self-renewal of human embryonic stem cells requires insulin - blood
TRANSCRIPT

STEM CELLS IN HEMATOLOGY
Self-renewal of human embryonic stem cells requires insulin-like growth factor-1receptor and ERBB2 receptor signaling
Linlin Wang,1 Thomas C. Schulz,2 Eric S. Sherrer,2 Derek S. Dauphin,1 Soojung Shin,3 Angelique M. Nelson,4
Carol B. Ware,4 Mei Zhan,5 Chao-Zhong Song,5 Xiaoji Chen,1 Sandii N. Brimble,2 Amanda McLean,6 Maria J. Galeano,2
Elizabeth W. Uhl,7 Kevin A. D’Amour,8 Jonathan D. Chesnut,3 Mahendra S. Rao,3 C. Anthony Blau,1 and Allan J. Robins2
1Division of Hematology, Department of Medicine, Institute for Stem Cell and Regenerative Medicine, University of Washington, Seattle; 2Novocell, Athens, GA;3Invitrogen, Carlsbad, CA; 4Department of Comparative Medicine, University of Washington, Seattle; 5Division of Medical Genetics, Department of Medicine,University of Washington, Seattle; 6Department of Animal and Dairy Science, University of Georgia, Athens; 7Department of Pathology, College of VeterinaryMedicine, University of Georgia, Athens; and 8Novocell, San Diego, CA
Despite progress in developing definedconditions for human embryonic stemcell (hESC) cultures, little is known aboutthe cell-surface receptors that are acti-vated under conditions supportive ofhESC self-renewal. A simultaneous inter-rogation of 42 receptor tyrosine kinases(RTKs) in hESCs following stimulationwith mouse embryonic fibroblast (MEF)conditioned medium (CM) revealed rapidand prominent tyrosine phosphorylationof insulin receptor (IR) and insulin-likegrowth factor-1 receptor (IGF1R); lessprominent tyrosine phosphorylation ofepidermal growth factor receptor (EGFR)
family members, including ERBB2 andERBB3; and trace phosphorylation of fi-broblast growth factor receptors. IntenseIGF1R and IR phosphorylation occurredin the absence of MEF conditioning (NCM)and was attributable to high concentra-tions of insulin in the proprietary Knock-Out Serum Replacer (KSR). Inhibition ofIGF1R using a blocking antibody or lenti-virus-delivered shRNA reduced hESC self-renewal and promoted differentiation,while disruption of ERBB2 signaling withthe selective inhibitor AG825 severelyinhibited hESC proliferation and pro-moted apoptosis. A simple defined me-
dium containing an IGF1 analog, heregu-lin-1� (a ligand for ERBB2/ERBB3),fibroblast growth factor-2 (FGF2), and ac-tivin A supported long-term growth ofmultiple hESC lines. These studies iden-tify previously unappreciated RTKs thatsupport hESC proliferation and self-renewal, and provide a rationally de-signed medium for the growth and main-tenance of pluripotent hESCs. (Blood.2007;110:4111-4119)
© 2007 by The American Society of Hematology
Introduction
In guiding human embryonic stem cell (hESC) technologytoward the clinic, 1 key issue to be addressed is a lack ofstandardization in the culture and maintenance of hESCs. In theabsence of mouse embryonic fibroblast (MEF) feeder layers,many researchers rely on “conditioning” in which medium isfirst exposed to MEFs to acquire soluble factors that support thepropagation of undifferentiated hESCs in culture. It has beendifficult to discern how MEF conditioning enables hESCs tomaintain an undifferentiated state. Other common features ofmore recently developed hESC culture conditions include thepresence of fibroblast growth factor-2 (FGF2), the absence ofserum, and the presence of a serum substitute such as KnockOutSerum Replacer (KSR, proprietary formulation; Invitrogen,Carlsbad, CA).1-3 Other factors suggested to play a role insupporting the maintenance of hESCs include transforminggrowth factor �1 (TGF�1),4 activin A (ActA),5,6 platelet-derivedgrowth factor (PDGF) and sphingosine-1-phosphate,7 BIO, asmall-molecule inhibitor of GSK3�,8 and neurotrophins.9 Sev-eral defined medium systems have been described for hESCsand are based upon FGF2 in combination with nodal,10 TGF�1,
GABA, and pipecolic acid, plus lithium chloride,11 Wnt3a plusApril/BAFF,12 or the N2/B27 supplements.13
Although these studies have focused on identifying growthfactors and conditions that support the proliferation of undifferenti-ated hESCs, little is known about the cell-surface receptors that areactivated when hESCs are exposed to conditions favorable forself-renewal. A number of receptor tyrosine kinases (RTKs) areexpressed at high levels on hESCs,14 including insulin-like growthfactor-1 receptor (IGF1R), fibroblast growth factor receptor(FGFR1), and EPHA1, as well as ERBB2 and ERBB3 (which aremembers of the epidermal growth factor receptor [EGFR] family),while expression of FGFR2 (EGFR) FGFR4, vascular endothelialgrowth factor receptor-2 (VEGFR2), IGFR2, KIT, and RET hasalso been detected.15,16 RTKs are likely to be central signalingeffectors17 that influence survival, apoptosis, proliferation, ordifferentiation decisions in pluripotent cells. To determine if any ofthese RTKs are involved in self-renewal, we simultaneouslyinterrogated the tyrosine phosphorylation status of 42 RTKs inhESCs grown in MEF–conditioned medium (CM) and developed adefined medium for hESC culture.
Submitted March 29, 2007; accepted August 16, 2007. Prepublished onlineas Blood First Edition paper, August 29, 2007; DOI 10.1182/blood-2007-03-082586.
The online version of this article contains a data supplement.
The publication costs of this article were defrayed in part by page chargepayment. Therefore, and solely to indicate this fact, this article is herebymarked ‘‘advertisement’’ in accordance with 18 USC section 1734.
© 2007 by The American Society of Hematology
4111BLOOD, 1 DECEMBER 2007 � VOLUME 110, NUMBER 12
For personal use only.on November 18, 2018. by guest www.bloodjournal.orgFrom

Materials and methods
Cell culture
The National Institutes of Health (NIH)–registered H1, BG01, BG02, andBG03 hESC lines, as well as CyT49, an hESC line isolated using humanfeeder cells under good manufacturing process (GMP) conditions (Novo-cell, San Diego, CA), were used in these experiments. Nonconditionedmedium (NCM) and CM were prepared as described previously,2 andhESCs were maintained in CM on plates coated with Matrigel (BDBiosciences, San Jose, CA) diluted 1:30, or in NCM on MEF feeder layerswhere indicated. Cultures were routinely passaged with collagenase IV2 ordispase.3 Karyotype analyses of hESCs grown in CM or defined conditions(DC) HAIF (Heregulin-1B, Activin A, Insulin like growth factor-1, FGF2)were performed using standard G-banding techniques.
Culture of hESCs in defined conditions
DC-HAIF consisted of DMEM/F12 (Invitrogen), 2% fatty acid-free Cohnfraction V bovine serum albumin (BSA; Serologicals, Norcross, GA),1 � nonessential amino acids, 50 U/mL penicillin, 50 �g/mL streptomycin,50 �g/mL ascorbic acid, 10 �g/mL bovine or human transferrin, 0.1 mM�-mercaptoethanol (all from Invitrogen), 1 times trace elements A, B, and C(Mediatech, Manassas, VA), 10 ng/mL HRG1� (Peprotech, Rocky Hill,NJ), 10 ng/mL ActA (R&D Systems, Minneapolis, MN), 200 ng/mLLR3-IGF1 (JRH Biosciences, Lenexa, KS), and 8 ng/mL FGF2 (Sigma, StLouis, MO) or R&D Systems). HESCs were cultured in DC-HAIF ongrowth factor–depleted Matrigel (BD Biosciences) diluted 1:200. Near-confluent hESC cultures were passaged by treating with 10 mg/mL(approximately 2000 U/mL) collagenase IV for 3 minutes, washing with0.2% BSA in DMEM/F12, and gently scraping to lift and break up thecolonies. Colony pieces were centrifuged at 200g, aspirated, gentlyresuspended in DC-HAIF, and typically split 1:3 to new plates. Theformulation of DC-HAIF is shown in Table S1 (available on the Bloodwebsite; see the Supplemental Materials link at the top of the online article);a detailed protocol for the maintenance of hESCs in DC-HAIF is availablein Document S1. Cell- and colony-counting experiments, the generationand analysis of teratomas, embryoid bodies, and defined differentiation toendodermal lineages are detailed in Document S1.
Human RTK arrays
Proteome Profiler human phospho-RTK antibody arrays (R&D Systems)were used according to the manufacturer’s instructions (Figure S1;Document S1). A total of 500 �g fresh protein lysates were incubatedovernight with nitrocellulose membranes dotted with duplicate spots for 42anti-RTK and control antibodies. Bound phospho-RTKs were detected witha pan antiphosphotyrosine antibody conjugated to horseradish peroxidaseusing chemiluminescence.
Receptor inhibition studies
For experiments using the IGF1R blocking antibody, H1 hESCs wereincubated with 10 �g/mL A12 (ImClone, New York, NY) or control humanIgG (Sigma) in CM. IGF1R expression was evaluated by flow cytometryafter 4 hours of incubation with A12 or control antibody. Colonies werecounted and graded morphologically on the indicated days (Document S1;Figure S2). Both differentiation and apoptosis were evaluated 3 days afterthe addition of A12. Morphologic assessment of differentiation wasconfirmed by SSEA3 staining using flow cytometry, and apoptosis wasassessed by TUNEL staining. Inhibition of ERBB2 with AG825 (Calbio-chem, San Diego, CA) used hESCs growing in CM on Matrigel diluted 1:30(Document S1). Cell-cycle and apoptosis assays were performed after4 days of culture in CM plus either AG825 or DMSO (control).
Flow cytometry, RT-PCR analysis, and immunofluorescence
Flow cytometry analyses or sorting were performed using a FACScan,FACSCanto, or FACS Aria cell sorter (BD Biosciences). The FlowTACS kit
(R&D Systems) was used to detect apoptotic cells, according to themanufacturer’s instructions. Antibodies used to detect cell-surface antigensfor flow and other immunofluorescence analyses are described in DocumentS1. Cell-cycle and DNA content analysis was performed using propidiumiodide staining and analysis with the Modfit 3.0 software (Verity HouseSoftware, Topsham, ME). Immunostaining of cell cultures used secondaryantibodies conjugated with Alexa-488 (green) or -594 (red) (Invitrogen).Nuclei were counterstained with DAPI (Sigma). Stained slides weremounted with Aqua Poly/Mount (Polysciences, Warrington, PA) andimages were captured using Nikon TE2000-S and -E inverted microscopes(Nikon, Tokyo, Japan), Q-Imaging Retiga cameras (Qimaging, Surrey, BC),and the Q-Capture imaging system. Histological sections were stained withhematoxylin and eosin (Sigma), or with primary antibody followed bydetection with alkaline phosphatase tagged secondary and counterstainingwith hematoxylin. Slides were mounted with Permount (Biomedia, FosterCity, CA), and images were captured with an Olympus BX41 lightmicroscope (Olympus, Center Valley, PA), an Olympus DP70 digitalcamera, and Image Pro Plus 6.0 software (MediaCybernetics, Bethesda,MD). Images were processed with Adobe Photoshop 6.0 (Adobe, San Jose,CA). Methods for RNA isolation, reverse transcription–polymerase chainreaction (RT-PCR) analyses, and primer sets are described in Document S1.
ShRNA inhibition of IGF1R
Design and construction of the IGF1R-targeting shRNA lentivirus vector,plasmid transfection, generation of viral supernatants, evaluation of titer,and transduction of H1 hESCs are all described in Document S1.Transduced cells were cultured for 22 days and evaluated periodically forgreen fluorescent protein (GFP) and IGF1R expression by flow cytometry.
Illumina bead array and transcriptional analysis
Methods for the preparation of samples, labeling, hybridization, washing,detection, and analysis have been described previously,18 and were carriedout according to the manufacturer’s instructions (Document S1). Analysiswas performed using parallel approaches described for other hESCsamples18 with Illumina Sentrix Human-6 Expression Beadchips containing47 296 transcript probes (Illumina, San Diego, CA). NIH gene expressionomnibus (GEO) accession number-GSE6645.
Results
The phosphorylation status of 42 RTKs was simultaneouslyinterrogated in hESCs using a membrane array of primary antibod-ies and detection with a pan-antiphosphotyrosine antibody (FigureS1). H1 cells cultured on Matrigel were starved overnight withDMEM/F12 and 0.5% BSA and then stimulated with CM for 5 or15 minutes. Following overnight starvation, variable faint ormoderate phosphorylation of several receptors, including IR,IGF1R, ERBB1 (EGFR), ERBB2, ERBB3, VEGFR2, and FGFR3,was detected (Figure 1A,B). Striking induction of IGF1R and IRphosphorylation occurred within 5 minutes of stimulation withCM, and persisted at 15 minutes (Figure 1A). Similarly, phosphor-ylation of ERBB2 increased at 5 minutes, and rose further at15 minutes. ERBB1 and ERBB3 were also phosphorylated, but to alesser extent, at 15 minutes. CM stimulation also elicited faint butconsistent phosphorylation of FGFR3. A handful of receptors,including Axl, VEGFR2, PDGFR�, EPHA2, FGFR2�, MER, andTie2 were phosphorylated faintly and inconsistently in response toCM. Of note, phosphorylated derivatives of FGFR1, and theneurotrophin receptors TRKB and TRKC, were not detectedfollowing stimulation with CM.
To discriminate between effects mediated by components ofNCM and effects resulting from conditioning, receptor phosphory-lation was compared after stimulating overnight-starved H1 cells
4112 WANG et al BLOOD, 1 DECEMBER 2007 � VOLUME 110, NUMBER 12
For personal use only.on November 18, 2018. by guest www.bloodjournal.orgFrom
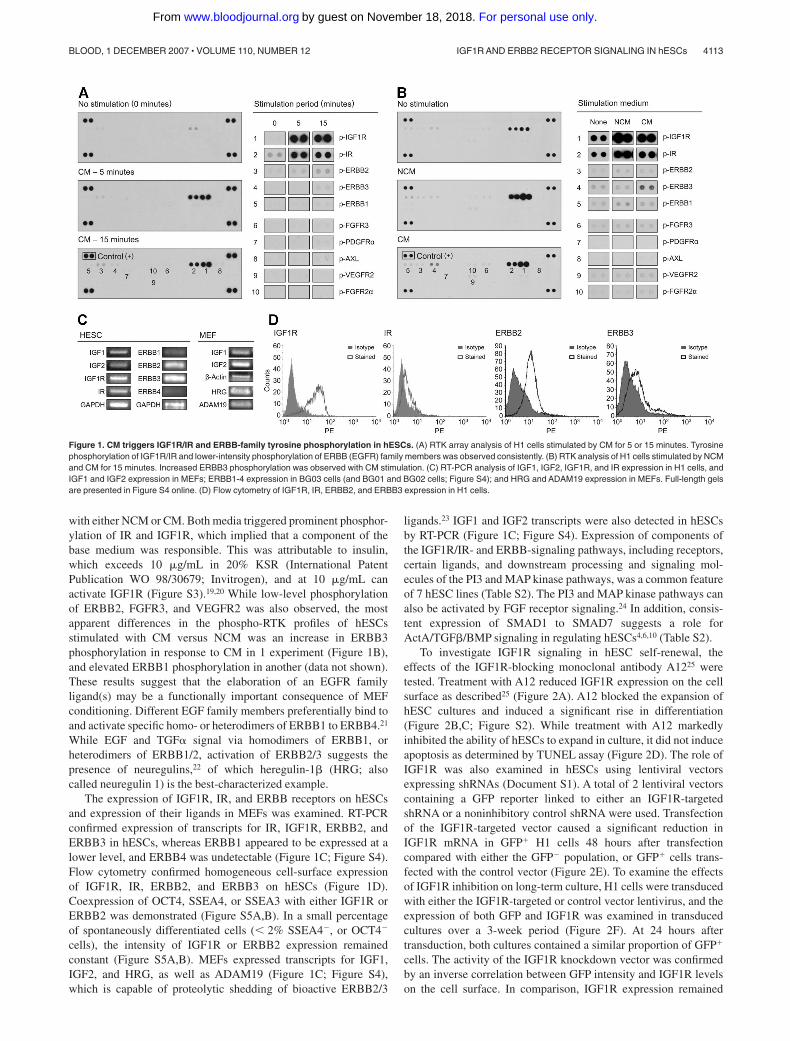
with either NCM or CM. Both media triggered prominent phosphor-ylation of IR and IGF1R, which implied that a component of thebase medium was responsible. This was attributable to insulin,which exceeds 10 �g/mL in 20% KSR (International PatentPublication WO 98/30679; Invitrogen), and at 10 �g/mL canactivate IGF1R (Figure S3).19,20 While low-level phosphorylationof ERBB2, FGFR3, and VEGFR2 was also observed, the mostapparent differences in the phospho-RTK profiles of hESCsstimulated with CM versus NCM was an increase in ERBB3phosphorylation in response to CM in 1 experiment (Figure 1B),and elevated ERBB1 phosphorylation in another (data not shown).These results suggest that the elaboration of an EGFR familyligand(s) may be a functionally important consequence of MEFconditioning. Different EGF family members preferentially bind toand activate specific homo- or heterodimers of ERBB1 to ERBB4.21
While EGF and TGF� signal via homodimers of ERBB1, orheterodimers of ERBB1/2, activation of ERBB2/3 suggests thepresence of neuregulins,22 of which heregulin-1� (HRG; alsocalled neuregulin 1) is the best-characterized example.
The expression of IGF1R, IR, and ERBB receptors on hESCsand expression of their ligands in MEFs was examined. RT-PCRconfirmed expression of transcripts for IR, IGF1R, ERBB2, andERBB3 in hESCs, whereas ERBB1 appeared to be expressed at alower level, and ERBB4 was undetectable (Figure 1C; Figure S4).Flow cytometry confirmed homogeneous cell-surface expressionof IGF1R, IR, ERBB2, and ERBB3 on hESCs (Figure 1D).Coexpression of OCT4, SSEA4, or SSEA3 with either IGF1R orERBB2 was demonstrated (Figure S5A,B). In a small percentageof spontaneously differentiated cells (� 2% SSEA4�, or OCT4�
cells), the intensity of IGF1R or ERBB2 expression remainedconstant (Figure S5A,B). MEFs expressed transcripts for IGF1,IGF2, and HRG, as well as ADAM19 (Figure 1C; Figure S4),which is capable of proteolytic shedding of bioactive ERBB2/3
ligands.23 IGF1 and IGF2 transcripts were also detected in hESCsby RT-PCR (Figure 1C; Figure S4). Expression of components ofthe IGF1R/IR- and ERBB-signaling pathways, including receptors,certain ligands, and downstream processing and signaling mol-ecules of the PI3 and MAP kinase pathways, was a common featureof 7 hESC lines (Table S2). The PI3 and MAP kinase pathways canalso be activated by FGF receptor signaling.24 In addition, consis-tent expression of SMAD1 to SMAD7 suggests a role forActA/TGF�/BMP signaling in regulating hESCs4,6,10 (Table S2).
To investigate IGF1R signaling in hESC self-renewal, theeffects of the IGF1R-blocking monoclonal antibody A1225 weretested. Treatment with A12 reduced IGF1R expression on the cellsurface as described25 (Figure 2A). A12 blocked the expansion ofhESC cultures and induced a significant rise in differentiation(Figure 2B,C; Figure S2). While treatment with A12 markedlyinhibited the ability of hESCs to expand in culture, it did not induceapoptosis as determined by TUNEL assay (Figure 2D). The role ofIGF1R was also examined in hESCs using lentiviral vectorsexpressing shRNAs (Document S1). A total of 2 lentiviral vectorscontaining a GFP reporter linked to either an IGF1R-targetedshRNA or a noninhibitory control shRNA were used. Transfectionof the IGF1R-targeted vector caused a significant reduction inIGF1R mRNA in GFP� H1 cells 48 hours after transfectioncompared with either the GFP� population, or GFP� cells trans-fected with the control vector (Figure 2E). To examine the effectsof IGF1R inhibition on long-term culture, H1 cells were transducedwith either the IGF1R-targeted or control vector lentivirus, and theexpression of both GFP and IGF1R was examined in transducedcultures over a 3-week period (Figure 2F). At 24 hours aftertransduction, both cultures contained a similar proportion of GFP�
cells. The activity of the IGF1R knockdown vector was confirmedby an inverse correlation between GFP intensity and IGF1R levelson the cell surface. In comparison, IGF1R expression remained
Figure 1. CM triggers IGF1R/IR and ERBB-family tyrosine phosphorylation in hESCs. (A) RTK array analysis of H1 cells stimulated by CM for 5 or 15 minutes. Tyrosinephosphorylation of IGF1R/IR and lower-intensity phosphorylation of ERBB (EGFR) family members was observed consistently. (B) RTK analysis of H1 cells stimulated by NCMand CM for 15 minutes. Increased ERBB3 phosphorylation was observed with CM stimulation. (C) RT-PCR analysis of IGF1, IGF2, IGF1R, and IR expression in H1 cells, andIGF1 and IGF2 expression in MEFs; ERBB1-4 expression in BG03 cells (and BG01 and BG02 cells; Figure S4); and HRG and ADAM19 expression in MEFs. Full-length gelsare presented in Figure S4 online. (D) Flow cytometry of IGF1R, IR, ERBB2, and ERBB3 expression in H1 cells.
IGF1R AND ERBB2 RECEPTOR SIGNALING IN hESCs 4113BLOOD, 1 DECEMBER 2007 � VOLUME 110, NUMBER 12
For personal use only.on November 18, 2018. by guest www.bloodjournal.orgFrom
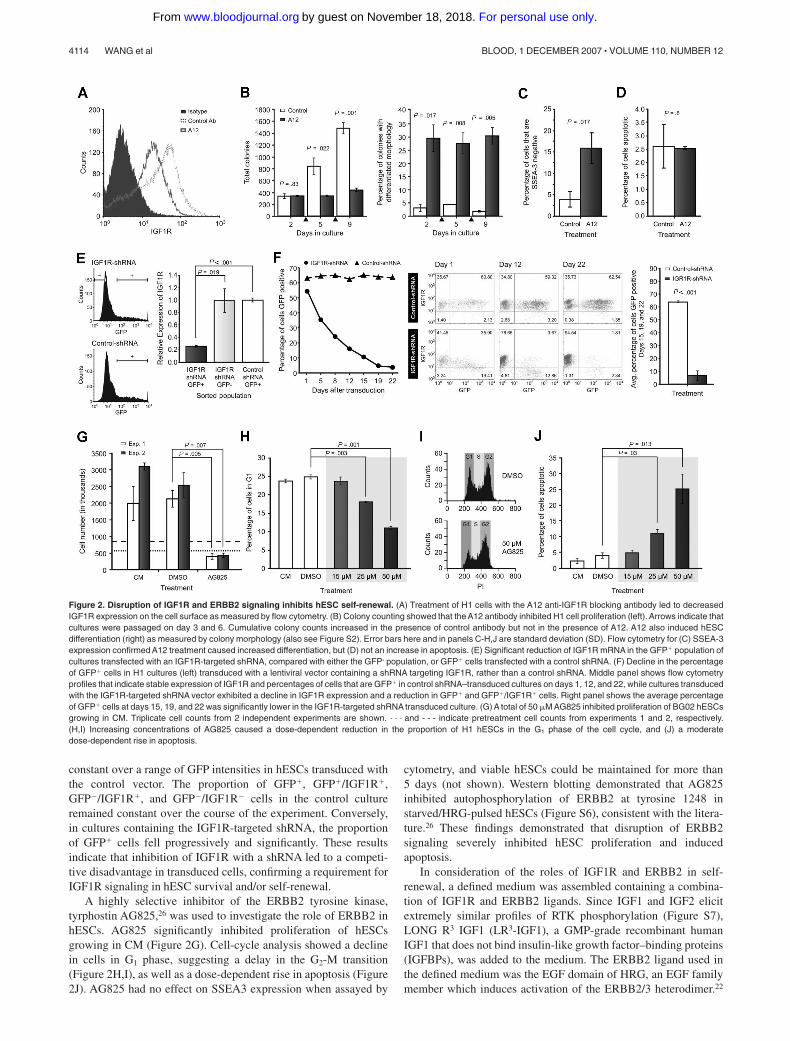
constant over a range of GFP intensities in hESCs transduced withthe control vector. The proportion of GFP�, GFP�/IGF1R�,GFP�/IGF1R�, and GFP�/IGF1R� cells in the control cultureremained constant over the course of the experiment. Conversely,in cultures containing the IGF1R-targeted shRNA, the proportionof GFP� cells fell progressively and significantly. These resultsindicate that inhibition of IGF1R with a shRNA led to a competi-tive disadvantage in transduced cells, confirming a requirement forIGF1R signaling in hESC survival and/or self-renewal.
A highly selective inhibitor of the ERBB2 tyrosine kinase,tyrphostin AG825,26 was used to investigate the role of ERBB2 inhESCs. AG825 significantly inhibited proliferation of hESCsgrowing in CM (Figure 2G). Cell-cycle analysis showed a declinein cells in G1 phase, suggesting a delay in the G2-M transition(Figure 2H,I), as well as a dose-dependent rise in apoptosis (Figure2J). AG825 had no effect on SSEA3 expression when assayed by
cytometry, and viable hESCs could be maintained for more than5 days (not shown). Western blotting demonstrated that AG825inhibited autophosphorylation of ERBB2 at tyrosine 1248 instarved/HRG-pulsed hESCs (Figure S6), consistent with the litera-ture.26 These findings demonstrated that disruption of ERBB2signaling severely inhibited hESC proliferation and inducedapoptosis.
In consideration of the roles of IGF1R and ERBB2 in self-renewal, a defined medium was assembled containing a combina-tion of IGF1R and ERBB2 ligands. Since IGF1 and IGF2 elicitextremely similar profiles of RTK phosphorylation (Figure S7),LONG R3 IGF1 (LR3-IGF1), a GMP-grade recombinant humanIGF1 that does not bind insulin-like growth factor–binding proteins(IGFBPs), was added to the medium. The ERBB2 ligand used inthe defined medium was the EGF domain of HRG, an EGF familymember which induces activation of the ERBB2/3 heterodimer.22
Figure 2. Disruption of IGF1R and ERBB2 signaling inhibits hESC self-renewal. (A) Treatment of H1 cells with the A12 anti-IGF1R blocking antibody led to decreasedIGF1R expression on the cell surface as measured by flow cytometry. (B) Colony counting showed that the A12 antibody inhibited H1 cell proliferation (left). Arrows indicate thatcultures were passaged on day 3 and 6. Cumulative colony counts increased in the presence of control antibody but not in the presence of A12. A12 also induced hESCdifferentiation (right) as measured by colony morphology (also see Figure S2). Error bars here and in panels C-H,J are standard deviation (SD). Flow cytometry for (C) SSEA-3expression confirmed A12 treatment caused increased differentiation, but (D) not an increase in apoptosis. (E) Significant reduction of IGF1R mRNA in the GFP� population ofcultures transfected with an IGF1R-targeted shRNA, compared with either the GFP- population, or GFP� cells transfected with a control shRNA. (F) Decline in the percentageof GFP� cells in H1 cultures (left) transduced with a lentiviral vector containing a shRNA targeting IGF1R, rather than a control shRNA. Middle panel shows flow cytometryprofiles that indicate stable expression of IGF1R and percentages of cells that are GFP� in control shRNA–transduced cultures on days 1, 12, and 22, while cultures transducedwith the IGF1R-targeted shRNA vector exhibited a decline in IGF1R expression and a reduction in GFP� and GFP�/IGF1R� cells. Right panel shows the average percentageof GFP� cells at days 15, 19, and 22 was significantly lower in the IGF1R-targeted shRNA transduced culture. (G) A total of 50 �M AG825 inhibited proliferation of BG02 hESCsgrowing in CM. Triplicate cell counts from 2 independent experiments are shown. . . . and - - - indicate pretreatment cell counts from experiments 1 and 2, respectively.(H,I) Increasing concentrations of AG825 caused a dose-dependent reduction in the proportion of H1 hESCs in the G1 phase of the cell cycle, and (J) a moderatedose-dependent rise in apoptosis.
4114 WANG et al BLOOD, 1 DECEMBER 2007 � VOLUME 110, NUMBER 12
For personal use only.on November 18, 2018. by guest www.bloodjournal.orgFrom
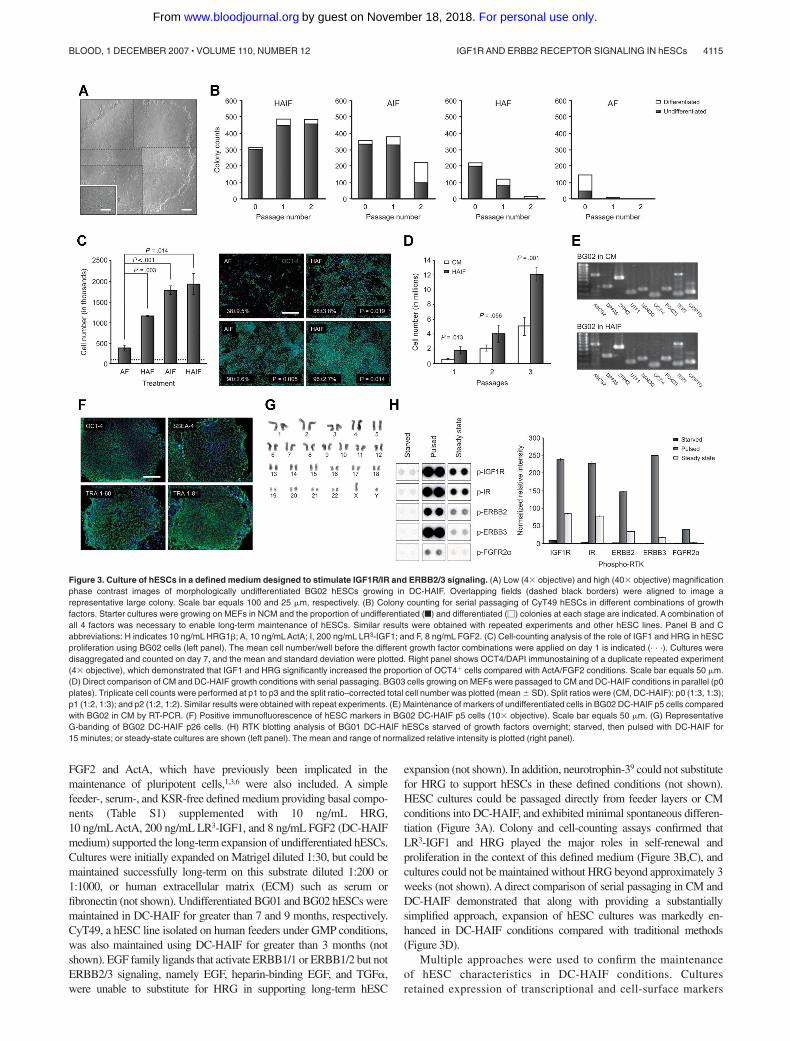
FGF2 and ActA, which have previously been implicated in themaintenance of pluripotent cells,1,3,6 were also included. A simplefeeder-, serum-, and KSR-free defined medium providing basal compo-nents (Table S1) supplemented with 10 ng/mL HRG,10 ng/mLActA, 200 ng/mL LR3-IGF1, and 8 ng/mL FGF2 (DC-HAIFmedium) supported the long-term expansion of undifferentiated hESCs.Cultures were initially expanded on Matrigel diluted 1:30, but could bemaintained successfully long-term on this substrate diluted 1:200 or1:1000, or human extracellular matrix (ECM) such as serum orfibronectin (not shown). Undifferentiated BG01 and BG02 hESCs weremaintained in DC-HAIF for greater than 7 and 9 months, respectively.CyT49, a hESC line isolated on human feeders under GMP conditions,was also maintained using DC-HAIF for greater than 3 months (notshown). EGF family ligands that activate ERBB1/1 or ERBB1/2 but notERBB2/3 signaling, namely EGF, heparin-binding EGF, and TGF�,were unable to substitute for HRG in supporting long-term hESC
expansion (not shown). In addition, neurotrophin-39 could not substitutefor HRG to support hESCs in these defined conditions (not shown).HESC cultures could be passaged directly from feeder layers or CMconditions into DC-HAIF, and exhibited minimal spontaneous differen-tiation (Figure 3A). Colony and cell-counting assays confirmed thatLR3-IGF1 and HRG played the major roles in self-renewal andproliferation in the context of this defined medium (Figure 3B,C), andcultures could not be maintained without HRG beyond approximately 3weeks (not shown). A direct comparison of serial passaging in CM andDC-HAIF demonstrated that along with providing a substantiallysimplified approach, expansion of hESC cultures was markedly en-hanced in DC-HAIF conditions compared with traditional methods(Figure 3D).
Multiple approaches were used to confirm the maintenanceof hESC characteristics in DC-HAIF conditions. Culturesretained expression of transcriptional and cell-surface markers
Figure 3. Culture of hESCs in a defined medium designed to stimulate IGF1R/IR and ERBB2/3 signaling. (A) Low (4� objective) and high (40� objective) magnificationphase contrast images of morphologically undifferentiated BG02 hESCs growing in DC-HAIF. Overlapping fields (dashed black borders) were aligned to image arepresentative large colony. Scale bar equals 100 and 25 �m, respectively. (B) Colony counting for serial passaging of CyT49 hESCs in different combinations of growthfactors. Starter cultures were growing on MEFs in NCM and the proportion of undifferentiated (f) and differentiated (�) colonies at each stage are indicated. A combination ofall 4 factors was necessary to enable long-term maintenance of hESCs. Similar results were obtained with repeated experiments and other hESC lines. Panel B and Cabbreviations: H indicates 10 ng/mL HRG1�; A, 10 ng/mL ActA; I, 200 ng/mL LR3-IGF1; and F, 8 ng/mL FGF2. (C) Cell-counting analysis of the role of IGF1 and HRG in hESCproliferation using BG02 cells (left panel). The mean cell number/well before the different growth factor combinations were applied on day 1 is indicated (. . .). Cultures weredisaggregated and counted on day 7, and the mean and standard deviation were plotted. Right panel shows OCT4/DAPI immunostaining of a duplicate repeated experiment(4� objective), which demonstrated that IGF1 and HRG significantly increased the proportion of OCT4� cells compared with ActA/FGF2 conditions. Scale bar equals 50 �m.(D) Direct comparison of CM and DC-HAIF growth conditions with serial passaging. BG03 cells growing on MEFs were passaged to CM and DC-HAIF conditions in parallel (p0plates). Triplicate cell counts were performed at p1 to p3 and the split ratio–corrected total cell number was plotted (mean � SD). Split ratios were (CM, DC-HAIF): p0 (1:3, 1:3);p1 (1:2, 1:3); and p2 (1:2, 1:2). Similar results were obtained with repeat experiments. (E) Maintenance of markers of undifferentiated cells in BG02 DC-HAIF p5 cells comparedwith BG02 in CM by RT-PCR. (F) Positive immunofluorescence of hESC markers in BG02 DC-HAIF p5 cells (10� objective). Scale bar equals 50 �m. (G) RepresentativeG-banding of BG02 DC-HAIF p26 cells. (H) RTK blotting analysis of BG01 DC-HAIF hESCs starved of growth factors overnight; starved, then pulsed with DC-HAIF for15 minutes; or steady-state cultures are shown (left panel). The mean and range of normalized relative intensity is plotted (right panel).
IGF1R AND ERBB2 RECEPTOR SIGNALING IN hESCs 4115BLOOD, 1 DECEMBER 2007 � VOLUME 110, NUMBER 12
For personal use only.on November 18, 2018. by guest www.bloodjournal.orgFrom
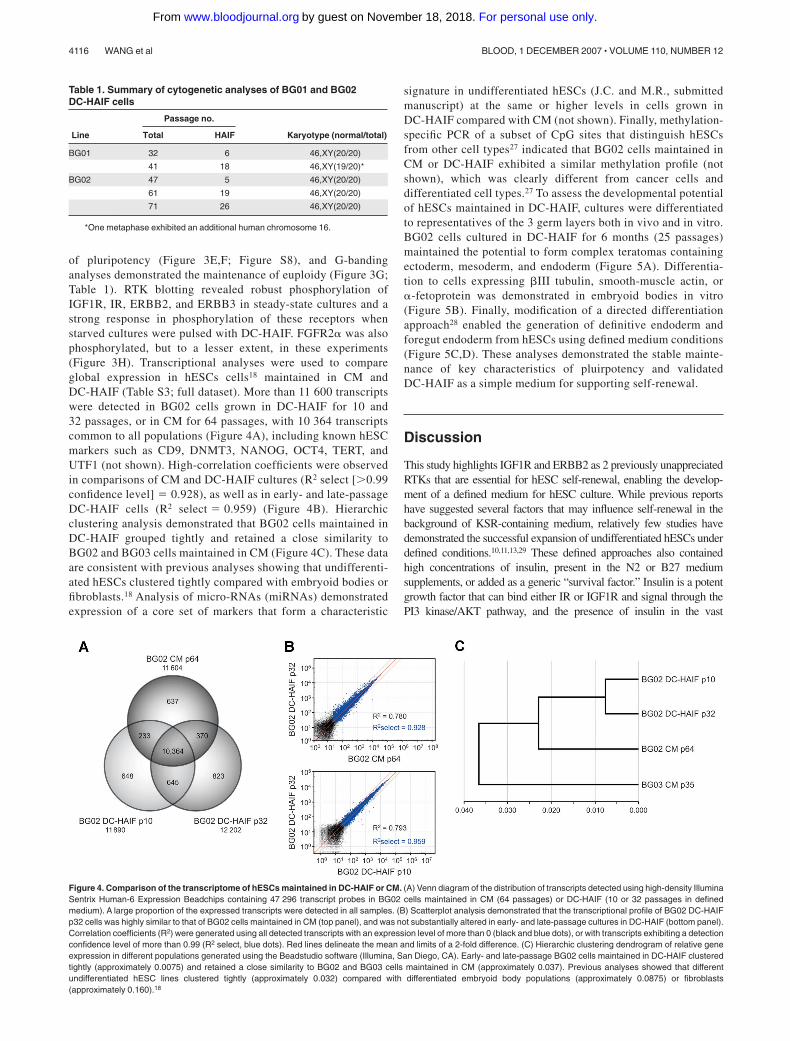
of pluripotency (Figure 3E,F; Figure S8), and G-bandinganalyses demonstrated the maintenance of euploidy (Figure 3G;Table 1). RTK blotting revealed robust phosphorylation ofIGF1R, IR, ERBB2, and ERBB3 in steady-state cultures and astrong response in phosphorylation of these receptors whenstarved cultures were pulsed with DC-HAIF. FGFR2� was alsophosphorylated, but to a lesser extent, in these experiments(Figure 3H). Transcriptional analyses were used to compareglobal expression in hESCs cells18 maintained in CM andDC-HAIF (Table S3; full dataset). More than 11 600 transcriptswere detected in BG02 cells grown in DC-HAIF for 10 and32 passages, or in CM for 64 passages, with 10 364 transcriptscommon to all populations (Figure 4A), including known hESCmarkers such as CD9, DNMT3, NANOG, OCT4, TERT, andUTF1 (not shown). High-correlation coefficients were observedin comparisons of CM and DC-HAIF cultures (R2 select [0.99confidence level] 0.928), as well as in early- and late-passageDC-HAIF cells (R2 select 0.959) (Figure 4B). Hierarchicclustering analysis demonstrated that BG02 cells maintained inDC-HAIF grouped tightly and retained a close similarity toBG02 and BG03 cells maintained in CM (Figure 4C). These dataare consistent with previous analyses showing that undifferenti-ated hESCs clustered tightly compared with embryoid bodies orfibroblasts.18 Analysis of micro-RNAs (miRNAs) demonstratedexpression of a core set of markers that form a characteristic
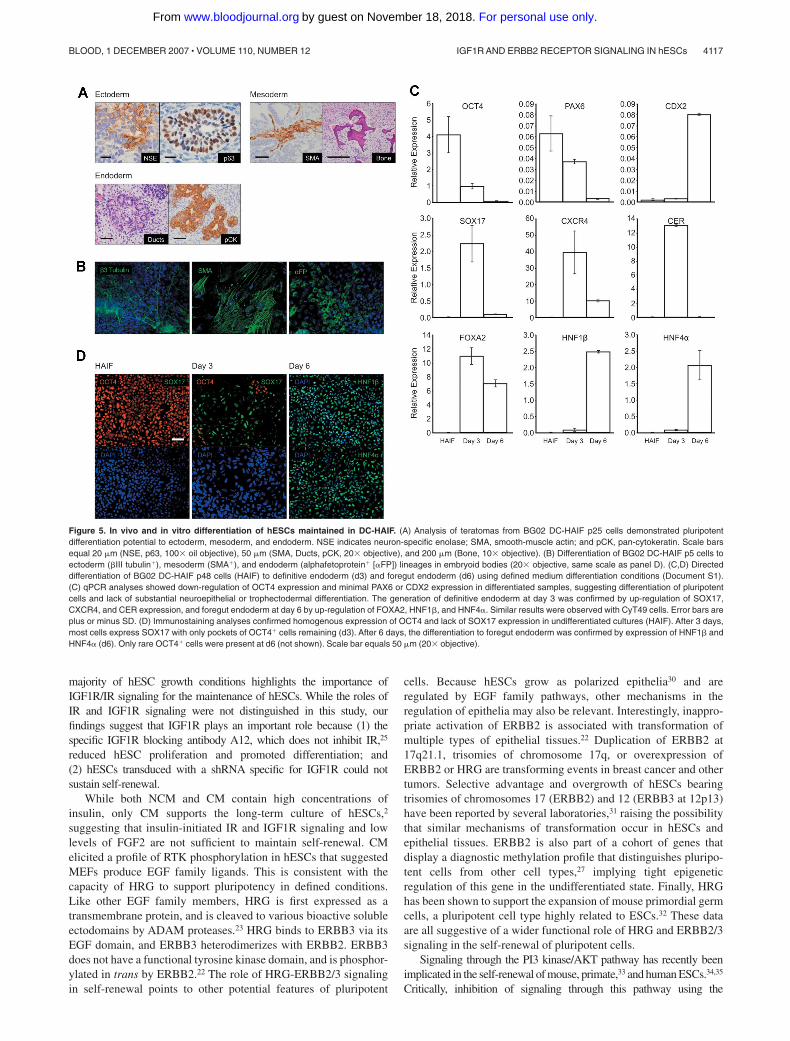
signature in undifferentiated hESCs (J.C. and M.R., submittedmanuscript) at the same or higher levels in cells grown inDC-HAIF compared with CM (not shown). Finally, methylation-specific PCR of a subset of CpG sites that distinguish hESCsfrom other cell types27 indicated that BG02 cells maintained inCM or DC-HAIF exhibited a similar methylation profile (notshown), which was clearly different from cancer cells anddifferentiated cell types.27 To assess the developmental potentialof hESCs maintained in DC-HAIF, cultures were differentiatedto representatives of the 3 germ layers both in vivo and in vitro.BG02 cells cultured in DC-HAIF for 6 months (25 passages)maintained the potential to form complex teratomas containingectoderm, mesoderm, and endoderm (Figure 5A). Differentia-tion to cells expressing �III tubulin, smooth-muscle actin, or�-fetoprotein was demonstrated in embryoid bodies in vitro(Figure 5B). Finally, modification of a directed differentiationapproach28 enabled the generation of definitive endoderm andforegut endoderm from hESCs using defined medium conditions(Figure 5C,D). These analyses demonstrated the stable mainte-nance of key characteristics of pluirpotency and validatedDC-HAIF as a simple medium for supporting self-renewal.
Discussion
This study highlights IGF1R and ERBB2 as 2 previously unappreciatedRTKs that are essential for hESC self-renewal, enabling the develop-ment of a defined medium for hESC culture. While previous reportshave suggested several factors that may influence self-renewal in thebackground of KSR-containing medium, relatively few studies havedemonstrated the successful expansion of undifferentiated hESCs underdefined conditions.10,11,13,29 These defined approaches also containedhigh concentrations of insulin, present in the N2 or B27 mediumsupplements, or added as a generic “survival factor.” Insulin is a potentgrowth factor that can bind either IR or IGF1R and signal through thePI3 kinase/AKT pathway, and the presence of insulin in the vast
Table 1. Summary of cytogenetic analyses of BG01 and BG02DC-HAIF cells
Line
Passage no.
Karyotype (normal/total)Total HAIF
BG01 32 6 46,XY(20/20)
41 18 46,XY(19/20)*
BG02 47 5 46,XY(20/20)
61 19 46,XY(20/20)
71 26 46,XY(20/20)
*One metaphase exhibited an additional human chromosome 16.
Figure 4. Comparison of the transcriptome of hESCs maintained in DC-HAIF or CM. (A) Venn diagram of the distribution of transcripts detected using high-density IlluminaSentrix Human-6 Expression Beadchips containing 47 296 transcript probes in BG02 cells maintained in CM (64 passages) or DC-HAIF (10 or 32 passages in definedmedium). A large proportion of the expressed transcripts were detected in all samples. (B) Scatterplot analysis demonstrated that the transcriptional profile of BG02 DC-HAIFp32 cells was highly similar to that of BG02 cells maintained in CM (top panel), and was not substantially altered in early- and late-passage cultures in DC-HAIF (bottom panel).Correlation coefficients (R2) were generated using all detected transcripts with an expression level of more than 0 (black and blue dots), or with transcripts exhibiting a detectionconfidence level of more than 0.99 (R2 select, blue dots). Red lines delineate the mean and limits of a 2-fold difference. (C) Hierarchic clustering dendrogram of relative geneexpression in different populations generated using the Beadstudio software (Illumina, San Diego, CA). Early- and late-passage BG02 cells maintained in DC-HAIF clusteredtightly (approximately 0.0075) and retained a close similarity to BG02 and BG03 cells maintained in CM (approximately 0.037). Previous analyses showed that differentundifferentiated hESC lines clustered tightly (approximately 0.032) compared with differentiated embryoid body populations (approximately 0.0875) or fibroblasts(approximately 0.160).18
4116 WANG et al BLOOD, 1 DECEMBER 2007 � VOLUME 110, NUMBER 12
For personal use only.on November 18, 2018. by guest www.bloodjournal.orgFrom
majority of hESC growth conditions highlights the importance ofIGF1R/IR signaling for the maintenance of hESCs. While the roles ofIR and IGF1R signaling were not distinguished in this study, ourfindings suggest that IGF1R plays an important role because (1) thespecific IGF1R blocking antibody A12, which does not inhibit IR,25
reduced hESC proliferation and promoted differentiation; and(2) hESCs transduced with a shRNA specific for IGF1R could notsustain self-renewal.
While both NCM and CM contain high concentrations ofinsulin, only CM supports the long-term culture of hESCs,2
suggesting that insulin-initiated IR and IGF1R signaling and lowlevels of FGF2 are not sufficient to maintain self-renewal. CMelicited a profile of RTK phosphorylation in hESCs that suggestedMEFs produce EGF family ligands. This is consistent with thecapacity of HRG to support pluripotency in defined conditions.Like other EGF family members, HRG is first expressed as atransmembrane protein, and is cleaved to various bioactive solubleectodomains by ADAM proteases.23 HRG binds to ERBB3 via itsEGF domain, and ERBB3 heterodimerizes with ERBB2. ERBB3does not have a functional tyrosine kinase domain, and is phosphor-ylated in trans by ERBB2.22 The role of HRG-ERBB2/3 signalingin self-renewal points to other potential features of pluripotent
cells. Because hESCs grow as polarized epithelia30 and areregulated by EGF family pathways, other mechanisms in theregulation of epithelia may also be relevant. Interestingly, inappro-priate activation of ERBB2 is associated with transformation ofmultiple types of epithelial tissues.22 Duplication of ERBB2 at17q21.1, trisomies of chromosome 17q, or overexpression ofERBB2 or HRG are transforming events in breast cancer and othertumors. Selective advantage and overgrowth of hESCs bearingtrisomies of chromosomes 17 (ERBB2) and 12 (ERBB3 at 12p13)have been reported by several laboratories,31 raising the possibilitythat similar mechanisms of transformation occur in hESCs andepithelial tissues. ERBB2 is also part of a cohort of genes thatdisplay a diagnostic methylation profile that distinguishes pluripo-tent cells from other cell types,27 implying tight epigeneticregulation of this gene in the undifferentiated state. Finally, HRGhas been shown to support the expansion of mouse primordial germcells, a pluripotent cell type highly related to ESCs.32 These dataare all suggestive of a wider functional role of HRG and ERBB2/3signaling in the self-renewal of pluripotent cells.
Signaling through the PI3 kinase/AKT pathway has recently beenimplicated in the self-renewal of mouse, primate,33 and human ESCs.34,35
Critically, inhibition of signaling through this pathway using the
Figure 5. In vivo and in vitro differentiation of hESCs maintained in DC-HAIF. (A) Analysis of teratomas from BG02 DC-HAIF p25 cells demonstrated pluripotentdifferentiation potential to ectoderm, mesoderm, and endoderm. NSE indicates neuron-specific enolase; SMA, smooth-muscle actin; and pCK, pan-cytokeratin. Scale barsequal 20 �m (NSE, p63, 100� oil objective), 50 �m (SMA, Ducts, pCK, 20� objective), and 200 �m (Bone, 10� objective). (B) Differentiation of BG02 DC-HAIF p5 cells toectoderm (�III tubulin�), mesoderm (SMA�), and endoderm (alphafetoprotein� [�FP]) lineages in embryoid bodies (20� objective, same scale as panel D). (C,D) Directeddifferentiation of BG02 DC-HAIF p48 cells (HAIF) to definitive endoderm (d3) and foregut endoderm (d6) using defined medium differentiation conditions (Document S1).(C) qPCR analyses showed down-regulation of OCT4 expression and minimal PAX6 or CDX2 expression in differentiated samples, suggesting differentiation of pluripotentcells and lack of substantial neuroepithelial or trophectodermal differentiation. The generation of definitive endoderm at day 3 was confirmed by up-regulation of SOX17,CXCR4, and CER expression, and foregut endoderm at day 6 by up-regulation of FOXA2, HNF1�, and HNF4�. Similar results were observed with CyT49 cells. Error bars areplus or minus SD. (D) Immunostaining analyses confirmed homogenous expression of OCT4 and lack of SOX17 expression in undifferentiated cultures (HAIF). After 3 days,most cells express SOX17 with only pockets of OCT4� cells remaining (d3). After 6 days, the differentiation to foregut endoderm was confirmed by expression of HNF1� andHNF4� (d6). Only rare OCT4� cells were present at d6 (not shown). Scale bar equals 50 �m (20� objective).
IGF1R AND ERBB2 RECEPTOR SIGNALING IN hESCs 4117BLOOD, 1 DECEMBER 2007 � VOLUME 110, NUMBER 12
For personal use only.on November 18, 2018. by guest www.bloodjournal.orgFrom

small-molecule antagonists LY294002 (PI3 kinase inhibitor) andAKT1-II(AKT1 inhibitor) induced differentiation of hESCs.34 Treatment ofhESCs with LY294002 was associated with a rapid collapse inphosphorylation of AKT1, pS6, pS6K/p70, and GSK3�.35 We haveconfirmed the importance role of this pathway by demonstrating that1 to 20 nM rapamycin, an inhibitor of FRAP1 (mTOR), severelyrestricted the proliferation of hESCs in DC-HAIF and induced differen-tiation in short term culture (not shown). Because the PI3 kinasepathway delivers both strong proliferative and antiapoptotic signals,IR/IGF1R signaling should not be overlooked when interpreting theeffects of factors that feed into this common pathway, including FGF2,PDGF, and neurotrophins.3,7,9,11 PI3 kinase signaling leads to theactivation of AKT and AKT-mediated inhibition of GSK3�, andtherefore the presence of insulin also complicates the interpretation thatsmall-molecule inhibitors of GSK3� are mimicking the effects of WNTsignaling.8 It is possible that multiple inputs are required for appropriatesignaling and regulation of self-renewal through PI3 kinase in hESCs,which in this study was driven by activation of IR, IGF1R, andERBB2/3.
This study identified IGF1R and ERBB2 as RTKs involved inself-renewal of hESCs and enabled the development of a definedmedium based around activation of these receptors. We have usedthis strategy to demonstrate the capacity for large-scale expansionof undifferentiated hESCs. A starting confluent culture of BG02cells in one 60-mm plate was expanded in DC-HAIF through4 passages over 20 days to generate more than 1010 cells (T.S.,A.R., unpublished data, December 2006). The ability to massivelyexpand undifferentiated hESC cultures, coupled with effectivedifferentiation approaches that also use defined conditions togenerate clinically relevant populations,28 lays important ground-work for eventual clinical applications.
Note added in proof: A requirement for IGF1R signaling inhESC self-renewal was also reported by another group36 while thismanuscript was under review.
Acknowledgments
We would like to thank Dr Steve Plymate for the A12 antibody;members of the Blau Lab for helpful discussions; Paul Fields forexpert assistance; and Dr Clifton Baile, Diane Hartzel, and the
Animal Facility, Animal and Dairy Science Department, Universityof Georgia, for assisting with generating teratomas.
This work was supported by grants from the NationalInstitute of General Medical Sciences at the NIH(1P01GM081619-01; C.A.B. and C.W.) and the National Insti-tute of Research Resources (9R24RR021313–04 to T.S.). Fed-eral funding was not used for non–NIH-registered hESC lines.
Authorship
Contribution: L.W. designed, performed, and supervised experi-ments, interpreted findings, and assisted in writing the manuscript.T.C.S. designed, performed, and supervised experiments, inter-preted findings, and wrote and assembled the manuscript. E.S.S.,S.N.B., A.M.L., and M.J.G. performed defined medium studies andother experiments. D.S.D. assisted with experiments and con-structed figures. S.S. performed and interpreted microarray andother molecular analyses. A.M.N. and C.B.W. assisted with thegeneration of hESC cultures. M.Z. and C.-Z.S. designed andconstructed lentiviral shRNA vectors. X.C. performed qPCRanalysis of shRNA experiments. E.W.U. performed histologicanalyses of teratomas. K.A.D. performed qPCR analysis ofendoderm differentiations. J.D.C. supervised and interpreted mi-croarray and other molecular analyses. M.S.R. conceived, directed,and interpreted microarray and molecular analyses. C.A.B. con-ceived, designed, and supervised investigation of MEF-CM, inter-preted findings, and assisted in writing the manuscript. A.J.R.conceived the defined medium study, assisted design and supervi-sion of experiments, interpreted findings, and assisted in writingmanuscript. L.W. and T.C.S. contributed equally to this manuscript.
Conflict-of-interest disclosure: T.C.S., E.S.S., S.N.B., M.J.G.,K.A.D., and A.J.R. are employees and have stock options in acompany that is not publicly traded (Novocell). S.S., J.D.C., andM.S.R. are employees and have ownership interest in a publiclytraded company (Invitrogen).
Correspondence: Allan J. Robins, Novocell Inc, 111 RiverbendRd, Athens, GA 30602; e-mail: [email protected]; orC. Anthony Blau, Mailstop 357710, K260 Health Sciences Build-ing, University of Washington, Seattle, WA 98195; e-mail:[email protected].
References
1. Xu C, Rosler E, Jiang J, et al. Basic fibroblastgrowth factor supports undifferentiated humanembryonic stem cell growth without conditionedmedium. Stem Cells. 2005;23:315-323.
2. Xu C, Inokuma MS, Denham J, et al. Feeder-freegrowth of undifferentiated human embryonic stemcells. Nat Biotechnol. 2001;19:971-974.
3. Levenstein ME, Ludwig TE, Xu RH, et al. Basicfibroblast growth factor support of human embry-onic stem cell self-renewal. Stem Cells. 2006;24:568-574.
4. Amit M, Shariki C, Margulets V, Itskovitz-Eldor J.Feeder layer- and serum-free culture of humanembryonic stem cells. Biol Reprod. 2004;70:837-845.
5. James D, Levine AJ, Besser D, Hemmati-Brivan-lou A. TGFbeta/activin/nodal signaling is neces-sary for the maintenance of pluripotency in hu-man embryonic stem cells. Development. 2005;132:1273-1282.
6. Beattie GM, Lopez AD, Bucay N, et al. Activin Amaintains pluripotency of human embryonic stemcells in the absence of feeder layers. Stem Cells.2005;23:489-495.
7. Pebay A, Wong RC, Pitson SM, et al. Essentialroles of sphingosine-1-phosphate and platelet-derived growth factor in the maintenance of hu-man embryonic stem cells. Stem Cells. 2005;23:1541-1548.
8. Sato N, Meijer L, Skaltsounis L, Greengard P,Brivanlou AH. Maintenance of pluripotency in hu-man and mouse embryonic stem cells throughactivation of Wnt signaling by a pharmacologicalGSK-3-specific inhibitor. Nat Med. 2004;10:55-63.
9. Pyle AD, Lock LF, Donovan PJ. Neurotrophinsmediate human embryonic stem cell survival. NatBiotechnol. 2006;24:344-350.
10. Vallier L, Alexander M, Pedersen RA. Activin/Nodal and FGF pathways cooperate to maintainpluripotency of human embryonic stem cells.J Cell Sci. 2005;118:4495-4509.
11. Ludwig TE, Levenstein ME, Jones JM, et al. Deri-vation of human embryonic stem cells in definedconditions. Nat Biotechnol. 2006;24:185-187.
12. Lu J, Hou R, Booth CJ, Yang SH, Snyder M. De-fined culture conditions of human embryonic stem
cells. Proc Natl Acad Sci U S A. 2006;103:5688-5693.
13. Yao S, Chen S, Clark J, et al. Long-term self-re-newal and directed differentiation of human em-bryonic stem cells in chemically defined condi-tions. Proc Natl Acad Sci U S A. 2006;103:6907-6912.
14. Sperger JM, Chen X, Draper JS, et al. Gene ex-pression patterns in human embryonic stem cellsand human pluripotent germ cell tumors. ProcNatl Acad Sci U S A. 2003;100:13350-13355.
15. Dvorak P, Dvorakova D, Koskova S, et al. Ex-pression and potential role of fibroblast growthfactor 2 and its receptors in human embryonicstem cells. Stem Cells. 2005;23:1200-1211.
16. Dvash T, Mayshar Y, Darr H, et al. Temporal geneexpression during differentiation of human embry-onic stem cells and embryoid bodies. Hum Re-prod. 2004;19:2875-2883.
17. Schlessinger J. Cell signaling by receptor ty-rosine kinases. Cell. 2000;103:211-225.
18. Liu Y, Shin S, Zeng X, et al. Genome wide profil-ing of human embryonic stem cells (hESCs), their
4118 WANG et al BLOOD, 1 DECEMBER 2007 � VOLUME 110, NUMBER 12
For personal use only.on November 18, 2018. by guest www.bloodjournal.orgFrom

derivatives and embryonal carcinoma cells to de-velop base profiles of U.S. Federal governmentapproved hESC lines. BMC Dev Biol. 2006;6:20.
19. Neely EK, Morhenn VB, Hintz RL, Wilson DM,Rosenfeld RG. Insulin-like growth factors are mi-togenic for human keratinocytes and a squamouscell carcinoma. J Invest Dermatol. 1991;96:104-110.
20. Li G, Barrett EJ, Wang H, Chai W, Liu Z. Insulin atphysiological concentrations selectively activatesinsulin but not insulin-like growth factor I (IGF-I)or insulin/IGF-I hybrid receptors in endothelialcells. Endocrinology. 2005;146:4690-4696.
21. Oda K, Matsuoka Y, Funahashi A, Kitano H. Acomprehensive pathway map of epidermalgrowth factor receptor signaling. Mol Syst Biol.2005;1:2005-0010.
22. Citri A, Skaria KB, Yarden Y. The deaf and thedumb: the biology of ErbB-2 and ErbB-3. Exp CellRes. 2003;284:54-65.
23. Seals DF, Courtneidge SA. The ADAMs family ofmetalloproteases: multidomain proteins with mul-tiple functions. Genes Dev. 2003;17:7-30.
24. Schlessinger J. Common and distinct elements incellular signaling via EGF and FGF receptors.Science. 2004;306:1506-1507.
25. Burtrum D, Zhu Z, Lu D, et al. A fully humanmonoclonal antibody to the insulin-like growthfactor I receptor blocks ligand-dependent signal-ing and inhibits human tumor growth in vivo. Can-cer Res. 2003;63:8912-8921.
26. Murillo H, Schmidt LJ, Tindall DJ. TyrphostinAG825 triggers p38 mitogen-activated proteinkinase-dependent apoptosis in androgen-inde-pendent prostate cancer cells C4 and C4–2. Can-cer Res. 2001;61:7408-7412.
27. Bibikova M, Chudin E, Wu B, et al. Human em-bryonic stem cells have a unique epigenetic sig-nature. Genome Res. 2006;16:1075-1083.
28. D’Amour KA, Bang AG, Eliazer S, et al. Produc-tion of pancreatic hormone-expressing endocrinecells from human embryonic stem cells. Nat Bio-technol. 2006;24:1392-1401.
29. Ludwig TE, Bergendahl V, Levenstein ME, Yu J,Probasco MD, Thomson JA. Feeder-independentculture of human embryonic stem cells. Nat Meth-ods. 2006;3:637-646.
30. Krtolica A, Genbacev O, Escobedo C, et al. Dis-ruption of apical-basal polarity of human embry-onic stem cells enhances hematoendothelial dif-ferentiation. Stem Cells. 2007;25:2215-2223.
31. Draper JS, Smith K, Gokhale P, et al. Recurrent
gain of chromosomes 17q and 12 in cultured hu-man embryonic stem cells. Nat Biotechnol. 2004;22:53-54.
32. Toyoda-Ohno H, Obinata M, Matsui Y. Membersof the ErbB receptor tyrosine kinases are in-volved in germ cell development in fetal mousegonads. Dev Biol. 1999;215:399-406.
33. Watanabe S, Umehara H, Murayama K, OkabeM, Kimura T, Nakano T. Activation of Akt signalingis sufficient to maintain pluripotency in mouseand primate embryonic stem cells. Oncogene.2006;25:2697-2707.
34. Armstrong L, Hughes O, Yung S, et al. The role ofPI3K/AKT, MAPK/ERK and NFkappabeta signal-ling in the maintenance of human embryonicstem cell pluripotency and viability highlighted bytranscriptional profiling and functional analysis.Hum Mol Genet. 2006;15:1894-1913.
35. McLean AB, D’Amour KA, Jones KL, et al. ActivinA efficiently specifies definitive endoderm fromhuman embryonic stem cells only when phospha-tidylinositol 3-kinase signaling is suppressed.Stem Cells. 2007;25:29-38.
36. Bendall SC, Stewart MH, Menendez P, et al. IGFand FGF cooperatively establish the regulatorystem cell niche of pluripotent human cells in vitro.Nature. 2007;448:1015-1021.
IGF1R AND ERBB2 RECEPTOR SIGNALING IN hESCs 4119BLOOD, 1 DECEMBER 2007 � VOLUME 110, NUMBER 12
For personal use only.on November 18, 2018. by guest www.bloodjournal.orgFrom

online August 29, 2007 originally publisheddoi:10.1182/blood-2007-03-082586
2007 110: 4111-4119
Rao, C. Anthony Blau and Allan J. RobinsMcLean, Maria J. Galeano, Elizabeth W. Uhl, Kevin A. D'Amour, Jonathan D. Chesnut, Mahendra S.Nelson, Carol B. Ware, Mei Zhan, Chao-Zhong Song, Xiaoji Chen, Sandii N. Brimble, Amanda Linlin Wang, Thomas C. Schulz, Eric S. Sherrer, Derek S. Dauphin, Soojung Shin, Angelique M. factor-1 receptor and ERBB2 receptor signalingSelf-renewal of human embryonic stem cells requires insulin-like growth
http://www.bloodjournal.org/content/110/12/4111.full.htmlUpdated information and services can be found at:
(166 articles)Stem Cells in Hematology (1930 articles)Signal Transduction
(3527 articles)Hematopoiesis and Stem Cells Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society
For personal use only.on November 18, 2018. by guest www.bloodjournal.orgFrom