diameter was much greater inmales than in females (9.60 6 0.07 mm vs. 6.87 6 0.05 mm;F 5 1027.24; P , 0.0001). Sex differences in the make-upof this region of neostriatum were very apparent (Fig. 4B).No uniform positional differences (left hemisphere com-pared with right hemisphere; lateral, intermediate, andmedial locations) were seen among males or females.
RA cell size
The distributions of soma sizes for all cell types withinmale nucleus RA and its matched location in female tissuewere examined for their potential to contribute to sexdifferences in archistriatal cytoarchitecture, as describedabove. A total of 2,102 cells were sampled in three males,and 2,184 cells were sampled in three females. Similar towhat was seen in the nucleus HVc, sex differences in thedistribution of cell sizes within RA were apparent (Ka 52.14; P 5 0.0002; Fig. 5A). The distribution of cell sizeswithin male RA clearly was bimodal, with the second peakcorresponding to the population of large (.14 mm in di-ameter), presumably projection neurons, similar to whatis seen in the zebra finch (Spiro et al., 1999). However, theaverage soma diameter was similar in males and females(9.23 6 0.10 mm vs. 9.05 6 0.09 mm; F 5 2.57; NS).Overall, male-female differences in RA cell sizes were lessstriking than those of nucleus HVc (Fig. 5B). Again, nouniform positional effects on cell sizes were seen in malesor females.
HVc neuron sizes
The size distribution of neurons (only) was sampled inall males (n 5 11 birds) and females (n 5 3 birds) exam-ined at three locations along the lateral-medial extent ofmale nucleus HVc or its presumed location in females.Strong male-female differences in the distribution of neu-ron sizes within nucleus HVc were found (Ka 5 6.83; P ,0.0001; Fig. 6A). The male HVc contained neurons thatwere larger on average than the neurons found in theregion of female neostriatum occupied by HVc (13.33 60.05 mm vs. 11.57 6 0.08 mm; F 5 276.54; P , 0.0001).Within each sex, neuron size varied strongly among indi-viduals (males, F 5 79.02; P , 0.0001; females, F 5 73.08;P , 0.0001). Variation among individuals in the size ofneurons within nucleus HVc potentially may have contrib-uted to differences in singing behavior. No hemisphericdifferences in neuron size were apparent for either sex.Likewise, no uniform positional differences (lateral, inter-mediate, or medial) in neuron size were evident.
RA neuron sizes
Strong sex differences in the size distribution of nucleusRA neurons also were apparent (Ka 5 4.15, P , 0.0001) asdescribed above. Neurons in male RA averaged 16.72 60.05 mm in diameter, whereas the average neuron diam-eter in the region of female archistriatum occupied bynucleus RA was significantly smaller (15.70 6 0.10 mm;F 5 95.55; P , 0.0001).
Differences in neuron size were apparent among indi-vidual males (F 5 27.24; P , 0.0001) and among individ-ual females (F 5 227.41; P , 0.0001). Differences among
352 P.M. NEALEN AND D.J. PERKEL

Fig. 4. A: Distribution of cell sizes (both neurons and glia) within the male (n 5 3) and female (n 53) nucleus HVc. B: Photomicrograph depicting Nissl-stained nucleus HVc in males and its matchedlocation in females.
353SEXUAL DIMORPHISM OF CAROLINA WREN SONG NUCLEI

individuals in the size of neurons within this SCR, asdescribed above, may reflect variation in singing behavior,because the size of nucleus RA neurons may be an impor-tant component of behavioral ability (Gurney, 1981). Po-sition within the nucleus influenced neuron size signifi-
cantly in both males (F 5 74.58; P , 0.0001) and females(F 5 37.62; P , 0.0001). In both sexes, neuron diameterwas greatest in the lateral portion of the nucleus anddecreased monotonically through the intermediate extentto the medial extent of the nucleus. Male neuron major
Fig. 5. A: Distribution of cell sizes (both neurons and glia) within the male (n 5 3) and female (n 53) nucleus RA. B: Photomicrograph depicting Nissl-stained nucleus RA in males and its matched locationin females.
354 P.M. NEALEN AND D.J. PERKEL

axis diameter decreased from a mean of 17.45 mm (6 0.09mm) in the lateral portion of the nucleus to 16.01 mm(6 0.08 mm) in medial RA, whereas female neuron sizedecreased from a lateral mean of 16.22 mm (6 0.18 mm) to14.84 mm (6 0.16 mm) in medial RA.
Neuron density within nucleus HVc
A total of 3,580 fields in nucleus HVc were sampled forneuron density in the 11 males examined, and 1,150 fieldswere sampled in three females. Because counts within anyone section are influenced uniformly by the thickness ofthat section, our initial analyses of these density countsmade use of section mean values. This is a conservativeapproach that reduces statistical power in detecting weakeffects but that also reduces the risk of spurious resultsdue to nonindependence of data. Density estimates arepresented as 104 neurons/mm3.
Across all animals, average neuron density within nu-cleus HVc was greater in males (14.97 6 0.28) than infemales (12.93 6 0.65; F 5 9.73; P 5 0.002). Differences inneuron density among individuals were apparent withinboth females (F 5 17.50; P , 0.0001) and males (F 5 5.44;P , 0.0001). Hemispheric differences in neuron densitywere not detected in either sex (males: F 5 0.92; NS;females: F 5 1.99; NS). Neuron density varied along thelateral-medial axis in females (F 5 6.42; P 5 0.0145; Fig.7) and tended to increase slightly from lateral positions tomedial positions (SAS CORR procedure: Pearson correla-tion coefficient [r] 5 0.28; n 5 55; P 5 0.042). No lateral-medial variation in neuron density was seen in males (F 51.75; NS).
Because of the systematic manner in which densitycounts were collected, we can ask how density varies overthree dimensions within the nucleus. By using the origi-
Fig. 6. Male-female comparisons of neuron size distributions within nucleus HVc (A) and nucleusRA (B).
355SEXUAL DIMORPHISM OF CAROLINA WREN SONG NUCLEI
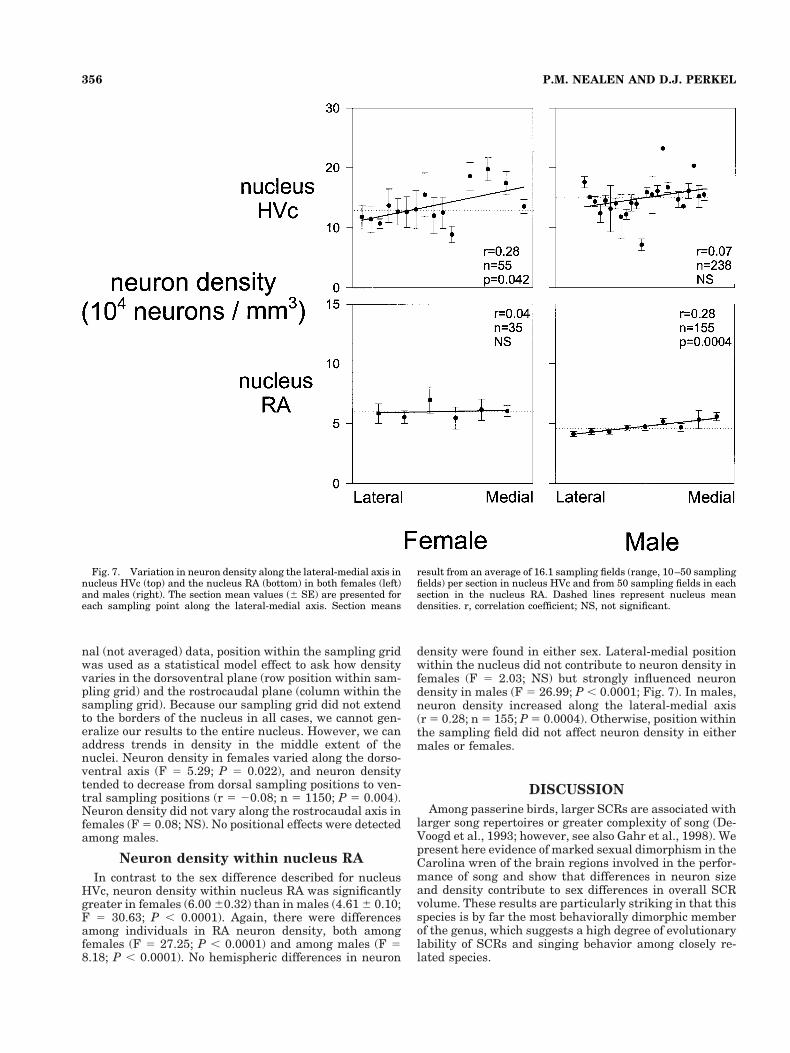
nal (not averaged) data, position within the sampling gridwas used as a statistical model effect to ask how densityvaries in the dorsoventral plane (row position within sam-pling grid) and the rostrocaudal plane (column within thesampling grid). Because our sampling grid did not extendto the borders of the nucleus in all cases, we cannot gen-eralize our results to the entire nucleus. However, we canaddress trends in density in the middle extent of thenuclei. Neuron density in females varied along the dorso-ventral axis (F 5 5.29; P 5 0.022), and neuron densitytended to decrease from dorsal sampling positions to ven-tral sampling positions (r 5 20.08; n 5 1150; P 5 0.004).Neuron density did not vary along the rostrocaudal axis infemales (F 5 0.08; NS). No positional effects were detectedamong males.
Neuron density within nucleus RA
In contrast to the sex difference described for nucleusHVc, neuron density within nucleus RA was significantlygreater in females (6.00 60.32) than in males (4.61 6 0.10;F 5 30.63; P , 0.0001). Again, there were differencesamong individuals in RA neuron density, both amongfemales (F 5 27.25; P , 0.0001) and among males (F 58.18; P , 0.0001). No hemispheric differences in neuron
density were found in either sex. Lateral-medial positionwithin the nucleus did not contribute to neuron density infemales (F 5 2.03; NS) but strongly influenced neurondensity in males (F 5 26.99; P , 0.0001; Fig. 7). In males,neuron density increased along the lateral-medial axis(r 5 0.28; n 5 155; P 5 0.0004). Otherwise, position withinthe sampling field did not affect neuron density in eithermales or females.
DISCUSSION
Among passerine birds, larger SCRs are associated withlarger song repertoires or greater complexity of song (De-Voogd et al., 1993; however, see also Gahr et al., 1998). Wepresent here evidence of marked sexual dimorphism in theCarolina wren of the brain regions involved in the perfor-mance of song and show that differences in neuron sizeand density contribute to sex differences in overall SCRvolume. These results are particularly striking in that thisspecies is by far the most behaviorally dimorphic memberof the genus, which suggests a high degree of evolutionarylability of SCRs and singing behavior among closely re-lated species.
Fig. 7. Variation in neuron density along the lateral-medial axis innucleus HVc (top) and the nucleus RA (bottom) in both females (left)and males (right). The section mean values (6 SE) are presented foreach sampling point along the lateral-medial axis. Section means
result from an average of 16.1 sampling fields (range, 10–50 samplingfields) per section in nucleus HVc and from 50 sampling fields in eachsection in the nucleus RA. Dashed lines represent nucleus meandensities. r, correlation coefficient; NS, not significant.
356 P.M. NEALEN AND D.J. PERKEL

Sexual dimorphism of SCR design:Male-dominated song
The boundaries of both nucleus HVc and nucleus RA, asdiscussed above, are readily apparent in Nissl-stainedtissue of the male Carolina wren (Fig. 2). This was ex-pected on the basis of their behavior: Carolina wren maleslearn and use large repertoires of song (Morton, 1987). Incontrast, the borders of these nuclei were not apparent inNissl-stained female tissue (see Results), and females donot sing (Haggerty and Morton, 1995; personal observa-tions). However, females of other passerine species inwhich males account for all or the vast majority of songnonetheless have recognizable SCRs that typically aremuch smaller than those of males (e.g., zebra finch: Not-tebohm and Arnold, 1976; white-crowned sparrowZonotrichia leucophrys: Baker et al., 1984; red-wingedblackbird Agelaius phoeniceus: Kirn et al., 1989). Thus,Carolina wrens represent the most extreme example ofneural, sexual dimorphism in the passerine song systemreported to date. No variation in cell size, density, andclustering that could indicate the boundaries of theseSCRs was evident in female tissue. It remains possiblethat other markers will detect SCR boundaries in femaleCarolina wrens that were not clearly delineated here byusing Nissl stain (as in the bush shrike Laniarius fune-bris: Gahr et al., 1998).
The role of SCRs in females that do not sing is unclear.Brenowitz (1991) and Del Negro et al. (1998) reported arole for nucleus HVc in the female perception of conspecificsong in canaries. However, lesions of nucleus HVc in fe-male zebra finches did not affect their ability to discrimi-nate between conspecific song and heterospecific song(MacDougall-Shackleton et al., 1998). If nuclei in the MPare necessary for female perception of male song, then theprediction could be made that nonsinging females of spe-cies characterized by large male repertoires may havelarge SCRs. However, Brenowitz et al. (1994) found nocorrelation between the song repertoire size of potentialmates and nuclear/cellular parameters of both nuclei HVcand RA in female marsh wrens Cistothorus palustris, al-though these nuclei were visible and could be measured.Thus, even a small amount of SCR tissue devoted to songperception in females may be sufficient (Brenowitz et al.,1994).
Given the large song repertoires of male Carolina wrensand the generally accepted role of male song repertoires inpasserine reproduction (see, e.g., Hasselquist et al., 1996),it seems unlikely that female Carolina wrens have no needfor song perception. Similarly, the finding that femaleCarolina wrens can respond with a rattle vocalization intime with song by their mate but not to that of neighboringmales (Haggerty and Morton, 1995), even if the mate isnot in immediate view (personal observations), suggeststhat song perception does occur and is of behavioral rele-vance. The lack of clear SCR boundaries in females of thisspecies, however, raises several alternative hypotheses forfemale SCR design and function. It is possible that nu-cleus HVc exists in females of this species but is morediffuse than in males or is not differentiated strongly (bythe size and spacing of cells) from surrounding neostria-tum. Alternatively, it is also possible that song perceptionin females of this species does not utilize a strongly orga-nized nucleus HVc but that song discrimination occursprincipally in auditory stations elsewhere, such as field L
(Lewicki and Arthur, 1996), the NCM (Vicario and Yohay,1995), or the NIf (Janata and Margoliash, 1999). Simi-larly, passerine nucleus RA serves a motor role in songproduction but also projects to respiratory motor neurons(for review, see Wild, 1997), presumably in both sexes.Again, it remains to be determined whether neurons of thepresumptive nucleus RA in female Carolina wrens fulfillthe latter task without being differentiated strongly fromsurrounding tissue. Given the clear delineation of both ofthese nuclei in male Carolina wrens as well as in femalesof other passerines characterized by male-only song (e.g.,zebra finch: Nottebohm and Arnold, 1976) and in bothsexes of congeners (T. nigricapillus, T. leucotis: Brenowitzet al., 1985), it seems unlikely that the song system of thefemale Carolina wren would be organized so differentlythat it contains no components of these regions.
Cytoarchitecture of nuclei HVc and RA
In addition to sexual dimorphism in the apparentboundaries of these nuclei in the Carolina wren, differ-ences in the cellular make-up of these regions is marked.Whereas the male to female ratios for SCR volumes havebeen published for a variety of species, the mean valuesfor basic neuronal parameters are less available. Nonethe-less, the values presented here are comparable to thosereported in males of other species that are characterizedby male-only or male-dominated song (Table 1; note thatdifferences in the methods used in tissue processing havethe potential to weaken an exact comparison of the re-ported values; however, we sought to place our resultswithin a broader context and, for this reason, have pro-vided raw values here). The distribution of cell sizes in thepresumptive female nucleus HVc is dominated by a classof small, nonneuronal cells with few large neurons andlittle of the clustering of neurons that is evident in themale nucleus HVc (Fig. 4). A class of large neurons isevident in male HVc, and, presumably, these neurons arethe two types of projection neurons in this nucleus (Dutaret al., 1998; Kubota and Taniguchi, 1998). Overall, neu-rons are both larger (Fig. 4) and more densely spaced (Fig.7) in male HVc than in female HVc. Likewise, neurons innucleus RA are larger in males than in the comparablesite in females (Fig. 5) but are more densely spaced infemales than in males (Fig. 7). This agrees with the de-scribed maturation pattern for nucleus RA in male song-birds, whereby adult nucleus size is achieved principallyby increases in neuron soma size and decreases in neurondensity during development rather than by increases inneuron number (Konishi and Akutagawa, 1985; Bottjer etal., 1986).
Comparative evidence from a variety of songbirds sug-gests a role for the size of nucleus HVc in singing versa-tility: Relatively large or complex song repertoires areassociated with a large nucleus (DeVoogd et al., 1993;Brenowitz, 1997; however, see also Brenowitz et al, 1995;Gahr et al., 1998). Presumably, both neuron size andneuron number in this nucleus are subject to evolutionarymodification. Likewise, large neurons in nucleus RA areassociated with singing behavior, as is a decreased densityof neurons in this nucleus. However, we remain relativelyuninformed about whether or how the size and density ofneurons alone contribute functionality. If individual neu-rons encode similar amounts of information, then the sizeof the song repertoire performed may be enhanced byincreases in neuron number (Brenowitz et al., 1994; Ward
357SEXUAL DIMORPHISM OF CAROLINA WREN SONG NUCLEI
et al., 1998) only if the amount of information that can becoded and used limits repertoire size. Similarly, a de-creased density of neurons in the nucleus RA seems to beassociated with the ability to sing (Gurney, 1981), but afunctional basis for enhanced performance through de-creased cell density has not been established. It is knownthat decreased neuronal density is associated with greaterdendritic lengths (DeVoogd and Nottebohm, 1981a,b),which suggests a greater synaptic density or number ofsynapses impinging on a given neuron. Because synapseshave been suggested as sites of information storage (see,e.g., Castellucci et al., 1970), such an explanation requiresthat synapse density somehow limits or contributes tobehavioral versatility.
Sexual dimorphism of Thryothorus wrens
The majority of Thryothorus wrens are tropical, duet-ting species (see Farabaugh, 1982), but the Carolina wrenT. ludovicianus stands as an exception in both aspects. Itis not known when duetting first evolved within this clade.Thryothorus wrens are allied most closely with Troglo-dytes and Thryomanes wrens (Sibley and Alquist, 1990);species within these groups do not duet. One parsimoni-ous interpretation would be that duetting is a basal char-acteristic within the Thryothorus wrens but has been lostby the Carolina wren. Alternatively, duetting may haveevolved after the divergence of the Carolina wren from itscongeners. Although females of this species do not sing,they do have the ability to attend and respond to song bytheir mates.
The relationships between sexual dimorphism in sing-ing behavior and several SCR characteristics have beenassessed in three tropical species of Thryothorus wrens(for summary, see Arnold et al., 1986). The Carolina wrenmale to female ratio for nucleus RA neuronal density(0.77) is less than that reported for the rufous-and-whitewren (1.02), the bay wren (0.89), and the buff-breastedwren (0.98), and it is greater than that reported for thezebra finch (0.20; Gurney, 1981). This is consistent with adecreased density of neurons in nucleus RA being a mas-culine characteristic associated with song production.
Similarly, the male to female ratio for RA neuron somalarea (1.18) is greater than that reported for the threeduetting species and less than that of the zebra finch(Arnold et al., 1986). This is consistent with RA neuronsize being an important component of the function of thisnucleus in species of male-only song (e.g., zebra finch:Gurney, 1981), but it contrasts with the morphology of theduetting Thryothorus wrens. In each of the duetting spe-cies, RA neuron size is monomorphic between the sexes,regardless of the degree of sexual dimorphism of behavior(Brenowitz et al., 1985; Arnold et al., 1986). This suggeststhat male-like RA neuron size is a necessary component ofany song production in this group. Overall, the character-istics of nucleus RA in both sexes of duetting Thryothoruswrens resemble more closely those of Carolina wren malesthan females (Table 1). Males and females of the duettingbay wren (Brenowitz and Arnold, 1985) and rufous-and-white wren (Brenowitz et al., 1996) also have also beenshown to be nondimorphic in their patterns of steroidhormone accumulation within several SCR, again sug-gesting that female song requires a male-like song system.
We also can assess the relative elaboration of SCRamong males of the Thryothorus species. The Carolinawren develops the largest song repertoire of the genus andpossesses a larger nucleus HVc than the congeneric bay,rufous-and-white, and buff-breasted wrens (1.85 mm3 vs.1.32, 1.24, and 1.00 mm3; latter values were estimatedfrom Brenowitz et al. [1985] and Brenowitz and Arnold[1986]). Likewise, nucleus RA is larger in male Carolinawrens (0.55 mm3) than in these congeners (0.31, 0.28, and0.26 mm3, respectively; latter values estimated from Bre-nowitz et al. [1985] and Brenowitz and Arnold [1986]).This is despite the much larger size of the rufous-and-white wren, in which the male body mass averages 50%larger than that of the Carolina wren (Dunning, 1993).Male bay and buff-breasted wrens also are larger thanCarolina wren males (17% and 12% respectively; Dun-ning, 1993). On the basis of body size alone, it may bepredicted that the Carolina wren has the smallest brain(and, hence, the smallest SCR) among these species; inactuality, its nuclei HVc and RA are the largest. These
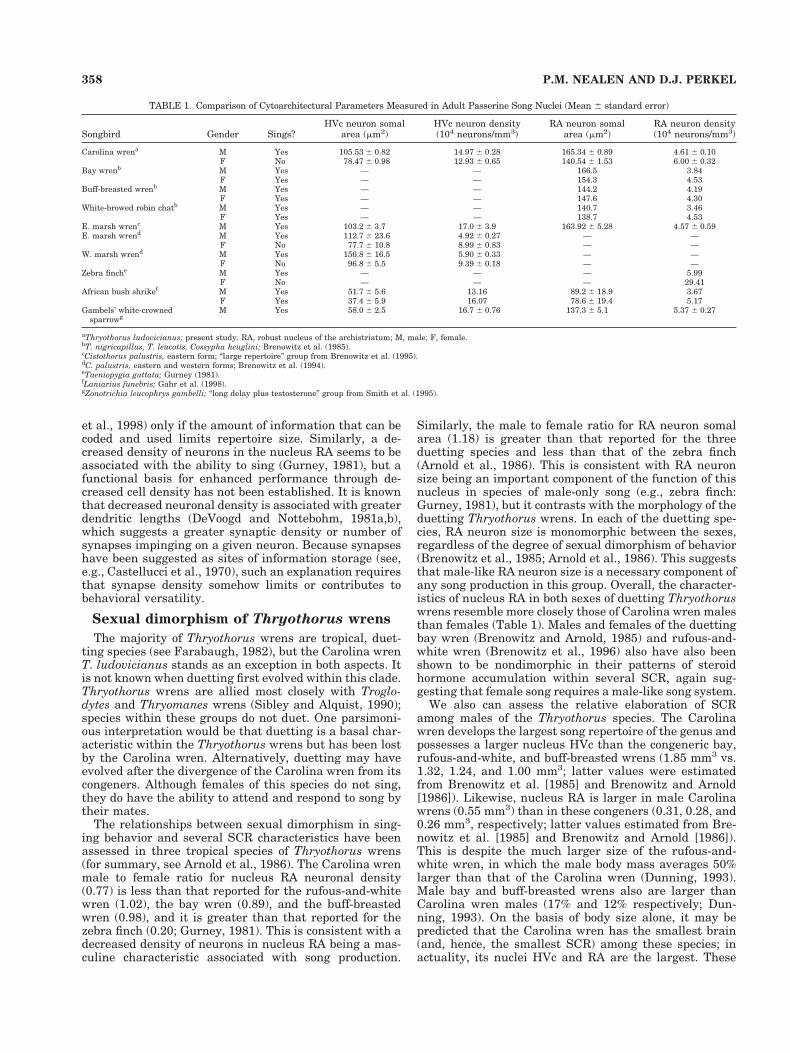
TABLE 1. Comparison of Cytoarchitectural Parameters Measured in Adult Passerine Song Nuclei (Mean 6 standard error)
Songbird Gender Sings?HVc neuron somal
area (mm2)HVc neuron density(104 neurons/mm3)
RA neuron somalarea (mm2)
RA neuron density(104 neurons/mm3)
Carolina wrena M Yes 105.53 6 0.82 14.97 6 0.28 165.34 6 0.89 4.61 6 0.10F No 78.47 6 0.98 12.93 6 0.65 140.54 6 1.53 6.00 6 0.32
Bay wrenb M Yes — — 166.5 3.84F Yes — — 154.3 4.53
Buff-breasted wrenb M Yes — — 144.2 4.19F Yes — — 147.6 4.30
White-browed robin chatb M Yes — — 140.7 3.46F Yes — — 138.7 4.53
E. marsh wrenc M Yes 103.2 6 3.7 17.0 6 3.9 163.92 6 5.28 4.57 6 0.59E. marsh wrend M Yes 112.7 6 23.6 4.92 6 0.27 — —
F No 77.7 6 10.8 8.99 6 0.83 — —W. marsh wrend M Yes 156.8 6 16.5 5.90 6 0.33 — —
F No 96.8 6 5.5 9.39 6 0.18 — —Zebra finche M Yes — — — 5.99
F No — — — 29.41African bush shrikef M Yes 51.7 6 5.6 13.16 89.2 6 18.9 3.67
F Yes 37.4 6 5.9 16.07 78.6 6 19.4 5.17Gambels’ white-crowned
sparrowgM Yes 58.0 6 2.5 16.7 6 0.76 137.3 6 5.1 5.37 6 0.27
aThryothorus ludovicianus; present study. RA, robust nucleus of the archistriatum; M, male; F, female.bT. nigricapillus, T. leucotis, Cossypha heuglini; Brenowitz et al. (1985).cCistothorus palustris, eastern form; “large repertoire” group from Brenowitz et al. (1995).dC. palustris, eastern and western forms; Brenowitz et al. (1994).eTaeniopygia guttata; Gurney (1981).fLaniarius funebris; Gahr et al. (1998).gZonotrichia leucophrys gambelli; “long delay plus testosterone” group from Smith et al. (1995).
358 P.M. NEALEN AND D.J. PERKEL

differences in SCR volume are in accord with differencesin the size of male song repertoires among species (Bre-nowitz and Arnold, 1986).
Thus, both sexes of the Carolina wren represent thebehavioral and neural extremes of the genus. We take thisas evidence for close linking of behavioral and neuralevolution within this clade. The potential exists for arelatively small number of developmental phenomena(such as circulating estrogen levels; Arnold et al., 1986) tohave effects on the degree of masculinization of both SCRand singing behavior. Only minimal evolutionary changein such a regulator could lead to this degree of markedbehavioral and neural differences among closely relatedspecies. Evidence supporting the concept of only-male oronly-female SCR design is the concerted manner in whichsexual differences in SCR design occur. In this behavior-ally dimorphic species, we do not see sex differences onlyin HVc neuron size, for example. Instead, the full suite ofmale-like SCR characteristics appears only in male Caro-lina wrens. Female Carolina wrens, despite their closeaffinity to species in which the females possess a male-likeSCR design, exhibit, at most, a dramatically reduced songsystem. Even the smallest song repertoire within thisclade (six songs [female rufous-and-white wren]; Breno-witz and Arnold, 1986) requires a masculinized song sys-tem. Thus, song system design among females does notappear to grade smoothly from a nonsong design to asinging design: Rather, some threshold amount of malecharacters may be necessary to support even minimalsong production. Among males of the genus, the basic songsystem design may be elaborated by enhanced nucleussize to support larger song repertoires. Overall, the highdegree of both intersexual and interspecific behavioraland neural diversity within this genus suggests that evo-lutionary lability of singing behavior and its neural basisis marked.
ACKNOWLEDGMENTS
We thank the editor and two reviewers for helpful com-ments.
LITERATURE CITED
Arnold AP, Bottjer SW, Brenowitz EA, Nordeen EJ, Nordeen KW. 1986.Sexual dimorphism in the neural vocal control system in song birds:ontogeny and phylogeny. Brain Behav Evol 28:22–31.
Baker MC, Bottjer SW, Arnold AP. 1984. Sexual dimorphism and lack ofseasonal changes in vocal control regions of the white-crowned sparrowbrain. Brain Res 295:85–89.
Bernard DJ, Eens M, Ball GF. 1996. Age- and behavior-related variation involumes of song control nuclei in male European starlings. J Neurobiol3:329–339.
Bottjer SW, Miesner EA, Arnold AP. 1984. Forebrain lesions disrupt de-velopment but not maintenance of song in passerine birds. Science224:901–903.
Bottjer SW, Miesner EA, Arnold AP. 1986. Changes in neuronal number,density and size account for increases in volume of song-control nucleiduring song development in zebra finches. Neurosci Lett 67:263–268.
Brenowitz EA. 1997. Comparative approaches to the avian song system.J Neurobiol 33:517–531.
Brenowitz EA. 1991. Altered perception of species-specific song by femalebirds after lesions of a forebrain nucleus. Science 251:303–305.
Brenowitz EA, Arnold AP. 1985. Lack of sexual dimorphism in steroidaccumulation in vocal control brain regions of duetting song birds.Brain Res 344:172–175.
Brenowitz EA, Arnold AP. 1986. Interspecific comparisons of the size of
neural song control regions and song complexity in duetting birds:evolutionary implications. J Neurosci 6:2875–2879.
Brenowitz EA, Arnold AP, Levin RN. 1985. Neural correlates of femalesong in tropical duetting species. Brain Res 343:104–112.
Brenowitz EA, Nalls B, Kroodsma DE, Horning C. 1994. Female marshwrens do not provide evidence of anatomical specializations of songnuclei for perception of male song. J Neurobiol 25:197–208.
Brenowitz EA, Lent K, Kroodsma DE. 1995. Brain space for learned song inbirds develops independently of song learning. J Neurosci 15:6281–6286.
Brenowitz EA, Arnold AP, Loesche P. 1996. Steroid hormone accumulationin song nuclei of a sexually dimorphic duetting bird, the rufous andwhite wren. J Neurobiol 31:235–244.
Brenowitz EA, Margoliash D, Nordeen KW. 1997. An introduction to bird-song and the avian song system. J Neurobiol 33:495–500.
Castellucci V, Pinsker H, Kupfermann I, Kandel E. 1970. Neuronal mech-anisms of habituation and dishabituation of the gill-withdrawal reflexin Aplysia. Science 167:1745–1748.
Coggeshall RE, Lekan HA. 1996. Methods for determining numbers of cellsand synapses: a case for more uniform standards of review. J CompNeurol 364:6–15.
Del Negro C, Gahr M, Leboucher G, Kreutzer, M. 1998. The selectivity ofsexual responses to song displays: effects of partial chemical lesion ofthe HVC in female canaries. Behav Brain Res 96:151–159.
DeVoogd TJ, Nottebohm F. 1981a. Sex differences in dendritic morphologyof a song control nucleus in the canary: a quantitative Golgi study.J Comp Neurol 196:309–316.
DeVoogd TJ, Nottebohm F. 1981b. Gonadal hormones induce dendriticgrowth in the adult avian brain. Science 214:202–204.
DeVoogd TJ, Krebs JR, Healy SD, Purvis A. 1993. Relations between songrepertoire size and the volume of brain nuclei related to song: compar-ative evolutionary analyses amongst oscine birds. Proc R Soc London(B) 254:75–82.
Dunning JB Jr. 1993. CRC handbook of avian body masses. Boca Raton,FL: CRC Press.
Dutar P, Vu H, Perkel DJ. 1998. Multiple cell types distinguished byphysiological, pharmacological, and anatomic properties in nucleusHVc of the adult zebra finch. J Neurophys 80:1828–1838.
Farabaugh SM. 1982. The ecological and social significance of duetting. In:Kroodsma DE, Miller EH, editors. Acoustic communication in birds, vol2. New York: Academic Press Inc. p 85–124.
Gahr M, Sonnenschein E, Wickler W. 1998. Sex difference in the size of theneural song control regions in a duetting songbird with similar songrepertoire size of males and females. J Neurosci 18:1124–1131.
Gurney ME. 1981. Hormonal control of cell form and number in the zebrafinch song system. J Neurosci 1:658–673.
Haggerty TM, Morton ES. 1995. Carolina wren (Thryothorus ludovicia-nus). In: Poole A, Gill F, editors. The birds of North America, no. 188.Philadelphia: The Academy of Natural Sciences; Washington, DC: TheAmerican Ornithologists’ Union.
Hasselquist D, Bensch S, Vonschantz T. 1996. Correlation between malesong repertoire, extra-pair paternity and offspring survival in the greatreed warbler. Nature 381:229–232.
Janata P, Margoliash D. 1999. Gradual emergence of song selectivity insensorimotor structures of the male zebra finch song system. J Neuro-sci 19:5108–5118.
Johnson F, Bottjer SW. 1995. Differential estrogen accumulation amongpopulations of projection neurons in the higher vocal center of malecanaries. J Neurobiol 26:87–108.
Konishi M, Akutagawa E. 1985. Neuronal growth, atrophy and death in asexually dimorphic song nucleus in the zebra finch brain. Nature 315:145–147.
Kubota M, Taniguchi I. 1998. Electrophysiological characteristics of classes ofneuron in the HVc of the zebra finch. J Neurophys 80:914–923.
Lewicki MS, Arthur BJ. 1996. Hierarchical organization of auditory tem-poral context sensitivity. J Neurosci 16:6987–6998.
MacDougall-Shackleton SA, Hulse SH, Ball GF. 1998. Neural bases of songpreferences in female zebra finches (Taeniopygia guttata). Neuroreport9:3047–3052.
Margoliash D, Fortune ES, Suffer ML, Yu AC, Wren-Hardin BD, Dave A.1994. Distributed representation in the song system of oscines: evolu-tionary implications and functional consequences. Brain Behav Evol44:247–264.
Morton ES. 1987. The effects of distance and isolation on song-type sharingin the Carolina wren. Wilson Bull 99:601–610.
359SEXUAL DIMORPHISM OF CAROLINA WREN SONG NUCLEI

Nottebohm F, Arnold AP. 1976. Sexual dimorphism in vocal control areasof the songbird brain. Science 194:211–213.
Nottebohm F, Nottebohm M. 1976. Left hypoglossal dominance in thecontrol of canary and white-crowned sparrow song. J Comp Physiol A108:171–192.
Nottebohm F, Stokes TM, Leonard CM. 1976. Central control of song in thecanary. J Comp Neurol 165:457–486.
Ritchison, G. 1986. The singing behavior of female Northern cardinals.Condor 88:156–159.
SAS Institute, Inc. 1996. SAS release 6.12. Cary, NC: SAS Institute, Inc.Scharff C, Nottebohm F. 1991. A comparative study of the behavioral
deficits following lesions of various parts of the zebra finch song system:Implications for vocal learning. J Neurosci 11:2896–2913.
Sibley CG, Alquist JE. 1990. Phylogeny and classification of birds. NewHaven, CT: Yale University Press.
Smith GT, Brenowitz EA, Wingfield JC, Baptista LF. 1995. Seasonalchanges in song nuclei and song behavior in Gambel’s white-crownedsparrows. J Neurobiol 28:114–125.
Smith WJ. 1991. Singing is based on two markedly different kinds ofsignaling. J Theor Biol 152:241–253.
Sohrabji F, Nordeen EJ, Nordeen KW. 1990. Selective impairment of songlearning following lesions of a forebrain nucleus in the juvenile zebrafinch. Behav Neural Biol 53:51–63.
Spiro JE, Dalva MB, Mooney R. 1999. Long-range inhibition within thezebra finch song nucleus RA can coordinate the firing of multipleprojection neurons. J Neurophys 81:3007–3020.
Vicario DS, Yohay KH. 1993. Song-selective auditory input to a forebrainvocal control nucleus in the zebra finch. J Neurobiol 24:488–505.
Ward BC, Nordeen EJ, Nordeen KW. 1998. Individual variation in neuronnumber predicts differences in the propensity for avian vocal imitation.Proc Natl Acad Sci USA 95:1277–1282.
West MJ. 1999. Stereological methods for estimating the total number ofneurons and synapses: issues of precision and bias. Trends Neurosci22:51–61.
Wild JM. 1997. Neural pathways for the control of birdsong production.J Neurobiol 33:653–670.
Yu AC, Margoliash D. 1996. Temporal hierarchical control of singing inbirds. Science 273:1871–1875.
360 P.M. NEALEN AND D.J. PERKEL