serotonin and neuroplasticity – investigated in vivo by
TRANSCRIPT

1
Serotonin and Neuroplasticity – investigated in vivo by
Positron Emission Tomography and structural
Magnetic Resonance Imaging
Doctoral thesis at the Medical University of Vienna
in the program Clinical Neurosciences
for obtaining the academic degree
Doctor of Medical Science
submitted by
Christoph Kraus, MD
Supervision by
Rupert Lanzenberger, Assoc. Prof. PD MD
NEUROIMAGING LABs (NIL) - PET & MRI & EEG & Chemical Lab
Department of Psychiatry and Psychotherapy
Medical University of Vienna
Waehringer Guertel 18-20, 1090 Vienna, Austria
http://www.meduniwien.ac.at/neuroimaging/
Vienna,
July 2015

2
I. Table of Contents
II. Declaration ................................................................................................................ 3
III. Abstract ..................................................................................................................... 4
IV. Kurzfassung .............................................................................................................. 5
V. List of Publicaitons .................................................................................................... 6
VI. Abbreviations ............................................................................................................. 7
VII. Acknowledgements and Project Funding ................................................................... 8
I. BACKGROUND ........................................................................................................... 9
1.1 General introduction .................................................................................................. 9
1.2 The role of neuroplasticity in health and disease ..................................................... 10
1.3 Mechanisms regulating neuroplasticity .................................................................... 11
1.4 Crosslinks between Serotonergic Neurotransmission and Neuroplasticity ............... 13
1.5 In Vivo Quantification of Brain Anatomy with MRI .................................................... 16
1.6 MRI quantification of Neuroplasticity: Voxel-based morphometry ............................ 18
1.7 In Vivo Quantification of Neuroplasticity in the Serotonergic System ....................... 20
1.8 PET radioligands for the Serotonin-1A receptor and Serotonin Transporter ............ 21
1.9 PET quantification: binding potential ....................................................................... 22
1.10 Neuroplasticity quantified with PET and MRI – previous results .............................. 25
1.11 Open questions ....................................................................................................... 27
II. AIMS of the THESIS ................................................................................................ 29
III. RESULTS ................................................................................................................. 30
3.1 First publication ....................................................................................................... 30
3.2 Second publication .................................................................................................. 54
3.3 Third publication ...................................................................................................... 82
IV. GENERAL DISCUSSION and RAISED QUESTIONS ............................................ 104
V. CONCLUSION and FUTURE PERSPECTIVES ..................................................... 107
VI. REFERENCES ....................................................................................................... 108
APPENDIX – CURRICULUM VITAE ...................................................................... 124

3
II. Declaration
This thesis was conducted at the NEUROIMAGING LABs (NIL) - PET & MRI & EEG &
Chemical Lab (http://www.meduniwien.ac.at/neuroimaging, head: Assoc. Prof. PD Dr. med.
Rupert Lanzenberger) at the Department of Psychiatry and Psychotherapy (head: O.Univ.
Prof. Dr. h.c.mult. Dr. med. Siegfried Kasper), Medical University of Vienna.
All PET measurements were performed at the Department of Biomedical Imaging und
Image-guided Therapy, Division of Nuclear Medicine, (http://www.radiopharmaceutical-
sciences.net, Leading co-investigator: Assoc.‐Prof. PD. Dr. Wolfgang Wadsak, PD. Dr.
Markus Mitterhauser), Medical University of Vienna.
All MRI measurements were performed at the MR Centre of Excellence in collaboration with
the Center for Medical Physics and Biomedical Engineering (co-investigator: Assoc. Prof. PD.
Dr. Christian Windischberger).

4
III. ABSTRACT
Neuroplasticity is defined as the sum of structural and functional neuronal adaptions in the
adult brain upon changes of external stimuli or the internal milieu. After neuroimaging
demonstrated restructuration in the brain associated to navigational expertise or learning to
juggle neuroplasticity has received significant scientific attention. Moreover, deficits in
neuroplasticity are thought to contribute to the pathogenesis of depression. Meanwhile,
neurobiological studies revealed many new insights into the mechanisms behind the
regulation of neuronal development, synaptogenesis or neurogenesis. Conclusive evidence
indicates a modulating influence of serotonin (5-HT) in triggering measurable morphological
changes of neuronal cells. While this notion emerged mainly from animal studies, there is a
lack of studies in humans on neuroplastic functions of 5-HT.
We therefore investigated the relationship between 5-HT and neuroplasticity in humans with
a multimodal neuroimaging approach using structural and functional high resolution MRI in a
combination with PET and imaging genetics in three consecutive studies. In a first study, we
demonstrate that binding potential of the 5-HT1A receptor, which is the main inhibitory
serotonergic receptor and which has been frequently linked with modulation of neuroplastic
processes, correlates with regional gray matter volumes (GMV) in distinctive brain regions.
Furthermore, we found a correlation between 5-HT1A autoreceptor binding in the dorsal raphe
nuclei (DRN), known to modulate the forebrain’s serotonergic tone, and cortical GMV. These
results indicate that 5-HT1A receptor densities in certain brain regions interrelate with the
volume of gray matter.
In the second study, we report a significant increase of gray matter after 10 days of oral
selective-serotonin reuptake inhibitor (SSRI) administration in the posterior cingulate cortex,
which is accompanied by an increase in functional neuronal connectivity in the same region.
Here, elevated synaptic 5-HT due to oral SSRI intake, which represent the most often
prescribed antidepressants, leads to dynamic alterations of brain structure and function as
measured by MRI in healthy humans.
In the third study we did not substantiate previously reported increases in 5-HT transporter
(SERT) or 5-HT1A bindings upon lifetime changes of brain-derived neurotrophic factor (BDNF)
function, as produced by a common single nucleotide polymorphism (SNP).
The work of this thesis provides ample evidence that many of serotonin’s neuroplastic effects,
which are highly active during the brain’s development, might be partly conserved throughout
lifetime. Under consideration that both deficits in neuroplasticity and serotonergic function are
important hypotheses in the etiopathogenesis of depression, this thesis offers solid
groundwork for approximating these pathomechanisms in future studies. Finally, the
relationship between 5-HT and neuroplasticity detailed in this work, could lead to further
insights in the brain’s ability to adapt itself.

5
IV. KURZFASSUNG
Nachdem Studien der bildgebenden Hirnforschung zeigten, dass die graue Substanz bei
Erwachsenen dynamischen Veränderungen unterliegt wurde der Neuroplastizität viel
wissenschaftliche Aufmerksamkeit zuteil. Außerdem stellt dysfunktionale Neuroplastizität
eine Haupthypothese der Neuropathogenese von Depression dar. Studien in Zellkulturen
oder Tiermodellen identifizierten zahlreiche Mechanismen der Neuroplastizität, welche
neuronale Entwicklung, Synaptogenese oder Neurogenese steuern können. Hier gibt es
deutliche Hinweise für einen Einfluss von Serotonin (5-Hydroxytryptamin, 5-HT) auf
morphologische Veränderungen von Neuronen. Während dies vor allem bei neuronalen
Netzwerken während der Embryonalphase und während der postnatalen Gehirnentwicklung
nachgewiesen werden konnte, fehlen Studien bei Erwachsenen.
Diese Arbeit setzte sich daher zum Ziel, das Verhältnis zwischen 5-HT und Neuroplastizität
in vivo bei Menschen mit struktureller und funktioneller Magnetresonanztomographie sowie
Positronen-Emissions-Tomographie (PET) und Genetik zu untersuchen. In der ersten Studie,
zeigen wir starke Korrelationen zwischen dem 5-HT1A Rezeptor, welcher neuroplastische
Aktivität vermitteln kann, und dem regionalen Volumen der grauen Substanz in bestimmten
Gehirnregionen. Wir fanden außerdem eine Korrelation zwischen 5-HT1A Autorezeptordichte
im dorsalen Raphe Nucleus, welcher die Aktivität des serotonergen Systems regelt, und
grauer Substanz in höheren kortikalen Regionen. Gemeinsam zeigen diese Ergebnisse,
dass die 5-HT1A Rezeptor Dichte mit mehr grauer Substanz einhergeht. In der zweiten Studie
finden wir starke Signalzunahmen grauer Substanz nach Einnahme eines selektiven
Serotonin-Wiederaufnahmehemmers, was wiederum mit einer Zunahme funktioneller
neuronaler Aktivität vergesellschaftet ist. Hierbei setzt eine vermehrte synaptische 5-HT
Konzentration molekulare Prozesse in Gang, die mit einer Umstrukturierung grauer Substanz
einhergehen. In der dritten Studie konnten wir zuvor berichtete Veränderungen des
Serotonintransporters oder 5-HT1A Rezeptors in Zusammenhang mit reduzierter
Verfügbarkeit des „brain-derived neurotrophic factors“ (BDNF) nicht bestätigen.
Diese Arbeit bietet wichtige Belege dafür, dass neuroplastische Effekte von Serotonin,
welche an der Entwicklung neuronaler Netzwerke beteiligt sind, auch in adulten humanen
Gehirnen aktiv sein könnten. Unter Berücksichtigung, dass sowohl Defizite im serotonergen
System als auch dysfunktionale Neuroplastizität zwei Haupthypothesen der
Neuroätiopathogenese der Depression darstellen, generiert diese Arbeit eine wichtige Basis
für eine Verbindung beider Hypothesen. Schließlich könnte der Zusammenhang zwischen
Serotonin und Neuroplastizität zu weiteren Einsichten über die Fähigkeit des Gehirns sich
selbst zu adaptieren führen.

6
V. LIST of PUBLICATIONS
Publications:
Kraus C, Hahn A, Savli M, Kranz GS, Baldinger P, Höflich A, Spindelegger C, Ungersböck J,
Häusler D, Mitterhauser M, Windischberger C, Wadsak W, Kasper S, Lanzenberger R.
Serotonin-1A receptor binding is positively associated gray matter volume – A multimodal
neuroimaging study combining PET and structural MRI. NeuroImage 2012 Nov
15;63(3):1091-1098. Epub 2012 Jul 23 [2014, IF: 6.35]
Kraus C, Ganger S, Losak J, Hahn A, Savli M, Kranz GS, Baldinger P, Windischberger C,
Kasper S, Lanzenberger R, Gray matter and intrinsic network changes in the posterior
cingulate cortex after selective serotonin reuptake inhibitor intake. NeuroImage 2014;
84:236-244. Epub 2013 Aug 26 [2014, IF: 6.35]
Kraus C, Baldinger P, Rami-Mark C, Gryglewsky G, Kranz GS, Haeusler D, Hahn A,
Wadsak W, Mitterhauser M, Rujescu D, Kasper S, Lanzenberger R. Exploring the impact of
BDNF Val66Met genotype on serotonin transporter and serotonin-1A receptor binding,
PLOS-One, 2014 Sep 4;9(9) [2014, IF: 3.23]
Related Publications:
Hahn A, Wadsak W, Windischberger C, Baldinger P, Höflich A, Losak J, Nics L, Philippe C,
Kranz GS, Kraus C, Mitterhauser M, Karanikas G, Kasper S, Lanzenberger R. Differential
modulation of self-referential processing in the default mode network via serotonin-1A
receptors. Proceedings of the National Academy of Sciences (PNAS) 2012 Feb
14;109(7):2619-24. [2014, IF 9.67]
Kranz GS, Hahn A, Baldinger P, Häusler D, Philippe C, Kaufmann U, Wadsak W, Savli M,
Höflich A, Kraus C, Vanicek T, Mitterhauser M, Kasper S, Lanzenberger R. Cerebral
serotonin transporter asymmetry in males and male-to-female transsexuals: a PET study
with [11C]DASB. Brain Structure and Function 2012 [2014, IF: 5.62]
Baldinger P, Hahn A, Mitterhauser M, Kranz G, Friedl M, Wadsak W, Kraus C, Ungersböck
J, Hartmann A, Giegling I, Rujescu D, Kasper S, Lanzenberger R. Impact of COMT genotype
on serotonin-1A receptor binding investigated with PET. Brain Structure and Function 2013.
Epub 2013 Aug 9. [2014, IF: 5.62]

7
VI. Abbreviations
5-HT – 5-hydroxytryptamine, serotonin
5-HT1A – serotonin-1A receptor
AMPA – α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor
BDNF – brain-derived neurotrophic factor
BOLD – blood oxygen level dependent
CNS – central nervous system
CREB – cyclic AMP response element-binding protein
CSF – cerebro spinal fluid
DARTEL – diffeomorphic anatomical registration using exponentiated lie algebra
ERK – extracellular-regulated kinase
FA – fractional anisotropy
GMV – gray matter volume
GRE – gradient echo sequences
NMDA – N-methyl-D-aspartate receptor
MAO – mono amino oxidase
MAPK – mitogen activated protein-kinase
MPRAGE – magnetization-prepared rapid acquisition of gradient echo
MRI – magnetic resonance imaging
PET – positron emission tomography
ROI – region of interest
SERT – serotonin transporter
SNP – single nucleotide polymorphism
SPM – statistical parametric mapping
STDP – spike-timing dependent plasticity
TE – echo time
TR – repetition time
VBM – voxel-based morphometry

8
VII. ACKNOWLEDGEMENTS and PROJECT FUNDING
I would like to express my dear thankfulness to my family and my friends. I could not exist
otherwise.
This work could not have been done without the direct participation and supportive energy of
young, motivated and funky researchers at the Neuroimaging Lab. With big support by it’s
fabulous members, this group was able to grow continuously as demonstrated by an
increasing number of scientific publications during the timespan this thesis was completed.
Therefore, I would like to express my gratitude to the “brain” behind this group, Rupert
Lanzenberger, Prof. MD, for supervision of this thesis. His friendly nature, scientific sharp-
mindedness and creative ideas were in concert a major stimulus for this work. Additionally, I
would like the thank the two members of the thesis committee Prof. Dr. Wolfgang Wadsak,
head of the Radiochemistry and Biomarker Development Unit at the Department of
Biomedical Imaging and Image-guided Therapy and especially Prof. Siegfried Kasper, MD,
head of the Department of Psychiatry and Psychotherapy, who is always eager support for
young scientists with his rich experience gained during his long and outstanding international
career.
Furthermore, I would like to thank the entire team of the Neuroimaging Lab, Dr. Andreas
Hahn for is support with computing, Dr. Marcus Savli for PET-modeling, as well as Dr. Pia
Baldinger, Dr. Anna Höflich, Dr. Georg Kranz and MSc. Sebastian Ganger for their clinical
and scientific support in a great variety of topics.
Regarding the hardships of simultaneous clinical training and scientific work, I would like to
use this chance to thank Prof. Dr. Richard Frey, Prof. Dr. Dietmar Winkler, Dr. Sandra Strnad
and Dr. Eva Resinger for supporting this scientific work and provide a series of releases of
duty to pursue the scientific studies invoked below and to attend international conferences
and scientific meetings.

9
I. BACKGROUND
1.1 General introduction
Neuroplasticity is defined as the sum of structural and functional neuronal changes in the
adult brain as a response to changes of external stimuli or the internal milieu (May, 2011).
Neuroplasticity mainly subsumes terms such as neurogenesis, synaptogenesis or dendritic
sprouting, while circulating synonyms for neuroplasticity, e.g., cytoarchitectural
restructuration or neuronal remodeling are a source of confusion.
During the past 20 years, neurobiological findings revealed remarkable new insights into the
underpinning mechanisms regulating neuroplasticity. This led to a reconsideration of the
traditional notion that the brain’s structural configuration is created during development from
early ages until early adulthood and remains stable during adulthood. Most prominently, it
was demonstrated that serotonin (5-hydroxytryptamine, 5-HT) is involved in memory
formation and learning by structural adaptation via second messenger proteins such as cyclic
AMP (Kandel, 2004; Kandel & Schwartz, 1982). Beyond this, dynamic neuronal
restructuration was recently demonstrated after motor learning, for example after learning to
juggle or musical training, but also upon enhanced experience of navigation, foreign
language learning and olfactory processing (Barkas et al, 2012; Delon-Martin et al, 2013;
Draganski et al, 2006; Hyde et al, 2009; Maguire et al, 2000; Martensson et al, 2012). These
studies were performed in humans with structural and functional neuroimaging and raised
criticism from many authors regarding the validity of these results (Fields, 2013; Thomas &
Baker, 2013). Researchers questioned, whether changes in neuroplastic MRI markers such
as gray matter volume (GMV) are caused by methodological shortcomings such as head
movement during scanning, realignment artifacts, and alterations in tissue water content or
regional cerebral blood flow changes other than neuronal activation. However, good
evidence in animal models shows significant neuronal remodeling by well-established
histological methods such as immunohistochemistry (Sagi et al, 2012; Tanti et al, 2012)
The molecular and cellular machinery behind neuroplasticity has been investigated with large
efforts and comprises numerous extracellular and intracellular proteins, signaling cascades,
transcription factors and genes, of which many are directly linked with 5-HT. But until now, it
remains unclear, which factors produce effects large enough to explain structural and
functional changes observed in human neuroimaging studies. To resolve open questions in
this regard, this thesis aims to further illuminate the cross-links between 5-HT and

10
neuroplasticity in humans. This was achieved by manipulation of 5-HT and measuring the
impact on neuroplasticity parameters obtained by functional and structural neuroimaging.
1.2 The role of neuroplasticity in health and disease
Neuroplasticity is a well-known key mechanism in learning and memory (Dayan & Cohen,
2011; Kandel, 2001), but besides altered neurotransmission, dysfunctional neuroplasticity is
a major pathogenetic factor in psychiatric disorders. In major depression two main etiologic
theories are well supported by a series of evidence, one of which is the so-called monoamine
hypothesis suggesting alterations in serotonergic and dopaminergic neurotransmission. The
second main etiologic theory is the “neuroplasticity hypothesis” indicating deficient neuronal
adaptation to external or internal stimuli as pathophysiologic factor. Both theories are
clinically relevant, because the mechanisms of action of current and potential future
treatments for psychiatric disorders are evaluated in this context.
The brain undergoes fundamental structural and functional changes from early ontology until
early adulthood, which is subsumed with the term developmental plasticity. Thereby, a
multiplicity of neurobiological changes drive physiologic transformations with far reaching
consequences for brain functionality and connectivity. During fetal brain development, early
precursor cells transform to neurons and migrate from the ventricular zone to form the
cerebral cortex. Dendrites and axons spread to establish a complex network of intercellular
communication via synapses (Rapoport & Gogtay, 2008). Interestingly, there is an initial
overproduction of neurons, which is followed by selective apoptosis leading to loss of about
50% of cortical neurons. During postnatal brain development, synaptic density increases
further to numbers above adult levels and further pruning happens during childhood and
adolescence to adult levels with primary sensory and motor regions maturing early in
comparison to more complex functions. Moreover, the brain needs stimulation by internal and
external environmental stimuli, similar to light stimulation for the developing eye (Rapoport &
Gogtay, 2008). Today it is known that the brain reaches ≈90% of its adult size around an age
of six, but changes constantly occur during adolescence (Brans et al, 2010). During the aging
process a consistent loss of brain tissue is observable (Good et al, 2001). While many of the
neurobiological mechanisms in control of neuroplastic changes during the brain’s
development seem to reduce their effects in adulthood, it remains unclear, which factors
remain significant and to what extent.
Malfunction of the molecular machinery regulating neuronal plasticity during developmental
plasticity may lead to malformations of cortical development, which are frequently associated
with mental retardation, epilepsy and congenital neurological deficits. The majority of these

11
disorders are now thought to share a genetic basis (Leventer et al, 2010), while many of the
underling mechanisms remain unclear. Furthermore, neurodevelopmental disorders such as
autism, fragile-X-syndrome, Down syndrome, motor disorders or schizophrenia (Lesch &
Waider, 2012) are associated with a distinctive serotonergic deficiency. Here, deficits in 5-HT
mediated synaptic signaling are thought to contribute to the pathophysiology and long-term
outcome of these patients.
In contrast to developmental plasticity the brain is able to adapt to internal or external
stressors or injury by compensatory neuroplasticity. Brain injury after stroke can result in a
series of well-studied events including intra-hemispheric changes in representational maps,
or inter-hemispheric balance shift whereby the uninjured hemisphere gains functions.
Furthermore, regional injury results in diffuse adaptive changes between functional network
nodes (Cramer et al, 2011). Similar forms of adaptive neuroplasticity have been described
during other forms of neuronal damage such as traumatic brain injury. In consideration of
age-related adaptive plasticity, it has been demonstrated that lesions in early age exhibit a
more efficient repair and children are able to handle trauma to brain areas of vision, motor,
auditory and language function considerably better than the adult CNS (Rapoport & Gogtay,
2008). Noticeably, adaptive neuroplasticity might not always have a positive impact on
clinical functioning. Neuronal reorganization after brain injury might lead to enhanced
disinhibition, a dysbalance between excitation and inhibition, even after months of the injury,
suggesting that the delayed onset is in relationship with slow axonal sprouting and the
formation of new neuronal connections. Maladaptive neuroplasticity is thought to cause
chronic pain and allodynia after amputation (Cramer et al, 2011).
1.3 Mechanisms regulating neuroplasticity
Ramon y Cayal and independently Sigmund Freud already postulated in 1894 that learning
might produce lasting changes in the effectiveness of synaptic connections (Kandel, 1981),
an idea that was not testable until decades later. Meanwhile several interdependent
mechanisms were found that regulate neuronal structure and function in neuroplastic
processes.
Neuroplasticity can be divided into large-scale morphological changes, such as axonal or
dendritic (neurite) sprouting or pruning and small-scale changes, such as synaptic formation
or pruning (Holtmaat et al, 2013). Due to methodological constraints, at present little is
known about longitudinal neurite and synaptic turnover in vivo in humans. From animal
studies it became clear that dendrite length and complexity are rather stable at low
magnification (Chow et al, 2009; Holtmaat & Svoboda, 2009; Kasai et al, 2010)

12
(Trachtenberg et al, 2002), while axonal length changes are more dynamic (De Paola et al,
2006). Most of the brain’s excitatory synapses are located on dendritic spines, which are
highly specialized, rapidly changing cellular substructures and often serve as proxies of
synapses. There are large, rather stable mushroom-like and small, dynamically changing
spines. During cortical development spine turnover is highest, but they remain under
homoeostatic control during lifetime, whereas stable spines stay in their form during lifetime
(Holtmaat et al, 2013). Short-term synaptic plasticity is essential to influence information
processing and high-pass or low-pass filtering of a synapse (Citri & Malenka, 2008)
according to initial transmitter release probability. Long-term spine plasticity is underlying
fundamental neurophysiologic processes such as long-term potentiation (LTP), long-term
depression (LTD) and spike-timing dependent plasticity (STDP), a subform of the first two
(Citri & Malenka, 2008).
Synaptic plasticity during aversive learning e.g. in fear conditioning located in brain areas
such as the hippocampus and the amygdala are thought to underlie LTP. Long-term
potentiation is considered an important factor for increasing synaptic strength. Repetitive,
simultaneous activation of excitatory synapses (Citri & Malenka, 2008; Jeffery & Reid, 1997)
induces a fast influx of Ca2+ into the postsynaptic cells. This is controlled by the excitatory
NMDA receptor of the glutamate neurotransmitter system and further modulated by AMPA
and metabotropic glutamate receptors. Long-term potentiation leads to an increased release
of neurotrophic factors, which enhance dendritic and synaptic strength (Malenka & Nicoll,
1999). Key players thereby are neurotrophins such as the brain derived neurotrophic factor
(BDNF) and it’s TrkB receptor.
The neurotrophin family consists of structurally related proteins, the nerve growth factor
(NGF), BDNF, neurotrophin 3 and neurotrophin 4. Each protein specifically binds at the TrkA,
TrkB, TrkC or the p75 receptors with receptor dimerization and structural modifications
enhancing specificity (Chao, 2003). Of all neurotrophins BDNF has gained most attention, as
it is associated with at least three intracellular signaling cascades not only directing synaptic
plasticity, but also cell survival and neuronal differentiation and neurite outgrowth (Black,
1999; Chao, 2003; Gentry et al, 2004). Due to the abundance of it’s functions BDNF, it’s
associated pathways and genes became one of the mostly investigated target structures in
neuropsychiatric research. Dysfunction of BDNF was linked with regional brain atrophy in
Alzheimer’s disease, neurodegenerative disorders as well as in depression [(Castrén, 2005;
Macqueen & Frodl, 2010)]. A polymorphism of the proBDNF’s promotor region consisting of a
valine to methionine substitution (val66met) in the codon 66 was found to cause significant
reductions in extracellular BDNF concentrations (Chen et al, 2004; Egan et al, 2003). This

13
polymorphism serves as a model of reduction of endogenous BDNF levels and has been
investigated in numerous neuropsychiatric studies.
Most of the above listed pathways are extracellular molecules circulating in the synaptic cleft
or membrane proteins. To fulfill the neuronal modifications upon cellular stimulation,
intracellular signaling cascades activate transcription factors that ultimately change protein
expression (McClung & Nestler, 2008). Many of the above mentioned structures lead to a
well-studied common transcription factor named cAMP response element-binding protein
(CREB), which transcriptional activity is fine-tuned by at least 30 other proteins. Major
second messenger proteins transducing receptor signals to CREB are protein kinases such
as the mitogen activated protein-kinase (MAPK). Further prominent transcription factors
regulating neuronal plasticity are located within the Fos family, including cFos, FosB, ΔFosB,
Fos-related antigen 1 (Fra1) and Fra2, which dimerize with Jun proteins (McClung & Nestler,
2008). Additionally, the rapid acting nuclear factor kappa-light-chain-enhancer of activated B
cells (NF-κB), plays an important role in synaptogenesis (Boersma et al, 2011). For LTP and
long-term neuroplastic changes, it was demonstrated that epigenetic modifications, histone
acetylation and DNA methylation, are required (Borrelli et al, 2008; McClung & Nestler,
2008). Post translational modifications e.g. by mRNA binding proteins such as
polyadenylation element-binding protein (CPEB) or mRNA regulation by micro RNAs
(miRNA) are intracellular mediators of neuroplasticity. Finally, it was demonstrated that a cell
cycle and differentiation regulating proteins of the S100 protein family, p11 is associated with
neurotransmitter transport, BDNF, neuroplasticity and 5-HT signaling (Warner-Schmidt et al,
2010).
1.4 Crosslinks between Serotonergic Neurotransmission
and Neuroplasticity
Besides histamine and the catecholamines adrenaline, dopamine and noradrenaline, 5-HT
belongs to the classical monoaminergic neurotransmitters. These are transmitters that are
built by one amino group, are connected to an aromatic ring by a two-carbon-chain (-CH2-
CH2-) and are synthesized form aromatic amino acids like phenylalanine, tyrosine or
tryptophan. The majority of the body’s serotonin is found in the enterochromaffine cells in the
digestive system where it controls gut movements. Furthermore, 5-HT is involved in bone
metabolism, vasoconstriction and exerts control in organ development. In the adult brain, 5-
HT is synthesized in the raphe nuclei of the midbrain and brainstem, from where serotonergic
neurons project to forebrain regions. Hence, the midbrain’s raphe are thought to possess a
major control function over the functionality of the serotonergic system. High 5-HT
concentrations were observed in the parahippocampus and hippocampus, amygdala,

14
cingulate cortex, temporal cortex as well as in the basal ganglia (Hornung, 2010). Hence, in
the brain 5-HT has multiple functions ranging from basal physiologic processes like appetite,
thermoregulation, sleep regulation, to “higher” functions such as emotion regulation,
impulsivity, reward processing (Akimova et al, 2009; Hoflich et al, 2012; Kranz et al, 2010;
Savli et al, 2012). The 5-HT system consists of at least 16 receptors and a serotonin
transporter (SERT) (Saulin et al, 2012).
A close interactive relationship between 5-HT and neuroplasticity is well established (Daubert
& Condron, 2010; Gaspar et al, 2003). Treatment with SSRIs is associated with changes in
the expression of BDNF (Koponen et al, 2005; Nestler et al, 2002). Increase of BDNF mRNA
in hippocampus and cortical brain regions have been reported following acute and chronic
administration of SSRIs (Kozisek et al, 2008; Nibuya et al, 1995). Research by E. Castren’s
group demonstrated that antidepressants activate BDNF mediated TrKB signaling and
subsequently CREB (Koponen et al, 2005; Rantamaki et al, 2007). Results from animal
(Karpova et al, 2011; Piubelli et al, 2011; Vetencourt et al, 2008; Vetencourt et al, 2011) or
human subjects (Nitsche et al, 2009; Serra-Millàs et al, 2011) conclusively suggest
enhancement of neuronal plasticity as a result of treatment with SSRIs. Furthermore, SSRIs
were demonstrated to improve motor recovery from ischemic stroke, which might arise from
these mechanisms (Chollet et al, 2011; Mead et al, 2012). Chronic administration of the SSRI
fluoxetine reinstates ocular dominance plasticity in adulthood and promotes the recovery of
visual functions in adult amblyopic animals (Vetencourt et al, 2008). These effects were
accompanied by increased expression of BDNF in the visual cortex (Vetencourt et al, 2011).
Serotonin and other monoamines are one the first neurotransmitters to emerge during
neuronal development (Rubenstein, 1998), where they first mediate autoregulatory effects in
growing serotonergic neurons (Whitaker-Azmitia, 1998), then catalyze the maturation of
astroglial cells (Whitaker-Azmitia, 1998) and finally influence target tissue maturation
(Whitaker-Azmitia et al, 1996). While transgenic mice completely lacking serotonergic
neurons exhibit high perinatal mortality rates and severe deficits in respiratory control
(Hodges et al, 2009), reversible inhibition of serotonin synthesis during early embryogenesis
(embryonic days (E) 12-17) results in long lasting alterations of cortical development (Vitalis
et al, 2007). Excess serotonin produces dystrophic serotonergic neurons (Daubert et al,
2010) as well as migration defects in retinal projection neurons (Upton et al, 1999),
thalamocortical axons (Vitalis et al, 2002) and cortical interneurons (Riccio et al, 2009).
Dystrophic serotonergic neurons were also reported in several neurodegenerative diseases
(Azmitia & Nixon, 2008) and serotonergic dysfunction is a characteristic of down syndrome
and autism (Whitaker-Azmitia, 2001).

15
Further investigations into the underlying molecular mechanisms revealed convergent
signaling pathways between serotonin and neuronal growth factors (Cowen, 2007; Polter &
Li, 2010). This evidence suggests that some serotonergic receptors, beyond their traditional
association with G proteins (all 5-HT receptors but the following) or ligand-gated ion channels
(5-HT3 receptor), are able to modulate the activity of signaling pathways involved in neuronal
plasticity such as extracellular-regulated kinase (ERK) and mitogen-activated protein kinase
(MAPK) (Cowen, 2007). Although knocking out SERT or MAO and genetic polymorphisms in
these enzymes impact on neuronal structure (Frodl et al, 2008; Karabeg et al, 2013; Singh et
al, 2013) effects seem to be less pronounced than manipulation of serotonergic receptors
(Benninghoff et al, 2012). The 5-HT1A, 5-HT1B, 5-HT1D, 5-HT2A, 5-HT2B, 5-HT2C, 5-HT4 and the
5-HT7 receptors are associated with ERK or Akt (proteinkinase B) in neuronal cells, which
indicates that intracellular serotonergic signals are involved in long-term cell protective
processes. The mechamism behind 5-HT’s neuroplastic properties are well examplified by
the 5-HT1A receptor. Starting from embryonic day (E) 12 in mouse embryos the 5-HT1A
receptor contributes to craniofacial development (Moiseiwitsch & Lauder, 1995) and peaks
from E14.5—E16.5 in the thalamus, hippocampus and the cortex (Bonnin et al, 2006). There
might be a time dependent expression pattern with peak expressions e.g. in the amygdala
throughout the developmental period and further peaks in regions maturing in postnatal
development (Bonnin et al, 2006; Mehta et al, 2007). Important neuromodulatory actions of
the 5-HT1A receptor are stimulation of neurogenesis and dentritic maturation in the
hippocampus (Yan et al, 1997) where neurogenesis in the dentate gyrus and the
subventricular zone remains life-long active (Gaspar et al, 2003). For example, treating mice
with a 5-HT1A agonist can reverse microencephaly induced by prenatal treatment with
cocaine (Akbari et al, 1994) and can reduce neuronal damage after ischemic stroke
(Mattson, 2008). Furthermore, 5-HT1A receptors on astroglial cells release a neurite
extension factor (S-100ß) and induce maturation of astrocytes (Azmitia, 2001). Astrocytic 5-
HT1A receptors in combination with S-100ß are responsible for maintenance of a mature
state in adult neurons (Azmitia, 1999; Wilson et al, 1998). Withdrawal of S-100ß leads to a
reduction of synaptic connections between neurons (Wilson et al, 1998) and goes along with
findings, that the 5-HT1A receptor is required for behavioral and neurogenic effects of the
selective serotonin reuptake inhibitor fluoxetine (Santarelli et al, 2003).
In summary, 5-HT is a neurotransmitter that exerts distinct neuromodulatory actions beyond
neurophysiological functions. Highly active in shaping the architecture of serotonergic
neurons during embryonic development and early postnatal neuronal maturation, this
neuroplastic role is partially conserved in specific brain regions throughout adulthood. Many
of these effects are mediated by the 5-HT1A receptor, through direct links to neuromodulatory

16
signaling pathways such as ERK and MAPK, neuronal cell maintenance by astrocytes or
linkages to neurotrophins such as BDNF.
1.5 In Vivo Quantification of Brain Anatomy with MRI -
Principles of Structural Magnetic Resonance Imaging
Magnetic resonance imaging (MRI) is a major clinical application of the nuclear magnetic
resonance phenomenon, which was discovered in 1939 by Isidor Rabi who described nuclei
in an electromagnetic field absorbing and re-emitting electromagnetic radiation. Thereby
every atomic isotope can be characterized by a specific resonance frequency depending on
the magnetic field strength. NMR allows identification of isotopes with an odd number of
protons and/or of neutrons by their unique magnetic moment and angular momentum or in
other words by a given, nonzero nuclear spin, whereas nuclides with even numbers of both
have a zero spin. Simply put, MRI is based on the interaction of a nuclear spin with an
external magnetic field (B0) (Haacke, 1999).
In more detail, MRI in humans relies on the manipulation of magnetic fields and the detection
of bulk precession of hydrogen spins in water, fat and organic tissue. While theoretically a
series of nuclei are detectable (e.g.: 2H, 6Li, 10B, 11B, 14N, 15N, 17O, 19F, 23Na, 29Si, 31P, 35Cl,
113Cd, 129Xe, 195Pt), mostly protons of 1H and 11C are used for MRI given their abundance in
organic material. Precession is defined as the change in the orientation of the rotational axis
of a rotating body. Nuclear spins are unique characteristics describing the circular movement
of a proton whereby it exhibits a loop of electric current around the axis about which it is
spinning (Haacke, 1999; Schneider & Fink, 2013). The electromagnetic current loop may
interact with external magnetic fields and is described by the magnetic dipole moment vector
𝜇 representing the spin axis and realigning, similar to a compass needle, along the force of
any external magnetic field, �⃗⃗�0. The process is influenced by the gyromagnetic ratio γ, which
is a constant unique to each particle given by the ratio of the magnetic dipole moment to its
angular momentum. E.g. the gyromagnetic ratio of a 1H proton has a value of 2.68×108
rad/s/Tesla (so that 𝛾 =γ
2π= 42.58 MHz/T). For a 7T magnetic field, the spins precess at a
radiofrequency of 7 × 42.58 = 289.06 MHz. This precession frequency is referred to as the
Larmor frequency and the relationship is expressed in the Larmor equation:
(1) ω0 = γ0B0
where ω0 is the Larmor frequency in megahertz (MHz), γ0 is the gyromagnetic ratio specific
to a particular nucleus and B0 is the strength of the magnetic field in Telsa (T).

17
To obtain a classical macroscopic MR-image, precession has to be generated by tipping the
magnetization vector, away from an external field. The magnetization is rotated away from its
longitudinal direction alignment with a radiofrequency (rf) pulse for a short time. The rf pulse
is a electromagnetic wave of the same frequency as the target nuclei’s’ Larmor frequency,
generated by a transmit coil. This process of energy absorption is known as excitation and
leads to longitudinal magnetization of the spin system. A pulse strong and long enough is
required to tip magnetization by exactly 90° (90° rf pulse) into a transverse plane. Nuclei are
hence rotated away from the z-axis towards the transverse plane (x,y-axis) perpendicular to
the direction of the main magnetic field, which produces an electrical current at the same
frequency as the Larmor frequency in a receiver coil. After the spin system is excited, the
magnetization vector starts to return to the state before excitation due to two independent
phenomena reducing transverse magnetization called spin-lattice interaction and spin-spin
interaction, which cause T1 relaxation and T2 relaxation. Longitudinal relaxation or T1
recovery describes the slow restoration of the spin vector to the z-axis, whereby the nuclei
emit their excess energy to their environment (i.e. lattice, hence spin-lattice). Transverse
relaxation subsumes an energy transfer between spins resulting from local changes in the
magnetic field. Spin-spin interactions are a consequence of differential durations of
dephasing between spins formerly in so-called phase coherence, which describes spins that
precess synchronously about the z-axis after excitation (T2 relaxation occurs at about 100-
300ms, T1 relaxation at about 0.5-5s, yet both processes are thought to occur independently)
(Weishaupt et al, 2006). Tissue contrast of brain MR-images depends on differential tissue
proton densities, whereby the CSF and fat exhibit highest densities followed by pathologies
such as a meningioma and gray matter. For delineation of T1-weighted tissue proton
densities, repetition time (TR) between iterative excitations is essential to MRI contrast, as
the number of newly excitable spins increases if more spins are already rotated back into the
z-plane. A long enough (over 1.5 s) TR ensures that differential relaxation times according to
tissue property are depicted. Tissue contrast in T2-weighted images is set by the echo time
(TE) of T2-weighted images, which is defined by the interval between deliverance of the
excitation pulse and collection of the MRI signal and in the range of only several hundred ms.
The digitized data are stored in a graphic matrix called k-space and afterwards Fourier
transformed. Signal-to-noise-ratio in MRI depends on multiple factors amongst them slice
thickness, field of view, number of acquisitions, scan parameters (TR, TE, flip angle), the
magnetic field strength or the selection of the transmit and receive coil (RF coil). For
structural MRI capable of in-depth neuroanatomical brain investigations mostly T1-weighted
gradient echo sequences (GRE) are applied, given their advantages in short imaging times
and hence less motion artifacts. Thereby, instead of pairs of RF pulses, GRE uses frequency
encoded gradients administered by gradient coils with alternating polarities. In functional MRI

18
(fMRI) typically T2*-weighted images are recorded, which are typically shorter than T2 and
obtain slightly more signal inhomogeneities at tissue borders or local magnetic fields (e.g. by
a high concentration of iron in several brain areas such as nucleus ruber). fMRI is based on
the effect that deoxygenated hemoglobin (Hb) exhibits shorter dephasing times than
oxygenated Hb. This blood oxygen level dependent (BOLD) contrast allows inference on
neuronal activity by a complex relationship expressed in the hemodynamic response
function.
1.6 MRI quantification of Neuroplasticity: Voxel-based
morphometry
Until recently neuroplasticity in humans could only be investigated by histologic samples
either obtained post mortem or during brain surgery. With improvements in neuroimaging
technologies beginning with computerized tomography scans demonstrating enlarged
ventricular size in schizophrenic patients (Weinberger et al, 1980; Weinberger et al, 1979),
early MRI-based manual brain volume measurements (Mathew & Partain, 1985) to
computational morphometry (Ashburner & Friston, 2000; Wright et al, 1995), in vivo brain
plasticity research has gained successively increased attention. High resolution or ultra-high
resolution structural MRI at field strengths of 3 respectively 7 Tesla and computerized
volumetric analyses are the current state-of-the-art techniques to quantify brain
microstructure at a minimal resolution around one mm3 (Lenglet et al, 2012; Lusebrink et al,
2013). This is achieved by a computational analysis of structural MRI data termed voxel-
based morphometry (VBM) (Ashburner & Friston, 2000) (a voxel is the cubic 3D analogue to
a pixel).
Voxel-based morphometry is a whole-brain automated technique for measuring regional
cerebral volume and tissue concentration differences in MRI. Simply put, it is the comparison
of local concentrations or volumes of gray matter between groups or within subjects
(Ashburner & Friston, 2000; Ashburner & Friston, 2009; Good et al, 2001). In more detail
VBM works with so-called mesoscopic anatomical differences in brain tissue density and
volume after discounting macroscopic differences, which are modeled after warping
individual brains to a common reference space (Ashburner & Friston, 2009). Although
criticized by some authors as susceptible to bias introduced by averaging individual brain
shapes to standardized brain templates (Bookstein, 2001) or lack of validity, because factors
such as head movement or cerebral blood flow may confound the MRI signal (Franklin et al,
2013), structural MRI exhibits an excellent test-retest reliability (Wonderlick et al, 2009) and
gray matter alterations observed with MRI were confirmed by post-mortem histologic analysis
(Vernon et al, 2011). Furthermore, VBM offers the advantage of fast calculations of regional

19
brain volumes, independent of anatomical delineation by manual region of interest (ROI)
drawing, which improves comparability of studies between study centers and improved
access to morphometric analysis, because detailed neuroanatomical and radiologic
knowledge is not a prerequisite.
Technically VBM is mostly performed with T1-weighted structural MRI images recorded with
inversion recovery gradient echo (GE) pulse sequences such as magnetization-prepared
rapid acquisition of GE (MPRAGE) (Mugler & Brookeman, 1990) or its advancement
MP2RAGE, which is thought to be more robust against field inhomogeneities at high static
magnetic fields (≥ 3 T) (Marques et al, 2010). Gradient echo images are designed to provide
the best contrast between gray and white matter with short acquisition times of only several
minutes. A good contrast between brain tissues is highly necessary to pre-process MRI data
in a standard VBM pipeline, which comprises segmentation in gray matter, white matter and
cerebrospinal fluid (CFS) images, registration or normalization of each individual’s brain
image to a standard stereotactic space, smoothing with an isotropic Gaussian kernel and
statistical analysis. The entire procedure is embedded in standard neuroimaging software
bundles implementing the general linear model such as statistical parametric mapping
(SPM). One of the currently available most accurate image segmentation and registration
algorithms is named DARTEL (“Diffeomorphic Anatomical Registration using Exponentiated
Lie algebra”) having significant advantages in tissue segmentation and normalization, due to
an increased number of realignment parameters (Ashburner, 2007; Klein et al, 2009). The
DARTEL-algorithm applies “flow fields” for each subject, which encode how the individual
images are warped or deformed to match best the average shape of a study specific,
iteratively improved template, and MRI data are so segmented into gray matter, white matter
and CSF images (Ashburner, 2009). In a further spatial transformation gray matter data,
which are mostly used for plasticity research, are matched to the canonical MNI (Montreal
Neurological Institute) brain, which is an internationally accepted standard template with a
coordinate system generated by averaging 152 subjects. To preserve the volume of tissue
from each voxel, gray matter data are modulated by Jacobian determinants encoding the
relative volumes of tissue before and after warping. This results in the main parameters in
neuroplasticity neuroimaging: GMV, a surrogate of the volume of gray matter in arbitrary units
and gray matter density (GMD) representing the density of tissue in a voxel. Both units are
usually smoothed by a Gaussian kernel between 8-12 mm to ensure normal distribution for
statistical processing. Now the ready pre-processed VBM data can fit a general linear model
(GLM) as implemented in SPM. Statistic analyses, regression models or longitudinal
analyses of variances (ANOVA) can be calculated with gray matter microstructure data at
around 1-3.5 mm3 resolution. This is the currently gold standard for in vivo assessment of
dynamic brain changes attributed to neuroplasticity.

20
1.7 In Vivo Quantification of Neuroplasticity in the
Serotonergic System with PET
Neurotransmitter release in the brain can be indirectly measured by a combination of
neuroreceptor quantification with radiotracers and positron emission tomography (PET) and
pharmacological challenges (Laruelle, 2000). The principles behind this approach are the
detection of changes in the availability of target receptors to radiotracer occupancy under
baseline and challenge conditions (Paterson et al, 2010). While this approach yielded highly
valuable results for the dopamine system (Laruelle, 2000) this did not yet translate to 5-HT
because only few currently available PET tracers (Selvaraj et al, 2012) were found to be
sensitive for 5-HT change, and these are still less potent as compared to measuring
endogenous dopamine release with [11C]raclopride or [11C]-(+)PHNO (Paterson et al, 2010).
Nevertheless, many important components of the 5-HT system in humans can be reliably
measured with PET and radioligands (Saulin et al, 2012).
A radioligand is composed by a radioactive isotope and a molecule binding selectively to a
specific target. The standard radioactive isotopes in use are 18Flour, 11Carbon 13Nitrogen or
15Oxygen with half-lives ranging from 2 minutes (15O) to almost 2 hours (18F). These emit a
positron, which reacts with an electron after travelling the positron range of about 1 mm. In
the process of positron annihilation two photons emerge exhibiting a charge of 511keV
spreading in opposite directions. The PET detector ring detects the two photons
simultaneously (coincidence), and stores the coincidence events as sinogram, from which
PET raw images are reconstructed (Turkheimer et al, 2014). The radioligand’s biochemical
properties determine the target structures at which the isotope should emit it’s signal. While
there are reversible and irreversible radioligands, in neuroreceptor and transporter mapping,
reversible tracer binding is preferred to mathematically model receptor binding (Laruelle et al,
2003). The chemical properties of the biological tracer to bind on brain structures must meet
several, occasionally contradictory demands making radioligand synthesis a quite
complicated task. First, it must be non-toxic, then it must be intravenously injectable in a
stable solution, exhibit a low peripheral metabolism, pass the blood-brain barrier and must
finally bind to the target structure with high selectivity and specificity. To yield proper binding,
the ligand should exhibit a sufficiently high affinity to the target protein depending on the
concentration of sites in the target region, which lies usually between 0.01 and 1 nM. The
lower the target density, the higher the required affinity (Innis et al, 2007; Laruelle et al,
2003), however a too high affinity will prolong scanning time, which impacts on decay
counting statistics for short-lived isotopes and patient-comfort. To pass the blood-brain barrier
lipophilicity is necessary, but a too high lipophilicity increases unspecific binding and
decreases the amount the free radioligand in the blood plasma. As far as metabolism

21
characteristics are concerned, no active metabolites crossing the blood-brain barrier are a
prerequisite. This multitude of necessary characteristics to synthetize a suitable radioligand
is the obvious reason why it is still a demanding task for radiochemists worldwide to provide
tailor made PET tracers.
During the PET neuroreceptor measurements, which vary dependent on the applied
radioligand between several minutes and about 2 hours, the subject’s head is placed parallel
to the orbitomeatal line guided by a laser beam in a polyurethane cushion. A standard PET
ring has a diameter of about 90 cm, where subjects are placed only with the head inside the
bore, hence claustrophobia is seldom an issue. Subjects are instructed not to talk, apart from
emergencies, and to minimize head and body movements. Subjects are intravenously
injected about 5 ml of the radioligand (using the bolus paradigm in most cases) in a stable
saline solution at the beginning of the PET measurement.
1.8 PET radioligands for the Serotonin-1A receptor and
Serotonin Transporter
For in vivo neuroplasticity research, the 5-HT1A receptor and the SERT are relevant
serotonergic structures, which both control neuroplastic processes and can be quantified with
excellent test-retest variability with PET radioligands. Several radioligands are commonly
used for PET and the 5-HT1A receptor: [3H]-WAY100635 (post-mortem), [carbonyl–11C]-
WAY100635, [18F]-FCWAY, [18F]-FPWAY, [18F]-MPPF, [11C]-CUMI-101 and various
derivatives (Billard et al, 2014), each one exhibiting advantages and disadvantages. One of
the most reliable compounds is [carbonyl–11C]-WAY100635, having excellent affinity,
specificity and selectivity (Martel et al, 2007) and was therefore selected for this work. It is
well synthesizable with simplified synthesis techniques (Rami-Mark et al, 2013; Wadsak W.,
2007). There are no relevant radioactive metabolites passing the blood-brain barrier which
leads to an excellent signal-to-noise-ratio (Pike, 2009; Wu et al, 2007).
Quantitative molecular imaging of the SERT was primarily focused on modification of SSRIs
based on their high affinity and specificity. This approach turned out to be not fruitful,
because in vitro measures proofed not to be translatable to in vivo performances (Huang et
al, 2010). In the following [128I]β-CIT was used in a number of SPECT-Studies, yet high non-
specific binding at the dopamine transporter (DAT) resulted in limited validity. Further
developments yielded radioligands with better specificity namely [11C]MADAM,
[11C]HOMADAM, [11C]DASB and [11C]AFM (Huang et al, 2010). Obtaining a high specific
binding, reversible high brain uptake and equilibrium within a short scanning time, [11C]DASB
emerged as most widely used PET ligand in studying the human SERT (Houle et al, 2000)

22
and was available for this work. Meanwhile a rapid automated preparation and purification of
[11C]DASB became available (Haeusler et al, 2009).
1.9 PET quantification: binding potential
To determine estimates of in vivo distribution of PET target proteins, PET raw data are
quantitatively analyzed by applying mathematical models. One of the most commonly used
outcome parameter is termed binding potential (BP) which is defined by the ratio of receptor
availability Bmax to radioligand dissociation constant KD at equilibrium, while KD is the
reciprocal of the affinity. The binding BP potential can be also viewed as the product of Bmax
and affinity (Innis et al, 2007):
(2) 𝐵𝑃 =𝐵max
𝐾D= 𝐵max
1
𝐾D= 𝐵max𝑎𝑓𝑓𝑖𝑛𝑖𝑡𝑦
The BP concept was originally defined by Mintun et al. (Mintun et al, 1984) and originates
from the Michaelis-Menten equation (Michaelis L., 1913),
(3) 𝐵 =𝐵max[𝑆]
𝐾D+[𝑆]
and was quickly incorporated in quantitative radioligand imaging analyses. B represents the
concentration of receptor bound ligand, Bmax the availability of receptors, S the concentration
of free substrate and KD is the dissociation constant. Here KD is equal to the substrate
concentration at which the reaction rate is half its maximal value (Berg et al, 2002). The
maximal rate Bmax is reached when catalytic sites on the enzyme are fully saturated with
substrate [S]. At very low substrate concentration [S] is much less than KD, yielding a rate
that is directly proportional to the substrate concentration:
(4) 𝐵 = 𝐵max
KD[𝑆]
In situations during radioligand imaging when [S] is very low, [S]≪KD, the Michaelis-Menten
equation (2) reduces to Mintun’s original definition of the binding potential (Bmax/KD) (1)
(Mintun et al, 1984) and equals the equilibrium ratio of specifically bound B ligand to its free
concentration [S].
(5) 𝐵𝑃 =𝐵max
𝐾D=
𝐵
[𝑆]

23
Of note, in vitro radioligand studies mostly use homogenized tissue where all receptors are
available to bind. Thereby, only one compartment is assumed and no distinction is made
between free concentrations in plasma or tissue. In contrast, proper modeling of in vivo
radioligand imaging in the brain demands two or three tissue compartment models with
additional compartments for non-specific binding. Additional compartments include the blood
plasma, and three compartments in the brain: a compartment of freely available ligand, a
non-specific and a specific bound compartment (Figure 1). In this model, the radioligand
binds to specific (the target structure) or non-specific targets (non wanted targets such as
other binding structures) in the brain after crossing the blood-brain barrier. The radioligand’s
free and the non-specific bound compartments are often combined, based on the notion that
the exchange between free and non-specific compartments is faster than between free and
specific compartments. This leads to a two-tissue model in the brain, instead of a three-
tissue compartment model, reduces model input and minimizes computation time. The
Figure 1: Compartment model assumptions commonly used for radioligand modeling. The ligand
enters the brain via the blood brain barrier. Here it is either freely available within the tissue,
specifically bound or non-specifically bound to target proteins. A three tissue compartment model is
reduced to a two-tissue compartment model by combining free and non-specific bound
compartments to a non-displaceable compartment model. The rate constants (k1-k6) describe
amount of ligand and time needed for the radioligand to transfer the compartments. Figure adapted
from (Slifstein et al, 2000).

24
assumptions contain a blood-brain coefficient of clearance into the brain (K1), a rate constant
of fractional clearance (k2) from the exchangeable pool of unbound radioligand in the brain
back to the systemic circulation and, most importantly, an association and a dissociation rate
constant towards and away from free ligand to specifically bound radioligands (k3, k4) and
two rate constants towards and away non-specific binding (k5, k6). To model BP either
volumes of distributions as in clinical pharmacology or rate constants as shown in Figure 1
can be estimated. In pharmacology the volume of distribution (V) reflects the relationship
between the amount of drug in the body at steady state and plasma drug concentration
(Hacker et al, 2009). In radioligand imaging V is the ratio of the concentration of radioligand
in a region of tissue to that in plasma (Innis et al, 2007). Depending on the kind of applied
rate constants (K1-k6) and concentrations in different volumes of distribution three BP models
are mostly used.
(6) 𝐵𝑃F = 𝐵avail 𝐾D⁄ = (𝑉T − 𝑉ND) 𝑓P⁄ =𝐾1𝑘3
𝑓p𝑘2𝑘4
BPF refers to the ratio at equilibrium of concentration of the specifically bound radioligand in
tissue to the concentration of free radioligand in plasma. VT and VND represent the volumes of
distribution for total radioligand in tissue and that of nondisplaceable tissue uptake. The free
fraction of plasma protein-bound radioligand 𝑓P needs to be measured by arterial blood
sampling, which itself underlies hardships (Parsey et al, 2000) such as arterial cannulation,
radioactive blood handling and quick high pressure liquid chromatography (HPLC). The in
vivo BPF is the most similar metric to the in vitro measurements of the relation of unoccupied
available receptors (Bavail) and KD, and many researchers argue that this reflects the most
accurate estimate of receptor distribution. But conditions of in vivo radioligand measurements
differ from in vitro measurements as far as temperature, multiple compartments, receptor
trafficking, phosphorylation state and competition with endogenous neurotransmitter are
concerned (Innis et al, 2007).
(7) 𝐵𝑃P = 𝑓P𝐵avail 𝐾D⁄ = 𝑉T − 𝑉ND =𝐾1𝑘3
𝑘2𝑘4
Another version of BP is BPP, which is similar to BPF but not corrected for 𝑓P, which provides
advantages if 𝑓P cannot be measured accurately or has a small difference between groups.
BPP is the ratio at equilibrium of specifically bound radioligand to that of total parent
radioligand in plasma. It equals VT - VND, yet without relation to 𝑓P, as well as the in vitro
analogue of the ratio between Bavail and KD corrected for 𝑓P . Both BPF and BPP can be
pronounced in mL∙cm-3.
(8) 𝐵𝑃ND = 𝑓ND𝐵avail 𝐾D⁄ = (𝑉T − 𝑉ND) 𝑉ND⁄ =𝑘3
𝑘4

25
Finally, the BPND refers to the ratio at equilibrium of specifically bound radioligand to that of
nondisplaceable radioligand in tissue. It uses a reference region with no target proteins for
non-specific binding, is methodically easier to implement and is computed in reference tissue
models (Gunn et al, 2001; Ichise et al, 2001; Logan et al, 1996). Hence, BPND is calculated
by brain data only and does not require the arterial input function, but it depends on the
assumption that nondisplaceable uptake is not influenced by between-group factors. For a
more in-depth mathematical discussion and consensus nomenclature the reader is referred
to an excellent review by Innis et al. (Innis et al, 2007).
1.10 Neuroplasticity quantified with PET and MRI –
previous results
In vivo studies applying structural MRI and VBM detected a wide array of internal and
external stimuli causing the brain to dynamically adapt itself, amongst them physiological
changes (Protopopescu et al, 2008), motoric training (Draganski et al, 2004; Draganski &
May, 2008; May, 2011; Pereira et al, 2007; Zatorre et al, 2012), playing instruments (Schlaug
et al, 2005) and pharmacological (Tardito et al, 2006; Vetencourt et al, 2008) and electro-
physiological interventions (May et al, 2006). Moreover, these techniques identified gray matter
changes intrinsic to a variety of brain pathologies. Beyond diseases where cerebral atrophy
is a known pathognomonic feature such as Alzheimer’s disease and other forms of dementia,
Huntington’s disease and other genetic disorders building-up toxic amounts of proteins in
neurons, inflammatory diseases such as multiple sclerosis or epilepsy, MRI and VBM
identified alterations in regional brain volumes in a number of psychiatric disorders such as
schizophrenia, depression, anxiety disorders, anorexia nervosa (Asami et al, 2012;
Johansen-Berg, 2012; van Tol et al, 2010). For example, patients with depression exhibit
smaller volumes of the basal ganglia, thalamus, hippocampus, frontal lobe, orbitofrontal
cortex and gyrus rectus while a smaller hippocampal volume is detectable in patients during
a depressive episode in comparison to patients during remission (Kempton et al, 2011).
Most importantly, to this work, the neurobiological correlates of changes in brain morphology
measured by MRI and VBM are completely unclear (Scholz et al, 2009; Tost et al, 2010).
Currently, there is a lack of exact models demonstrating which cellular compounds
correspond to the signal strength in one voxel to what extent. Many of these effects could be
mediated by serotonergic structures such as the 5-HT1A receptor, the SERT, neurotrophins
such as BDNF or reciprocal interaction with other neurotransmitters and neurotrophins.
There are a number of unknown variables, for example to what percentage brain vasculature
or cerebrospinal fluid contributes to the gray matter changes observed in VBM. Furthermore,
changes in proton density might alter proton-based MRI analyses. However, neuronal cell

26
bodies and glial cells are assumed to contribute mostly to gray matter visible on T1-weighted
MRI scans. Even at ultra high-resolution MRI and voxel edge lengths below 1 mm, there are
still ten thousands of interconnected neurons packed in one single voxel. Thus, more data
providing information at a molecular level and subsequently linking these with macroscopic
brain changes are necessary to determine what molecular processes gain large enough
effects to be detectable with structural MRI.
In an attempt to combine molecular with structural neuroimaging previous studies mainly
focused on interactions between brain glucose consumption and cerebral atrophy in
Alzheimer’s disease. Hereby, usually PET with the radioligand 18FDG (2-Fluor-2-desoxy-D-
glucose) and VBM are combined, to reveal cerebral hypometabolism and it’s association with
GMV alterations (Chételat et al, 2008; Ishii et al, 2005; Kanda et al, 2008). These studies
identified hypometabolism exceeding atrophy in many altered brain regions in Alzheimer’s
dementia, confirmed frontal and temporal lobe anomalies in frontotemporal dementia. A
similar approach demonstrated that anterior hippocampal formation volume and the posterior
cingulate glucose metabolism are at least altered in normal aging (Kalpouzos et al, 2009).
Several studies investigated associations between brain amyloid content as measured by
PET and the radioligand 11C-PiB (carbon-11-Pittsburgh compound B) and cerebral atrophy
(Jack et al, 2008; Oh et al, 2010; Villemagne et al, 2013). Furthermore, the relationship
between cerebral morphology and dopamine D2/D3 receptor distribution was shown by PET
and [18F]fallypride (Woodward et al, 2009). This body of evidence does not provide hints
towards molecular mechanisms behind dynamic gray matter alterations in healthy subjects or
gray matter atrophy in abovementioned psychiatric disorders. In that regard, two studies
aimed to closer investigate the relationship of molecular makers of neuronal density, GABAA
receptors labeled with 18F-flumazenil, and neuronal density as measured with MRI (Duncan
et al, 2013; la Fougère et al, 2010). The results did not find a linear correlation but rather
indicate a differential relationship between cortex thickness and cortical surface thickness
and neuronal density.
In two pioneering reviews Johansen-Berg and colleagues recently outlined basically four
potential biological mechanisms underlying dynamic gray matter alterations: neurogenesis,
gliogenesis, synaptogenesis and vascular changes (Johansen-Berg, 2012; Zatorre et al,
2012). As potential underlying molecular mechanisms the authors only refer to BDNF related
signaling. The authors finally argue that structural plasticity as measured with MRI should be
given a place in assessment of functional brain changes in learning and recovery and that
more research is urgently needed to identify molecular mechanisms leading to structural
rearrangement of the brain.

27
1.11 Open questions
While the previous sections could not detail all aspects of the entanglement of 5-HT in
neuroplasticity due to limited space, the introducing section provides a solid groundwork for
the work of this thesis. Serotonin beyond physiological functions as regulator of appetite,
thermal control, emotions, impulsivity and reward – just to name a few – is highly involved in
early embryonic and postnatal cytoarchitectural organization of the central nervous system.
The 5-HT1A, 5-HT1B, 5-HT1D, 5-HT2A, 5-HT2B, 5-HT2C, and 5-HT4 receptors are tightly linked to
signaling cascades involved in neuronal restructuration such as ERK, MAPK, and to
neurotrophic systems such as BDNF and their transcriptions factors (e.g., CREB) (Azmitia &
Nixon, 2008; Cowen, 2007; Polter & Li, 2010). An elaborated regulation of 5-HT in
neuroplasticity is further supported by animal models exhibiting dystrophic neurons upon
excessively elevating 5-HT (Daubert et al, 2010; Homberg, 2012). Additionally, treatment with
SSRIs, the most commonly described antidepressants, elevates 5-HT and subsequently
increases dendritic spine numbers (Hajszan et al, 2005), promotes neurogenesis (Mahar et
al, 2014; Pilar-Cuellar et al, 2013), interacts with synapse formation (Getz et al, 2011) and
enhances BDNF signaling (Pittenger & Duman, 2008; Rantamaki et al, 2007; Vetencourt et
al, 2011). The close interplay of 5-HT and neuroplasticity is further demonstrated by
pronounced serotonergic deficits in neurodevelopmental disorders like autism, fragile-X-
syndrome, Down syndrome, motor disorders or schizophrenia (Lesch & Waider, 2012). While
most findings arose from animal models, mostly as a consequence of limited invasiveness
into the human brain in vivo, only little is known about the mechanisms of neuroplasticity in
humans.
Meanwhile a series of studies applying structural magnetic resonance imaging and VBM
demonstrated dynamic in vivo gray matter alterations in adult brains. This was observed due
to a broad variety of internal and external stimuli such as navigation (Maguire et al, 2006),
language learning (Dorsaint-Pierre et al, 2006), musical expertise (Gaser & Schlaug, 2003),
rehabilitation (Sarkamo et al, 2014) but as well in neurological and psychiatric brain
conditions like multiple sclerosis (Eshaghi et al, 2014), depression or anxiety disorders (van
Tol et al, 2010). Explanatory mechanisms for these structural brain alterations are unclear.
Many of the neuroplastic properties of 5-HT might be conserved throughout adulthood and,
linked with in vivo neuroimaging experiments, provide testable hypothesis on the
mechanisms underlying dynamic gray matter changes in the adult brain. Although
information from structural MRI is frequently used for anatomical co-registration
(Henningsson et al, 2009; Lan et al, 2014) or bias-correction (Greve et al, 2014; Matuskey et
al, 2012) of PET data, until now, no study investigated the relationship between the
distribution of serotonergic receptors and the regional volume of gray matter. Additionally,

28
while there are numerous studies investigating the effects of elevated 5-HT related to SSRIs
on brain functionality as investigated with fMRI, there is a lack of data investigating the
influence of 5-HT challenge on gray matter. Finally, the link between 5-HT and BDNF is well
established in preclinical models, but rarely investigated in humans.

29
II. AIMS of the THESIS
Based on the open questions three main aims of this thesis were generated:
The first aim was to detail associations between a serotonergic receptor with neurotrophic
properties such as the 5-HT1A receptor as measured by PET with
[carbonyl−11C]WAY−100635 and regional volumes of gray matter as measured by structural
MRI. This question is treated in the first publication listed below.
A second aim of this thesis was to investigate dynamic alterations of gray matter after 5-HT
challenge with SSRIs in healthy adult subjects, which is the objective of the second
publication.
Finally, the third target was to investigate in more detail the relation between BDNF and
neurotrophic structures of the serotonergic system such as the 5-HT1A receptor and the
SERT. This was the subject of the third publication. The aims can be pointed out in detail as
follows:
To investigate the relationship between 5-HT1A heteroreceptor distribution in the
human brain measured with PET and the radioligand [carbonyl−11C]WAY−100635 and
the regional volume of gray matter measured with MRI and VBM.
To test the association of 5-HT1A autoreceptor binding in the dorsal raphe nuclei and
whole brain regional GMV.
To test the influence of elevated 5-HT as consequence of treatment with a widely
used SSRI (escitalopram) on gray matter.
To elucidate functional neuronal network changes after 5-HT challenge by
escitalopram with fMRI.
To test the impact of a functional polymorphism of BDNF and Val66Met on 5-HT1A
receptor distribution in healthy subjects and SERT binding in patients with major
depression.

30
III. RESULTS:
3.1 First publication: Serotonin-1A receptor binding is
positively associated with gray matter volume – A
multimodal neuroimaging study combining PET and
structural MRI
Christoph Krausa, Andreas Hahna, Markus Savlia, Georg S. Kranza, Pia Baldingera,
Anna Höflicha, Christoph Spindeleggera, Johanna Ungersboeckb, Daniela Haeuslerb,
Markus Mitterhauserb, Christian Windischbergerc, Wolfgang Wadsakb, Siegfried Kaspera,
Rupert Lanzenbergera*
a Department of Psychiatry and Psychotherapy,
b Department of Nuclear Medicine, PET Center,
c MR Center of Excellence, Center for Medical Physics and Biomedical Engineering,
Medical University of Vienna, Austria
Published in
NeuroImage, 2012 63(3):1091-1098,
[2014, IF: 6.35]
*Corresponding author:
A/Prof. Rupert Lanzenberger, MD
Department of Psychiatry and Psychotherapy
Functional, Molecular and Translational Neuroimaging - PET & MRI
Medical University of Vienna
Waehringer Guertel 18-20, 1090 Vienna, Austria
http://www.meduniwien.ac.at/neuroimaging

31
ABSTRACT
Animal models revealed that the serotonin-1A (5-HT1A) receptor modulates gray matter
structure. However, there is a lack of evidence showing the relationship between 5-HT1A
receptor concentration and gray matter in the human brain in vivo. Here, to demonstrate an
association between the 5-HT1A receptor binding potential, an index for receptor
concentration, and the local gray matter volume (GMV), an index for gray matter structure,
we measured 35 healthy subjects with both positron emission tomography (PET) and
structural magnetic resonance imaging (MRI). We found that regional heteroreceptor binding
was positively associated with GMV in distinctive brain regions such as the hippocampi and
the temporal cortices in both hemispheres (R2 values ranged from 0.308 to 0.503, p < 0.05
cluster-level FDR-corrected). Furthermore, autoreceptor binding in the midbrain raphe region
was positively associated with GMV in forebrain projection sites (R2 = 0.656, p = 0.001). We
also observed a broad range between 5-HT1A receptor binding and GMV. Given the
congruence of altered 5-HT1A receptor concentrations and GMV reduction in depression or
Alzheimer’s disease as reported by numerous studies, these results might provide new
insights towards understanding the mechanisms behind GMV alterations observed in these
brain disorders.
Key words: positron emission tomography, structural magnetic resonance imaging, 5-HT1A
receptor

32
INTRODUCTION
Growing evidence shows distinctive neuromodulatory properties of serotonin (5-
hydroxytryptamine, 5-HT) in developing and mature brain networks (Daubert and Condron,
2010; Gaspar et al., 2003). Early alterations in the 5-HT system are associated with life-long
changes in cognitive and behavioral functioning and the neuronal organization in
neuropsychiatric diseases (Gaspar et al., 2003). The 5-HT1A receptor, one of at least 16
receptors in the serotonergic system, is directly linked to signaling cascades mediating
neuroplasticity (Azmitia, 2001). Structural neuroimaging techniques revealed increased
amounts of gray matter volume (GMV) as surrogate for enhanced neuroplasticity in relation
to motoric training, cognitive performance or treatment with the antidepressant fluoxetine, a
selective serotonin reuptake inhibitor (Draganski et al., 2004; Kanai and Rees, 2011;
Vetencourt et al., 2008). On the other side, GMV loss as measured with high-resolution
structural magnetic resonance imaging (MRI), is a key feature of neuropsychiatric brain
disorders, whereby the hippocampal formation was demonstrated to be especially vulnerable
to volumetric alterations (Benninghoff et al., 2010; Geuze et al., 2005).
Serotonin-1A autoreceptors are located presynaptically on serotonergic neurons in the raphe
nuclei where they reduce tonic cell firing, thus autoinhibiting 5-HT release (Hall et al., 1997).
Postsynaptically, 5-HT1A heteroreceptors are expressed on glutamatergic and GABAergic
neurons and mediate an inhibitory serotonergic response (Amargós-Bosch et al., 2004; Hall
et al., 1997; Puig et al., 2005). Neurobiological studies identified a vast number of second
messenger pathways that exert neuroplastic changes (Citri and Malenka, 2008; Pittenger
and Duman, 2008) triggerd by 5-HT via 5-HT1A receptors (Azmitia, 2001; Tardito et al., 2006).
To sum up, 5-HT1A receptors might be involved in altering GMV, thereby offering a possible
explanation for gray matter atrophy observed in several brain disorders.
Dysfunctional neuronal organization is an important contributor to the pathogenesis of
Alzheimer’s disease (Mesulam, 1999), schizophrenia (Lewis and González-Burgos, 2008)
and depressive disorder (Pittenger and Duman, 2008), however the underlying molecular
mechanisms, leading to gray matter loss in these disorders are complex and not fully
understood. Interestingly, positron emission tomography (PET) studies demonstrated
alterations of 5-HT1A receptors in patients suffering from these disorders (Kepe et al., 2006),
(Mamo et al., 2007); (Kasper et al., 2002; Lanzenberger et al., 2007; Savitz et al., 2009). This
congruence and a lack of data in human brains in vivo lead us to investigate the relationship
between 5-HT1A receptor concentration and GMV with a multimodal neuroimaging approach.

33
MATERIAL AND METHODS
Participants
We examined 35 healthy adults, 18 males and 17 females (age range = 21-52, mean = 26.6
±6.8 years, Table 1), with at least general qualification for university entrance as lowest
educational level. All subjects were recruited via advertisement at the Medical University of
Vienna, Austria and underwent a general physical and neurological examination at the
screening visit including medical history, electrocardiogram and routine laboratory tests.
Inclusion criteria were age between 18 and 60, ability to perform study procedures and
absence of any acute or chronic disease. Exclusion criteria compromised any history of
severe disease, any psychiatric or neurologic disorder, previous drug abuse, pregnancy as
assessed by urine pregnancy tests and any continuous medication for three months prior to
the study. All participants provided written informed consent after written and oral
presentation of a general intelligible information form and received reimbursement after
participation. The institutional review board of the Medical University of Vienna, Austria, gave
approval to all study procedures. The pooled study sample consisted of subjects who were
part of positron emission tomography (PET) and magnetic resonance imaging (MRI) studies
previously published by our group (Hahn et al., 2010; Spindelegger et al., 2009).
Magnetic Resonance Imaging and Image Preprocessing
Structural magnetic resonance imaging was performed at the MR Center of Excellence at the
Medical University of Vienna, Austria, with a 3 Tesla whole-body MEDSPEC S300 MR-
scanner (Bruker BioSpin, Ettlingen, Germany) using a magnetization-prepared rapid gradient
echo (MPRAGE, T1-weighted) sequence (128 slices, 256 × 256 matrix, slice thickness 1.56
mm, voxel size 0.78 × 0.86 mm). To optimize image-preprocessing quality we used the
DARTEL (Diffeomorphic Anatomical Registration using Exponentiated Lie algebra) algorithm
(Ashburner, 2007), which ranked top in a comparison of 14 image registration algorithms
(Klein et al., 2009). The major advantage of the DARTEL algorithm is an increase in the
accuracy of inter-subject alignment by a high number of parameters derived from
deformation fields. T1-weighted images of all 35 subjects in our study were manually re-
oriented and segmented using the New Segment option in SPM8 (2009, Wellcome Trust
Centre for Neuroimaging, Institute of Neurology, University College London, London, United
Kingdom, http://www.fil.ion.ucl.ac.uk/spm/software/spm8/) to generate rigid-body aligned
gray matter, white matter and CSF-images. After segmentation all images were visually
checked for major artifacts. The DARTEL algorithm consecutively generates six individual
templates based on deformation fields calculated during segmentation, where the last

34
template produced (number 6) was used for normalization. Each individual’s segmented gray
matter image together with each deformation field and the template was normalized to
standard Montreal Neurological Institute (MNI) space at a voxel size of 1.5 × 1.5 × 1.5mm. To
correct for nonlinear spatial normalization, images were modulated by multiplication with the
Jacobian determinants of the deformation fields in order to preserve the actual amount gray
matter within each structure before normalization. Based on this, the modulated images are
further referred to as gray matter volume (GMV). The resultant values represent a
quantitative measure of gray matter tissue volume per unit volume of the spatially normalized
images (Ashburner and Friston, 2009). Finally, GMV images were smoothed with an 8-mm
full-width at half-maximum Gaussian kernel. Such smoothing is considered sufficient to
increase the stability of segmented images with respect to small registration errors.
Radiochemistry
The 5-HT1A receptor specific radioligand [carbonyl-11C]WAY-100635 was prepared at the
Cyclotron Unit of the PET center at the Department of Nuclear Medicine of the Medical
University of Vienna, Austria according to the optimized synthesis instruction proposed by
Wadsak et al. (2007). [Carbonyl-11C]WAY-100635 was prepared in a multistep radiosynthesis
starting from cyclotron-produced [11C]CO2 and purified by high-performance liquid
chromatography and solid-phase extraction. [carbonyl-11C]WAY-100635 was dissolved in a
phosphate-buffered saline solution and injected at a target dose of 5.4 MBq/kg bodyweight,
further details of radiochemical variables are given in Table 1.
Positron Emission Tomography (PET) Measurements
PET was performed at the Department of Nuclear Medicine of the Medical University of
Vienna, Austria with a GE advance full-ring scanner (General Electric Medical Systems,
Milwaukee, WI). Each subject’s head was placed in the scanner parallel to the orbitomeatal
line guided by a laser beam system to ensure full coverage of the neocortex and the
cerebellum in the field of view (FOV). A polyurethane cushion and head straps kept the head
in position to minimize head movement and to guarantee a soft head rest during the whole
scanning period. Initially, a 5-minute transmission scan in two-dimensional mode was
conducted to correct for tissue attenuation with a retractable 68Ge ring source. Dynamic PET
scans started simultaneously with the intravenous bolus injection of the radioligand
[carbonyl-11C]WAY-100635. PET scans lasted for 90 minutes per subject and were acquired
in a three-dimensional mode. The overall dynamic scan time was divided in 30 successive

35
time frames (15 × 1 minute, 15 × 5 minutes). The emission data were scatter- and
attenuation corrected based on the data from the transmission scans and reconstructed
using an iterative filtered back-projection algorithm (FORE + ITER). The final spatial
resolution of the reconstructed volume was 4.36 mm full-width at half maximum at the center
of the FOV. We did not perform realignment for head movement upon visual inspection of
PET-data quality. All 30 dynamic PET image frames were summed (PETADD) for co-
registration to the MRI.
Quantification of 5-HT1A Receptor Binding Potential
We assessed in vivo receptor density as indexed by 5-HT1A receptor binding potential (BPND),
the ratio at equilibrium of specifically bound radioligand to that of nondisplaceable radioligand
in tissue (Innis et al., 2007). Binding was computed using the voxel-wise modeling tool in the
PMOD software package (v3.1, 2009, for Linux, PMOD Technologies, Zurich, Switzerland,
http://www.pmod.com) and applying the two-parameter linearized reference tissue model
(MRTM2) (Ichise et al., 2003). Compared to other models such as the simplified reference
tissue model (SRTM), MRTM2 leads to lower BPND bias and hence a better signal-to-noise-
ratio, especially for whole-brain voxel-by-voxel analysis. We modeled 5-HT1A BPND as
previously described by our group using the insula and the cerebellum taken from an
automated anatomical labeling-based (AAL) region of interest (ROI) (Tzourio-Mazoyer et al.,
2002) atlas, as receptor-rich and receptor-poor region, respectively. The cerebellum
excluding cerebellar vermis served as reference region. This was done for the voxel-by-voxel
analysis as well as the ROI-based multimodal analysis.
Image Co-Registration for Multimodal Data Analysis
We combined the advantages of PET in quantifying receptors at the molecular level with
structural MRI, which provides data on brain structure such as cortical folding, regional
cortical thickness and volume or gray/white matter contrast. This was achieved by co-
registration of each individual’s PET image to the corresponding structural MRI image. We
used SPM8 to apply the transformation matrix of the structural scans obtained during
normalization to the PETADD images. As the structural scans were already normalized to
standard MNI space, this step also brought the PET data to MNI space resulting in whole-
brain dynamic [carbonyl-11C]WAY-100635 maps co-registered to the structural MRI images.

36
Quantification of 5-HT1A Receptor Binding Potential in Anatomical Regions of Interest
With this post-hoc analysis we aimed to investigate the area-specific relationship between 5-
HT1A and GMV to further confirm our primary voxel-by-voxel results using a different
approach. The ROI-based analysis also served to test, whether there exists a network
association between 5-HT1A receptors and gray matter in the projection areas of the one of
the main raphe nuclei, the dorsal raphe nucleus. For the ROI-based network analysis the
DRN was manually delineated on an averaged PETADD image in PMOD 3.1. Our DRN ROI
consisted of a sphere, 4 mm in diameter, comprising three slices on the averaged PETADD
image. Each individual’s 5-HT1A BPND value in the DRN was then obtained from individual
time-activity curves averaged across subjects (again using cerebellar gray matter as
reference). For further post-hoc analysis we quantified 5-HT1A receptor BPND values and
GMV values in MNI standard space in 48 ROIs, taken from the AAL atlas covering a broad
range of brain regions as previously shown (Stein et al., 2008). A ratio between GMV and
rescaled 5-HT1A BPND values was calculated by dividing the first through the latter.
Statistical analyses
Demographics
Sex differences in biological demographical and radiochemical variables were calculated to
assess study sample characteristics with either independent sample t-tests or Mann-Whitney
U tests where appropriate, using IBM SPSS Statistics (v19.0, 2010, SPSS, Inc., an IBM
Company, Chicago, United States of America) assuming a significance level of α = 0.05.
Multimodal Analysis
Multimodal image analysis was divided into two parts: we calculated regional voxel-by-voxel
associations between 5-HT1A receptor distribution and gray matter in the whole-brain.
Second, we assessed the associations between 5-HT1A receptor binding in a single area, the
DRN, and GMV in projection sites of the DRN. The DRN was chosen because of its central
role in the regulation of serotonergic firing and neurotransmission.
We thus calculated a multiple linear regression model with 5-HT1A receptor BPND values as
independent and GMV values as dependent variable for every voxel in the entire gray matter.
In this regression model age, sex and total GMV served as controlling variables. This was
done to adjust for age related gray matter alteration, varying brain sizes, and sex differences
(as outlined in Table 1) for. In two further models, we also considered the two radiochemical
variables specific activity (SA) and injected dose (ID) as factors. However, given that the

37
number of control variables should not exceed n/10 and the results (data not shown) were
virtually identical with the primary model, we did not include SA and ID in our further
analyses. The voxel-by-voxel regression model was set up in the Biological Parametric
Mapping (BPM) toolbox for SPM8 (Casanova et al., 2007), which is designed to calculate
voxel-by-voxel statistics for multiple imaging modalities. More precisely, multiple regression
was calculated in each voxel (average voxel number across all subjects = 216,741.2) with
one value for GMV and one for 5-HT1A BPND (in arbitrary units). We used 0.1 as absolute
threshold and a level of statistical significance of α = 0.001. Due to multiple comparisons and
the concomitant high chance of false positives the obtained results were corrected with the
cluster-level false discovery rate (FDR) at a significance level of α = 0.05. Correlation
coefficients were calculated with cluster-wise means (in arbitrary units) in Matlab (v. r2010b,
The MathWorks, Inc., Natick, United States of America).
For the analysis of serotonergic projections from the DRN, we calculated a regression model
in SPM8 using 5-HT1A BPND values of the DRN ROI as independent variables and whole
brain GMV as dependent variable. Sex, age and total GMV were control variables for the
reasons mentioned above. Further, GMV of the DRN was added in the regression model to
eliminate potential confounding effects of DRN gray matter and whole brain gray matter
interactions. GMV values were obtained from the DRN ROI overlaid on the MRI images. We
excluded voxels exhibiting BPND or GMV voxel values below 0.1. The level of statistical
significance was set at α = 0.001 and only results with a cluster size over 100 voxels are
reported.
For the analysis of smoking status on BPND a regression analysis was set up in SPM8 using
5-HT1A BPND values as independent variables and smoking status or number of smoked
cigarettes as dependent variables, respectively, controlling for sex, age and GMV. Age effects
on GMV were calculated with a regression analysis using GMV as independent variable and
age as dependent variable controlling for sex and GMV. In both analyses an uncorrected α =
0.001 was accepted as level of significance.
RESULTS
5-HT1A receptor binding positively correlated with gray matter volumes within
distinctive brain regions
In this pooled study sample, male study subjects significantly differed from females in GMV,
weight and total injected radiotracer dose (Table 1). In line with previous results of our group,
5-HT1A BPND, an index for receptor density, peaked in the parahippocampal gyri, the temporal
poles and the insula (Figures 1A, Figure 3, Table S1 and [Stein et al., 2008]).

38
Serotonin-1A BPND strongly correlated with GMV in the hippocampus (the cluster in the right
hippocampus spread from the posterior hippocampus to the parahippocampus), the posterior
medial temporal cortex, the posterior inferior temporal cortex, the medial occipital cortex and
the pericalcarine region in each hemisphere (R2 values ranged from 0.308 to 0.503, p < 0.05
cluster-level false discovery rate [FDR]) corrected, see Figures 1B, 1C and Table 2). In other
words, 5-HT1A heteroreceptor binding strongly correlated with relative volumes of gray matter
in these specific regions. Negative correlations between 5-HT1A BPND and GMV were
restricted to two regions in the cerebellum (Table 2).
Figure 1 Serotonin-1A (5-HT1A) receptor binding is positively associated with regional gray matter.
(A) 5-HT1A receptor distribution in vivo measured with positron emission tomography displayed with
the surface-rendering algorithm used by the visualization program MRIcro
(http://www.cabiatl.com/mricro/mricro/mricro.html) (B) T maps showing that 5-HT1A binding potential
(BPND) strongly correlates with gray matter volume (GMV). Significant positive correlations were
superimposed on MR images (p < 0.05, FDR cluster-level corrected, see Table 2), coordinates
correspond to the standard Montreal Neurological Institute (MNI) stereotactic system. (C) Regression
graphs between GMV and 5-HT1A BPND (multiple regression analysis controlled for sex, age and total
GMV, adjusted values in arbitrary units) correspond to cluster means of each subject (in red circles
(B), N = 35).
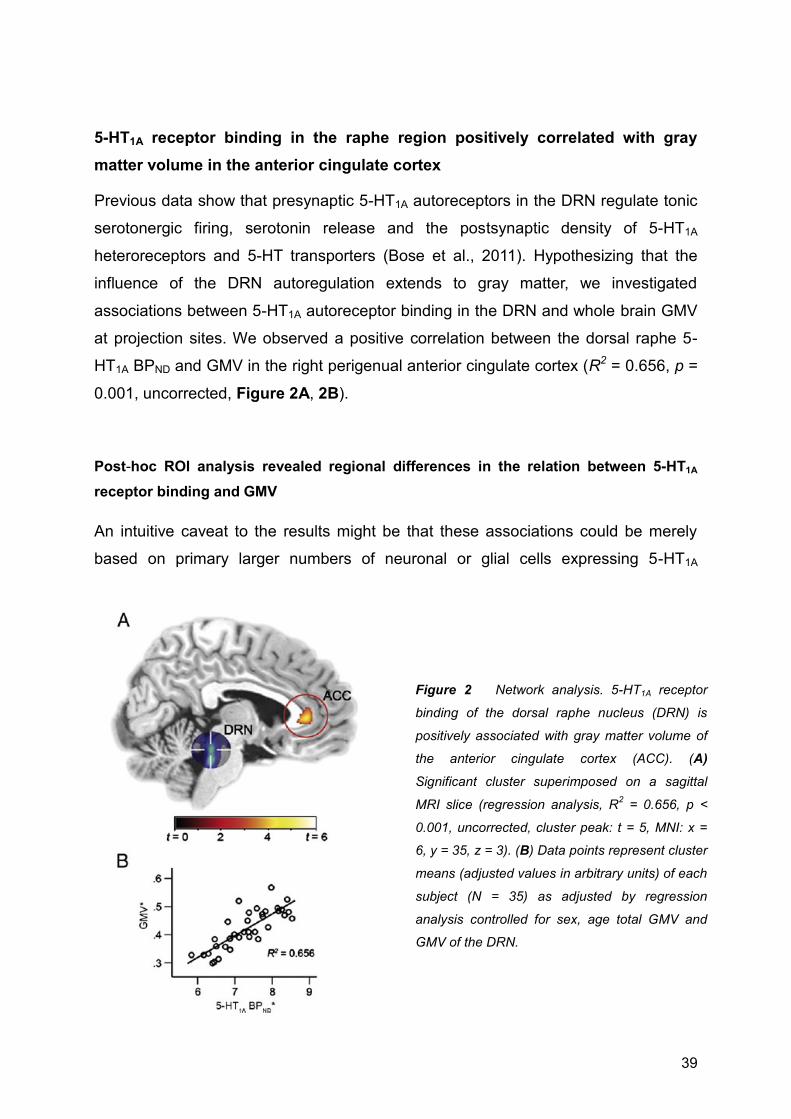
39
5-HT1A receptor binding in the raphe region positively correlated with gray
matter volume in the anterior cingulate cortex
Previous data show that presynaptic 5-HT1A autoreceptors in the DRN regulate tonic
serotonergic firing, serotonin release and the postsynaptic density of 5-HT1A
heteroreceptors and 5-HT transporters (Bose et al., 2011). Hypothesizing that the
influence of the DRN autoregulation extends to gray matter, we investigated
associations between 5-HT1A autoreceptor binding in the DRN and whole brain GMV
at projection sites. We observed a positive correlation between the dorsal raphe 5-
HT1A BPND and GMV in the right perigenual anterior cingulate cortex (R2 = 0.656, p =
0.001, uncorrected, Figure 2A, 2B).
Post-hoc ROI analysis revealed regional differences in the relation between 5-HT1A
receptor binding and GMV
An intuitive caveat to the results might be that these associations could be merely
based on primary larger numbers of neuronal or glial cells expressing 5-HT1A
Figure 2 Network analysis. 5-HT1A receptor
binding of the dorsal raphe nucleus (DRN) is
positively associated with gray matter volume of
the anterior cingulate cortex (ACC). (A)
Significant cluster superimposed on a sagittal
MRI slice (regression analysis, R2 = 0.656, p <
0.001, uncorrected, cluster peak: t = 5, MNI: x =
6, y = 35, z = 3). (B) Data points represent cluster
means (adjusted values in arbitrary units) of each
subject (N = 35) as adjusted by regression
analysis controlled for sex, age total GMV and
GMV of the DRN.

40
receptors and thus a priori higher GMV values. Therefore, we investigated if the
resulting clusters were exclusively situated in regions with high regional GMV and
quantified the BPND and GMV values of 48 ROIs covering the whole brain. We found
several regions, such as the cingulate cortex or the amygdala, which despite high
regional GMV and 5-HT1A BPND values did not exhibit significant positive associations
in the voxel-by-voxel analysis (Figure 3). Furthermore, we calculated ratios between
GMV and BPND values to assess regional proportions between 5-HT1A receptor
binding and gray matter in the whole brain. These GMV/BPND ratios ranged from 0.54
in the temporal pole to 4.8 in the caudate region, suggesting high regional variability
within the ratio of regional GMV and 5-HT1A BPND (Figure 3).
Following that, to confirm the associations between 5-HT1A BPND and GMV obtained
by voxel-by-voxel analysis, we repeated the regression analysis within two ROIs. The
ROIs should have similar GMV and 5-HT1A BPND values, one exhibiting and one
lacking the associations as obtained by voxel-by-voxel analysis. Out of the 48
quantified ROIs, the hippocampus and the insula were the only two regions meeting
the selection criteria (GMV/BPND: insula = 0.63, hippocampus = 0.64, GMV: insula
and hippocampus = 0.53, BPND: insula = 0.84, hippocampus = 0.83, see Figure 3
and Table S1). In the voxel-by-voxel analysis, the hippocampus exhibited significant
positive associations between 5-HT1A receptor BPND and GMV, but in the insula,
despite similar values, 5-HT1A BPND did not correlate with GMV. Congruent to the
voxel-by-voxel analysis, in the post-hoc ROI analysis a significant positive correlation
was observed in the hippocampus (r = 0.41, p = 0.02) but not in the insula (r = - 0.03,
p = 0.87).
No effects of age or smoking status
To rule out cortical atrophy due to aging was somehow related to the results, we analyzed
our dataset for age-related effects. Multiple regression analysis in SPM8 revealed a negative

41
Figure 3 Area-specific differences in the relation between 5-HT1A receptor binding and gray matter
volume. Gray matter volume (GMV, red) and 5-HT1A binding potential (BPND, blue), in arbitrary units,
quantified in 48 brain regions of interest (ROI) covering the whole brain. This demonstrates high
variabilities between 5-HT1A receptor densities and regional volumes of gray matter (also see Table
S1).
correlation for GMV and age in the left medial occipital cortex (t = 4.24, p < 0.001,
uncorrected, x = -27, y = -81, z = 26) near the angular gyrus. BPND was negatively correlated
with age in a cluster around the left postcentral gyrus (t = 3.82, p < 0.001, uncorrected; x = -
22, y = -27, z = 62). These results indicate that an effect of aging in our data occurred in
different brain areas than the main results. Smoking status was available for 34 participants,
out of which 14 were smokers (6 female, mean cigarettes per day = 7.1 ± 4.8). Multiple

42
regression analysis revealed that neither smoking status nor number of smoked cigarettes
was associated with 5-HT1A BPND (all p > 0.001).
DISCUSSION
Our results demonstrate positive associations between 5-HT1A receptor binding and gray
matter. In distinctive regions of both hemispheres, as in the hippocampi and in temporal
cortices, 5-HT1A receptor binding was strongly correlated with gray matter. These results
were not just based on a priori higher regional values of gray matter, because we
demonstrated that in regions such as the insula, in contrast to the hippocampus, there were
no significant positive associations, although having comparable gray matter and 5-HT1A
receptor binding. We observed a large variability between 5-HT1A binding and gray matter in
the whole brain. We also found that 5-HT1A autoreceptor binding in the DRN was positively
associated with gray matter in the anterior cingulate cortex. The results were not affected by
cortical atrophy due to aging or smoking status. A large number of previous findings in animal
models (Daubert and Condron, 2010; Gaspar et al., 2003) show direct links between
serotonergic receptors like the 5-HT1A receptor and neuroplasticity. Furthermore, there is
evidence that allows direct inference from MRI-based measurements to changes of the
underlying neuronal structures (la Fougère et al., 2010). Therefore, we propose that the
discovered associations provide valuable insights into the relationship between 5-HT1A
receptor binding and gray matter cytoarchitecture in adult human brains in vivo.
Serotonin is highly active in shaping neurons during embryonic development and early
postnatal neuronal maturation, and this neuroplastic role is partially conserved in specific
brain regions throughout adulthood (Gould, 1999). Downstream cytosolic signaling kinases
from membrane-bound small G proteins (Ye and Carew, 2010), that activate transcription
factors (McClung and Nestler, 2008) and epigenetic mechanisms (Borrelli et al., 2008) were
suggested to effect neuronal reconfigurations. Serotonergic-1A receptors are able to
modulate the activity of these pathways (Cowen, 2007; Polter and Li, 2010). Recently a study
using hippocampal cell cultures could show, that 5-HT1A receptors are essential for normal
synaptogenesis (Mogha et al., 2012). Blockade of astrocytic 5-HT1A receptors leads to a
reduction of synaptic connections between neurons (Wilson et al., 1998) and fits well to
findings demonstrating that the 5-HT1A receptor is required for behavioral and neurogenic
effects of the selective serotonin reuptake inhibitors (Santarelli et al., 2003). From a
neurobiological perspective, we suggest that neuroplastic effects of 5-HT1A receptors might
contribute to the observed association between 5-HT1A binding and alterations of regional
gray matter.

43
Nevertheless, this interpretation must be considered with caution, because one voxel in high-
resolution structural MRI contains too many neuronal cells to reliably link our results with
mechanisms observed in cell cultures or animal models (for review see [(May, 2011)]). To
better pin down how 5-HT1A receptor-mediated neuroplasticity might affect gray matter in the
living brain, further longitudinal investigations are needed. A possible alternative explanation
to our results could simply be, that regional 5-HT1A binding was elevated through primary
higher regional amounts of gray matter. However, considering results from animal models,
we have several arguments against this.
Firstly, we predominately found positive associations. In a similar study using structural MRI,
PET and the D2/D3 receptor ligand [18F]fallypride, Woodward et al. (Woodward et al., 2009)
previously pointed out that negative associations are unexpected. Hypothetically, the density
of 5-HT1A receptors should vary with the amount of gray matter within a region, in other
words the more gray matter, the more receptors it can support and vice versa. A recent study,
however, nicely demonstrates divergent values between 5-HT1A binding and neuronal
densities in humans as measured by stereology and autoradiography (Underwood et al.,
2008). This result is congruent to the broad variability in the range between 5-HT1A binding
and GMV shown in our study. The predominantly positive associations in our study go well
along with the finding, that astrocytic 5-HT1A receptors (via S-100ß) are necessary for
maintaining the neuronal integrity (Whitaker-Azmitia, 2001). In the absence of S-100ß a
mature neuron can regress its major processes and even enter apoptosis (Whitaker-Azmitia,
2001). Secondly, the associations between 5-HT1A binding and gray matter were obtained
symmetrically in both hemispheres, which indicates validity. The distinctive regional pattern
could be explained by varying strengths of 5-HT1A mediated neuroplastic effects (Cowen,
2007). Thirdly, the insula, even with similar 5-HT1A and GMV values as the hippocampus, did
not exhibhit significant associations. This further suggests a region-specific mechanism and
might indicate that the observed associations were not based on higher numbers of regional
neuronal and glial cells, both associated with GMV. According to the current state of
knowledge 5-HT1A mediated neroplasticity is more active in the hippocampus than in the
insula (Santarelli et al., 2003). In the hippocampus 5-HT1A receptors were demonstrated to
stimulate neurogenesis and dendritic maturation (Yan et al., 1997). Finally, we demonstrated
an association between 5-HT1A receptors in one of the major serotonergic nuclei, the DRN,
and gray matter at serotonergic axon terminals in the anterior cingulate cortex.
Hypothetically, interregional correlation between 5-HT1A auto- and heteroreceptors (Hahn et
al., 2010) could foster the observed association between 5-HT1A receptors and GMV in this
study. The autoregulatory influence of the DRN on serotonergic heteroreceptors at axon
terminals in the forebrain, by neuroplastic properties of 5-HT1A receptors, might thus extend
to GMV. Patients suffering from mood disorders exhibit both significantly reduced GMV of the

44
anterior cingulate cortex and altered 5-HT1A receptor density in the raphe nucleus (Salvadore
et al., 2011; Savitz and Drevets, 2009; van Tol et al., 2010). We could speculate here, that a
disturbance in this association might contribute to the reduction of GMV in the anterior
cingulate cortex.
In summary, the associations between 5-HT1A binding and GMV could theoretically result
from a priori higher regional amounts of gray matter or other unknown mechanisms. But
given the high amount of clear evidence, we suggest that neuroplastic actions of 5-HT1A
receptors should be taken into account as explanatory model for this dataset. The 5-HT1A
receptor, in addition, could prove to be an interesting target in clinical studies on altered
neuroplasticity in brain disorders, due to well known behavioral behavioral functions such as
mediating mood (Savitz et al., 2009), anxiety (Akimova et al., 2009) or cognition (Ogren et
al., 2008).
Limitations
This dataset does not imply that the observed associations can be causally attributed to
neuroplastic actions of 5-HT1A receptors. For such a deduction a longitudinal, interventional
and translational approach in a future study would be more favorable, for which this dataset
provides excellent justification.
Furthermore, as previously pointed out (Tost et al., 2010) the neurobiological correlates of
changes in brain morphology measured by structural neuroimaging are not sufficiently
resolved, for an excellent recent review see (Zatorre et al., 2012). Even at high-resolution
MRI, there are still ten thousands of interconnected neuronal and glial cells packed in one
single voxel. Thus, more translational cell studies on neuroplasticity are necessary to exactly
determine what cellular processes are mediated by serotonin and the 5-HT1A receptor that
could gain effects, large enough to be detectable by structural MRI (May, 2011).
Finally, we did not use correction for partial volume effects (PVC) of the PET data. Although
this may be an obvious issue, PVC is typically carried out by using the corresponding
segmented MRI, namely, the gray and white matter probability maps. More precisely, the GM
values represent the denominator of the PVC algorithm (Muller-Gartner et al., 1992). This
implies that the PET activity concentrations are adjusted for individual differences in the GM
volume. However, the current study particularly aims to investigate the association between
individual differences in 5-HT1A binding and GM volume. Hence, MRI-based PVC would
include the effect of interest as nuisance variable, which in turn cancels the association.
Accordingly, no PVC was carried out in the similar investigation of Woodward et al. (2009).

45
Conclusions
Our results demonstrate that 5-HT1A receptor binding is positively associated with gray
matter in specific regions such as the hippocampus and the temporal cortices in both
hemispheres. Furthermore 5-HT1A autoreceptor binding in the midbrain is positively
associated gray matter in the anterior cingulate cortex. Currently, it is hard to pin down the
molecular mechanisms underlying our results, mostly because, there are no exact models
which cellular compounds correspond to the signal strength in a single voxel. To increase the
validity of neuroimaging studies, this issue must be an objective of further studies.
With regard to translational neuroscience, assessments of processes underlying networking
and reorganization of neurons as well as early surrogate markers to predict and monitor
treatment response were demanded (Cramer et al., 2011). With combinations of structural
and molecular neuroimaging, as performed in this multimodal study, dysfunctional
neuroregulatory processes leading to loss of gray matter might be investigated at early
stages in clinical populations. This could lead to a more comprehensive understanding of
neurodegenerative diseases as Alzheimer’s disease, schizophrenia and mood disorders and
ultimately to a better diagnostic assessment and therapeutic evaluation of patients with these
highly life impairing disorders.
ACKNOWLEDGEMENTS
This research was partly supported by grants from the Austrian Science Fund, and the
Austrian National Bank (P 11468) to R. L. A. Hahn is recipient of a DOC-fellowship of the
Austrian Academy of Sciences at the Department of Psychiatry and Psychotherapy. We are
grateful to the technical and medical teams of the PET and High-Field MRI Centre, Medical
University of Vienna, especially to K. Kletter, R. Dudczak, E. Moser, L.-K. Mien, and F. Gerstl.
Furthermore, we would like to thank U. Moser, M. Fink, and P. Stein for medical support and
A. Saulin for help with the manuscript.

46
REFERENCES
Akimova, E., Lanzenberger, R., Kasper, S., 2009. The serotonin-1A receptor in anxiety disorders. Biol Psychiatry 66, 627-635.
Amargós-Bosch, M., Bortolozzi, A., Puig, M.V., Serrats, J., Adell, A., Celada, P., Toth, M., Mengod, G., Artigas, F., 2004. Co-expression and in vivo interaction of serotonin1A and serotonin2A receptors in pyramidal neurons of prefrontal cortex. Cereb Cortex 14, 281-299.
Ashburner, J., 2007. A fast diffeomorphic image registration algorithm. NeuroImage 38, 95-113.
Ashburner, J., Friston, K.J., 2009. Computing average shaped tissue probability templates. NeuroImage 45, 333-341.
Azmitia, E.C., 2001. Modern views on an ancient chemical: serotonin effects on cell proliferation, maturation, and apoptosis. Brain Res Bull 56, 413-424.
Benninghoff, J., Gritti, A., Rizzi, M., Lamorte, G., Schloesser, R.J., Schmitt, A., Robel, S., Genius, J., Moessner, R., Riederer, P., Manji, H.K., Grunze, H., Rujescu, D., Moeller, H.-J., Lesch, K.-P., Vescovi, A.L., 2010. Serotonin depletion hampers survival and proliferation in neurospheres derived from adult neural stem cells. Neuropsychopharmacology 35, 893-903.
Borrelli, E., Nestler, E.J., Allis, C.D., Sassone-Corsi, P., 2008. Decoding the epigenetic language of neuronal plasticity. Neuron 60, 961-974.
Bose, S.K., Mehta, M.A., Selvaraj, S., Howes, O.D., Hinz, R., Rabiner, E.A., Grasby, P.M., Turkheimer, F.E., Murthy, V., 2011. Presynaptic 5-HT1A is Related to 5-HTT Receptor Density in the Human Brain. Neuropsychopharmacology.
Casanova, R., Srikanth, R., Baer, A., Laurienti, P.J., Burdette, J.H., Hayasaka, S., Flowers, L., Wood, F., Maldjian, J.A., 2007. Biological parametric mapping: A statistical toolbox for multimodality brain image analysis. NeuroImage 34, 137-143.
Citri, A., Malenka, R.C., 2008. Synaptic plasticity: multiple forms, functions, and mechanisms. Neuropsychopharmacology 33, 18-41.
Cowen, D.S., 2007. Serotonin and neuronal growth factors - a convergence of signaling pathways. J Neurochem 101, 1161-1171.
Cramer, S.C., Sur, M., Dobkin, B.H., O'Brien, C., Sanger, T.D., Trojanowski, J.Q., Rumsey, J.M., Hicks, R., Cameron, J., Chen, D., Chen, W.G., Cohen, L.G., deCharms, C., Duffy, C.J., Eden, G.F., Fetz, E.E., Filart, R., Freund, M., Grant, S.J., Haber, S., Kalivas, P.W., Kolb, B., Kramer, A.F., Lynch, M., Mayberg, H.S., McQuillen, P.S., Nitkin, R., Pascual-Leone, A., Reuter-Lorenz, P., Schiff, N., Sharma, A., Shekim, L., Stryker, M., Sullivan, E.V., Vinogradov, S., 2011. Harnessing neuroplasticity for clinical applications. Brain 134, 1591-1609.
Daubert, E.A., Condron, B.G., 2010. Serotonin: a regulator of neuronal morphology and circuitry. Trends Neurosci 33, 424-434.
Draganski, B., Gaser, C., Busch, V., Schuierer, G., Bogdahn, U., May, A., 2004. Neuroplasticity: changes in grey matter induced by training. Nature 427, 311-312.
Gaspar, P., Cases, O., Maroteaux, L., 2003. The developmental role of serotonin: news from mouse molecular genetics. Nat Rev Neurosci 4, 1002-1012.

47
Geuze, E., Vermetten, E., Bremner, J.D., 2005. MR-based in vivo hippocampal volumetrics: 2. Findings in neuropsychiatric disorders. Mol Psychiatry 10, 160-184.
Gould, E., 1999. Serotonin and hippocampal neurogenesis. Neuropsychopharmacology 21, 46S-51S.
Hahn, A., Lanzenberger, R., Wadsak, W., Spindelegger, C., Moser, U., Mien, L.-K., Mitterhauser, M., Kasper, S., 2010. Escitalopram enhances the association of serotonin-1A autoreceptors to heteroreceptors in anxiety disorders. J Neurosci 30, 14482-14489.
Hall, H., Lundkvist, C., Halldin, C., Farde, L., Pike, V.W., McCarron, J.A., Fletcher, A., Cliffe, I.A., Barf, T., Wikstrom, H., Sedvall, G., 1997. Autoradiographic localization of 5-HT1A receptors in the post-mortem human brain using [3H]WAY-100635 and [11C]way-100635. Brain Res 745, 96-108.
Ichise, M., Liow, J.-S., Lu, J.-Q., Takano, A., Model, K., Toyama, H., Suhara, T., Suzuki, K., Innis, R.B., Carson, R.E., 2003. Linearized Reference Tissue Parametric Imaging Methods: Application to [11C]DASB Positron Emission Tomography Studies of the Serotonin Transporter in Human Brain. Journal of Cerebral Blood Flow & Metabolism, 1096-1112.
Innis, R.B., Cunningham, V.J., Delforge, J., Fujita, M., Gjedde, A., Gunn, R.N., Holden, J., Houle, S., Huang, S.-C., Ichise, M., Iida, H., Ito, H., Kimura, Y., Koeppe, R.A., Knudsen, G.M., Knuuti, J., Lammertsma, A.A., Laruelle, M., Logan, J., Maguire, R.P., Mintun, M.A., Morris, E.D., Parsey, R., Price, J.C., Slifstein, M., Sossi, V., Suhara, T., Votaw, J.R., Wong, D.F., Carson, R.E., 2007. Consensus nomenclature for in vivo imaging of reversibly binding radioligands. J Cereb Blood Flow Metab 27, 1533-1539.
Kanai, R., Rees, G., 2011. The structural basis of inter-individual differences in human behaviour and cognition. Nature reviews Neuroscience.
Kasper, S., Tauscher, J., Willeit, M., Stamenkovic, M., Neumeister, A., Kufferle, B., Barnas, C., Stastny, J., Praschak-Rieder, N., Pezawas, L., de Zwaan, M., Quiner, S., Pirker, W., Asenbaum, S., Podreka, I., Brucke, T., 2002. Receptor and transporter imaging studies in schizophrenia, depression, bulimia and Tourette's disorder--implications for psychopharmacology. World J Biol Psychiatry 3, 133-146.
Kepe, V., Barrio, J.R., Huang, S.C., Ercoli, L., Siddarth, P., Shoghi-Jadid, K., Cole, G.M., Satyamurthy, N., Cummings, J.L., Small, G.W., Phelps, M.E., 2006. Serotonin 1A receptors in the living brain of Alzheimer's disease patients. Proc Natl Acad Sci U S A 103, 702-707.
Klein, A., Andersson, J., Ardekani, B.A., Ashburner, J., Avants, B., Chiang, M.-C., Christensen, G.E., Collins, D.L., Gee, J., Hellier, P., Song, J.H., Jenkinson, M., Lepage, C., Rueckert, D., Thompson, P., Vercauteren, T., Woods, R.P., Mann, J.J., Parsey, R.V., 2009. Evaluation of 14 nonlinear deformation algorithms applied to human brain MRI registration. NeuroImage 46, 786-802.
la Fougère, C., Grant, S., Kostikov, A., Schirrmacher, R., Gravel, P., Schipper, H., Reader, A., Evans, A., Thiel, A., 2010. Where in-vivo imaging meets cytoarchitectonics: The relationship between cortical thickness and neuronal density measured with high-resolution [(18)F]flumazenil-PET. NeuroImage.
Lanzenberger, R.R., Mitterhauser, M., Spindelegger, C., Wadsak, W., Klein, N., Mien, L.-K., Holik, A., Attarbaschi, T., Mossaheb, N., Sacher, J., Geiss-Granadia, T., Kletter, K., Kasper, S., Tauscher, J., 2007. Reduced serotonin-1A receptor binding in social anxiety disorder. Biol Psychiatry 61, 1081-1089.

48
Lewis, D.A., González-Burgos, G., 2008. Neuroplasticity of Neocortical Circuits in Schizophrenia. Neuropsychopharmacology 33, 141-165.
Mamo, D., Graff, A., Mizrahi, R., Shammi, C.M., Romeyer, F., Kapur, S., 2007. Differential effects of aripiprazole on D(2), 5-HT(2), and 5-HT(1A) receptor occupancy in patients with schizophrenia: a triple tracer PET study. Am J Psychiatry 164, 1411-1417.
May, A., 2011. Experience-dependent structural plasticity in the adult human brain. Trends Cogn Sci 15, 475-482.
McClung, C.A., Nestler, E.J., 2008. Neuroplasticity mediated by altered gene expression. Neuropsychopharmacology 33, 3-17.
Mesulam, M.M., 1999. Neuroplasticity failure in Alzheimer's disease: bridging the gap between plaques and tangles. Neuron 24, 521-529.
Mogha, A., Guariglia, S., Debata, P., Wen, G., Banerjee, P., 2012. Serotonin 1A receptor-mediated signaling through ERK and PKCa is essential for normal synaptogenesis in neonatal mouse hippocampus. Translational Psychiatry 2.
Muller-Gartner, H.W., Links, J.M., Prince, J.L., Bryan, R.N., McVeigh, E., Leal, J.P., Davatzikos, C., Frost, J.J., 1992. Measurement of radiotracer concentration in brain gray matter using positron emission tomography: MRI-based correction for partial volume effects. J Cereb Blood Flow Metab 12, 571-583.
Ogren, S.O., Eriksson, T.M., Elvander-Tottie, E., D'Addario, C., Ekström, J.C., Svenningsson, P., Meister, B., Kehr, J., Stiedl, O., 2008. The role of 5-HT(1A) receptors in learning and memory. Behav Brain Res 195, 54-77.
Pittenger, C., Duman, R.S., 2008. Stress, depression, and neuroplasticity: a convergence of mechanisms. Neuropsychopharmacology 33, 88-109.
Puig, M.V., Artigas, F., Celada, P., 2005. Modulation of the activity of pyramidal neurons in rat prefrontal cortex by raphe stimulation in vivo: involvement of serotonin and GABA. Cereb Cortex 15, 1-14.
Salvadore, G., Nugent, A.C., Lemaitre, H., Luckenbaugh, D.A., Tinsley, R., Cannon, D.M., Neumeister, A., Jr, C.A.Z., Drevets, W.C., 2011. Prefrontal cortical abnormalities in currently depressed versus currently remitted patients with major depressive disorder. NeuroImage 54, 2643-2651.
Santarelli, L., Saxe, M., Gross, C., Surget, A., Battaglia, F., Dulawa, S., Weisstaub, N., Lee, J., Duman, R., Arancio, O., Belzung, C., Hen, R., 2003. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 301, 805-809.
Savitz, J., Lucki, I., Drevets, W.C., 2009. 5-HT(1A) receptor function in major depressive disorder. Prog Neurobiol 88, 17-31.
Savitz, J.B., Drevets, W.C., 2009. Imaging phenotypes of major depressive disorder: genetic correlates. Neuroscience 164, 300-330.
Spindelegger, C., Lanzenberger, R., Wadsak, W., Mien, L.K., Stein, P., Mitterhauser, M., Moser, U., Holik, A., Pezawas, L., Kletter, K., Kasper, S., 2009. Influence of escitalopram treatment on 5-HT 1A receptor binding in limbic regions in patients with anxiety disorders. Mol Psychiatry 14, 1040-1050.

49
Stein, P., Savli, M., Wadsak, W., Mitterhauser, M., Fink, M., Spindelegger, C., Mien, L.K., Moser, U., Dudczak, R., Kletter, K., Kasper, S., Lanzenberger, R., 2008. The serotonin-1A receptor distribution in healthy men and women measured by PET and [carbonyl-11C]WAY-100635. Eur J Nucl Med Mol Imaging 35, 2159-2168.
Tardito, D., Perez, J., Tiraboschi, E., Musazzi, L., Racagni, G., Popoli, M., 2006. Signaling pathways regulating gene expression, neuroplasticity, and neurotrophic mechanisms in the action of antidepressants: a critical overview. Pharmacol Rev 58, 115-134.
Tost, H., Braus, D.F., Hakimi, S., Ruf, M., Vollmert, C., Hohn, F., Meyer-Lindenberg, A., 2010. Acute D2 receptor blockade induces rapid, reversible remodeling in human cortical-striatal circuits. Nat Neurosci 13, 920-922.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello, F., Etard, O., Delcroix, N., Mazoyer, B., Joliot, M., 2002. Automated Anatomical Labeling of Activations in SPM Using a Macroscopic Anatomical Parcellation of the MNI MRI Single-Subject Brain. NeuroImage 15, 273-289.
Underwood, M.D., Mann, J.J., Huang, Y.-Y., Arango, V., 2008. Family history of alcoholism is associated with lower 5-HT2A receptor binding in the prefrontal cortex. Alcohol Clin Exp Res 32, 593-599.
van Tol, M.-J., van der Wee, N.J.A., van den Heuvel, O.A., Nielen, M.M.A., Demenescu, L.R., Aleman, A., Renken, R., van Buchem, M.A., Zitman, F.G., Veltman, D.J., 2010. Regional brain volume in depression and anxiety disorders. Arch Gen Psychiatry 67, 1002-1011.
Vetencourt, J.F.M., Sale, A., Viegi, A., Baroncelli, L., De Pasquale, R., O'Leary, O.F., Castrén, E., Maffei, L., 2008. The antidepressant fluoxetine restores plasticity in the adult visual cortex. Science 320, 385-388.
Wadsak W., M.L.K., Ettlinger D.E., Lanzenberger R., Haeusler D., Dudczak R., Kletter K., Mitterhauser M., 2007. Simple and fully automated preparation of [carbonyl-11C]WAY-100635. Radiochimica Acta 95, 1-6.
Whitaker-Azmitia, P.M., 2001. Serotonin and brain development: role in human developmental diseases. Brain Res Bull 56, 479-485.
Wilson, C.C., Faber, K.M., Haring, J.H., 1998. Serotonin regulates synaptic connections in the dentate molecular layer of adult rats via 5-HT1a receptors: evidence for a glial mechanism. Brain Res 782, 235-239.
Woodward, N.D., Zald, D.H., Ding, Z., Riccardi, P., Ansari, M.S., Baldwin, R.M., Cowan, R.L., Li, R., Kessler, R.M., 2009. Cerebral morphology and dopamine D2/D3 receptor distribution in humans: a combined [18F]fallypride and voxel-based morphometry study. NeuroImage 46, 31-38.
Yan, W., Wilson, C.C., Haring, J.H., 1997. 5-HT1a receptors mediate the neurotrophic effect of serotonin on developing dentate granule cells. Brain Res Dev Brain Res 98, 185-190.
Ye, X., Carew, T.J., 2010. Small G protein signaling in neuronal plasticity and memory formation: the specific role of ras family proteins. Neuron 68, 340-361.
Zatorre, R.J., Fields, R.D., Johansen-Berg, H., 2012. Plasticity in gray and white: neuroimaging changes in brain structure during learning. Nat Neurosci 15, 528-536.
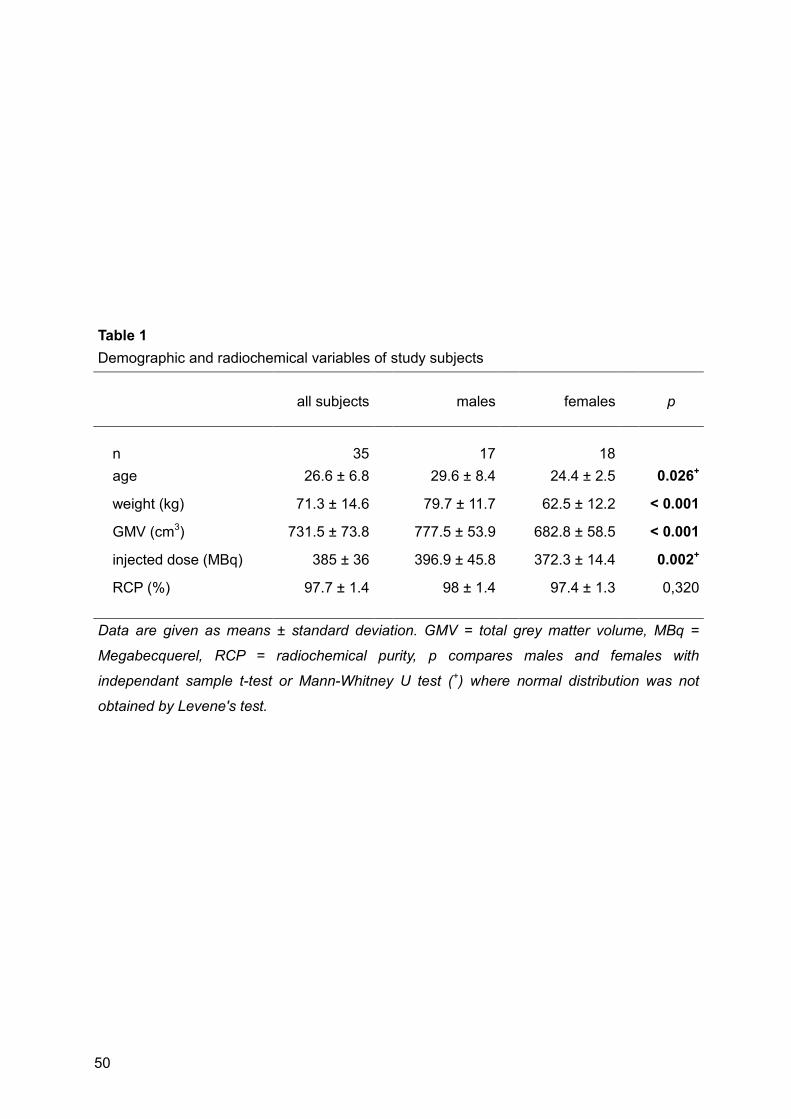
50
Table 1 Demographic and radiochemical variables of study subjects
all subjects
males
females
p
n 35
17
18
age 26.6 ± 6.8
29.6 ± 8.4
24.4 ± 2.5
0.026+
weight (kg) 71.3 ± 14.6
79.7 ± 11.7
62.5 ± 12.2
< 0.001
GMV (cm3) 731.5 ± 73.8
777.5 ± 53.9
682.8 ± 58.5
< 0.001
injected dose (MBq) 385 ± 36
396.9 ± 45.8
372.3 ± 14.4
0.002+
RCP (%) 97.7 ± 1.4
98 ± 1.4
97.4 ± 1.3
0,320
Data are given as means ± standard deviation. GMV = total grey matter volume, MBq =
Megabecquerel, RCP = radiochemical purity, p compares males and females with
independant sample t-test or Mann-Whitney U test (+) where normal distribution was not
obtained by Levene's test.

51
Table 2
Statistical results as obtained by Statistical Parametric Mapping (SPM8).
region
peak
cluster
x y z t z R2 p−FWE p−FDR voxels
positive correlation
right posterior medial temporal
42 −32 0
6.7 5.2
0.487 < 0.001 < 0.001 952
right hippocampus/ parahippocampus
29 −39 0
6.2 4.9
0.503 < 0.001 < 0.001 850
right medial occipital
38 −92 −5
4.9 4.1
0.308 < 0.001 < 0.001 541
right inferior orbitofrontal
51 20 −8
4.9 4.2
0.403 0.002 0.001 434
right posterior inferior temporal
59 −32 −15
5.4 4.5
0.436 0.002 0.002 416
left posterior medial temporal
−44 −26 −6
6.7 5.2
0.386 0.003 0.002 400
right superior parietal
24 −53 48
6.0 4.8
0.449 0.004 0.002 381
left posterior inferior temporal
−53 −42 −14
5.1 4.3
0.302 0.004 0.002 374
left medial occipital
−21 −101 9
4.9 4.2
0.217 0.006 0.002 349
left pericalcarine
−27 −57 5
6.1 4.9
0.424 0.009 0.003 323
left precentral
−33 5 41
5.7 4.7
0.183 0.031 0.01 253
right pericalcarine
30 −57 8
7.2 5.4
0.489 0.043 0.013 234
right inferior occipital
38 −65 −11
5.5 4.5
0.308 0.089 0.026 195
left hippocampus
−24 −27 −14
5.3 4.4
0.428 0.127 0.035 176
negative correlation
left posterior lobe of cerebellum
−26 −57 −23
−6 −4.83
0.422 0.102 0.176 363
left cerebellar crus
−50 −69 −23
−5.4 −4.48
0.461 0.349 0.35 10
Voxel−wise regression analysis results between whole−brain 5−HT1A binding potential (BPND) and whole−brain grey
matter volume (GMV). Stereotactical coordinates (x. y. z) represent cluster peaks in standard Montreal Institute of
Neurology (MNI) space. FWE = family wise error. FDR = false discovery rate. Note that R2 values were calulated
cluster−wise between 5−HT1A BPND and GMV and therefore do not correspond to peak t − or z − values.

52
Table S1
Binding potential and gray matter volume values for 48 regions of interest as quantified with an AAL based atlas (data sorted by GMV/nBPND)
Region BPND nBPND GMV GMV/nBPND
temporal pole, middle temporal gyrus 4,98 0,87 0,47 0,54
temporal pole, superior temporal gyrus 5,26 0,92 0,50 0,54
parahippocampus 5,72 1,00 0,55 0,55
superior frontal, medial 3,48 0,61 0,34 0,56
superior frontal, orbital 3,61 0,63 0,37 0,59
superior parietal 2,96 0,52 0,31 0,59
rectus 4,53 0,79 0,48 0,61
inferior temporal 4,83 0,84 0,52 0,62
superior frontal 3,21 0,56 0,35 0,62
Heschl's gyrus 3,99 0,70 0,43 0,62
orbitofrontal 4,05 0,71 0,44 0,62
insula 4,82 0,84 0,53 0,63
hippocampus 4,73 0,83 0,53 0,64
superior temporal 3,99 0,70 0,45 0,65
supplemantary motor area 3,04 0,53 0,36 0,67
fusiform 4,90 0,86 0,57 0,67
middle temporal 4,32 0,76 0,51 0,68
olfactory sulcus 4,50 0,79 0,54 0,69
paracentral 2,88 0,50 0,35 0,70
medial orbitofrontal 3,47 0,61 0,43 0,70
precuneus 3,16 0,55 0,39 0,71
postcentral 2,70 0,47 0,35 0,73
medial occipital 3,36 0,59 0,43 0,74
inferior frontal, operculum 3,44 0,60 0,45 0,74
inferior orbitofrontal 3,48 0,61 0,45 0,75
inferior occipital 3,46 0,60 0,45 0,75
superior occipital 2,67 0,47 0,35 0,75
amygdala 4,21 0,74 0,55 0,75
midbrain 1,44 0,25 0,19 0,75
supramarginal 3,75 0,66 0,50 0,76
dorsal raphe nucleus 2,33 0,41 0,31 0,76
inferior parietal 3,37 0,59 0,45 0,76
medial frontal 3,36 0,59 0,45 0,77
angular 3,50 0,61 0,49 0,80
cuneus 2,59 0,45 0,36 0,80
precentral 2,49 0,43 0,35 0,81
inferior frontal, triangular 3,09 0,54 0,44 0,81
anterior cingulate 3,79 0,66 0,56 0,85
subgenual cingulate 3,76 0,66 0,56 0,86
medial cingulum 3,17 0,55 0,53 0,96
lingual 2,92 0,51 0,51 1,00
calcarine 2,17 0,38 0,40 1,06

53
posterior cingulate 2,51 0,44 0,49 1,11
Nucleus accumbens 1,45 0,25 0,54 2,11
thalamus 0,70 0,12 0,30 2,46
striatum 0,73 0,13 0,37 2,91
putamen 0,75 0,13 0,42 3,20
caudatus 0,64 0,11 0,54 4,80
ROI = region of interest, AAL = automatic anatomical labelling, BPND = binding potential, GMV = gray
matter volume. BPND and GMV values represent means across both hemispheres. For ROI
regression analysis values from one hemisphere were chosen. nBPND (normalized Binding Potential)
values were rescaled for visualization in Figure 3 by dividing through the highest BPND value (5.72 in
the parahippocampus). To further demonstrate a positive association (hippocampus) and a lack of it
(insula) the regions in bold were selected for ROI regression analyses based on their similarity of
GMV and BPND values.

54
1.12 Second publication: Gray matter and intrinsic network
changes in the posterior cingulate cortex after
selective serotonin reuptake inhibitor intake
Christoph Kraus1,2, Sebastian Ganger1,2, Jan Losak1,2, Andreas Hahn1,2,
Markus Savli1,2, Georg S. Kranz1,2, Pia Baldinger1,2, Christian Windischberger3,4,
Siegfried Kasper1, Rupert Lanzenberger1,2
1 Department of Psychiatry and Psychotherapy,
2 Functional, Molecular and Translational Neuroimaging Lab – PET & MRI,
3 Center for Medical Physics and Biomedical Engineering, 4 MR Centre of Excellence,
Medical University of Vienna, Austria
Published in
NeuroImage, 2014; 84:236-244
[2014, IF: 6.35]
* Correspondence to: Rupert Lanzenberger, Assoc. Prof., MD Functional, Molecular and Translational Neuroimaging Lab
Department of Psychiatry and Psychotherapy Medical University of Vienna Waehringer Guertel 18-20, 1090 Vienna, Austria
Phone (Fax): +43 1 40400 3825 (3099) [email protected]
Running Title: Gray matter and functional changes after SSRI intak

55
ABSTRACT
Preclinical studies have demonstrated that serotonin (5-HT) challenge changes neuronal
circuitries and microarchitecture. However, evidence in human subjects is missing.
Pharmacologic magnetic resonance imaging (phMRI) applying selective 5-HT reuptake
inhibitors (SSRIs) and high-resolution structural and functional brain assessment is able to
demonstrate the impact of 5-HT challenge on neuronal network morphology and functional
activity. To determine how SSRIs induce changes in gray matter and neuronal activity, we
conducted a longitudinal study using citalopram and escitalopram. Seventeen healthy
subjects completed a structural and functional phMRI study with randomized, cross-over,
placebo-controlled, double-blind design. Significant gray matter increases were observed
(amongst other regions) in the posterior cingulate cortex (PCC) and the ventral precuneus
after SSRI intake of 10 days, while decreases were observed within the pre- and postcentral
gyri (all P<0.05, family wise error [FWE] corrected). Furthermore, enhanced resting functional
connectivity (rFC) within the ventral precuneus and PCC was associated with gray matter
increases in the PCC (all FWE Pcorr<0.05). Corroborating these results, whole-brain
connectivity density, measuring the brain’s functional network hubs, was significantly
increased after SSRI-intake in the ventral precuneus and PCC (all FWE Pcorr<0.05). Short-
term administration of SSRIs changes gray matter structures, consistent with previous work
reporting enhancement of neuroplasticity by serotonergic neurotransmission. Furthermore,
increased gray matter in the PCC is associated with increased functional connectivity in one
of the brain’s metabolically most active regions. Our novel findings provide convergent
evidence for dynamic alterations of brain structure and function associated with SSRI
pharmacotherapy.

56
1. INTRODUCTION
Magnetic resonance imaging (MRI) and voxel-based morphometry (VBM) studies in patients
with depression and obsessive-compulsive disorder (OCD) showed gray matter
enhancements after treatment with selective serotonin reuptake inhibitors (SSRIs) (Hoexter
et al., 2012; Smith et al., 2012). Moreover, depressive patients homozygous for the LA-allele
in the SERT gene (rs25531) seem to be more susceptible to gray matter atrophy (Frodl et al.,
2008), this polymorphism also seems to affect gray matter in healthy subjects (Frodl et al.,
2008). Remarkably, at least 3 of the 16 known 5-HT receptors (5-HT1A, 5-HT1B and 5-HT2A)
(Gaspar et al., 2003) and the 5-HT transporter (SERT) (Benninghoff et al., 2012) are involved
in neuroplasticity processes (Gould, 1999; Mogha et al., 2012; Vitalis et al., 2002). Tight links
between the neurotrophin system and 5-HT have previously been shown (Castrén and
Rantamäki, 2010) demonstrating the role of 5-HT in regulating neuronal morphology and
circuitry (Daubert and Condron, 2010).
Selective 5-HT reuptake inhibitors represent the first line medication for depression (Bauer et
al., 2007), anxiety disorder, OCD, post-traumatic stress disorder and eating disorders (Aigner
et al., 2011; Bandelow et al., 2008). At the serotonergic synapse, SSRIs bind to a binding site
at the SERT and block reuptake of 5-HT (Kasper et al., 2009; Stahl, 1998). Treatment with
SSRIs results in rapid 20-fold increase in 5-HT levels within the midbrain raphe nuclei (Tao et
al., 2000), increases 5-HT binding at 5-HT1A autoreceptors there, which subsequently alters
neuronal firing rates and promotes desensitization of 5-HT1A receptors (Stahl, 1998; Zimmer
et al., 2004). The resulting lack of autoinhibition triggers 5-HT release at axon terminals
(Gibbons et al., 2012).
Most of the existing studies using phMRI applied functional MRI and investigated task related
blood oxygen dependent level (BOLD) responses. Evidence from phMRI and VBM, which
addresses the impact of pharmaceuticals on gray matter structure, is scarce. Yet, this
technique is a powerful tool that is able to detect morphological alterations in vivo at high
resolution. Recent work has confirmed gray matter alterations detected by MRI with ex vivo

57
MRI scans as well as post-mortem volumetric analysis (Vernon et al., 2011), and structural
MRI exhibits an excellent test-retest reliability (Wonderlick et al., 2009). In vivo gray matter
changes found by MRI and VBM were validated by post mortem findings (Hornberger et al.,
2012).
Taken together, a convergent line of evidence demonstrates that 5-HT is involved in
development and regulation of gray matter morphology through a series of mechanisms
associated with neuroplasticity. Treatment with SSRIs might thus trigger gray matter
changes, yet confirmation in healthy subjects is missing and the impact of regionally altered
gray matter on neuronal functionality is hardly known. Hence, the aim of this study was to (1)
investigate the influence of 5-HT on gray matter and (2) to elucidate the associated functional
neuronal network changes. This was accomplished by administration of SSRIs to healthy
subjects followed by structural and functional phMRI with quantification of gray matter
changes through VBM and assessment of neuronal networks through resting functional
connectivity (rFC) analyses.
2. MATERIALS AND METHODS
2.1. Subjects
A longitudinal, crossover, double-blind, placebo-controlled study design was used. The study
sample is part of a previously published fMRI study (Windischberger et al., 2010), yet all
analyses were previously not considered. Twenty-four healthy adult subjects were recruited
by advertisement at community boards at the General Hospital in Vienna, four subjects did
not meet inclusion criteria or refused to participate, 20 subjects were randomized, two
subjects dropped out (not related illness, non-compliance) and structural data from one
subject was not available for all three points. Hence, structural MRI datasets were available
from 17 healthy Caucasian subjects (6 female, 11 male 26.5±6.1 years, mean±SD, see Table
1). All subjects provided written informed consent and received reimbursement after

58
participation. All subjects underwent a medical examination at the screening visit that
included medical history, electrocardiogram and routine blood tests. Exclusion criteria were
history of severe disease, any psychiatric (according to assessment by Structured Clinical
Interview for DSM-IV Axis I+II Disorders, SCID I+II) or neurological disorder, drug abuse
including anabolic steroids, psychiatric medication, use of hormonal contraceptives for the
past 6 months, and a positive urine pregnancy test. All subjects were naïve to SSRIs and
psychotropic medication. No particular menstrual phase for scanning of female subjects was
defined. The interventions ended with a final check-up visit for each participant. All study
related procedures were approved by the Ethics Committee of the Medical University of
Vienna.
2.2. Study design and medication
All study subjects received 10 mg escitalopram (S-citalopram), an equivalent dosage of
20 mg citalopram (the 1:1 racemic mixture of R-citalopram and S-citalopram) and placebo, in
randomized order respectively, for 10 days prior to MRI scanning. This period of medication
intake was chosen to reach the plasma steady-state condition (Kasper et al., 2009; Klein et
al., 2007). Study subjects consecutively underwent three MRI scanning sessions (one after
citalopram, escitalopram and placebo) with an average interval of 21.8±13.0 (mean, SD)
between screening visit and MRT1 (no wash-out period), 33.8±6.5 days between MRT1 and
MRT2 and 33.1±4.9 between MRT2 and MRT3. According to the half-lives of citalopram and
escitalopram (Bezchlibnyk-Butler et al., 2000; Rao, 2007), visit intervals have provided
enough time to ensure previous drug/placebo washout. Treatment adherence was
ascertained by announcing control of medication intake through measurements of plasma-
levels at any given time point during study duration. Color-matched dextrose tablets were
used as placebo. In order to blind all study personnel and participants to medication group
assignment, independent pharmacists prepared the medication in accordance with a
computer generated randomization list and each blister was encoded with a unique number

59
to prevent inferences on treatment type and subject. For quantification of plasma levels,
blood samples were taken from each subject approximately 10 min before each fMRI
session. Plasma was frozen at -20 °C and shipped for analysis (Quintiles Analytical Services,
Sweden).
2.3. MRI Measurements and Image Analyses
Structural MRI measurements were performed on 3 Tesla (T) whole−body MEDSPEC S300
MR−scanner (Bruker BioSpin, Ettlingen, Germany) using a standard quadrature single-loop
transmit/receive birdcage head coil at the MR Center of Excellence at the Medical University
of Vienna, Austria. The imaging protocol comprised a magnetization−prepared rapid gradient
echo (MPRAGE, T1−weighted) sequence yielding 128 slices, a 256×256 matrix, at a slice
thickness of 1.56 mm and a voxel size of 0.78×0.86 mm.
Additionally, all study subjects underwent fMRI with a facial expression task (described
below), of which the fMRI results were published previously (Hahn et al., 2009). In the same
MRI session a single-shot gradient-recalled echo planar imaging (GR-EPI) sequence was
applied, optimized for imaging blood oxygen dependent (BOLD) contrast. This EPI-sequence
was done at a TE=31 ms, TR=1000 ms and a matrix size=128×91, which resulted in a total
slab width of 34.5 mm with 10 axial slices of 3 mm thickness aligned to the AC-PC line (0.5
mm slice gap).
2.4. Voxel-based morphometry
In order to test the main hypothesis of this study, which was to detect alterations of gray
matter after 5-HT reuptake inhibition, we used VBM for structural brain assessment. All
analyses of images were performed with statistical parametric mapping (SPM8, Wellcome
Trust Centre for Neuroimaging, Institute of Neurology, University College London, London,

60
United Kingdom, http://www.fil.ion.ucl.ac.uk/spm/software/spm8/) and MATLAB 7.10 (Math
Works, Natick, MA). An optimized VBM protocol was used, applying the DARTEL
(Diffeomorphic Anatomical Registration using Exponentiated Lie algebra) algorithm
(Ashburner, 2007). The images were segmented into gray matter, white matter, and
cerebrospinal fluid (CSF) compartments and successfully passed visually checking for major
artifacts. Subsequently, the gray matter maps obtained by this procedure were separately
normalized to a gray matter template representing the stereotaxic standardized Montreal
Neurological Institute (MNI) space at a voxel size of 1.5 × 1.5 × 1.5 mm. Based on
deformation fields calculated during segmentation, a template was generated by the
DARTEL algorithm. The Jacobian determinants derived from the spatial normalization were
used during nonlinear spatial transformations, which modulated the regional differences of
partitioned gray matter on images from relative to absolute amounts (volume). Unmodulated
images constitute gray matter density (GMD), while modulated images constitute gray matter
volume (GMV). Gray matter images were smoothed with a Gaussian filter of 8 × 8 × 8 mm
full width at half maximum (FWHM).
2.5. Preprocessing Resting Functional Connectivity
Resting functional connectivity was determined using a method to separate network-specific
low frequency BOLD (LF-BOLD) signals from task-evoked BOLD responses (Fair et al.,
2007). Intrinsic connectivity networks are present during task performance (Fox et al., 2006)
so that the obtained resting activity mimics real-life setting, where the brain is usually
continuously engaged in task processing. So gathered intrinsic network activity at rest offers
a reliable alternative to resting-state fMRI at the advantage of controlling for inter- and
intraindividual variations in thought processes and mentation (Shih et al., 2011).
Functional MRI data preprocessing comprised slice-timing correction, realignment, spatial
normalization and spatial smoothing as implemented in SPM8. An in-house, scanner-specific
EPI template was created from previous data by spatial normalization of each individual scan

61
to the SPM template which was followed by averaging across individuals. The advantage of
using an in-house template is that potential local field inhomogeneities of individual scans
match those of the template, which in turn improves the spatial normalization. This template
was used for normalization to MNI-space and a Gaussian smoothing kernel of 8 × 8 × 8 mm
FWHM for spatial smoothing.
Resting functional connectivity was extracted from a blocked facial fMRI paradigm (Hahn et
al., 2009), consisting of 20s baseline blocks, during which a black screen with a white fixation
cross at the center was presented. Baseline blocks were alternated by 20s presentation of
faces as active task blocks, whereby the facial expression and attractiveness had to be
rated. The entire paradigm consisted of 5 alternating baseline and active paradigm blocks
leading to total paradigm duration of 200 s. In accordance with Fair et al (Fair et al., 2007),
the active task periods were removed by cutting out the five 20 s baseline periods. In
consideration of the delay in the hemodynamic response function, cuts were set 5 s after
active and resting blocks, respectively, leading to a time shift of 5 s. This number was used to
maximize the number of frames within steady-state data.
2.6. Resting Functional Connectivity
Because both SSRIs and brain changes underlying VBM findings have previously been
demonstrated to alter brain connectivity (Reetz et al., 2012; van Marle et al., 2011), we were
particularly interested in differences in brain rFC associated with a principal VBM finding
between the SSRI and placebo group. Hence, seed voxel correlation analysis was performed
with resting data extracted from the FEDT fMRI paradigm as described above. Resting data
were corrected as published in an earlier study (Weissenbacher et al., 2009). Linear
regression was applied to correct for changes in white-matter, ventricular and global signal.
Then, data were band-pass filtered by a 12-term finite impulse response (FIR) filter to 0.009-
0.08 Hz with Matlab. Connectivity maps were calculated by cross-correlation between the
BOLD time course from the seed region obtained by VBM (see results section) and the time

62
course from the remaining voxels of the entire brain. For group comparisons, correlation
maps were converted to z-values using Fisher’s r-to-z transformation.
2.7. Resting Functional Connectivity Density
The brains’ resting activity has recently been shown to be subdivided into functional “hubs”
representing network nodes with high functional connectivity (Bullmore and Sporns, 2009),
which differ between their resting activity and anatomical topology (Tomasi and Volkow,
2011). We hypothesized that the brain’s functional hubs are changed themselves by 5-HT
reuptake inhibition. To investigate this question we applied resting functional connectivity
density (rFCD) mapping. This recently developed functional connectivity analysis to define
functional network hubs is a fast voxel-wise data-driven approach sensitive to the number of
local functional connections in brain regions (Tomasi and Volkow, 2010; Tomasi and Volkow,
2011). Technically, functional hubs represent network nodes with a large number of edges as
defined by graph theory. This approach yields FCD, in other words the node degree, and
resembles the cross-correlation function of every voxel. Functional connectivity density has a
‘‘scale-free’’ distribution in the brain (Tomasi and Volkow, 2010; Tomasi and Volkow, 2011),
with few hubs and numerous weakly connected nodes, consistent with the emergence of
scaling in neural networks (He et al., 2010).
To reduce spatial dimensionality for more efficient computation of the BOLD time series,
rFCD maps were downsampled by spline interpolation to a resolution of 4 × 4 × 4 mm. Only
gray matter voxels were further processed using a gray matter mask. White-matter,
ventricular and global signal were regressed-out and the time series were detrended by a 4th
order polynomial function, which was chosen because of optimal results after visually
inspecting the plots of the time-series. For FCD, a connectivity matrix with Pearson
correlation coefficients was generated and a threshold was set at the 1% strongest
correlations in each graph, the remaining voxels were set to zero (Fornito et al., 2012).
Functional connectivity density maps were generated by summing the number of

63
connections within the connectivity matrix for each voxel separately, corresponding to global
FCD as defined previously (Tomasi and Volkow, 2010).
2.8. Statistical Analyses
To decrease the variance in the verum condition, we combined citalopram and escitalopram
groups by averaging the effects within repeated-measure analysis of variance (rmANOVA)
design (i.e., the average of the two verum conditions was equally weighted as the placebo
condition). Three main analyses were performed: one for VBM (1), rFC (2) and rFCD (3),
respectively.
(1) In the main VBM statistical analysis a contrast for gray matter density (GMD) differences
between structural MRI scans after SSRIs (pooled citalopram and escitalopram scans) and
those after placebo was defined by rmANOVA as implemented in SPM8. In this analysis we
controlled for total GMD (tGMD) and sex as two additional factors. Results were corrected
applying the voxel-level family-wise error (FWE) rate at a significance level of Pcorr<0.05.
Only results reaching a cluster-size above 100 voxels are reported. An identical analysis was
performed with gray matter volume (GMV), stronger results are reported.
(2) Proceeding from the major VBM finding, the cluster coordinates were used as seed in
functional connectivity statistics. Seed voxel rmANOVA with rFC maps was performed in
SPM8 and contrasted between SSRI intake (pooled) and placebo condition. Total GMD and
sex were controlled as two additional factors. Results were corrected for false positives using
FWE-correction at a significance level of Pcorr<0.05.
(3) Finally, group differences in rFCD maps from scans after SSRI intake (pooled) and
placebo intake were calculated with rmANOVA in SPM8 and again controlled for sex and
tGMD. Results were corrected for false positives using FWE-correction at a significance level
of Pcorr<0.05.

64
Follow-up Pearson correlations were calculated between principal voxel-based morphometry
(VBM) finding and selective serotonin reuptake inhibitor (SSRI) plasma levels, to test the
relationship between plasma levels and gray matter changes. Furthermore, to investigate a
more global effect of SSRI plasma levels on VBM-results, we performed an identical
rmANOVA controlling for SSRI plasma levels, only. Sex and gray matter density (GMD) were
not added here, because the number of variables roughly should not exceed N/10 to avoid
overfitting.
To test the validity of the extracted resting data and subsequent rFCD analysis, we aimed to
depict major FCD hubs as previously demonstrated (Tomasi and Volkow, 2011). Therefore, in
an intermediate step before SSRI vs. placebo analysis we calculated a one-sample t-test
over rFCD maps of subjects receiving placebo only. Results were corrected with voxel-level
FWE-correction at a significance level of Pcorr<0.05.
Additional statistical tests were used: to rule out cortical atrophy as source of variance and to
investigate global gray matter differences between SSRIs and placebo. Subject gender
differences were calculated with independent sample T-test or Mann-Whitney U test where
appropriate.
We aimed to exclude cortical atrophy due to aging as a potential source of variance. Since
constant covariates across the scans are not part of the general linear model in SPM and
could therefore not be included in the main rmANOVA analyses, the linear effect was
computed ([citalopram+escitalopram]/2-placebo) between the different scanning sessions
resulting in a separate map for every subject. The subsequent one-sample t-test across
these maps is equal to the contrast in the rmANOVA reported above. Hence, the linear effect
maps could be used in a regression analysis with age as independent variable to investigate
the effects of age on VBM. Age effects were corrected using voxel-level FWE-correction at a
significance level of Pcorr<0.05.
Additionally, we aimed to compare global gray matter across treatment with SSRIs or
placebo. Therefore, we performed a repeated-measure analysis of variance (rmANOVA) in

65
SPSS 19.0 comparing total GMD after SSRIs (tGMD=citalopram+escitalopram/2) or placebo
intake using sex as between-subjects variable and age as covariate. A statistical level of
significance was accepted at Puncorr<0.001.
3. RESULTS
Age, body mass index, total brain gray matter, alcohol and cigarette consumption were not
significantly different between males and females (all P>0.1, Table 1). No significant effect of
age was observed between the linear-effect maps of SSRI vs. placebo condition and age (all
FWE Pcorr>0.05. Comparison of total gray matter revealed no significant effect of SSRI
treatment on total brain GMD (Puncorr=0.471).
3.1. Voxel-based morphometry differences between SSRIs and Placebo
The main VBM analysis revealed significant increases of GMD after oral intake of SSRIs
compared to placebo. Gray mater increases were found in the posterior cingulate cortex
(PCC), the ventral precuneus (peak T-values=16.6), the fusiform gyrus, the insula and the
medial superior frontal cortex (all regions bilaterally, 8.1<T<16.6, all FWE Pcorr <0.05; for all
regions see Figure 1 and Table 2). There was no significant correlation between GMD and
SSRI plasma levels (r=0.27, P=0.278) in the PCC cluster. Topologically similar results were
obtained after correcting for SSRI plasma levels (data not shown). However, in the PCC and
precuneus, T-values were attenuated and cluster size decreased (T=6.3, FWE Pcorr=0.012)
Furthermore, significant decreases of GMD after SSRI intake were observed bilaterally in the
precentral gyrus, the cerebellum and the cingulate cortex (8.4<T<13.4, all FWE Pcorr<0.05,
see Table 3). Gray matter density and GMV results were topologically comparable (data not
shown).

66
Figure 1: Increases of gray matter density in 17 healthy subjects after 10 days of SSRI intake
compared to placebo in the bilateral posterior cingulate cortex and adjacent ventral precuneus as
measured with structural MRI and voxel-based morphometry. Color bar represents T-values displayed
at P<0.05 (FWE-corrected). Numbers represent coordinates in MNI standard space at the location of
the crosshair, warm color tones represent increases.
3.2. Seed-voxel rFC differences between SSRIs and Placebo
The PCC is a known hotspot of neuronal activity and exhibited gray matter increases in the
VBM results. Hence, the PCC cluster was chosen for further analysis and subsequent
discussion. The PCC result from VBM served as seed region (VBM peak MNI: x,y,z = 8, -45,
38; 3261 voxels) for calculating differences in rFC between the SSRI and placebo group. In
the voxel-wise rFC analysis, enhanced connectivity from the PCC seed region was observed
after SSRI intake compared to placebo in the bilateral PCC spreading to the ventral
precuneus (T=5.7, FWE Pcorr<0.05, Figure 2A). The resulting rFC cluster in the PCC/ventral
precuneus was located overlapping and caudally adjacent to the VBM seed region (Figure
2C). No other regions exhibiting significant positive or negative rFC alterations. Applying less
stringent correction, increased rFC was obtained in the cuneus (Puncorr<0.001, T=3.6, 40
voxels).
3.3. FCD connectivity hub changes under SSRI administration
The extraction of resting activity and subsequent rFCD analysis at baseline levels of block
design fMRI data in subjects after intake of placebo yielded typical rFCD hubs as reported

67
Figure 2: Increased resting functional connectivity (rFC) and resting functional connectivity density
(rFCD) associated with increased gray matter (VBM) in the PCC after SSRI intake compared to
placebo. (A) Increased seed-voxel rFC associated with increased gray matter in the PCC (VBM peak
MNI: x,y,z = 8, -45, -19; 3261 voxels) to gray matter within small parts of the PCC and caudally
adjacent in gray matter of the ventral precuneus. (B) Increased rFCD in subjects after SSRI intake
compared to placebo in the PCC and ventral precuneus. (C) Triple overlay indicating close proximity of
increased gray matter, rFC and rFCD in the PCC and ventral precuneus, known to represent one of
the brain’s metabolically most active region (Gusnard and Raichle, 2001). Blue color bars represent
rFC T-values at P<0.05 (FWE-corrected), displayed at Puncorr<0.001. Purple color bars represent rFCD
T-values at P<0.05 (FWE-corrected), displayed at Puncorr<0.001. Spectral color bar represents VBM T-
values displayed at P<0.05 (FWE-corrected). Numbers represent coordinates in MNI standard space
at the location of the crosshair.
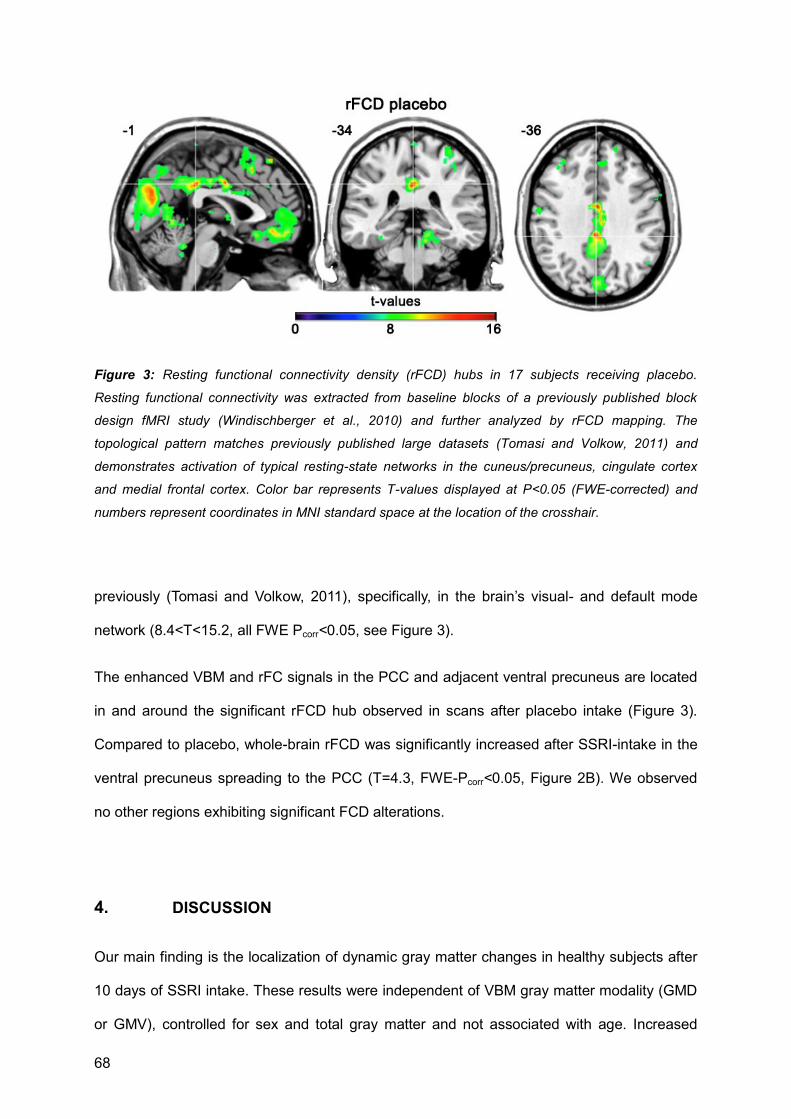
68
Figure 3: Resting functional connectivity density (rFCD) hubs in 17 subjects receiving placebo.
Resting functional connectivity was extracted from baseline blocks of a previously published block
design fMRI study (Windischberger et al., 2010) and further analyzed by rFCD mapping. The
topological pattern matches previously published large datasets (Tomasi and Volkow, 2011) and
demonstrates activation of typical resting-state networks in the cuneus/precuneus, cingulate cortex
and medial frontal cortex. Color bar represents T-values displayed at P<0.05 (FWE-corrected) and
numbers represent coordinates in MNI standard space at the location of the crosshair.
previously (Tomasi and Volkow, 2011), specifically, in the brain’s visual- and default mode
network (8.4<T<15.2, all FWE Pcorr<0.05, see Figure 3).
The enhanced VBM and rFC signals in the PCC and adjacent ventral precuneus are located
in and around the significant rFCD hub observed in scans after placebo intake (Figure 3).
Compared to placebo, whole-brain rFCD was significantly increased after SSRI-intake in the
ventral precuneus spreading to the PCC (T=4.3, FWE-Pcorr<0.05, Figure 2B). We observed
no other regions exhibiting significant FCD alterations.
4. DISCUSSION
Our main finding is the localization of dynamic gray matter changes in healthy subjects after
10 days of SSRI intake. These results were independent of VBM gray matter modality (GMD
or GMV), controlled for sex and total gray matter and not associated with age. Increased

69
gray matter did not correlate with SSRI plasma levels, statistical significance was, however
attenuated upon correcting for plasma levels. Total gray matter was not significantly different
between treatment groups. Furthermore, we observed increased functional connectivity after
SSRI intake associated with increased gray matter in the PCC within the PCC and to
adjacent gray matter in the ventral precuneus. Connectivity density, a measure for the brain’s
functional network hubs, was increased after SSRI intake in a topologically identical region in
the PCC/ventral precuneus. Taken together, our results provide consistent evidence that
SSRI intake is associated with changes of gray matter and neuronal functionality as
measured by phMRI.
Our findings are striking for two apparent reasons. First, although differently calculated, both
increased rFCD and rFC spatially overlap in the PCC/ventral precuneus. Enhanced
functional connectivity seeded from increased PCC gray matter was validated by whole brain
connectivity density, which resembles whole brain correlational strength of LF-BOLD signals
without the need of seeds. Second, these brain structures exhibit large neuronal densities
and are amongst the brain’s metabolically most active (Gusnard and Raichle, 2001). Peak
values in blood flow, metabolic activity and oxygen turnover are located there, which all might
impact changes of structural and functional MRI signals.
We observed several regions with significant gray matter increases, such as the fusiform
gyrus, the insula, and the frontal cortex, and the posterior cingulate cortex where
serotonergic neurotransmission might impact on structural plasticity as measured with MRI.
Furthermore, several decreases were found, such as in the pre- and postcentral gyri. The
principal neurobiological mechanisms effecting volumetric changes in the living brain as
reported by numerous MRI studies are currently being investigated with large effort. The
findings revealed by this study appear very interesting because they indicate a role of 5-HT in
changing gray matter and functional neuronal activity. Decreased metabolism was
discovered at one week and reversal towards increased metabolism after 6 weeks of
treatment in the posterior cingulate cortex, hippocampus, insula, putamen as well as in

70
temporal and prefrontal cortices (Mayberg et al., 2000). This was explained by the authors to
be caused by receptor downregulation or changes in second messenger systems and further
undermines the notion that SSRIs impact on energy metabolism (Webhofer et al., 2011).
Furthermore, response dependent differences in regional blood flow were demonstrated in
depressive patients in the PCC (Joe et al., 2006). Beyond this, previous evidence from
animal research enables a brief discussion on 5-HT mediated neuroplasticity. Hypothetically,
SSRI-dependent altered plasticity might occur due to binding at serotonergic receptors,
which are linked to second messenger systems effecting restructuration. Candidate
molecules potentially mediating such remodeling in the PCC and ventral precuneus are 5-
HT1A, 5-HT1B and 5-HT2A receptor (Savli et al., 2012) and cross-links to the neurotrophin
system with prominent targets such as BDNF and CREB. Unfortunately, MRI data at the
current spatial resolution do not allow inference on the kind of neuronal reactivity or the type
of cellular remodeling that cause gray matter signal changes. A number of factors such as
altered neuronal or glial reactivity (Lesch and Waider, 2012), liquor circulation, blood flow and
angiogenesis (Seevinck et al., 2010) might likewise account for brain volume changes and
subsequently altered neuronal functionality.
Our data show that changes of regional gray matter MRI signals are also associated with
altered functional activity. Higher amount of gray matter in the PCC was associated with
increased resting activity in the PCC itself and the adjacent ventral precuneus. This region is
a key node of the default mode network and differentially active during learning, memory,
reward and task engagement (for review see (Pearson et al., 2011)). We demonstrated that
the brain’s functional connectivity in our dataset is compartmentalized in resting network
hubs at locations in line with earlier studies (Tomasi and Volkow, 2010; Tomasi and Volkow,
2011). The anatomical localization and functional activation of these hubs is consistently
reported (Gusnard and Raichle, 2001; Tomasi and Volkow, 2010; Tomasi and Volkow, 2011),
though, according to our study, SSRIs might alter the activity of such hubs. In consideration
of the fact that multiple imaging modalities detected increases in gray matter, altered
functional connectivity and hub activity in the PCC/ventral precuneus, this study provides

71
robust evidence that SSRIs interfere with a broad cascade of mechanisms affecting several
physiological brain systems. Moreover, these data are indicative that functional changes
occur in concert with structural alterations, even after short treatment periods. However, our
study design does not allow inference on the length of time the observed network changes
last. Previous reports indicate that structural modifications usually take more time than
changes in functionality, yet demand further research on the structure-function relationship of
brain networks (Bullmore and Sporns, 2009), which emphasizes the relevance of our
findings.
We are not aware of a similar study in healthy subjects investigating the impacts of SSRIs on
neuronal structure and function by combining structural and functional MRI. Increases of gray
matter have been recently published in studies using phMRI in psychiatric patients
undergoing SSRI therapy. A coinciding longitudinal study using the DARTEL algorithm shows
increases of gray matter in the dorsolateral prefrontal cortex after intake of sertraline (Smith
et al., 2012). Yet herein the authors investigated depressed patients over a time course of 12
weeks. Moreover, gray matter increases were reported for the left putamen after 12 week
intake of fluoxetine in patients with obsessive compulsive disorder (Hoexter et al., 2012).
Another coinciding study investigated patients with posttraumatic stress disorder and found
hippocampal volume increases after 9-12 months continuous treatment with paroxetine
(Vermetten et al., 2003). Gray matter increases after SSRI intake are further supported by a
study in first-episode drug-naïve depressive disorder and panic disorder, detailing moderate
increases in subcortical structures such as the nucleus accumbens, the putamen and the
hippocampus (Lai and Wu, 2011). On the other side, a cross-sectional study with patients
suffering from depression and anxiety disorders did not reveal a protecting effect of stable
SSRI treatment on gray matter reductions observed in these disorders (van Tol et al., 2010).
In consideration of neuroplastic effects of 5-HT, the observed decreases of gray matter in our
study are challenging to interpret, but potentially the same mechanisms mediating gray
matter increases could be involved. Only one study reported gray matter decreases, located
in the superior temporal cortex in patients with social anxiety disorder after a 12 week-long

72
intake of 20 mg escitalopram (Cassimjee et al., 2010). Taken together, different patient
collectives drug types and duration in relation to our study impairs comparability.
Until now, rFC alterations upon SSRI challenge were only scarcely investigated. A study in
healthy subjects identified decreased functional coupling between the amygdala and the
ventral medial prefrontal cortex (McCabe and Mishor, 2011). Enhanced functional coupling
was reported between the amygdala and the anterior insula after duloxetine intake (van
Marle et al., 2011), here 19 healthy subjects were analyzed. VBM-based seed regions were
not considered in either of these studies. Noteably, rFC and rFCD changes after SSRI intake
were regionally restricted to intrinsic areas around increased gray matter in the PCC.
Cognitive load adherent in blocked design task activation was previously located at the PCC
(Newton et al., 2011). Furthermore, transient BOLD responses at block transitions occur at
the PCC and many other regions (Fox et al., 2005), yet inserted intervals accounting for
delay in the hemodynamic response function limit these transitions. Variance from cognitive
load and transitions might theoretically spill into the analyzed baseline blocks, which should
be taken into account upon comparison of our data with traditionally obtained resting-state
data. Hence, it remains intriguing that whole-brain connectivity density analysis, which is not
related to seed-based connectivity, identifies the same region as connectivity from increased
gray matter signal. The mechanisms that SSRIs interfere with in this region are therefore
likely to be associated with several factors altering substrates detectable both in T1-weighted
as well as EPI MRI sequences. The existing gap between underlying molecular mechanisms
and alterations of voxel-intensity values are vigorously debated (for critical review and
comments see (Draganski and Kherif, 2013; Erickson, 2013; Fields, 2013; Thomas and
Baker, 2013), so that strong gray matter changes as shown in this study emphasize the need
for more translational work on molecular players mediating in vivo structural and functional
changes of neuronal networks as measured by MRI.
The following study limitations must be reported. Though we analyzed a rather low subject
number, sample sizes of active groups within previous studies have been even lower than in

73
our study (Anand et al., 2005; McCabe and Mishor, 2011). Therefore, this factor, though
indeed a limitation, remains a common feature in many pharmacologic neuroimaging studies.
In addition, subjects were not balanced according to sex, this issue was however addressed
by including sex as nuisance variable.
In summary, we found that study subjects after SSRI intake exhibited significant gray matter
changes. Moreover, almost identical locations of increased resting functional connectivity and
connectivity density associated with gray matter increases in the PCC provide evidence for
the involvement of SSRIs in multiple mechanisms changing brain structure and functionality.
When taken together, these results point towards plastic changes of brain structure and
function as neuronal substrate of effects associated with SSRI intake and offer a paradigm
for further exploration of these mechanisms in psychiatric patients.
5. ACKNOWLEDGEMENTS
Data have been measured within a project funded by an investigator-initiated and
unrestricted research grant from H. Lundbeck A/S, Denmark to S. Kasper. The sponsors and
funders did not participate in the design and conduct of the study and were not involved in
the preparation, review, or approval of the manuscript. The study protocol has been planned
by the authors who retained full academic control. In the study presented here we applied
new data analysis approaches in structural and functional magnetic resonance imaging
recently available beyond the scope of a study already published (Windischberger et al.,
2010). The work of C. Kraus has been funded by an intramural grant of the research cluster
between the Medical University of Vienna and the University of Vienna (FA103FC001) to R.
Lanzenberger and C. Lamm. A. Hahn was funded by a DOC fellowship of the Austrian
Academy of Sciences at the Department of Psychiatry and Psychotherapy. The authors are
grateful to C. Spindelegger, U. Moser, P. Stein, M. Fink, L. Pezawas, A. Erfurth, and M. Willeit
for their medical support, and to A. Holik, S. Friedreich, F. Gerstl, and E. Moser for technical

74
support. We thank M. Spies for native English editing. The study is part of C. Kraus’ thesis
“Serotonin and Neuroplasticity” supervised by R. Lanzenberger in the Clinical Neurosciences
PhD program at the Medical University of Vienna, Austria. Parts of this study have been or
will be presented by P. Baldinger at the 19th European Congress of Psychiatry (EPA), March
12-15, 2011, Vienna, Austria, by M. Savli at the 24thEuropean College of
Neuropsychopharmacology (ECNP) Congress, September 3-7, 2011, Paris, France, and by
C. Kraus at the 11th World Congress of Biological Psychiatry (WFSBP), June 23-27, 2013,
Kyoto, Japan.
6. CONFLICT OF INTEREST
Without any relevance to this work, S. Kasper declares that he has received grant/research
support from Eli Lilly, Lundbeck A/S, Bristol-Myers Squibb, Servier, Sepracor,
GlaxoSmithKline, Organon, and has served as a consultant or on advisory boards for
AstraZeneca, Austrian Sick Found, Bristol-Myers Squibb, GlaxoSmithKline, Eli Lily, Lundbeck
A/S, Pfizer, Organon, Sepracor, Janssen, and Novartis, and has served on speakers’
bureaus for AstraZeneca, Eli Lilly, Lundbeck A/S, Servier, Sepracor and Janssen. R.
Lanzenberger received travel grants and conference speaker honoraria from AstraZeneca
and Lundbeck A/S.

75
REFERENCES Aigner, M., Treasure, J., Kaye, W., Kasper, S., Disorders, W.T.F.O.E., 2011. World Federation of Societies of Biological Psychiatry (WFSBP) guidelines for the pharmacological treatment of eating disorders. World J Biol Psychiatry 12, 400-443.
Anand, A., Li, Y., Wang, Y., Wu, J., Gao, S., Bukhari, L., Mathews, V.P., Kalnin, A., Lowe, M.J., 2005. Antidepressant effect on connectivity of the mood-regulating circuit: an FMRI study. Neuropsychopharmacology 30, 1334-1344.
Ashburner, J., 2007. A fast diffeomorphic image registration algorithm. NeuroImage 38, 95-113.
Bandelow, B., Zohar, J., Hollander, E., Kasper, S., Moller, H.J., Wfsbp Task Force on Treatment Guidelines for Anxiety, O.-C., Post-Traumatic Stress, D., Allgulander, C., Ayuso-Gutierrez, J., Baldwin, D.S., Buenvicius, R., Cassano, G., Fineberg, N., Gabriels, L., Hindmarch, I., Kaiya, H., Klein, D.F., Lader, M., Lecrubier, Y., Lepine, J.P., Liebowitz, M.R., Lopez-Ibor, J.J., Marazziti, D., Miguel, E.C., Oh, K.S., Preter, M., Rupprecht, R., Sato, M., Starcevic, V., Stein, D.J., van Ameringen, M., Vega, J., 2008. World Federation of Societies of Biological Psychiatry (WFSBP) guidelines for the pharmacological treatment of anxiety, obsessive-compulsive and post-traumatic stress disorders - first revision. World J Biol Psychiatry 9, 248-312.
Bauer, M., Bschor, T., Pfennig, A., Whybrow, P.C., Angst, J., Versiani, M., Möller, H.-J., Disorders, W.T.F.o.U.D., 2007. World Federation of Societies of Biological Psychiatry (WFSBP) Guidelines for Biological Treatment of Unipolar Depressive Disorders in Primary Care. World J Biol Psychiatry 8, 67-104.
Benninghoff, J., van der Ven, A., Schloesser, R.J., Moessner, R., Moller, H.J., Rujescu, D., 2012. The complex role of the serotonin transporter in adult neurogenesis and neuroplasticity. A critical review. World J Biol Psychiatry 13, 240-247.
Bezchlibnyk-Butler, K., Aleksic, I., Kennedy, S.H., 2000. Citalopram--a review of pharmacological and clinical effects. J Psychiatry Neurosci 25, 241-254.
Bullmore, E., Sporns, O., 2009. Complex brain networks: graph theoretical analysis of structural and functional systems. Nat Rev Neurosci 10, 186-198.
Cassimjee, N., Fouche, J.-P., Burnett, M., Lochner, C., Warwick, J., Dupont, P., Stein, D.J., Cloete, K.J., Carey, P.D., 2010. Changes in regional brain volumes in social anxiety disorder following 12 weeks of treatment with escitalopram. Metab Brain Dis 25, 369-374.
Castrén, E., Rantamäki, T., 2010. The role of BDNF and its receptors in depression and antidepressant drug action: Reactivation of developmental plasticity. Dev Neurobiol 70, 289-297.
Daubert, E.A., Condron, B.G., 2010. Serotonin: a regulator of neuronal morphology and circuitry. Trends Neurosci 33, 424-434.
Draganski, B., Kherif, F., 2013. In vivo assessment of use-dependent brain plasticity-Beyond the "one trick pony" imaging strategy. NeuroImage 73, 255-259.
Erickson, K.I., 2013. Evidence for structural plasticity in humans: Comment on Thomas and Baker (2012). NeuroImage 73, 237-238.
Fair, D.A., Schlaggar, B.L., Cohen, A.L., Miezin, F.M., Dosenbach, N.U., Wenger, K.K., Fox, M.D., Snyder, A.Z., Raichle, M.E., Petersen, S.E., 2007. A method for using blocked and

76
event-related fMRI data to study "resting state" functional connectivity. NeuroImage 35, 396-405.
Fields, R.D., 2013. Changes in brain structure during learning: Fact or artifact? Reply to Thomas and Baker. NeuroImage 73, 260-264.
Fornito, A., Zalesky, A., Pantelis, C., Bullmore, E.T., 2012. Schizophrenia, neuroimaging and connectomics. NeuroImage 62, 2296-2314.
Fox, M.D., Snyder, A.Z., Barch, D.M., Gusnard, D.A., Raichle, M.E., 2005. Transient BOLD responses at block transitions. NeuroImage 28, 956-966.
Fox, M.D., Snyder, A.Z., Zacks, J.M., Raichle, M.E., 2006. Coherent spontaneous activity accounts for trial-to-trial variability in human evoked brain responses. Nat Neurosci 9, 23-25.
Frodl, T., Koutsouleris, N., Bottlender, R., Born, C., Jäger, M., Mörgenthaler, M., Scheuerecker, J., Zill, P., Baghai, T., Schüle, C., Rupprecht, R., Bondy, B., Reiser, M., Möller, H.-J., Meisenzahl, E.M., 2008. Reduced gray matter brain volumes are associated with variants of the serotonin transporter gene in major depression. Mol Psychiatry 13, 1093-1101.
Gaspar, P., Cases, O., Maroteaux, L., 2003. The developmental role of serotonin: news from mouse molecular genetics. Nat Rev Neurosci 4, 1002-1012.
Gibbons, R.D., Hur, K., Brown, C.H., Davis, J.M., Mann, J.J., 2012. Benefits from antidepressants: synthesis of 6-week patient-level outcomes from double-blind placebo-controlled randomized trials of fluoxetine and venlafaxine. Arch Gen Psychiatry 69, 572-579.
Gould, E., 1999. Serotonin and hippocampal neurogenesis. Neuropsychopharmacology 21, 46S-51S.
Gusnard, D.A., Raichle, M.E., 2001. Searching for a baseline: functional imaging and the resting human brain. Nat Rev Neurosci 2, 685-694.
Hahn, A., Holik, A., Gerstl, F., Savli, M., Stein, P., Akimova, E., Angleitner, P., Windischberger, C., Kasper, S., Lanzenberger, R., 2009. Anxiety Scores are Related to Amygdala Activity Induced by Facial Attractiveness and Emotional Expressions. NeuroImage 47 48S.
He, B.J., Zempel, J.M., Snyder, A.Z., Raichle, M.E., 2010. The temporal structures and functional significance of scale-free brain activity. Neuron 66, 353-369.
Hoexter, M.Q., de Souza Duran, F.L., D'Alcante, C.C., Dougherty, D.D., Shavitt, R.G., Lopes, A.C., Diniz, J.B., Deckersbach, T., Batistuzzo, M.C., Bressan, R.A., Miguel, E.C., Busatto, G.F., 2012. Gray matter volumes in obsessive-compulsive disorder before and after fluoxetine or cognitive-behavior therapy: a randomized clinical trial. Neuropsychopharmacology 37, 734-745.
Hornberger, M., Wong, S., Tan, R., Irish, M., Piguet, O., Kril, J., Hodges, J.R., Halliday, G., 2012. In vivo and post-mortem memory circuit integrity in frontotemporal dementia and Alzheimer's disease. Brain 135, 3015-3025.
Joe, A.Y., Tielmann, T., Bucerius, J., Reinhardt, M.J., Palmedo, H., Maier, W., Biersack, H.J., Zobel, A., 2006. Response-dependent differences in regional cerebral blood flow changes with citalopram in treatment of major depression. J Nucl Med 47, 1319-1325.
Kasper, S., Sacher, J., Klein, N., Mossaheb, N., Attarbaschi-Steiner, T., Lanzenberger, R., Spindelegger, C., Asenbaum, S., Holik, A., Dudczak, R., 2009. Differences in the dynamics of

77
serotonin reuptake transporter occupancy may explain superior clinical efficacy of escitalopram versus citalopram. Int Clin Psychopharmacol 24, 119-125.
Klein, N., Sacher, J., Geiss-Granadia, T., Mossaheb, N., Attarbaschi, T., Lanzenberger, R., Spindelegger, C., Holik, A., Asenbaum, S., Dudczak, R., Tauscher, J., Kasper, S., 2007. Higher serotonin transporter occupancy after multiple dose administration of escitalopram compared to citalopram: an [123I]ADAM SPECT study. Psychopharmacology (Berl) 191, 333-339.
Lai, C.H., Wu, Y.T., 2011. Duloxetine's modest short-term influences in subcortical structures of first episode drug-naive patients with major depressive disorder and panic disorder. Psychiatry Res 194, 157-162.
Lesch, K.P., Waider, J., 2012. Serotonin in the modulation of neural plasticity and networks: implications for neurodevelopmental disorders. Neuron 76, 175-191.
Mayberg, H.S., Brannan, S.K., Tekell, J.L., Silva, J.A., Mahurin, R.K., McGinnis, S., Jerabek, P.A., 2000. Regional metabolic effects of fluoxetine in major depression: serial changes and relationship to clinical response. Biol Psychiatry 48, 830-843.
McCabe, C., Mishor, Z., 2011. Antidepressant medications reduce subcortical-cortical resting-state functional connectivity in healthy volunteers. NeuroImage 57, 1317-1323.
Mogha, A., Guariglia, S.R., Debata, P.R., Wen, G.Y., Banerjee, P., 2012. Serotonin 1A receptor-mediated signaling through ERK and PKCalpha is essential for normal synaptogenesis in neonatal mouse hippocampus. Transl Psychiatry 2, e66.
Newton, A.T., Morgan, V.L., Rogers, B.P., Gore, J.C., 2011. Modulation of steady state functional connectivity in the default mode and working memory networks by cognitive load. Hum Brain Mapp 32, 1649-1659.
Pearson, J.M., Heilbronner, S.R., Barack, D.L., Hayden, B.Y., Platt, M.L., 2011. Posterior cingulate cortex: adapting behavior to a changing world. Trends Cogn Sci 15, 143-151.
Rao, N., 2007. The clinical pharmacokinetics of escitalopram. Clin Pharmacokinet 46, 281-290.
Reetz, K., Dogan, I., Rolfs, A., Binkofski, F., Schulz, J.B., Laird, A.R., Fox, P.T., Eickhoff, S.B., 2012. Investigating function and connectivity of morphometric findings--exemplified on cerebellar atrophy in spinocerebellar ataxia 17 (SCA17). NeuroImage 62, 1354-1366.
Savli, M., Bauer, A., Mitterhauser, M., Ding, Y.S., Hahn, A., Kroll, T., Neumeister, A., Haeusler, D., Ungersboeck, J., Henry, S., Isfahani, S.A., Rattay, F., Wadsak, W., Kasper, S., Lanzenberger, R., 2012. Normative database of the serotonergic system in healthy subjects using multi-tracer PET. NeuroImage 63, 447-459.
Seevinck, P.R., Deddens, L.H., Dijkhuizen, R.M., 2010. Magnetic resonance imaging of brain angiogenesis after stroke. Angiogenesis 13, 101-111.
Shih, P., Keehn, B., Oram, J.K., Leyden, K.M., Keown, C.L., Muller, R.A., 2011. Functional differentiation of posterior superior temporal sulcus in autism: a functional connectivity magnetic resonance imaging study. Biol Psychiatry 70, 270-277.
Smith, R., Chen, K., Baxter, L., Fort, C., Lane, R.D., 2012. Antidepressant effects of sertraline associated with volume increases in dorsolateral prefrontal cortex. J Affect Disord.

78
Stahl, S.M., 1998. Mechanism of action of serotonin selective reuptake inhibitors. Serotonin receptors and pathways mediate therapeutic effects and side effects. J Affect Disord 51, 215-235.
Tao, R., Ma, Z., Auerbach, S.B., 2000. Differential effect of local infusion of serotonin reuptake inhibitors in the raphe versus forebrain and the role of depolarization-induced release in increased extracellular serotonin. J Pharmacol Exp Ther 294, 571-579.
Thomas, C., Baker, C.I., 2013. Teaching an adult brain new tricks: A critical review of evidence for training-dependent structural plasticity in humans. NeuroImage 73, 225-236.
Tomasi, D., Volkow, N.D., 2010. Functional connectivity density mapping. Proc Natl Acad Sci U S A 107, 9885-9890.
Tomasi, D., Volkow, N.D., 2011. Association between Functional Connectivity Hubs and Brain Networks. Cereb Cortex, 1-11.
van Marle, H.J., Tendolkar, I., Urner, M., Verkes, R.J., Fernandez, G., van Wingen, G., 2011. Subchronic duloxetine administration alters the extended amygdala circuitry in healthy individuals. NeuroImage 55, 825-831.
van Tol, M.-J., van der Wee, N.J.A., van den Heuvel, O.A., Nielen, M.M.A., Demenescu, L.R., Aleman, A., Renken, R., van Buchem, M.A., Zitman, F.G., Veltman, D.J., 2010. Regional brain volume in depression and anxiety disorders. Arch Gen Psychiatry 67, 1002-1011.
Vermetten, E., Vythilingam, M., Southwick, S.M., Charney, D.S., Bremner, J.D., 2003. Long-term treatment with paroxetine increases verbal declarative memory and hippocampal volume in posttraumatic stress disorder. Biol Psychiatry 54, 693-702.
Vernon, A.C., Natesan, S., Modo, M., Kapur, S., 2011. Effect of chronic antipsychotic treatment on brain structure: a serial magnetic resonance imaging study with ex vivo and postmortem confirmation. Biol Psychiatry 69, 936-944.
Vitalis, T., Cases, O., Gillies, K., Hanoun, N., Hamon, M., Seif, I., Gaspar, P., Kind, P., Price, D.J., 2002. Interactions between TrkB signaling and serotonin excess in the developing murine somatosensory cortex: a role in tangential and radial organization of thalamocortical axons. J Neurosci 22, 4987-5000.
Weissenbacher, A., Kasess, C., Gerstl, F., Lanzenberger, R., Moser, E., Windischberger, C., 2009. Correlations and anticorrelations in resting-state functional connectivity MRI: a quantitative comparison of preprocessing strategies. NeuroImage 47, 1408-1416.
Windischberger, C., Lanzenberger, R., Holik, A., Spindelegger, C., Stein, P., Moser, U., Gerstl, F., Fink, M., Moser, E., Kasper, S., 2010. Area-specific modulation of neural activation comparing escitalopram and citalopram revealed by pharmaco-fMRI: a randomized cross-over study. NeuroImage 49, 1161-1170.
Wonderlick, J.S., Ziegler, D.A., Hosseini-Varnamkhasti, P., Locascio, J.J., Bakkour, A., van der Kouwe, A., Triantafyllou, C., Corkin, S., Dickerson, B.C., 2009. Reliability of MRI-derived cortical and subcortical morphometric measures: effects of pulse sequence, voxel geometry, and parallel imaging. NeuroImage 44, 1324-1333.
Zimmer, L., Riad, M., Rbah, L., Belkacem-Kahlouli, A., Le Bars, D., Renaud, B., Descarries, L., 2004. Toward brain imaging of serotonin 5-HT1A autoreceptor internalization. NeuroImage 22, 1421-1426.
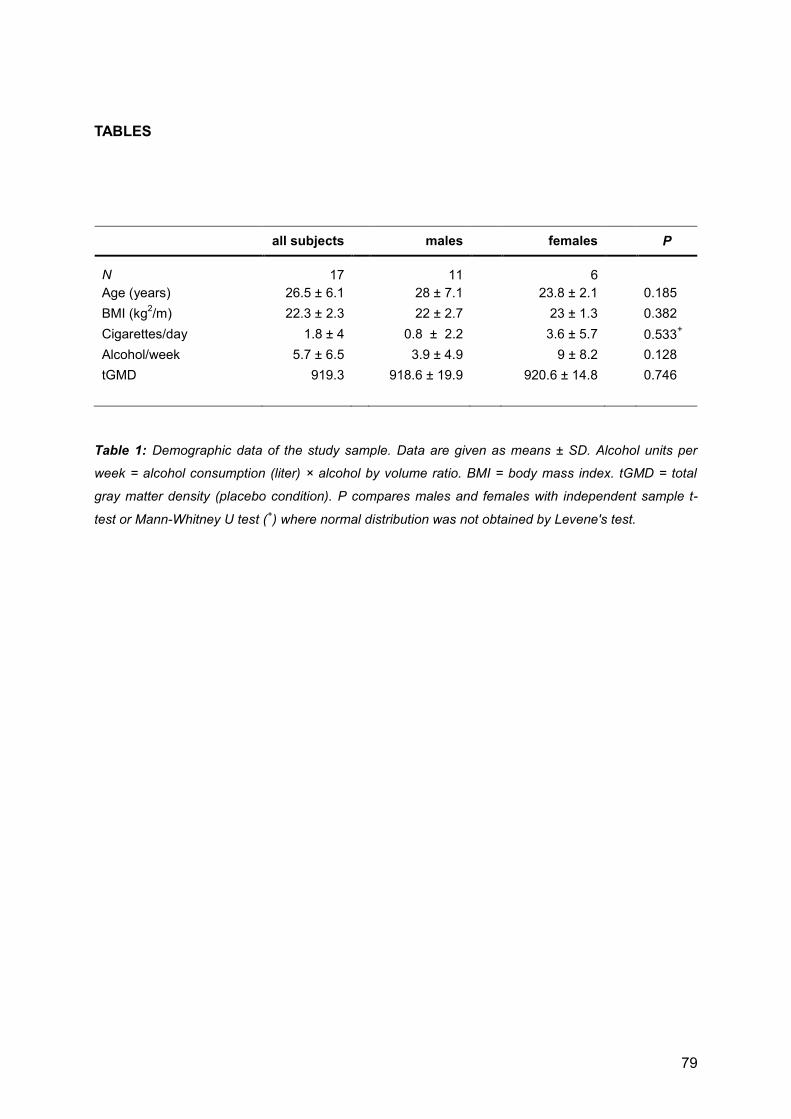
79
TABLES
all subjects
males
females
P
N 17
11
6
Age (years) 26.5 ± 6.1
28 ± 7.1
23.8 ± 2.1
0.185
BMI (kg2/m) 22.3 ± 2.3
22 ± 2.7
23 ± 1.3
0.382
Cigarettes/day 1.8 ± 4
0.8 ± 2.2
3.6 ± 5.7
0.533+
Alcohol/week 5.7 ± 6.5
3.9 ± 4.9
9 ± 8.2
0.128
tGMD 919.3
918.6 ± 19.9
920.6 ± 14.8
0.746
Table 1: Demographic data of the study sample. Data are given as means ± SD. Alcohol units per
week = alcohol consumption (liter) × alcohol by volume ratio. BMI = body mass index. tGMD = total
gray matter density (placebo condition). P compares males and females with independent sample t-
test or Mann-Whitney U test (+) where normal distribution was not obtained by Levene's test.
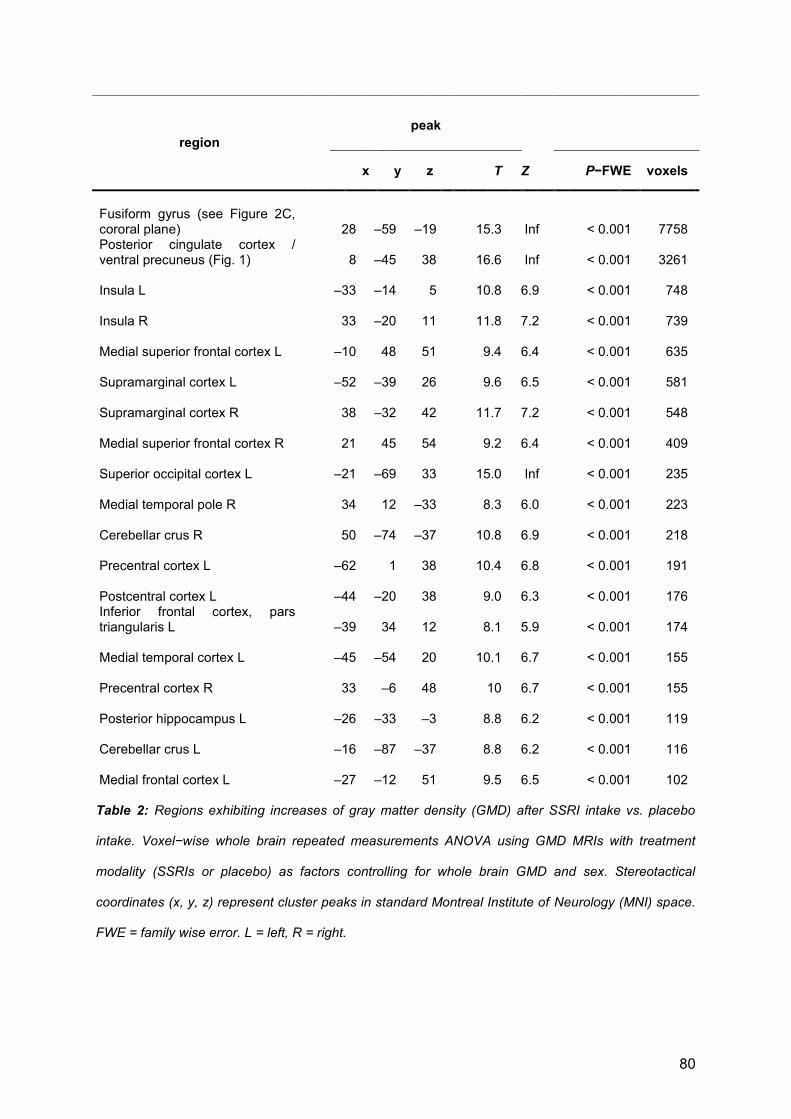
80
Table 2: Regions exhibiting increases of gray matter density (GMD) after SSRI intake vs. placebo
intake. Voxel−wise whole brain repeated measurements ANOVA using GMD MRIs with treatment
modality (SSRIs or placebo) as factors controlling for whole brain GMD and sex. Stereotactical
coordinates (x, y, z) represent cluster peaks in standard Montreal Institute of Neurology (MNI) space.
FWE = family wise error. L = left, R = right.
region
peak
x y z
T Z P−FWE voxels
Fusiform gyrus (see Figure 2C, cororal plane)
28 –59 –19
15.3 Inf
< 0.001 7758
Posterior cingulate cortex / ventral precuneus (Fig. 1)
8 –45 38
16.6 Inf
< 0.001 3261
Insula L
–33 –14 5
10.8 6.9
< 0.001 748
Insula R
33 –20 11
11.8 7.2
< 0.001 739
Medial superior frontal cortex L
–10 48 51
9.4 6.4
< 0.001 635
Supramarginal cortex L
–52 –39 26
9.6 6.5
< 0.001 581
Supramarginal cortex R
38 –32 42
11.7 7.2
< 0.001 548
Medial superior frontal cortex R
21 45 54
9.2 6.4
< 0.001 409
Superior occipital cortex L
–21 –69 33
15.0 Inf
< 0.001 235
Medial temporal pole R
34 12 –33
8.3 6.0
< 0.001 223
Cerebellar crus R
50 –74 –37
10.8 6.9
< 0.001 218
Precentral cortex L
–62 1 38
10.4 6.8
< 0.001 191
Postcentral cortex L
–44 –20 38
9.0 6.3
< 0.001 176 Inferior frontal cortex, pars triangularis L
–39 34 12
8.1 5.9
< 0.001 174
Medial temporal cortex L
–45 –54 20
10.1 6.7
< 0.001 155
Precentral cortex R
33 –6 48
10 6.7
< 0.001 155
Posterior hippocampus L
–26 –33 –3
8.8 6.2
< 0.001 119
Cerebellar crus L
–16 –87 –37
8.8 6.2
< 0.001 116
Medial frontal cortex L
–27 –12 51
9.5 6.5
< 0.001 102
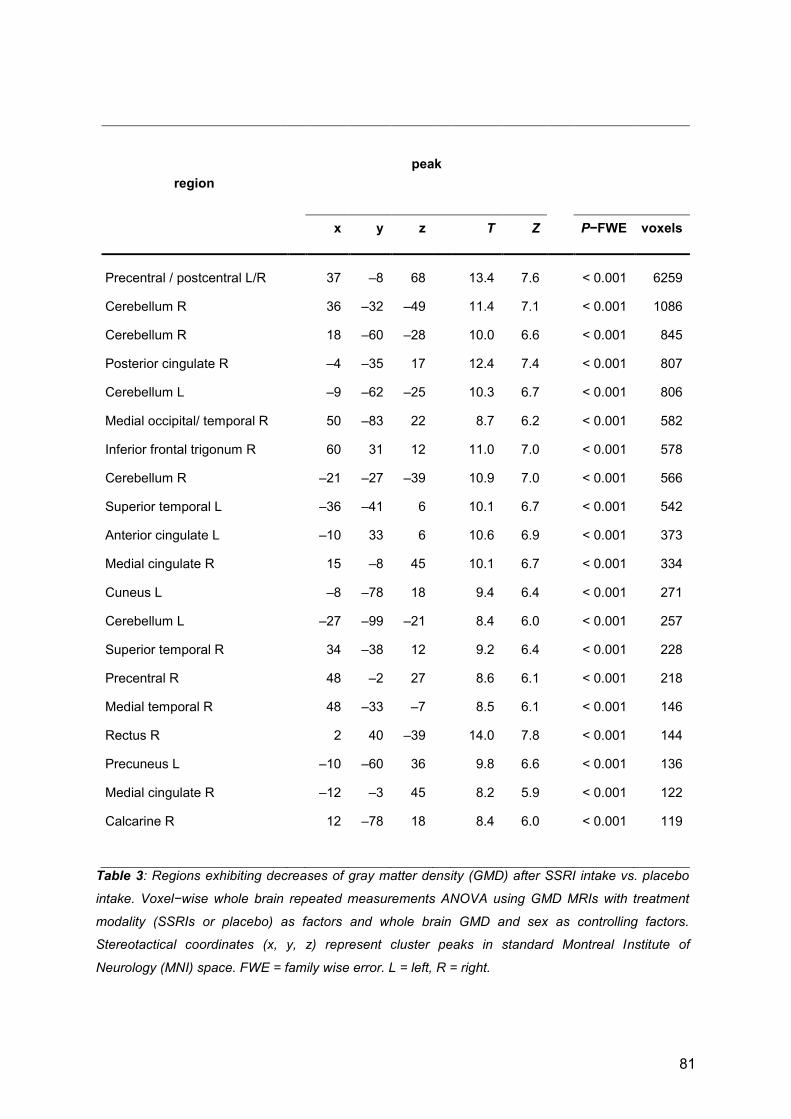
81
region
peak
x y z T Z P−FWE voxels
Precentral / postcentral L/R
37 –8 68
13.4 7.6
< 0.001 6259
Cerebellum R
36 –32 –49
11.4 7.1
< 0.001 1086
Cerebellum R
18 –60 –28
10.0 6.6
< 0.001 845
Posterior cingulate R
–4 –35 17
12.4 7.4
< 0.001 807
Cerebellum L
–9 –62 –25
10.3 6.7
< 0.001 806
Medial occipital/ temporal R
50 –83 22
8.7 6.2
< 0.001 582
Inferior frontal trigonum R
60 31 12
11.0 7.0
< 0.001 578
Cerebellum R
–21 –27 –39
10.9 7.0
< 0.001 566
Superior temporal L
–36 –41 6
10.1 6.7
< 0.001 542
Anterior cingulate L
–10 33 6
10.6 6.9
< 0.001 373
Medial cingulate R
15 –8 45
10.1 6.7
< 0.001 334
Cuneus L
–8 –78 18
9.4 6.4
< 0.001 271
Cerebellum L
–27 –99 –21
8.4 6.0
< 0.001 257
Superior temporal R
34 –38 12
9.2 6.4
< 0.001 228
Precentral R
48 –2 27
8.6 6.1
< 0.001 218
Medial temporal R
48 –33 –7
8.5 6.1
< 0.001 146
Rectus R
2 40 –39
14.0 7.8
< 0.001 144
Precuneus L
–10 –60 36
9.8 6.6
< 0.001 136
Medial cingulate R
–12 –3 45
8.2 5.9
< 0.001 122
Calcarine R
12 –78 18
8.4 6.0
< 0.001 119
Table 3: Regions exhibiting decreases of gray matter density (GMD) after SSRI intake vs. placebo
intake. Voxel−wise whole brain repeated measurements ANOVA using GMD MRIs with treatment
modality (SSRIs or placebo) as factors and whole brain GMD and sex as controlling factors.
Stereotactical coordinates (x, y, z) represent cluster peaks in standard Montreal Institute of
Neurology (MNI) space. FWE = family wise error. L = left, R = right.

82
1.13 Third publication: Exploring the impact of BDNF
Val66Met genotype on serotonin transporter and
serotonin-1A receptor binding
Christoph Kraus1, MD, Pia Baldinger1, MD, Christina Rami-Mark2, MSc, Gregor Gryglewski1,
Georg S. Kranz1, PhD, Daniela Haeusler2, PhD, Andreas Hahn1, PhD,
Wolfgang Wadsak2, Assoc. Prof. PD PhD, Markus Mitterhauser2, Assoc. Prof. PD,
Dan Rujescu3, Prof. MD, Siegfried Kasper1, Prof. MD, Rupert Lanzenberger1*, Assoc. Prof. PD MD
1 Department of Psychiatry and Psychotherapy,
2 Department of Biomedical Imaging und Image-guided Therapy, Division of Nuclear Medicine,
Medical University of Vienna, Austria
3 Department of Psychiatry, Medical University of Halle, Germany
published in:
PLOS-One, 2014 Sep 4;9(9)
[2013: 3.53]
* Correspondence to: Rupert Lanzenberger, Assoc. Prof. PD, MD
Functional, Molecular and Translational Neuroimaging Lab
Department of Psychiatry and Psychotherapy
Medical University of Vienna
Waehringer Guertel 18-20, 1090 Vienna, Austria
Phone (Fax): +43 1 40400 3825 (3099)
http://www.meduniwien.ac.at/neuroimaging

83
ABSTRACT
Background: The brain-derived neurotrophic factor (BDNF) Val66Met polymorphism
(rs6265) may impact on the in-vivo binding of important serotonergic structures such as the
serotonin transporter (5-HTT) and the serotonin-1A (5-HT1A) receptor. Previous positron
emission tomography (PET) studies on the association between Val66Met and 5-HTT and 5-
HT1A binding potential (BPND) have demonstrated equivocal results.
Methods: We conducted an imaging genetics study investigating the effect of Val66Met
genotype on 5-HTT or 5-HT1A BPND in 92 subjects. Forty-one subjects (25 healthy subjects
and 16 depressive patients) underwent genotyping for Val66Met and PET imaging with the
5-HTT specific radioligand [11C]DASB. Additionally, in 51 healthy subjects Val66Met
genotypes and 5-HT1A binding with the radioligand [carbonyl-11C]WAY-100635 were
ascertained. Voxel-wise and region of interest-based analyses of variance were used to
examine the influence of Val66Met on 5-HTT and 5-HT1A BPND.
Results: No significant differences of 5-HTT nor 5-HT1A BPND between BDNF Val66Met
genotype groups (val/val vs. met-carrier) were detected. There was no interaction between
depression and Val66Met genotype status.
Conclusion: In line with previous data, our work confirms an absent effect of BDNF
Val66Met on two major serotonergic structures. These results could suggest that altered
protein expression associated with genetic variants, might be compensated in vivo by
several levels of unknown feed-back mechanisms. In conclusion, Val66Met genotype status
is not associated with changes of in-vivo binding of 5-HTT and 5-HT1A receptors in human
subjects.

84
INTRODUCTION
The brain-derived neurotrophic factor (BDNF) is the most prominent member in the
neurotrophin family and involved in development and activity-dependent regulation of
neuronal structures [1]. Cumulating evidence demonstrated a functional interplay between
BDNF and the neurotransmitter serotonin (5-HT), constituting common intracellular signaling
pathways and transcription factors, BDNF control over the development and function of
serotonergic neurons as well as serotonergic regulation of BDNF gene expression and
signaling [2].
Briefly, BDNF is linked with at least three major intracellular signaling cascades: the
phosphoinositide-3 kinase pathway enabling cell survival, the phospholipase-gamma
pathway effecting synaptic plasticity and the mitogen-activated protein kinase pathway
associated with neuronal differentiation and neurite outgrowth [3]. Beside the p75
neurotrophin receptor, which is activated by proBDNF and all other neurotrophins, BDNF
releases it’s effects by binding to tropomyosin-kinase related receptor B (TrkB) [4-6].
Thereby, BDNF is a major factor in the proper development and plastic regulation of the
central nervous system and highly active in limbic structures such as the hippocampus and
the amygdala, where long-term potentiation, learning and memory are facilitated [7].
However, it should be stated here that most of the evidence of BDNF in this context is based
on rodent data.
The BDNF gene is located at chromosome 11p13-14, including many splice sites and
promoters. All BDNF mRNAs are initially translated into proBDNF and are then cleaved into
mature BDNF [8]. The most investigated polymorphism of the BDNF gene exists in the
codon 66 of proBDNF (Val66Met, rs6265) and consists of a valine to methionine substitution,
which is associated with reduced intracellular proBDNF trafficking, synaptic secretion of
BDNF, and thus a lower extracellular BDNF concentration in met-allele carriers [9]. Thought

85
to trigger deficits in neuronal development and plasticity, the Val66Met polymorphism is of
major interest in neuropsychiatric research [2, 7].
Interestingly, in humans the molecular connections between 5-HT and BDNF, and how
alterations in one system affect the other are hardly known. Due to the lack of current
methods to measure BDNF, TrkB or p75 in the living human brain, in vivo research in
humans mainly focuses on the investigation of alterations of serotonergic structures thought
to be mediated via changes in BDNF. In imaging genetics studies, serotonergic markers are
labeled by radioligands and their binding is measured using PET. As yet, there exist three
studies investigating alterations of BDNF, as represented by the Val66Met polymorphism,
and it’s association with binding of 5-HT1A, 5-HT2A receptors as well as the 5-HTT in the
human brain [10-12]. Two previous studies failed to detect links between Val66Met and
binding of 5-HT1A and 5-HT2A receptors. On the other side, a recently published study
reports lower 5-HT1A binding in healthy subjects carrying the met-allele compared to val-
homozygotes, a difference which was not observed in depressed subjects [12]. As far as 5-
HTT is concerned, in one study, applying the serotonin transporter (5-HTT) specific
radioligand [11C]-MADAM (N=25) with PET and [123I]-ß-CIT (N=18) with single photon
emission tomography (SPECT) in two independent samples, the authors found increased 5-
HTT binding in val-homozygote male subjects and compared to met-allele carriers [10]. On
the other hand applying the radioligand [11C]DASB (N=49), the second study failed to detect
any effect of Val66Met genotype status on 5-HTT binding [11].
To resolve contradictory results we conducted an imaging genetics study investigating the
association between 5-HTT binding using PET with the radioligand [11C]DASB and the
Val66Met genotype status in healthy subjects as well as in depressive patients. We also
measured 5-HT1A receptor binding in healthy subjects genotyped for Val66Met, in order to
resolve two equivocal findings. We hypothesized, that Val66Met impacts on 5-HTT binding in
patients with major depression and healthy subjects. Furthermore, we hypothesized that

86
significant differences are detected between BDNF genotype status and 5-HT1A binding in
healthy subjects.
METHODS
Subjects
In a neuroimaging genetics study with a cross-sectional design in total 92 subjects, aged 18-
65 years were included. The study was divided into two groups, in the first one 51 healthy
adult volunteers (37 female) were included and measured with [carbonyl-11C]WAY-10063. In
the second group 25 healthy subjects (HS) and 16 currently depressed patients with an
Hamilton Depression Rating Scale ≥ 16 (HAMD: 19.7±3.5, mean±SD) were included (for
further details see table 1) and measured with [11C]DASB. None of the subjects received
both radioligands. The study population originates from a pooled sample, which is part of
previously published studies [13-16]. Genotyping data of BNDF were previously not
published. All subjects underwent a psychiatric screening by the help of the complete
Structured Clinical Interview for DSM-IV type disorders (SCID I+II), physical and neurological
examination, clinical history, ECG, routine laboratory analysis, urinary drug and pregnancy
screening. All subjects were at least three months free of any psychotropic medication.
Every study subject was enrolled in study participation after detailed oral information about
all study procedures and subsequent signing of a written informed consent form. The study
and all study related procedures were approved by the Ethics Committee of the Medical
University of Vienna.
BDNF Genotyping
All procedures were performed as previously described [13]. Briefly, DNA was isolated from
peripheral blood mononuclear cells by the QIAamp DNA Mini-Kit (QIAGEN®, Hilden,

87
Germany). Genotyping of BDNF rs6265 single nucleotide polymorphism (SNP) was
conducted with the MassARRAY platform (SEQUENOM®, San Diego, CA) as described
elsewhere [17]. PCR-primers were generated with the Assay Designer 4.0 software
(SEQUENOM®). Multiplex PCR reactions were performed with 12.5ng of genomic DNA,
500μM dNTPs (ABgene®, Hamburg, Germany), 100nM PCR primers, 1.625mM MgCl2 and
0.5U HotStar Taq polymerase (QIAGEN®). Shrimp alkaline phosphatase (SAP) treatment,
an iPLEX reaction cocktail with extension primers (7-14μM), a iPLEX termination mix and an
iPLEX enzyme (SEQUENOM®) were added to the PCR-products. The resulting extension
products were desalted using SpectroCLEAN resin (SEQUENOM®), then spotted on
SpectroCHIPs GenII (SEQUENOM®) and analyzed with the MassARRAY MALDI-TOF mass
spectrometer. Typer 3.4 Software was used to identify allele specific extension products and
resulting genotypes (SEQUENOM®). For genotyping quality assurance CEU HapMap Trios
(Coriell Institute for Medical research, Camden, NJ) were included and compared with the
HapMap-CEU population (www.hapmap.org). For all analyses val/val homozygotes (=GG-
carriers) were compared against met-carriers (AG- and AA-carriers).
Radiochemistry of [11C]DASB and [carbonyl-11C]WAY-100635 and PET Procedures
Radioligand synthesis and all PET measurements were conducted at the Department of
Biomedical Imaging und Image-guided Therapy, Division of Nuclear Medicine at the Medical
University of Vienna. PET measurements were performed with a GE Advance full ring PET
scanner (General Electric Medical Systems, Waukesha, WI, USA). Subjects were placed
with their head parallel to the orbitomeatal line guided by a laser beam system to ensure full
coverage of the neocortex and the cerebellum in the field of view (FOV). A polyurethane
cushion and head straps were used to minimize head movement and to guarantee a soft
head rest during the whole scanning period.

88
For a complete description of [11C]DASB radioligand synthesis see [18]. Mean injected dose
was 358.97±70.47 MBq, specific activity at time of injection was 49.00±38.10 MBq/nmol and
radiochemical purity was above 95%. After a 5 min transmission scan with retractable 68Ge
rod sources the 3D dynamic emission measurement was initiated simultaneously with the
intravenous bolus injection of the radioligand [11C]DASB. The total acquisition time (35
slices) was 90 min and reconstructed images comprised a spatial resolution of 4.36 mm full-
width at half-maximum (FWHM).
For a complete description of [carbonyl-11C]WAY-100635 please see [19, 20]. Mean injected
dose was 312.04±105.84 MBq, specific activity at time of injection was
285.47±251.22GBq/µmol and radiochemical purity was above 95%. Again, a 5 min
transmission scan (68Ge) was followed by 90 min dynamic scanning per subject at a spatial
resolution of 4.36 mm FWHM.
Data preprocessing and calculation of binding potential
PET preprocessing was done in SPM8 (Wellcome Trust Centre for Neuroimaging, London,
UK, http://www.fil.ion.ucl.ac.uk/spm/) using standard algorithms and parameters unless
stated differently. After realignment to the mean image (quality = 1) scans of the entire time
series were summed up and spatially normalized (affine regularization = average sized
template) to a tracer-specific template within standard MNI-space (Montreal Neurological
Institute). Thereafter, the resulting transformation matrix was applied to each time frame.
We assessed in vivo target structure density as indexed by 5-HT1A receptor and 5-HTT
binding potentials (BPND), which represent the ratio at equilibrium of specifically bound
radioligand to that of nondisplaceable radioligand in tissue [21]. All binding potentials were
computed using the voxel-wise modeling tool in the PMOD 3.3 software package (PMOD
Technologies, Ltd., Zurich, Switzerland) and applying the two-parameter linearized reference

89
tissue model (MRTM2) [22], which provides advantages in signal-to-noise-ratio, especially
for whole-brain voxel-wise analysis.
We modeled 5-HT1A BPND as previously described by our group using the insula as receptor-
rich region and the cerebellum as receptor-poor region [23]. The cerebellar gray matter
excluding cerebellar vermis and venous sinus served as reference region. Serotonin
transporter BPND were modeled using the MRTM2 as previously described [16]. In short, k2’
was estimated from the striatum as 5-HTT-rich region and the cerebellar gray matter (excl.
vermis and venous sinus) as 5-HTT-poor region. The cerebellar gray matter was chosen
because it represents an optimal reference region for the quantification of the serotonin
transporter with [11C]DASB [24, 25]. Regions of interest (ROI) for both radioligands were
taken from an automated anatomical labeling-based (AAL) atlas [26] after normalization of
BPND maps to standard MNI-space. Values were averaged across both hemispheres. Due to
inherent smoothness of PET data of the scanner and temporary smoothing during
normalization we did not smooth during statistical processing.
Statistical Analysis
For normally distributed demographic variables and clinical measures student’s t-tests, for
nominal variables chi-squared tests were performed. Significance was determined as p<0.05
and all tests were two-sided.
Differences of 5-HT1A and 5-HTT BPND between BDNF Val66Met genotype groups were
calculated using a voxel-wise and a ROI-based approach. For the voxel-wise analysis both
in the 5-HTT and the 5-HT1A – groups an ANOVA was performed as implemented in SPM8.
Grouped genotype status (val/val, vs. met-carrier = GG vs. A-carrier) served as factor and
radioligand specific activity, sex and age served as covariates. In the 5-HTT-collective
diagnosis was added as additional factor in a second step analysis. F-tests and group-wise
post-hoc t-tests between genotype groups were calculated and contrasted in SPM8.

90
Additionally, in the 5-HTT-group an interaction between diagnosis and genotype status was
contrasted by weighting contrast vectors in SPM according to group size. An absolute image
threshold was set at 0.1 BPND to remove voxels with low signal-to-noise ratio and a cluster
threshold was set at 50 voxels. A statistical level of p<0.05 corrected for multiple
comparisons by the family-wise error rate (FWE) at voxel-level was considered significant,
for subsequent explorative analysis an uncorrected threshold of p<0.001 was accepted.
In the ROI-based analyses differences between genotypes groups (val/val vs. met carrier)
were calculated with a linear mixed model in SPSS 19 (IBM Corp. Released 2010. IBM
SPSS Statistics for Windows, Version 19.0. Armonk, NY: IBM Corp.). Thereby, subject
served as the random effect and BDNF genotype status, region, sex and age served as
fixed effects. Ten representative regions were chosen due to their a priori known high
distribution of 5-HT1A receptors and 5-HTT and implications in psychiatric disorders (see
tables 1,2 and figures 1,2). Diagnosis was taken as additional factor in the 5-HTT-study
collective. Significance was determined as p<0.05. Post-hoc t-tests were conducted two-
sided in 10 AAL ROIs (see tables 1,2 and figures 1,2).
RESULTS
Out of the 51 HS in the 5-HT1A-group 30 carried GG, 18 carried AG and 3 AA. The 5-HTT-
group had 25 HS with 19 carrying GG, 5 carried AG and 1 AA, whereas in the MDD group
with 16 depressed patients 13 carried GG, 3 carried AG and 0 the AA allele (table 1). Allele
frequencies of the BDNF gene in all study groups were distributed in accordance with the
Hardy-Weinberg equilibrium [5-HT1A-group: 2=0.02, p=0.891, 5-HTT-group HS: 2=0.72
p=0.4, MDD patients 2=0.17 p=0.68). The AA and AG+GG study groups did not differ in
demographical, clinical measures or radiopharmaceutical measures (table 1). The allelic
distribution was not associated with diagnosis in the 5-HTT-group (2=0.157, p=0.692).

91
In the voxel-wise analysis there was no significant association of BDNF genotype (GG vs. A-
carrier) status with 5-HT1A BPND (F-test: all p>0.05 FWE corr. and all p>0.001 uncorr.).
Furthermore, there was no significant association of BDNF genotype (GG vs. A-carrier) with
5-HTT BPND (F-test: all p>0.05 FWE corr. and all p>0.001 uncorr.). There was no interaction
between BDNF genotype status, diagnosis or sex and 5-HTT BPND (t-test: all p>0.05 FWE
corr. and all p>0.001 uncorr.).
The mixed model analyses of ROIs in the 5-HT1A-group, controlling for potential effects of
sex, age and specific radioligand activity, yielded no significant difference of 5-HT1A BPND in
selected ROIs between GG homozygotes and A-allele carriers (F=0.342, df=1,45, p=0.562).
In the 5-HTT-group, the mixed model revealed no significant difference between 5-HTT BPND
in the selected ROIs between GG homozygotes and A-allele carriers (F=0,41, df=1,33,
p=0.526). There was no interaction between diagnosis and allele in the statistical model
(p=0.989). Post-hoc t-tests and average BPND values for both study groups are shown in
Figure 1: Bar chart plotting serotonin-1A binding potential (5-HT1A BPND) according to BNDF
Val66Met genotype status. Values at the y-axis represent 5-HT1A BPND separated for val/val and
met-carrier, respectively, x-axis shows regions of interest. Regions and values correspond to table
2. ACC: anterior cingulate cortex, AMY: amygdala, MCC: medial cingulate cortex, HIPP:
hippocampus, INS: insula, paraHIPP: parahippocampus, PCC: posterior cingulate cortex,
TempPole: temporal pole, DRN: dorsal raphe nucleus.

92
table 2 and table 3, BPND-values of allele groups are displayed in figure 1 and figure 2. Here,
in the 5-HTT-group, a significant difference between GG and A-carriers was observed in HS
in the midbrain (p=0.040, uncorr., table 3) as well as between GG in HS and GG in MDD
patients (p=0.034, uncorr.), with BPND increases in GG-carriers, respectively. All other post-
hoc tests (5-HT1A: GG vs. A-carrier; 5-HTT HS: GG vs. A-carrier, MDD GG vs. A-carrier, HS
vs. MDD GG, HS males GG vs. HS males A-carrier) did not yield significant results (all
p>0.05 uncorr.).
DISCUSSION
In a voxel-wise analysis as well as in a ROI-based approach, we did not observe significant
differences of 5-HT1A-receptor BPND nor of 5-HTT BPND according to BDNF genotype status.
Figure 2: Bar chart plotting serotonin transporter binding potential (5-HTT BPND) according to
BNDF Val66Met genotype status. Values at the y-axis represent 5-HTT BPND in pooled healthy
subjects and depressive patients. Binding potential is separated for val/val and met-carriers,
respectively, x-axis shows regions of interest. Because healthy subjects and depressive patients
were pooled here, regions do, but values do not correspond to table 3. ACC: anterior cingulate
cortex, AMY: amygdala, MCC: medial cingulate cortex, HIPP: hippocampus, CAUD: caudatum,
PUT: putamen, THAL: thalamus, STRIA: striatum, MID: Midbrain, NACC: nucleus accumbens.

93
There was no interaction between MDD diagnosis or sex and 5-HTT BPND. In the midbrain,
weak increases of 5-HTT-BPND in healthy subjects between val-homozygotes and met-
carriers were found. Furthermore, weak increases of 5-HTT BPND were observed in the
midbrain in val-homozygote healthy subjects compared to val-homozygote MDD patients.
There was no association between allelic distribution and major depression. To sum up, all
voxel-wise and ROI-based testing yielded negative results and none of the post-hoc tests
survived correction.
Our results are in concordance with a previous PET study applying [11C]DASB in 49 healthy
subjects, where the authors neither detected differences in 5-HTT binding in relation to
BDNF genotype nor a correlation between blood BDNF levels and central 5-HTT binding
[11]. Additionally, no effect on 5-HT2A binding was shown in this work. Here, the authors
calculated the radiotracer BPND similar to our study by applying a fully automated reference
region model (MRTM2) [22] and an automated ROI-delineation. The only other currently
published human PET-study investigating the impact of BDNF polymorphisms on 5-HTT
binding reports differences in men and shows no effect of genotype status on 5-HT1A binding
[10]. Men homozygous for the val-allele exhibited significantly higher 5-HTT binding in
regions such as the hippocampus, insula or dorsal raphe compared to met-carrier, while this
effect was absent in women. Furthermore, reductions of 5-HTT binding in met-carrier (n=3)
compared to val-homozygotes (n=6) in an independent [123I]-ß-CIT-study with male suicide
attempters were demonstrated, but this reduction was absent when pooled with healthy
controls. The authors also used a reference region model with [11C]-MADAM, a tracer
exhibiting a comparable 5-HTT affinity to [11C]DASB [27], the ROIs were manually delineated
on individual magnetic resonance images (MRI). Notably, our group previously reported
strong correlations of BPND values between automatically and manually delineated ROIs
[23]. The radioligand and the method of ROI generation are on these grounds an unlikely
source of variance leading to alternative results. Importantly, in search of arguments for this
difference, one must mention that the number of male met-carriers in that collective was low

94
(n=4), which makes the analysis vulnerable to outliers and hence may increase type-I errors.
Likewise, our study exhibits a subgroup with a low subject number and indeed we saw an
outlier in the MDD met-carrier group (n=3) when we plotted the individual BPND values (data
not shown). Hence, our results in depressed patients have to be interpreted with caution. But
the fact that both the study by Klein et al., which exhibits a large sample size of healthy
volunteers, as well as our study did not reproduce higher 5-HTT binding in val-homozygote
healthy subjects, rather speaks for an absent effect of BDNF Val66Met on 5-HTT binding.
Apart from this, our study agrees with the data by Henningsson et al., on an absent effect of
Val66Met on 5-HT1A receptor binding in healthy subjects [10]. Both studies apply the same
radioligand, i.e. [carbonyl-11C]WAY-100635, exhibit an almost identical number of subjects
(n=53 in Henningsson et al.), and modeled 5-HT1A binding by a reference region model
(BPND). These results are in contradiction to a recent finding reporting 5-HT1A reductions in
healthy met-allele carriers [12], which is not present in MDD patients. In this study 50 healthy
subjects and 50 MDD patients were measured with the radioligand [carbonyl-11C]WAY-
100635, yet 5-HT1A binding was calculated by an arterial input function (BPF). Most
interestingly, when the authors repeated their analysis with BPND values, the reduction of 5-
HT1A binding in healthy met-carriers was not detectable, suggesting that this finding was
associated with the method of radioligand modeling. Following the discussion of the authors,
one cannot rule out that Val66Met causes differences of radioligand binding in the blood
leading to a bias in the arterial input function. Although, our results are in agreement with all
previous studies on 5-HT1A binding using reference tissue models [10, 12], validation by a
different tracer not susceptible to modeling methodology is further needed. Taken together,
while there are currently contradicting findings on the in vivo effect of BDNF Val66Met
genotypes on 5-HTT binding [10, 11], this study adds data emphasizing the absence of such
an effect. Moreover, this work corroborates previous results by reference tissue models
demonstrating no association between BDNF Val66Met genotype status and 5-HT1A

95
receptor binding [10, 12] and is in contradiction with a study reporting binding values
modeled with arterial blood sampling [12].
Preclinical data report that BDNF promotes development and function of serotonergic
neurons by enhancing survival and differentiation [28], increasing local 5-HT [29] modifying
the firing pattern of serotonergic raphe neurons [28, 30] and altering the function of
serotonergic receptors such as the 5-HT1A and 5-HT2A receptors and the 5-HTT [2, 29, 31].
Vice-versa, raised extracellular 5-HT levels occurring upon administration of SSRIs are
thought to increase local BDNF levels by enhanced phosphorylation of serotonergic receptor
coupled cAMP response element-binding (CREB) protein [32-34], a common target of BDNF
and G protein-coupled serotonergic receptors [2]. Confronted with this evidence, one is
puzzled upon the lack of strong evidence for an association between BDNF and
serotonergic structures in humans in vivo. However, preclinical studies are not consistent
and negative results regarding the expression of 5-HT receptors and transporter are
reported [31, 35]. Although the interaction between the BNDF and 5-HT provides a
promising bridge between structural and functional neuronal activity, and serves as
explanatory hypothesis for neuronal plasticity deficits in neuropsychiatric disorders, exact
mechanisms underlying the regulation of the cross connection between BDNF and 5-HT in
humans remain unresolved [36]. Our data in concert with above referred work speak for a
similar expression of 5-HTT and 5-HT1A receptors upon life-time BDNF reduction, but
unfortunately do not illuminate the mechanisms leading to this observation. Theoretically,
counter-regulatory or compensatory effects may have altered 5-HTT and 5-HT1A expression.
Furthermore, it is possible that not absolute numbers but functional activity of serotonergic
structures is altered by BDNF.
The evidence on connections between depression and BNDF genotype status is
inconsistent as well. Meta-analytical research suggested an association of Val66Met with
major depressive disorder antidepressant treatment response or hippocampal volume and a
role of gender and ethnicity [37-39]. However, recent meta-analyses refuted these

96
associations and detected power deficits in many trials [40-42]. Low serum levels of BDNF
were suggested as potential peripheral marker of depression and increase of serum BDNF
as response to the appropriate first-line treatment with selective 5-HT reuptake inhibitors
(SSRIs). Likewise, this association is weaker than initially thought and there is no
relationship between symptom severity and BDNF serum concentration [43]. Our results
suggest no association between allelic distribution and diagnosis. Our small number of MDD
subjects remain a limiting factor in that regard.
LIMITATIONS
Unfortunately a common problem of human PET studies is weak power resulting from low
subject numbers, owed to the large effort of conducting PET-imaging. This is even more
intrinsic to genetic PET studies reporting results based on genotype subgroups [44] and in
SNP neuroimaging studies where pooling of rare genotype groups is common practice. The
low subject number in the MDD met-carrier group could therefore be a limitation of our study.
One elegant way to circumvent this issue in future studies would be pooling data between
PET centers, which is already common in MRI studies. Second, mean age of genotype
groups is heterogeneous, yet controlled for in all statistical analyses. Finally, we did not
model PET data with an arterial input function [45], because arterial blood data were not
collected. This would have been useful to confirm reported differences according to the
methodology for calculating 5-HT1A binding with [carbonyl-11C]WAY-100635, an issue we are
trying to resolve in future studies [46].
CONCLUSION
Although others have investigated the effects of the BDNF gene on 5-HTT and 5-HT1A
binding with PET, this study adds data to the ongoing discussion about the cross connection

97
between 5-HT and BDNF. While previous work in humans demonstrated contradicting
results, due to this work the conclusion of an absent influence of Val66Met on 5-HTT and 5-
HT1A has gained substantial support.
ACKNOWLEDGEMENTS
The authors are grateful to U. Moser, E. Akimova, P. Stein, M. Fink, C. Spindelegger, A.
Höflich, I. Hofer-Irmler, S. Zgud, S. Pichler, A. Kautzky and D. Winkler for medical and
administrative support, and M. Savli for technical support. We thank the PET team,
especially G. Karanikas, T. Traub-Weidinger, L.-K. Mien, J. Ungersboeck, K. Kletter, L. Nics,
and C. Philippe for technical support. Further, we thank the genetics team of D. Rujescu,
especially M. Friedl, A. Hartmann, I. Giegling.
The study is part of C. Kraus’ thesis “Serotonin and Neuroplasticity” supervised by R.
Lanzenberger in the Clinical Neurosciences PhD program at the Medical University of
Vienna, Austria. Parts of this study have been presented by P. Baldinger at the 19th at the
11th World Congress of Biological Psychiatry (WFSBP), June 23rd – 27th, 2013, Kyoto, Japan.
REFERENCES
1. Johnson JE, Barde YA, Schwab M and Thoenen H (1986) Brain-derived neurotrophic factor supports the survival of cultured rat retinal ganglion cells. J Neurosci 6: 3031-3038.
2. Martinowich K and Lu B (2008) Interaction between BDNF and serotonin: role in mood disorders. Neuropsychopharmacology 33: 73-83.
3. Chao MV (2003) Neurotrophins and their receptors: a convergence point for many signalling pathways. Nat Rev Neurosci 4: 299-309.
4. Gentry JJ, Barker PA and Carter BD (2004) The p75 neurotrophin receptor: multiple interactors and numerous functions. Prog Brain Res 146: 25-39.
5. Gan WB (1999) TrkB works at postsynaptic sites. Neuron 24: 491-492.
6. Black IB (1999) Trophic regulation of synaptic plasticity. J Neurobiol 41: 108-118.

98
7. Castrén E (2005) Is mood chemistry? Nature reviews Neuroscience 6: 241-246.
8. Liu QR, Walther D, Drgon T, Polesskaya O, Lesnick TG, et al. (2005) Human brain derived neurotrophic factor (BDNF) genes, splicing patterns, and assessments of associations with substance abuse and Parkinson's Disease. Am J Med Genet B Neuropsychiatr Genet 134B: 93-103.
9. Chen ZY, Patel PD, Sant G, Meng CX, Teng KK, et al. (2004) Variant brain-derived neurotrophic factor (BDNF) (Met66) alters the intracellular trafficking and activity-dependent secretion of wild-type BDNF in neurosecretory cells and cortical neurons. J Neurosci 24: 4401-4411.
10. Henningsson S, Borg J, Lundberg J, Bah J, Lindstrom M, et al. (2009) Genetic variation in brain-derived neurotrophic factor is associated with serotonin transporter but not serotonin-1A receptor availability in men. Biol Psychiatry 66: 477-485.
11. Klein AB, Trajkovska V, Erritzoe D, Haugbol S, Madsen J, et al. (2010) Cerebral 5-HT2A receptor and serotonin transporter binding in humans are not affected by the val66met BDNF polymorphism status or blood BDNF levels. J Cereb Blood Flow Metab 30: e1-7.
12. Lan MJ, Ogden RT, Huang YY, Oquendo MA, Sullivan GM, et al. (2014) Genetic variation in brain-derived neurotrophic factor val66met allele is associated with altered serotonin-1A receptor binding in human brain. NeuroImage.
13. Baldinger P, Hahn A, Mitterhauser M, Kranz GS, Friedl M, et al. (2013) Impact of COMT genotype on serotonin-1A receptor binding investigated with PET. Brain Struct Funct.
14. Lanzenberger R, Kranz GS, Haeusler D, Akimova E, Savli M, et al. (2012) Prediction of SSRI treatment response in major depression based on serotonin transporter interplay between median raphe nucleus and projection areas. NeuroImage.
15. Lanzenberger R, Wadsak W, Spindelegger C, Mitterhauser M, Akimova E, et al. (2010) Cortisol plasma levels in social anxiety disorder patients correlate with serotonin-1A receptor binding in limbic brain regions. Int J Neuropsychopharm: 1-15.
16. Hahn A, Haeusler D, Kraus C, Hoflich AS, Kranz GS, et al. (2014) Attenuated serotonin transporter association between dorsal raphe and ventral striatum in major depression. Hum Brain Mapp.
17. Oeth P, Beaulieu M, Park C, Kosman D, del Mistro G, et al. (2006) iPLEXTM Assay: Increased Plexing Efficiency and Flexibility for MassARRAY® System Through Single Base Primer Extension with Mass-Modified Terminators. Application Note. Sequenom®.
18. Haeusler D, Mien LK, Nics L, Ungersboeck J, Philippe C, et al. (2009) Simple and rapid preparation of [11C]DASB with high quality and reliability for routine applications. Appl Radiat Isot 67: 1654-1660.
19. Wadsak W. MLK, Ettlinger D.E., Lanzenberger R., Haeusler D., Dudczak R., Kletter K., Mitterhauser M. (2007) Simple and fully automated preparation of [carbonyl-11C]WAY-100635. Radiochimica Acta 95: 1-6.

99
20. Rami-Mark C, Ungersboeck J, Haeusler D, Nics L, Philippe C, et al. (2013) Reliable set-up for in-loop (1)(1)C-carboxylations using Grignard reactions for the preparation of [carbonyl-(1)(1)C]WAY-100635 and [(1)(1)C]-(+)-PHNO. Appl Radiat Isot 82: 75-80.
21. Innis RB, Cunningham VJ, Delforge J, Fujita M, Gjedde A, et al. (2007) Consensus nomenclature for in vivo imaging of reversibly binding radioligands. J Cereb Blood Flow Metab 27: 1533-1539.
22. Ichise M, Liow J-S, Lu J-Q, Takano A, Model K, et al. (2003) Linearized Reference Tissue Parametric Imaging Methods: Application to [11C]DASB Positron Emission Tomography Studies of the Serotonin Transporter in Human Brain. Journal of Cerebral Blood Flow & Metabolism: 1096-1112.
23. Savli M, Bauer A, Mitterhauser M, Ding YS, Hahn A, et al. (2012) Normative database of the serotonergic system in healthy subjects using multi-tracer PET. NeuroImage 63: 447-459.
24. Parsey RV, Kent JM, Oquendo MA, Richards MC, Pratap M, et al. (2006) Acute occupancy of brain serotonin transporter by sertraline as measured by [11C]DASB and positron emission tomography. Biol Psychiatry 59: 821-828.
25. Meyer JH (2007) Imaging the serotonin transporter during major depressive disorder and antidepressant treatment. J Psychiatry Neurosci 32: 86-102.
26. Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, et al. (2002) Automated Anatomical Labeling of Activations in SPM Using a Macroscopic Anatomical Parcellation of the MNI MRI Single-Subject Brain. NeuroImage 15: 273-289.
27. Laruelle M, Slifstein M and Huang Y (2003) Relationships between radiotracer properties and image quality in molecular imaging of the brain with positron emission tomography. Mol Imaging Biol 5: 363-375.
28. Djalali S, Holtje M, Grosse G, Rothe T, Stroh T, et al. (2005) Effects of brain-derived neurotrophic factor (BDNF) on glial cells and serotonergic neurones during development. J Neurochem 92: 616-627.
29. Guiard BP, David DJP, Deltheil T, Chenu F, Le Maître E, et al. (2008) Brain-derived neurotrophic factor-deficient mice exhibit a hippocampal hyperserotonergic phenotype. Int J Neuropsychopharmacol 11: 79-92.
30. Yu H, Wang DD, Wang Y, Liu T, Lee FS, et al. (2012) Variant brain-derived neurotrophic factor Val66Met polymorphism alters vulnerability to stress and response to antidepressants. J Neurosci 32: 4092-4101.
31. Daws LC, Munn JL, Valdez MF, Frosto-Burke T and Hensler JG (2007) Serotonin transporter function, but not expression, is dependent on brain-derived neurotrophic factor (BDNF): in vivo studies in BDNF-deficient mice. J Neurochem 101: 641-651.
32. Nibuya M, Morinobu S and Duman RS (1995) Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J Neurosci 15: 7539-7547.

100
33. Khundakar AA and Zetterstrom TS (2006) Biphasic change in BDNF gene expression following antidepressant drug treatment explained by differential transcript regulation. Brain Res 1106: 12-20.
34. Coppell AL, Pei Q and Zetterstrom TS (2003) Bi-phasic change in BDNF gene expression following antidepressant drug treatment. Neuropharmacology 44: 903-910.
35. Szapacs ME, Mathews TA, Tessarollo L, Ernest Lyons W, Mamounas LA, et al. (2004) Exploring the relationship between serotonin and brain-derived neurotrophic factor: analysis of BDNF protein and extraneuronal 5-HT in mice with reduced serotonin transporter or BDNF expression. J Neurosci Methods 140: 81-92.
36. Castren E (2013) Neuronal network plasticity and recovery from depression. JAMA Psychiatry 70: 983-989.
37. Zou YF, Ye DQ, Feng XL, Su H, Pan FM, et al. (2010) Meta-analysis of BDNF Val66Met polymorphism association with treatment response in patients with major depressive disorder. Eur Neuropsychopharmacol 20: 535-544.
38. Verhagen M, van der Meij A, van Deurzen PA, Janzing JG, Arias-Vasquez A, et al. (2010) Meta-analysis of the BDNF Val66Met polymorphism in major depressive disorder: effects of gender and ethnicity. Mol Psychiatry 15: 260-271.
39. Hajek T, Kopecek M and Hoschl C (2012) Reduced hippocampal volumes in healthy carriers of brain-derived neurotrophic factor Val66Met polymorphism: meta-analysis. World J Biol Psychiatry 13: 178-187.
40. Molendijk ML, van Tol MJ, Penninx BW, van der Wee NJ, Aleman A, et al. (2012) BDNF val66met affects hippocampal volume and emotion-related hippocampal memory activity. Transl Psychiatry 2: e74.
41. Gyekis JP, Yu W, Dong S, Wang H, Qian J, et al. (2013) No association of genetic variants in BDNF with major depression: a meta- and gene-based analysis. Am J Med Genet B Neuropsychiatr Genet 162B: 61-70.
42. Cole J, Weinberger DR, Mattay VS, Cheng X, Toga AW, et al. (2011) No effect of 5HTTLPR or BDNF Val66Met polymorphism on hippocampal morphology in major depression. Genes Brain Behav 10: 756-764.
43. Molendijk ML, Spinhoven P, Polak M, Bus BA, Penninx BW, et al. (2013) Serum BDNF concentrations as peripheral manifestations of depression: evidence from a systematic review and meta-analyses on 179 associations (N=9484). Mol Psychiatry.
44. Willeit M and Praschak-Rieder N (2010) Imaging the effects of genetic polymorphisms on radioligand binding in the living human brain: A review on genetic neuroreceptor imaging of monoaminergic systems in psychiatry. NeuroImage: 1-15.
45. Ichise M, Meyer JH and Yonekura Y (2001) An introduction to PET and SPECT neuroreceptor quantification models. J Nucl Med 42: 755-763.
46. Parsey RV, Ogden RT, Miller JM, Tin A, Hesselgrave N, et al. (2010) Higher Serotonin 1A Binding in a Second Major Depression Cohort: Modeling and Reference Region Considerations. Biol Psychiatry 68: 170-178.
101
Table 1. Demographic variables of the entire study sample.
val/val
met-carrier
p
healthy subjects
[carbonyl-11C]WAY-100635
N (=51) 30
21
Age (years) 43.8 ± 13.1
45.1 ± 12.36
0.737
Sex (f/m) 21/9
16/5
0.626*
weight 72.9 ± 17.1
67.1 ± 10.5
0.169
SA 296.9 ± 269.1
285.7 ± 197.3
0.702
[11
C]DASB
N (=25) 19
6
Age (years) 31.0 ± 8.8
33.0 ± 13.2
0.672
Sex (f/m) 8/11
1/5
0.258*
weight 76.7 ± 12.1
80.2 ± 10.8
0.537
SA 44.1 ± 47.7
25.6 ± 25.4
0.378
MDD patients
[11
C]DASB N (=16) 13
3
HAMD 19.4 ± 3.6
21± 3.5
0.495
Age (years) 41.1 ± 8.9
46.7 ± 7.5
0.34
Sex (f/m) 9/4
3/0
0.267*
weight 77.7 ± 21.3
61.3 ± 2.5
0.251
+
SA 63.9 ± 22.6
62.5 ± 16.7
0.925
Data are given as means ± standard deviations (SD). P-values compare pooled
BNDF Val66Met genotype groups with independent sample t-test, chi-square(*) or
Mann-Whitney U test (+) where appropriate.

102
Table 2. Post-hoc t-tests comparing serotonin-1A receptor (5-HT1A) binding
potential (BPND) according to BDNF Val66Met genotype status in 51 healthy
subjects.
healthy subjects [carbonyl-11C]WAY-100635
region val/val (n=30)
met-carrier (n=21)
p
Anterior cingulate cortex 3.54 ± 1.14
3.63 ± 0.86
0.758
Amygdala 3.98 ± 1.23
4.17 ± 1.03
0.559
Medial cingulate cortex 2.9 ± 0.97
2.98 ± 0.65
0.723
Hippocampus 3.64 ± 1.14
4.12 ± 0.94
0.118
Insula 4.46 ± 1.33
4.64 ± 0.91
0.596
Parahippocampus 5.41 ± 1.64
5.60 ± 1.14
0.596
Posterior cingulate cortex 2.2 ± 0.79
2.25 ± 0.58
0.822
Subgenual anterior cingulate 3.51 ± 0.96
3.85 ± 1.1
0.247
Temporal pole 4.65 ± 1.5
4.75 ± 0.93
0.786
Dorsal raphe nucleus 2.33 ± 0.87
2.29 ± 0.74
0.857
Regions of interest (ROIs) in standardized MNI space (Montreal Neurological Institute) were
calculated by automatic anatomical labeling in both hemispheres and averaged. Data are
given as 5-HT1A BPND means ± standard deviations (SD) for each ROI and compared by
post-hoc student’s t-tests, values correspond to bar charts in Fig.1.
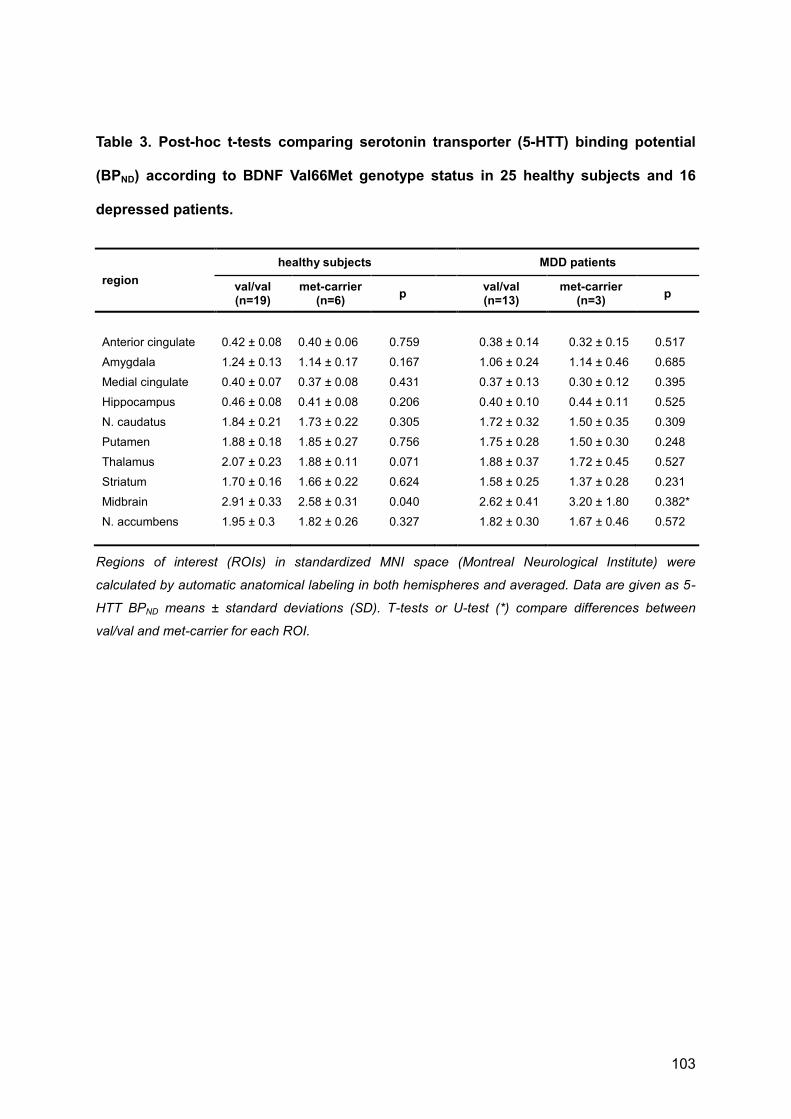
103
Table 3. Post-hoc t-tests comparing serotonin transporter (5-HTT) binding potential
(BPND) according to BDNF Val66Met genotype status in 25 healthy subjects and 16
depressed patients.
region
healthy subjects
MDD patients
val/val (n=19)
met-carrier (n=6)
p
val/val (n=13)
met-carrier (n=3)
p
Anterior cingulate 0.42 ± 0.08 0.40 ± 0.06 0.759
0.38 ± 0.14 0.32 ± 0.15 0.517
Amygdala 1.24 ± 0.13 1.14 ± 0.17 0.167
1.06 ± 0.24 1.14 ± 0.46 0.685
Medial cingulate 0.40 ± 0.07 0.37 ± 0.08 0.431
0.37 ± 0.13 0.30 ± 0.12 0.395
Hippocampus 0.46 ± 0.08 0.41 ± 0.08 0.206
0.40 ± 0.10 0.44 ± 0.11 0.525
N. caudatus 1.84 ± 0.21 1.73 ± 0.22 0.305
1.72 ± 0.32 1.50 ± 0.35 0.309
Putamen 1.88 ± 0.18 1.85 ± 0.27 0.756
1.75 ± 0.28 1.50 ± 0.30 0.248
Thalamus 2.07 ± 0.23 1.88 ± 0.11 0.071
1.88 ± 0.37 1.72 ± 0.45 0.527
Striatum 1.70 ± 0.16 1.66 ± 0.22 0.624
1.58 ± 0.25 1.37 ± 0.28 0.231
Midbrain 2.91 ± 0.33 2.58 ± 0.31 0.040
2.62 ± 0.41 3.20 ± 1.80 0.382*
N. accumbens 1.95 ± 0.3 1.82 ± 0.26 0.327
1.82 ± 0.30 1.67 ± 0.46 0.572
Regions of interest (ROIs) in standardized MNI space (Montreal Neurological Institute) were
calculated by automatic anatomical labeling in both hemispheres and averaged. Data are given as 5-
HTT BPND means ± standard deviations (SD). T-tests or U-test (*) compare differences between
val/val and met-carrier for each ROI.

104
IV. GENERAL DISCUSSION and RAISED QUESTIONS
With this work we investigated a series of issues on the association of neuroplasticity with
serotonin in the living human brain. To this goal, neuroimaging methods such as structural
and functional MRI and PET with the radioligands [11C]DASB and [carbonyl-11C]WAY-100635
were used for the to probe relationships between elevated 5-HT levels, gray matter,
important serotonergic proteins and the neurotrophine brain derived neurotrophic factor. The
here presented results underline that some of the known functions of 5-HT shaping the
neuronal architecture of the brain in embryonic and early post-natal periods are very likely, at
least partly, conserved throughout adulthood.
While neuroplastic effects of the 5-HT1A receptor function were previously discovered in
animals, we here translated this feature into in vivo human research by demonstrating strong
associations between the distribution of the 5-HT1A receptor and regional gray matter
volume. This result was mainly observed in brain regions with high 5HT1A receptor density,
yet restricted to certain regions and not present in others with equaly high 5-HT1A densitiy,
suggesting regional differences in neuroplastic functions. Furthermore, a correlation of 5-
HT1A autoreceptors in the dorsal raphe nuclei, one of the midbrain’s major serotonergic
nucleus, and cortical gray matter in the anterior cingulate cortex undermined the regulatory
function of the raphe nuclei. With the limitation of beeing correlational, these results in any
case justify further longitudinal investigations e.g. into how pharmacological manipulations of
5-HT1A receptor function would be able to effect changes of gray matter. Secondly, we found
that elevated 5-HT levels after selective serotonin reuptake inhibitor administration leads to
strong enhancements of cortical gray matter in the posterior cingulate cortex, which are in
turn associated with altered functional neuronal activity in this region. While this finding goes
along with previous work showing gray matter inceases after selective serotonin reuptake
inhibitor intake in psychiatric patients (Hoexter et al, 2012; Smith et al, 2012; Vermetten et al,
2003), gray matter increases in healthy subjects constitute a novel finding, as well as
adjacent functional alterations as consequence of enhanced gray matter. These results raise

105
many questions. Given that healthy subjects do not exhibit major neuropsychological
changes after 10 days of SSRI intake, one might ask what are the phenotypical correlates of
such strong signal increases. Consequently, this work demands closer investigations of the
biological correlate of VBM results. Several authors question the sensitivity regarding
neuronal changes in VBM studies (Bookstein, 2001; Franklin et al, 2013) and suspect
changes of perfusion and difusion of cerebral blood flow to underlie VBM findings. Indeed,
the huge gap between biological mechanisms and findings of MRI-based structural
neuroimaigng techniques are currently discussed controversially (Draganski & Kherif, 2013;
Erickson, 2013; Fields, 2013; Thomas & Baker, 2013). This work adds to the ongoing
discussion that neuroplastic serotonergic receptors might contribute to structural MRI signal
changes. This supposition provides new testable hypoptheses, that could combine
neurobiological data on neuroplasticity with neuroimaging-based in vivo information.
Finally, we found no evidence for an association between SERT or 5-HT1A availability upon
lifetime alterations of BDNF as produced by a common single nucleotid polymorphism
(Val66Met). This finding is controversial, because two previous studies demonstrated an
effect of this polymorphism on the expression of SERT and 5-HT1A receptors (Henningsson
et al, 2009). The dicrepancy might arise from methodological differences in quantification
using the radiligand [carbonyl-11C]WAY-100635, especially due to reference region data from
an aterial input function by other groups. Given the reported evidence in the introduction
section, BDNF and 5-HT posess many molecular crossconections, but previous studies
failed to report strong effects in one system upon deficites of the other. Reciprocal
compensatory mechanisms might be able to counter-regulate single weak points, and this
fits well to clinical studies generally reporting no clearcut evidence of “single-deficite”-
proteins in psychiatric conditions such as depression, in which BDNF and 5-HT are
considered relevant pathogenetic factors. In these disorders, pathophysiological concepts
comprise entaglements between biological vulnerability by genetical predisposition, multi-hit
neuropathological defictes and environmental adversities (Krishnan & Nestler, 2010;

106
Pittenger & Duman, 2008; Schmidt et al, 2011). Both 5-HT and BDNF would provide ample
neurobiological targets connecting these three concepts, yet until now only a very limited
number of studies encompass this phenomenologically heterogenous spectrum. Especially
studies applying imaging genetics and information on environmental adversities could better
unrevel the impact of these two neurophysiologic systems to pathomechanisms of
depression (Agren et al, 2012; Rabl et al, 2014; Witte et al, 2012). With this latter study we
have provided a groundwork and justification of such studies.
A limitation of this project constitutes the still insufficient resolution of neuroimaging
techniques and the lack of knowledge about exact neurobiological correlates of structural
MRI-based methods. Therefore, due to methodological constraints, it is currently not
possible to detect in vivo in humans how exactly 5-HT exerts it’s neuroplastic functions.
Nevertheless, a combination of MRI and PET in multimodal neuroimaging, as applied in the
first study, is elegantly capable of adding molecular information to the same voxel and thus
bringing neuroimaging closer to neurobiology. The next step, would be study human brain
function with combinations of structural, functional and molecular neuroimaging and in
translational animal models with additional post-mortem data, for examples see (Sagi et al,
2012; Vernon et al, 2011). Additional factors might have influenced our results, including low
power in the molecular imaging genetics study, where statistical power is a trade-off to high
efforts in conducting PET studies and subsequent splitting of study subjects into genotype
groups. Finally, we have to state in this section that binding potential results of [carbonyl-
11C]WAY-100635 vary according to the chosen reference region expecially under
consideration of the arterial input function, and that discrepent results between groups are
thought to be susceptible to heterogenous methodological aproaches, for review see
(Shrestha et al, 2012).

107
V. CONCLUSION and FUTURE PERSPECTIVES
This doctoral thesis was conducted to investigate neuroplastic functions of 5-HT in vivo in
healthy humans. By a combination of structural and functional MRI with molecular imaging of
the serotonergic system including imaging genetics, this work assess the links between 5-
HT and neuroplasticity with multiple methodological approaches. The central statements of
this thesis lies in demonstrating the relationship between structural, molecular, functional
and genetic properties of the human serotonergic system as measured with neuroimaging.
While this improved our knowledge on structural and functional properties of this
neurotransmitter system, future work on pathological alterations of serotonin’s neuroplastic
capabilites is needed. While previous knowledge revealed that 5-HT is implicated in the
pathogenesis of psychiatric diseases such as depression, and serotonergic receptors and
the 5-HT transporter are important targets in psychopharmacology, little is known about the
exact pathomechanisms of depression and the mechanisms of action of antidepressants.
Investigations of deficites in 5-HT mediated neuroplasticity in depression could hence
provide a unification of two previously competing major pathogenetic hypotheses, the
“neuroplasticity hypothesis” and the “monoamine hypothesis”.
Additionally, this work gives rise to still unresolved problems regarding the neurobiology
behind MRI-based neuroimaging methods. Bringing neuroimaging closer to neurobiology
could thereby reduce the gap between sophisticated neuroscientific “bench” aproaches in
animals such as optogenetics or stem cell research and low resolution human “bed side” in
vivo methods. Certainly, a better understanding of neurobiological alterations in vivo would
generate big leaps foreward in the understanding and treatment of psychiatric diseases such
as depression.

108
VI. REFERENCES
Agren T, Furmark T, Eriksson E, Fredrikson M (2012) Human fear reconsolidation and allelic differences in serotonergic and dopaminergic genes. Transl Psychiatry 2: e76
Akbari HM, Whitaker-Azmitia PM, Azmitia EC (1994) Prenatal cocaine decreases the trophic factor S-100 beta and induced microcephaly: reversal by postnatal 5-HT1A receptor agonist. Neurosci Lett 170: 141-144
Akimova E, Lanzenberger R, Kasper S (2009) The serotonin-1A receptor in anxiety disorders. Biol Psychiatry 66: 627-635
Asami T, Bouix S, Whitford TJ, Shenton ME, Salisbury DF, McCarley RW (2012) Longitudinal loss of gray matter volume in patients with first-episode schizophrenia: DARTEL automated analysis and ROI validation. Neuroimage 59: 986-996
Ashburner J (2007) A fast diffeomorphic image registration algorithm. Neuroimage 38: 95-113
Ashburner J (2009) Computational anatomy with the SPM software. Magn Reson Imaging 27: 1163-1174
Ashburner J, Friston KJ (2000) Voxel-based morphometry--the methods. NeuroImage 11: 805-821
Ashburner J, Friston KJ (2009) Computing average shaped tissue probability templates. Neuroimage 45: 333-341
Azmitia EC (1999) Serotonin neurons, neuroplasticity, and homeostasis of neural tissue. Neuropsychopharmacology 21: 33S-45S
Azmitia EC (2001) Modern views on an ancient chemical: serotonin effects on cell proliferation, maturation, and apoptosis. Brain research bulletin 56: 413-424
Azmitia EC, Nixon R (2008) Dystrophic serotonergic axons in neurodegenerative diseases. Brain Res 1217: 185-194
Barkas L, Redhead E, Taylor M, Shtaya A, Hamilton DA, Gray WP (2012) Fluoxetine restores spatial learning but not accelerated forgetting in mesial temporal lobe epilepsy. Brain 135: 2358-2374
Benninghoff J, van der Ven A, Schloesser RJ, Moessner R, Moller HJ, Rujescu D (2012) The complex role of the serotonin transporter in adult neurogenesis and neuroplasticity. A critical review. World J Biol Psychiatry 13: 240-247

109
Berg JM, Tymoczko JL, Stryer L (2002) Biochemistry, 5th ed edn. New York: W.H. Freeman.
Billard T, Le Bars D, Zimmer L (2014) PET radiotracers for molecular imaging of serotonin 5-HT1A receptors. Curr Med Chem 21: 70-81
Black IB (1999) Trophic regulation of synaptic plasticity. J Neurobiol 41: 108-118
Boersma MC, Dresselhaus EC, De Biase LM, Mihalas AB, Bergles DE, Meffert MK (2011) A requirement for nuclear factor-kappaB in developmental and plasticity-associated synaptogenesis. J Neurosci 31: 5414-5425
Bonnin A, Peng W, Hewlett W, Levitt P (2006) Expression mapping of 5-HT1 serotonin receptor subtypes during fetal and early postnatal mouse forebrain development. Neuroscience 141: 781-794
Bookstein FL (2001) "Voxel-based morphometry" should not be used with imperfectly registered images. Neuroimage 14: 1454-1462
Borrelli E, Nestler EJ, Allis CD, Sassone-Corsi P (2008) Decoding the epigenetic language of neuronal plasticity. Neuron 60: 961-974
Brans RG, Kahn RS, Schnack HG, van Baal GC, Posthuma D, van Haren NE, Lepage C, Lerch JP, Collins DL, Evans AC, Boomsma DI, Hulshoff Pol HE (2010) Brain plasticity and intellectual ability are influenced by shared genes. J Neurosci 30: 5519-5524
Castrén E (2005) Is mood chemistry? Nature reviews Neuroscience 6: 241-246
Chao MV (2003) Neurotrophins and their receptors: a convergence point for many signalling pathways. Nat Rev Neurosci 4: 299-309
Chen ZY, Patel PD, Sant G, Meng CX, Teng KK, Hempstead BL, Lee FS (2004) Variant brain-derived neurotrophic factor (BDNF) (Met66) alters the intracellular trafficking and activity-dependent secretion of wild-type BDNF in neurosecretory cells and cortical neurons. J Neurosci 24: 4401-4411
Chételat G, Desgranges B, Landeau B, Mézenge F, Poline JB, de la Sayette V, Viader F, Eustache F, Baron J-C (2008) Direct voxel-based comparison between grey matter hypometabolism and atrophy in Alzheimer's disease. Brain 131: 60-71
Chollet F, Tardy J, Albucher JF, Thalamas C, Berard E, Lamy C, Bejot Y, Deltour S, Jaillard A, Niclot P, Guillon B, Moulin T, Marque P, Pariente J, Arnaud C, Loubinoux I (2011) Fluoxetine for motor recovery after acute ischaemic stroke (FLAME): a randomised placebo-controlled trial. Lancet Neurol 10: 123-130

110
Chow DK, Groszer M, Pribadi M, Machniki M, Carmichael ST, Liu X, Trachtenberg JT (2009) Laminar and compartmental regulation of dendritic growth in mature cortex. Nat Neurosci 12: 116-118
Citri A, Malenka RC (2008) Synaptic plasticity: multiple forms, functions, and mechanisms. Neuropsychopharmacology 33: 18-41
Cowen DS (2007) Serotonin and neuronal growth factors - a convergence of signaling pathways. J Neurochem 101: 1161-1171
Cramer SC, Sur M, Dobkin BH, O'Brien C, Sanger TD, Trojanowski JQ, Rumsey JM, Hicks R, Cameron J, Chen D, Chen WG, Cohen LG, deCharms C, Duffy CJ, Eden GF, Fetz EE, Filart R, Freund M, Grant SJ, Haber S, Kalivas PW, Kolb B, Kramer AF, Lynch M, Mayberg HS, McQuillen PS, Nitkin R, Pascual-Leone A, Reuter-Lorenz P, Schiff N, Sharma A, Shekim L, Stryker M, Sullivan EV, Vinogradov S (2011) Harnessing neuroplasticity for clinical applications. Brain 134: 1591-1609
Daubert EA, Condron BG (2010) Serotonin: a regulator of neuronal morphology and circuitry. Trends Neurosci 33: 424-434
Daubert EA, Heffron DS, Mandell JW, Condron BG (2010) Serotonergic dystrophy induced by excess serotonin. Mol Cell Neurosci 44: 297-306
Dayan E, Cohen LG (2011) Neuroplasticity subserving motor skill learning. Neuron 72: 443-454
De Paola V, Holtmaat A, Knott G, Song S, Wilbrecht L, Caroni P, Svoboda K (2006) Cell type-specific structural plasticity of axonal branches and boutons in the adult neocortex. Neuron 49: 861-875
Delon-Martin C, Plailly J, Fonlupt P, Veyrac A, Royet JP (2013) Perfumers' expertise induces structural reorganization in olfactory brain regions. NeuroImage 68: 55-62
Dorsaint-Pierre R, Penhune VB, Watkins KE, Neelin P, Lerch JP, Bouffard M, Zatorre RJ (2006) Asymmetries of the planum temporale and Heschl's gyrus: relationship to language lateralization. Brain 129: 1164-1176
Draganski B, Gaser C, Busch V, Schuierer G, Bogdahn U, May A (2004) Neuroplasticity: changes in grey matter induced by training. Nature 427: 311-312
Draganski B, Gaser C, Kempermann G, Kuhn HG, Winkler J, Buchel C, May A (2006) Temporal and spatial dynamics of brain structure changes during extensive learning. J Neurosci 26: 6314-6317

111
Draganski B, Kherif F (2013) In vivo assessment of use-dependent brain plasticity-Beyond the "one trick pony" imaging strategy. NeuroImage 73: 255-259
Draganski B, May A (2008) Training-induced structural changes in the adult human brain. Behav Brain Res 192: 137-142
Duncan NW, Gravel P, Wiebking C, Reader AJ, Northoff G (2013) Grey matter density and GABAA binding potential show a positive linear relationship across cortical regions. Neuroscience 235: 226-231
Egan MF, Kojima M, Callicott JH, Goldberg TE, Kolachana BS, Bertolino A, Zaitsev E, Gold B, Goldman D, Dean M, Lu B, Weinberger DR (2003) The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 112: 257-269
Erickson KI (2013) Evidence for structural plasticity in humans: Comment on Thomas and Baker (2012). NeuroImage 73: 237-238
Eshaghi A, Bodini B, Ridgway GR, Garcia-Lorenzo D, Tozer DJ, Sahraian MA, Thompson AJ, Ciccarelli O (2014) Temporal and spatial evolution of grey matter atrophy in primary progressive multiple sclerosis. Neuroimage 86: 257-264
Fields RD (2013) Changes in brain structure during learning: Fact or artifact? Reply to Thomas and Baker. NeuroImage 73: 260-264
Franklin TR, Wang Z, Shin J, Jagannathan K, Suh JJ, Detre JA, O'Brien CP, Childress AR (2013) A VBM study demonstrating 'apparent' effects of a single dose of medication on T1-weighted MRIs. Brain Struct Funct 218: 97-104
Frodl T, Koutsouleris N, Bottlender R, Born C, Jäger M, Mörgenthaler M, Scheuerecker J, Zill P, Baghai T, Schüle C, Rupprecht R, Bondy B, Reiser M, Möller H-J, Meisenzahl EM (2008) Reduced gray matter brain volumes are associated with variants of the serotonin transporter gene in major depression. Mol Psychiatry 13: 1093-1101
Gaser C, Schlaug G (2003) Brain structures differ between musicians and non-musicians. J Neurosci 23: 9240-9245
Gaspar P, Cases O, Maroteaux L (2003) The developmental role of serotonin: news from mouse molecular genetics. Nat Rev Neurosci 4: 1002-1012
Gentry JJ, Barker PA, Carter BD (2004) The p75 neurotrophin receptor: multiple interactors and numerous functions. Prog Brain Res 146: 25-39

112
Getz A, Xu F, Zaidi W, Syed NI (2011) The antidepressant fluoxetine but not citalopram suppresses synapse formation and synaptic transmission between Lymnaea neurons by perturbing presynaptic and postsynaptic machinery. Eur J Neurosci 34: 221-234
Good CD, Johnsrude IS, Ashburner J, Henson RN, Friston KJ, Frackowiak RS (2001) A voxel-based morphometric study of ageing in 465 normal adult human brains. Neuroimage 14: 21-36
Greve DN, Svarer C, Fisher PM, Feng L, Hansen AE, Baare W, Rosen B, Fischl B, Knudsen GM (2014) Cortical surface-based analysis reduces bias and variance in kinetic modeling of brain PET data. Neuroimage 92: 225-236
Gunn RN, Gunn SR, Cunningham VJ (2001) Positron emission tomography compartmental models. J Cereb Blood Flow Metab 21: 635-652
Haacke EM (1999) Magnetic resonance imaging : physical principles and sequence design, New York: Wiley.
Hacker MP, Messer WS, Bachmann KA. (2009) Pharmacology principles and practice. Academic Press/Elsevier,, Amsterdam ; Boston, pp. 1 online resource (xi, 594 p.).
Haeusler D, Mien LK, Nics L, Ungersboeck J, Philippe C, Lanzenberger RR, Kletter K, Dudczak R, Mitterhauser M, Wadsak W (2009) Simple and rapid preparation of [11C]DASB with high quality and reliability for routine applications. Appl Radiat Isot 67: 1654-1660
Hajszan T, MacLusky NJ, Leranth C (2005) Short-term treatment with the antidepressant fluoxetine triggers pyramidal dendritic spine synapse formation in rat hippocampus. Eur J Neurosci 21: 1299-1303
Henningsson S, Borg J, Lundberg J, Bah J, Lindström M, Ryding E, Jovanovic H, Saijo T, Inoue M, Rosén I, Träskman-Bendz L, Farde L, Eriksson E (2009) Genetic variation in brain-derived neurotrophic factor is associated with serotonin transporter but not serotonin-1A receptor availability in men. Biological Psychiatry 66: 477-485
Hodges MR, Wehner M, Aungst J, Smith JC, Richerson GB (2009) Transgenic mice lacking serotonin neurons have severe apnea and high mortality during development. J Neurosci 29: 10341-10349
Hoexter MQ, de Souza Duran FL, D'Alcante CC, Dougherty DD, Shavitt RG, Lopes AC, Diniz JB, Deckersbach T, Batistuzzo MC, Bressan RA, Miguel EC, Busatto GF (2012) Gray matter volumes in obsessive-compulsive disorder before and after fluoxetine or cognitive-behavior therapy: a randomized clinical trial. Neuropsychopharmacology 37: 734-745
Hoflich A, Baldinger P, Savli M, Lanzenberger R, Kasper S (2012) Imaging treatment effects in depression. Rev Neurosci 23: 227-252

113
Holtmaat A, Randall J, Cane M (2013) Optical imaging of structural and functional synaptic plasticity in vivo. Eur J Pharmacol 719: 128-136
Holtmaat A, Svoboda K (2009) Experience-dependent structural synaptic plasticity in the mammalian brain. Nat Rev Neurosci 10: 647-658
Homberg JR (2012) Serotonin and decision making processes. Neurosci Biobehav Rev 36: 218-236
Hornung JP (2010) The neuroanatomy of the serotonergic system. In Handbook of the behavioral neurobiology of serotonin, pp 51-64. London: Academic
Houle S, Ginovart N, Hussey D, Meyer JH, Wilson AA (2000) Imaging the serotonin transporter with positron emission tomography: initial human studies with [11C]DAPP and [11C]DASB. Eur J Nucl Med 27: 1719-1722
Huang Y, Zheng MQ, Gerdes JM (2010) Development of effective PET and SPECT imaging agents for the serotonin transporter: has a twenty-year journey reached its destination? Current topics in medicinal chemistry 10: 1499-1526
Hyde KL, Lerch J, Norton A, Forgeard M, Winner E, Evans AC, Schlaug G (2009) The effects of musical training on structural brain development: a longitudinal study. Ann N Y Acad Sci 1169: 182-186
Ichise M, Meyer JH, Yonekura Y (2001) An introduction to PET and SPECT neuroreceptor quantification models. J Nucl Med 42: 755-763
Innis RB, Cunningham VJ, Delforge J, Fujita M, Gjedde A, Gunn RN, Holden J, Houle S, Huang SC, Ichise M, Iida H, Ito H, Kimura Y, Koeppe RA, Knudsen GM, Knuuti J, Lammertsma AA, Laruelle M, Logan J, Maguire RP, Mintun MA, Morris ED, Parsey R, Price JC, Slifstein M, Sossi V, Suhara T, Votaw JR, Wong DF, Carson RE (2007) Consensus nomenclature for in vivo imaging of reversibly binding radioligands. J Cereb Blood Flow Metab 27: 1533-1539
Ishii K, Sasaki H, Kono AK, Miyamoto N, Fukuda T, Mori E (2005) Comparison of gray matter and metabolic reduction in mild Alzheimer's disease using FDG-PET and voxel-based morphometric MR studies. Eur J Nucl Med Mol Imaging 32: 959-963
Jack CR, Lowe VJ, Senjem ML, Weigand SD, Kemp BJ, Shiung MM, Knopman DS, Boeve BF, Klunk WE, Mathis CA, Petersen RC (2008) 11C PiB and structural MRI provide complementary information in imaging of Alzheimer's disease and amnestic mild cognitive impairment. Brain 131: 665-680
Jeffery KJ, Reid IC (1997) Modifiable neuronal connections: an overview for psychiatrists. Am J Psychiatry 154: 156-164

114
Johansen-Berg H (2012) The future of functionally-related structural change assessment. Neuroimage 62: 1293-1298
Kalpouzos G, Chételat G, Baron J-C, Landeau B, Mevel K, Godeau C, Barré L, Constans J-M, Viader F, Eustache F, Desgranges B (2009) Voxel-based mapping of brain gray matter volume and glucose metabolism profiles in normal aging. Neurobiol Aging 30: 112-124
Kanda T, Ishii K, Uemura T, Miyamoto N, Yoshikawa T, Kono AK, Mori E (2008) Comparison of grey matter and metabolic reductions in frontotemporal dementia using FDG-PET and voxel-based morphometric MR studies. Eur J Nucl Med Mol Imaging 35: 2227-2234
Kandel ER (1981) Calcium and the control of synaptic strength by learning. Nature 293: 697-700
Kandel ER (2001) The molecular biology of memory storage: a dialogue between genes and synapses. Science 294: 1030-1038
Kandel ER (2004) The molecular biology of memory storage: a dialog between genes and synapses. Biosci Rep 24: 475-522
Kandel ER, Schwartz JH (1982) Molecular biology of learning: modulation of transmitter release. Science 218: 433-443
Karabeg MM, Grauthoff S, Kollert SY, Weidner M, Heiming RS, Jansen F, Popp S, Kaiser S, Lesch KP, Sachser N, Schmitt AG, Lewejohann L (2013) 5-HTT deficiency affects neuroplasticity and increases stress sensitivity resulting in altered spatial learning performance in the Morris water maze but not in the Barnes maze. PLoS One 8: e78238
Karpova NN, Pickenhagen A, Lindholm J, Tiraboschi E, Kulesskaya N, Agustsdottir A, Antila H, Popova D, Akamine Y, Bahi A, Sullivan R, Hen R, Drew LJ, Castren E (2011) Fear erasure in mice requires synergy between antidepressant drugs and extinction training. Science 334: 1731-1734
Kasai H, Fukuda M, Watanabe S, Hayashi-Takagi A, Noguchi J (2010) Structural dynamics of dendritic spines in memory and cognition. Trends Neurosci 33: 121-129
Kempton MJ, Salvador Z, Munafo MR, Geddes JR, Simmons A, Frangou S, Williams SC (2011) Structural neuroimaging studies in major depressive disorder. Meta-analysis and comparison with bipolar disorder. Arch Gen Psychiatry 68: 675-690
Klein A, Andersson J, Ardekani BA, Ashburner J, Avants B, Chiang M-C, Christensen GE, Collins DL, Gee J, Hellier P, Song JH, Jenkinson M, Lepage C, Rueckert D, Thompson P, Vercauteren T, Woods RP, Mann JJ, Parsey RV (2009) Evaluation of 14 nonlinear deformation algorithms applied to human brain MRI registration. Neuroimage 46: 786-802

115
Koponen E, Rantamaki T, Voikar V, Saarelainen T, MacDonald E, Castren E (2005) Enhanced BDNF signaling is associated with an antidepressant-like behavioral response and changes in brain monoamines. Cell Mol Neurobiol 25: 973-980
Kozisek ME, Middlemas D, Bylund DB (2008) Brain-derived neurotrophic factor and its receptor tropomyosin-related kinase B in the mechanism of action of antidepressant therapies. Pharmacol Ther 117: 30-51
Kranz GS, Kasper S, Lanzenberger R (2010) Reward and the serotonergic system. Neuroscience 166: 1023-1035
Krishnan V, Nestler EJ (2010) Linking molecules to mood: new insight into the biology of depression. Am J Psychiatry 167: 1305-1320
la Fougère C, Grant S, Kostikov A, Schirrmacher R, Gravel P, Schipper H, Reader A, Evans A, Thiel A (2010) Where in-vivo imaging meets cytoarchitectonics: The relationship between cortical thickness and neuronal density measured with high-resolution [(18)F]flumazenil-PET. Neuroimage
Lan MJ, Ogden RT, Huang YY, Oquendo MA, Sullivan GM, Miller J, Milak M, Mann JJ, Parsey RV (2014) Genetic variation in brain-derived neurotrophic factor val66met allele is associated with altered serotonin-1A receptor binding in human brain. NeuroImage
Laruelle M (2000) Imaging synaptic neurotransmission with in vivo binding competition techniques: a critical review. J Cereb Blood Flow Metab 20: 423-451
Laruelle M, Slifstein M, Huang Y (2003) Relationships between radiotracer properties and image quality in molecular imaging of the brain with positron emission tomography. Mol Imaging Biol 5: 363-375
Lenglet C, Abosch A, Yacoub E, De Martino F, Sapiro G, Harel N (2012) Comprehensive in vivo mapping of the human basal ganglia and thalamic connectome in individuals using 7T MRI. PLoS One 7: e29153
Lesch KP, Waider J (2012) Serotonin in the modulation of neural plasticity and networks: implications for neurodevelopmental disorders. Neuron 76: 175-191
Leventer RJ, Jansen A, Pilz DT, Stoodley N, Marini C, Dubeau F, Malone J, Mitchell LA, Mandelstam S, Scheffer IE, Berkovic SF, Andermann F, Andermann E, Guerrini R, Dobyns WB (2010) Clinical and imaging heterogeneity of polymicrogyria: a study of 328 patients. Brain 133: 1415-1427
Logan J, Fowler JS, Volkow ND, Wang GJ, Ding YS, Alexoff DL (1996) Distribution volume ratios without blood sampling from graphical analysis of PET data. J Cereb Blood Flow Metab 16: 834-840

116
Lusebrink F, Wollrab A, Speck O (2013) Cortical thickness determination of the human brain using high resolution 3T and 7T MRI data. Neuroimage 70: 122-131
Macqueen G, Frodl T (2010) The hippocampus in major depression: evidence for the convergence of the bench and bedside in psychiatric research? Mol Psychiatry
Maguire EA, Gadian DG, Johnsrude IS, Good CD, Ashburner J, Frackowiak RS, Frith CD (2000) Navigation-related structural change in the hippocampi of taxi drivers. Proc Natl Acad Sci U S A 97: 4398-4403
Maguire EA, Woollett K, Spiers HJ (2006) London taxi drivers and bus drivers: a structural MRI and neuropsychological analysis. Hippocampus 16: 1091-1101
Mahar I, Bambico FR, Mechawar N, Nobrega JN (2014) Stress, serotonin, and hippocampal neurogenesis in relation to depression and antidepressant effects. Neurosci Biobehav Rev 38: 173-192
Malenka RC, Nicoll RA (1999) Long-term potentiation--a decade of progress? Science 285: 1870-1874
Marques JP, Kober T, Krueger G, van der Zwaag W, Van de Moortele P-F, Gruetter R (2010) MP2RAGE, a self bias-field corrected sequence for improved segmentation and T1-mapping at high field. Neuroimage 49: 1271-1281
Martel J-C, Leduc N, Ormière A-M, Faucillon V, Danty N, Culie C, Cussac D, Newman-Tancredi A (2007) WAY-100635 has high selectivity for serotonin 5-HT(1A) versus dopamine D(4) receptors. Eur J Pharmacol 574: 15-19
Martensson J, Eriksson J, Bodammer NC, Lindgren M, Johansson M, Nyberg L, Lovden M (2012) Growth of language-related brain areas after foreign language learning. NeuroImage 63: 240-244
Mathew RJ, Partain CL (1985) Midsagittal sections of the cerebellar vermis and fourth ventricle obtained with magnetic resonance imaging of schizophrenic patients. Am J Psychiatry 142: 970-971
Mattson MP (2008) Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann N Y Acad Sci 1144: 97-112
Matuskey D, Pittman B, Planeta-Wilson B, Walderhaug E, Henry S, Gallezot JD, Nabulsi N, Ding YS, Bhagwagar Z, Malison R, Carson RE, Neumeister A (2012) Age effects on serotonin receptor 1B as assessed by PET. J Nucl Med 53: 1411-1414

117
May A (2011) Experience-dependent structural plasticity in the adult human brain. Trends Cogn Sci 15: 475-482
May A, Hajak G, Ganssbauer S, Steffens T, Langguth B, Kleinjung T, Eichhammer P (2006) Structural Brain Alterations following 5 Days of Intervention: Dynamic Aspects of Neuroplasticity. Cerebral Cortex 17: 205-210
McClung CA, Nestler EJ (2008) Neuroplasticity mediated by altered gene expression. Neuropsychopharmacology 33: 3-17
Mead GE, Hsieh CF, Lee R, Kutlubaev MA, Claxton A, Hankey GJ, Hackett ML (2012) Selective serotonin reuptake inhibitors (SSRIs) for stroke recovery. The Cochrane database of systematic reviews 11: CD009286
Mehta M, Ahmed Z, Fernando SS, Cano-Sanchez P, Adayev T, Ziemnicka D, Wieraszko A, Banerjee P (2007) Plasticity of 5-HT 1A receptor-mediated signaling during early postnatal brain development. J Neurochem 101: 918-928
Michaelis L. MML (1913) Die Kinetik der Invertinwirkung. Biochem Z: 333-336
Mintun MA, Raichle ME, Kilbourn MR, Wooten GF, Welch MJ (1984) A quantitative model for the in vivo assessment of drug binding sites with positron emission tomography. Annals of neurology 15: 217-227
Moiseiwitsch JR, Lauder JM (1995) Serotonin regulates mouse cranial neural crest migration. Proc Natl Acad Sci U S A 92: 7182-7186
Mugler JP, 3rd, Brookeman JR (1990) Three-dimensional magnetization-prepared rapid gradient-echo imaging (3D MP RAGE). Magnetic resonance in medicine : official journal of the Society of Magnetic Resonance in Medicine / Society of Magnetic Resonance in Medicine 15: 152-157
Nestler EJ, Barrot M, DiLeone RJ, Eisch AJ, Gold SJ, Monteggia LM (2002) Neurobiology of depression. Neuron 34: 13-25
Nibuya M, Morinobu S, Duman RS (1995) Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J Neurosci 15: 7539-7547
Nitsche MA, Kuo M-F, Karrasch R, Wächter B, Liebetanz D, Paulus W (2009) Serotonin affects transcranial direct current-induced neuroplasticity in humans. Biol Psychiatry 66: 503-508
Oh H, Mormino EC, Madison C, Hayenga A, Smiljic A, Jagust WJ (2010) β-Amyloid affects frontal and posterior brain networks in normal aging. Neuroimage

118
Parsey RV, Slifstein M, Hwang DR, Abi-Dargham A, Simpson N, Mawlawi O, Guo NN, Van Heertum R, Mann JJ, Laruelle M (2000) Validation and reproducibility of measurement of 5-HT1A receptor parameters with [carbonyl-11C]WAY-100635 in humans: comparison of arterial and reference tisssue input functions. J Cereb Blood Flow Metab 20: 1111-1133
Paterson LM, Tyacke RJ, Nutt DJ, Knudsen GM (2010) Measuring endogenous 5-HT release by emission tomography: promises and pitfalls. J Cereb Blood Flow Metab 30: 1682-1706
Pereira AC, Huddleston DE, Brickman AM, Sosunov AA, Hen R, McKhann GM, Sloan R, Gage FH, Brown TR, Small SA (2007) An in vivo correlate of exercise-induced neurogenesis in the adult dentate gyrus. Proc Natl Acad Sci U S A 104: 5638-5643
Pike VW (2009) PET radiotracers: crossing the blood-brain barrier and surviving metabolism. Trends Pharmacol Sci 30: 431-440
Pilar-Cuellar F, Vidal R, Diaz A, Castro E, dos Anjos S, Pascual-Brazo J, Linge R, Vargas V, Blanco H, Martinez-Villayandre B, Pazos A, Valdizan EM (2013) Neural plasticity and proliferation in the generation of antidepressant effects: hippocampal implication. Neural plasticity 2013: 537265
Pittenger C, Duman RS (2008) Stress, depression, and neuroplasticity: a convergence of mechanisms. Neuropsychopharmacology 33: 88-109
Piubelli C, Vighini M, Mathe AA, Domenici E, Carboni L (2011) Escitalopram affects cytoskeleton and synaptic plasticity pathways in a rat gene-environment interaction model of depression as revealed by proteomics. Part II: environmental challenge. Int J Neuropsychopharmacol 14: 834-855
Polter AM, Li X (2010) 5-HT1A receptor-regulated signal transduction pathways in brain. Cellular Signalling 22: 1406-1412
Protopopescu X, Butler T, Pan H, Root J, Altemus M, Polanecsky M, McEwen B, Silbersweig D, Stern E (2008) Hippocampal structural changes across the menstrual cycle. Hippocampus 18: 985-988
Rabl U, Meyer BM, Diers K, Bartova L, Berger A, Mandorfer D, Popovic A, Scharinger C, Huemer J, Kalcher K, Pail G, Haslacher H, Perkmann T, Windischberger C, Brocke B, Sitte HH, Pollak DD, Dreher JC, Kasper S, Praschak-Rieder N, Moser E, Esterbauer H, Pezawas L (2014) Additive gene-environment effects on hippocampal structure in healthy humans. J Neurosci 34: 9917-9926
Rami-Mark C, Ungersboeck J, Haeusler D, Nics L, Philippe C, Mitterhauser M, Willeit M, Lanzenberger R, Karanikas G, Wadsak W (2013) Reliable set-up for in-loop (1)(1)C-carboxylations using Grignard reactions for the preparation of [carbonyl-(1)(1)C]WAY-100635 and [(1)(1)C]-(+)-PHNO. Appl Radiat Isot 82: 75-80

119
Rantamaki T, Hendolin P, Kankaanpaa A, Mijatovic J, Piepponen P, Domenici E, Chao MV, Mannisto PT, Castren E (2007) Pharmacologically diverse antidepressants rapidly activate brain-derived neurotrophic factor receptor TrkB and induce phospholipase-Cgamma signaling pathways in mouse brain. Neuropsychopharmacology 32: 2152-2162
Rapoport JL, Gogtay N (2008) Brain neuroplasticity in healthy, hyperactive and psychotic children: insights from neuroimaging. Neuropsychopharmacology 33: 181-197
Riccio O, Potter G, Walzer C, Vallet P, Szabó G, Vutskits L, Kiss JZ, Dayer AG (2009) Excess of serotonin affects embryonic interneuron migration through activation of the serotonin receptor 6. Mol Psychiatry 14: 280-290
Rubenstein JL (1998) Development of serotonergic neurons and their projections. Biol Psychiatry 44: 145-150
Sagi Y, Tavor I, Hofstetter S, Tzur-Moryosef S, Blumenfeld-Katzir T, Assaf Y (2012) Learning in the fast lane: new insights into neuroplasticity. Neuron 73: 1195-1203
Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S, Weisstaub N, Lee J, Duman R, Arancio O, Belzung C, Hen R (2003) Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 301: 805-809
Sarkamo T, Ripolles P, Vepsalainen H, Autti T, Silvennoinen HM, Salli E, Laitinen S, Forsblom A, Soinila S, Rodriguez-Fornells A (2014) Structural changes induced by daily music listening in the recovering brain after middle cerebral artery stroke: a voxel-based morphometry study. Front Hum Neurosci 8: 245
Saulin A, Savli M, Lanzenberger R (2012) Serotonin and molecular neuroimaging in humans using PET. Amino Acids 42: 2039-2057
Savli M, Bauer A, Mitterhauser M, Ding YS, Hahn A, Kroll T, Neumeister A, Haeusler D, Ungersboeck J, Henry S, Isfahani SA, Rattay F, Wadsak W, Kasper S, Lanzenberger R (2012) Normative database of the serotonergic system in healthy subjects using multi-tracer PET. NeuroImage 63: 447-459
Schlaug G, Norton A, Overy K, Winner E (2005) Effects of music training on the child's brain and cognitive development. Ann N Y Acad Sci 1060: 219-230
Schmidt HD, Shelton RC, Duman RS (2011) Functional biomarkers of depression: diagnosis, treatment, and pathophysiology. Neuropsychopharmacology 36: 2375-2394
Schneider F, Fink GR (2013) Funktionelle MRT in Psychiatrie und Neurologie, 2., überarbeitete und aktualisierte Auflage 2013 edn. Berlin, Heidelberg: Springer.

120
Scholz J, Klein MC, Behrens TEJ, Johansen-Berg H (2009) Training induces changes in white-matter architecture. Nat Neurosci 12: 1370-1371
Selvaraj S, Turkheimer F, Rosso L, Faulkner P, Mouchlianitis E, Roiser JP, McGuire P, Cowen PJ, Howes O (2012) Measuring endogenous changes in serotonergic neurotransmission in humans: a [11C]CUMI-101 PET challenge study. Mol Psychiatry 17: 1254-1260
Serra-Millàs M, López-Vílchez I, Navarro V, Galán A-M, Escolar G, Penadés R, Catalán R, Fañanás L, Arias B, Gastó C (2011) Changes in plasma and platelet BDNF levels induced by S-citalopram in major depression. Psychopharmacology
Shrestha S, Hirvonen J, Hines CS, Henter ID, Svenningsson P, Pike VW, Innis RB (2012) Serotonin-1A receptors in major depression quantified using PET: Controversies, confounds, and recommendations. NeuroImage 59: 3243-3251
Singh C, Bortolato M, Bali N, Godar SC, Scott AL, Chen K, Thompson RF, Shih JC (2013) Cognitive abnormalities and hippocampal alterations in monoamine oxidase A and B knockout mice. Proc Natl Acad Sci U S A 110: 12816-12821
Slifstein M, Parsey RV, Laruelle M (2000) Derivation of [(11)C]WAY-100635 binding parameters with reference tissue models: effect of violations of model assumptions. Nucl Med Biol 27: 487-492
Smith R, Chen K, Baxter L, Fort C, Lane RD (2012) Antidepressant effects of sertraline associated with volume increases in dorsolateral prefrontal cortex. J Affect Disord
Tanti A, Rainer Q, Minier F, Surget A, Belzung C (2012) Differential environmental regulation of neurogenesis along the septo-temporal axis of the hippocampus. Neuropharmacology 63: 374-384
Tardito D, Perez J, Tiraboschi E, Musazzi L, Racagni G, Popoli M (2006) Signaling pathways regulating gene expression, neuroplasticity, and neurotrophic mechanisms in the action of antidepressants: a critical overview. Pharmacol Rev 58: 115-134
Thomas C, Baker CI (2013) Teaching an adult brain new tricks: A critical review of evidence for training-dependent structural plasticity in humans. NeuroImage 73: 225-236
Tost H, Braus DF, Hakimi S, Ruf M, Vollmert C, Hohn F, Meyer-Lindenberg A (2010) Acute D2 receptor blockade induces rapid, reversible remodeling in human cortical-striatal circuits. Nat Neurosci 13: 920-922
Trachtenberg JT, Chen BE, Knott GW, Feng G, Sanes JR, Welker E, Svoboda K (2002) Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex. Nature 420: 788-794

121
Turkheimer FE, Veronese M, Dunn J (2014) Experimental design and practical data analysis in positron emission tomography.
Upton AL, Salichon N, Lebrand C, Ravary A, Blakely R, Seif I, Gaspar P (1999) Excess of serotonin (5-HT) alters the segregation of ispilateral and contralateral retinal projections in monoamine oxidase A knock-out mice: possible role of 5-HT uptake in retinal ganglion cells during development. J Neurosci 19: 7007-7024
van Tol M-J, van der Wee NJA, van den Heuvel OA, Nielen MMA, Demenescu LR, Aleman A, Renken R, van Buchem MA, Zitman FG, Veltman DJ (2010) Regional brain volume in depression and anxiety disorders. Arch Gen Psychiatry 67: 1002-1011
Vermetten E, Vythilingam M, Southwick SM, Charney DS, Bremner JD (2003) Long-term treatment with paroxetine increases verbal declarative memory and hippocampal volume in posttraumatic stress disorder. Biol Psychiatry 54: 693-702
Vernon AC, Natesan S, Modo M, Kapur S (2011) Effect of chronic antipsychotic treatment on brain structure: a serial magnetic resonance imaging study with ex vivo and postmortem confirmation. Biol Psychiatry 69: 936-944
Vetencourt JFM, Sale A, Viegi A, Baroncelli L, De Pasquale R, O'Leary OF, Castrén E, Maffei L (2008) The antidepressant fluoxetine restores plasticity in the adult visual cortex. Science 320: 385-388
Vetencourt JFM, Tiraboschi E, Spolidoro M, Castrén E, Maffei L (2011) Serotonin triggers a transient epigenetic mechanism that reinstates adult visual cortex plasticity in rats. Eur J Neurosci 33: 49-57
Villemagne VL, Burnham S, Bourgeat P, Brown B, Ellis KA, Salvado O, Szoeke C, Macaulay SL, Martins R, Maruff P, Ames D, Rowe CC, Masters CL, Australian Imaging B, Lifestyle Research G (2013) Amyloid beta deposition, neurodegeneration, and cognitive decline in sporadic Alzheimer's disease: a prospective cohort study. Lancet Neurol 12: 357-367
Vitalis T, Cases O, Gillies K, Hanoun N, Hamon M, Seif I, Gaspar P, Kind P, Price DJ (2002) Interactions between TrkB signaling and serotonin excess in the developing murine somatosensory cortex: a role in tangential and radial organization of thalamocortical axons. J Neurosci 22: 4987-5000
Vitalis T, Cases O, Passemard S, Callebert J, Parnavelas JG (2007) Embryonic depletion of serotonin affects cortical development. Eur J Neurosci 26: 331-344
Wadsak W. MLK, Ettlinger D.E., Lanzenberger R., Haeusler D., Dudczak R., Kletter K., Mitterhauser M. (2007) Simple and fully automated preparation of [carbonyl-11C]WAY-100635. Radiochimica Acta 95: 1-6

122
Warner-Schmidt JL, Chen EY, Zhang X, Marshall JJ, Morozov A, Svenningsson P, Greengard P (2010) A role for p11 in the antidepressant action of brain-derived neurotrophic factor. Biological Psychiatry 68: 528-535
Weinberger DR, Bigelow LB, Kleinman JE, Klein ST, Rosenblatt JE, Wyatt RJ (1980) Cerebral ventricular enlargement in chronic schizophrenia. An association with poor response to treatment. Arch Gen Psychiatry 37: 11-13
Weinberger DR, Torrey EF, Wyatt RJ (1979) Cerebellar atrophy in chronic schizophrenia. Lancet 1: 718-719
Weishaupt D, Köchli VD, Marincek B (2006) How does MRI work? : an introduction to the physics and function of magnetic resonance imaging, 2nd edn. Berlin ; New York: Springer.
Whitaker-Azmitia PM (1998) Role of the neurotrophic properties of serotonin in the delay of brain maturation induced by cocaine. Ann N Y Acad Sci 846: 158-164
Whitaker-Azmitia PM (2001) Serotonin and brain development: role in human developmental diseases. Brain research bulletin 56: 479-485
Whitaker-Azmitia PM, Druse M, Walker P, Lauder JM (1996) Serotonin as a developmental signal. Behav Brain Res 73: 19-29
Wilson CC, Faber KM, Haring JH (1998) Serotonin regulates synaptic connections in the dentate molecular layer of adult rats via 5-HT1a receptors: evidence for a glial mechanism. Brain Res 782: 235-239
Witte AV, Kurten J, Jansen S, Schirmacher A, Brand E, Sommer J, Floel A (2012) Interaction of BDNF and COMT polymorphisms on paired-associative stimulation-induced cortical plasticity. J Neurosci 32: 4553-4561
Wonderlick JS, Ziegler DA, Hosseini-Varnamkhasti P, Locascio JJ, Bakkour A, van der Kouwe A, Triantafyllou C, Corkin S, Dickerson BC (2009) Reliability of MRI-derived cortical and subcortical morphometric measures: effects of pulse sequence, voxel geometry, and parallel imaging. Neuroimage 44: 1324-1333
Woodward ND, Zald DH, Ding Z, Riccardi P, Ansari MS, Baldwin RM, Cowan RL, Li R, Kessler RM (2009) Cerebral morphology and dopamine D2/D3 receptor distribution in humans: a combined [18F]fallypride and voxel-based morphometry study. Neuroimage 46: 31-38
Wright IC, McGuire PK, Poline JB, Travere JM, Murray RM, Frith CD, Frackowiak RS, Friston KJ (1995) A voxel-based method for the statistical analysis of gray and white matter density applied to schizophrenia. Neuroimage 2: 244-252

123
Wu S, Ogden RT, Mann JJ, Parsey RV (2007) Optimal metabolite curve fitting for kinetic modeling of 11C-WAY-100635. J Nucl Med 48: 926-931
Yan Y, Sheppard PC, Kasper S, Lin L, Hoare S, Kapoor A, Dodd JG, Duckworth ML, Matusik RJ (1997) Large fragment of the probasin promoter targets high levels of transgene expression to the prostate of transgenic mice. Prostate 32: 129-139
Zatorre RJ, Fields RD, Johansen-Berg H (2012) Plasticity in gray and white: neuroimaging changes in brain structure during learning. Nat Neurosci 15: 528-536

124
APPENDIX – CURRICULUM VITAE
Dr. med. univ. Christoph Kraus
Schilfgasse 2, 2452 Mannersdorf Weinbergstraße 6, 7100 Neusiedl am See mobile: +43 664 4147931 Email: [email protected] Department of Psychiatry and Psychotherapy Medical University of Vienna Waehringer Guertel 18-20 A – 1090 Vienna, Austria
Personal Details: 1.7.1983 born in Eisenstadt
unmarried
Austrian
Primary Education:
1989-1993 Primary School
1993-2001 Gymnasium Neusiedl am See, Matura, 7.6.2001
Higher Education:
2002-2009 Medical Studies at the Medical University of Vienna
2007-2009 Diploma thesis at the Department of Psychoanalysis and
Psychotherapy under supervison from Ao. Univ. Prof. Mag. Dr. Jandl-
Jager Elisabeth entitled “Violence in childhood and recognition in the
medical system of Austria”.
17.9.2009 M.D. degree of the Medical University of Vienna
2012 – present PhD Thesis in Clinical Neurosciences (N790). Title: Serotonin and
Neuroplasticity – Investigated in vivo by Positron Emission
Tomography and structural Magnetic Resonance Imaging. Supervisor:
A/Prof. Rupert Lanzenberger, Mentors: Prof. Siegfried Kasper, A/Prof.
Wolfgang Wadsak

125
Professional Education:
1.9.2011–present Clinical Training, Medical University of Vienna, Department of
Psychiatry and Psychotherapy
1.10.2012–1.10.2013 Clinical Training, Medical University of Vienna with emphasis on
treatment of forensic psychiatric Patients in the Prison “Justizanstalt
Josefstadt”
1.3.2014–30.9.2014 Psychiatric consultant service at the Vienna General Hospital (AKH)
1.10.2014-present Training in Neurology, Department of Neurology, Medical University of
Vienna, as required for specialist training
Scientific Collaborations:
2009 – present staff member in the „NEUROIMAGING LABs (NIL)“ (head Assoc.-Prof. PD Dr. Rupert Lanzenberger), Department of Psychiatry and Psychotherapy, MUW. Collaboration and Coinvestigator in clinical projects focused on neuroimaging and genetics:
2009 – 2011 Coinvestigator: Networks of Anxiety: Connectivity Analysis in Social Phobia using Functional Magnetic Resonance Imaging, OeNB procet number 12982, EK 619/2007, Principal Investigator: Assoc.-Prof. Priv.-Doz. Dipl.-Ing. Dr. Christian Windischberger, MUW Austria.
2009 – 2011 Coinvestigator: The influence of hormone replacement therapy on the cerebral serotonin-1A receptor distribution and mood in postmenopausal women. A longitudinal study using Positron Emission Tomography (PET) and the radioligand [carbonyl-11C]WAY-100635. Principal Investigator: O. Univ. Prof. Dr. h.c. mult Dr. med. S. Kasper, MUW Austria.
2010 – 2011 Coinvestigator: Effects of electroconvulsive therapy on serotonin-1A receptor binding in major depression. A longitudinal study using Positron Emission Tomography (PET) and the radioligand [carbonyl-11C]WAY-100635. Principal Investigator: Ao. Univ. Prof. Dr. Richard Frey, MUW Austria.
2010 – 2014 Coinvestigator: The influence of sex steroid hormones onserotonin transporter bindingin the human brain investigated by PET. OeNB project number 13214, EK 620/2008. Principal Investigator: Assoc.-Prof. Priv.-Doz. Dr. Rupert Lanzenberger, MUW Austria.
2011 – 2014 Coinvestigator: The Serotonin Transporter in Attention Deficit Hyperactivity Disorder Investigated with Positron Emission Tomography. OeNB project number AP13675ONB, EK 784/2009, Principal Investigator: Priv. Doz. Mag. Dr. Markus Mitterhauser, Austria.

126
2012 – present Coinvestigator: Multimodal Assessment of Neurobiological Markers for Psychiatric Disorders (MAN-BIOPSY). Research cluster “Multimodal Neuroimaging in clinical neurosciences (MMI-CNS), Medical University Vienna, University of Vienna. Principal Investigators: Assoc.-Prof. Priv.-Doz. Dr. Rupert Lanzenberger, Univ. Prof. Mag. Dr. Claus LammPhD
Other Universitary Acitvities:
2008 Initiation of the course: „Medizinsoziologie mit wechselndem Schwerpunkten: Balint-Gruppe, wissenschaftliche Methoden, Traumforschung“, together with Ao. Univ. Prof. Mag. Dr. Jandl-Jager in winterterm 08.
2009 Initiation of the course “Talks about death and dying“, starting in winterterm 09/10, togehter with Univ. Prof. Dr. Pötter, Univ. Prof. Dr. Watzke, Mag. Dr. Hladschik-Kermer, Mag. Kirchheiner
2012 – present Representative of resident psychiatrists at the Department of Psychiatry and Psychotherapy
Further Education:
2009 – 2012 Psychotherapeutic propaedeutics – H.O.P.P. Vienna 2012 Statistical Parametric Mapping Course, Center for Experimental
Medicine, Department of System Neuroscience UKE Eppendorf, Hamburg.
2012 PMOD basic Application and Small Animal Imaging Processing
Courses, Zürich 2013 – present Training in Cognitive Behavioural Therapy
1. FELLOWSHIPS
Young Scientist Association (YSA) of the Medical University of Vienna
Austrian Medical Chamber
Austrian Psychiatric and Psychotherapeutic Association
European College of Neuropsychopharmacology (ECNP)
2. TEACHING
2013 – now Student courses in clinical psychiatry and psychopharmacology at the Medical University of Vienna

127
2014 – now Nurse Training in clinical psychiatry at the School of Nursery of the Viennese Hospital Association
2015 Thesis Completion of Dr. Patrick Köck, Thesis supervision cand. med. Clauddia Winkler
3. REVIEWER FOR MANUSCRIPTS:
Molecules [IF 2014: 2.42]
World Journal of Biological Psychiatry [IF 2014: 4.23]
International Journal of Neuropsychopharmacology [IF 2014: 4.0]
Journal of Psychiatry and Neuroscience [IF 2014: 7.49]
4. PRIZES
Best Poster Award, 7th PhD Sympusium MUW, 2011
Best Poster Award, 8th PhD Symposium MUW, 2012
BSM – ÖGN – Mallinckrodt Förderungspreis Nuklearmedizin, 2012
Rafaelsen Young Investigator’s award by the International College of
Neuropsychopharmacology (CINP) 2014
WFSBP Educational Grant, 2015
ECNP Travel Award, 2015
5. CHAIRS Co-Chair, Serotonin and Neuroplasticity at the WFSBP Conference 2015
Young Programme Sub-committee for the Seoul Congress
6. SCIENTIFIC FIELD OF WORK
Serotonin and Neuroplasticity, Neuroimaging of Neuropsychiatric disorders: ADHD,
Depression, Anxiety Disorders, Genetics in Neuroimaing

128
7. PUBLICATION LIST
7.1 Articles
7.1.1 1ST Authorships:
1. Baldinger P, Kraus C, Rami-Mark C, Gryglewsky G, Kranz GS, Haeusler D, Hahn A, Wadsak W, Mitterhauser M, Rujescu D, Kasper S, Lanzenberger R. Interaction between 5-HTTLPR and 5-HT1B genotype status enhances serotonin-1A receptor binding. NeuroImage 2015 May 15;111:505-512. Epub 2015, [2014, IF: 6.36]
2. Kraus C, Baldinger P, Rami-Mark C, Gryglewsky G, Kranz GS, Haeusler D, Hahn A, Wadsak W, Mitterhauser M, Rujescu D, Kasper S, Lanzenberger R. Exploring the impact of BDNF Val66Met genotype on serotonin transporter and serotonin-1A receptor binding, PLOS-One, 2014 Sep 4;9(9) [2014: 3.23]
3. Kraus C, Ganger S, Losak J, Hahn A, Savli M, Kranz GS, Baldinger P, Windischberger C, Kasper S, Lanzenberger R, Gray matter and intrinsic network changes in the posterior cingulate cortex after selective serotonin reuptake inhibitor intake. NeuroImage 2014; 84:236-244. Epub 2013 Aug 26 [2014, IF: 6.36]
4. Kraus C, Hahn A, Savli M, Kranz GS, Baldinger P, Höflich A, Spindelegger C, Ungersböck J, Häusler D, Mitterhauser M, Windischberger C, Wadsak W, Kasper S, Lanzenberger R. Serotonin-1A receptor binding is positively associated gray matter volume – A multimodal neuroimaging study combining PET and structural MRI. NeuroImage 2012 Nov 15;63(3):1091-1098. Epub 2012 Jul 23 [2014, IF: 6.36]
5. Kraus C, Jandl-Jager E. Awareness and knowledge of child abuse amongst physicians - a descriptive study by a sample of rural Austria. Wien Klin Wochenschr. 2011 Jun;123(11-12):340-9. Epub 2011 May 4. [2014, IF: 0.84]
7.1.2 Co-authorships:
6. Seidel EM, Pfabigan D, Hahn A, Sladky R, Grahl A, Paul K, Kraus C, Küblböck M, Kranz G, Hummer A, Lanzenberger R, Windischberger C, Lamm C. Uncertainty during pain anticipation: The adaptive value of preparatory processes. Human Brain Mapping,. 2014 Oct 16. [Epub ahead of print], [2014, IF: 5.97]
7. Pfabigan D, Seidel EM, Sladky R, Hahn A, Paul K, Grahl A, Küblböck M, Kraus C, Hummer A, Kranz G, Windischberger C, Lanzenberger R, Lamm C, P300 amplitude variation is related to ventral striatum BOLD response during gain and loss anticipation: An EEG and fMRI experiment. NeuroImage 2014 Aug 1;96:12-21, Epub 2014 Apr 6 [2014, IF: 6.36].
8. Hahn A, Haeusler D, Kraus C, Höflich A, Kranz GS, Baldinger P, Savli M, Mitterhauser M, Wadsak W, Karanikas G, Kasper S, Lanzenberger R. Attenuated serotonin transporter association between dorsal raphe and ventral striatum in major depression. Human Brain Mapping Hum Brain Mapp. 2014 Aug;35(8):3857-66. Epub 2014 Jan 17. [2014, IF: 5.97]

129
9. Sladky R, Höflich A, Küblböck M, Kraus C, Baldinger P, Moser E, Lanzenberger R, Windischberger C. Disrupted efective connectivity between the amygdala and orbitofrontal cortex in social anxiety disorder during emotion discrimination revealed by dynamic causal modeling for fMRI, Cerebral Cortex, Epub 2013 Oct 9 [2014, IF: 8.66]
10. Baldinger P, Hahn A, Mitterhauser M, Kranz G, Friedl M, Wadsak W, Kraus C, Ungersböck J, Hartmann A, Giegling I, Rujescu D, Kasper S, Lanzenberger R. Impact of COMT genotype on serotonin-1A receptor binding investigated with PET. Brain Structure and Function 2014 Nov;219(6):2017-28. Epub 2013 Aug 9. [2014, IF: 5.62]
11. Hahn A, Kranz GS, Seidel EM, Sladky R, Kraus C, Küblböck M, Pfabigan DM, Hummer A, Grahl A, Ganger S, Windischberger C, Lamm C, Lanzenberger R. Comparing neural response to painful electrical stimulation with functional MRI at 3 and 7 Tesla. NeuroImage 2013 Nov 15;82:336-343. Epub 2013 Jun 12 [2014, IF: 6.36]
12. Kranz GS, Hahn A, Baldinger P, Häusler D, Philippe C, Kaufmann U, Wadsak W, Savli M, Höflich A, Kraus C, Vanicek T, Mitterhauser M, Kasper S, Lanzenberger R. Cerebral serotonin transporter asymmetry in males and male-to-female transsexuals: a PET study with [11C]DASB. Brain Structure and Function. 2014 Jan;219(1):171-83. Epub 2012 Dec 9 [2014, IF: 5.62]
13. Hahn A, Nics L, Baldinger P, Wadsak W, Savli M, Kraus C, Birkfellner W, Ungersboeck J, Haeusler D, Mitterhauser M, Karanikas G, Kasper S, Frey R, Lanzenberger R. Application of image-derived and venous input functions in major depression using [carbonyl-11C]WAY-100635. Nuclear Medicine and Biology 2013 Apr;40(3):371-7 [2014, IF: 2.41].
14. Hahn A, Wadsak W, Windischberger C, Baldinger P, Höflich A, Losak J, Nics L, Philippe C, Kranz GS, Kraus C, Mitterhauser M, Karanikas G, Kasper S, Lanzenberger R. Differential modulation of self-referential processing in the default mode network via serotonin-1A receptors. Proceedings of the National Academy of Sciences (PNAS) 2012 Feb 14;109(7):2619-24. Epub 2012 Jan 30. [2014, IF: 9.67]
15. Sladky R, Höflich A, Atanelov J, Kraus C, Baldinger P, Moser E, Lanzenberger R, Windischberger C. Increased neural habituation in the amygdala and orbitofrontal cortex in social anxiety disorder revealed by FMRI. PLOS One. 2012;7(11):e50050. Epub 2012 Nov 29. [2014, IF: 3.23]
7.2 Abstracts
7.2.1 1ST Authorships:
1. Kraus C, Savli M, Hahn A, Baldinger P, Höflich A, Mitterhauser M, Wadsak W, Windischberger C, Kasper S, Lanzenberger R. Serotonin-1A binding in the subgenual anterior cingulate cortex is associated with regional grey matter volume in striatum and temporal areas. 19th European Congress of Psychiatry (EPA), March 12-15, 2011, Vienna, Austria, European Psychiatry 2011, 26(1): P02-338

130
2. Kraus C, Hahn A, Savli M, Mitterhauser M, Wadsak W, Windischberger C, Kasper S, Lanzenberger R. A positive Correlation Between Inhibitory Serotonergic Neurotransmission and Grey Matter Volume. 7th PhD Symposium, 15-16th June 2011, Vienna Austria
3. Kraus C, Savli M, Hahn A, Höflich A, Baldinger P, Mitterhauser M, Wadsak W, Kasper S, Lanzenberger R. Molecular imaging of the serotonergic system – Serotonin, an important effector in modulating emotions. 12th International Neuropsychoanalysis Congress, June 24 – 26, 2011, Berlin, Germany
4. Kraus C, Hahn A, Savli M, Höflich A, Baldinger P, Kranz GS, Mitterhauser M, Wadsak W, Kasper S, Lanzenberger R. Serotonin-A receptor binding in the dorsal raphe nucleus is associated with hippocampal grey matter volume. 24th European College of Neuropsychopharmacology (ECNP) Congress, 3-7 September 2011, Paris, France European Neuropsychopharmacology,Vol 21, Suppl. 3, S318-319
5. Kraus C, Hahn A, Savli M, Baldinger P, Höflich A, Kranz GS, Losak J, Mitterhauser M, Wadsak W, Windischberger C, Kasper S, Lanzenberger R. Multimodal neuroimaging detects serotonin-1A receptor mediated neuroplasticity in humans. 24th IGB Workshop, Regulation of Neural Gene Expression from Development to Disease, 16-19 October 2011 Capri, Italy
6. Kraus C, Savli M, Hahn A, Baldinger P, Höflich A, Ungersboeck J, Mitterhauser M, Windischberger C, Wadsak W, Kasper S, Lanzenberger R. Serotonin-1A receptor related morphogenic signaling is associated with regional brain volumes and network neuroplasticity 20th EPA European Congress of Psychiatry, Prague, 3-6 March 2012
7. Kraus C, Vanicek T, Baldinger P, Hartmann A, Wadsak W, Lanzenberger R. Relationship between 5-HT1B receptor SNPs and 5-HT1A receptor BPND in healthy subjects measured by PET. 8th International Imaging Genetics Conference, Irvine, California, USA, January 16th–17th, 2012
8. Kraus C, Savli M, Hahn A, Höflich A, Baldinger P, Wadsak W, Windischberger C, Mitterhauser M, Kasper S, Lanzenberger R. Multimodal neuroimaging with PET and MRI to investigate the relation between serotonergic neurotransmission and regional brain volumes. ECNP Workshop on Neuropsychopharmacology for Young Scientists in Europe, 15-18 March 2012, Nice, France European Neuropsychopharmacology, Vol xx, Suppl. x, March 2012, Sxx(P.x.xxx)
9. Kraus C, Mitterhauser M, Bauer A, Ding Y-S, Henry S, Rattay F, Savli M, Lanzenberger R. A Normative database of the serotonergic system in healthy subjects using multi-tracer PET 10th PhD Symposium, June 13-14, 2012, Vienna, Austria

131
10. Kraus C, Vanicek T, Wadsak W, Baldinger P, Hartmann A, Mitterhauser M, Ungersboeck J, Rujescu D, Kasper S, Lanzenberger R. 5-HT1B receptor gene status alters 5-HT1A binding as investigated in vivo by PET and [carbonyl-11C]WAY-100635 25th European College of Neuropsychopharmacology (ECNP) Congress, 13-17 October 2012, Vienna, Austria European Neuropsychopharmacology,Vol 2x, Suppl. x, Sxxx-xxx
11. Kraus C, Ganger S, Losak J, Hahn A, Savli M, Spies M, Baldinger P, Windischberger C, Kassper S, Lanzenberger R. Rapid gray matter increases and resting state network changes after selective serotonin reuptake inhibitor administration. 11th World Congress of Biological Psychiatry (WFSBP), June 23rd- 27th, 2013, Kyoto, Japan
12. Kraus C, Kranz GS, Küblböck M, Pfabigan D, Hahn A, Sladky R, Seidel EM, Hummer A, Paul K, Ganger S, Windischberger C, Lamm C, Lanzenberger R. Reward anticipation maps comparing high and ultrahigh field functional MRI. 19th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 16-20, 2012, Seattle, USA
13. Kraus C, Baldinger P, Hahn A, Rami-Mark C, Wadsak W, Mitterhauser M, Kasper S, Lanzenberger R. Brain derived neurotrophic factor genotype status impacts on hippocampal serotonin-1A receptor binding. 13th Meeting of the Austrian Neuroscience Association (ANA) Vienna, 16–19 September 2013
14. Kraus C, Kranz GS, Pfabigan DM, Hoffmann A, Hahn A, Seidel EM, Küblböck M, Spies M, Paul K, Sladky R, Kasper S, Windischberger C, Lamm C, Lanzenberger R. Parahippocampal and insular gray matter volume correlates with empathy. 29th
CINP–World Congress of Neuropsychopharmacology, 22-26 June 2014, Vancouver, Canada
15. Kraus C, Stürkat IL, Sladky R, Hahn A, Pfabigan D, Tik M, Köck P, Windischberger C, Lamm C, Lanzenberger R. Altered structural plasticity in acute and remitted depressive patients investigated with ultra-high field magnetic resonance imaging. 28th ECNP Congress 29.8. – 1.9. 2015, Amsterdam.
7.2.2 Co-authorships:
1. Baldinger P, Savli M, Kranz GS, Höflich A, Kraus C, Windischberger C, Kasper S, Lanzenberger R. Are there structural brain changes following 10 days of SSRI administration investigated by voxel-based morphometry? 19th European Congress of Psychiatry (EPA), March 12-15, 2011, Vienna, Austria European Psychiatry 2011, 26(1): P02-317

132
2. Höflich A, Philippe C, Savli M, Baldinger P, Kranz GS, Müller S, Häusler D, Zgud S, Kraus C, Wadsak W, Mitterhauser M, Lanzenberger R, Kasper S. Prediction of steady-state occupancy of the serotonin transporter based on single-dose occupancy: A [11C]DASB PET study. 19th European Congress of Psychiatry (EPA), March 12-15, 2011, Vienna, Austria European Psychiatry 2011, 26(1): P02-333
3. Sladky R, Kraus C, Tröstl J, Kasper S, Lanzenberger R, Moser E, Windischberger C. Orbitofrontal hyperactivity in social anxiety disorder patients: An fMRI study. 19th European Congress of Psychiatry (EPA), March 12-15, 2011, Vienna, Austria European Psychiatry 2011, 26(1): P01-179
4. Tröstl J, Sladky R, Hummer A, Kraus C, Moser E, Kasper S, Lanzenber R, Windischberger C. Reduced connectivity in the uncinate fiber tract between the frontal cortex and limbic subcortical areas in social phobia. 19th European Congress of Psychiatry (EPA), March 12-15, 2011, Vienna, Austria European Psychiatry 2011, 26(1): P01-182
5. Savli M, Hahn A, Häusler D, Baldinger P, Höflich A, Kraus C, Wadsak W, Mitterhauser M, Lanzenberger R, Dudczak R, Kasper S. Can the Median Raphe Nucleus predict Clinical Outcome in Patients with Major Depression? A [11C]DASB PET study Talk (Young Scientists award session) and Abstract (CD) YS-02-001 10th World Congress of Biolog. Psychiatry (WFSBP), 29.6.– 2.7.2011, Prague, Czech Republic
6. Hahn A, Lanzenberger R, Häusler D, Philippe C, Savli M, Baldinger P, Höflich A, Kraus C, Akimova E, Mitterhauser M, Wadsak W, Kasper S. Reduced Serotonin Transporter Association between Raphe Region and Ventral Striatum in Major Depressive Disorder. Talk (Free Communication) and Abstract (CD) FC-06-002 10th World Congress of Biolog. Psychiatry (WFSBP), 29.6.– 2.7.2011, Prague, Czech Republic
7. Savli M, Hahn A, Häusler D, Philippe C, Baldinger P, Höflich A, Kraus C, Kranz GS, Zgud S, Akimova E, Wadsak W, Mitterhauser M, Lanzenberger R, Dudczak R, Kasper S. The Impact of Software Motion Correction on PET Drug Occupancy Studies. 10th International Conference on Quantification of Brain Function with PET, May 24-28, 2011, Barcelona, Spain
8. Kranz GS, Kaufmann U, Ungersböck J, Hahn A, Stein P, Baldinger P, Höflich A, Zgud S, Kraus C, Losak J, Mitterhauser M, Wadsak W, Kasper S, Lanzenberger R. Estrogen and progesterone treatment affects serotoninergic neurotransmission in postmenopausal women. 17th Annual Meeting of the Organization for Human Brain Mapping, June 26-30, 2011, Quebec City, Canada
9. Sladky R, Kraus C, Tröstl J, Kasper S, Lanzenberger R, Moser E, Windischberger C. Orbitofrontal hyperactivity and habituation in social anxiety disorder patients: an

133
fMRI study. 17th Annual Meeting of the Organization for Human Brain Mapping, June 26-30, 2011, Quebec City, Canada
10. Tröstl J, Sladky R, Hummer A, Kraus C, Moser E, Kasper S, Lanzenberger R, Windischberger C. White Matter Alterations in Social Anxiety Disorder: a DTI study. 17th Annual Meeting of the Organization for Human Brain Mapping, June 26-30, 2011, Quebec City, Canada
11. Höflich A, Hahn A, Kraus C, Baldinger P, Kranz GS, Windischberger C, Kasper S, Lanzenberger R. Resting-state functional connectivity of the raphe nuclei. 24th European College of Neuropsychopharmacology (ECNP) Congress, 3-7 September 2011, Paris, France European Neuropsychopharmacology,Vol 21, Suppl. 3, S319-320
12. Savli M, Baldinger P, Kranz GS, Höflich A, Kraus C, Losak J, Windischberger C, Kasper S, Lanzenberger R. Rapid gray matter density changes after selective serotonin reuptake inhibitor administration revealed by voxel-based-morphometry. 24th European College of Neuropsychopharmacology (ECNP) Congress, 3-7 September 2011, Paris, France European Neuropsychopharmacology,Vol 21, Suppl. 3, S312
13. Savli M, Hahn A, Häusler D, Baldinger P, Höflich A, Kraus C, Wadsak W, Mitterhauser M, Lanzenberger R, Dudczak R, Kasper S. The Impact of Median Raphe Nucleus Serotonin Transporter Binding on Depression: A [11C]DASB PET study. (Abstract and Talk). Annual Meeting of the International Society of NeuroImaging in Psychiatry (ISNIP), 07-10.09.2011, Heidelberg, Germany.
14. Tröstl J, Sladky R, Hummer A, Kraus C, Moser E, Kasper S, Lanzenberger R, Windischberger C. DTI of White Matter Alterations in the Uncinate Fasciculus of Social Phobia Patients. (Talk) 28th Annual Scientific Meeting of the European Society for Magnetic Resonance in Medicine and Biology (ESMRM), October 6-8, 2011, Leipzig, Germany.
15. Baldinger P, Kraus C, Friedl M, Haeusler D, Hartmann A, Mitterhauser M, Kranz G, Rujescu D, Kasper S, Lanzenberger R. 5-HT1A receptor genotype is associated with 5-HT1A receptor binding in the healthy human brain measured by PET. ECNP Workshop on Neuropsychopharmacology for Young Scientists in Europe, 15-18 March 2012, Nice, France European Neuropsychopharmacology, Vol xx, Suppl. x, March 2012, Sxx(P.x.xxx)
16. Hahn A, Wadsak W, Windischberger C, Baldinger P, Höflich A, Losak J, Nics L, Ungersböck J, Kranz GS, Kraus C, Mitterhauser M, Karanikas G, Kasper S, Lanzenberger R. Default mode network is modulated by serotonin-1A receptors. 18th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 10-14, 2012, Beijing, China NeuroImage Volume xx, Supplement x, August 2012, Pages Sxxx-Sxxx

134
17. Sladky R, Höfich A, Tröstl J, Kraus C, Baldinger P, Moser E, Lanzenberger R, Windischberger C. Increased neural habituation in the amygdala and orbitofrontal cortex in social anxiety disorder revealed by fMRI. 18th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 10-14 2012, Beijing, China NeuroImage Volume xx, Supplement x, August 2012, Pages Sxx-Sx
18. Lanzenberger R, Hahn A, Ungersböck J, Friedl M, Baldinger P, Philippe C, Nics L, Kranz GS, Kraus C, Häusler D, Hartmann A, Savli M, Vanicek T, Mitterhauser M, Wadsak W, Rujescu D. Kasper S. A genetic variation of the serotonin-1B receptor affects serotonin-1A receptor in vivo binding. 18th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 10-14 2012, Beijing, China NeuroImage Volume xx, Supplement x, August 2012, Pages Sxx-Sx
19. Lanzenberger R. Mitterhauser M, Hahn A, Baldinger P, Friedl M, Kraus C, Pichler S, Rujescu D, Wadsak W, Kasper S. Molecular imaging genetics of the serotonin-1A receptor investigating the common rs6295 single nucleotide polymorphism. The 9th International Symposium on Functional Neuroreceptor Mapping of the Living Brain, August 9-11, 2012, Baltimore, Maryland, USA Journal of Cerebral Blood Flow and Metabolism 2012, Suppl. Xx
20. Baldinger P, Hahn A, Wadsak W, Friedl M, Kraus C, Mitterhauser M, Ungersböck J, Rujescu D, Kasper S, Lanzenberger R. Association between Catechol-O-methyltransferase Genotyp and Serotonin-1A Receptor Binding measured via Positron Emission Tomography 25th European College of Neuropsychopharmacology (ECNP) Congress, 13-17 October 2012, Vienna, Austria European Neuropsychopharmacology,Vol 2x, Suppl. x, Sxxx-xxx
21. Baldinger P, Hahn A, Mitterhauser M, Kraus C, Wadsak W, Rujescu D, Kasper S, Lanzenberger R. Genotyp of serotonin-1B receptor affects serotonin-1A receptor
binding in vivo 10th PhD Symposium, June 13-14, 2012, Vienna, Austria
22. Höflich A, Hahn A, Atanelov J, Baldinger P, Kraus C, Windischberger C, Kasper S, Lanzenberger R. Influence of ketamine on resting-state functional connectivity in healthy volunteers - A fMRI study. 10th PhD Symposium, June 13-14, 2012, Vienna, Austria
23. Seidel EM, Pfabigan D, Hahn A, Sladky R, Grahl A, Kraus C, Kueblboeck M, Kranz G, Hummer A, Lanzenberger R, Windischberger C, Lamm C. Uncertainty during pain anticipation – An fMRI and EEG experiment. 19th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 16-20, 2012, Seattle, USA
24. Hahn A, Kranz G, Seidel EM, Sladky R, Kraus C, Kueblboeck M, Pfabigan D, Hummer A, Grahl A, Ganger S, Windischberger C, Lamm C, Lanzenberger R.

135
Imaging the periaqueductal gray during pain processing at 3 and 7 Tesla functional MRI. 19th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 16-20, 2012, Seattle, USA
25. Pfabigan D, Seidel EM, Sladky R., Hahn A, Paul K, Kueblboeck M, Kraus C, Hummer A, Kranz G, Windischberger C, Lanzenberger R, Lamm C. A multimodal study on gain and loss anticipation combining fMRI and EEG. 19th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 16-20, 2012, Seattle, USA
26. Sladky R, Höflich A., Kuebelboeck M, Kraus C, Baldinger P, Moser E, Lanzenberger R, Windischberger C. Disrupted effective connectivity between amygdala and OFC in social anxiety disorder revealed by DCM. 19th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 16-20, 2012, Seattle, USA
27. Ganger S, Hahn A, Sladky R, Kueblboeck M, Kranz G, Höflich A, Kraus C, Losak J, Spies M, Windischberger C, Lanzenberger R. Comparing techniques for resting state extraction from task data. 19th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 16-20, 2012, Seattle, USA
28. Küblböck M, Hummer A, Hahn A, Hoffmann A, Kraus C, Woletz M, Komorowski A, Lanzenberger R, Lamm C, Windischberger C. Reduction in vascular confounds of 3T and 7T fMRI group analysis results using the RESCALE method. 20th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 8-12, 2014, Hamburg, Germany.
29. Hoffmann A, Sladky R, Spies M, Küblböck M, Höflich A, Hummer A, Kranz GS, Woletz M, Lamm C, Lanzenberger R, Windischberger C. The Default Mode Network’s Frequency-dependency. 20th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 8-12, 2014, Hamburg, Germany.
30. Woletz M, Hoffmann A, Ganger S, Seiger R, Hahn A, Lamm C, Lanzenberger R, Windischberger C. Slice-timing correction for multi-band images in SPM. 20th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 8-12, 2014, Hamburg, Germany.
31. Sladky R, Minkova L, Höflich A, Kraus C, Baldinger P, Moser M, Lanzenberger R., Windischberger C. Task-dependent modulation of amygdalar connectivity in social anxiety disorder patients and healthy subjects. 20th Annual Meeting of the Organization for Human Brain Mapping (HBM), June 8-12, 2014, Hamburg, Germany.
32. Sladky R , Kraus C, Stürkat IL, Hoffmann A, Lamplmair D, Tik M, Spies M, Pfabigan D, Lamm C, Lanzenberger R, Windischberger C. Effective connectivity of amygdalar sub regions and OFC in acute and remitted MDD patients at 7T. 21st Annual Meeting of the Organization for Human Brain Mapping (HBM), June 14-18 2015, Honolulu, Hawaii, USA

136
33. The influence of high‐ dose estradiol administration on limbic brain structures and
the ventricular system Seiger R, Hahn A, Hummer A, Kranz GS, Ganger S, Woletz M, Kraus C, Sladky R, Kasper S, Windischberger C, Lanzenberger R. DGPPN Kongress – Deutsche Gesellschaft für Psychiatrie und Psychotherapie, Psychosomatik und Nervenheilkunde, November 25‐ 28 2015, Berlin
7.3 Lectures
1. Transkranielle Magnetstimulation als neue Behandlungsmethode bei Sozial Phobie. Fortbildungsveranstaltung der Klinischen Abteilung für Allgemeine Psychiatrie in Wien (Scientific seminar at the Department for Psychiatry and Pschotherapy, 21.10.10).
2. Einfluss der serotonergen Transmission auf die Struktur der Grauen Substanz. Winterseminar “Biologische Psychiatrie”, Oberlech 21.3.11 (Scientific seminar at the at the Wintersymposium “Biological Psychiatry, Oberlech, 21.03.11).
3. Serotonin-1A receptor distribution in the subgenual anterior cingulate cortex is associated with regional grey matter volume in the striatum and temporal areas. 19th European Congress of Psychiatry (EPA), March 12-15, 2011, Vienna, Austria, e-Poster presentation.
4. Inhibitory serotonergic neurotransmission correlates positively with cortical grey matter volume as revealed by PET and MRI. World Congress of Biological Psychiatry (WFSBP), May 31st, 2011, Prague, Czech Republic, free communication.
5. Serotonin-1a receptor related morphogenic signaling is associated with regional brain volumes and network neuroplasticity. 20th European Congress of Psychiatry, March 3-6, 2012, Prague, oral presentation.
6. Gen–Umwelt Interaktionen – Erfahrungsbedingte Plastizität in Gesundheit und psychiatrischen Erkrankungen, Winterseminar “Biologische Psychiatrie”, Oberlech 11.3.2013 (Scientific seminar at the at the Wintersymposium “Biological Psychiatry, Oberlech).
7. Rapid gray matter increases and resting state network changes after selective serotonin reuptake inhibitor administration, 11th World Congress of Biological Psychiatry (WFSBP), 26.6.2013, Kyoto, Japan, free communication.
8. Brain derived neurotrophic factor genotype status impacts on hippocampal serotonin-1A receptor binding, 13th Meeting of the Austrian Neuroscience Association (ANA),Vienna, 16.9.2013, oral presentation
9. Die Bedeutung des glutamatergen Systems bei psychiatrischern Erkrankungen, ÖGBP, 15.11.2013, Official Meeting of the Austrian Society for Neuropsychopharmacologgy and Biological Psychiatry.
10. Interaktionen zwischen Serotonintransporter und Serotonin-1B Rezeptor-Genotypen auf die Expression des Serotonin-1A Rezeptors, Winterseminar “Biologische Psychiatrie”, Oberlech (Scientific seminar at the at the Wintersymposium “Biological Psychiatry, Oberlech, 23.03.2014

137
11. Interaction between serotonin transporter and 1B receptor genotype status impacts on serotonin 1A receptor binding, Oral Presentation, 22.5.2014, The 10th International Symposium on Functional NeuroReceptor Mapping of the Living Brain, Egmond aan Zee, The Netherlands
12. Serotonin and Neuroplasticity, Young Scientists Section at DEVELAGE-Symposium, Medical University of Vienna, 27.11.2014
13. Prädiktion des Behandlungserfolges bei Depression mit funktioneller MRT, (Scientific seminar at the at the Wintersymposium “Biological Psychiatry, Oberlech, 17.3.2015
14. Serotonin and Neuroplasticity, Interdisciplinary Meeting at the Center of Brain Science, 27.3.2015
02/07/2015