sessile a - university of hawaii...the two coral patch reefs utilized in this study are located in...
TRANSCRIPT

ABSTRACT
Name: Clark R. Lewis Department: Biological Sciences
Title: Sessile Invertebrate Colonization of a Coral Patch Reef: A Study of Two Reefs in Kaneohe Bay, Hawaii
Major: Biological Sciences Degree: Master of Science
Approved by: Date:
NORTHERN ILLINOIS UNIVERSITY

ABSTRACT
Marine invertebrate colonization for a complete annual cycle was
examined on two coral patch reefs in Kaneohe Bay, Oahu, Hawaii. Poly
vinyl chloride panels provided the substratu~ for settlement and their
placement on the reefs was along ~indward to leeward (upstream to
downstream) transects. Counts of individual organisms and area covered
by colonies provided data for site by site and inter-reef comparisons
of temporal and spatial colonization trends.
Over 80% of the total invertebrate settlements could be ascribed
to five taxonomic groups: oysters, barnacles, serpulid worms,
bryozoans, and tunicates. The patterns of colonization exhibited by
these five groups are analyzed and discussed in detail. The greatest
numbers of new settlements consistently occurred at the shallow wind
ward site of each reef, whereas the least amount of colonization took
place in the middle of the study reefs. These colonization phenomena
are discussed with respect to the influence of various physical and
biological factors.
Five months into the study, all of the fishes were removed from
the smaller of the two patch reefs, providing at least temporarily, e
means of examining the effects of fish on invertebrate colonization.
Visual transects were used prior to and after fish removal to assess
the resident fish population. Due to the rapid recolonization of the
reef t particularly by dominant herbivores, major effects on inverte
brate color,izat~on patterns were not detected.

NORTHERN ILLINOIS UNIVERSITY
SESSILE INVERTEBRATE COLONIZATION
OF A CORAL PATCH REEF:
A STUDY OF TWO REEFS IN KANEOHE BAY, HAWAII
A THESIS SUBMITTED TO THE GRADUATE SCHOOL
IN PARTIAL FULFILLMENT OF THE REQUIREMENTS
FOR THE DEGREE
MASTER OF SCIENCE
DEPARTMENT OF BIOLOGICAL SCIENCES
BY
CLARK R. LE~.JI S
DEKALB, ILLINOIS
AUGUST 1980

Certification: In accordance with departmental and Graduate
School policies, this thesis is accepted in
partial fulfillment of degree requirements.
Thesis Director
Date

iii ACKNOWLEDGEMENTS
Facilities and logistical support for this study were provided
by the Hawaii Institute of Marine Biology with partial funding from
grant #R803983 of the Environmental Protection Agency. Additional in
formation and services were supplied by the Hawaii Coastal Zone Data
Bank.
Many persons assisted in the various stages of this study and,
to all of them, I am very grateful. I especially wish to thank Dr.
Richard Brock, Dr. Stephen Smith, and Mr. Paul Jokiel for their thought
ful input and helpful encouragement throughout the course of this work.
I am also indebted to Dr. Carl von Ende for his contributions and his
willingness to serve as my advisor despite obvious difficulties in com
municating over a vast geographical distance. Most of all, my deepest
appreciation goes to my wife, Kay, for her patience, understanding, and
consistent support of my efforts.

TABLE OF CONTENTS
INTRODUCTION . . . . . . . . . . · . . . . • • • • •
MATERIAL AND METHODS • . . . . . . . . . . . . . . . . Physiography of Study Location .
Study Reefs ••••••••••
· . - . . . . . . · . . . . . .
Fouling Panels.
Experimental Reef
RESULTS . . . . . . . . . .
. . . . .
• • • · . . . . . . . . . . . .
· . . . . . . . . • • • . . . Climatology · . . . . . . · . . . . . . . . . . . . . . Long Term Communities ••••
Algal Production and Grazing.
· . . . . . . . . · . . . ~ . . . . .
Colonization of Panels ••••••• · . . . . . . . . . . . Oysters •••• • • • • . •• . . . . Barnacles
Serpu1 ids
Tun; cates
· . . . . · . . . . . . . . . . . . · . . . . . . . . . . . . . . . . .
Bryozoans
Other Invertebrates . . . . . . . . . . . . . . . Experimental Reef - Defaunation
DISCUSSION AND CONCLUSIONS •
BIBLIOGRAPHY • • . . . ..
APPENDICES • . . . . . . . . . . . .
· . . . . . . . . · . .
· . . . . . . . . · . . .. . .
Page
1
4
4
9
11
20
24
24
25
28
32
37
44
51
58
63
64
69
76
85
93
iv

v
LIST OF TABLES
Table Page
1 Summary of long term communities. · · 27
2 Nonparametric ANOVA for chlorophyll a . • · · · 31
3 Three factor ANa VA for numbers of oysters · · · · 43
4 Mean numbers of Balanus · · . · • · · 49
5 Mean numbers of Hldroides . · · · • . . 52
6 Mean numbers of spirobids . · · · • · 53
7 Mean numbers of Ascidia sidneiensis 60
8 Mean numbers of didemnid tunicates . . . • · 61
9 Monthly mean numbers of tunicates 62
10 Feeding categories of fishes collected by l'otenone 70
A-l Diffusion factors (\'Iater mot ion) at study sites 96
A-2 Fouling organisms after is month exposure . . . . 101
A-3 Raw panel set data • . . . · · · . . . . . 105
A-4 Fishes collected from experimental reef 126

Figure
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
A-l
A-2
A-3
A-4
A-5
LIST OF FIGURES
Page
Hawaiian Islands •••••••• . . · . . . 5
7 Kaneohe Bay and study reefs . . . . . . . . Fouling panels . . . . • . . . . . • · · · · · 13
Panel placements • . . · • · · • · 15
Microgram-atoms of chlorophyll a . • · · • · 29
Grazing indi ces - experimental reef · · · · · 33
Grazing indices - control reef · · • 35
Numbers of oysters - upper panel surface 38
Numbers of oysters - lower panel surface · • · 40
Numbers of Bal anus - upper panel surface . 45
Numbers of Balanus - lower panel surface • · 47
Numbers of H;tdroides - lower panel surface 54
Numbers of spi rob; ds - lower panel surface 56
Numbers of Microporelia - lower panel surface · · • · 65
Numbers of Bugula neritina - lower panel surface. 67
Reef fish colonization curve
Current patterns in Kaneohe Bay
· . . . . . . . 73
94
97
99
Ambient light conditions for Kaneohe Bay ..
Wind speed and direction at Coconut Island •.
Water temperatures in Kaneohe Bay
Rainfall in Kaneohe Bay watershed
. . . . . . . . . . .
• 129
• 131
vi

INTRODUCTION
Renowned for their biological complexities, coral reefs are among
the most diverse ecosystems in the world (Day, 1963; Connell, 1978).
Sessile marine invertebrates such as sponges, barnacles, certain
molluscs, annelids, bryozoans, hydrozoans, and tunicates are extremely
abundant and represent components of the marine community that con
tribute significantly to the complex structure of coral reefs (Wells,
1957; Kohn, 1971). Charles Darwin (1842) was among the first to note
the diversity of sessile invertebrates on coral reefs and Dana (1875)
gave a detailed account of coral reefs in which he noted the impor
tance of hydrozoans and bryozoans to the overall reef structure.
The attachment of sessile marine invertebrates to hard substratum
is commonly termed IIfouling", primarily due to the adverse effects
that accumulations of invertebrates have on man-made structures. As a
phenomenon that man has had to deal with for centuries, invertebrate
fouling has been the subject of a vast amount of scientific research.
Fouling studies, however, have generally tended to focus on either the
biology of a specific organism or on the nature of the fouling as it
relates to human activities in the marine environment. Studies
specifically concerned with the community role of sessile inverte
brates and particularly with their function in determining and
maintainlng the structure of coral reefs are limited.
Investigations of fouling that deal with community aspects of
marine invertebrate colonization have primarily been concentrated

2
in temperate regions. Examples of this work come from the west coast
of the United States (Cae, 1932; Johnson and Mii1er, 1935; Coe and
Allen, 1937; Boyd, 1972), Alaska (Long, 1972), the easter'n U.S.
coastal regions (Cowles, 1930; Grove, 1933; McDougall, 1943; Mook,
1976; Osman, 1977) and Australia (Allen and Wood, 1950; Dew and Wood,
1955; Wisely, 1959; Stephenson and Searies, 1960). Related studies in
tropical and subtropical waters are less common but have been conducted
in Hawaii (Edmondson, 1937, 1946; Edmondson and Ingram, 1939; Long,
1974), Asia (Vrijmoed, 1975) and Central America (Birkeland et ~.,
1976). Recently, Schoener et~. (1978) compared invertebrate cOloni
zation at seven geographic locations ranging from tropicai to subarctic
regions. References of other related fouiing studies can be found in
Woods Hole Oceanographic Institute (1952), Boyd (1972) and Schoener
(1974). By far, the majority of these studies have dealt with the
colonization of surfaces set apart from the natural benthic substratum.
None of the previously mentioned works has dealt with sessile inverte
brate colonization on coral reefs or the role that such colonization
has in maintaining the structure of coral reef communities.
The influence of fishes on benthic invertebrate communities has
been noted within a number of relatively recent papers (Stephenson
and Searles, 1960; Bakus, 1964, 1966, 1969; Sutherland, i974; Vine,
1974; Kaufman, 1977; Ogden and Lobel, 1978; Virnstein, 1977).
Because these authors have suggested that fishes play an important
role in determining the success of various invertebrate components
within the benthic community, reef fishes may be an important factor
in determining the overall community structure of coral reefs.

3
The purpose of this study was to collect information on the
developmental patterns within a coral reef community and to elucidate
some of the mechanisms responsible for these patterns. To achieve \
this goal, there were two aspects to this study: (1) a documentation
of patterns, both temporal and spatial, of sessile invertebrate coloni
zation on coral patch reefs and (2) an experimental perturbation
designed to determine the role of coral reef fishes in influencing
these patterns.
Two patch reefs were selected for this study because of their
similarities and location within a large embayment. To observe
invertebrate colonization, fouling panels were placed in specific
areas on each of these two reefs. The patch reefs represented simpler
and more easily manipulated microcosms of much larger coral reefs and,
as such, information from them could be extrapolated to the larger
ecosystems.
One of the patch reefs was subjected to the experimental pertur-
bation which consisted of removing the entire fish community on that
particular reef. Invertebrate colonization on the panels was recorded
for both reefs prior to and foilowing this perturbation to assess the
role of reef fishes and to relate this to the development and mainten
ance of the coral patch reef co~unity.

4
MATERIALS AND ~1ETHODS
Physiography and Climate of Study location
This study was conducted in Kaneohe Bay, Oahu in the Hawaiian
Islands from March, 1977 to June, 1978. Kaneohe Bay is situated in a
semi-tropical region of the Pacific Ocean at 21 0 28 1 north latitude
and 1570 48' west longitude (Figure 1). The Bay is primarily
characterized by the presence of a barrier reef extending across its
mouth and cut by two navigable channels (Figure 2). The inner portien
of Kaneohe Bay covers an area of approximately 28 sq km and is composed
of a relatively flat mud-sand lagoon floor with a mean depth of about
15 m. This lagoon is broken by numerous patch reefs in its north
western and central regions and by a fringing reef extending along
most of its shoreline. Within the past 20 to 30 years, the environ
ment of the bay has been significantly stressed and altered to some
extent by human activities such as fishing, dredging, and sewage
discharge (Smith et £1.,1973; Banner, 1974; Devaney et ~., 1976;
Smith, 1977). Prominent features of the Bay include four islands, a
gravel bar, and a sand bar; the largest of these formations being
Coconut Island (Figure 2).
During this study, meteorological data were recorded daily at
the Hawaii Institute of Marine Biology (HIMB) located on Coconut
Island. Sunlight, wind speed and direction, amount of rainfall, and
water temperature were monitored as possible factors influencing

FIGURE 1. The Hawaiian Islands showing the location of
Kaneohe Bay, Oahu (from Smith et ~., 19i3).
5

6
z ~
\
... ; %
Q C 0 ... Q ...
:."
• • ... . • .! .. i e
..! ;;
-' .. • . -:: Q Q . z • . ... ~ ~
0 f;;:.:=-c
c Q; ~
~
II <>
c ~ ~ • • ~ ,=,- ~--.. ..
• • ~ .. .::: 'b
~
t: ... ~
!..:. III
~ oi I.)
c Va ... 0
: ~ '"' J
u ;;:: U ~
..: , ;; C :
~ ~
i -:E-
i ':! ~ • I - :. I -

FIGURE 2. Prominent features of Kaneohe Bay, Hawaii shm·!ing
the location of the two study reefs. Areas \'Iithin
the Bay that are above the high tide line are
indicated by solid black.
7

8

9
invertebrate colonization.
The Study Reefs
The two coral patch reefs utilized in this study are located in
the central sector of Kaneohe Bay, approximately 230 m apart and about
150 to 200 m into the lagoon from the barrier reef flat (Figure 2).
Unlike other areas within the Bay, the reefs in this sector have not
been subjected to the same high levels of stress from stream runoff
and sewage discharge (Smith, 1977). The two study reefs were primarily
selected because of their reasonably small size and relative isolation
from surrounding reef areas. Both of these reefs are, furthermore, in
an area of the Bay that is subject to unidirectional current flow
during trade wind conditions (Appendix Figure A-l).
The smaller and more northern of the two patch reefs was the site
of a previous study on the community structure of the resident fish
populations (Wass, 1967). Largely because of this previous work, the
northernmost reef became the experimental reef for this study and the
site of the fish removal.
The experimental reef is roughly circular having a mean diameter
at the top of 26 m. At the 6 m depth contour, the reef has a cir
cumference of 111 m giving it a total area of approximately 1200 sq m
to this depth. The uppermost portion of the reef is relatively flat
and lies at a mean depth of just under a meter, although it is
occasionally exposed during periods of very low spring tides. The
majority of the reef ;s coral rubble with about ten percent of the top
and upper slopes covered by live heads of Porites compressa. Other

10
corals present but less common include Fungia scutaria, Pccillopora
damicornis, Montipora flabellata, Montipora patula, and Pavona
varians. By far, the dominant alga is Oictyosphaeria cavernosa, that
covers nearly 40% of the uppermost areas on the reef. Much less
abundant are algae of the genera Hypnea, Ceramium, Lyngbya, and
Porolithon.
Aside from the coral, the most common invertebrates on the experi
mental reef include the sea cucumbers, Holothuria atra and Ophiodesoma
spectabolis, mantid shr.imps of the genera Gonodactylus and Squilla,
the colonial hydrozoan, Pennaria, the oyster, Ostrea sandvichensis,
and numerous annelids, tunicates, bryozoans, and porifera. The
slopes of the reef below about six meters are generally steeper than
45° and consist primarily of unstable rubble surfaces intergrading
into the mud-sand of the lagoon floor.
The control reef used in this study is also roughly circular with
a relatively flat top of 40 m in mean diameter. With a circumference
of 171 m at the six meter depth contour, the total area encompassed
by this reef is approximately 2300 sq m. Much like the experimental
reef in overall appearance, the control reef is largely coral rubble
interspersed with areas of living Porites comDressa. All of the
corals previously mentioned are also present on this reef with the
addition of Psammocora stellata. The green alga, Oictyosphaeria
cavernosa, and its congener, Q. verslusii, cover about 30% of the
upper reaches of the control reef. Again, the algal genera Hypnea,
Ceramium, Lynqbya, Porolithcn are represented. The invertebrates are
much the same as those found on the experimental reef although the sea

11
urchin, Tripneustes gratilla, is also encountered. As before, the
reef below about six meters supports little in the way of conspicuous
organisms owing to the unstable nature of the rubble slopes.
The specific direction and relative amount of water flow on the
two reefs was studied using fluorescein dye and plaster-of-paris
"clod ll cards. Fluorescein dye was released into the water next to the
reefs under varying wind and tide conditions and the resultant obser
vations provided the basis for upstream to downstream placement of the
fouling panels. Clod cards, designed and employed following the
methods of Doty (197l), were placed along these windward to leeward
transects. The clod cards, consisting of plaster-of-paris cubes glued
onto plastic cards, were weighed both before and after placement in
the field. Their subsequent loss of weight directly relates to the
amount of water motion and diffusion at the particular locations
(Muus, 1968). The diffusion factors derived from the clod cards give
an indication of the magnitude of greater water motion which occurred
at the windward crests and along the tops of both reefs (Appendix
Table A-l).
Fouling Panels
Grey polyvinyl chloride (PVC) fouling panels were utilized
throughout this study. Although various authors have demonstrated
that settling by marine invertebrates can be affected by such sub
stratum attributes as color (Visscher and Luce, 1928; Pomerat and
Reiner, 1942; Gregg, 1945; Weiss, 1947; Smith, 1948), type of material
(Pomerat and Weiss, 1946; Crisp, 1965, 1967) and surface texture

12
(Pomerat and Weiss, 1946; Barnes, 1955, 1956), these phenomena were
beyond the scope of this present study. PVC has recently been used in
similar situations where it has proven to be a suitable substratum for
the attachment of many, if not most, marine invertebrates. The
preparation of the study paneis consisted of lightly sanding their
surfaces and soaking them in seawater at least 24 hours to remove any
manufacturing residues.
The fouling panels measured 20 X 18 X 0.3 cm and were mounted
along plastic tracks in sets of three to concrete blocks (Figure 3).
This mounting arrangement provided for a stable placement of the panels
on the reef while allowing for weekly removal and replacement of
single plates. Whereas the tracks limited the exposed surface area
of each panel to just over 260 sq cm, Mook (1976) previously demon
strated that 225 sq cm of planar surface were sufficient to collect
85% of the probable fouling species within a given location. All
surfaces of the panels used in this study were fully exposed with the
exception of the underside of the middle panel in each set. To
minimize any edge effects across the upper surfaces of the panels,
the panel sets were oriented perpendicular to the direction of the
prevailing currents.
The panel sets were installed on the two study reefs in May,
1977 and were removed 15 months lat2r in July, 1978. Five panel S2tS
were placed on each reef along the previously mentioned windward to
leeward (upstream to downstream) transect (Figure 4). There was a
certain amount of site biasing to insure stable p1acement of t~e panel
sets, but every effort was made to keep from deviating from the

FIGURE 3. Fouling panel sets. Photo shows construction of
panel holders and orientation of the PVC panels.
13

14

15
FIGURE 4. Diagrammatic illustration of panel placements.
The triangular symbols refer to panel locations.

MEAN
3 DEPTH ... 0 ,
~
PREVAILING P 11.0 8.8
A~ F M M ~C?(9)w \'V • X" ~ CURRENT
6
EXPERIMENTAL REEF \ t8 .~
SIDE VIEW ~ 'C14 at
~-- .. 2 .. ~~ ~ '""4 .... '"'.~. ~------}o T./""" --......v
3
PREVAILING
CUR~EHT
~
- --1--- 16.4 ~ M 6'1
CONTROL REEF SIDE VIEW
"<9 ~ >-
2
4
6
6
114M ..... m

17
straight line transect while keeping to selected depths.
Because initial colonization by invertebrates was the major con
cern of this study, the time of exposure for each panel was considered
critical. Optimum exposure was judged to be the period of time ending
just before significant species interactions occurred that would
affect species survivorship. Wilson (1969) termed this the period of
non-interactive species equilibrium. Preliminary investigations on
the two patch reefs indicated that a three week time interval would
be within this non-interactive period and thus near the optimum
exposure. A lesser time was not enough to allow sufficient develop
ment of many of the organisms whereas by the fourth week, enough
community development had taken piace that species interactions
(primarily competition) were becoming obvious and were beginning to
noticeably affect the various colonizers. The arrangement of three
panels per set made it possible to stagger the placement of the panels
so that each week, one panel having a three week exposure could be
removed and replaced at each site. The weekly results reported in
this study are based on these three week exposure times.
As each panel was collected in the field, it was placed into a
plastic tray containing seawater. The panels were brought back to the
laboratory totally immersed, and care was taken not to disturb their
surfaces. Because the development of marine invertebrate communities
has been associated with a1gal production (Sakus, 1964; Vine, 1974;
Brock, 1978; Ogden and Lobel, 1978), a sample of algae was taken from
each fouling panel. Immediate1y after being brought in from the reefs.
a 5 X 5 em area of one of the upper surface corners was scraped. The

18
material was washed into an opaque vial containing 15 to 20 m1 of 95%
acetone. These vials were stored in a freezer for subsequent phyto
pigment analyses according to the techniques and calculations outlined
in Strickland and Parsons (1968).
As the panels were being scraped, they were examined for con
spicuous soft-bodied animals such as anemones; notes were taken on
these as well as on any macroscopic organisms that had fallen off the
panel surfaces during transport. The panels were then kept in the
plastic holders and allowed to air dry. ~~hen time permitted, further
and more extensive examinations were conducted by positioning each
panel under a grid marked off in 1 sq cm segments. The panels were
viewed using a Wild binocular microscope at 120X magnification and
numbers of organisms were counted along a transect of 70 sq cm of the
grid. These values were extrapolated to express numbers of organisms
per 100 sq cm of panel area. Colonial organisms. with the exception
of didemnid tunicates, sponges, and hydrozoans, were counted as indi
viduals under the assumption that the settlement of a single larval
specimen initiated the development of the colony. The area of
coverage for calcareous algae as well as for the colonial organisms
noted was determined and has been expressed as a percentage of the
total panel area.
When the field portion of this study was terminated in July,
1978, the panel mounting blocks had been exposed for a total of 15
months. The concrete mount; ng bl acks ... ,ere rerooved from thei r pas i
tions on the reef and the fouling communities present on these blocks
were analyzed to determine the composition of the long-term

19
communities at each location. Again, numbers of discrete individuals
were counted and the percentage of area covered by colonial organisms
was noted.
Grazing by both fishes and invertebrates was evident on the upper
surfaces of virtually all panels brought in from the fieid. Secause
of this and the relative importance to invertebrate colonization, an
attempt was made to quantify the amount of grazing.
Two types of marks on the panels made it easy to distinguish
grazing by invertebrates from the grazing by fishes. Visual observa
tions led to the discovery that small, linear "bite" marks less than
one cm long were produced by juvenile parrotfishes (Scaridae).
Although it was never substantiated, circular sucker-like marks
appeared to be the result of grazing by gastropod molluscs, possibly
of the family Cypraeidae.
Ten intersection points were selected at random on the counting
grid and the type of grazing mark present under each intel~section
point was noted. A mark occurring under one or two grid points
resulted in a grazing index of one, marks at three to four points gave
an 'index of two, and so on to the maximum grazing index of five.
These index values were recorded along with an estimate of the per
centage of grazing attributable to the fishes.
Data obtained from the fouling panels were analyzed with the aid
of computer programs of the Statistical Analysis System (SAS; Barr et
al., 1976) and the P-Series of Biomedical Computer Programs (BMDP;
Dixon and Brown, 1977). Temporal patterns were deduced from time
series plots drawn for the most abundant groups of invertebrate

20
colonizers.
The fouling panel data primari1y consisted of counts of organisms
and, as such, contained many zero values. The data for most of the
invertebrate groups were, therefore, not normally distributed and
severely violated assumptions necessary for conducting parametric
statistical tests. Although the experimental design of this study
suggested a three-way analysis of variance (ANOVA) with the two reefs,
the two panel surfaces, and the five sites as factors in the analysis,
in most instances the count data made this type of statistical test
invalid. Multiple factor nonparametric ANOVA tests are possible (e.g.
Wilson, 1971), but these tests have not received widespread acceptance
and the results obtained from them are, at best, very difficult to
interpret. For this reason, analysis of the fouling panel data con
sisted mostly of qualitative evaluations based on the time-series
plots. Mean numbers of individuals and their 95% confidence intervals
were tabulated for some invertebrate groups when these values proved
useful for better depicting spatial colonization patterns. Data for
colonization of the lower surface of the middle panel within each set
were excluded from the analyses because its close proximity to the
concrete mounting block (see Figure 3) resulted in exposure conditions
different from the other panels.
The Experimental Reef
After five months of monitoring invertebrate colonization on the
two patch reefs, virtually all of the fishes were removed from the
experimental reef. Although this was part of a larger study (Smith

21
et ii., 1978; Brock, 1978; Brock ~~.t 1979), it provided an
opportunity to examine the role of fishes in determining invertebrate
colonization patterns. l Any shifts in these patterns on the experi
mental reef would have been an indication of the extent that reef
fishes influence sessile invertebrate colonization.
Following the methods of Wass (1967), rotenone was used as a non
selective means of removing the fish on the experimental reef. Smith
(1973) has detailed the use of this ichthyocide on coral patch reefs
and, along with various other authors (Randall, 1963; Clark et !l., 1968; Smith and Tyler, 1972; Nagelkerken, 1974; Russell ~!l., 1977)
has demonstrated its effectiveness in quantitative sampling of reef
fishes. Russell et ii. (1977) have shown that the effectiveness of
rotenone is enhanced and the results improved when the ichthyocide is
applied within the confines of some type of net enclosure.
On October 12, 1977, the experimental reef was enclosed with a
net measuring li m X 150 m with a stretched mesh size of 1 em. This
net was laid from a small skiff and then positioned along the nine
meter depth contour by a team of SCUBA divers. As the divers were
securing the net, 54 kg of 5% powdered rotenone was mixed with sea-
water in plastic bags to form a heavy paste. The ichthyocide was then
applied to the reef starting near the bottom of the net and working
upward along the reef slopes. Almost immediately, smail fishes such
lCaging of the panels was additionally considered but discarded because of the virtual certainty of attracting attention to the Danel sets and increasing the chances of someone disturbing the experiment in this popular area of Kaneohe Bay.

22
as gobies, e1eotrids, and apogonids began swimming erratically in the
water column. These fishes were quickly collected by divers using
hand nets and then put into buckets in boats waiting nearby. Within
an hour after the application of rotenone, most of the fishes on the
reef had succumbed to the ichthyocide; within six hours, the majority
of these had been collected.
The fishes were transported back to the laboratory, sorted into
groups, and then further separated by species. Each individual fish
was subsequently weighed, measured for standard lengths, and then
preserved in 10% formalin. Approximately 40 persons assisted in this
process as well as in the actual collection and other related portions
of the fish removal.
Visual transects were conducted prior to and after the fish
removal in an attempt to assess the changes in the resident fish
population. Visual methods for sampling reef fishes have been widely
applied to a variety of situations (Brock, 1954; Odum and Odum, 1955;
Bardach, 1953, 1959; Clark et &.., 1968; Risk, 1972; Collette and
Talbot, 1972; Key, 1973; Hobson, 1974; Chave and Ekert, 1974; Nolan
~!l., 1975; Gundermann and Popper, 1975; Grovhoug and Henderson,
1976). Shortcomings of visual censusing techniques have been dis
cussed by several authors (Wass, 1967; Chapman et ~., 1974; Jones
and Chase, 1975; Jones and Thompson, 1978; Russell et ~., 1977) and
basically consist of underestimating populations of nocturnal or
cryptic species. In this particular study, howe'/er, the collection of
fish from the reef provided the means for confirming the accuracy of
preceding fish counts. Brock (1978) estimated that 577 parrot fishes

23
(Scaridae) were in· residence on the day before defaunation and noted
that 545 were actually removed from the experimental reef. Visual
censusing for the majority of fish species on the experimental reef
was probably very accurate because of its small overall size and
isolation, both factors that tended to restrict movements of most of
the fish species.
Complete censusing of the experimental reef began two weeks
before the application of rotenone and continued through December 1978.
The usual procedure for censusing was for two divers to take separate
fish counts on the reef on the same day. Estimates for the overall
fish populations were derived from a comparison of these separate fish
counts.

24
RESULTS
Climatology
The meteorological data gathered during this study showed that
sunlight, wind speed and direction, and water temperature did not vary
dramatically from their long term norms. The amount of rainfall,
however, was less than usual.
The incident solar radiation values, recorded continuously
throughout the day, were highly variable as the result of rapid
changes in the amount of cloud cover. Because of this, monthly mean
values were calculated as a better indicator of the prevailing light
conditions for Kaneohe 8ay (Apoendix Figure A-2). An overall mean of
379 cal per cm2 per day was de~ived for the period from June 15, 1977
to June 15, 1978. This value agrees favorably with the long term mean
solar radiation value of 356 cal per cm2 per day calculated by
Yoshihara and Ekern (1978).
Because Kaneohe Bay is located within the zone of the northeast
trade winds, both the wind speed and direction are fairly constant the
year round (Appendix Figure A-3). The median windspeed during the
period of this study was 17.9 km per hour as compared to a calculated
long term median (1976 to 1979) of 18.9 km per hour for this area of
Oahu.
The water temperature at a mid-bay site approximately 150 m
downwind of the two study reefs followed the usual annual cycle

25
f 23 10 C t 27.5° C was (Appendix Figure A-4). The year's range a . a
typical for this subtropical embayment (Chave, 1973).
Monthly rainfall values were notably less than the average
monthly values recorded for previous years (Appendix Figure A-S). The
total accumulated rainfall from June 15, 1977 through June 15, 1978
was 78 cm, whereas 100 to 150 cm has been reported as the average
annual rainfall for the Kaneohe Bay watershed (Chave, 1973). Because
only an excess amount of rainfall would significantly alter the water
conditions in the central sector of the Bay, it is doubtful that the
lack of rainfall had any effect on invertebrate colonization on the
two patch reefs. Daily mean light conditions during the study period
were, if anything, slightly above average.
Long Term Communities
An analysis of the long term invertebrate communities was made to
substantiate trends of community composition along the windward to
leeward transects on the two patch reefs. The communities established
on the concrete blocks and the plastic tracks were used for this
analysis because of their convenience and easily quantifiable surface
areas. Although some authors have stated that invertebrate communities
do not reach equilibrium within one year (Sutherland and Karlson,
1977), others have concluded that stable communities are established
in less than 12 months (Osman, 1977; Schoener, 1974; Schoener, ~sl.,
1978). In this study, based on observations made at the time the
panel sets were taken off the reefs, the invertebrate communities
present after a 15 month exposure were similar to the contiguous reef

26
communities.
Numerous difficulties involved with the identification of all of
the fouling organisms down to the species level required that many
organisms be grouped into higher taxonomic divisions. This precluded
the possibility of calculating species diversity for the various sites,
but comparisons between the reefs and reef sites that involved
relative differences were still possible. Forty seven taxonomic
groups were represented on the panel sets after the 15 month exposures
(Appendix Table A-2) and a summary of these data shows that two trends
were apparent (Table 1). First, a greater variety (diversity) of
organisms occurred on the windward side of each reef and second, the
control reef sites possessed a greater variety of organisms than
comparable sites on the experimental reef. The strongest overall
pattern was for the windward sides of the reefs to accumulate a higher
percentage of the total taxonomic groups.
Counts of individual organisms or the area covered by colonials
revealed few patterns in the long term communities. Although corals
contributed significantly to the windward to leeward zonation on both
study reefs, this was not reflected by their settlement and growth on
the panel sets. Small heads of Poeillooora damicornis (less than 3 em
in diameter) appeared only slightly more numerous on the windward
sides of the two reefs and were most abundant on the panel set located
in the middle of the control reef. The on1y other organism that
showed a more favorable growth on the windward reef areas was a
species of didemnid tunicate. The calcareous alga, Poro1ithon
onkodes, was most prevalent on the middle and shallow leeward sides of

TABLE 1. Summary of taxonomic groups present on the panel sets after a 15 month exposure.
TOTAL NUMBER
OF GROUPS
PRESENT
% OCCURRENCE
OF TOTAL
(47) TAXA
SITE
1
2
3
4
5
1
2
3
4
5
EXPERIMENTAL REEF
32
32
25
26
24
68.1
68.1
53.2
55.3
51.1
CONTROL REEF
35
39
26
31
29
74.5
83.0
55.3
66.0
61.7
27

28
both patch reefs. Spirobid worms were most numerous on the shallow
leeward panel sets whereas the tube worm, Petaloconchus keenae, seemed
to favor the shallow sets on both sides of the reefs. The counts of
barnacles, bryozoans, and oysters did not show any definite patterns.
Algal Production and Grazing
Because chlorophyll a is a phytopigment common to all marine
algal groups (Dawson, i966), the results from the analyses for this
pigment were used as .indicators of algal production on the fouling
panels. Inter-reef and site by site comparisons of algal production
were made by utilizing the chlorophyll a quantities as relative
measures of algal biomass (Figure 5).
Statistical analysis of the possible comparisons for chlorophyll
a was performed using nonparametric one-way analysis of variance
(ANOVA). The use of this nonparametric method, the Kruskal-Wallis
Test, was necessary after Levene's Test for equality of variances
demonstrated that the variances within these data were not homogeneous
(P<.Ol). Levene's Test was considered aopropriate because this test
can be applied to data that are not normally distributed and it is
more robust than the often used Bartlett's Test (Levene, 1960; Zar,
1974). The Kruskal-Wallis Test showed that chlorophyll a values were
significantly different for the ten reef sites (P<.Ol; Table 2).
Post-test comparisons of the chlorophyll a values at each site
were performed to elucidate patterns in the data. A nonparametric
multiple range test suggested by Zar (1974) showed that tne amount of
chlorophyll a was not significantly different at the windward crests

FIGURE 5. Microgram-atoms of chlorophyll a present on
100 cm2 of upper panel surface area.
(-----control reef, --.....texperimental reef)
29

sao
200
100
• n
UPPER PANEL SURFACE SITE 1
, , , \ , \ , , , , , .... I ..
, I I' I , J'" './"
04---~~~~--~--~~---r--~--r-~---r~
a: 400
...J
...J >- 300 :c 0-0 200 a: o ...J :c 100 U
SITE 2
~ O~--~~~~--~--~~~~--~--~~---r~ en 200 L o t- 100 a:
I L a: a: 200 C!) o ~ 100 ~
:::::
SITE 3
SITE 4
O~--~~---r--~--~~---r--~--r-~---r~
200 SITE 5
100
0~--~-7~~--r-~---r--~~---r--~,--,--, JUN AUG OCT DEC fEB APR JUN
1977 1978
30

TABLE 2. Kruska1-Wa11is non-parametric ANOVA and the corresponding multiple comparison test for chlorophyll a data.
Kruskal-Wa11is Test (Chi-square approximation) = 168.49
X2.01,9 = 21.67 P < .01
---------------------------------------------------------------------------------------------------
n 50 49 50 52 50 48 49 51 49 52
REEF:SITE C:5 E:5 E:1 C:4 C:3 £:3 E:4 C:l ~ E:2 C:2
RANK SUM 6505 9164 9578.5 10312.5 10986.5 11401.5 11447.5 16662 19312 19880
Underlined sites indicate no significant difference at the .01 level
w --'

32
of both study reefs (P<.Ol). Chlorophyll a values were significantly
different at the deep windward and deep leeward sites on the control
reef but were not significantly different among all of the other reef
locations (P<.Ol). Overall, algal production and the relative amounts
of algal biomass were highest at the windward reef crests and signifi
cantly less at all other patch reef sites.
The relationship between the distribution of algae on coral reefs
and grazing by both fishes and invertebrates has been demonstrated by
several authors (Stephenson and Searles, 1960; Randall, 1961; Bakus,
1967, 1969; Dart 1972; Vine, 1974). To indicate the relative grazing
pressures exerted by fishes and invertebrates at each study site, the
grazing index data were aVeraged to give single monthly values. These
monthly grazing indices clearly illustrate the fact that the least
amount of grazing took place at the windward crests of the two study
reefs and the greatest grazing pressures occurred on the two mid-reef
sites (Figures 6 and 7). Fishes were the major agent of these grazing
pressures, but invertebrates played a significant role in the grazing
at both of the deep leeward panel locations.
Colonization of Panels
More than 80% of the invertebrate colonization on the two patch
reefs could be attributed to five taxonomic groups: oysters, barnacles,
serpulid worms, tunicates, and bryzoans. The temporal and spatial
patterns of invertebrate colonization on the study reefs are described
on the basis of these five groups.

FIGURE 6. Histograms of monthly grazing indices for the
upper panel surfaces on the experimental reef.
33

-l.!..J 0::
. 0.X W
x w a :z -C!> z ...... N a: 0:: C!>
5
• 3
2
1
0
:1 3
2
1
a 5
• 3
2
1
0 J J
!IIIIJ FISH 0 INVERTEBRATES
, A S (j N 0 J FHA H J
1977 1978
W I-....... <n
W I-....... <n
34

FIGURE 7. Histograms of monthly grazing indices for the
upper panel surfaces on the control reef.
35

-I..L... 3
t:j 2 a:: 1
. ~ 5 :z: 4: U
3
x 2 w
lIIII1 FISH D INVERTEBRATES
-W
II-...... U')
W t-...... en
W t-...... en
o 1 :z _ O~~~~~~~~~~~~~~
e:> 5 :z: 4: -N S a: CC 2 e:>
1
1 .4
3
2
1
O~~~~~~~~~~~~~~ 11 A Ii J
.1977 19713
If)
W t-....... en
36

37
Oysters
Bivalve molluscs, especially oysters, were the most abundant
sessile invertebrates to colonize the panel surfaces. During the year
of sampling, four species of bivalves were recognized among the more
mature specimens on the panels: Ostrea sandvichensis, Anomia nobilis, .
Hiatel1a hawaiiensis, and a species of Isognomon. Of these four
species, the only frequent colonizer was the oyster Q. sandvichensis.
Significant numbers of !i. havraiiel'1sis settled only occasionally on the
panels, whereas specimens of A. nobilis and IsoQnomon were very seldom
encountered. The frequency of occurrence makes it practical to dis
cuss the colonization trends of only the oysters.
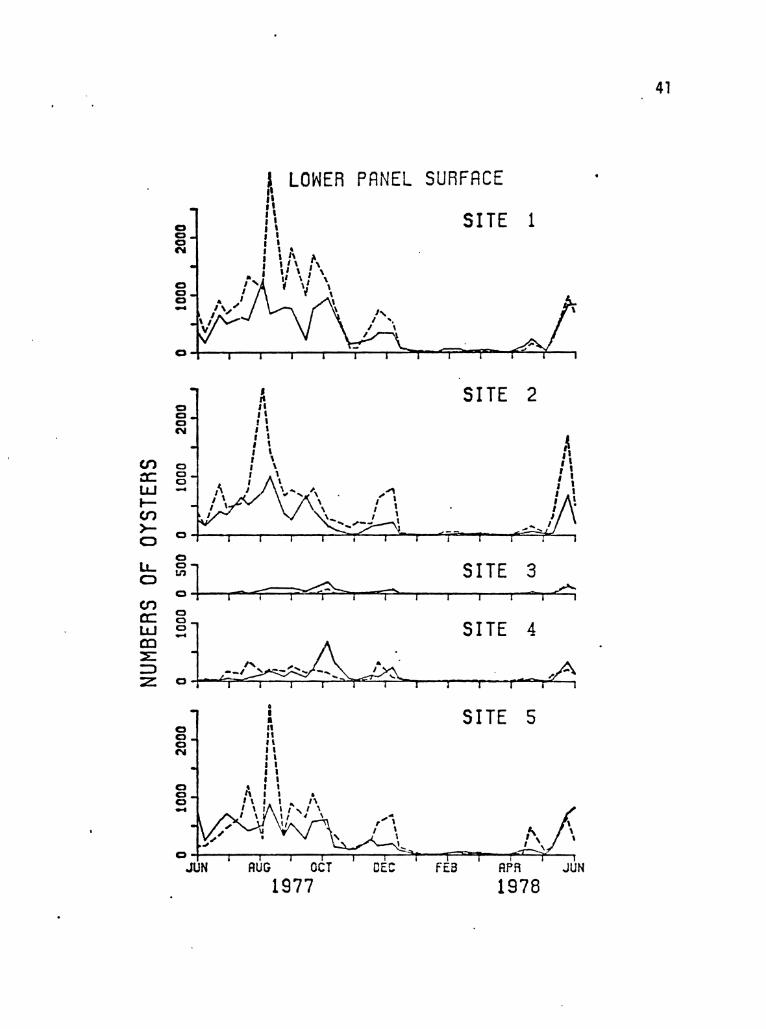
Several patterns are appare~t ~n the oyster data (Figures 8 and
9). First, virtually no colonization took place at any of the reef
locations during the months of January, February, March, and April.
Secondly, essentially no colonization occurred on the upper panel
surfaces at sites three and four, the middle and shallow leeward reef
locations. These two sites did incur some settling by oysters on
their lower surfaces, but generally this was much less·than at Jny of
the other sites on the two reefs.
To completely describe the colonization patterns of oysters on
the study reefs, it was deemed appropriate to determine whether there
were any significant differences between sites one, two, and five, and
whether there was a significant differe~ce in colonization between the
two reefs or between the upper and lower panel surfaces.
After excl udi ng the months of January, February, ~larch, and

38
FIGURE 8. Numbers of oysters per 100 sq cm found sett ling en
the upper panel surfaces after a three week exposure.
(------control reef, ---eexperimenta 1 reef)

en a: w ~ en >o IJ.. o en a: w. (Q
~ :::J Z
o o c C'oI
o o C) -
UPPER PANEL SURFACE
, il • t, ,>.I
1\:,1 'J II. " I~ , ,. J ' I,
I "' ~,
'\' 'V ~ \ \ " t'lr' It A \ , " " 0 .... ~
o o o N
o o o -
o o o ("If
o o o -
SITE 1
SITE 2
SITE 5
o4-~~~~~--~~~~~~--r-~---r--~~ JUH AUG OCT DEC FEB APR JUH
1977 1978
39

FIGURE 9. Numbers of oysters (Ostrea sandv;chens;s) per
100 sq cm on lower panel surfaces after a three
week exposure. The middle panel of each set has
been excluded.
(-----control reef, --~experimental reef)
40
41

42
April and sites three and four from the analysis, the remaining data
cells contained sufficient variability to make possible a parametric
three-factor analysis of variance (Table 3). Using dates for repeti
tive measures (see Winer, 1971), this analysis led to the conclusion
that the numbers of oysters colonizing both reefs were not signifi
cantly different (P>.Ol). In addition, the numbers of oysters were
not significantly different for both panel surfaces (P>.Ol). The
quite obvious inter-site differences have already been stated and were
the basis for excluding sites three and four, but the analysis pointed
out a significant difference in the amount of colonization at the
remaining three sites (P<.01). Because sites one and two, on the
windward side, accumulated more oysters than anywhere else on the
reefs, this difference may be attributed to the lower numbers of
oysters settling at the deep leeward sites. All second and third
order interactions between the reefs, sites, and panel surfaces were
significant (P<.Ol) indicating that within a particular site or a
particular reef, differences existed in the numbers of oysters colo
nizing the two panel surfaces. Conversely, Significant differences
occurred between the control reef and the experimental reef with
respect to the panel surfaces. The importance of site specific
conditions is indicated by these significant interactions.
Overall, the oysters showed a strong tendency to colonize dif
ferent sites on the two study reefs and seemed to parti cul arly favor
the windward areas. With a couple of exceptions, there did not appear
to be any general tendency for oysters to colonize upper panel
surfaces more often than lower panel surfaces or vice versa.
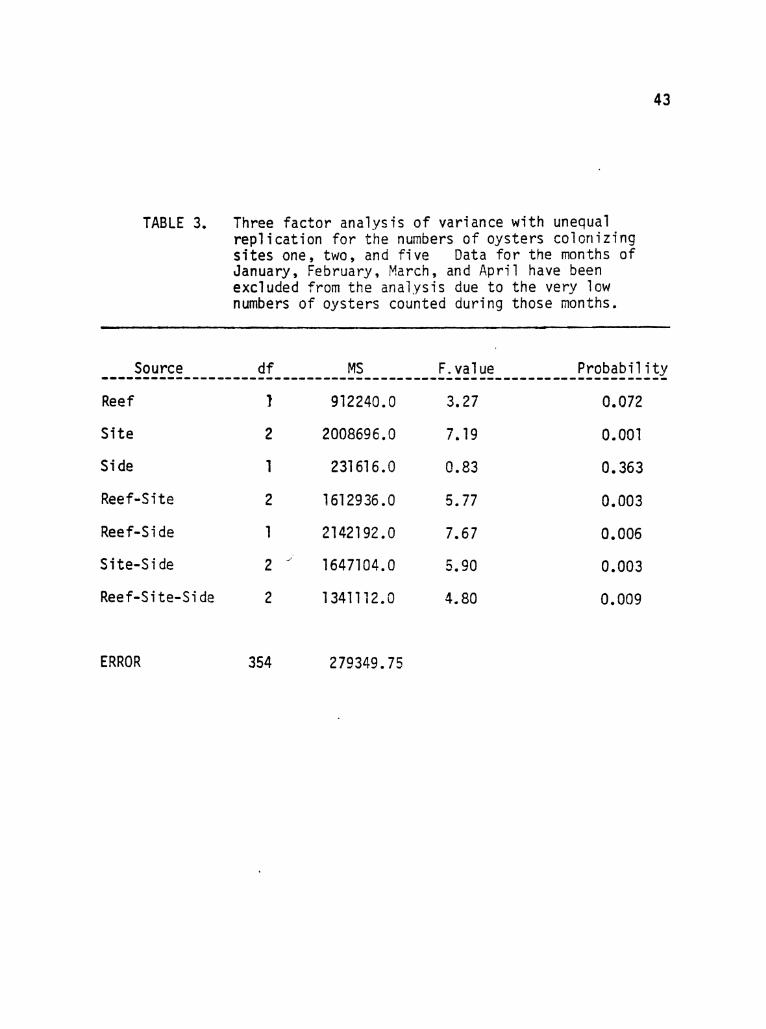
Reef
Site
Side
TABLE 3. Three factor analysis of variance with unequal replication for the numbers of oysters colonizing sites one, two, and five Data for the months of January, February, March, and April have been excluded from the analysis due to the very low numbers of oysters counted during those months.
1 912240.0
2 2008696.0
1 231616.0
Reef-Site 2 1612936.0
3.27
7.19
0.83
5.77
7.67
0.072
0.001
0.363
0.003
0.006
0.003
0.009
Reef-Side
Site-Side
Reef-Si te-Si de
ERROR
1 2142192.0
2 -' 1647104.0
2 1341112.0
354 279349.75
5.90
4.80
43

44
Similarly, although differences in settling existed at specific sites,
over the entire reef transects oysters colonized both reefs equally on
a per unit area basis. The colonization of the patch reefs in Kaneohe
Bay apparently followed an annual reproductive cycle with seasonal
peaks occurring in September and Octcber and very little settling by
the oyster larvae from January through April.
Barnacles
Barnacles are among the most widespread and common fouling
organisms in marine waters of the world. On the study panels, these
highly specialized crustaceans were represented by at least three
species of Balanus: !. amphitrite, £. eburneus, and B. reticulatus.
During the course of this study, a fourth species, ~. triqonus, was
occasionally encountered. Based on the frequency of occurrence of
mature specimens, the most abundant barnacle species to appear on the
panel surfaces was ~. amphitrite. The difficulties involved with
identification of newly settled barnacles made it necessary to group
all of the individuals into the single genus.
The sporadic appearance of very high numbers of barnacles required
the use of a semi-logarithmic scale to best represent the colonization
trends (Figures 10 and 11). Although this scale accentuated low
numbers of individuals and de-emphasized the peak periods of coloni
zation, the overall trends were still apparent. The panels at sites
three and four accumulated very few newly settled barnacle cyprids.
The panels at these sites seldom collected more than ten individua1s,
whereas counts of barnacles at the other three sites were often more

FIGURE 10. Numbers of Balanus per 100 sq em of upper panel
surface area.
(-----contro1 reef, ---t:!experimental reef)
45

1000
100
(f') 10 l1J -l
UPPER PRNEL SURFACE
SITE 1
SITE 2
~ 1·~,~~~~~~--~~~-r~~~T-~~~~~--~
~1000 a:: a: 100 CO
l.1.. 10 (:)
SITE 3
\ (f') l~--~~--~--~~~~~~~~--~--T-~~~
ffi 1000 CD ::E: 100 :::a z:
10
SITE- 4
", .t. I " • \ , \' I.:::..!...l...,
, \ I \ Y ,,\ l+---~~---r~~~~~~~~~~~--~~---'
1000 SITE 5
100
10
1+L~~~~-r~~~~~~~~~~~~~----~
JUH RUG OCT DEC fEB RPA JUU 1977 1978
46

FIGURE 11. Numbers of Balanus per 100 sq em on the lower
side of the foul~ng panels.
(-----control reef, ---eexperimental reef)
47
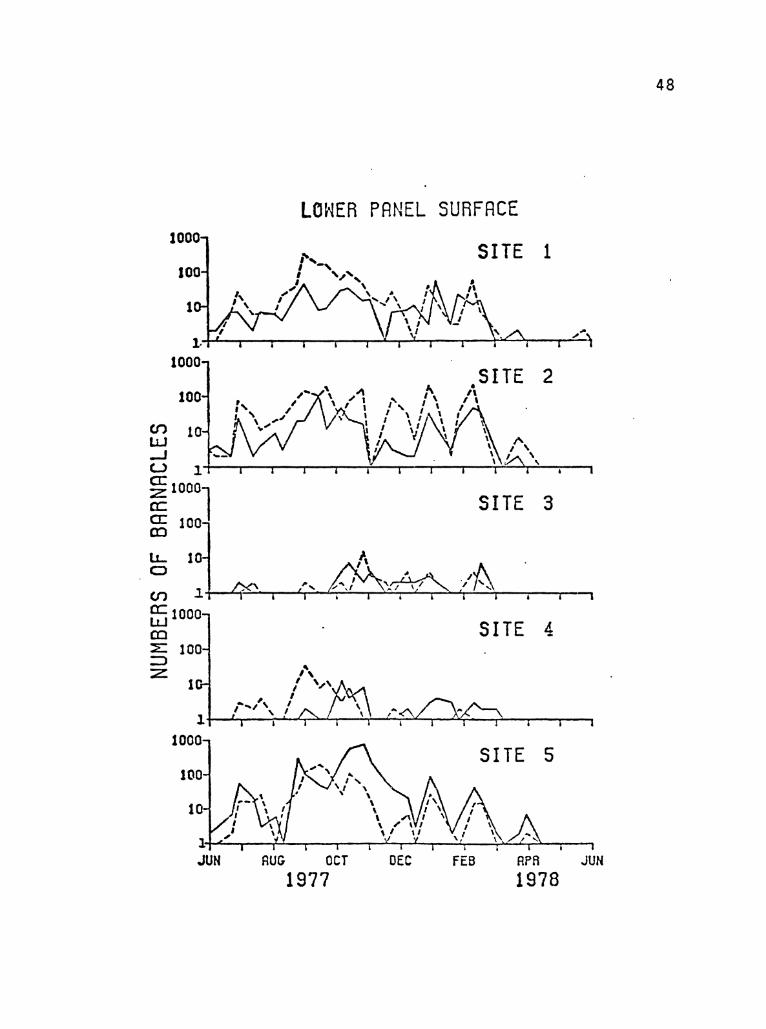
48
LOWER PANEL SURFACE 2000
SITE 1
1

TABLE 4.
UPPER
PANEL
SURFACE
Mean numbers ± 95% confidence intervals of Balanus per 100 sq cm on the fouling panels during the one year sampling period.
SITE EXPERIMENTAL REEF CONTROL REEF
1 • S± .31 2.2±1.29
2 3. 7±1. 74 9.1±S.08
3 • 1 ± .05 .1± .13
4 .2± .14 .S± .38
S 4.2±2.20 1. l± .60
49
---------------------------------------------------------------------
LOWER
PANEL
SURFACE
1
2
3
4
5
9.0±4.36
12.0±6.98
.7± .25
.9± .75
72.6±S3.50
32.0±22.31
45. 9±21. 73
.9± .82
2.2± 2.01
22.2±14.80

50
than ten, especially on the lower panel surfaces and during the months
from late August to March.
Further patterns of colonization were apparent from comparisons
of the mean numbers of barnacles for each site and for each side of
the panels (Table 4). More barnacle larvae tended to colonize the
lower surfaces of the panels at all sites and the lack of cyprids
settling at sites three and four was obvious. The significance of
these patterns was indicated by the fact that the 95% confidence
intervals for these comparisons did not overlap. Other trends in the
data were also suggested. For example, barnacles tended to be more
numerous on the windward sites of the control reef, whereas more
Balanus settled on the deep leeward site of the experimental reef.
With the exception of the deep leeward site, more barnacle cyprids
settled out over a given area on the control reef than on the experi
mental reef.
In summary, barnacles attached more often on the undersurfaces
of the fouling panels than on the upper sides and there was a general
propensity for barnacle cyprids to settle along the windward areas of
the two patch reefs. Significant colonization also occurred on the
1eeward sides of the study reefs at a depth of three meters. At the
other sites, the control reef accumulated more individuals per unit
area than the experimental reef. Recruitment of barnacles to the
reefs was heaviest from September through March with noticeably fewer
cyprid larvae settling out of the water column between the months of
April and June.

51
Serpulid Worms
Calcareous tube-dwelling anne1ids (family Serpulidae) are
another prevalent and quite often abundant group of sessile marine
invertebrates. On the study panels, serpulid worms were usually
among the more conspicuous fouling organisms. Two species of
Hydroides, ~. elegans and ~. lunulifera, were the largest of these
polychaetes with the more mature specimens forming tubes 15 to 20 mm
in length. Three members of the subfamily Spirobinae, Janua pseudo
corrugata, Janua steuri, and Pileo1aria pseudomi1itaris were the
smallest of the calcareous tube worms, forming tiny spirals less than
2 mm in diameter. Other species of serpulids occasionally were found
on the panels but their numbers and frequency of occurrence made them
an insignificant part of the total fouling communities. For purposes
of this study, the serpulid worms were split into two groups; the
Hydroides spp. and the Spirobinae.
The mean numbers of Hydroides and Spirobinae show that very
little colonization by both types of serpulids occurred on the upper
panel surfaces (Tables 5 and 6). There was no overlap of the 95%
confidence intervals for this comparison. Because of this, the data
for serpulids on the upper panel sides have been omitted.
Two conspicuous temporal patterns were noticeable in the
calcareous tube worm data (Figures 12 and 13). First, Hydroides
appeared to exhibit a six to eight month cycle of peak abundance
(Figure 12). Care must be taken in interpreting this pattern because
the data represent two distinct species and the apparent periodicity

TABLE 5. Mean numbers ± 95% confidence intervals of Hydroides per 100 sq em on the fouling panels during the one year sampling period.
52
SITE EXPERIMENTAL REEF CONTROL REEF
1 1.5±.86 1.9±1.25
UPPER 2 .S±.51 .4± .25
PANEL 3 O±O • 1± .04
SURFACE 4 O±O • l± .04
5 1.3±.73 1.0± .65
---------------------------------------------------------------------
1 17.8±6.05 9.7±3.77
LOWER 2 24.7±8.25 15.8±6.15
PANEL 3 12.0±7.97 2. 3±1. 58
SURFACE 4 9.0±3.67 2. 9±l. 63
5 19.6±7.71 11. O±4. 95

53
TABLE 6. Mean numbers ± 95% confidence intervals of Spirobinae per 100 sq cm on the fouling panels during the one year sampling period.
UPPER
PANEL
SURFACE
LOWER
PANEL
SURFACE
SITE
1
2
3
4
5
1
2
3
4
5
EXPERIMENTAL REEF
.2± .31
.4± .76
O±O
• 1± .04
1.1±1.93
274.3± 57.70
90.S± 28.77
227. 3± 11 O. 19
8S.2± 33.06
lS7.6± 38.78
CONTROL REEF
.2±.17
• 1 ±. 04
O±O
• 1 ±. as
o±o
12.6± 3.17
16.9±12.28
2S.3±14.75
B.6± 2.83
Sl.0±27.73

FIGURE 12. Numbers of Hydroides per 100 sq em on the under
side of the fouling panels.
( experimental reef, -----control reef)
54

55
LOWER PANEL SURFACE 100 SITE 1 7S
50
25
a 100 SITE 2 75
50
(J") 25 w Cl a ...... o 100 a: ~ 75 ::r:: I.l.... 50 0
25 (J")
a: 0 w CD 100 :L SITE 4 => :z 75
50
2S
a 100
5 75
50
25
0 JUN AUG OCT DEC FEB APR JUN
1977 1978

FIGURE 13. Numbers of Spirobinae per 100 sq em on the under
side of the fouling panels.
( experimentai reef, -----eontrol reef)
56

57
LOWER PANEL SURFACE 750 SITE 1
sao
250
a -,-----.., .......... ----- -- -------. 500 SITE 2
250
a I.LJ 1250 SITE 3 < z -ۤ 1000 c: -0-en 150 u.. 0
sao t.f) c: UJ a:) 250 ~ ::l • z: " ,,.,--,"--....
... , I
0 ." .... _-500 SITE 4
250
0 - --- -- -_ ........... . 500 SITE 5
250
,-0 JUN AUG OCT DEC FES APR JUN
1977 1978

58
could represent separate peaks for each species. The second temporal
pattern, particularly dramatic at sites three, four, and five was the
increase in numbers of Spirobinae toward the end of the study period
(Figure 13). This trend does not appear to be a seasonal phenomenon,
at least not with respect to the time scale of this study.
In most instances, more Spirobinae and more Hydroides were found
on the experimental reef than on the same r~spective sites of the
control reef. This is a reversal of the pattern shown by the barna
cles. Although definite differences occurred between sites, there was
no real obvious windward to leeward colonization pattern for any of
the serpulid worms, nor was there a clear pattern of settling that
related to the depths of the fouling plates.
Reiterating the stronger and more consistent trends, serpulid
worms preferred the shaded undersurfaces of the panels for their
colonization. Hydroides displayed some seasonality whereas the
Spirobinae colonization was more aperiodic. Furthermore, the data
indicated serpulid worms were more abundant on the experimental reef
than on the control reef.
Tunicates
Tunicates are another group of cosmopolitan marine invertebrates
that were usually present and conspicuous on the study panels. Both
colonial and solitary forms were represented and were among the
largest organisms found on the three-week-old panels. It was not
unusual for single colonies to cover 4 to 6 sq cm, whereas solitary
tunicates reached sizes of just over 1 cm. Solitary tunicates were

59
usually a species of Ascidia; either A. sidneiensis or A. interruota.
Of these two, A. sidne1ens1s was the most conmon. Colonial tunicates
included Botryloides ~. and Polyclinum ~~, Trididemnum savignii,
Didemnum edmondsoni and Didemnum candidum. Neither the Botryloides
or the Polyclinum species occurred very frequently, but the didemnid
tunicates were almost always prominent components of the newly formed
invertebrate communities. For purposes of this study, the only colo
nization trends analyzed were those of the most abundant tunicates,
the didemnids (taking all species collectively) and A. sidneiensis.
Virtually no colonization occurred for Ascidia sidneiensis on the
upper panel surfaces (Table 7). The colonial forms, the didemnid
tunicates, were also noticeably absent from the top sides of the
panels (Table 8). Both colonial and solitary tunicates were more
prevalent on the panels of the control reef than they were on the
experimental reef.
Colonization by the solitary tunicate, Ascidia sidneiensis,
reached a peak in late June to early July with possibly a smaller peak
about six months later in January and February (Table 9). Most of the
solitary tunicates appeared on the windward sides of the reefs and
only once did A. sidneiensis settle on the underside of the mid-reef
panels (Table 7).
The didemnid tunicates appeared on both reefs from June through
November and displayed a peak around September (Table 9). Colonies of
didemnids were essentially absent on all of the panels except for
those at the windward reef sites (Table 8).

TABLE 7. Mean numbers ± 95% confidence intervals of Ascidia sidne;ensis per 100 sq em of surface area on the fouling panels.
60
SITE
1
EXPERIMENTAL REEF
o
CONTROL REEF
o
UPPER
PANEL
SURFACE
UPPER
PANEL
SURFACE
2
3
4
5
1
2
3
4
5
O.2±.25
o
o
o
2. 1±2. 35
3.1±3.11
O.O±.06
O.S±.68
1.O±1. 16
o
o
o
O.2±.32
6.1±5.40
2.8±2.13
o O.7±.99
1.l±1.22

61
TABLE 8. Mean numbers ± 95% confidence intervals of didemnid tunicates per 100 sq em of surface area on the fouling
UPPER
PANEL
SURFACE
LOWER
PANEL
SURFACE
panels.
SITE
1
2
3
4
5
1
2
3
4
5
EXPERIMENTAL REEF
o
o
o
o
o
O.l±.ll
O.S±.34
O.0±.06
0
0
CONTROL REEF
o
o
o
o
o
1.0±.36
2. 3± 1. 27
O.O±.06
0
O.O±.O6

TABLE 9. Monthly mean numbers of tunicates on the lower panel surfaces from June 1977 to June 1978. The didemnid tunicates are expressed as percent coverage.
ASCIDIA SIDNEIENSIS DIOEMNIDS
62
EXPT. REEF CNTL. REEF EXPT. REEF CNTL. REEF
JUNE 13 15 .20 .50
JULY 5 10 .07 1.13
AUGUST 1 1 • 13 1.67
SEPTEMBER 0 0 .33 1.80
OCTOBER 1 0 .66 .80
NOVEMBER 0 0 .07 .80
DECEMBER 0 1 .00 .53
JANUARY 1 0 .00 .20
FEBRUARY 0 3 .00 .13
MARCH 0 0 .00 .67
APRIL 0 0 .00 .00
MAY 0 1 .00 .07
JUNE 0 1 .00 .10

63
Bryozoans
Bryozoans were the single most diverse taxonomic group of organ
isms found on the study panels. They were present throughout the year
and were usually very abundant. The bryozoans were represented by a
variety of growth forms ranging from small, rather inconspicuous
colonies of Aetea truncata to larger patches (approx. one sq cm) of
encrusting Schizoporella species. In between this range, specimens of
Bugula neritina, Bugula californica, HoloDorella ~., Microporel1a
~., Savignyel1a lafont;, and Watersipora~. appeared en the panel
surfaces. Although an attempt was made to count the more easily
recognized species, most of the bryozoans were difficult to distinguish
and their presence was usually only noted as being of a particular
growth form.
Bugula neritina, Savignyella lafonti, and Microporel1a~. were
three of the more readily identifiable species. Other bryozoan's were
counted as either encrusting or erect forms. During the course of this
study, essentially none of the bryozoans ever settled on the upper
panel surfaces. This is consistent with the fact that they are
usually found semi-concealed in cracks and crevices of coral reefs.
Savignyella lafont;, although relatively easy to identify, was
never very abundant on the study panels. The maximum number of 312
colonies of this bryozoan occurred on site two of the control reef in
late September. With the exception of this one count, the number of
colonies of i. lafonti never exceeded 150 on any panel and were
usually less than ten for a11 sites during the year. The peak co10n;-

64
zation period for i. lafonti was from mid-August to early November
with the colonies only appearing on the windward sites (sites one and
two) of both study reefs.
Colonies of Microporella were consistently more numerous on the
experimental reef than on the control reef (Figure 14). Although this
bryozoan was never very abundant at any time during the year, it
reached its peak colonization during the period from September to
February. All of the sites on the reef recruited approximately the
same numbers of Microporella except for site three where the colonies
were rarely present.
Like the previous two species of bryozoans, colonies of Bugula
neritina were never particularly numerous (Figure 15). When they were
present, these colonies consistently appeared in greater quantities
on the control reef than on the experimental reef. Bugula neritina
was most prevalent on the deep windward sides of the reefs and
exhibited peaks at what appeared to be approximately three month
intervals. These peaks were not sharpiy defined and may have been an
artifact of the sampling during this study.
Other Invertebrates
Aside from the five invertebrate groups just discussed, sponges
and hydrozoans were the only other sessile invertebrates of any con
sequence to appear on the study panels. Sponges, mostly encrusting
types, were occasionally conspicuous components of the three week old
communities. Hydrozoans, noticeable on freshly collected panels,
generally did not remain intact during the drying process and so were

FIGURE 14. Numbers of Microporel'a spp. per laO sq em on the
lower side of the fouling panels.
(-----control reef, ---eexper;mental reef)
65

LOWER PANEL SURFACE 100 SITE 1 7S
so
2S
o+---~~~~=-~~~--~~~~r-~
100
• Il- 75 Il-(f)
a: -'
so
-' 25 UJ a:: o Il-
SITE 2
§ 2:]+~~~~_<)~~~~~IQ~~i _S~:_T_E~3~-, L 75 lJ... o
50
100
75
50
25
SITE 4
SITE 5
O+--r~--~~=-~~~--~~~~~~
JUN AUG OCT DEC FEB APR JUN
1977 1978
66

FIGURE 15. Numbers of Bugula neritina per 100 sq em on the
lower side of the fouling panels.
(-----control reef, --~experimental reef)
67

125
100
75
50
25 a: :::z: -t- O -125 a: w :::z: a: 100 --J ::> c!) 75 ::> CD
IJ.. 50 0
en a: 25 w CD ~ ::> 0 :::z:
] ] ]
JUN
LOWER PANEL SURFACE SITE 1
I -I, '1 I, I,
I I I. , . •• • • I I I • I I f • I • : ' ....
J.: \,,, f, I 'II I I I ,I I I \ I •
, " I , "
• • • " f. " t f • ~ I • .' I I ,\ , , f ' I , • I , • I I I , I '
: ~ l\ : \:, I I '\ I I I , I I I \ I '..t " :\
, ,I , I
: f ~I I 1.1, I J "I , , "\ I , I
I " 'I " J" ... J '. __ ... ' .., I ,
,
SITE 2 , = ,I
" " " I' I' I' ,
• I I • • • , I , I I , • • I I I
SITE 3
eC'> .---I r
SITE 4 ~>=t~
i , t '" -4 .--' .... < '1-
AUG OCT DEC 1977
SITE 5 ~ < .... , -,..,....<,~
iii
FEB RPR 1978
JUN
68

69
noted only as being present or as being present and abundant.
Motile invertebrates such as amphipods, copepods, and gastropod
molluscs were almost always found on the panels and were sometimes
very abundant (Appendix Table A-3). Although their presence on the
panel surfaces almost certainly had some influence on the overall
fouling communities, this aspect of invertebrate colonization was
beyond the scope of this study and no attempt was made to analyze the
data for motile invertebrates.
Experimental Reef - Oefaunation
More than 4500 fishes were removed from the experimental reef on
October 12t 1977 (Appendix Table A-4). A visual survey conducted on
the day after the fish removal supported the contention that more than
95% of the resident fishes had been collected. Eight small octopus
and four banded coral shrimp (Stenopus hispidus) were killed but the
rest of the invertebrates on the reef appeared to be unaffected by
the rotenone. The fish collected from the experimental reef were
identified and assigned to feeding categories based on the work of
other authors (primarily Hobson, 1974 and Gosline and Brock, 1960) or
on stomach contents (Table 10).
In te~s of biomass, planktivores dominated the patch reef com
munity cow.prising over one-half (56%) of the total weight of fishes
on the reef while accounting for over one-fourth (29%) of the numbers
of individuals. Most of the planktivores (81%) were either the
butterflyfish, Chaetodcn miliaris, or the damsel fish, Abudefduf
abdominal is. Together these two species comprised 90% of the total

TABLE 10. Summary of fishes collected by rotenone from the experimental reef on October 12, 1977. The feeding categories were based on previous literature (primarily Hobson, 1974) or stomach analysis.
No. of No. of Wt. % by % by Species Ind. (kg) No. Wt.
CARNIVORES 50 1586 42.46 34.3 30.8
PLANKTIVORES 10 1336 77.08 28.9 55.9
HERBIVORES 11 620 14.92 13.4 10.8
OMNIVORES 2 58 2.12 1.3 1.5
DETRITIVORES 3 1007 1.22 21.8 0.9
CORAL POLYP OR MUCOUS FEEDERS 1 1 0.01 0.02 0.01
UNDEFINED FEEDING TYPES 4 18 0.01 0.4 0.1
70
---------------------------------------------------------------------TOTAL 81 4626 137.94 100. 100.

71
weight of planktivores. Overall, 24% of the fishes collected and 50%
of the total wet weight of fishes removed from the experimental reef
were either £. miliaris or A. abdominal is.
Carnivores were the most numerous feeding type collected both in
terms of numbers of individuals and in numbers of species. By weight,
they comprised 31% of the total fish population and numerically, 34%
of the fishes were carnivores. No single species of carnivore con-
tributed much more than 5% to the overall fish population in terms of
weight, but the cardinal fish, Faa brachyqramma, accounted for 24% of
the total number of fishes collected. As a group, the eels (family
Muraenidae) made up 13% of the total fish weight on the experimental
reef.
Approximately one out of every seven fishes on the experimental
reef (13%) was a herbivore. Parrotfishes (family Scaridae) were the
dominant herbivore collected from the reef and accounted for 72% of
the wet weight within this group. Overall, herbivores comprised 11%
of the total weight of fishes in the collection.
Recolonization of the reef in the weeks following the removal of
the fishes was surprisingly rapid (Figure 16). The curve resulting
from the visual transect data may be described by the following
equation:
n
where Y ;s the expected number of species, Noo is the asymptotic number
of species, K is a constant, and t ;s the elapsed time in number of
days. This equation is mentioned by MacArthur and Wilson (1967) as
best describing the colonization curve for insular situations. It was

72
fitted to the data using an iterative least-squares computer program
of the Statistical Analysis System (SAS). Starting points for the two
parameters, Noo and K, were initially estimated using a method described
by Rafail (1973) and were used as inputs to the program. The follow
ing equation resulted for numbers of species recruited to the reef:
Y = 70 (1_e-·0036t)
The goodness of fit for this equation was determined with the method
given by Zar (1974) for calculating the coefficient of determination,
~:
~ = 5S regression 5S total
This curve provided an excellent fit to the visual transect data,
yielding a value for the coefficient of determination of .92.
The fitted colonization curve predicted that 645 days were
required to recruit 90% of the fish species to the experimental reef.
Although this time period was relatively long with respect to the
duration of the study and invertebrate colonization, initial recruit-
ment to the reef occurred more rapidly than predicted (Figure 16).
More significantly, recolonization of the reef by certain species of
fish occurred much faster. The visual transects revealed that in less
than one month after removal, over 400 individual fish, representing
14 different species, had returned to the reef. More than 75% of
these individuals were small parrotfishes, the dominant species of
herbivore. The numbers of these parrotfishes on the reef 28 days

FIGURE 16. Colonization curve based on visual counts of fishes
present on the experimental reef after complete
removal of the popuiation by the application of
rotenone.
73

II , \ , , , , , , , , , , ,
\ , \ , , \ , , El , ,
\ , , , , IJ \ , , ,
.\ \ , l\ , ,
\ , \ \ EJ , ,
C \ ... \ C ILl , Q> \ -= \EJ ... ~ uu ... \ ... 2: , :z: a , ~;: , ... co: " ... , ..J- , a:: :z: , is " _0
, >&.0 ,
EJ , " , ,
" II " " , EI
II EJ
II EJ
0 0 0 (0 ~ N
S3I:J3dS .dO 838WnN
" , " " " " " " " " I ElEl " " Elq.:
0
o C) ~
0 C) (r)
0 C) ("\1
C)
0 -
0
74
z C) -t-a: z :::> a: lL. w 0
W U Z ....... en en >-a: Cl

75
after the fish removal (365 counted) exceeded the number actually
collected from the reef. The parrotfishes overshot their original
population numbers and reached a peak of 963 individuals on the
experimental reef 258 days after the fish removal. The influence of
these herbivores was noticeable in the grazing marks that they left
on the panel surfaces. Because of the rapid return of herbivores,
the monthly grazing indices were not markedly affected by the fish
removal (see Figures 6 and 7). The planktivores, carnivores, and
other feeding types returned to the experimental reef much more
slowly. After 28 days, there were only 15 Abudefduf abdominalis on
the reef and two Chaetodon miliaris. This represented less than 3% of
the original populations of these dominant planktivores.
A more extensive discussion of the recolonization data, including
the colonization rate curve and a discussion of the stability of the
fish population on the experimental reef may be found in Brock, ~~.
(1979).

76
DISCUSSION AND CONCLUSIONS
The data accumulated during this study show that sessile marine
invertebrates exhibited definite patterns in their colonization of
coral patch reefs. These colonization trends can be broken down into
four basic categories: (1) seasonal or periodic variations, (2)
differences in panel exposure (upper versus lower surfaces), (3) vari
ations between reefs, and (4) site by site differences. Many of the
colonization patterns that have emerged during this study are not
surprising in light of the previous research on marine fouling
communities.
In Edmondson and Ingram's (1939) paper on fouling organisms in
Hawaii, they determined that the principal organisms responsible for
most of the fouling in Kaneohe Bay were barnacles, bryozoans, serpulid
worms, oysters, and tunicates. These same five groups of organisms
represented the major colonizers of the panels on the two study reefs.
Edmondson and Ingram (1939) pointed out several settling patterns
exhibited by these taxa including fewer barnacle settlements from
January through March, a lower abundance of Bugula neritina during the
winter months, the sporadic appearance of serpulid worms, minimal
reproduction of oysters in winter and the overall propensity for most
of the benthic invertebrate taxa to colonize shaded surfaces. With
the exception of the time of peak colonization for barnacles, all of
these trends have been substantiated by the data collected in the
present study.

77
The disparity in season of attachment for barnacle cyprids can
De attr1 Duted to ttle di rrerences in the 1ocation of the study sites
within Kaneohe Bay. Edmondson and Ingram (1939) suggest that the
decrease in settling by barnacles from January through March may be
primarily due to the heavy seasonal rainfall and stream runoff during
this period that causes an increase in turbidity and a decrease in
salinity of the surface waters. Their fouling panels, by being
placed in the surface waters in the southeast sector of the bay, were
much more susceptible to being influenced by these conditions than
were the present study panels.
Although seasonal trends unquestionably occur in tropical and
subtropi ca 1 communities, they are genera 11y not regarded wi th as much
consequence as those occurring in higher latitudes. Seasonality must
be demonstrated by the existence of annual repetition. The interpre
tation of seasonal colonization trends in this study was made difficult
by the fact that only one complete annual cycle was followed. In
studies where more than one year of colonization have been investi
gated, Sutherland and Karlson (1973) and Sutherland (1974) have
reported significant year to year variations in panel recruitment for
sessile marine invertebrates. Osman (1977) noted that the species
composition within a sessile invertebrate community did not repeat
itself between successive years. He suggested that there may be
replacement of species on a one-to-one basis. In light of these
examples, it would appear to be even less practical to extrapolate
seasonal trends based on the limited amount of data available in this

78
study. Peak periods of colonization, however, did appear for most of
the invertebrate groups under consideration. Because the data was
collected for the initial colonization phase of the community develop
ment, these peaks were generally a reflection of high levels of
invertebrate reproduction and larval availability.
Long (1974) briefly discussed seasonality in data from an
Hawaiian fouling panel study and noted that no seasonal trends were
observed because most species appeared in an inconsistent manner
throughout the year. Direct comparisons with his data are not pos
sible because his fouling panels were vertically oriented and were
either affected by waters from Pearl Harbor, a large estuarine port,
or exposed to open ocean water conditions at depths between 15 and
30 m. In either case, exposure conditions were significantly different
from those of the panels in this study.
It has already been noted that relatively small geographical
differences could account for apparent differences in peak periods of
colonization. The fluctuations in numbers of barnacles on the patch
reefs were most likely in response to varying environmental conditions
such as salinity, water temperature, and day length. The peak period
for oysters occurred about one to three months prior to the peak for
barnacles and could have also been contingent on these same factors.
Because most of these environmental parameters exhibit some seasonal
cycling even in tropical and subtropical regions, responses of
invertebrates to these parameters may be expected to repeat annuaily.
Didemnid tunicates and the bryozoans responded in a manner similar to

79
the oysters and barnacles and disp1ayed single peak periods of coloni
zation during the year. The multiple peaks or aperiodic appearances
of the other invertebrate groups suggest that their reproduction and
subsequent larval availability were in response to other, non-seasonal,
factors.
The colonization patterns for upper versus lower panel surfaces
were much more evident than seasonal trends and were better supported
by previous investigations. The tendency for many sessile marine
invertebrates to colonize shaded surfaces is a well known and well
documented phenomenon (Crisp, 1974). Birkeland (1977) speculates that
the causal mechanisms for these sciaphilous tendencies is in the
greater protection afforded by shaded environments. Jokiel (1980)
cites the harmful effects of ultraviolet radiation as a major contrib
uting factor in the distribution of shallow coral reef epifauna.
The distribution of barnacles on the study panels and on coral
reefs in general can primarily be explained by the tendencies to avoid
lighted surfaces. Visscher and Luce (1928) demonstrated that barnacle
cyprids show negative phototropic behavior at the time of attachment.
Other researchers have since shown that the relative amount of illumi
nation of the substratum ;s one of the more important factors
influencing colonization by barnacles (Gregg, 1945; Pomerat and Reiner,
1942; Weiss, 1947; Smith, 1948; Forbes, et ~., 1971).
Larval oysters are indifferent toward light just prior to attach
ment, but they generally aggregate and mature on shaded surfaces
(Crisp, 1974). In the present study, however, there was no significant

80
tendency for these molluscs to colonize on one side of the panels more
often than on the other. This was possibly a particular character
istic of the species observed (Ostrea sandv;chensis), or it could have
resulted from local environmental conditions such as turbidity or
amount of light reflected from the reef surface. It is likely that
the settl i ng behavi or of Q. sandvi chens is was the resul t of i nfl uence
from factors other than the protective mechanisms discussed by
Birkeland (1977).
Differences in the numbers of colonizers per unit area between
the two reefs were noted for the barnacles, serpul i d wonns, and
bryozoans. In all cases except barnacles, more individuals were
counted per unit area on the experimental reef than on the control
reef. These differences can best be attributed to the specific
locations of the two study reefs within Kaneohe Bay. The current
patterns around the two reefs are not identical and plankton samples
taken in the mid-bay area have indicated the abundance of merop1ankters
can vary widely between the two locations (Kimmerer, pers. comm.). By
being somewhat closer and downstream from the large central bay reef
flat, the experimental reef was better situated to receive more of
the invertebrate larvae with short-lived planktonic stages than the
control reef. The larval stages of the spirobids, tunicates, and
bryozoans all settle out of the plankton very shortly after being
released (Thorsen, 1961). Barnacle cyprids, however, can remain in
the plankton for relatively long periods of time and are capable of
traveling long distances (Sche1tema, 1971). This longer residence

81
time in the water column tends to disperse the larvae more and
minimize the effects of distance on colonization success. Primarily
because of this phenomenon, colonization by barnacles was less
affected by the specific locations of the study reefs in the bay than
was the colonization by other invertebrate groups.
The increase in numbers of spirobids on the study panels was an
alternate example of colonization being affected by planktonic life
span. As individuals colonized and matured on the concrete blocks of
the panel sets, their propagules became more and more abundant on the
nearby panels. The short-lived spirobid larvae were able to take
advantage of the new habitat space made available every week when the
panels were changed. This was supported by th~ fact that many of the
newly settled sp;robids were observed on the panels in closely grouped
patches, apparently after being released by a mature adult in the
surrounding community.
Differences between sites amounted to the greatest colonization
occurring on the windward sites and particularly on the windward
crests of the two study reefs. The long term data revealed that not
only greater abundances but a greater variety of organisms occurred
at the windward locations. The causes for this distribution most
likely involve an interplay of various physical and biological
factors. The fact that the windward sites were the first areas
encountered by meroplankters traveling onto the reef may have trig
gered settling responses in many of the invertebrate larvae.
The higher numbers of invertebrate colonizers on the windward
sites coincided with greater algal growth. Water motion has been

82
shown to be a major factor in the productivity on coral reefs (Doty,
1971) and could account for the differences in algal growth as well as
invertebrate colonization patterns on the study panels. Basically due
to the physical characteristics of greater water motion and near
optimal light conditions, the windward crests of both patch reefs were
the most favorable sites for algal production. The growth of algae
was further aided at the windward crest sites by the fact damsel fishes
(Stegastes fasciolatus) established territories in these areas on both
reefs and effectively limited the grazing by other fishes. The site
selection by these damsel fish was, in turn, probably a response to
the favorable physical environment, habitat and food availability, and
optimal algal production.
The increased amount of algal growth on the windward crests,
however, was not necessarily the precursor for successful recruitment
of sessile invertebrates. Vine (1974) has shown that the development
of a thick algal mat tends to inhibit rather than enhance the settle
ment and growth of many sessile invertebrate species.
That the most abundant invertebrate colonization coincided with
increased algal growth was not a contradiction of this phenomenon, but
was more a result of the experimental design and the particular
influence of Stegastes fasciolatus. By constantly being replaced at
three week intervals, the panels were not exposed for long enough
periods of time to develop thick algal mats. Furthermore, large
schools of herbivores (primarily juvenile parrotfishes) were
occasionally capable of overdhelm;ng individua1 S. fasciolatus and
invading the territories to feed on the algae. Stegastes fasciolatus

83
themselves continually grazed on the algae present within their
territories. Algal development was thus limited at the windward
crests while predation pressure on the invertebrate communities at
these sites remained at moderate levels. Grazing pressure and sub
sequent predation was higher at the other reef sites which, in
combination with a less favorable physical environment, resulted in
less algal growth and fewer invertebrate colonizers on those panels.
This result is in agreement with the hypothesis that increasing levels
of predation first cause an increase in benthic community diversity
before causing a decrease (Paine and Vadas, 1969; Menge and
Sutherland, 1976; Day, 1977; and Brock, 1978). The influence of
S. fasciolatus could also have manifested itself in the differences
in invertebrate colonization between the two study reefs. Because
juvenile parrotfish and other grazers were more numerous on the
larger control reef, there was a greater likelihood of them invading
the damsel fish territories. Grazing and predation levels were thus
more intense along the windward crests of the control reef which would
have led to a different response from the invertebrate colonizers.
The removal of fishes from the experimental reef resulted in a
very temporary change in grazing patterns and predation pressure on
the study panels. The rapid return of the dominant herbivores to the
experimental reef coincided with the return of Stegastes fasciolatus
which quickly re-established territories on the windward reef crests.
Because the panels were of three week duration and the return of
important components of the fish community occurred in less than one
month, the invertebrate colonization did not reflect this perturbation.

84
In conclusion, the invertebrate colonization trends relating to
panel exposure (upper versus lower surface areas) and of windward to
leeward locations were strong, consistent, and predictable. Slight
local environmental differences led to variations in colonization
between reefs apparently as the result of differences in life span of
certain meroplankters. Windward reef crests were the best developed
areas on the reefs primarily because of favorable physical character
istics and the maintenance of benthic communities through consistent
recruitment. Qualitative data from this study suggest that
herbivorous fishes are important in determining sessile invertebrate
colonization patterns and are heavily influenced by the presence of
aggressive, territorial fish species. Because of the establishment of
territories on windward reef crests, such fishes appear to promote
and help to perpetuate the community structure and resulting habitats
on coral patch reefs. The development of larger reef structures such
as coral atolls and islands may be due to many of the same factors
exemplified in this study.

REFERENCES CITED
Allen, F.E. and E. J. Ferguson Wood. (1950) Investigations on underwater fouling, Aust. J. Mar. Freshw. Res., 1(1):92-105.
85
Bakus, G.J. (1964) The effects of fishgrazing on invertebrate evolution in shallow tropical waters, Allan Hancock Foundation Pub. Occ. Pap., No. 27,27 pp.
Bakus, G.J. (1966) on coral reefs,
Some relationships of fishes to benthic organisms Nature, 210:280-284.
Bakus, G.J. (1967) The feeding habits of fishes and primary production at Eniwetok Marshall Islands, Micronesica 3:135-149.
Bakus, G.J. (1969) Effects of the feeding habits of coral reef fishes on the benthic biota, Proc. Symp. Corals and Coral Reefs, Mar. Bio1. Assoc. India, 445-448.
Banner, A.H. (1974) Kaneohe Bay, Hawaii: urban pollution and a coral reef ecosystem, Proc. 2nd Internt1. Symp. Coral Reefs, Brisbane, 2:685-702.
Barr, A.J., J. H. Goodnight, J.P. Sall and J. T. Helwig (1976) A user's guide to SAS-76, SAS lnst. Inc., Raleigh, North Carolina, 329 pp.
Barnes, H. (1955) Further observations on rugophilic behavior in Balanus balanoides (L.) Urdensk. Medd. Dansk. Nanek., 117:341-348.
Barnes, H. (1956) Surface roughness and the settlement of Balanus ba 1 anoi des, (L.) Arch. Soc. "Vanamo ll , 10: 164-168.
Bardach, J.E. f; shes,
(1958) On the movements of certain Bermuda reef Ecology, 39:139-146.
Bardach, J.E. (1959) The summer standing crop of fish on a shallow Bermuda reef, Limno1. and Oceanogr., 4:77-85.
Birkeland, C., A.A. Reiner and J. R. Young (1976) Survey of marine communities in Panama and experiments with oil, U.S. Environ. Protect. Agency, Ecol. Res. Series #600-3-76-028, Washington, D.C.

86
Birkeland, C. (1977) The importance of rate of biomass accumulation in early successional stages of benthic communities to the survival of coral recruits, In:Third Internat. Coral Reef Symp., Miami Vol. I, pp. 15-21.
Boyd, M. (1972) Fouling community structure and development in Bodega Harbor, Calif., Ph.D. dissertation, Un;v. Calif., Davis.
Brock, R.E. (1978) The effects of grazing by parrotfishes (Family Scaridae) on selected shallow Hawaiian marine communities, Ph.D.' dissertation, College of Fisheries, Univ. Washington., 190 pp.
Brock, R.E., C. R. Lewis and R. C. Wass (1979) Stability and structure of a fish community on a coral patch Hawaii, Mar. Biol. 54:281-292.
Brock, V.E. (1954) A preliminary r~port on a method of estimating reef fish populations, J. Wildlife Management, 18:297-308.
Chapman, C.J., A.D.F. Johnstone, J.R. Dunn and D.J. Creasy (1974) Reactions of fish to sound generated by diver's open-circuit underwater breathing apparatus, Mar. Biol., 27:357-366.
Chave, E.H. and D.B. Eckert (1974) Ecological aspects of the distribution of fishes at Fanning Island, Pac. Science, 28 ( 3) : 296-3 17.
Chave, K.E. (1973) Setting and surroundings of Kaneohe Bay, In:S.V. Smith, K.E. Chave and D.T.O. Kam (eds), Atlas of Kaneohe Bay: a reef ecosystem under stress, Honolulu, Univ. Haw. Sea Grant Pub. TR-72-01, pp. 1-8.
Clark, E., A. Ben-Tuvia and H. Steinitz (1968) Observations on a coastal fish community, Dah1ak A,rchipelago, Red Sea, Bull. Sea Fish. Res. Sta. Haifa, No. 49, pp. 15-31.
Coe, W.R. (1932) Season of attachment and rate of growth of sedentarv marine organisms, Scripps Inst. Oceanogr. Bull., Tech. Series, ~ 3:37-86.
Cae, W.R. and W .... E. Allen (1937) Gro\</th of sedentary marine organisms on experimental blocks and plates for nine successive years, Scripps Inst. Oceanogr. Bull., Tech. Series, 4:101-136.
Collette, B.B. and F.H. Talbot (1972) Activity patterns of coral reef fishes with emphasis on nocturnal-diurnal changeover, In: B.B. Collette and S.A. Earle (eds), Results of the Tektite Program: ecology of coral reef fishes, Bull. Nat. Hist. Mus. Los Angeles, 14:1-9.

Connell, J.H. (1978) Diversity in tropical rain forests and coral reefs, Science, 199:1302-1310.
Cowles, R.P. (1930) A biological study of the offshore waters of Chesapeake Bay, U.S. Bur. Fish. Bull., 46:277-381.
87
Crisp, D.F. (1965) Surface chemistry, a factor in the settlement of marine invertebrate larvae, Proc. 5th Mar. Biol. Symp. Botanica Gothoburgensia, 3:51-65.
Crisp, D.J. (1967) Chemical factors inducing settlement in Crassostrea virginica (Gmelin), J. Anim. Ecol., 36:329-335.
Crisp, D.J. (1974) Factors influencing the settlement of marine invertebrate larvae, In: Chemoreception in Marine Organisma, Grant, P.T. and A.M. Mackie (eds.l, Academia Press, London pp. 177-265.
Dana, J.D. (1875) Corals and coral islands, Sampson, Low, Marston, Low and Searle, London, xx + 348 pp.
Darwin, C. (1842) On the structure and distribution of coral reefs, Lon don t 8 vo 1 s .
Dart, J.K.G. (1972) Echinoids, algal lawn and coral reco1onisation, Nature, 239:50-51.
Dawson, Y.E. (1966) Marine botany: an introduction, Holt, Rinehart and Wilson, Inc., New York, 371 pp.
Day, J.H. (1963) The complexity of the biotic environment, In: Harding, J.P. and N. Tebble (eds), SpeCiation in the Sea, System. Assoc., London, 3i~49.
Day, R.W. (1977) Two contrasting effects of predation on species richness in coral reef habitats, Mar. Biol., 44:1-5.
Devaney, D.M., M. Kelly, P.J. Lee and L.S. Motteler (1976) Kaneohe: a history of change (1778-1950), B. P. Bishop Museum, Honolulu, 271 pp.
Dew, B. and E.J.F. Wood (1955) Observations on periodicity in marine invertebrates, Austr. J. Mar. Freshw. Res., 6(3):469-478.
Dixon, W.J. and M.B. Brown (1977) Biomedical computer programs Pseries, Univ. Calif. Press, Berkeley, Calif., 880 pp.
Doty, M.S. (1971) Measurement of water movement in reference to benthic algal growth, Botanica Marina, 14:32-35.

88
Edmondson, C.H. (1937) Studies of fouling organisms in Kaneohe Bay, B.P. Bishop Mus. Occ. Pap., 14:251-300.
Edmondson, C.H. (1946) Reef and shore fauna of Hawaii, B.P. Bishop Mus. Spec. Pub., No. 22, ii + 381 pp.
Forbes, l., M.J.B. Seward and D.J. Crisp (1971) Orientation to light and the shading response in barnacles, In: Fourth Europ. Mar. Bio1. Sym., D.J. Crisp (ed.), Cambridge Univ. Press, Cambridge, pp. 539-58.
Gosline, W.A. and V.E. Brock (1960) Handbook of Hawaiian Fishes, University of Hawaii Press, Honolulu, Hawaii 372 pp.
Gregg, J.H. (1945) Background illumination as a factor in the attachment of barnacle cyprids, Biol. Bull., 88:44-49.
Grove, B.H. (1933) Rate of growth, age at sexual maturity, and duration of life of certain marine organisms at Woods Hole, Massachusetts, Biol. Bull., 65:375-386.
Grovhoug, J.G. and R.S. Henderson (1976) Distribution of inshore fishes at Canton Atoll, In: S.V. Smith and R.S. Henderson (eds), An environmental survey of Canton Atoll lagoon, 1973, San Diego, Naval Undersea Center, pp. 99-157.
Grovhoug, J.G. (1978) Pit; power plant intake survey: November 1977, San Diego, Naval Ocean Systems Center, TR 288, 58 pp.
Gundermann, N. and D. Popper (1975) Some aspects of recolonization of coral rocks in Ei1at (Gulf of Agaba) by fish populations after poisoning, Mar. Bio1., 33:109-117.
Hobson, E.S. (1974) Feeding r~lationships of teleostean fishes on coral reefs in Kana, Hawaii, Fish. BulL, 72:915-1031.
Johnson, M.W. and R.C. Miller (1935) The seasonal settlement of shipworms, barnacles, and other wharf-pile organisms at Friday Harbor, Washington, Univ. Wash. Pub. Oceanogr., 2:1-18.
Jokiel, P.L. (1980) Solar ultraviolet radiation and coral reef epifauna, Science, 207:1069-1071.
Jones, R.B. and J.A. Chase (1975) Community structure and distribution of fishes in an enclosed high island iagoon in Guam, Micronesica, ll(1):127-i48.
Jones, R.S. and ~.J. Thompson (1978) Comparison of Florida reef fish assemblages using a rapid visual technique, Bull. Mar. Sci., 28:159-172.

Kaufman, L. (1977) The three spot damsel fish: effects on benthic biota of Caribbean coral reefs, Proc. Third Interntl. Coral Reef Symp., Miami, 1 :559-
Key, G.S. (1973) Reef fishes in the bay, In: S.V. Smith, K.E. Chave and D.T.O. Kam (eds), Atlas of Kaneohe Bay: a reef ecosystem under stress, Honolulu, Univ. Haw. Sea Grant Pub. TR-72-01, pp. 1-8.
89
Kahn, A.J. (1971) Diversity, utilization of resources, and adaptive. radiation in shallow-water marine invertebrates of tropical oceanic islands, Limnol. Oceanogr., 16(2):332-348.
Levene, H. (1960) Robust tests for equality of variances, In: Contributions to Probability and Statistics, I. Olkin (ed.), Stanford Univ. Press, Stanford, Calif., pp. 278-292.
Long, E.R. (1972) Studies of marine fouling and boring off Kodiak Island, Alaska, Mar. Biol., 14:52-57.
Long, E.R. (1974) Marine fouling studies off Oahu, Hawaii, Veliger, 17:23-36.
MacArthur, R.H. and E.O. Wilson (1967) The theory of island biogeography, Princeton University Press, Princeton, New Jersey 203 pp.
McDougall, K. D. (1943) Sess i1 e marine invertebrates of Beaufort, North Carolina: a study of settlement, growth, and seasonal fluctuations among pile-dwelling organisms, Ecol. ~lonogr., 13:321-374.
Menge, B.A. and J.P. Sutherland (1976) Species diversity gradients: synthesis of the roles of predation, competition, and tempora1 heterogeneity, Amer. Nat. 110(973):351-369.
Mook, D. (1976) Studies on fouling invertebrates in the Indian River, 1. Seasonality of settlement, Bull. Mar. Sci., 26(4):610-615.
Muus, B.J. (1968) A field method for measuring "exposure" by means of plaster preliminary account, Sarsia, 34:61-68.
Nagelkergen, W.P. (1974) On the occurrence of fishes in relation to corals in Curacao, Stud. Fauna Curacao, 147:118-141.
Nolan, R.S., R.R. McConnaughey and C.R. Stearns (1975) Fishes inhabiting two small nuclear test craters at Eneweta~ Atoll, Marshall Islands, Micronesica, 11:205-217.

90
Odum, H.T. and E.P. Odum (1955) Trophic structure and productivity of a windward coral reef community on Eniwetok Atoll, Ecol. Monogr., 25:291-320.
Ogden, J.e. and P.S. lobel (1978) The role of herbivorous fishes and urchins in coral reef communities, Env. Biol. Fish., 3(1):49-63.
Osman, R.W. (1977) The establishment and development of a marine epifaunal community, Ecol. Monogr., 47:37-63.
Paine, R.T. and R.L. Vadas (1969) The effects of grazing by sea urchins, Strongylocentrotus spp. on benthic algal population, limnol. Oceanogr., 14:710-719.
Pomerat, C.M. and C.M. Weiss (1946) The influence of texture and composition of surface on the attachment of sedentary marine organisms, Biol. Bull., 91:57-65.
Rafail, S.Z. (1973) A simple and precise method for fitting a Von Bertalonffy growth curve, Marine Biology, 19:354-358.
Randall, J.E. (1963) An analysis of the fish populations of artificial and naturai reefs in the Virgin Islands, Caribb. J. Sci., 3:31-47.
Randall, J. (1965) Grazing effect on sea-grasses by herbivorous fishes in the West Indies, Ecology, 46:255-260.
Rastetter, E.B. and W.J. Cooke (1979) Responses of marine fouling communities to sewage abatement in Kaneohe Bay, Oahu, Hawaii, Marine 8iology, 53(3):271-280.
Risk, M.J. (1972) Fish diversity on a coral reef in the Virgin Islands, Atoll Res. Bull., 153:1-6.
Russell, B.C., F.H. Talbot and S. Oomm (1974) Patterns of colonisation of artificial reefs by coral reef fishes, In: Proc. 2nd Int. Coral Reef Symp., Brisbane, Great Barrier Reef Committee, 1:207-215.
Russell, B.C., G.R.V. Anderson and F.H. Talbot (1977) Seasonality and recruitment of coral reef fishes, Aust. J. Mar. Freshw. Res., 28:521-528.
Schoener, A., E.R. Long and J.R. DePalma (1978) Geographic variation in artificial island colonization curves, Ecology, 59(2):367-382.
Scheltema, R.S. (1971) The dispersal of the larvae of shoal-water benthic invertebrate species over long distances by ocean currents In: Crisp, D.J. (ed), Fourth Europ. Symp. Mar. Biol., Cambridge, pp. 7-28.

91
Smith. C.L. and J.C. Tyler (1972) Space resource sharing in a coral reef fish community, In: B.B. Collette and S.A. Earle (eds), Results of the Tektite Program: ecology of coral reef fishes, Bull. Nat. Hist. Mus. Los Angeles, 14:125-170.
Smith, C.L. (1973) Small rotenone stations: a tool for studying coral reef fish communities, Amer. Mus. Novit., 2512:1-21.
Smith, F.G.W. (1948) Surface illumination and barnacle attachment, Biol. Bull., 94:33-39.
Smith, S.V., K.E. Chave and D.T.D. Kam (1973) Atlas of Kaneohe Bay: a reef ecosystem under stress, Univ. Haw. Sea Grant Pub. TR-72-01,128 pp.
Smith, S.V. (1977) Kaneohe Bay, a preliminary report on the responses of a coral estuary ecosystem to the relaxation of sewage stress, In: Third Internat. Coral Reef Symp. Miami Vol. II, pp. 577-584.
Stephenson, W. and R.B. Searles (1960) Experimental studies on the ecology of intertidal environments at Heron Island I. Exclusion of fish from beach rock, Austral. J. Mar. Freshw. Res., 11{2}:24l-267.
Strickland, J.D.H. and T.R. Parsons (1968) A practical handbook of seawater analysis, Bull. Fish. Res. Sd. Can., Vol. 167, 311 pp.
Sutherland, J.S. and R.H. Karlson (1973) Succession and seasonal progression in the fouling community at Beaufort, North Carolina, In: Third Interntl. Congress on Marine Corrosion and Fouling, Northwestern Univ. Press, Evanston, Illino;s, 906-929.
Sutherland, J.S. (1974) Multiple stable points in natural communities Amer. Nat., 108:859-873.
Thorson, G. (1961) Length of pelagic larval life in marine bottom invertebrates as re1ated to larval transport by ocean currents, In: Oceanography, Amer. Assoc. Adv. Sci. Publ. 67:455-474.
Vine, P.J. (1974) Effects of algal grazing and aggressive behavior of the fishes Pomacentrus lividus and Acanthurus soka1 on coral reef ecology, Mar. 8iol., 24(2):131-136.
Virnstein, R.W. (1977) The importance of predation by crabs and fishes on benthic infauna in Chesapeake Bay, Ecology, 58:1199-1217.
Visscher, J.P. and R.H. Luce (1928) Reactions of the cyprid larvae of barnacles to light with special reference to spectral colors, Bio1. Bull., 54:336-350.

92
Vrijmoed, L. (1975) An analysis of surface fouling organisms in the coastal waters of Hong Kong, In: B. Morton (ed), Special Symposium on Marine Sciences, Pacific Science Assoc., Hong Kong, 129-135.
Wass, R.C. (1967) Removal and repopulation of the fishes on an isolated patch coral reef in Kaneohe Bay, Oahu, Hawaii, M.S. thesis, Univ. Hawaii, 77 pp.
Weiss, C.M. (1947) The effect of illumination and state of tide on the attachment of barnacle cyprids, Biol. Bull., 93:240-249.
Wells, J.W. (1957) Coral reefs, In: National Research Council Treatise on Marine Ecology and Paleoecology, New York, Vol. 1, Ecology, ed. by J.W. Hedgepeth, (Geol. Soc. Amer., Mem. 67), pp. 609-631.
Wilson, E.O. (1969) The species equilibrium, In: Diversity and Stability in Ecological Systems, Brookhaven Symp. in Biol., 22:38-47.
Wilson, K.V. (1956) A distribution-free test of analysis of variance hypotheses, Psych. Bull. 53(1):96-101.
Winer, B.J. (1971) Statistical Principals in Experimental Design 2nd ed., New York, McGraw-Hill.
Wiseley, B. (1959) Factors influencing the settling of the principal marine fouling organisms in Sydney Harbour, Aust. J. Freshw. Mar. Res., 10:30-44.
Woods Hole Ocean. Inst. (1952) Marine fouling and its prevention, U.S. Naval Institute, Washington, D.C., 388 pp.
Yoshihara, T. and P.C. Ekern (1978) Assessment of the potential of solar energy in Hawaii, final report, Univ. Hawaii, Hawaii Natural Energy Institute, 49 pp. + .
Zar, J.H. (1974) Biostatistical Analysis, Prentice-Hall Inc .• Englewood Cliffs, New Jersey, 619 pp.

93
APPENDICES

FIGURE A-l. Primary current patterns in Kaneohe Bay
(from Bathen, 1978).
94
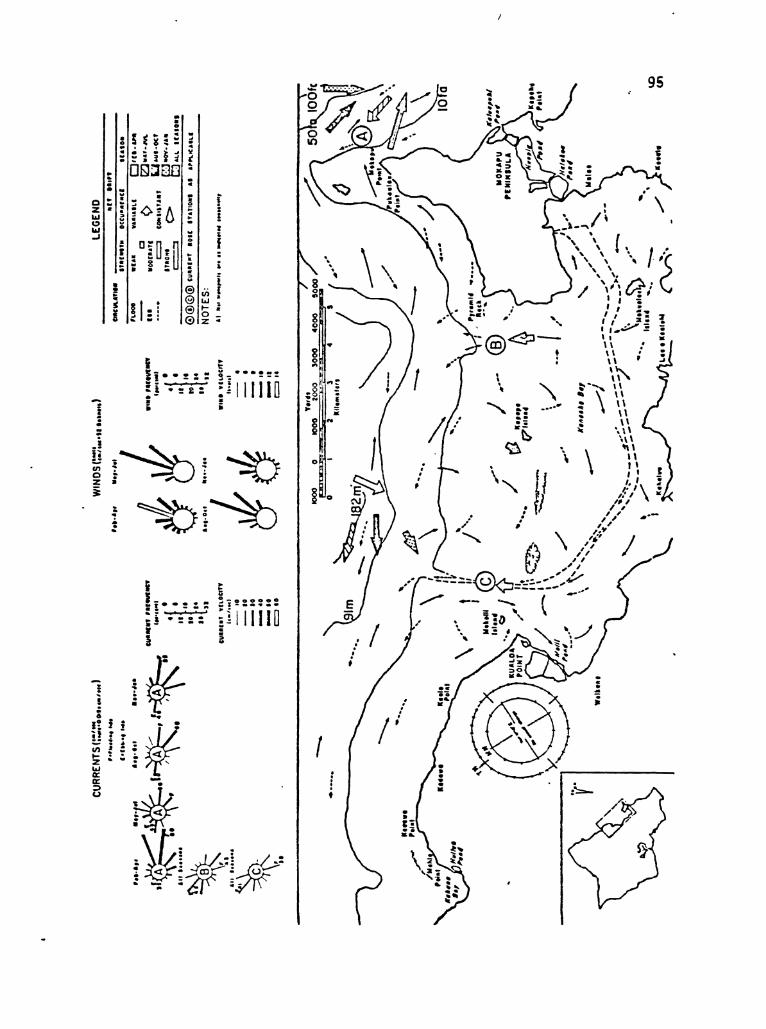
. ~
~ .. w 0 .
! ! §I-: .. _. ~ ...
i 1_ ••• :. = =! ; .. ... :, .. -.. ;! - - ... , !
~¥: . , •
- -I! i! #f .. -i !
'} r • hit: \ en .. ..... <:( ,... ! :..: .... Z « W a:: _
a:: •
a }*
f I ! 11
i :
.. S ••• !!!'!
~!IIIIIO .. . ii
f'i{)J;". : e-i:#.: ,tf. :",-.--
: ~.: u . <t _ . ; -: --=
g '" 51,
o
, : : t
95

Table A-I. Diffusion fac~ors derived from plaster-af-paris ·clod cards" (see Doty, 1971). The higher diffusion factors indicate a greater amount of water motion.
Initial Final Wt. Loss Diffusion Site Nt. (g) Wt. (g) per 24hr Factor -------------------------------------------------------------Exp-l 42.3 37.0 5.3 2.5
4l.2 36.3 4.9 2.3
Exp-2 43.2 33.9 9.3 4.4 42.1 32.9 9.2 4.4
Exp-3 43.1 29.8 1·3.3 6.3 40.6 27.9 12.7 6.0
Exp-4 42.2 33.6 8.6 4.1 39.8 31. 7 8.1 3.9
Exp-S 42.7 38.0 4.7 2.2 42.5 37.9 4.6 2.2
Con-l 40.8 35.5 5.3 2.5 41.2 36.1 5.1 2.4
Con-2 42.1 34.4 7.7 3.7 43.6 35.9 7.7 3.7
Con-3 43.1 28.2 14.9 7.1 39.6 25.2 14.4 6.9
Con-4 40.9 33.5 7.4 3.5 43.0 35.8 7.2 3.4
Con-5 42.4 37.9 4.5 2.2 40.9 36.5 4.4 2.2
96

FIGURE A-2. Ambient light conditions in Kaneohe Bay
(Smith, unpubl.).
97

o :::z a: ...J en ...... .=> z o u o u
I:I:: c!) ...... ...J
I:z w o ...... u :z -
o o (,0
Z!J!:¥l4iQ> ;:...,. -- -.-'-.-~.-
o o 1.[')
______ _i'
~~},.;.,::::~
o o (T')
o o N
JJ:JO tr3d SJ.3l~NtJl
98
z (J)
J
:E: en
:::::::: c-01 ---, ..-.
:
~ :z: (.()
~ ---, ::::::::
~ CO
:::::::: ["-
-l ~ 01
j ..-. . ::z
~ (f')
J
Z c-
:::2:: C-O)
J -=I
:z: (f)
J
:::::::: (D
:::::::: C-O;
J -0

FIGURE A-3. Wind speed and direction at Coconut Island,
Kan~ohe Bay (Smith, unpubl.).
99

-(,) CD
~ E -
0 w w a.. CI)
-CD
E -..... o
~ o -z o lt.)
W 0:: -o
100
8
6
4 ;A
JFMAMJJASONDJFMAMJJASONDJFMAMJJASOND
1976 1977 1978

TABLE A-2. Checklist of organisms present on panel sets after 15 month exposure.
EXPERIJ.'tE,:'lAJ> REEF SITES CONTROL REEF SITES
1 2 3 4 5 1 2 3 4 5
-----------------------------------------------------------------------------------------------------------CIIWROPIlYTA
Enteromorpha ap. X X X
ClillYSOPUYTA tlavicula sp. X X X X X
RHODOPIlY'fA PoroU thon onkodes X X X X X X X X X X
PORIFERA Unid. sponges X X X X X X X
CI1IDAHIA Hydrozoa
Ilnid. hydrozoans (Pennaria?) X X J\nlho~oa
Actinurin !u,tll~ i04 l'ulcltelJ a • X X X X R: .. lirmtll\\fi coolwi X
l-ladrepor:1.rb !~cillo~~ d:1.~icorni6 X X X X X X X X X l<ontipcra ve~~ X Pori b~,; ~l'..~ X Tubilutrea aurea X
PI.ATYlIWIItI'l'HES Unid. flatworOlI1 X X X X X X X X X ....
0 -

TP.BLE A-2. (Continued) ---
EXPERIMENTAL REEF SITES CONTROL REEF SITES
1 2 3 4 5 1 2 3 4 5 ------------------------------------------------------------------------------------------------------------ANNELIDA
Polychaeta Errantia
Nereidae spp. X X X X X X X X X Sedental'ia
Subellidae spp. X X Sp.irobinao BPP. X X X X X X X X X X Hydroidcs spp. X X X X X X X X X X
ARTImOPODA Cru.atacea
Bilh~ spp. X X X X X X X X X X n;11.:mU3 amphi tri te X X X X X X X X X x n:~] i1IlUS eblirneua X X X X X X X x Di1J3nus rcticulatua X X
Isopod-n---Isopoda app. X X X X X X X X X X
ArnphipodOl Amphipoda spp. X X X X X X X X X X Caprell idae app. X X X X X X X
Decapoda Alpltcua sp. X Porlunidae sp. X Xanthidae ap. X X
HOLLUSCA Gastropoda
Unid. snails X X X X X X X X X X
-o N

TABLE A-2. (Continued)
EXPERIHEtITAL REEF SITES CONTROl. REEF SITES
1 2 3 4 5 1 2 3 4 5
-----------------------------------------------------------------------------------------------------------Vermetidae
VcrmctUlJ alii X X X X X X X X X X hltJ ocanchus Iteer.ae X X X X' X X X X X X
Calyptn.eillne CrcniduLI aculenta X X X X X X X X CruciiJ\;lum cpinosum X X
Nudibri,nchi a Unid. nudibranchs X X X X X X
Pelecypodu (Biv!llvb) Or;trea condvicenois X X X ·X X X X X X X Anomia -noiJilis X ----
IDl'OPROCTA (Bryozoa) Unid. bryozoans X X X X X X X X X X /I.tc:J. trllflcClta X X X X X X X X X B\i701]_~ [;!;;:- X X X X X X X X X X Swi;':nydla Infant! X X X X X X X lIo]opordla spp. X X X X X X X X X X llict'oporell~ 6p. X X X X X X X X X X Schizoporella sp. X X X X X X X X X X
a:HINODERMA'l'i\ Echinoidea
!!ipncustes ~_atil1a (1 juv.) X X
-<:)
w

TABLE A-2. (Continued)
EXPERIl1ENTAL REEF SITES
12345
CONTROL REEF SITES
'I 2 3 4 5
------------------------------------------------------------------------------------------------------------CHORDATA
Phnllusidae Accidia spp. Aacidia intcrrupta
Didcmnlche IJidernnurn ap.
Polyclinidlle 1'01 yel inurn cOl'lBtellatum
Do tr y :tiTd';e---Botryloides ap.
x X
X
x X
X
X
x X
X
x
X
X
X
x X
X
x X
X
y.
x
x
x X
X
X
x
x
-----------------------------------------------------------------------------------------------------------
PERCENT OCCURRENCE OF TOTAL TAXA (47)
68.1 68.1 53.2 55.3 51.1 74.5 83.0 55.3 66.0 61.7
..... o ~

• "I:
<.I
0 0 -s.. cu Co
ea cu S-ea S-O
en s-cu
.Q E ::3 :z:
• en !t-
CU (I) S-
~ "C ::3
+.> en 0 3
+.> (I)
..r::. +.>
S-O !t-
ea +.> m
"C
+.> s::: (I) E (I) -+.> +.> (I) en
-CU s::: m
Q.
· ('t') I ex:
LLJ -I a:l ex: I-
-% ~
:l
5 :II c
c
Z ;;: .., " 0 u
u
~
% OJ ·A .J 1: :to
:to
~
? "J JI
" cr 12.
~
""' z
, z
III ... ., z
!)-O .. t"Z~7
... :>z-..., ..... .JI
1lc:>_'~z .. 1.!J
_V1.-:2n.::f'J
105
~~~~OOO~~4~N4~~~~~~N~_N4~ __ ~O~~NG~_~OOOOOOOO .- -- -
~"~~O~'~~~~~~OO~~OO?~O~OO~?ft??OO~~O~?~?OOO~O -I/\!-'" ~~~o~on~04_~=~~_.~~~~N~.~·~~NC~~~O~-O?O~~~OG
~~~_~~N~~~# N~-
~OR~._~~~~~~~-N_'C,'-.~-~.~~~~~~~~O~O~~O~~ftO ~~ __ ~~N~nn_~~:~~~~c. __ V~N-_~NN_
~OO~.~~~_4~ON~N~_O __ N~O~_n_"~_NON~o~oo~onoo~ ,., - ... -
~a~~~.o~~"-~-~~-r~~O.'"-NQ~~N~e~~N~~~O=~N •• _ ~~~~_~~~~~~ft~~.'~.~.NN ~DN.. _NM~n_ --~._ ~-~n~~~~~~~'~4--~~~ -~ ~~~ ,
~-~~~~.'~~~~~_~~~-.o __ ._._.ft~~_~"~.?~?o~OOO ~.··~G"~'~.GN~.~-_~.a=.~_~~_=.>_ON_
-V-'~~.~~~JI~~--- - ______ ~'~N.'~.M_ ~~.~~~40.~·JI~~,.~-_~~~~~--~.~~N~:#~_~ono __ ~o ~N-- -~_.~~~N ~.- N
c':tc.%-tlO~ O.o"' .... o"oo,..n"'o::tt")oo"oe"'~O~OOOI")O'5.,(JO'!tl"3_ ... _N'O_C"~ .. N "IN
"., - ~ . .:J
--~~-~~~O.~O~ .. ~OG~~~~~-~.~,-~~O,~O~O~OOQOOO ~~ N~-_ ~ N __
-----------------------------------------~--G~ca~~~~ ~~~a~~~~~~~~~~~1~~3~~~Q~~~~~~~~~~~~~ .)(J('I( .. )I)C'x~,,'>(v'liJ()I()(wJ("<l(~ .... )(lII,,"C ... 1(.If.."':rt>(j(<<I(J(l()()("'1t()(
W \&I ..., at 11.1 tu W 1.1 y,; W !.L' '.al lH ill ... IU .oJ ~I.! U.I flY .L' L.J 'JJ r:I U 'J • .J 11' ..:J 1.J IU 1..J -.: l!.I ;';"1 161 JJ '" W I.IJ I!.J 1.&1 I" !ll
~,·····~······.····.~~~ora~~~=~~~:~~ ....... ~ ~ •.•••••.......•....... ~ .•••...•.....•••• ~ .. ~~~~~~~S~~~::~~~~~~~S~~~3~~~~l~;::;§~~~~~~~g ~~~~,.<.~~~===%~~~==~~~~~~~~ .•• ~~>"~",~,,. ~.~M.~ •• • •• ~~~~~···-"-~-=W-~~~~_"· ••• rN~~~~~~ -NO-Nn-Nft-N~-No-~nNN __ OONn_~e_~O-~~-_NN'_NNO

--0 OJ :::l r:: -+-l s::: 0 u -. (Y')
I < UJ -J a:l < ~
c-ouzz::tt
... ~ Z -.1 C ... l!.I
.. z .. ~ z ~ .. • ~
; 0 ~ ~
" ... • ... .. Z 'lol ... ., .. a.
2--~'~a.oo
'i ,~
:It ... ao: c·o..:uc"".., Q. .. :I Z
~ z -,~ .. ~ z l1~OZ"'"
Q. C Z hl..l
106
.~~~~~d.~4~~,~~~~,~~ft~~~~.CD-~~-~.~~?"~.-~~. ~ 0 _. ~~~~- ~~~ -- - n-_N~ __ -
.ft~~ .• ~~~~e~-~~~-.-~.~_~_o __ ~_n._~~~.~"'.GO~~ ~._~~.~_~~ •• ~~~NM~_.~'" ~__ _N_~~~~
N~~~~~~_·~:~~ -N_ ~4
--------------------------------------------e. 1\ " 0.. ., '\ ~ I". ~ ~ a. ." ~ ~ ~ Q. e. ~ =- 0 :::. e ~ t:.. Q. "_ 0. t! ':.. 0. ~ ~ ~ 1"_ ~ : l'" Cl 0. 0: c.. e. a. I'\. • ~ ~ ~ v ~ • ~ x ~ ~ ~ ~ ~ < ~ ~ ~ X K • ~ X K X ~ ~ K X X ~ X X • X K ~ K K • X ~ • K U w W W !i.I ~ fU UJ UJ W ...... tlJ IoU d ..J ~ u.. UJ I.lJ W II! _ LJ W :.;J lI,.. III W l,1 I,' \"II W .... \:. lU U IU \OJ ~I W ... I.IJ 'l.I tIJ
~,~,~~~,~~"'~,,'~,~,~~~~~~~~~~~~~~G~~~~~~eO ,~~,',~,~",'~,,'~,~,~"~~~~~~~,,~"',~,~~~,
~ ~ ~ ~ ~ ~ ~ ~ ~ ~ - - > » > > > U v U y ~ = Z = = ~ = = 1 ~ : = = ~ ~ ~ ~ > ~ ~ ~ ~ Z :t ;l ':) :: I., .;J :U ... J ...; ........ ,--1 C :; :: 'J -= ..J ~: ;:1 _' ~ ... <t < ..J .... ..J .,J wi o.l , -.l .. "". ~ :.. Co ., -< <l ." <I( :J ••• <~~~~=~~~~z~z~'cc~·~~~~~.~7~Y-' ••• CZTXT.' ?~._~._~~~ft~N'<~~~.-n._p- __ '~~-ft~N~~N~G~~·._~ --N.~_.N?--Ne~-N~O-~N~ __ Ne~_Nao-N~e--N~--~~~

.. % .. 0 z ;:) III .. • <C
; ~
" " '" u
• u
· .. % : .. .. '" % S
.. .. z .... VI w a: Jl. .. 0 z
-"0 Z (I) ::l W C .. ..... 0 +-J :: C 0
(..) -•
(") I
< LAJ -oJ CO < .....
IoIOII.UQ.COVI
%"'oa:~-c.tU
-tnOt\O-:2
oc ... w
107
O~~~~N~~~~~N~_~~~~.O~_~~~N~.~~~_~~~N_O~OO~OO _~. d ___ N_ -- - -~-.
Q~~~O~~~~OO~O~4_0000~~~O~OO~"O?O~O~~~OOOOOOO .. -OOOOOOO.~~O_~~~_=_~O~=O~~"~"O"O~_N~OOOOOOOOO
~~"~.~._. N -~~-_
O~~O~·_O_M4~.~~4~~.~_~~4M4~~~~~~·.~·.OO~OOOO _N_ ~ ••• ~ ___ 'N~ ~-~~N~--- ~ ~~ -
oo~o~a~~~~4~~~~nooooo~~oo~~~~~O~OO~N~~~~e~~Q NN~~~ -
.4nN~·-~.n~~~~~~~~"-N~MN~~OD.·~ ••• ~N~'-~~~4--~~~~.'_Q~~.~~~--~.N_ ~N' .Q~~~~~~'~~~n N~-~-~~'O'NO._ __N AN ... ~~~
~Q-~~~4.N~~.~.-"~~-"-- •• ~==4~~O~~~~~·OOO~O~O _~N~'N~O_~_~>D ___ N~ M-N- ~ .... ~,~.=.=
-~-~-~-' N-
~~~'N~.n~~N~.O~ON~N~".~Don-~.~N-.N=Z'.OOOO-O .- -~~~ .. ~.~~- ~-.= ... --~-
~~~~~O~~O~~O~OQO~_~O~OOO~OO~O~O~QOONO?~O~OG~ ., ~
Q~"-M-"eN~~~-~-~~~N __ ~NNN~~NO_OOOOOOO~~OOO~ • _N~_~~_ ~ __ .~
~Qa~aa~=~a~~a~~~e~~~n~~~~a~~~~oe~~~~~~~a~~aa ~ •• ~KK~_W.~K •••• <~~x.~ •• <.KX~<.~X •• X~~K~.KWW ~W~W~~Wm~~~wwmmw~w~uwWWWWW~WWWWWWw~W~~WWW~WW
D~~~~~~~~~'~~'~~~~~"en~eC~~G~CC~~~~~~~~~~~~ ~~~~~'~'h~"~~~'~~""~"~~"'~~~~~'~~~~~~~~ :: : ;: ...I ...I .2 .:" ,. '': .... '!. c.. ~ ... - > .... > U \J U Z Z ;;: 't' ':l :' .;:. 0. :r ,J .:: .. )0 )0 :- =- z 2 -: .J .J .,.J ..J ~ : : ::0 ~ -:: :> :- :- ',J ~ I.. L ~ V ::' ~ ::: ....: 1.1.' '..J < ot t" .. ' ~ .~ ~ rl C !:!. C " ~ <C: ., :> .., :: ::: ::Ii :: ::l ::l ~~'~~~<4·~~~=-~::Z=O~~~~~_·" •• <Y7·~~~~"'~' .~~~~·~~.~.~~~O~D"·_~_~_r~_~~~~~n·.·.~Na~~O· __ ~~_NO_~~_.n_~n_nONN __ ~~No_No_Nn_N~ __ NNO_NN

-"C QJ :::J C -+0) C 0
U -•
('t') I
<: IoU -I CO < to-
.CI~ .. ':)Z-'"
.. :r ~ :I
~ 11 C
C
· 7! 0
" ~ \J
• oJ
· .. z 'U ... .. .. A
'1
Z ... .. ::,: c: =. .. 0 Z
t .. .. 0 z
!l.cz .....
108
,~OOO~" __ O~O_OOO~,O~~~~ __ .O"_~O~~~~~~~CA~~~_ -- ~~ .. ~~.~~~~~O~D~~~4-~~~~~~N~O~~~~~.na~~-~~~~~-m~ .... -# N \I') ",., ~ ,.. ;,."" ;,;~ _ # """ ;,. _ .. , .:; .::J .., ,"lit "0 .. ", '" - C'- (\.:,;. - - i1 • :'.I _ N .. , .... '" - 1) ." ... "" "') Tl :P ~ _ .. "ft N ----:'I,I:\I-'"'-'V_I'\I __
o~~o~~ooo~o~o~o~~oao~~o~~~'~~~~O~O~~Q~OO,~~o
'"
.~Ne •• rN~~4·~"-~O~~-OOOOO~_O~~O~~~_~~.~_~~~_ "'~~Nnll ~NN-~ ~.~ ~
~~-O.-.~~4_~_~~~~ ___ O~_~L4~4.~~40~~_OO~_a~~" ~- ~-
¢~~~Ge~~~a~~~~~~a~~~~~~~~~~o~~~~~~~~~~~e~~~~ .. li( )( Jill J( l( x .. lit ... " 1( 'II .... l( .oC ...... 1C II' II( )( " "II: 'II( )( :0( lit l( JC • 1( lI( ,.. 11( :0( lC 1( .. 11( ]I( )( II(
~~WY~wwU~U~~W~wwww~~~~~~~~~~w~~~~~~~~~~W~~~w
_-~~~~~~~~'~'~·~~'~~~"".~~~~eG~~~~e~~~~~~cac~ ~~~~~~~~~~~~ ...... ~ ... ~~, ... ~,~~,,~~~,,~,,~, ... ~,,~,~~ ~~~~~~~Q'~~~~'»~>vVVW~~=7==~~~'~~7~~~~> •••• ~~~~=~~ww~~uv,===r~www ••• <.w~~ ••••• ~QOQ •••• c •••••• ~~~=~~J2 ,~~z~c~~~~~~£~~4.···~.c ••• ~~.% ~~·._·._Z~~~~N'~~~·._C._~~_E~N_r~~~~N>4~"·~_ o--N~O-N~~ __ N~O_N~~_NNO __ ~~Q_NOe_N~C __ Nn __ N~

.. z .. '!I Z :) ~ .. 4
· Z \::I ,. S ., v
• OJ
· .. Z ... " III 'lI: ~
• :1-
· .. Z ... .. III
= Cl .. ~ •
Cl - z "0 H
CV III .. :;:, 0 c: z ..-+-l c: 0 u -
• M , ex: L&J ....J CO c:t I-
.. ;):_u~ .. ",
.... :J If(
.. , 4. "" Z ~ w
...... Yo!
109
~~N~O~_~~_':~_~n~~~m~~eO~N~~_~~~~~NR_~OO~O~_
N- • ~O>~~O~V~_~.~ -~ -~:
""~ -~
~~_~Ne~_~~NOft'~_._Nm~~.~~_~'~'N~~e~~.~o~n~~o -~-~N~'~NII~. ~~~4~_~~~ •• ~~~
_~_"''''=''~''II.",.~~ ... V''I
OO~~~~-~-~_N"'4~~~a~~~~o~~'O'='~Nooo~~nO~oo~ --~ ". ... 00- :-,
a~~~e~~~~~~~~~fte~a~~~"~n~~a~~e~~o~e~~d~~e~a~ x • • • ~ W ~ ~ x ~ x ~ ~ _ • x • • x ~ x x ~ x ~ • ~ ~ ~ ~ x ~ x ~ ~ ~ x ~ • ~ ~ • • • ... UU ... ~~W ... WW~W~W~~ ... ~W~~W~~ ... ~U~U~WW~WWW~WW ...... W ......
.e··.~"··'.·····.· .. '~~D.~~~r~~c.~~~o·~.~.· ~~~~~~~,~~~~~~~~~~,~~~~,~~~~~~~,~~~~~~~~~~~~
~ ~ 7 7 ~ ~ ~ .~ l~ .~ ~ ~ ~ - - ~ , ~ ~ ~ u V 7 ~ ~ ~ - T ~ ~ ~ = ~ ~ > > ~ % ~ ~ Z ~ J ~ !) ~ ~ :"t .. ~ ~ ., ? :- .. .&.1 tt' V V V C'I ~ ~ .;J I!J '.1 >4 fI!l Ia.I • ..: 'U *f • ~ s. ~ .. < ..... ':: :" '::I ") ... ~ :l .., .. .., -. .., .. ~ ., .., <'( ... " .. "t ", ~ '::' .., :! ~ ~ ::: '") r: -, .., ........... '7 ~ ~ < , .. ::I ... ,. ., ... ~ ., ... ~ ~ ., ~~~~G~·~·.·~~n~~.a"·_~_~_T~_.N~~Q~·.'.~N~Q~G ~ __ Nn_N~_N~_NO_~O_~~~N __ ~~N~_NO_~~_MO __ NN~_N

.. : :0 z :l JI 4
• -C
· Z 0 ,. 7 .~
'J
V
· .. .• ~U
'" _-J
" ~
!l .. 7
'" '" '.1 .. :l.
~
0 Z
0. - Z
" OJ '" ::s I-0 s:: ." ....
-+oJ s:: 0 u -
• M
I c:(
I.lJ -l CO c:( I-
110
~~~O_~~.O __ 4~~O.~~~.O~ •• O ___ OO.~_.~._.~~_."O ~- ~-
"o&U"JlIr"Q
Wla .... :z-y
... 'Z-u<c~.u
"Ul"rrtt.-t"'IJ _.~.~_~o~~.~~.~~_~_.~~~_~~~o~o~~~~~_~ __ ~o~~_ :-, -N
e :.a'!1e.~Q.aa."1a.e.e.:..a.:.c.~ :.c.ec..I1.c.'\.~~Q.~~ ~c:.c:.:'''O~o..~c.a.l\aa. e ~ ~ ~ ~ ~ x • ~ ~ • ~ ~ ~ v ~ x x ~ ~ ~ x ~ x ~ ~ ~ x • ~ ~ ~ x ~ ~ w • ~ ~ x ~ ~ x ~ x ~ W K ~~~W~~~"'w~w.w~.~~~w~ww.~~~ww~~~W~~~.~_~~WW~W
~····~······~····~····~~~:~o~~~n~~C~GGGo~eaG ,... ............ ""' ................ ,.. ............. ,... ........ ,... ....... ~ ... """ ......... "'"' ................ " ............. -,... ............... ~ '.: -!" ," .) ,~, n. :l ,..~ ::. - ,.. - ~ :> ~ "10 )0 :> 'J .) v v :! = ;: .~ ~ ": -= tt "t 1: :: It n. :. :. ::' a: )00 .. :- ~
e ~ ~ tI.I ':l ~ ~ '::t :) :I ,~ 'If I,! .... I,) v u u ;:) ~ .; ~ ::: 'J.) td i..J 'lJ .. 4 ~ 4 'JI I.J : .. : UJ .... ." -t .. "" c. ~ a. :l. .. <0( '" ~ ~ 4 • ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ! : • ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ _ ~ • ~ ~ • ~ ~ 4 4 ~ ~ ~ r ~~~·.-·.-.~~~~~·~-~·--~.-~~-~rN-~~~~~~~C~O •• NO--N"O-N~O--~O~-NnO-~~n __ ~~C_~~O_N~O __ ~~ __ ~

Z 4 0
3 » ~
~
· z 0 % Z 0 y
• y
• z ~ ~ ~
i • ~
~ ~ ~ ~ : ~
· 0 z
~ Z
~
~ ~
w ~ ~
~ 0
c z ~
+J C 0 ~ ~
• ~
I c(
~ ~ co c( ~
111
~~30n~~~~~_~~~~o~~~~~>~~~_,_~~#~ •• ~·R~~OOOOO ~-"~4 ~O~_~MN_.~~~N~~~~M.-~C-~~~~
~~OOOO~~O_~~_#~~~'~~~~'~h~~N_~~ __ o~~~~~eQ~~~ N ~-N~- ~- __ -~~ N
o~,~~oo~,~~·· .•• _··~·~~~._~.~~~~.~~_#·~o~oooO ~ •• ~ __ '~'~~~~~NN~~~~~NN_·N
.~~~~~ __ ~.~~·'-~ __ ?3=~~ __ ~_~~~ __ ~_~~·~~~_'~_
-~-~~~-~~4~-~-~~~~"" ~~ ~~. N4n ~ ~_
~3R.~~~~~~_~M~~~~-ft~'.NO.·"~~·~~~'~·_~·O~OOO -NN.~.~-~~~-~- N ___ ~~~-,~"ft._~~~~.o
----~-~~~~~
~'~~--G~-.~ON •• -N.~-_O'~_~~~~N •• • ___ NN.~~03' - ___ ~.N~ --
OOftO~OOOOoooooooooooooe~oeo~o~ooooooo~oooooo
.............•... A ~ Q ~ ~ e 0 ~ ~ ~ ~ ~ 0 ~ ~ a ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ = ~ ~ 1 ~ ~ ~ ~ ~ a ~ a ~ ~ ~ ~ a K ~ X • K ~ ~ x ~ < ~ x x W K W ~ ~ x x x x < x ~ ~ x ~ ~ ~ K X X X v x X X X X K ~ X X ~w~~w~~w~~~~~~_wu~wu~~~~wu~~~u~w~u~W~~WWMWWW
e~.~~~'~~~,~~"'~~~~~~~~~~~~Q~O~~~~~~~~~~~~~ ~,~,~~,~,~,~~,~~~~,~~~~,~~~,~,~~~~~",'~~~~~
=~§~~~~~~~~~~~~::~~~~~~~~a~S~!~~~~:::~~~~§~~ ~~,~,~"·<c~~~==~====Q='~ ••• Txrcc.~~=,~~~,,, -··~~~"~~·.~·~~~~~wo·-m-~-.N_~~~·.~~·.·.~N~.~ "O--N~-NO-~Q-NO-~O-~ON~ __ ~QNO_~~_N~_~C __ NNQ_

.. z ~ ~ i
.or
; 0 ~
! ;:;
'J . !; :Ai '" ... '!' .. ~
... :: .. Jt
'" OS II.
i
..-.. lI.
"0 'Z
cu I:J ::l ... c: ..,
...... z +0) c: 0 u "'-"
. ('I')
I <:: LI.J -I CO <:: to-
IoIQI.U'I.QQ<II
U&,,"."'''O
.: ..... :.z .. u
yc.,Juc .... ..,.c
:IlQ.-I%O~-O
::~OIZO .... ::C
112
uo~eNOOOOOOOCCOOO~OOOOOOOOOOOOOOOOOO~OOOOOOO
~"~~4._~.NCN-~_~~~"_=._~~ __ ~~~~_~C~O~3_0~_Nft ~~ -~M4~-~ N-- N
......... ~ ............•...............•..••• G. 0. e. :1 c.. a a. e. :1. ~ :. 0. a. 0. ~ e.. :1. 0. ~ "l. ':. ~ e.. c.. 0. " 0. 0. ~ '1 e: :1 0. :. ~ e. ~ e.. 0. Cl 1'_ a a. a. ~ ~ w w x • x ~ • x ~ ~ • ~ • • • x w • • x x • • K X ~ k X _ • w x x ~ x ~ x • ~ • • • ~~w~~~~~~~~~~~u~ ... u~~~uw~~~~w~w.~wuwwu~w~~w~w
~·~~~~"~,~·~~~~,~~~~~'~~ao~~~=~~~~~~~~~~~eo ~~~~,~~~~,~~~~~~,~~,~,~~"~~~,~,~~,-,,~-~,~~
~J~~~~~~~s.-- __ »»>uvuur~~z~-=~~~a~~~a~a~p .. :» ~ ~ :) ':!' ~ ~ ~I L.J ...,j 1..1 \",I U ..J U = =:; :: ~ :l .... ;,u w 'u ........ u .. :.1' 'I' I:J .. of ..r ...... '1 "'.. 'l. 0. .... of/( ~ ~ - ~ c < • ~ ~ n ~, c ~ ~ ~ = z = z z = ~ ~ 0 ~ ~ ~ , ~ ~ ~ ~ • ~ ~ • ~ .. .. • .. y • s o ... ,... .... '"" .. - "'"' .. - =: './' "'"t .,. .: ". :" .oJ "'" ,., ... 4# _ «:. • _ ~ ,r. _ .... or'! ~! - 'f' \t'I <'\t "" .. "I '" l7' .c ~ 0 .... NNO--~"Q-~N"--NOO_~~O_~NO __ NoO_NQn_NNO __ NO __

~ z • ~ z ~ • • • · z 0
i ~ ~
~
· ~ z ~ ~
~
i
A
· -% ~ • ~ = A ~
~ Z
~ i ~ -~
~ ~ ~
C ~
.~ Z
~ C 0 ~ ...,
• ~
". ~ LU ~ CO ~ ~
113
OO~O~~OOOO~_.N •• _~'.~O~~· ••• ~~O~MN-40~OOOOOO ~-_N~ __ N~~~_" ~~# - ~
?~~~~O'~~Q __ ~~~4~4~~~_~~·rD~~~~~~~._NN~_OO~~ --.~_~.~~~a~2':!~~~~~ ~~~~. ~~:
_~~OOOO'~~O_~N"N~~~N.~eO~QON~~On~OOOnoOOOOaO ~ ~
~--_'~~~,~~~n_·r.~' __ ·~~·N~.ftr~~~~~_~~~~Q~~_ ~ft~-~_~'N.d'_"~O'~~'_ -~a"'~~ -~~-~NN 'N~~~.a~~.N.~- _N__ _~e
oo-o=ftrft.'~~O~DNV'.~ __ ~Q~~M~_~3. __ '~ •• _~OOOO _~N~~ •• ~~._·.··~·~'~~~~~~o'~~.~~·N' __ ~_~ ____ N_~~~. ~~N
~~~o'~r-~~O'NN-4~_ft.Q~··~O'~~~.~~.~N_'n._oo_ __ N_ _ __ N_D'~~_N~._~
OOOO_~~.~N~n~'~'~·'N'~~N~~_~N'_O_COOOOOOOOOO
~- ~~~~~~~-~~- 0" ~A~~
~e~~~d~~~~~~~e~~~d~~~~~~~~~~~~~~~~~~~~d~~~~~ • ~ x ~ • ~ • ~ x x x • < x • x W. x x x x x x x ~ ~ X K ~ X ~ X ~ ~ ~ K X ~ X ~ X X X ~ w~uuwwww~u~ww~~~~wwuw~~wu~uuw~u~w~~~"uw~w~w~
eD~~~~~~~~~~'~~~~~~~,~'~~~~~ac=~~~~~c~~e~~,~ ~~~~~,~~~~~~~~~~~,~~,~,~~~,~~~~~~~,~,~~~,~~~
~~z=~=~~w~'~~~~.-_>'>uVV77=:=Z~~~=~~ •• ZZZZzw _ 4 ~ ~ ~ ~ ~ ~ ~ ~ : ~ w ~ ~' u u ~ ~ 0 = ~ ~ w ~ ~ oJ ~ W • 4 4 : ~ ~ < , c ~ ~ ~ ~ ~ ~ ~~'~~~~'~.«.m~~~~-z~~n='~~~~~~,·«c.r~"",,~ .-·.~~Q~~"·.~.~~~~~~D~-~-w-~N-n~~~~~~ ••• ~~~~ ~~~--~O-NO-N~-~Q-~O-~~NN--O?NO-N~_NO_NO __ NNO

~
~ ~ ~ C .~
~ C 0 ~ -.;
. ~
• c(
~ -J CO c( ~
~
% , ~ z , ~ ~
-. ~ % ~ ~ ~
~
~ % ~ ~ ~ ~ ~
~ . ~
Z M A ~ Z ~ . 0
·Z
& Z
~ . 0 Z
C4~U
114
.>O~_N~~._D~~~~ __ ~~O~#~~~ •• ~~O __ N~N~:.~n~'~O __ ~.~U_=~~._.~~~">.M.~~~ M ~ -~
r--.~~~~~~4~~~
O-O-OO~~.~O~-OC-4~4~~--~O---N-D~.~~OCO~C~~OO _ - -~ _ _"N
~ ~ ~ ~ ~ ~ ~ ~ i ! ~ ~ ; S ~ ; ; ; ; ~ ~ ~ ~ ~ ; ; ~ ~ ~ ~ ~ ~ ; ~ ~ ; ~ i ~ ~ ; ~ ~ ~ ~~U~~W~~~U~~~~~~U~~UU~U~~WW~~~UUUU~U~WWUWWWU
~·.·.· ••• ••• •• •• •• ···.·.·cc~ee~eeeD~~~oecOO. ~~,~~~,~~~~,~~,~~"~~~~~,~~",~~~"~~~~"~~,
~~~;~~~~~~~l~~~~;~~;~~~~~;:;;3E~S;;~=~~:[I=: ~ ~ ~ 4 ~ 4 C • ~ ~ ~ ~ ~ - ~ C ~ ~ Z 7 ~ = coO ~ ~ , ~ ~ ~ ~ _ ~ ~ T ~ Y • • c ~ ~ ~ "-.~~·._·._~a~~~N~~~O·._~._~~_~~N_~~N~~N~C"O _~NO __ N~"_N~O __ N~~_N~O_~~O __ N~O_NOO_NNO __ NO_

z .. a ~ ~ :II c
c . " i s ~ u
u
.. Z II III ~ ... ~
~
• :r .. A .. IS A .. 0 z
~
--- z
" <LI ... .. :::s ':I C ~ .....
-4-> C 0
U -•
(\")
• c:x: LU ...J CO c:x: ~
",oo,/oIQ.QQI/I
III%U"OCX"'':
.. :>z-u .... '"
':)cA,.:III_n
.,. Q. - a 0.1) -.'l
-"IOIlO=-
c,"",:t-I\OO
:"~a::o ... Oc
115
~~O"~~~N~O?O_~~~_~O?_OO~_O~_~?~~~OQOO-NOOaOQ ~N~~ ~
o~a~oOOOON_MN_~~~n~.'N#O~I/I~#O~~-I/IOn'·NA=~O~O __ 4_QNn n-~ # ~~-
OO~O~O~~~~?~~~4~~~~~4~~~O~~-~~~O~~~.~~~~N~ftO ~-o.n ~-~ - ~4-_~_ N~-n~N N
OO~~OQoooo~~n-nO=nn~ON.30~O"~NO-nO~OOOOOOOftO N
N·N~~.D~N .•. ~~N._~~~~.·_.N_~~·~.~_~_M~N~.~n. n-~·~ .• ~~~~~N~->~~;'~~.-. .~N.~~~O.
~~n.~'~~~--~Z~~N. ..~ N~~ __ ~ - -.¥j---.~OO~O-~_ •• N~_.D~I/I.4D_n~~~._~o~~~~_~ •• ~nooo • __ N N -n-N-.- N
nO--~4'-.nnDon~~I/I_~~n~N·_·CD4DO·4~ __ ~_nN~~._ --__ ~ ._.N -N
-~oo~o_~.~_~~~~_~_nN~ON~~NnOONN~O __ NO ___ Noon ~.-"' ___ "'N(\,f.\I_
.~-~"n-~~"O~OOO~~OO~OO~OOOOOO~~OOonoooooo~~o N"'N
ooOoono~~~I/I~O~~~~O~~'OD~O'.NN~D-O-OOOOO_OO~O N M~~~DGn.-_N ~_ 1/1
~ --
~~~~~---------------------------------------~j~~~j~~~~~~~~~~J~~~~j~~~jj~~j~j~jjj~~~ ~aa4~_~~~ _____ -~_~ _____ ~~~ ___ • __ ~~~ __ ~~ __ ~~~
w.XW_Z~~~~~~7=ZZZ~=~===~ZZ~Z=%==~ZZZZ2ZZZZZZ ~~WWwuuuuuUUUUUUUUJuuvUUUUUuUUUUUUUUUUUUuUVU
~~~~G'·~~'~'~~~~~~'~~"~~~~~~e:=e~~~c~~~~~~~ ~~h~~~~~~~~~'~~'~~~~~'~~~~~~'~~~~~~.~~~~~~~~
,.. ~ ~ ~ Z z .:: ..J j ...; !,!' \,'t ..., " ~ ~ - .... .... > :> ... ...,: v ~ l ~ Z :;, .., -: e C'" t" ~ 't ('P )0 ~ )0 Z Z 'Z Z Z "" 0( ..: :) ~ :) ~ ::: = ~ :) ~ ~ '.j ~ '.!..' 'J U ..... -= '.::' ~ I.iJ l" 1:,1 • I( .~ ~'t'· , .... :':. 0. ':. .. ~ ~ ~ ~ ::;l ::J ::> 7' 'T :" , ........ ., .., ~ .., -<r .. -r . .., .... ~., = r: -::: :: -:" ~ ::- : -'I .., '"" :- IJ. 3· 'T T ~ ., < ~ '7 7' ... ., , .., ... ·.-·.~~Q~·~·.·.~~~4~~~·_M_=-.N-:N~N~~· ••• ~N~ _N~Q __ NO_NO_NO_~O_NO_~O~N __ OO~O_NO_~O_NO __ NN

.. 'It -~ ; 11 <
., z 0 'It :r 0 u
u
.. Z III .: III .. ~
~
.. z: 'II '11
'" '" I\. .. 0 z • ~ - z
~ CIJ '" = .. c: :I
Z -+.) c: 0 u -•
("I') f
c::t LLJ ....J CO c::t I-
"".,JUC,J':'''
<II ... ·U C
%-c ... ..: ... ..Jc
,.. L"~ I'f '.&1-0
crt"\.-crO!l-O
116
~"~~~~~~_4~~0444D~~4e~~.~N~-~'O~NO-~~.~-~~.n._.O~_N.~~~~4.-~~~-"."N"- ~QN. n~-'--~-- •• -
"'...... ~fW- ..
~_o~o~_ouoo~o~o~_~~~~e __ .no_.~_~~~~~~~~._~.~ ., - ..,'"
-_.~O~.~~~_N.~_.~~~~.OQ~.Q~.:;_ •• n.~N~~.4~~-O~ ~~~~.,~~~.~~ __ ~_~~~ •• _.~~~~ _nN_ N.~.~_~~~~~~~~.~.~_~ -"~~. _____ NN
on--O-~-ONQe.N"ONo-.onoo-_~_~oo_.~oooooo~~oo - ~- - -
-----------.-------------~------------------~~~~~~~J~~~~j~~~~J~..JJJJJ~~~~~~~JJ~~jJ~JJjJ~j
~~~~~~~~~~~~~---~----~~-~-~~---~~----~~-~-~-3~~3555555355533~3~3355~55355353a~5533335t35
~~~~~~~~~"~~~,~~,~,~~'~~~~~e=~~~=e~~~~eGeo~ ,~~~,~~~~~~~~,~,"~,~~"~"'~,~"~~~~~,~~,~~
:i "'" .oJ ...I ,~ "' \.:J \j ".4 c:. : ... :. - "- - ~ ..." " "> "Jo >- V U '",,;, z;::'" 1: ,... "!' = - '= :: ~ "':" n :z :r a. e: ~ _ ::"'I ~ :: :;l ::: ~ ~ ::0 ~ '.J.. "JI ... U ..J V V 0 "! e -::: -::: .oJ 1',1 W Io,i -'t *' ... -f j!' L' ~, '.J -r ..... -1 '" '!. "\ '1 t".. otr , ~ , , '" ~ ~ ~ .. ~ ~ ~ ~ = ~ = ~ ~ Z Z = = ~ ~ ~ ~ ~ ~ , , ~ ~ ~ ~ X ~ Y y Y 4 '" ~ '" T .~o.~n~. __ ._~~~~~~~.~- .. -r._n~_r~N_;~N~~N~~~ O_NNO __ N~O-NNO __ ~OO_N~U-N~ft--NO~-NOO-N~O __ NO

",a"'''IDIl''O
t-::.z-u-r_au
.. z ., ., Z :2
" c
• • . z 0 ~ :r ~ \J
\J .. Z tV o/t II. :r ~
IS. . ... z .. '" " .. :I. .. 0 z
- ! ~ (1.J u ::::I ... s:::: z -~ s:::: 0 u -
• (Y)
I c=:: L.U ...J IXl c=:: I-
117
OO~~?004~~~~~_~?N~~~ __ ~.n4_~~NN~.N.-n.~14-0? ~~ ~ ____ ~~N_._~~~_ N__ --~ 0 N --
o~~o~~~~o~,oo.~oo-~~oo- __ oo~~~oooooono~~~_oo ...
~'~~~?10~O~_04~ •• -~.~o~_~~ __ e_ ... ___ ~_~_o~o~o~ _ ~n~._N - ~ - - ~~
~~QO~'~~O~Q~N~~GN~40_~~~~~~?O?~?~'O~OO-~~02~ ;,.1"1-.- .... _ .,
oo~o_oo~on~04_~~_~~~O ___ ~oc~oe~~OQ~O?_O~~OOo
" O~.~ ... Q~~.~4-~~~ •• a_ro __ ~~~.~~Q~~ .. NN~~~~_ •• ~
~ ~~.~.~-~ ..• " .. ~N~.-. ~4-.~ ~~M~~_~O .,_O.~·~.~·~ONN_N_~~ ~Q~--
. . "'-O.,1e~~~Q.~ ... ~ • .,.N~-Q=~~.~~W_."'~=-~~~-··.~~ooe _ _____ ____ N-~"
~
OO~6~~"'"'.N~~'~N'Q'N_~4.""N~~a·Q~.04_N~.~_"'O .~~---~~N ~~~. -- -
oooo~ot~~=~~~-~~-n~~-~N~N~O~~~~~~C~O~_~N~~ftO ~ --.-_NN U --
~-.OO-~"~-~~ftO~ocooooe~oeoooOoooooe-o~oo-o~~ .....
oe~~~ON--~:~~~.~~--~~O-~~~~~-N~~~~~~QOO~OOO~ ~~_NN=~e~~~~ ~~~ ~~ ~~N
.. N
~~~~~~~~~~J~J~~~~~~~~~~~~~~J~~~~~~~~~~~~~~~~
------~~-~~~~--~-~~--------------------~-~--7..%"'~Z7.~"'7.::'?:7"7::''?:!'l'';'!"!Z:::':--'''Z:!~?-::':'''?77'?Z':!ZZZ'Z'%ZZ-VWUW~WWVWUWUUWUWWVWJ~UU~VWVUw~w~UUWUVwVUUUUU
eDG~o~~'~~-~~~~~~~~"~~~~~~~ec~~~c~c~~~e~~~~ ,~~~~~~~~~~~,~~~,~,~,~~~~-~~,~~-~~~~-~~~,~~~
~. ~. Z ~ = = ~ ~ ~ ~ ~ ~ ~ ~ ~ - ~ ~ > ~, 0 ~.J 77 = = ~ ~ ~ ~ C ! = ~ ~ ~ Z Z Z % fI( .. C ·t ~ :- :", :; ~ == ,) ;) :) ., '"' ....J u " U \oJ .;) = :! 1..1 ~ ...J ... ~ I,J". .... -J "" 00{ -C ~ Q ~ 0( 4 '" :I ::;) :l ::l '·'·~-'~"' •. ~~~~~~=7="~~D_~~_.'··~ •• r""" -:> "'" • - "" If!' .. -. N " ....., ... ~ ,.. ..... ., "1: '~, =- .0 ::" ~ 0 .... _ -::. _ If" _:- , .... _ I/" .'.1 I/'" "<I ..:. .., .... <# " • ., f'J. __ N~O __ NO_NO_~O_~n_NO_~ONN __ O~~O_~O_NO_~O __ N

~
z ~
~ .. c
; S ~ <.> ... "oJ . ~
z '" ,~ .' i • Q.
~ Z ' .. '" '" z ~ .. 0 z
- it Z
"C cv w :::s ~
c: :)
..... z ~ c: 0 u -. ('W")
I <: L&J -' £Xl <: t-
IIIcu .. e.,..a
.. O..J",%-",
.. ~z-uc ... u
".,c~-·~z,. ...
%_C"'W.l..lC
ut="-\Z01t-O
r .. o~o-e!£l
118
~OOOOO~~OOOOOOOOOOOOOO~OOOOOO~OO~OQOOOOOOOOO
?OO"O~O~OOOOOOQ~OOOOOOOOOOOO~~OOO~~~'~OOOOOO
~_~~~~oooo~o~no~?~ooo~~a~_o~~~~_OD._~~~Q_~~~ ~ _ N
_~_.'_~~N .. _~_~~.~.~~~~ .. _~C~_~O~~O= __ ~_>Q.G "" ~ .c c) At ~ .. 1 'T (\j ... -;) ... , !"II • .., 1"1 _ ... ., ., Q ..... "" ,", -- .... oW ..... :.-, v - Q ~_ .~.~~~.~N- ___ ._N__ N __ _
.~~~-.~~~N.N~~-~~.~~~~-.~_~_~ __ O_~N~_~~_O~~ • • ~'~~~N~-~-~~-~~ - N-
OO-.~-O~O._O#N.N·OO~~~O_~~' __ ~N_~~.~_O_O __ OO ~ __ N ~~ _ N ~~_
j~~,jJ~~~~~~~~~~~~J~~~J~,j~,j~~,j~~~~J~J,jjJJ~~~~
~~~~~-~-~~-~~-----~----~~---~--~---------~~~ % Z ~ r z ~ = z z ~ z z ~ ~ ~ z z ~ ~ : 7 ~ = ~ z ~ z ~ ~ z ~ ~ z z ~ z z z ~ z ~ z z z ............ v ... v ... uu~WWVWVWVUWUVwVUuwwvuuwuwwuuvwuuuu ...
~~,~~~~~,~~~~~"~"'~~~~,~,~~~~~~¢~~ee=~eaD. ~~'~h~~~'~~~~,~~~~~,~"~,~,~,~~~"~~,'~,~~~~
Z~~~J~"0~~~~:'--~-»>~>UUyu:·:z~~-~~~~3~xa«~ ~ ~ =- ~ ~ :) ~ =' :: ::; I' f,.1 ....J ,.; '.J ,.J V \..I :: :: :"' :: ~ ""' .u 'J II. .. .0( <tC -« <...i l,J ~ ~ J ..... # « ..,. Q. C :l. 0. ~~~~~.4cc.~~A~~~=~:~Z=~====~~~~~~~~~S~Z2C.CC ~Q~~.~Q •• - •• -~~-~C~~~-"·.-~.-~~_~~N_r~N~~N~~ NO_N~o __ ~~n_~NO __ N~O_N~O_~NO __ Non_~OO_~NO __ N

o
" f ~
01"4Y 10 'O~.Y/O
I , ...... V '" 24 ... .lY1U
31 ,uy 18 o lj'JN 1ft
,'tJU"410
'~JIJN/7
22J\.u1'
06J'JL " !JJUl" 2: I.I\ .. L 7 1 0) 4 'JC. 17
I I .ut; I' 24 "IJ(~ I' O;~i:.P7'
a ,,':'i:P 11
"-,~L~71
("!.oIl": f "
,')Ue"" 2(U(I/1
"~'lllV 11
't.",uv " l();~ .HI " ~1ut.( 11 2Jl)f..( If 2e )t(" r, •• JII r .. 19
IItJ~N'8
O.':C'lltl O·}t=~d ,,,
22FEd1d OP"Il"of 16
'~"'HI16 Zl,Ut( (1, l)!j AIIG 79
'""~I~ 18 2GL~t" 'I;
f,)] ... AvlfJ
11'14" 1d
Z"'''Vl& 01 J.)" 18 • 4JIJN 78
1~JUN1T
TABLE A-3.
A
e: E f
,NfL (..l1l
(NfL
CN'L C·ftL C .. 'L
'''It.,. CNIL e."rL
''''L CulL CNIL ( .. Tl ( NIL
C"'I. CNtL CUlL ( fllt.. CNIL C.'HL CulL CNIL Crj Tl CNIL (..141l
CNTl (till
(t"L CH'L PHI.
L"'L Ct-4 ft.
U"L fUlL eU lL , ... ll
,,.IL CtnL ("IL
C'HL (NfL
CNIL CNTL CNfL
Ii I f e:
1/ 1/ 1/ 1/
II II 1/ 3 :J
J 3 l :I J 1 J J l l ;)
l :I )
)
1 J J l J l
l 3 1 3 :J ;)
3 J J J ;)
]
]
5 I D
t
u u U U
U U
\J
L L L L
L L L L L L L L
L L L
L L
L L L
L L
L L L
L L l L L
L L L L L L U
(Continued) NOTEI ~ • ~OT PHLStNT, P • PAESE~f. C • CuMMUN ..... AaUND4NT
p
" ~ E L
" 8 t ... U (. .. C .. C .. C .. , ... C
" C ... C II C
" C ... C A C ... , L
" C
"C
" C
" t
"C
" C
s P r. Ii
G E
(>
o o o o Q
IJ
<l o o Q
\l ;)
Q
Q
Q
Q
C o ~ o ()
o Q
Q
II Q
;)
Q
;)
Q
o Q
II
o o o Ii
" I)
I)
o o o
II A L
" "I
U
S
;)
o o o o a II Q
o Q
II I ;)
o u o
;)
()
I I)
U 2 I o oJ o l I OJ I)
l I U I)
o (I
I)
U
o II
o o :J
H
V U II
'I
"U .. I~" I' I.P Nil NP
"II'
NP NP Nil
Nf'
NI' t,U.J
NP "I' NI' N,>
Nf' til:»
Nil
UP Ni'
N" Nil
I' .... P I' I' II P
t ... II "II'
" I' II
" I' II
"I"
"I' NI'
"" N"
" M II H I p
o o
1 J
2~
I (I
o (I
o
" " I o o u u
" o o I)
o Q
o \I oJ Q
U
U I)
U <)
" o 0;
a o o
" o ., (j
o o o
4 II P
H
C .. 5 t
o o o o o o o J
o II I)
Q
Q
o o
" I o II II fa ()
o
J o ~
o I)
o o o o I)
u o \l
II o
o o
"
C 4 ... 11 t L L u
o o I)
o 3 II o o II oj
" II .. o 3 U
" Il o G I)
II 11 u U Q
o oj
II I)
o Q
~ o iJ o o Q
II Q
" o o II
I 5 U P o U
" o J l a II II II )
\l U
tI o J ;)
II I o I U
I Q
" I 1 .)
I Q
il ()
o a Q
J o I o J.
" U
o I)
II
H
Y ()
R o I U
E
o g
" I)
)
I o ()
Q
:J
iI I Q
I o o II o I)
Q
II I I)
!.i
I Q
:1
:J
15 !>
2l u to J. Z
o I II o
" " Q
r. P I H IJ It I II
o o o o ~
oJ II ,)
o I o I ;)
l 5 l l 6 a ., I 2 :! .... , (I
III
" IT It tJ
11 1>1
I" I' ...
1"8 h U.
1.18
Il' 110
g
[J
5 T fI
E I [)
2, I ..
" 91 1\1
241
" I) u Il .. I ,. II l 2
Iii JtI
l
fI' l l 2
" i!5 8'1
I I o 1 II
2 c)
U
1 ;J
J I!I
I 2
1& IhD 'l
I
II
I
" T f L L
" o I)
I (I
II II I)
o
" o l II o o o u
" I)
" " o ')
~
o o II I)
I)
U
I II II ;)
0)
')
J II
" " o I)
o o o
G
" S T P o p
o
II II II I I)
II :)
II
" o I)
" I)
o 'J
" I)
u
" " a
" II o
" o II o o
" I)
" o
" " ., " 'J
" II 0)
,)
" CI
H tI
'i U L
" I)
I)
o II ;)
II
" " " o
" II II I)
o o o I)
o 1
" o o o u o o II
" U
II 1
" II .J
o 0)
I o o I)
" T Ii .. "p H" Nfl
.. " ~,.
H'"II' , .... til'
t .. P'
Nfl
rd' tiP
N" ... " P
"P to r> C
" A C A
t
" C C
C .... "" e "p t (;
t P r.P .... N" " Nt> Nf")
NI'
3
" II
I 10 N y
E
i)
II II o J o iI U J ;)
U I)
(\
I)
:)
II II o o o
" ()
a o i)
()
o oJ .)
o o II a o o o II l I)
" " J o o
N C
" 5 r
IJ .. y
o o i)
II 11 ., I}
o o o iI o o o I)
o
I o I o o II
I
" I)
u o o o u , o o oj
I)
iI o o o o Q
o o
It
I C R
L.I P o H
o o
" Q
" " 3 I)
.l
.1
U I)
o 1.1 II o
" 1 I)
o o (\
II
o J U oj
II I)
I)
:J
" I)
u
u I)
.)
o oj
u '1 I)
I)
f U H
I C
" T
E
o o o U
1 U a oj
<)
o U Il o I)
Q
Il
" " " ;)
o
" o <)
l II
" o Q
o U
II U i) .. o o o o II o \I o Q
C Ao
L C
" L .. " o U II
" Q
" <)
o ... o
" U \I
" " " " " ;)
" I)
Q ., I)
I)
" o i)
\I ;)
" ;j
o o II o U I)
II I)
:I ;)
o o
o
• I)
e: M
" U .. o
" II
" J
" J o o i)
o 1 o I)
o o u
" J d o o o
" )
o o o o ,)
o o II o .. :I o o o I)
,)
o o o
• o L T U
~
I C
I:' II C T o II V
o
I) 0
" 0 I) 0 o II o 0 o 0
" 0 o 0 I) 0 0) I)
.. " ) I)
u " o 0 a 0
" 0 !> ;)
t. I I) 0
" 0 o 0 2 0
" 0 2 0 :I 0 o 0 o 0 1: 0
" II ') :I I) 0
o " II 2 ., 0 .) 0 2 I)
I) 0 o I)
o I)
o 0
I " II 0
:2 " " 0
c
" II I! P Q
o S
U .J .l 1 1 1 8
o 2 J o I 2 o 2 , 2
~
2 16 ..
6 10
" a !> I 6 "
1/ Q
1 I I 6 I .1 .1 i)
" o o
-----1.0

--c Q) ::s ~ -+) ~ 0 u .....-
. ('t)
I c:l;
UJ -I co c:l; t-
I-
~ 5 ., • <t
· 5 ,.. ! J
.. · .. ~ .. l :.
· .. z .. .. .... :: !l-
I-;) Z
'1 Z . .. ;; z
"."""'~IZ>Q
""2-U.~tJ
120
oooo~ooooooaooooooooo~o~OOO~OOOOOOOOO~OOOOOO
OOOOOOOOOOO~O~O~OOO~OOQoo~~on~O~OOOOQOO~~O~O
~~~J~JJ~JJJJJ4JJJJJj~J~~~JJJ~JJ~JJJJJJJJ~JJ~
~-~-~-~~~~-----~------------~---~--~--~--~-~ ZZtl.:;:zzzzZZZ====-'LZ=ZZ=Z;;::'zzz;:zz.£:!zzzzzzzzzzzz UUVVVvYvUwvuvuuvvvvu~vwvuuuwuuvvuuuuvuuuuuuv
~~~~~~~~~'~~~~~~~~~~~'~~·~·~~~~~e~~C~~~~OG.e ~~~~~~~~~~~~~~~~.'~~~"~'~'~~'~"'~~'~~'-~~~
Z Z J .J ..J -I '"' l" I" ., t~ :. :!. ':. ~ ,... ... to. ,... :> > > :II > U u v \,J Z Z Z ~ ~ -r ::: ~ u C' 7 :r Z C% '% Cf ~~~~~~=~=~~~~~_UUVLO~~~~~~~W~~CC.~W~ ••• ,.~~~ "'~"C~ ••• ~~~~=~~=z==~~=a~='~~~~~~L7·T7~CC. N~4"~··3~'-·.-~~'~~N'~~~·--~._=n_~~N_~~N~~~~ ~NO-NN~ __ ~~ __ NN~ __ ND~_NMO_NNn __ N~n_NO~_N~O __

.. z .. Q
~ '" c
c
· Z C
" ~ Q \J
• U
· .. z .. VI ... " Il
C.
-Z ." VI
"' " Il .. 0 Z
!l. Z -~ '" (1) ..
:::s " c: z -.., c: 0 u -
• ('t')
I ex: I.IJ -l al ex: I-
,,"' ...... oc.o
:-c_~~~c
%'>O"O-!:lU:
:>-QI%QNQC
121
O __ O~_~~Nft~·~.ru~-~_~~G~eN~NONO_N~.~~Q~·~~.NO _~N~~.~~ •• #~.= -.N4~ • _O~
-_~_~_N___ ~ -~-
O~~OQ~~nOO_~~~~~~"~O~~=~~O~~Q~~~~Q_~O~~~~~~~ ~ __ N_~ _~M_
~~~~~_~~~"_NN~~NNoO~3_0~O_~~N~~~~O_~N~=~ON~ _ _ N
ao~ooOOO~O~N-~OON~~-N~O~~-OOOO~-OOO=O=OOOOOO
-'"'
~~j~~~~~j~~~~~~~~~~~~~~~~Jj~J~~~~~J~~~~~~~j~
~~~~-~--~~~~-~~~~~-~-~~.---~----------~~--~~ Z ~ Z : 2 : Z Z Z ~ r z z Z 7 ~ r r = ~ = = ~ Z = ~ Z % ~ = Z = = : 4 ~ z ~ ~ 4 ~ Z ~ Z WVVVVw~~vuwwvvvv~VVVWW~UWUUUUUUVWWuVVvuVV\J\J\J
Gr~~O~~G'~~~'~~~~~'~~~~~h~~'~~~~~~=~~c=e~co. ,~~,','~,~-,~~~~,',~~~~~~~~~~,~~,~,~~"~~~,~
CE )00 ,.. > ,.. • ~ ..,. Z ..,. ~ ~ .J ! ':' U ':] ~ ." ...... _ :)0 '> , U V '.J Z ~ "":' = -: :;. ...z: ~ "! rz "'!' >- :J- )0 Z % C. .- ... ~ ... = ~ ':' ::" ~ :"I ":" :" ~ :' 1_' td II ''''; U U ~ -: ':' .4./ !U !lJ .. .r 'J.. w u .. c of .... ".. e. .., ." .... ~ ~ ~ • ~ ~ ~ ~ ~ , ~ ~ ~ , ~ ~ < ' " ~ ~ = = ~ = ~ ~ ~ = : ~ ~ ~ ~ ~ ~ T , •• C Y , ~ ~ ~ ~~O·.-~.~N~~·~ •• ·.~~~.~~M·_~_~_a~_~N~N.~ •• • • • O-_N~O __ N~_~O_NO_N~_NO_~ON~ __ OONO_NO_NO_~O_

-'0 Cl) :::J c:: .... ~ c:: 0
u -. M
I c;(
L&.J ...J CXl c;( I-
lIIa.., ...... o
~-uClO~Otr
~
% .. ~ z :) 0:1 ~
.. · z '=' lr • .~
\I
\I
· .. % III .. ~ i
• :&.
Z ;oj 1/ .. ::: ~ -:: z • i .. .. 0:1 1!
122
OOQOOOOOOOOOOOOOO~OOOOOOOOOOOOO~OOOOOOQOO~OO
OO~OOOOOO~OOOO~~~O~OOOOOQOO~oooooaooooo~oooo
~~~~C~"~-~=_aN~~~r~~~._~_~~_~oo~~eou~~o_O_o? -"4 _'" N
ooooo~~eo~oooo~~~~ocoooooooeOO~~Gooeoooooooo
jj~j~~~~~~~~~J~~~JJ~~JJ~J~J~J~JJ~JJ~J~JJjJ~~
~~--.---~~~-----~~~--~-----------.--~~--~--~ Z Z % = Z Z = z : z z z ~ = z z : : z = = z z z z z z = z ~ = = z ~ ~ L ~ = z : z : z z uu~uuuwuuwvuuuvvuuwwu~u~vuwuuwvu~~vvvuvwvvuv
~·.·.·~.·'.·~."' ••••• ' ••• • •• ~D:~~QQ~~~COODO ~~~~~~~~~~~'~~~~~'~~~~~~~~~"~~~~"~'~~'~~'~
Z:~J~~~~"~~:O~~'~-~~»»>UuvuZ77=~~=~~~"%~%~ :> ::l :' ~ :: ::t :;:0 ~ :1 ::> ~ ~ '..,) ..... ___ ~ _ ~ ...) = ~ .::. ~ .:) :.oJ I<.: U .... .of oCt ~ '" t- ...., !"., W < " oil o.L '" ~ a.. ~~~~"'C4C~."~~~==~=Z=Z7=~Q~""'~ •• ~~~%·X.c ~N~4~O·"~~.-~._-~~~G-~o~~~._~._.~_~d~_C~N~~N -NN~-NNO--.~O-NN~ __ N~O_N~O_NNO __ NOO_N~O_NNO_

~
z ~
i ~ ~ c
c
z e z • ~ ~
~ . -z ~ ~ ~ % ~
• ~
: ~ ~ ~ ~ ~
~
0 z
~ ~ z
" " ~ ~
~ ~
0 ~ %
or-~ ~ 0 ~ ~
• ~
I <C
~ ~ CO c( ~
123
~OO~OO~O~~O~OOM~~O._N.~_o~~~~n~~~~~MN~O-~~O. -N~4~~~~-N-~ -~-
~ __ O~_~O_ft~~~~.'_~'C~~_~.N~~N~~~~~'N~~'~4~·_ ~ •• u~,·~~m.fta.=~~~~~.- ~~-.- ~.~ •• __ ~.~_~~~~.~.~ _N~4_ • ~~
N
~ftO-~OOOO~~~~~·-~~N~-'~ •• ·G~~Q~~N~.N".-»~~__ ~_r~._~_NN~ __ N ~-~-~-~-~~~~~~~ .-O~O~OO_O~~~~~_4~~_N~OO_~NNM~O~¢~O··~.4_~ __ M~
~- --N~N~~-
~~OOOODoeo~-.~~o~~'~~~~-.ONQ~~N-~.~O~~-~OOOO
--~ -n-,~~o.- ~------o~~ooo~~oo~o~~oo~~o~~~eo~oooo~ooooooo~~coooo
~~~~~~~~~~~~~~~~~~~~~~J~J~J~~~~~~j~J~J~J~~~~
••••••••• ~~~~~~~~~~n~a~~~~~~~.~.~~~ft~~dOftdd~
~jJJ~JJJ~J~~J~JJJ~~~J~~~JJJJ~~J~~~JJJ~JJJ~J~
~~~-~-~-~~-~~~~~~--~-~--~-------~---~-~~~~-~ z z : ? r z z z z ~ = z ~ z ~ = z z z z z z z z z z ~ = z ~ ~ ~ z = ~ z z z z z z z z : UVYUuUVUUUUUUUU~V~U~~UUUUUyv~VVUUVuvuvuuVVUU
c=q~C~~~~~~~'~~'~~~~~~'~~~~'~~~~~=e~~~~Q~~~= ~~~~~~~~~,~~~~~~~,~~~,~~~~"~~"~~~~~~~~~~~~
f~::=:~§~~§~~~g~~1~~~~:~;;~~~5~5S~~~~~~~:::! ~.~~T~F~~~~~~~c.C~~~J~~z==~=~~~~~~··zccc~%r, ~~~O·.-'.~N~~·~····:~~~~wC~-E-r-~~-r~~N~M~.~ _~O __ N~O __ N~_~O_NO_NO_~O_~Q~N __ OeN~_N~_NO_~O

-"'0 Q) ;:, G:: -+oJ t: 0
U -. M
t c:t: L&J ....J CO c:t: t-
ftO., .. :)%-",
ioftc>_"z,..:u .. % ... ~ % ~ .:: -<
• • . %
~ % .:I
" " z .. Il't ~ -!Z ':)!J - 0
'" ... It '1
:>-::::11-0'.11
I\.
Z III
'" '" It I\.
to ,;) %
! :>OIZO .... OC
III ... c '"
124
~O~O~ •• O __ O_O_O_.~ __ ~ •• O_.OO_O_~OQ_._4.0_ •• ~ N ... _--
ooaaOOOOOOOC~OOOOOOO~OOOQOOOOOOOOOOOOOOOOOOO
NO~Ooooooono~oooo~ooooooo~oooonooooooooooooo
~o~oooooooooaoooOooo~OOOOOOOOOOOOOOOOOOOO~OO
O~QOO~O~OO~OOOOO~OOOOOOOOOOOO~OOOOOOOO~10000
~N~~~~~~~-··~-·.~a~~n~~~~4.-~N~~-OOOO~.~40.-0- D._~~~~~~n~N~.~N~~_ ~ -40_-N ~NO~ ~~ ••• ~~N~-
~~~ ~~ ~~~~·a~~~~e~~~~~~~~~~a~~~~~a~~~~~e~~~ zzz~zz~zz=z=z::~:ZZ%~z:z=Z%ZZZZZZZZZZZZZZZZZ
~~~~~~~~~~~~~~~~~~j~~~j~~~j~~~~~j~~~~jJj~~~~
~~~~~.-.~~~~~~~-~~--~~--~---~~~-~~~---~-~-~~ : z z : • : z z = z ~ ~ = ~ 4 ~ Z ~ = z z ~ z , = = z ~ ~ ~ z ~ z z z z z z z z z z z Z uuuw~wuvvvuv~~vv~uv~v¥~YUVw~~w~~v~~~uuuvuv~v
=L~~'~~~~~~'~~~~'~~~~'~~'~~~~'~~~=~~~=~4=GG. ~~~~~~~~~~~,~"~~~~~~~,~,~,~~~~~,~~~~~~~~~~~
'Z Z "2 Z ~ .oJ ...J ~ ".),~ \ .. ~ ~ :. 'l. :. - - - - ., :> '> > )0 v '- v .,J ::: ? :: ;: ~ = :: ~ 2 I c.;. ': '" II ~ ~ ~ ~ :: j ., ~ -::: :"I :" '::I !) 11.1 10J .. ' :ow V u ,.j ..... :; = = .., ) .,.. U ...J .u ., 4 ~ -< <.J ....: u.. '"" ~ .. ~ -< 4It Q. ~ ~ ~ ~ , ~ , ~ < ! ~ • • ~ ~ ~ ~ ~ ~ c = z = ~ ~ ~ ~ ~ c 0 ~ ~ ~ , ~ ~ ~ ~ ~ 7 ~ 7 X c .~N~~~o.n~~.- •• -:~~~~~~~~o .• -~._=~_r~N_~~~~~ --NN~-~NO--N~O-~~)--~OO-N~O_~~O __ ~OO_NOO_~NO

TABLE A-3. (Continued) ,<on: I NFo • tiU, "RESENT. P • PAESENT. e • (O .... UN.
" " " C tf !o It G a ., II Ie " V p 0 I ..
5 " 0 p P P I 0 I I " S
" P L A .. H " 5 H ;0 T , 0 R 5 5 A U A 0 I ( E U U 0 ,., E A A- t I I N N N Z P A I. I' I Ii E L 0 T L , 0 E (', II U 0 5 I. U 0 I I I. P E f E E I. E 5 A U E D 0 E 0 0 " 0
U4P,f '8 ,HTL !'> U ~ 0 0) .... 0 0 0 0 , 0 0 0) 0 ....... PR 18 eH'L s u It II 0) ,,:. 0 0 0 0 0 0 , i) 0 21> A ... n 18 (;1111. S U e 0) 0)
"''' 0 0 0 0 0) 0 to 0) 0 03;40 V 1~ eH'L s u .. 0 0 "'i' 0 ~ " I ~ .:I .... 0 0 IO .. AY'8 ("TL !j U D 0 0 Hi' 0 0 0 0) 0 .) I) 0 1 , .... Y 7,. (NTL S U ( 0 0 .... 0 0 0 0 I) 0 1 " 0 24,l1(Ay 1t1 (N'L 5 U " 0 0 Np 0 0 0 I 0 0 0 0 0 31 .. "V711 e .. TL 5 II 8 I) 0 Hi' 0 0 0 0 0 0 6 0 I 07,iUN18 e"'L 5 u C 0) 0 NP 0 0 I 0 0 0 20 0) 0 ._ .iU"'. (NTL S U A 0 0 N" 0 0 0 0 :I 0 .n 0 0
" • 4 .. UNO\NT
5 H ... , " C I U .. v w ( If
II I S R I G " G T ., C U , OJ It P " I. E y R 0 T
" " E y R E
0 HI' J 0 0 ~
0 NP 0 0 0 0 I)
"''' 0 0 0 0 0 H" 0 0 0 0 I .. P 0 0 0) 0 0 .... .) 0 0 0 0 NP 0 0 0 0 .) No' 0 0 0 0 0 NI> 0 0 0 0 0 NI> 0 0 0 0
c 0 A
" I 0 I. 0 I. C C , II N U I. N N G U I A- N C
0 0) 0)
0 0 0)
0 0 0)
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
E Ii C T D Ii y U
Q
0 0 0 0 0 0 0 0 0
C 0 P £ P D U S
0 0
• I :J ;I .)
0 I a
-.a N (J1

126
TABLE A-4. List of fishes collected by rotenone from the experimental reef on October 12, 1977. The feeding categories are assigned to each species on the basis of previous literature or stomach analysis. C = carnivore, P = planktivore, CP = coral polyp or mucous feeder, 0 = omnivore, H = herbivore, and 0 = detritivore. A question mark is noted for those species whose feeding habits are not well known or defined.
Total Mean Std. Feeding Nc. Wt.(!;') Wt.(e) Length ( c:n ) Cateeo~y
~ 5'P ~ S:P
CLUPEIDAE 8 792 Sardir.ella m~~uesensis 8 792 98.9 ;j: 8.2 19.1;1:0.5 P
SYNODONTIDAE 24- 1704 Saurida r,racilis 23 1657 68.6:' 80.7 15.5: 6.4 ~
" S;IEodus v1lrie~3.tus 1 4-7 46.8 15.0 c
HURAlliIDAE 212 17807 Uro-pteryc::inus ti~inus 2 910 255.0 t 278.5 89.0±5.7 c U. fUSCO~U~~~tU3 4 103 25.e±12.2 26.1 ± 3.~ C Echidna zebra 6 4178 696.3 t 213.7 73.C::: 9.S C E. polyz~ 1 56 55.7 30.5 C ., GY1nl'lothc!""lx eurost:.s 19 2223 121.3 ± 98.9 33.2 ± ,3.1 C G. steinr.~ch~er: ., 96 95.6 33.0 c Q:. flilvi:r:Clr::-i:1"ltc:s 3 252 84.0 ± 41.7 36.7:: 5.3 c !!. jav;mict:S 1 1912 1911.5 89.0 c G. !r.clea<-:ris 1 257 25E.5 43.C c G. undul;;'tus 164 7631 46.5 ± 177.3 24.6 1 9.7 c Anarchbs leuc'E'-IS 7 107 15.3:i; 5.0 25.1 ± .2.1 C ~
G. sracilcaud~:us 1 39 38.9 28.0 c G. buroer..sis 1 38 37.9 28.0 C Gymnothcr:u: sp. 1 5 5.0 5.6 C
CONGRIDAE 8 400 Con~er ciner('us 8 400 50.0: 56.2 28.1±11.2 C
AUIDSTO:!IDAE 24 1142 AulOGto:::us chinensis 24 1142 47.6128.7 26.91 6.0 C
SYNG:'lATH'DAE 2 1 Do~:'h::tL'phus r.:elanonleura 2 1 P
HOLCC:::lTnnAE 5 200 Adior:r.c l~ctco~utt:lt"..!S 2 20 9.6 1 3.7 7.'1± 2.8 C ,a. di3.ce!:".a 3 180 60.1 ± 17.2 12.5± 1.3 C
BOTHIDAZ 1 35 Botr.us mancus 1 35 34.5 11.5 c
PLL.""'UrtONECTIDAS 2 22 Sat:".ar i 5 C '-IS t!"iocellutus 2 22 10.8;1; 2.5 8.If: 1.1 C ?
II

127
TABLE A-4. ( Continued)
Total Hean Std. Feeding
No. Wt.(g) Wt.(g) Length(cm) Category
PRIACANTHIDAE ? 368 Priacanthus cruentatus ? 368 .52.6:11.0 12.3 =1.2 P
APCGONIDAE 1216 5089 ~ brachy~a~na 1090 2066 1.9·1.5 3.2 ± .7 c Apogon erythr~~us 3 7 2.3 ± 1 .0 3.5 ± .8 c !. snydori 123 3016 24.5:12.7 8.8:2.1 p
CARANGIDAE 11 917 Caranx me lar.roysus 11 917 c.J.4 ; 9.6 15.2 ± .5 ...
" MULLIDAE 25 2866
Mulloidichthys flavolineatus 12 2002 168.5: 49.2 19.5:1.8 C M. vanicolcn.sis 9 633 70.3:: 11.0 14.9 ± .6 c Paruneneus :Jor:Jhyre~lS . . 2 192 96.1 :1013.9 14.7:1; .4 C E. multifasci~t~ 2 39 19.2 ± 12.9 9.3 ± 2.5 c
CHAETOOOr.'TIDAE 428 18330 Chaetodon auri~a 5 208 41.6=8.3 10.1 =.5 C c. ornatissir::us 1 10 9.5 5.8 CP C. miliaris 421 18063 42.9 ± 20.4 9.3:1.J P Heniochus acuminat~ 1 49 49.3 9.8 p
POMAC~rTRrDAE 819 56567 Dascyllus albise~la 99 3708 37.5 -:16.2 8.111.8 p Abudefduf abdominalis 666 50916 76.5 =25.4 11.7=1.5 P Chromis hanui 2 24 12.1 : 2.0 6.2 ± .4 p
£. ovali-s-- 7 139 18.8 ;!; 6.4 7.3 ;; .8 p Plectror,;lY't)nidodon johnstoniamz:: 1 10 9.7 5.6 C -Eupomacentrus fasciol~tus 44 1770 40.3 ±9.6 9.0: .9 0
LABRIDAE 110 1.;004 Bodianus bilunulatus 5 180 35.9 ;29.6 1305±11.4 C Cheilinus r~odcc~rcus 4 85 21.3±26.1 8.5 ±3.8 ...
" ThalasGorr:a d~1'c"e~ .. 1.... 67 2845 42.8 ±24.5 11.6 =2.3 C Gomphosus varlt:S 25 748 29.9 =17.0 11.7 =2.1 C Stethojulis balteata " 85 14.1 :9.0 7.6 ± 1 .4 C 0
Macrooharz~~cdon ~eoffroyi 3 61 20.2 ± 16.1 7.9" %2.5 C
SCARIDAE 545 10683 Scarus persnicil1at~ 19 2861 150.6 "i:.77.2 14.9 :;1;2.8 H s. sordidus 142 3246 22.e±11.4 8.0 ± 1.4 H S. taeniarus 40 1928 48.2 = 16.2 10.6 :I; 1.5 H Scarus spp. ( j uvenHes) 344 2648 7.7",6.6 5.7 !i:1.5 H
ZANCLIDAE 3 275 Zanclus cnnescens ('"'.-, ....... ., 3 275 91.5 ;-£4.3 10.8 J::3.9 C

128
TABLE A-4. (Continued)
Total Mean Std. Feeding No. 'it.(g) 'it.(g) Length(cr.l) Category
ACANTHURIDAE 88 4933 Acanthurus nilTofucC"us 2 22 11.2;1;2.1 6.9;1; .9 H !. Eioste'ius sanci'licensis 7 104 14.9;1;12.3 6.8 ;I; 1 .6 H A. xanthoc:ter::s 4 255 63.7 ± 37.2 10.8 :1:3.1 H Ctenochaetus scri~osus 15 710 47.3=33.5 9.1 =2.5 D Zebraser.,(l. flavesce!1S 54 2856 52.8 ; 17.2 10.2 ;!: 1.6 H z. velife!"urn 5 938 187.5 ! 60.0 16.8 :t 2.1 H E. unicornis 1 48 47.9 11.1 H
ELEOTRIDAE 988 511 Asterronteryx s~~ipunctatus 988 511 .5 D ?
GOBIIDAE 23 15 BathI:sobius cottice-::>5 6 8 1.3 ? B. fuscus 5 2 .4 C Gnathele:lis sp. 7 4 .6 ?
1 0.2 ? Gobiidae sp. 4 1 .3
BLENNIDAE 6 17 Cirriuectus variolosus 2 16 8.0 H I6tible~illius zeora 4 , .3 D
BROTU-LIDAE 4 1525 Brotula multibarbata 4 1525 381.2 ± 143.1 32.8 :4.8 C
SCORP A.E!\IDAE 31 1582 Dendrochirus brach~terus 11 326 29.6 ± 28.2 8.0 =3.1 C ScorpaenO"DSlS iSiobo~ 15 1061 70.7;67.5 10.9 ± 3.4 C s. cacot:sis 3 148 49.3 ± 37.9 9.8 ± 4.4 C Sco~paena conierta 2 47 23.5;1;1.4 7.8; .4 C
MONACANTHIDAE 14 349 Pervagor suiloso~a 14 349 24.9:.t: 5.4 8.7 ± .6 0
OSTRACIONTIDAE 1 9 Ostracion melear.ris 1 9 8.5 4.4 c
CANT!lIGASTERmAE 15 128 Canthigaster jact~ 15 128 8.5;1; 2.4 4.8 •• 5 c
DIODOtlTIDAE 4- 7513 Diodon hystrix 3 7112 2370.7 * 900.9 36.3;1;9.1 c E. holocanthus 1 401 401.0 17.0 C
ANTENrIAR I In ,\.E 2 41 Antenr.arit.:.S dror:.bus 1 14 14.2 5.7 C !. polucce:-..s:'s 1 27 27.0 6.8 c

FIGURE A-4. Water temperatures at four locations in
Kaneohe Bay (Smith, unpub1.). Site "CE"
was in the central sector of the Bay
approximately 150 m southwest of the
two study reefs.
129

130

FIGURE A-S. Rainfall in the Kaneohe Bay watershed (Smith t
unpubl.) .
131
lLJ Z a: ::.::: -a: :::;c:
t.n OJ OJ
o C) N -
C) C) CO
C) C) - C)
lLJ -l C)
:c a: a: :::;c:
I
c-en co
C) C) CO
a: :x:: :::l a: :::L:
lLJ :c C) lLJ Z a: ::.:::
I
co c-
(SH313WIllIWl ll~JNI~H llHINOW
C) C)
ton c-en
~ c-en
U"'J <.0 en
= = en
132