seed biology and rehabilitation in the arid zone: a study in the … · shark bay salt (sbs) has...
TRANSCRIPT

Seed biology and rehabilitation in the arid zone:
a study in the Shark Bay World Heritage Area,
Western Australia
Lucy Commander
BSc (Hons)
This thesis is presented for
the degree of Doctor of Philosophy of
The University of Western Australia
School of Plant Biology
2008


i
DECLARATION FOR THESES CONTAINING PUBLISHED WORK AND/OR WORK
PREPARED FOR PUBLICATION
The examination of the thesis is an examination of the work of the student. The work must have been substantially conducted by the student during enrolment in the degree. Where the thesis includes work to which others have contributed, the thesis must include a statement that makes the student’s contribution clear to the examiners. This may be in the form of a description of the precise contribution of the student to the work presented for examination and/or a statement of the percentage of the work that was done by the student. In addition, in the case of co-authored publications included in the thesis, each author must give their signed permission for the work to be included. If signatures from all the authors cannot be obtained, the statement detailing the student’s contribution to the work must be signed by the coordinating supervisor. Please sign one of the statements below.
1. This thesis does not contain work that I have published, nor work under review for publication.
2. This thesis contains only sole-authored work, some of which has been published and/or prepared for publication under sole authorship. The bibliographical details of the work and where it appears in the thesis are outlined below.
3. This thesis contains published work and/or work prepared for publication, some of which has been co-authored. The bibliographical details of the work and where it appears in the thesis are outlined below. The student must attach to this declaration a statement for each publication that clarifies the contribution of the student to the work. This may be in the form of a description of the precise contributions of the student to the published work and/or a statement of percent contribution by the student. This statement must be signed by all authors. If signatures from all the authors cannot be obtained, the statement detailing the student’s contribution to the published work must be signed by the coordinating supervisor. The publications arising from the thesis are original work undertaken by the student (Lucy Commander), with guidance from her three supervisors (Kingsley Dixon, David Merritt and Deanna Rokich). Signature................................................................................................................................
Signature................................................................................................................................
Signature................................................................................................................................
Signature................................................................................................................................

ii
Statement of candidate contribution
This thesis contains published work and/or work prepared for publication, which has
been co-authored. The publications arising from the thesis are original work undertaken
by the student (Lucy Commander), with guidance from her three supervisors (Kingsley
Dixon, David Merritt and Deanna Rokich). The bibliographical details and location of
the work are outlined below.
One of the chapters from this thesis is published. The text in this chapter appears
as published.
CHAPTER 4:
Commander LE, Merritt, DJ, Rokich DP, Flematti GR, Dixon KW (2008) Seed
germination of Solanum spp. (Solanaceae) for use in rehabilitation and commercial
industries. Australian Journal of Botany 56 333-341
Two of the chapters from this thesis are manuscripts in review. The text in these
chapters appears as in the manuscripts.
CHAPTER 2:
Commander LE, Merritt DJ, Rokich DP, Dixon KW Seed germination of arid zone
species: Germination biology of 18 Australian species. Journal of Arid Environments
(submitted)
CHAPTER 3:
Commander LE, Merritt DJ, Rokich DP, Dixon KW Storage conditions can influence
after-ripening of arid zone seeds. Plant Ecology (submitted)
Two of the chapters from this thesis are in preparation for publication.
CHAPTERS 6 + 7:
Commander LE, Rokich DP, Merritt DJ, Dixon KW Seed broadcasting and greenstock
planting for mine rehabilitation in the Australian arid zone (in prep)

iii
Abstract
Research into seed biology and restoration ecology of areas disturbed by mining is
crucial to their revegetation. Shark Bay Salt, a solar salt facility in the Shark Bay World
Heritage Area in Western Australia has several areas of disturbance as a result of
‗soil borrowing‘. Soil from these areas termed ‗borrow pits‘ was used to create
infrastructure such as the roads and embankments surrounding the evaporation ponds.
Many of the pits contain little to no vegetation after >10 years since disturbance ceased,
hence research into their restoration is now essential.
A vegetation survey at the site established the key species in the undisturbed vegetation,
and investigated the vegetation in borrow pits subject to natural migration and topsoil
replacement. The vegetation communities in the borrow pits were vastly different to
those in the undisturbed vegetation, highlighting the need for research into revegetation.
An investigation into the use of ‗borrowed‘ topsoil on a small scale showed that
seedling recruitment from ‗borrowed‘ topsoil was generally similar in the donor site
(natural vegetation) and the borrow pits. Due to the absence of topsoil for further
revegetation, it was necessary to understand seed germination and dormancy
characteristics to establish seed pre-treatments prior to seed broadcasting and seedling
(greenstock) planting.
An investigation into seed germination and dormancy characteristics of 18 common
species revealed that most species germinated equally well at 26/13oC and 33/18
oC,
however seven species had improved performance at 26/13oC. Untreated seeds of seven
species exhibited high germination. Seeds of two species had low imbibition, which
increased with hot-water treatment, and hence require scarification for germination.
Germination of seeds of three species substantially increased with gibberellic acid
(GA3), smoke water (SW) and karrikinolide (KAR1, a butenolide isolated from smoke).
Seeds of the remaining six species had low germination regardless of treatment. As a
result, species were classified as likely to be non-dormant (44%), physiologically
dormant (44%) or physically dormant (11%).
Physiological dormancy of three species was at least partly alleviated by dry after-
ripening, whereby moisture content of seeds was adjusted to 13% or 50% equilibrium
relative humidity and seeds were stored at 30oC or 45
oC for several months. All

iv
after-ripening conditions increased germination percentage and rate of two species with
one only germinating when treated with GA3 or KAR1. The germination of the third
species was dependent on after-ripening temperature and seed moisture content. After-
ripening had a limited effect on three additional species tested.
Germination of one of the non-dormant species, Solanum orbiculatum was stimulated
by GA3, SW and KAR1. Given that this result was not reflective of other Solanum
species responses, this genera became a case-study genera. Germination characteristics
of seven other Australian Solanum species were not all similar, with high germination of
two species without treatment, high germination of two additional species with GA3,
SW and KAR1, and stimulation of germination of the remaining three species with GA3
only, highlighting species specific responses within genera.
Once seed germination and dormancy were understood, seed pre-treatments (GA3,
KAR1, SW, hot water) were developed, this enabled investigation into optimising seed
broadcasting and greenstock survival. Seed broadcasting was affected by both year and
season, with seedling emergence higher in the wetter year (2005) compared to the drier
year (2006) and higher when seeds were sown in autumn compared with winter. In the
wetter year, soil ripping and raking increased seedling emergence. There were
differences in survival of greenstock between the two years, which may partly be due to
rainfall, and also possibly due to differences in seedling quality. In the second year,
several treatments were investigated to optimise the growth and survival of greenstock,
and soil ripping was the treatment with the largest positive effect on survival. The other
treatments had minimal effect on the two focus species, apart from pruning, which had a
detrimental effect on survival of Atriplex bunburyana, and fertiliser, which had a
positive effect on growth of Atriplex bunburyana. Survival of Atriplex bunburyana was
substantially higher than the Acacia tetragonophylla and this may be at least partly due
to the high soil electrical conductivity in two of the borrow pits, given that Atriplex
bunburyana is a member of the salt-tolerant genus Atriplex.
This study provides information to assist with the restoration of borrow pits at SBS, and
findings may be applied to other arid areas in Western Australia, and elsewhere.

v
Table of Contents
Statement of candidate contribution
Abstract
Table of Contents
Acknowledgements
i
iii
v
vii
CHAPTER 1. General introduction and literature review
1.1
1.2
1.3
1.4
1.5
Study site
Rehabilitation
Seed germination
Seed dormancy
Rational aim and thesis outline
1
8
16
22
29
SECTION I. Seed Biology
CHAPTER 2. Seed biology of 18 species used for rehabilitation
2.1
2.2
2.3
2.4
2.5
Abstract
Introduction
Methods
Results
Discussion
31
32
34
37
46
CHAPTER 3. Storage conditions can influence after-ripening
of arid zone seeds
3.1
3.2
3.3
3.4
3.5
Abstract
Introduction
Methods
Results
Discussion
51
52
55
58
65
CHAPTER 4. Case study: Seed germination of Solanum spp.
from arid Australia
4.1
4.2
4.3
4.4
4.5
Abstract
Introduction
Methods
Results
Discussion
69
70
72
76
82

vi
SECTION II. Rehabilitation
CHAPTER 5. Assessment of borrow pit vegetation and
surrounding undisturbed vegetation
5.1
5.2
5.3
5.4
5.5
Abstract
Introduction
Methods
Results
Discussion
87
88
89
96
103
CHAPTER 6. Topsoil replacement and broadcast seeding in
borrow pits
6.1
6.2
6.3
6.4
6.5
Abstract
Introduction
Methods
Results
Discussion
107
108
110
112
120
CHAPTER 7. Greenstock treatments to optimise plant growth
and survival in borrow pits
7.1
7.2
7.3
7.4
7.5
Abstract
Introduction
Methods
Results
Discussion
125
126
130
135
149
CHAPTER 8. General Discussion
8.1
8.2
8.3
8.4
8.5
Introduction
Floristics
Investigation into seed germination requirements
and dormancy alleviation
Restoration ecology of disturbed land at Shark Bay
Salt
Conclusion
157
159
159
166
174
References
Appendices
175
193

vii
Acknowledgements
Firstly, thanks to my supervisors, Kingsley Dixon, David Merritt and Deanna Rokich,
without whom this research would not have been possible.
Funding for this research was provided by Shark Bay Salt whose staff also provided
logistical support for the collection of some seeds used in this study, and assistance in
implementing the field trials. In particular, thanks go to Peter Newstead, Max Le
Clercq and Colin Thomas.
A huge thanks goes to the following people:
Jeff Walck, Siti Hidayati and Shane Turner for their comments to improve the
manuscript.
Staff, students and volunteers at Kings Park who assisted me on my numerous field
trips.
The Kings Park Master Gardeners for their assistance with seed cleaning.
Philip Commander and Craig Miskell for their assistance in developing Figure 1.5
Karrikinolide was provided by Gavin Flematti of The University of Western Australia.
Seed of Solanum centrale was kindly donated by Alice Springs Desert Park and seed of
Solanum chippendalei was kindly donated by Kim Courtenay of Broome TAFE.
Thanks also go to Bronwynne York for technical support.
Thanks to the Minerals and Energy Research Institute of Western Australia (MERIWA)
for awarding me a post-graduate scholarship.
Thanks to my parents for their support throughout my education.
A special thanks to Craig Miskell.

viii

1
C H A P T E R 1
General introduction and literature review
1.1 Study site
1.1.1 Background
Shark Bay Salt (SBS) has been operating a solar salt facility since 1965 at the small
townsite of Useless Loop, located approximately 900 km north of Perth on the Edel
Peninsula in the Shark Bay World Heritage Area (Fig. 1.1). In 2004, SBS produced
1,397,000 tonnes of raw salt from the 6,031 ha of primary ponds, 397 ha of secondary
ponds and 489 ha of crystalliser ponds. These ponds are bounded by roads and bunds
(Appendix 1, 2). Over the years, soil for the construction of these roads and bunds has
been excavated from a series of pits, which had been cleared of their vegetation, and are
termed ‗borrow pits‘ (Appendix 1, 2). There are currently 31 borrow pits encompassing
an area of 83.7 ha. While some of these borrow pits have naturally revegetated to some
extent over time and have been left untreated, others have been subjected to topsoil
replacement, seeding or brushing (Fig. 1.2). In all instances (except topsoil
replacement), there is evidence of limited or no re-vegetation by natural means, and
consequently, borrow pits will need to be enriched with a great diversity of flora to
reflect the pre-existing vegetation. To date there is little legal obligation for SBS to
rehabilitate the borrow pits. However, the Shark Bay Solar Salt Industry Agreement Act
(1983) states: ‗The Joint Venturers shall in respect of the matters referred to in Clause 7
which are the subject of approved proposals under this Agreement, carry out a
continuous programme of investigation and research including monitoring and the study
of sample areas to ascertain the effectiveness of the measures they are taking pursuant to
their approved proposals for the protection and management of the environment‘. As a
result, SBS is committed to undertaking restoration research to improve the diversity
and health of the ecosystems that are impacted by borrow pit construction, particularly
given the area‘s World Heritage status.
Little is known about the conservation and rehabilitation needs and research into
restoration capacity of the World Heritage Area. Furthermore, the use of rehabilitation
principles developed from other disturbance activities (e.g. mining) may not always be

2
appropriate since the nearest available restoration sites within a mining context are 600
km south and are in a different biome (Southwest Australian Floristic Region (Hopper
and Gioia 2004)). In addition, rehabilitation principles may not be applicable due to the
site‘s unique characteristics, including its variable rainfall patterns given the close
proximity to the summer-winter rainfall line, its calcareous soils, and its vegetation
complexes, which are not often subject to rehabilitation.
Figure 1.1. Location of Useless Loop in Shark Bay, Western Australia
To begin setting rehabilitation targets at SBS, it will be necessary to understand the
species diversity at the site by undertaking a vegetation survey. In addition, surveying
the vegetation in the borrow pits will be necessary to quantify the extent to which the
vegetation has returned, and identify key species that are absent.
To enable rehabilitation principles to be developed, seed germination and dormancy
research should be undertaken in order to determine pre-treatments for seeds that will be
directly hand sown (broadcast) to site and/or used to produce seedlings. Limited
understanding of seed issues will severely hamper broadcast seedling and planting
efforts. Finally, the soil environment should be investigated and procedures to optimise
seedling growth and survival will also need to be researched if successful rehabilitation
standards are to be employed.

3
Figure 1.2. (clockwise from left) pit K, an active pit; pit Q, a pit that has partly
regenerated through natural migration; undisturbed vegetation; pit L, a pit spread with
topsoil.
1.1.2 Site description
Climate
The Shark Bay region has an arid climate with an average annual rainfall of 216 mm.
Most of this rain falls in the winter with occasional summer cyclonic activity (Fig. 1.3,
1.4). Winter is cooler than summer, with the winter exhibiting an average maximum
and minimum temperature of 21oC and 15
oC, respectively, and summer exhibiting an
average maximum and minimum temperature 33oC and 23
oC, respectively (Fig. 1.3a).
Rainfall from year to year is extremely variable (Fig. 1.3b, c) and over the last 24 years
has ranged from 59 to 392 mm (Fig. 1.4). The lowest rainfall on record was received in
2006.

4
a)
b)
c)
Figure 1.3. a) Monthly maximum and minimum air temperatures (averaged over 8
years) and rainfall (averaged over 25 years). Bars indicate standard error. b) Long term
variability: Box plot indicating the lowest and highest monthly values, the 10th
, 25th
,
75th
and 90th
percentiles and the median value over 25 years. c) Short term variability:
Jan Feb Mar April May June July Aug Sept Oct Nov Dec
Rain
fall
(mm
)
0
10
20
30
40
50
60
70
Tem
pera
ture
(oC
)
0
10
20
30
40
50
60
70
Rainfall
Min Temp
Max Temp
Jan Feb Mar April May June July Aug Sept Oct Nov Dec
Rain
fall
(mm
)
0
40
80
120
160
200
240
280
3202005
2006
2007
2008
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Rain
fall
(mm
)
0
40
80
120
160

5
Monthly rainfall across the four years of the study (for 2008, only rainfall from January
to March is shown).
1984
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
Rain
fall
(mm
)
0
100
200
300
400
500
Rainfall (mm)
Average rainfall
Figure 1.4. Annual rainfall from 1984 to 2007 at Shark Bay Salt (bars) and average
rainfall across this period (line).
Geology, geomorphology and soils
The Shark Bay region is within the Carnarvon sedimentary basin, which consists of a
lower sequence, the Silurian-Early Devonian Kalbarri Group, overlain by the Early
Cretaceous Winning Group (Hocking et al. 1987; Hocking 1990) (Fig. 1.5). These
rocks are not exposed, but the Birdrong Sandstone at the base of the Winning Group has
been intersected in the Useless Loop artesian bore (Wills and Dogramaci 2000). The
surface geology on the Edel Peninsula where SBS is situated is made up of Middle to
Late Pleistocene Tamala Limestone, which is a shelly eolian dune sand. Dissolution
and redeposition of the calcium carbonate has formed hard limestone within the dunes.
The overlying soils are mainly reddish-brown calcareous sands (DEP 2001; CALM
2005).

6
Figure 1.5. Geological cross section from Useless Inlet to Freycinet Reach through the
evaporation ponds. Original image developed by the authors, Philip Commander and
Craig Miskell from data from the Useless Loop town bore log.
Vegetation
The Edel peninsula is on the transition zone between the South West Botanical Province
and the Eremaean Province (DEP 2001). It is located in the northernmost extreme of
the Geraldton Sandplains (GS) IBRA bioregion (Fig. 1.6). As a transitional zone, the
area has high species diversity with 823 species of flowering plants. Fifty-six of these
species are at their southern limit and 229 are at their northern limit. There is also a
high level of endemism (53 species) with the flora providing a habitat for rare or
threatened fauna (DEP 2001).

7
Figure 1.6. Interim Biogeographic Regionalisation for Australia (Thackway and
Cresswell 1995, Environment Australia 2000) taken from Western Australian
Herbarium (1998–).
At SBS, four vegetation complexes have been described; shrublands, herblands,
halophytic and coastal (Mattiske 1996). Within the four complexes there are 17
vegetation associations (Appendix 3). The shrublands complex (associations 1-12) is
dominated by Acacia spp., Melaleuca spp., Atriplex spp. and Thryptomene baeckeacea.
The herblands (associations 13, 14) are present on the birridas (salt and claypans) and
are dominated by Muellerolimon salicorniaceum, Frankenia setosa, Olearia axillaris
subsp. obovata and Wilsonia humilis. The halophytic complex (association 15) is
dominated by Halosarcia species. Lastly, the coastal complex (association 16, 17) has
one association containing Spinifex longifolius, Acanthocarpus preissii and Nitraria
billardierei and the other association is dominated by Avicennia maritima (mangrove)
(Mattiske 1996). The vegetation at Shark Bay Salt is similar to nearby Dirk Hartog
Island as described by Burbidge and George (1978). These authors identified five
communities on the island, and the community that has the most similarities with the
shrublands complex is the ‗low closed/open-heath with hummock grasses‘. This
vegetation type contained Triodia plurinervata, Thryptomene baeckeacea, Melaleuca
cardiophylla and Acacia ligulata. As Shark Bay is in a transitional zone between the
South West Botanical Province and the Eremaean Province, some species found in the
area by Mattiske (1996) occur in both provinces, for example Acacia ligulata, Acacia
tetragonophylla, Alyogyne cuneiformis, Atriplex bunburyana, Diplolaena grandiflora
and Melaleuca cardiophylla (Burbidge and George 1978). Others only occur in the

8
South West Botanical Province and these include Acanthocarpus preissii, Halgania
littoralis, in particular Aphanopetalum clematideum is at its northern limit. However,
other species only occur in the Eremaean Province such as Brachycome latisquamea,
Thryptomene baeckeacea and Triodia plurinervata (Burbidge and George 1978).
Species distribution could have an effect on seed germination ecology.
It is unknown whether, following disturbance, the vegetation at SBS conforms to the
initial floristic model or the relay floristic model (Egler 1954). The initial floristic
model states that following disturbance, all species recruit together, and due to varying
life-spans, species gradually disappear from the ecosystem. In ecosystems that follow
the relay floristic model, groups of species replace each other over time. These models
can be used to predict recruitment requirements, for example, the jarrah forest in
southwest Western Australia follows the initial floristic model, so after disturbance
caused by mining, restoration practitioners aim to establish all species in the ecosystem
immediately (Koch and Ward 1994; Norman et al. 2006a).
Animals
The macro fauna in the area is dominated by feral animals including foxes, cats, rabbits
and house mice (Short and Turner 2000). These animals pose a problem for
rehabilitation efforts. These feral animals may compete with the native fauna for food,
change the vegetation structure by selective and over grazing, and disturb the soil
environment through their daily activities. In particular, the diet of rabbits on Heirisson
Prong includes Acacia ligulata, Acacia tetragonophylla, Alyogyne cuneiformis,
Eremophila spp. Melaleuca cardiophylla and Pimelea microcephala (Robley et al.
2001). Also, cats and foxes predating the native fauna is a particular problem
considering the high number of threatened or endangered native species (DEP, 2001).
1.2 Rehabilitation
The mining and extraction of minerals and energy are major activities in Western
Australia, and usually involve clearing vegetation and disturbing soil profiles (Gardner
2001). Clearing vegetation and soil disturbance not only reduce aesthetic values, but
also lead to erosion, loss of biodiversity and loss of habitats for native fauna (Clemente
et al. 2004). A thorough understanding of rehabilitation procedures and techniques is
essential for success in reinstating biodiversity. These procedures may include site and

9
soil preparation, and plant replacement through 1) topsoil removal, storage and
replacement, 2) seed broadcasting and 3) seedling planting (Gardner 2001; Burke 2003;
Rokich and Dixon 2007). Some procedures developed in other parts of Western
Australia and the world may be applied, whereas others may be specific to the site or
species due to the unique aspects of site, climate variables and the endemic nature of the
taxa. Indeed, Hobbs and Cramer (2007) indicate that restoration practices developed in
the Northern Hemisphere may not be applicable to other areas, including
Mediterranean-type areas such as Western Australia. The vegetation community
resulting from the rehabilitation must be sustainable, and thrive without constant
intervention.
Rehabilitation, particularly in arid areas, is challenging (Anderson and Ostler 2002) due
to a number of issues. For example, erosion problems may be exacerbated by high
winds or intense rainfall events. In addition, seed germination and seedling survival
may be limited by low and erratic rainfall (Glenn et al. 2001; Snyman 2003). Seedling
survival may also be limited by grazing pressure from feral animals such as rabbits.
Seedling growth may be limited by low soil nutrients (Milton 2001; Williamson and
Neilsen 2003) and soil compaction.
1.2.1 Site preparation
The soil environment of post-disturbed areas must undergo appropriate restoration to
enable successful revegetation (Bradshaw 1997). Seedling mortality in post-mined
areas may be related to the soil environment (Enright and Lamont 1992), soil
compaction in particular. Soil compaction can be a result of the use of heavy machinery
or natural processes (Rokich 1999). Soil compaction can restrict root penetration, and
limit water movement within the soil (Ashby 1997). A result of soil compaction is an
increase in soil bulk density which has been shown to decrease growth of Eucalyptus
seedlings (Williamson and Neilsen 2003). However, ripping the soil can alleviate
compaction (Rokich et al. 2001) and has been shown to increase saturated hydraulic
conductivity (therefore increasing the water infiltration capacity), reduce bulk density
(Luce 1997) and reduce soil impedance (Rokich et al. 2001). Ripping can have a
variable effect on plant growth. For example, Ashby (1997) found trees planted in soil
that was ripped to a depth of 1.2m were taller than those planted in un-ripped areas after
5 years. In addition, Yates et al. (2000) found four out of five species planted in ripped
sites had longer shoot lengths compared with those in un-ripped sites. However, a study

10
in California found little effect of ripping on plant establishment two years after sowing
(Montalvo et al. 2002). In another study (Rokich et al. 2001), ripping did not have an
effect on shoot or root weights of most species, and reduced root and shoot weights of
one species. Although root weights were not affected, root architecture of tap and
lateral roots differed between ripped and un-ripped sites. In five out of the eight species
tested, tap roots were longer and there were fewer lateral roots in ripped compared with
un-ripped sites (Rokich et al. 2001). Clearly, simple practices adopted in rehabilitation
programs elsewhere may require detailed research to establish procedures and benefits
at this site.
1.2.2 Plant replacement
Topsoil
Topsoil is generally considered to be the most important source of seeds for
rehabilitation purposes (Koch et al. 1996; Rokich et al. 2000), and a source of micro-
organisms, nutrients and organic matter (Ward et al. 1996; Gardner 2001; Williamson
and Neilsen 2003) when handled correctly. Seedling recruitment from respread topsoil
can be affected by several factors. These factors include the depth to which topsoil is
stripped and spread, the time of stripping and spreading and whether material is
stockpiled or direct-returned (i.e. direct reinstatement not stockpiled) (Koch et al. 1996;
Rokich et al. 2000).
Depth
The depth at which topsoil is stripped has an effect on seedling emergence due to the
distribution of seed in the soil. In general, as depth from the surface increases, the
amount of seeds in the soil seed bank decreases. For instance, in one study (Tacey and
Glossop 1980) in the jarrah forest, topsoil was sampled from depth intervals of 0-1, 1-2,
2-5 and 5-10 cm and seedling emergence was found to be greatest from the 0-1 and 1-2
cm fractions. Another jarrah forest study (Koch et al. 1996) found greater soil seed
content in the 0-2 and 2-5 cm fractions compared with 5-10 cm. Consequently,
stripping to a greater depth simply dilutes the topsoil with seedless soil as found for
Banksia woodland, where stripping to 10 cm resulted in 254 seedlings per 5 m2
compared with 81 seedlings per 5 m2 after stripping to 30 cm (Rokich et al. 2000).
The depth to which topsoil is re-spread also affects seedling emergence. For example,
the optimal burial depth for most of the twelve species used in one study on jarrah forest

11
rehabilitation was between 0 and 2 cm (Grant et al. 1996). Only three species emerged
from 15 cm, but emergence percentages at this depth were suboptimal. This implies
that if topsoil is spread too thickly, then seeds will become buried and will be unable to
emerge. This finding is confirmed by Rokich et al. (2000) who found that topsoil
stripped at 10 cm and spread to 10 and 30 cm produced similar seedling recruitment,
with an extra 20 cm of topsoil containing seed failing to increase seedling numbers as it
is likely that many seeds are unable to emerge from the greater depth.
Timing of topsoil removal
The timing of stripping and spreading topsoil has an effect on seedling recruitment.
Some authors recommend that topsoil should be stripped, spread and ripped when it is
still dry (Ward et al. 1996; Koch et al. 1997). Handling wet soil may be detrimental as
seeds may have commenced germination and may be damaged or lose viability (Rokich
et al. 2000). Also, if soil is spread in the winter, valuable time for germination and
establishment may have been lost. For example, in Banksia woodland with wet winters
and dry summers, stripping and direct return of topsoil in autumn resulted in higher
seedling recruitment (73.3 seedlings per 5m2) compared with stripping and spreading in
winter (5 seedlings per 5m2) (Rokich et al. 2000).
Stockpiling
Stockpiling topsoil can adversely affect seed density, seedling recruitment and species
richness. For example, in one study, clearing, stockpiling and respreading topsoil
decreased seed density to 13.4% of the original density compared with a decrease to
31% of control levels following clearing and direct return topsoil (Koch et al. 1996). In
addition, several studies (Tacey and Glossop 1980; Ward et al. 1996; Rokich et al.
2000) found that species richness was lower from soil that was stockpiled than direct-
return topsoil. For instance, Rokich et al. (2000) found no seedling recruitment after
stockpiling for three months compared with 73.3 seedlings from 18 species per 5m2
after direct return in the autumn. Similar benefits were reported by Ward et al. (1996)
who found direct return topsoil had 94 species and four times greater density compared
with respread stockpiled topsoil, which returned 75-81 species.
Seed broadcasting
While topsoil return is an important practice in post-mine restoration, seed broadcasting
is necessary to enable replacement of species that do not store their seed in the soil, but

12
instead store seeds in their canopy (Rokich et al. 2002), or when topsoil is unavailable.
The advantages of seed broadcasting compared with planting seedlings may include the
lower cost and minimal labour inputs (Turner et al. 2005b). Also, broadcasting seed
over topsoil can increase both seedling recruitment and species richness (Rokich et al.
2002). Many factors affect the success of seed broadcasting including the timing of
sowing (Turner et al. 2006c), seed burial (Rokich and Dixon 2007), seed predation (Ord
2007), wind activity (Ord 2007) and soil moisture. In addition, it is essential to
overcome seed dormancy prior to seed broadcasting (Norman et al. 2006b) as unknown
germination requirements and dormancy mechanisms can limit the success of seed
broadcasting (see sections 1.3 and 1.4).
Incorporating broadcast seeds under the surface of the soil by raking may increase
seedling emergence. Seed burial through raking may increase emergence through
reducing predation or wind or water erosion (Rokich and Dixon 2007). For example,
raking soil after seed broadcasting increased seedling emergence from 1.2% to 11.8% at
a post-sand extraction site in Banksia woodland (Turner et al. 2006c). Also, scarifying
soil increased establishment of eucalypts and legumes (but not understory species), and
this was attributed to increased seed-soil contact (Ward et al. 1996). In contrast,
disturbance (using a four tyne scarifier) similar to raking did not have any effect on the
establishment of Themeda triandra in Eucalyptus woodland (Cole et al. 2005).
The timing of seed broadcasting can have an effect on seedling emergence as shown by
two studies in southwest Western Australia. Firstly, at a restoration site in Banksia
woodland, seedling emergence of four out of the nine species utilised was greater after
sowing in May compared with July (Turner et al. 2006c). Secondly, in a post-mined
site in the Jarrah forest, broadcasting time affected legume and Eucalyptus species
(Ward et al. 1996). Out of plots sown in December, February, April and June, the
highest number of legumes was present on the April plots, and the highest number of
Eucalyptus species on the April and June plots.
Seed germination can be slow and erratic with respect to emergence time and seedling
vigour (Heydecker et al. 1973; Karssen et al. 1989). One treatment that can result in
earlier germination, improve the germination rate, increase uniformity of germination
and increase germination percentages under optimal and sub optimal conditions is
priming (Heydecker et al. 1973; Heydecker and Coolbear 1977; Karssen et al. 1989).

13
Priming involves soaking seeds in water (hydropriming) or an osmoticum such as
sodium chloride or polyethylene glycol (osmopriming) and then drying them prior to
incubation. During priming, the seed imbibes and begins metabolism, but it is dried just
prior to radical emergence. Priming duration and temperature affect germination
(Hardegree 1996). Equally, the availability of water may be a key factor in seed
germination and seedling establishment for sites where rainfall amounts are variable and
timing is unpredictable. Irrigation may therefore enhance germination. For example, at
a minesite in New South Wales, irrigation of plots seeded with Themeda australis
increased seedling density (Windsor and Clements 2001), however, irrigation of direct
seeded plots in Arizona did not increase plant cover (Glenn et al. 2001). Irrigation may
be expensive, and water availability limited, hence this may not be practical in some
environments.
Plant treatments
Planting seedlings raised in a nursery is another method of revegetation. This method
may be more reliable than broadcast seeding, but more costly (Barrett-Lennard et al.
1991; Glenn et al. 2001; Anderson and Ostler 2002; Barrett-Lennard et al. 2003). Also,
for species that perform poorly when direct seeded (due to insufficient seed, unknown
dormancy mechanisms and/or low seed viability – see sections 1.3 and 1.4) propagation
from cuttings, seed or tissue culture then planting as seedlings, may be the only method
of replacing these species to site (Gardner 2001). As factors such as soil nutrients, soil
moisture and root development affect plant survival, various treatments can be
employed to enhance the success of these seedlings. These treatments include the
application of fertiliser (Williamson and Neilsen 2003) and water holding polymer gels
(Woodhouse and Johnson 1991) and the use of pots that encourage good root
architecture (Burdett et al. 1983).
Disturbed soil may be low in nutrients due to vegetation and topsoil removal. If
nutrients are limiting, fertiliser applied at planting may assist plant establishment (Close
et al. 2005b). For example, fertiliser is used routinely in rehabilitation areas in the
southwest Western Australia, at a rate of 500 kg ha-1
(Gardner 2001). One study has
postulated that there may be a lower nutrient supply in subsurface soil due to the finding
that organic matter decreased with increasing soil depth (Williamson and Neilsen 2003).
As a result, Eucalyptus seedlings grown on subsurface soil (10-30 cm) had lower above-
ground dry weight compared with seedlings grown on topsoil (0-10 cm). Addition of a

14
combination of nitrogen and phosphorus fertiliser increased the above ground dry
weight of seedlings grown on the subsurface soil (Williamson and Neilsen 2003). Also,
organic fertiliser supplements have been shown to increase percentage plant cover
compared to control plots at roadcut slopes in Mesa Verde National Park, an arid region
of Colorado (Paschke et al. 2000). However, a study at a limestone quarry in Portugal
showed fertiliser response to be species specific with the use of a slow release NPK
fertiliser increasing shoot length and stem diameter in Ceratonia siliqua, but not Olea
europaea or Pistacia lentiscus (Clemente et al. 2004). Also, a study on Eucalyptus
grandis in South Africa found the effect of fertiliser to be site specific based on the
mineralizable-N status of the study soils (Herbert 1990).
While fertiliser applied in the nursery can benefit growth after planting, the regime of
application also has the potential to make significant differences. Conventional
fertiliser regimes in nursery production apply constant amounts of fertiliser from when
the seed germinates, to when the seedling leaves the nursery. However, growth is not
always constant. Plant growth follows a sigmoidal growth curve, that is, there is a lag
phase, followed by a period of exponential growth (Fig. 1.7), then a steady state.
Exponential growth rates imply that plants may benefit from exponential additions of
fertiliser, rather than constant additions. Constant fertiliser additions may over-supply
fertiliser when the seedling is young, and under-supply later in plant growth (Timmer
1997). Nutrient loading uses high inputs of fertiliser to increase the consumption of
nutrients from a level that is sufficient consumption to luxury consumption. Nutrient
loading can be used to increase nutrient concentrations in the plant, but not change plant
dry mass. Although nutrient loaded seedlings have been shown to be similar sized at
planting compared with those conventionally fertilised, loaded seedlings have higher
tissue nutrient content, and are 44% taller, and achieve 37% more biomass (Timmer
1997). Exponential nutrient loading aims to deliver a stable internal concentration of
nutrients during this phase of exponential growth (Timmer 1997). For example, under
an exponentially increasing nutrient application regime applied in the nursery, total
biomass of Eucalyptus globulus seedlings was 1.5 to 2-fold higher, compared with
conventional fertiliser application, five months after planting (Close et al. 2005a).
Lack of water can also be detrimental to planted seedlings as it can cause drought stress
(Close et al. 2005b). Irrigation and polymer gels (also called hydrogel, which are
polymers that expand to form a gel, and hold water which plant roots can access)

15
(Woodhouse and Johnson 1991) represent useful but expensive options under broadacre
applications. Superabsorbent polymers have been shown to be effective when used with
potting media with increases in dry weight and water use efficiency of lettuce and
barley (Woodhouse and Johnson 1991) and prolonged survival and increased growth of
Pinus halepensis seedlings under drought conditions (Hüttermann et al. 1999).
However, polymer gels did not improve plant establishment in a restoration study in a
semi-arid region of the US, but in that study, the polymer was trialed only via
incorporation into field soils prior to direct seeding rather than in the growing medium
under nursery conditions (Paschke et al. 2000). Polymer gel also did not affect shoot
growth of three shrubs grown in a Mediterranean quarry restoration site (Clemente et al.
2004).
Figure 1.7. Dry matter production and nitrogen (N) uptake of mesquite seedling shoots
at single dose fertilization regimes during the growing season (from Imo and Timmer
(1992) as cited in Timmer (1997)).
Root morphology can affect the stability of the plant, root growth (Burdett et al. 1983)
and survival (Rokich et al. 2001). Root morphology is not only affected by
environmental factors such as soil compaction (see section 1.2.1) (Rokich et al. 2001)
but also seedling production methods (Burdett et al. 1983). Root pruning is a method
used to improve root morphology of seedlings prior to transplanting. Without root
pruning, lateral roots have been shown to grow down the side of the container wall, and
when planted, they continue growing downwards instead of horizontally. Chemical root
pruning (dipping pots in paint mixed with cupric carbonate powder) has been shown to
prevent lateral roots from growing down the sides of the container wall (Burdett et al.
1983). When planted, these roots continue to grow horizontally, hence stabilising the
tree and accessing moisture and nutrients close to the soil surface with root pruned

16
seedlings exhibiting greater shoot growth four years after planting (Burdett et al. 1983).
In addition to chemical root pruning, another method of root pruning is to make slits in
the sides of pots (Zahreddine et al. 2004), and these pots are sometimes termed ‗air
pruned pots‘.
1.3 Seed germination
1.3.1 Seed morphology and viability
A seed is often the most important resource to ensure the survival and continuation of a
species. In restoration, seeds are either used for seed broadcasting or for the
development of seedlings for planting. Seeds contain an embryo which is often
surrounded by a seed coat or testa, and may contain endosperm or perisperm to supply
nutrients to the embryo. The embryo consists of the embryonic axis and one
(monocotyledon), two (dicotyledon) or more cotyledons (Bewley and Black 1994).
Embryos can be a variety of different sizes and shapes and occupy different positions
within the seed (Martin 1946). Understanding embryo morphology can assist with
dormancy classification.
Martin (1946) developed an embryo classification system (Fig. 1.8). He described three
main types (divisions) of embryo position: basal, peripheral and axile, and these
divisions are further classified into 12 types. Baskin and Baskin (2007) have recently
revised Martin‘s classification system and removed the dwarf classification from the 12
types (Fig. 1.8). They have renamed micro as undifferentiated. In addition, they have
amended Martin‘s linear and spatulate types to be named fully developed linear and
fully developed spatulate (Fig 1.8). Baskin and Baskin (2007) have added two new
types, underdeveloped linear and underdeveloped spatulate. The fully developed types
have an E:S (embryo to seed) ratio >0.5 whereas the underdeveloped types have an E:S
ratio ≤0.5.
Not all seeds contain a live embryo. Those seeds that contain a live embryo are termed
viable and have the potential to germinate, while those seeds that do not contain a live
embryo and therefore cannot germinate, are termed non-viable. There are several
methods to test seed viability. These include tetrazolium chloride, excised embryo,
float and cut tests (Moore 1972; Paynter and Dixon 1990; Sweedman and Merritt 2006;
ISTA 2007). In addition, seed fill can be determined by x-ray analysis (Goodman et al.

17
2006). The principle of the tetrazolium test is that the seed will imbibe the colourless
2,3,5-tryphenyl tetrazolium chloride, and living tissues within the seed will react to
produce tryphenyl-formazan – a red substance. The non-living parts of the seed do not
stain red, and hence remain colourless. Seeds may also partially stain, and these are
classified as either viable or non-viable depending on the position of the stained areas
(ISTA 2007). The excised embryo test involves excising the embryo from the seed and
incubating it on growth media. Embryos that grow are considered viable (Sweedman
and Merritt 2006). This test may not be applicable to all species, as viable embryos
with certain types of dormancy will not grow until their dormancy is broken. For a float
test, seeds are soaked in water, and those that float are considered non-viable, whereas
those that sink are viable (Paynter and Dixon 1990). When these three tests were
performed on Geleznowia verrucosa the results were similar, indicating that they are all
good indicators of viability (Paynter and Dixon 1990). The final viability test is a cut
test. To perform a cut test, the seed coat is removed and the embryo is examined
(Sweedman and Merritt 2006). Viable embryos are generally white and turgid. Non-
viable embryos may be brown or black and possibly withered. Some seeds do not
contain an embryo at all. As well as a cut test, seed fill (determining whether seeds
contain an embryo) can be identified using x-ray analysis. X-ray intensity and exposure
time can be specified during analysis. Digital imaging technology produces an image of
the x-rayed seeds. On the digital image, seed tissue is white, while empty space is
black. Using this method, unfilled seeds can be distinguished quickly and easily from
filled seeds. X-ray analysis is efficient because it is less time consuming than cut tests,
and is non-destructive, so that seeds can be returned for use in other purposes such as
germination testing (Goodman et al. 2006).
1.3.2 Seed germination
Seed germination ‗begins with water uptake by the seed and ends with the start of
elongation by the embryonic axis, usually the radicle‘ (Bewley and Black 1994) (Fig.
1.9). Germination is usually considered complete when the radicle protrudes through its
covering structures. Three environmental factors are the minimum necessary
requirements for seed germination – water, oxygen and incubation at an appropriate
temperature (Bewley and Black 1994). In addition to these, other environmental factors
such as light quality or quantity, smoke, nitrates and a variety of other chemicals can
stimulate germination (Vleeshouwers et al. 1995). The requirement for these stimulants
singly or in combination is very species dependent.

18
Figure 1.8. Seed types based on embryo size and position (modified from Martin,
1946). Martin‘s seed types are hand written and modifications to these seed types by
Baskin and Baskin (2007) are typed in brackets. Note that linear underdeveloped and
spatulate underdeveloped have been added and dwarf has been removed.

19
Water
Water uptake is the first step towards germination (Bewley and Black 1994) and it is
triphasic (Fig. 1.9). The first phase (imbibition) is due to the lower water potential of
the seed compared with the water surrounding it. Imbibition occurs in seeds that are
non-dormant, dormant (except physical dormancy, see section 1.4.3), non-viable and
viable. During the second phase (lag phase), very little water is taken up by the seed,
but seeds are metabolising. The third phase is simultaneous with radical elongation, and
hence is only achieved by non-dormant seeds. Germination is completed when the
radicle protrudes from the covering tissues.
Figure 1.9. Time course of major events associated with germination and subsequent
postgerminative growth (from Bewley (1997)).
Oxygen
Oxygen is necessary for respiration. Respiration rates are low in seeds that have not
commenced imbibition, but as soon as seeds imbibe, oxygen consumption increases.
This increase is followed by a lag phase. The lag phase ceases as soon as the radicle
emerges, which coincides with another increase in oxygen consumption. The embryo
continues to consume oxygen as it grows, and eventually there is a decrease in oxygen
consumption in the seed‘s storage tissues as they are expended and senesce (Bewley and
Black 1994).

20
Temperature
Seeds can sense temperature in a wide variety of ways – both in the dry or imbibed
state. Seed germination occurs over a range of temperatures (sometimes wide,
sometimes narrow) and this temperature usually corresponds with the season of water
availability in the natural environment (Bell et al. 1993; Bell 1999). For example,
germination of 18 out of 43 species from the southwest of Western Australia tested by
Bell et al. (1995) was affected by temperature, and all but one of those had higher
germination at 15oC compared with 23
oC. Temperature at the beginning of winter is
around 15oC, and winter is the period of reliable rainfall in the southwest of Western
Australia. In addition, germination of eight taxa of Asteraceae from arid inland
Australia was optimal from 10 to 20oC, indicating winter germination, although
germination of one species, Rhodanthe humboldtiana was optimal from 10 to 30oC,
indicating germination whenever water is available, hence possibly has aseasonal
germination (Plummer and Bell 1995). Similarly, two Eucalyptus species from the
southwest of Western Australia exhibited different temperature responses. Eucalyptus
wandoo displayed highest germination in the range 15-20oC, whereas E. rudis exhibited
highest germination at 20-30oC. Eucalyptus rudis (flooded gum) occurs in wetter areas
along streams, and hence moisture is available over a wider range of temperatures (Bell
and Bellairs 1992). In contrast to the southwest of Western Australia where there is one
season of water availability, in arid Australia, rain falls in both summer and winter
events. Mott (1972) observed that monocotyledons tended to germinate at the
temperature range of 25-30oC which corresponded to summer, and dicotyledons at 15 -
20oC, corresponding to a winter growth cycle. Shark Bay is also located in the arid
zone. Most of the rain falls in the winter, but the area has occasional summer cyclonic
activity. This rainfall pattern has a major impact on seed germination, as water is
potentially available both in the summer and winter, hence, seeds may be able to
germinate at summer and/or winter temperatures. Also, rainfall in the arid zone can be
sporadic and unpredictable and this may also have an effect on germination strategies.
Germination stimulants
The distinction between germination stimulants and dormancy breaking agents is
inconsistent. For instance, Finch-Savage and Leubner-Metzger (2006) consider that
light and smoke are able to release dormancy. Conversely, this author and others
consider light and smoke products (aerosol smoke, smoke water and karrikinolide) to be

21
germination stimulants (Merritt et al. 2007; Ooi 2007; Rokich and Dixon 2007) rather
than dormancy-releasing agents. For further explanation see section 1.4.1.
Seeds of some species require light for germination, whereas light inhibits germination
of other species. For example, Eucalyptus marginata, Eucalyptus calophylla, Acacia
drummondii subsp. candolleana and Kennedia prostrata exhibited lower percentage
germination when incubated under white light compared with darkness (Rokich and
Bell 1995). In contrast, germination of several Australian Asteraceae species is
promoted by light (Plummer and Bell 1995; Plummer et al. 2001; Schütz et al. 2002;
Merritt et al. 2006). In particular, seven of the 10 taxa of Asteraceae from arid inland
Australia tested by Plummer and Bell (1995) showed higher germination percentages
when incubated in the light. This light requirement was replaced by exogenously
applied gibberellic acid (GA3) in six of those species. A light requirement may indicate
that seeds need to be on, or close to the soil surface for germination (Fenner and
Thompson 2005). Seeds may also be able to detect light quality such as where light is
filtered through leaves in a canopy, as the canopy absorbs red wavelengths, therefore
altering the red:far-red ratio compared with direct sunlight. For example, a number of
species possess seeds that require sunlight for germination, and where germination is
inhibited by light filtered through a canopy (Fenner and Thompson 2005).
Smoke is a major and universal stimulant of germination for many species, particularly
Australian plants (Dixon et al. 1995; Roche et al. 1997b; Rokich and Dixon 2007). In
one study, 48% of Australian species tested responded to smoke water (Dixon et al.
1995), and in another, 69% responded (Roche et al. 1997a). Smoke can be applied in
aerosol form or dissolved in water, (Dixon et al. 1995; Roche et al. 1997a; Roche et al.
1997b) to produce smoke water (SW). Although SW may be more convenient for
broad-scale use, aerosol smoke has been found to be more effective at enhancing
seedling recruitment in Banksia woodland restoration (Rokich et al. 2002) and
Eucalyptus marginata forest restoration (Roche et al. 1997b). The concentration of SW
has an effect on germination and some studies have proposed that SW at high
concentrations may have an inhibitory effect on germination (Roche et al. 1997a;
Flematti et al. 2004; Baker et al. 2005a). For example, concentrations of 0.1% and 30%
SW increased germination of Actinotus leucocephalus above the control, but
germination was optimal when seeds were treated with concentrations of between 1%
and 20% (Baker et al. 2005a). In addition, undiluted SW decreased germination of

22
lettuce seeds (Lactuca sativa L. cv. Grand Rapids) compared with the control, whereas a
1 in 10 dilution elicited a two-fold increase in germination (Flematti et al. 2004).
Smoke response can have an interaction with the age of seeds (Roche et al. 1997a), the
season of application (Roche et al. 1998), exposure to high temperatures (Tieu et al.
2001a), soil burial (Tieu et al. 2001b; Baker et al. 2005c) and warm stratification
(Merritt et al. 2007). These interactions are often species specific and may depend on
the dormancy status of the seed.
Karrikinolide (KAR1 (3-methyl-2H-furo[2,3-c]pyran-2-one), formerly known as
butenolide) (Flematti et al. 2004; Dixon et al. 2008) is a recently discovered chemical
isolated from smoke and shown to be the key agent responsible for germination of fire
following species. Due to its recent discovery, there are few studies on the stimulatory
effect of the KAR1 alone on Australian species. KAR1 stimulates germination of lettuce
seeds (Lactuca sativa L. cv. Grand Rapids) incubated in the dark and the Australian
species, Conostylis aculeata, Stylidium affine (Flematti et al. 2004), Angianthus
tomentosus and Myriocephalus guerinae, in both light and darkness, and Podolepis
canescens in the dark (Merritt et al. 2006). Kulkarni et al. (2007) and Daws et al.
(2007) have shown similar and often pronounced responses of seed for a variety of
native and weed species.
1.4 Seed dormancy
1.4.1 Definition
Many authors define dormancy as ‗the failure, or block of a viable seed to complete
germination under normally favourable conditions‘ (Bewley 1997; Koornneef et al.
2002; Kucera et al. 2005; Hilhorst et al. 2006). However, other authors do not consider
dormancy to be an absence of germination, leading to the definition of ‗dormancy is a
seed characteristic, the degree of which defines what conditions should be met to make
the seed germinate‘ and ‗dormancy relieving factors cause a widening of the range of
conditions that allow germination‘ (Vleeshouwers et al. 1995). Nevertheless, even
those using this definition have differing interpretations. For instance, Vleeshouwers et
al. (1995) consider that temperature is the only environmental factor that breaks
dormancy. Therefore, light does not break dormancy, but stimulates germination as it
fails to widen the range of conditions allowing germination. On the other hand, Finch-
Savage and Leubner-Metzger (2006) agree with the above definition of dormancy, but

23
interpret it to mean that light breaks dormancy so that the seed is able to germinate in
darkness, therefore widening the range of conditions allowing germination. Yet another
opinion is that temperature may alter dormancy and light sensitivity, but light indicates
the suitability of conditions to germination. Hence, this review and others (Baskin and
Baskin 2004c; Fenner and Thompson 2005) consider that light, along with nitrates and
smoke (Merritt et al. 2007; Ooi 2007; Rokich and Dixon 2007), are environmental
factors that may be pre-requisites of non-dormant seeds to germinate. However,
temperature is not the only environmental condition that affects dormancy. Although
Vleeshouwers et al. (1995) state that soil moisture does not affect dormancy, the
hydration status of the soil (or relative humidity under laboratory conditions) affects the
water content of the seed, and this, combined with temperature, affects dormancy
(Baskin and Baskin 1998).
A new definition of seed dormancy is that ‗a dormant seed does not have the capacity to
germinate in a specified period of time under any combination of normal physical
environmental factors (temperature, light/dark etc.) that is otherwise favourable for its
germination‘ (Baskin and Baskin 2004c). This definition indicates that light is an
environmental factor that may be necessary for germination. Also, this definition
specifies a period of time, after which non-germinating seeds may be considered
dormant. This is useful for experimental purposes as seeds may germinate over a long
period of time under test conditions. For these reasons, I have chosen to use this
definition in my study.
1.4.2 Classification
An early classification of dormancy by Harper (1957) cited in Mott and Groves (1981)
and Vleeshouwers (1995) classifies dormancy into three states. The first state, innate
dormancy, refers to seeds that are dormant at dispersal as a result of immature embryo
or seed-based germination inhibitors. The second state, enforced dormancy, is due to an
external environmental factor, and dormancy is present until the factor is removed.
Induced dormancy is the third state and is caused by an external factor. It is developed
after seeds are dispersed – however, dormancy is still present when this factor is
removed. This classification is similar to that of Nikolaeva (2001) who classifies
dormancy as either organic, which is a seed property (like innate and induced), or
imposed, which is due to environmental factors (like enforced). Due to the current
definition of dormancy adopted here, lack of favourable environmental conditions

24
(imposed dormancy) is no longer considered dormancy. Nikolaeva, classified organic
dormancy into three groups each with sub groups – exogenous (physical, chemical and
mechanical), endogenous (morphological, physiological and a combination of the two)
and combined (a combination of exogenous and endogenous dormancy).
Nikolaeva‘s classification has been refined by Baskin and Baskin (1998; 2004c) to
recognise five classes of dormancy. These classes are physical, physiological,
combinational, morphological and morphophysiological, and each class may contain
levels and types. To classify seeds into these classes, Baskin and Baskin (1998; 2004c)
have created a dichotomous key based on seed characteristics of water permeability of
the seed coat and embryo morphology (Table 1.1). A description of the five classes is
as follows.
1. Seeds that are impermeable to water exhibit physical dormancy.
2. Physiological dormancy occurs when the fully developed embryo lacks the
growth potential to push through the endosperm and/or seed coat.
3. Seeds with combinational dormancy have both physical and physiological
dormancy.
4. Morphologically dormant seeds have undifferentiated and/or underdeveloped
embryos that must grow before the seeds germinate.
5. Morphophysiologically dormant seeds exhibit both morphological and
physiological dormancy.
Many scientists, particularly those investigating the model species Arabidopsis thaliana,
Avena fatua, Nicotiana sp. and Solanum sp., do not use the classification system
proposed by Baskin and Baskin (1998; 2004c), but instead either do not classify
dormancy, or classify dormancy as coat or embryo-based dormancy (Hilhorst and
Karssen 1992; Bewley and Black 1994; Fennimore and Foley 1998; Kermode 2005).
These model species all exhibit non-deep physiological dormancy (Baskin and Baskin
2004c). In addition, it has been postulated that the mechanism of non-deep
physiological dormancy has a coat and/or embryo component (Finch-Savage and
Leubner-Metzger 2006). Hence, coat and embryo dormancy are not classifications per
se, but are used to separate the two mechanisms. Although separating coat and embryo
components of dormancy may be useful when studying the mechanism of non-deep
physiological dormancy, the Baskin and Baskin (2004c) classification describes a
greater diversity of species and provides more direction as to how to release dormancy,

25
and hence is more useful for ecological studies. The wide application to ecological
studies is the reason why this study will be using the Baskin and Baskin (2004c)
classification system.
Table 1.1. Dichotomous key to dormancy types (reproduced from Baskin and Baskin
(2003b))
1. Seed or fruit coat not permeable to water………………... 2
2. Germination occurs within 2 weeks when seed or fruit
coat is scarified…………………………………...…..
Physical
2. Germination does not occur within 2 weeks when seed
or fruit coat is scarified………………………………
Combination of Physical
and Physiological
1. Seed or fruit coat permeable to water……………………. 3
3. Embryo either un-differentiated, or underdeveloped
(small)………………………………………………..
4
4. Embryo not differentiated……………………………. Specialised type of
Morphological
4. Embryo differentiated but underdeveloped…………... 5
5. Seeds germinate within 30 days at habitat
temperatures……………………………………
Morphological
5. Seeds do not germinate within 30 days at habitat
temperatures…………………………………....
Morphophysiological
3. Embryo differentiated and fully developed
(elongated)………….…………………………………
6
6. Seeds do not germinate within 30 days at habitat
temperatures…………………………………………
Physiological
6. Seeds germinate within 30 days at habitat
temperatures…………………………………………
Non-dormant
Another classification often used is the distinction between primary and secondary
dormancy (Hilhorst and Karssen 1992; Bewley and Black 1994; Vleeshouwers et al.
1995). Primary dormancy (similar to innate dormancy) is exhibited by freshly matured
seeds. Secondary dormancy (similar to induced dormancy) develops in non-dormant
seeds. However, Baskin and Baskin (1998; 2004c) do not consider that dormancy can
develop in seeds that are non-dormant at maturity and do not recognise primary and
secondary dormancy as classes of dormancy. They do, however, recognise that seeds
exhibiting non-deep physiological dormancy may cycle in and out of dormancy in
response to temperature. That is, seeds with non-deep physiological dormancy exhibit
primary dormancy at dispersal, then may lose this dormancy, and if they become
dormant again, this is termed secondary dormancy. The state between primary
dormancy and non-dormancy, and between non-dormancy and secondary dormancy is
termed conditional dormancy. This study will be mainly testing freshly dispersed seeds
(where available), hence if they are dormant, they will be in a primary dormant state.

26
1.4.3 Overcoming dormancy
Physical dormancy
Species with physical dormancy are known to occur in 16 families including the
Fabaceae, Surianaceae and Malvaceae (Baskin et al. 2000; Baskin et al. 2006b). As
physical dormancy is caused by a seed/fruit coat that is impermeable to water, any
treatment that increases permeability should break dormancy. These treatments can
include hot (or boiling) water, mechanical scarification, acid scarification and exposure
to high temperatures. For example, physical dormancy of six Australian Rhamnaceae
species was alleviated by hot water treatment (88-92oC for 5 min) (Turner et al. 2005a).
Soaking in boiling water also increased germination of Senna marilandica and S.
obtusifolia (Baskin et al. 1998) however, there was an optimal boiling time which was
different for the two species. Either side of this optimal time, germination was lower.
Other treatments that increased germination of Senna marilandica and S. obtusifolia
were mechanical or acid scarification and soaking in ethanol (Baskin et al. 1998).
However, exposure to high temperatures (50-140oC for 1-60 min) did not break
dormancy in Senna marilandica, and had limited success in Senna obtusifolia. In
contrast, high temperatures have been used to break physical dormancy of pasture
legumes (Quinlivan 1961; Quinlivan 1966). Exposing legume seeds to constant
temperature of 140oF (60
oC) for 5 months increased the permeability of the seed coats
to water and permeability was further increased by fluctuating temperatures of 60-140oF
(15-60oC) (Quinlivan 1961).
Physiological dormancy
Embryos within physiologically dormant seeds lack the growth potential to grow
through the covering tissue (Baskin and Baskin 2004c). In the laboratory, treatments
which remove or disrupt the covering tissue around the embryo tip (radicle) may
improve germination of physiological dormant seeds. Scarifying (nicking) the seed coat
or excising the seed or embryo from its covering structures are two such treatments
(Baskin and Baskin 2004c).
In the natural environment, seed dormancy is affected by temperature and seed water
content (dictated by the moisture content of the soil). Dormancy breaking treatments
therefore may be determined from the lifecycle of the seed, that is, the season of seed
maturation and germination, and the environmental conditions (temperature and
moisture) experienced between these states (Baskin and Baskin 2004b). For example,

27
seeds that mature in spring, and are exposed to summer temperatures before
germinating in autumn, may require a period of warm and wet conditions (warm
stratification) or warm and dry conditions (dry after-ripening) to overcome dormancy
(Baskin et al. 1998; Baskin and Baskin 2004b; Baskin and Baskin 2004c). Seeds that
mature in autumn and germinate in spring may need cold stratification to mimic winter.
Some seeds that disperse in spring may not germinate in the first autumn, but in the
following autumn, hence would need a period of warm followed by cold stratification.
In Shark Bay, seed maturation of most species is in late spring, and although
germination may occur with sporadic summer rainfall, it is likely to occur with the more
reliable winter rainfall. Hence, warm, dry conditions (dry after-ripening) like those
experienced during summer may overcome dormancy. After-ripening has been shown
to overcome dormancy of several Australian species, in particular several species from
the arid zone (Mott and Groves 1981; Jurado and Westoby 1992; Bell 1999; Plummer et
al. 2001; Tieu et al. 2001a; Schütz et al. 2002; Turner et al. 2006a; Merritt et al. 2007;
Hoyle et al. 2008). After-ripening can be affected by temperature, seed water content
and time. Dormancy loss by after-ripening occurs faster as temperatures increase, and
dormancy loss increases with time (Steadman et al. 2003). However, seeds after-ripen
at intermediate water contents, as they do not after-ripen at low water contents and lose
viability at high water contents (Baskin and Baskin 1979).
Combinational dormancy
Combinational dormancy is a combination of both physical and physiological dormancy
(Baskin and Baskin 1998). The water-impermeable seed coat prevents imbibition, so
both a treatment to increase permeability as for physical dormancy, and a dormancy
breaking treatment as for physiological dormancy are necessary for germination. For
example, germination of Diplopeltis huegelii which exhibits combinational dormancy
was increased by soaking in hot water, and further increased by storage at 23oC for six
weeks or storage at room temperature (~22oC) for 13 months (Turner et al. 2006a).
Morphological and Morphophysiological dormancy
Morphologically dormant seeds have a small (underdeveloped) but differentiated
embryo – that is, the radicle and cotyledon(s) are recognisable (Baskin and Baskin
1998). Embryos need to grow within the seed before they are able to germinate. In
addition, seeds with morphophysiological dormancy have underdeveloped but

28
differentiated embryos, and physiological dormancy. Embryos must grow within the
seed, and need a dormancy breaking treatment such as cold or warm stratification as for
physiological dormancy (Baskin and Baskin 2004c). For example,
morphophysiological dormancy of Actinotus leucocephalus can be overcome by after-
ripening followed by wet/dry cycles at 37oC (Baker et al. 2005b).
1.4.4 Mechanism of non-deep physiological dormancy
Non-deep physiological dormancy is the most common class of dormancy worldwide
and is the type of dormancy exhibited by many model species used to understand
dormancy mechanisms (Baskin and Baskin 2004c). One theory to explain the
mechanism of physiological seed dormancy is the ‗hormone balance‘ theory. The two
main hormones perceived to be involved in seed dormancy are abscisic acid (ABA) and
gibberellic acid (GA). Under this theory, ABA is responsible for maintaining
dormancy, and GA is responsible for releasing it (Baskin and Baskin 2004c). However,
not all studies consider that GA breaks dormancy. The controversy over whether or not
GA breaks dormancy is sometimes due to differing definitions of dormancy. For
example, GA has been shown to replace the after-ripening requirement of Nicotiana
plumbaginifolia (Grappin et al. 2000). After-ripened seeds had lower ABA content
compared with fresh (dormant) seeds. Addition of GA to dormant seeds decreased their
ABA content, and enabled earlier germination. It is unclear as to whether lower ABA
content of after-ripened seed is due to increased GA content or sensitivity. However,
dormancy in Nicotiana plumbaginifolia was defined as the delay in seed germination,
which is different from the definition of dormancy described by Baskin and Baskin
(2004c) which is that dormant seeds are unable to germinate within a specified period of
time (30 days). As ‗dormant‘ seeds germinated within 21 days, and showed similar
final germination percentages to ‗non-dormant‘ seeds, it is more likely that addition of
GA stimulates germination and that seeds were non-dormant or conditionally dormant.
This is confirmed by Bewley (1997) and others (Hilhorst and Karssen 1992) who
postulate that GA does not control dormancy, rather, ABA inhibits germination, and
once this is overcome, GA can stimulate germination. This second theory is referred to
as the ‗remote control‘ model (Baskin and Baskin 2004c).

29
1.5 Rational aim and thesis outline
Given the lack of topsoil, which contains the soil-stored seed bank, and which is
considered to be the most important source of plants for rehabilitation purposes, this
study investigated seed germination and dormancy mechanisms of the identified
dominant species. Once dormancy was classified, seed pre-treatments to stimulate
germination and overcome dormancy were understood, and germination was optimised,
seed broadcasting (direct to site) and planting techniques were investigated to determine
the potential for plant return. Site and seedling treatments were investigated in order to
optimise seedling recruitment (from broadcast seeding) and seedling survival and
growth (from planted seedlings).
Chapter 2 investigated germination requirements of the dominant species. Germination
requirements included appropriate incubation temperature and smoke products.
Dormancy of these species, if present, was classified and potential dormancy breaking
treatments were recommended.
Chapter 3 focused on six species with physiological dormancy, and determined whether
after-ripening overcame dormancy. The effect of seed moisture content, storage
temperature and time on dormancy loss was investigated.
Chapter 4 was a case study of eight Solanum species in arid Australia, one of which is a
key species occurring at Shark Bay Salt. Germination requirements were investigated
and dormancy mechanisms resolved.
Chapter 5 contained a vegetation survey of the undisturbed vegetation at SBS and four
borrow pits, one of which was subject to topsoil replacement. Dominant species in the
undisturbed vegetation, and dominant species absent from the borrow pits were
identified.
Chapter 6 examined soil properties in undisturbed areas and in the borrow pits requiring
rehabilitation. Site treatments for optimal seedling recruitment from broadcast seeding
were described.
Chapter 7 described site and seedling treatments investigated in order to optimise
seedling survival and growth from planted seedlings.

30
Chapter 8 presented a synthesis of the findings in this study in terms of rehabilitation of
arid zone ecosystems.

31
C H A P T E R 2
Seed biology of 18 species used for rehabilitation
2.1 Abstract
Revegetation of land following disturbance, particularly in arid environments, is often
hindered due to low establishment of seedlings following high levels of intervention.
Information on seed biology and germination cues of keystone species is lacking,
particularly in the arid zone of Australia which is a major zone for mining
developments. This study investigated the seed characteristics and germination cues of
18 common species required for rehabilitation of disturbed areas at Shark Bay Salt in
the Shark Bay World Heritage Area of Western Australia. Untreated seeds of seven
species (Aphanopetalum clematideum, Atriplex bunburyana, Austrostipa elegantissima,
Melaleuca cardiophylla, Pembertonia latisquamea, Rhagodia baccata, Salsola tragus)
exhibited high germination. Seeds of two species (Acacia tetragonophylla, Stylobasium
spathulatum) had low imbibition, which increased with hot-water treatment, and hence
require scarification for germination. Germination of seeds for three species
(Anthocercis littorea, Diplolaena grandiflora, Solanum orbiculatum) substantially
increased with gibberellic acid, smoke water and karrikinolide (a butenolide isolated
from smoke). Seeds of the remaining six species (Dioscorea hastifolia, Eremophila
oldfieldii, Nitraria billardierei, Ptilotus exaltatus, Thryptomene baeckeacea,
Zygophyllum fruticulosum) had low germination regardless of treatment. Most species
germinated equally well at 26/13oC and 33/18
oC, however seven species had improved
performance at 26/13oC. This study is of significance to land managers and
conservation agencies with an interest in optimising germination rates of arid zone seeds
used in restoration.

32
2.2 Introduction
Land rehabilitation in arid zones presents numerous challenges (Anderson and Ostler
2002). In particular, strong seasonal aridity, low and erratic rainfall (Glenn et al. 2001;
Snyman 2003) and high wind activity hamper revegetation of areas following
disturbance, together with a paucity of knowledge on the restoration ecology of native
plants. Compounding the challenges in the arid zone of Australia is the old landscape,
highly evolved flora with a high level of speciation, low dispersal capability of plants
and a high degree of seed dormancy. As a result, revegetation of areas following
disturbance in arid Australia requires a high level of human intervention.
Understanding the seed biology of species used in revegetation projects is vital for
ensuring establishment of seedlings. For example, pre-treating seeds prior to seed
broadcasting to stimulate germination or overcome seed dormancy is the first step in
overcoming the numerous challenges and may increase the likelihood of seed
germination and seedling establishment in these low rainfall ecosystems. However, in
certain areas, little information is available on the seed biology of species needed for
rehabilitation. Limited knowledge on seed germination is currently hampering
rehabilitation efforts in areas following disturbance within Australia (Baskin and Baskin
2003a). One such area is Shark Bay Salt (SBS), a solar salt facility in Western
Australia.
SBS, located in the Shark Bay World Heritage Area, is committed to rehabilitating ca.
84 ha of ‗borrow pits‘ at their solar salt facility. The Shark Bay World Heritage Area is
renowned for its outstanding natural features and serves as a habitat for many
endangered species (DEP 2001). Shark Bay is located in an arid climate that is
transitional between the Mediterranean climate of the southwest and the arid interior of
the country. Most of the 220 mm of rainfall occurs in winter (Shark Bay Salt,
unpublished data); however, the area is occasionally subject to rainfall from summer
cyclonic systems since it is close to the summer/winter rainfall line that bisects the
Australian continent from west to east (Gentilli 1979). In addition, Shark Bay borders
the South West and the Eremaean Biogeographic Regions where southwestern flora
meets northwestern and arid zone flora (CALM 2005). Some species that occur in the
area are restricted to the southwest or northwest of the state, whereas others occur only
on the coast or throughout the state.

33
Seed germination usually occurs during the season of highest water availability (Bell et
al. 1993; Bell 1999), and optimal germination temperature(s) coincide with this period.
The optimum range of germination temperatures is 20-30oC for tropical Australian
species that experience summer rainfall (Bellairs and Ashwath 2007), whereas it is 13-
18oC for Mediterranean-type species in the southwest of Western Australia with winter
rainfall (Bell 1994). Central Australia is subject to aseasonal rainfall, suggesting that
germination can occur at any time of the year. For example, germination of two
Australian arid zone Asteraceae was high at incubation temperatures of 26/13oC and
33/18oC, indicative of winter and summer temperatures (Merritt et al. 2006). Since
Shark Bay is located in a transitional floristic and climatic zone with winter rain and
also sporadic summer rain, we predicted that optimal germination temperatures could
vary among Shark Bay‘s species.
In addition to requiring moisture and an appropriate incubation temperature for
germination, some seeds require a germination stimulant. Gibberellic acid, smoke water
and karrikinolide (formerly known as butenolide (Dixon et al. 2008)) have been used to
promote germination for seeds of numerous species, both in Australia and other
ecosystems (Bell et al. 1995; Dixon et al. 1995; Plummer and Bell 1995; Flematti et al.
2004; Merritt et al. 2006; Kulkarni et al. 2007). Along with the appropriate
temperature, these stimulants may be useful as pre-treatments for broadcast seed or for
seedling production under nursery conditions.
Research on seed biology is limited for arid zone species in Western Australia, and is
scarce for species within the Shark Bay World Heritage Area. Some of the common
Shark Bay species are in genera for which problematic germination is typical, e.g.
Thryptomene (Beardsell et al. 1993), Ptilotus (Williams et al. 1989), Solanum
(Stefaniski 1998; Ahmed et al. 2005) and Stylobasium (Baskin et al. 2006b). Moreover,
no information on germination requirements is available for other species in the genera
Aphanopetalum, Diplolaena and Rhagodia. Thus, the aim of the present study was to
investigate the seed biology of 18 common species within the solar salt facility of Shark
Bay. These 18 species were selected as representative of 14 families and a variety of
life forms (tree, shrub, climber, herb and grass), and those that produced ample seed
quantities. In addition, many of these species are dominant members of the vegetation
(Chapter 5). For each species, we determined (a) fruit, seed and embryo characteristics,
(b) water uptake by un-scarified seeds, and if none, by scarified seeds, (c) germination

34
responses to simulated summer and winter temperatures and (d) the effects of
germination promoting treatments (gibberellic acid, smoke water, karrikinolide).
2.3 Materials and Methods
2.3.1 Seed collections
Fruits were collected within the SBS lease (26o07‘53‖S, 113
o22‘58‖E) in Shark Bay,
Western Australia between September and December 2004 (for 16 species), November
2005 (Melaleuca cardiophylla) or November 2006 (Anthocercis littorea) (Table 2.1).
After collection, seeds were removed from fleshy or dehiscent fruits, and all seeds and
indehiscent fruits were air dried and stored at ambient laboratory conditions (ca. 22oC,
50% RH) until germination testing commenced in February 2005, February 2006 and
January 2007, respectively. Experiments were undertaken on seeds (Acacia
tetragonophylla F.Muell., Anthocercis littorea Labill., Aphanopetalum clematideum
(Harv.) Domin, Atriplex bunburyana F.Muell., Austrostipa elegantissima (Labill.)
S.W.L.Jacobs & J.Everett, Dioscorea hastifolia Endl., Diplolaena grandiflora Desf.,
Melaleuca cardiophylla F.Muell., Rhagodia baccata (Labill.) Moq., Salsola tragus L.,
Solanum orbiculatum Poir., Zygophyllum fruticulosum DC.), intact indehiscent fruits
(Eremophila oldfieldii F.Muell., Pembertonia latisquamea (F.Muell.) P.S.Short, Ptilotus
exaltatus Nees, Stylobasium spathulatum Desf., Thryptomene baeckeacea F.Muell.) or
indehiscent fruits with the fleshy mesocarp removed (Nitraria billardierei DC.).
Hereafter, indehiscent fruits are referred to as seeds. Distribution maps of the recorded
locations of the species are displayed in Appendix 4.
2.3.2 Fruit, seed and embryo characteristics
The types of fruits (dispersal units) were recorded for each species. Dispersal method
and storage strategy (soil-stored or canopy-stored) were inferred from the fruit type.
Seeds were dissected to observe and classify the embryo type according to Martin
(1946) and Baskin and Baskin (2007), and the presence or absence of endosperm was
recorded. Embryo:seed (E:S) ratio was calculated using the following formula: E:S
ratio = embryo length / seed length. Embryo type and E:S ratios were used to determine
if seeds contained a fully developed versus underdeveloped embryo (Baskin and Baskin
2007). A cut test using three replicates of 20 seeds of each species assessed seed
viability. In this test, imbibed seeds were cut in half and inspected. Seeds with firm,

35
white embryos were considered viable and empty seeds, or those with shrivelled or
brown/black embryos, were non-viable (Baskin and Baskin 1998).
2.3.3 Imbibition
For each species, three replicates of >0.03 g of seeds were weighed and placed on moist
germination test paper (Advantec, 84 mm) in Petri dishes at ambient laboratory
conditions (ca. 22oC, 50% RH). After 5 min on moist test paper, seeds were removed,
patted dry with a paper towel to absorb surface moisture and re-weighed, to determine
the initial weight. Seeds were replaced on moist test paper and re-weighed after 1, 2, 4,
6, 24, 48 and 72 h of imbibition. Percent water uptake was determined gravimetrically.
Species that exhibited low percent water uptake (Acacia tetragonophylla and
Stylobasium spathulatum) were subsequently treated with hot (ca. 95oC) water for 2 min
and the imbibition test performed again, with seeds weighed after 1, 2, 4, 6, 24, 48, 72
and 168 h.
2.3.4 Germination requirements
Seeds of all species were soaked for 24 h in four treatment solutions: 2.89 mM
gibberellic acid (GA3), smoke water (SW) (10% v/v), 0.67 μM karrikinolide (KAR1)
(the butenolide, 3-methyl-2H-furo[2,3-c]pyran-2-one) or water (control). Prior to these
treatments, Acacia tetragonophylla and Stylobasium spathulatum seeds were treated
with hot (ca. 95oC) water for 2 min. SW was prepared with straw using the process
described by Dixon et al. (1995) and KAR1 was synthesised in pure form following
Flematti et al. (2005). After soaking, seeds were surface sterilised in 2% (w/v) calcium
hypochlorite (Ca(OCl)2) for 30 min, then rinsed three times with sterilised deionised
water. Seeds were incubated in Petri dishes on water agar (0.7% w/v) at 12/12 h
alternating temperature regimes of 26/13oC or 33/18
oC. These two temperatures
approximate winter and summer temperatures, respectively, in the Shark Bay World
Heritage Area. Petri dishes were wrapped in aluminium foil to exclude light, but were
unwrapped when germination was assessed in the laboratory under ambient lighting.
Four replicates of 25 seeds each were used in all treatments. Germination was defined
as the emergence of the radicle, and was scored regularly until no further germination
was observed (for at least 2 weeks). Time to 50% of the maximum germination was
interpolated from cumulative germination curves for each replicate. Time to final
germination for each replicate also was recorded.

36
2.3.5 Statistical analyses
Germination percentages were arcsine transformed prior to analysis. Germination data
(percentage, time to 50% germination, time to final germination) for each species was
analysed by two-way analyses of variances (ANOVAs) using Genstat 8.1 (Copyright
2005, Lawes Agricultural Trust). Factors included in the ANOVAs were germination
promoting treatment and temperature. If significant differences were detected by
ANOVAs, Fisher‘s LSD was used as the multiple comparison test. Untransformed data
appears in all figures and tables. Viability adjusted germination (VAG) is presented for
species with <65% seed viability (Ptilotus exaltatus, Rhagodia baccata, Salsola tragus,
Thryptomene baeckeacea) and was calculated using the following formula (Sweedman
and Merritt 2006): VAG = (germination (%) / viability (%)) x 100.

37
2.4 Results
2.4.1 Fruit, seed and embryo characteristics
A variety of fruit types was observed among the 18 species, with capsule, utricle and
drupe being the most common (Table 2.1). Many of the seeds had aerodynamic
appendages, suggesting wind dispersal. Nearly all species shed seeds to the soil seed
bank, the exceptions being Melaleuca cardiophylla (canopy-stored seed) and Salsola
tragus (keeps some seeds on the plant and sheds some to the soil). All embryos were
differentiated (showing both a radicle and cotyledons). Seven types of embryos were
identified: linear, peripheral, spatulate, investing, lateral, bent and capitate. Seeds of
several species were exendospermous; hence, the embryos filled the seed. For
endospermous seeds, E:S ratios ranged from 0.16 to 0.90. A species with lateral
embryos (Austrostipa elegantissima) had an E:S ratio of 0.16, and one species with
capitate embryos (Dioscorea hastifolia) had a ratio of 0.39. Seeds of all other species
exhibited an E:S ratio >0.80. Viability of seeds varied from 12% to 100% (Table 2.1).
The majority of species had high viability (>87%), and only four species had <65%
viability. Eremophila oldfieldii was the only species with multiple seeds per fruit; 27 ±
3% (mean ± se) of fruits contained at least one seed and 12 ± 2% of locules per fruit
was filled.
2.4.2 Imbibition
Imbibition studies demonstrated seeds of 16 species readily took up water during 3 days
of imbibition (data not shown). Seeds of Acacia tetragonophylla and Stylobasium
spathulatum did not imbibe water unless they had previously been soaked in hot water
(Fig. 2.1).
a) Acacia tetragonophylla
Time (hours)
0 24 48 72 96 120 144 168
Incre
ase
in
ma
ss (
%)
0
20
40
60
80
100Control
Hot water
b) Stylobasium spathulatum
Time (hours)
0 24 48 72 96 120 144 168
Control
Hot water
Figure 2.1. Percent increase in mass (mean ± SE) of seeds of two species with and
without hot water treatment (ca. 95oC for 2 min).

38
Table 2.1. Fruit, seed and embryo characteristics of 18 species from the Shark Bay World Heritage Area in Western Australia. Family Species Fruit type
Dispersal
method of seeds
Storage strategy of
seeds
Embryo
type
Endosperm
presencea
E:S ratiob
Viability of
seeds (%)
Mimosaceae Acacia tetragonophylla Legume Ballistic, ants Soil-stored Investing No ~1 100 ± 0
Solanaceae Anthocercis littorea Capsule Wind Soil-stored Linear Yes 0.83 ± 0.003 100 ± 0
Cunoniaceae Aphanopetalum clematideum Nut Wind Soil-stored Linear Yes 1.40 ± 0.04 100 ± 0
Chenopodiaceae Atriplex bunburyana Utricle Wind Soil-stored Peripheral Yes >1 88 ± 2
Poaceae Austrostipa elegantissima Caryopsis Wind Soil-stored Lateral Yes 0.16 ± 0.003 97 ± 3
Dioscoreaceae Dioscorea hastifolia Capsule Wind Soil-stored Capitate Yes 0.39 ± 0.015 93 ± 1
Rutaceae Diplolaena grandiflora Schizocarp Gravity Soil-stored Linear Yes 0.90 ± 0.002 100 ± 0
Myoporaceae Eremophila oldfieldii Drupe Wind Soil-stored Linear Yes 0.88 ± 0.01 27 ± 3c
Myrtaceae Melaleuca cardiophylla Capsule Gravity Canopy-stored Linear No ~1 100 ± 0
Zygophyllaceae Nitraria billardierei Drupe Birds Soil-stored Spatulate No ~1 87 ± 7
Asteraceae Pembertonia latisquamea Cypsela Wind Soil-stored Spatulate No ~1 98 ± 2
Amaranthaceae Ptilotus exaltatus Utricle Wind Soil-stored Peripheral Yes >1 46 ± 8
Chenopodiaceae Rhagodia baccata Berry Birds, animals Soil-stored Peripheral Yes >1 63 ± 5
Chenopodiaceae Salsola tragus Utricle Wind Soil & canopy-stored Peripheral No >1 59 ± 2
Solanaceae Solanum orbiculatum Berry Animals Soil-stored Linear Yes >1 95 ± 3
Surianaceae Stylobasium spathulatum Drupe Gravity Soil-stored Spatulate No ~1 98 ± 2
Myrtaceae Thryptomene baeckeacea Nut Gravity Soil-stored Bent No ~1 12 ± 4
Zygophyllaceae Zygophyllum fruticulosum Capsule Wind Soil-stored Spatulate Yes 0.89 ± 0.02 100 ± 0 a Endosperm or perisperm
b Embryo:seed (E:S) ratio of seeds without endosperm is indicated as ~1 since the embryo completely fills the seed. E:S ratio of seeds with a curved or coiled
embryo that is longer than the seed is indicated as >1. c Multiple seeds per fruit; 27 ± 3% (mean ± se) of fruits contained at least one seed and 12 ± 2% of locules per fruit was filled.
38

39
2.4.3 Germination requirements
Responses (as judged by at least one of the three germination parameters – final
germination, time to 50% germination, time to final germination) of Acacia
tetragonophylla, Anthocercis littorea, Atriplex bunburyana, Austrostipa elegantissima,
Dioscorea hastifolia, Diplolaena grandiflora, Melaleuca cardiophylla, Pembertonia
latisquamea and Solanum orbiculatum seeds to germination promoting treatments were
highly dissimilar between temperatures (treatment x temperature, P ≤0.041). Whereas
temperature only affected all three parameters for Aphanopetalum clematideum,
treatment only did so for two parameters of Ptilotus exaltatus (P ≤0.008). In contrast,
treatment and temperature effects (but not their interaction) were shown to be
significant in at least one parameter for Nitraria billardierei, Rhagodia baccata, Salsola
tragus and Stylobasium spathulatum (P ≤0.036). Neither treatment nor temperature
affected Eremophila oldfieldii, Thryptomene baeckeacea and Zygophyllum fruticulosum
germination.
Final germination for seeds of eight species (Acacia tetragonophylla, Aphanopetalum
clematideum, Atriplex bunburyana, Austrostipa elegantissima, Melaleuca cardiophylla,
Pembertonia latisquamea, Rhagodia baccata, Salsola tragus) soaked in water (control)
was high (>65%) when incubated at one or both temperature regimes (Fig. 2.2). Final
germination for the remaining species was <60% when incubated at both temperatures
including Eremophila oldfieldii, which failed to germinate in any treatment.
Final germination of ten species was (further) enhanced by GA3. For four of these
species, final germination was promoted by GA3 at both incubation temperatures.
Anthocercis littorea seeds exhibited >70% germination at both 26/13oC and 33/18
oC
with the addition of GA3, whereas untreated seeds failed to germinate (Fig. 2.2b).
Diplolaena grandiflora and Ptilotus exaltatus seeds showed a two-fold increase in final
germination when treated with GA3 compared to seeds treated with water (Fig. 2.2g, l).
Also, GA3 decreased time to 50% germination of Diplolaena grandiflora when
incubated at 33/18oC (Table 2.2). With the addition of GA3, final germination of
Solanum orbiculatum seeds increased from 10% to 75% at 26/13oC and from 26% to
90% at 33/18oC (Fig. 2.2o). In addition, time to 50% decreased from 21 to 4 days at
26/13oC (Table 2.2). Final germination of the other six species was only promoted by
GA3 at one incubation temperature. GA3 increased germination of Aphanopetalum
clematideum, Austrostipa elegantissima, Pembertonia latisquamea, Rhagodia baccata

40
and Thryptomene baeckeacea when seeds were incubated at 33/18oC, but not at 26/13
oC
(Fig. 2.2c, e, k, q). Conversely, GA3 increased final germination of Dioscorea hastifolia
seeds at 26/13oC, but not at 33/18
oC (Fig. 2.2f). For the remaining eight species, GA3
had no effect on final germination, but of these, four species (Austrostipa elegantissima,
Dioscorea hastifolia, Melaleuca cardiophylla, Salsola tragus) demonstrated quicker
germination with GA3 (either time to 50% or time to final) in at least one temperature
(Table 2.2).
SW and/or KAR1 increased final germination of six species. Both of these stimulants
increased final germination of Anthocercis littorea and Solanum orbiculatum seeds
compared with the control at both incubation temperatures. Untreated seeds of
Anthocercis littorea did not germinate, but both KAR1 and SW elicited 40-71%
germination (Fig. 2.2b). For Solanum orbiculatum, final germination of KAR1 treated
seeds was greater than SW treated seeds at 26/13oC (Fig. 2.2o). Both SW and KAR
decreased time to 50% germination of Solanum orbiculatum at 26/13oC. Germination
of Diplolaena grandiflora and Pembertonia latisquamea seeds was promoted at either
one or both temperatures, depending on the treatment. SW increased final germination
of Diplolaena grandiflora seeds at both 26/13oC and 33/18
oC, and decreased the time to
50% germination at 33/18oC, but KAR1 only increased final germination at 33/18
oC
(Fig. 2.2g). KAR1 increased final germination of Pembertonia latisquamea seeds at
both 26/13oC and 33/18
oC, while SW only increased final germination at 33/18
oC (Fig.
2.2k). Final germination of Aphanopetalum clematideum and Ptilotus exaltatus seeds
was only increased at one temperature. SW increased final germination of
Aphanopetalum clematideum seeds at 33/18oC (Fig. 2.2b). Both SW and KAR1
increased final germination of Ptilotus exaltatus seeds at 26/13oC (Fig. 2.2l) and
decreased the time to final germination at both temperatures (Table 2.2). SW and KAR1
had no effect on the final germination of the remaining twelve species.
Percentages of germination for ten species were unaffected by incubation temperature
(Acacia tetragonophylla, Anthocercis littorea, Dioscorea hastifolia, Diplolaena
grandiflora, Eremophila oldfieldii, Ptilotus exaltatus, Salsola tragus, Stylobasium
spathulatum, Thryptomene baeckeacea, Zygophyllum fruticulosum) (Fig. 2.2).
Incubation temperature also showed no effect on the speed of germination (time to 50%
germination) for four of those species (Ptilotus exaltatus, Salsola tragus, Thryptomene
baeckeacea, Zygophyllum fruticulosum) (Table 2.2). Except for Salsola tragus, where

41
time to final germination was slowest at 33/18oC, time to final germination was
independent of temperature for the other three species (Table 2.2). Germination of
Anthocercis littorea, Dioscorea hastifolia and Diplolaena grandiflora seeds was slower
at 33/18oC compared with 26/13
oC for some treatments, whereas germination of Acacia
tetragonophylla and Stylobasium spathulatum seeds was slower at 26/13oC compared
with 33/18oC (Table 2.2).
Conversely, germination for seeds of the other eight species was affected by
temperature. Seeds of seven species (Aphanopetalum clematideum, Atriplex
bunburyana, Austrostipa elegantissima, Melaleuca cardiophylla, Nitraria billardierei,
Pembertonia latisquamea, Rhagodia baccata) exhibited higher germination when
incubated at 26/13oC, compared with 33/18
oC (Fig. 2.3). Aphanopetalum clematideum
seeds showed substantially higher germination at 26/13oC (range of 94-96%) compared
with 33/18oC (range of 6-15%), and germination was faster at 26/13
oC compared with
33/18oC (Fig. 2.2c, Table 2.2). Atriplex bunburyana seeds had almost a two-fold higher
germination when incubated at 26/13oC compared with 33/18
oC but germination speed
was the same (Fig. 2.2d, Table 2.2). Germination of Austrostipa elegantissima,
Pembertonia latisquamea and Rhagodia baccata seeds was greater when seeds were
incubated at 26/13oC compared with 33/18
oC, but the speed of germination of Rhagodia
baccata was the same at both temperatures (Fig. 2.2e, k, m, Table 2.2). Germination of
control, SW and KAR1 treated Melaleuca cardiophylla seeds was slightly higher when
incubated at 26/13oC and the speed of germination was higher at 26/13
oC than 33/18
oC
(Fig. 2.2h, Table 2.2). Seeds of Nitraria billardierei germinated only slightly better at
26/13oC (Fig. 2.2j, Table 2.2) but with SW germinated faster at 33/18
oC. Seeds of the
remaining species (Solanum orbiculatum) exhibited higher germination when incubated
at 33/18oC with GA3 or SW but the speed of germination was the same at both
temperatures, except for the control where it was slower at 26/13oC (Fig. 2.2o, Table
2.2).

42
Ge
rmin
atio
n (
%)
0
20
40
60
80
100
a) Acacia tetragonophyllaG
erm
ina
tio
n (
%)
0
20
40
60
80
100
c) Aphanopetalum clematideum
Ge
rmin
atio
n (
%)
0
20
40
60
80
100
aa
dcd cd
bc
a
b
c
a
c
b
c
b
c
ab
g) Diplolaena grandiflora
d) Atriplex bunburyana
b
a
b
a
b
a
b
a
e) Austrostipa elegantissima
Ge
rmin
atio
n (
%)
0
20
40
60
80
100
a
b
aa
c c cc
i) Melaleuca cardiophylla
Control GA SW KAR
Ge
rmin
atio
n (
%)
0
20
40
60
80
100c
abbc bc bc
a a
c
f) Dioscorea hastifolia
a
d
ab
cdbcd
abc abc a
b) Anthocercis littorea
c
cc
b
b
b
a a
h) Eremophila oldfieldii
j) Nitraria billardierei
Control GA SW KAR
aabc aabc bcdcddd

43
n) Salsola tragus (VAG)
r) Zygophyllum fruticulosum
Control GA SW KAR
p) Stylobasium spathulatum
a
abab
bbababab
o) Solanum orbiculatum
Ge
rmin
atio
n (
%)
0
20
40
60
80
100
l) Ptilotus exaltatus (VAG)
m) Rhagodia baccata (VAG)
Ge
rmin
atio
n (
%)
0
20
40
60
80
100
q) Thryptomene baeckeacea (VAG)
Control GA SW KAR
Ge
rmin
atio
n (
%)
0
20
40
60
80
100
a ab
c
bc bcbc
bcbc
a
a
b
cdbc
e e e
bc
a
c
b
bc
a
bc
a
a a
b
ab
ab
abab ab
26/13C
33/18C
26/13C
33/18C
26/13C
33/18C
k) Pembertonia latisquamea
Ge
rmin
atio
n (
%)
0
20
40
60
80
100
c
a
cc
b
b
b
d
26/13oC 33/18
oC
Figure 2.2. Germination (mean ± SE) for seeds of 18 species treated with water
(control), gibberellic acid (GA), smoke water (SW) or karrikinolide (KAR) and
incubated at 26/13oC or 33/18
oC. Dissimilar letters indicate significant differences
(P≤0.05) among treatments and temperatures within each species. Species without
letters on the graph did not exhibit treatment differences. VAG indicates germination is
presented as viability adjusted germination.

44
Table 2.2. Time to 50% germination and time to final germination of seeds treated with
water (control), gibberellic acid (GA3), smoke water (SW) or karrikinolide (KAR1) and
incubated at 26/13oC or 33/18
oC. Numbers in brackets indicate standard error, and
dissimilar letters indicate significant differences (P≤0.05) among treatments and
temperatures within a parameter for each species.
Speciesa Treatment Time to 50%
germination (days)
Time to final
germination (days)
26/13oC 33/18oC 26/13oC 33/18oC
Acacia Control 5 (1.0) ab 5 (1.6) a 23 (6.2) abc 14 (0.5) a
tetragonophylla GA3 7 (0.5) bc 5 (0.6) ab 26 (4.5) cd 15 (0.9) a
SW 8 (0.5) c 6 (1.5) ab 32 (4.0) d 22 (8.9) abc
KAR1 8 (1.7) c 6 (0.2) ab 17 (2.1) ab 25 (8.7) bcd
Anthocercis Control -b - b - b - b
littorea GA3 18 (1.7) ab 17 (0.7) a 54 (7.4) bc 61 (3.4) c
SW 18 (3.0) a 23 (0.8) bc 34 (0.0) a 58 (4.4) c
KAR1 18 (0.7) a 28 (2.0) c 43 (5.5) ab 58 (3.4) c
Aphanopetalum Control 10 (0.4) a 51 (18.6) b 20 (2.5) a 59 (21.2) b
clematideum GA3 10 (0.2) a 45 (12.7) b 20 (2.5) a 80 (10.5) b
SW 9 (0.4) a 50 (7.0) b 20 (5.1) a 62 (12.6) b
KAR1 12 (0.3) a 62 (16.4) b 19 (0.8) a 62 (16.4) b
Atriplex Control 6 (0.3) a 7 (0.5) a 18 (3.3) a 18 (2.8) a
bunburyana GA3 6 (0.0) a 6 (0.1) a 22 (3.5) a 22 (2.5) a
SW 6 (0.5) a 10 (1.4) b 18 (3.3) a 25 (2.0) a
KAR1 7 (0.2) a 7 (0.8) a 19 (3.0) a 20 (3.9) a
Austrostipa Control -c - c 25 (2.5) c 29 (1.4) cd
elegantissima GA3 - - 9 (0.4) a 16 (0.5) b
SW - - 14 (0.7) ab 27 (0.0) cd
KAR1 - - 31 (3.8) d 42 (0.0) e
Dioscorea Control 17 (2.6) a 91 (0.0) d 46 (32.4) a 162 (0.0) c
hastifolia GA3 18 (2.2) a 43 (29.4) b 113 (61.1) bc 144 (35.5) c
SW 27 (10.3) ab 70 (14.0) c 74 (59.0) ab 162 (0.0) c
KAR1 21 (6.2) a 88 (20.6) cd 46 (21.6) a 162 (0.0) c
Diplolaena Control 18 (1.6) a 46 (22.2) b 46 (23.2) a 81 (7.0) b
grandiflora GA3 16 (1.7) a 17 (2.7) a 44 (22.2) a 81 (7.0) b
SW 17 (0.9) a 22 (3.8) a 38 (10.6) a 56 (14.0) ab
KAR1 22 (3.3) a 41 (11.8) b 58 (30.7) ab 81 (7.0) b
Melaleuca Control 7 (0.1) a 13 (0.3) c 16 (1.8) ab 34 (1.5) c
cardiophylla GA3 6 (0.4) a 9 (0.4) b 9 (0.3) a 37 (4.2) cd
SW 7 (0.2) a 17 (0.2) d 12 (0.9) ab 44 (5.0) d
KAR1 7 (0.0) a 17 (1.1) d 21 (4.8) b 39 (3.5) cd
Nitraria Control 13 (1.5) abc 11 (0.0) ab 19 (3.1) abc 11 (0.0) a
billardierei GA3 7 (1.4) a 9 (0.3) ab 14 (7.4) ab 9 (0.4) a
SW 19 (4.3) c 10 (3.4) ab 29 (4.7) cd 13 (1.5) a
KAR1 -b 16 -d -b 28 -d
Ptilotus Control 5 (2.1) ab 6 (2.8) b 30 (2.0) c 33 (3.4) c
exaltatus GA3 4 (0.5) ab 4 (0.2) ab 10 (3.2) ab 20 (5.0) b
SW 5 (0.7) ab 4 (0.8) ab 15 (5.0) ab 9 (1.4) a
KAR1 3 (0.2) ab 2 (0.1) a 9 (1.4) a 9 (2.3) a
Rhagodia Control 6 (1.0) a 8 (2.6) ab 28 (5.0) a 20 (5.8) a
baccata GA3 5 (0.7) a 7 (0.4) ab 16 (1.7) a 27 (4.6) a
SW 6 (0.8) a 8 (0.6) ab 21 (5.1) a 17 (3.7) a
KAR1 6 (0.4) a 10 (2.0) b 24 (5.0) a 26 (5.8) a
Salsola Control 3 (0.2) a 4 (0.0) a 20 (5.3) bc 8 (0.8) a
tragus GA3 3 (0.6) a 3 (0.6) a 9 (0.3) a 9 (0.6) a
SW 4 (0.5) a 4 (0.3) a 23 (5.0) c 13 (2.7) ab
KAR1 4 (0.5) a 3 (0.4) a 20 (3.2) bc 10 (1.8) a

45
Speciesa Treatment Time to 50%
germination (days)
Time to final
germination (days)
26/13oC 33/18oC 26/13oC 33/18oC
Solanum Control 21 (15.3) b 5 (0.6) a 48 (6.8) b 15 (7.1) a
orbiculatum GA3 4 (0.29) a 3 (0.2) a 14 (1.6) a 10 (2.9) a
SW 4 (0.29) a 3 (0.5) a 14 (2.6) a 12 (3.8) a
KAR1 4 (0.29) a 3 (0.2) a 17 (4.0) a 11 (3.2) a
Stylobasium Control 79 (8.1) d 55 (8.0) bc 130 (0) c 91 (0.0) abc
spathulatum GA3 84 (6.2) d 37 (10.2) ab 120 (9.8) bc 79 (26.6) ab
SW -c 18 (2.7) a -e 73 (10.7) a
KAR1 67 (11.9) cd 34 (5.9) ab 110 (20.3) abc 70 (8.6) a
Zygophyllum Control 5 (0.4) a 6 (2.9) ab 8 (0.5) a 18 (8.4) abc
fruticulosum GA3 5 (0.4) a 10 (4.1) ab 8 (1.3) ab 19 (7.1) abc
SW 8 (1.1) ab 14 (4.8) b 27 (7.0) bc 28 (6.8) c
KAR1 10 (3.0) ab 6 (3.0) a 27 (9.1) bc 22 (5.9) abc
a Time to 50% germination and time to final germination are not reported for Eremophila oldfieldii (given that seeds
failed to germinate in all treatments), Pembertonia latisquamea, and Thryptomene baeckeacea (due to low
germination). b Time to 50% germination and time to final germination are not recorded as there was no germination in these
treatments. c Time to 50% germination was not recorded. d No standard error is recorded because germination occurred in only one replicate. e Time to final germination was not recorded.

46
2.5 Discussion
Our study represents the first to investigate the seed biology for a wide variety of
species from the arid zone of Western Australia, focusing on the Shark Bay World
Heritage Area. Using the information from the seed and embryo characteristics,
imbibition studies and germination requirements enabled us to categorise the class of
dormancy found in the species (sensu Baskin and Baskin 2004c). With the exception of
two species (Acacia tetragonophylla, Stylobasium spathulatum), seeds of all species
readily imbibed water. Water uptake by Acacia tetragonophylla and Stylobasium
spathulatum seeds was very low but it was greatly increased by treatment with hot
water, indicating that untreated seeds are impermeable to water and physical dormancy
is present. Acacia spp. are well known to exhibit physical dormancy and soaking in hot
water has previously been found to alleviate dormancy (Bell et al. 1993). Stylobasium
spathulatum also has been shown to exhibit physical dormancy (Baskin et al. 2006b).
However, in our study, hot water treatment elicited germination of <50% of the seeds.
For the ungerminated seeds, either the hot water treatment did not alleviate dormancy of
the entire seed lot or they exhibit combinational dormancy (physical and physiological),
as postulated by Baskin et al. (2006b).
Seeds of six species (Dioscorea hastifolia, Eremophila oldfieldii, Nitraria billardierei,
Ptilotus exaltatus, Thryptomene baeckeacea, Zygophyllum fruticulosum) had low or
slow germination regardless of treatment, and are therefore likely to exhibit
physiological dormancy since they had fully developed embryos. Although the E:S
ratio is small (<0.5) in Dioscorea hastifolia seeds, they have capitate embryos and do
not exhibit morphological or morphophysiological dormancy (Baskin and Baskin 1998).
Other studies have also found slow seed germination of Dioscorea species (Terui and
Okagami 1993; Albrecht and McCarthy 2006), low (and slow) germination of Nitraria
billardierei seeds (Noble and Whalley 1978), low germination of Ptilotus exaltatus
seeds (Williams et al. 1989) and no germination (of fresh seeds) of Thryptomene
calycina (Beardsell et al. 1993). Three levels of physiological dormancy are recognised
that are differentiated partly on responses to GA3, with germination of non-deep and
intermediate levels being stimulated by GA3 (Baskin and Baskin 2004c). However,
GA3 only stimulated germination of Ptilotus exaltatus (at both incubation temperatures)
indicating that the other species may exhibit deep physiological dormancy.

47
Low germination was also observed for three other species (Anthocercis littorea,
Diplolaena grandiflora, Solanum orbiculatum), but germination was substantially
enhanced by smoke products – SW and KAR1. Our finding is analogous to other
studies which have categorised Anthocercis as a post-fire ephemeral (Pate et al. 1985;
Bell et al. 1993) and to the observation that the spread of Solanum species is encouraged
by fire (Latz 1995). However, since seeds of the Australian Solanum centrale did not
respond to SW (Stefaniski 1998), but responded to aerosol smoke after nicking, it was
concluded that they have a water impermeable seed coat (Ahmed et al. 2005). In our
study, Solanum orbiculatum seeds readily imbibed water (see also Chapter 4). Solanum
orbiculatum seeds treated with SW or KAR1 had high (82-97%) final germination, and
time to 50% germination was only 3-4 days. Thus, seeds of this species were
apparently non-dormant when germination testing commenced, and smoke acted as a
germination stimulant (Merritt et al. 2007; Ooi et al. 2007). In contrast, Anthocercis
littorea and Diplolaena grandiflora seeds had low to moderate germination (23-76%)
and time to 50% germination was 17-41 days. Apparently, a portion of these seed lots
were physiologically dormant when germination testing started. Additional storage
(and after-ripening) may have increased the percentages and speed of germination,
particularly for Anthocercis littorea (see also Chapter 3).
Untreated and treated seeds of seven species had high final germination: Atriplex
bunburyana, Aphanopetalum clematideum, Austrostipa elegantissima, Melaleuca
cardiophylla, Pembertonia latisquamea, Rhagodia baccata and Salsola tragus. At the
time of testing, seeds of these species were apparently non-dormant. Although it is
possible that these seeds exhibited physiological dormancy when they were collected,
which was relieved by after-ripening during storage. Germination for only three of
these genera (Atriplex, Austrostipa, Melaleuca) has been studied in Australia. A study
of two other Western Australian Austrostipa species concluded that seeds were either
non-dormant or physiologically dormant (Baker et al. 2005a). However, after-ripening
at 23oC and 50% RH increased germination in Austrostipa elegantissima, indicating that
physiological dormancy is present in fresh seeds (Turner et al., unpublished data).
Melaleuca cardiophylla stores its seed in the canopy, and seeds of many canopy-stored
species are non-dormant (Baskin and Baskin 2003a). Germination of Melaleuca
cardiophylla was similar to Melaleuca acerosa, a Western Australian species that
exhibited high (>90%) germination at three incubation temperatures (10, 15, 20oC) with
no other treatment (Bellairs and Bell 1990). Seeds of most Atriplex spp. are categorised

48
as having physiological dormancy (Baskin and Baskin, 1998). Germination of Atriplex
spp. increases when bracts (or bracteoles) are removed from seeds, which was done in
our study, although it is not always maximised with this method. For example, whereas
germination of Atriplex undulata was ≥76% after bract removal, that of Atriplex
amnicola and Atriplex nummularia was ≤60% (Stevens et al. 2006).
Seven of our study species had substantially higher germination at 26/13oC than at
33/18oC. Suppression of germination at high temperatures could prevent germination
during summer months when occasional cyclonic rainfall may provide a transient period
of conditions suitable for germination, but uncertain conditions for establishment. In
the Shark Bay region, which is located south of the summer-winter rainfall line for
Australia (Gentilli 1979), most precipitation occurs in winter. Germination of these
species in winter is analogous to findings of Mott and Groves (1981) and Jurado and
Westoby (1992). They concluded that forbs and dicotyledonous species generally
germinate with winter rains, whereas summer rainfall elicits germination of grasses and
other monocotyledons. However, 30% of the species in Jurado and Westoby‘s (1992)
study did not show a temperature preference. Similarly, 10 species in our study had
similar germination percentages between the two temperature regimes. Interestingly,
the two species that store all (or a portion) of their seeds in capsules on the canopy (Bell
and Bellairs 1992; Borger et al. 2007), thus implying that germination is controlled by
capsule dehiscence, were either indifferent to temperature (Salsola tragus) or had
slightly higher germination at the lower temperature (Melaleuca cardiophylla). Our
results on the higher temperature responses of Solanum orbiculatum are consistent with
those of the arid Solanum centrale. Ahmed et al. (2005) found that incubation
temperatures of 12, 20 and 28oC (corresponding to winter, spring/autumn and summer)
had no effect on germination of this species, and they postulated that germination was
linked with moisture cues rather than seasonal temperature. Likewise, species showing
no temperature preference for germination also may rely on moisture rather than
temperature per se to regulate the timing of germination in nature.
Jurado and Westoby (1992) hypothesised that quick germinating species would take
advantage of a single rainfall event, whereas slow germinating ones would wait for
several rainfall events ensuring adequate moist soil over a considerable length of time.
As such, they found that arid zone species germinated more quickly than species from
other ecosystems in Australia. Moreover, Cochrane et al. (2002) demonstrated that

49
emergence times are slower across a wide and taxonomically diverse range of taxa from
the (non-arid) southwest of Western Australia. In contrast, Fenner and Thompson
(2005) suggested that species exhibiting fast germination are likely to be favoured in
drought free ecosystems, whereas species with slow germination are tolerant of
droughts. In our study, germination speeds (i.e., time to 50% germination) were highly
variable: <10 days (nine species), 11-20 days (four species) and >20 days (one species).
Our study site was at the very arid northern extremity of the Southwest. Thus, the
majority of our species germinated relatively quickly, which would allow a rapid
response to sporadic and infrequent rainfall events that characterise the Shark Bay
region. With only 33% of species responsive to the smoke stimulant, fire may play a
secondary role to germination, after moisture.
A variety of dormancy classes are exhibited among habitats, but the proportion of them
differs between biogeographical zones. For example, deserts have a relatively high
proportion of species with physiological dormancy, moderate with physical dormancy
and low of non-dormant, morphological and morphophysiological dormancy (Baskin
and Baskin 1998). Although we cannot definitively categorise the class of dormancy
found in all of our study species, several important comments can be made in
comparison to Baskin and Baskin‘s (1998) biogeographical survey. Physiological
dormancy is common among shrubs and herbs in deserts, and our study categorised
44% of species (all shrubs and herbs) as exhibiting physiological dormancy. Physical
dormancy is most common in desert trees, and the only tree in our study, Acacia
tetragonophylla, had this class of dormancy. We did not document any species with
morphological and morphophysiogical dormancies supporting the rarity of these
dormancy classes in arid ecosystems (Baskin and Baskin 1998). In contrast, in the
temperate region of south-eastern Australia proportions of physical and physiological
dormancy are nearly equal (ca. 40% each) and are greater than morphophysiological
dormancy (17%) and morphological dormancy (<5%) (Ooi 2007).
Our study has important implications for successful germination and nursery production
of species for restoration of mined sites in the arid zone of Western Australia. Before
sowing, species with physical dormancy will need to be scarified to allow imbibition.
Following storage in the laboratory, seeds of most species germinated to moderate to
high percentages particularly at 26/13oC. In fact, we would recommend this
temperature regime for incubation since it was effective for the majority of species.

50
Moreover, seeds should be sown in the field just prior to winter when moisture and
(low) temperature conditions are most favourable for germination. Application of GA3,
SW and/or KAR1 significantly improved germination in several species, and we suggest
pre-treating seeds with these chemicals prior to sowing. A few of our study species
germinated to low percentages, regardless of treatment or temperature. Additional
studies are necessary to further explore the requirements for dormancy break.

51
C H A P T E R 3
Storage conditions can influence after-ripening
of arid zone seeds
3.1 Abstract
The effects of after-ripening (storage under warm, dry conditions) on seed dormancy
loss and germination was examined on six Western Australian arid zone plant species,
which represent key life form types and important components of rehabilitation
objectives: Acanthocarpus preissii Lehm., Anthocercis littorea Labill., Dioscorea
hastifolia Endl., Eremophila oldfieldii F.Muell., Thryptomene baeckeacea F.Muell. and
Zygophyllum fruticulosum DC.. After-ripening was investigated by adjusting seed
moisture content to 13%, 50% and 75% equilibrium relative humidity (eRH) at 20oC,
and storing seeds at two temperatures (30oC and 45
oC) from 1 to 18 months. Following
storage, seeds were incubated in water, gibberellic acid (GA3) or karrikinolide (KAR1)
(the butenolide, 3-methyl-2H-furo[2,3-c]pyran-2-one). All after-ripening conditions
increased germination percentage and rate of Anthocercis littorea and Dioscorea
hastifolia, with Anthocercis littorea only germinating when treated with GA3 or KAR1.
The germination of Zygophyllum fruticulosum was dependent on after-ripening
temperature and seed moisture content. After-ripening had a limited effect on the
remaining three species. The ecological and restoration implications of the findings are
discussed.

52
3.2 Introduction
Understanding seed germination biology is crucial to seed conservation and
rehabilitation efforts (Merritt et al. 2004). This is particularly relevant in the Australian
arid zone, given the old landscape, highly evolved flora, high level of flora speciation,
poor seed dispersal and ultimately a high level of seed dormancy in seeds of many
native plant species. Further, unlike many other ecosystems, there is a paucity of
knowledge on seed germination and dormancy characteristics of Australian arid zone
species.
In desert environments, the majority of plant species (>80%) produce seeds that are
dormant at maturity, with the most common type being physiological dormancy (Baskin
and Baskin 1998). Dormancy type is generally overcome when seeds are exposed to
environmental conditions in their natural habitat. However, if seeds are collected at the
time of dispersal and stored ex-situ prior to use in rehabilitation procedures, the storage
conditions may not be conducive to dormancy loss. Seeds destined for use in
rehabilitation are usually used within a few months or 1-2 years following collection,
and storage of these seeds at 5oC or -18
oC (conditions more conducive to prolonging
longevity) may not be appropriate for maximising dormancy loss / germination in the
short term (Merritt et al. 2004). Understanding how to overcome or accelerate
dormancy loss of stored seeds is therefore useful to optimise rehabilitation procedures
such as seed broadcasting and nursery seedling production.
Concepts for alleviating seed dormancy can be derived from the environmental
conditions (primarily temperature and soil/seed moisture content) between seed
dispersal and germination (Baskin and Baskin 2004a; Merritt et al. 2007; Hoyle et al.
2008). Seed dispersal at Shark Bay, an arid region in Western Australia occurs in
spring, and seeds are exposed to a hot, dry summer (with occasional cyclonic systems),
a warm autumn, and a cool, wet winter. Average daily maximum temperatures in
summer are >60oC and the average daily temperatures are 32-34
oC. If seeds germinate
during the winter period of reliable rainfall, then warm, dry conditions during summer
may contribute to dormancy loss. The loss of dormancy under warm (≥15oC) and dry
conditions is known as after-ripening (or dry storage) (Baskin and Baskin 2004b).
After-ripening is well known to alleviate dormancy / promote germination of many
Australian species, including some from the arid zone (Mott and Groves 1981; Jurado

53
and Westoby 1992; Bell 1999; Plummer et al. 2001; Tieu et al. 2001a; Schütz et al.
2002; Turner et al. 2006a; Merritt et al. 2007; Hoyle et al. 2008).
Seed dormancy loss of Australian native plant species via after-ripening under ambient
laboratory conditions can be a slow process, requiring several months or years (Tieu et
al. 2001b; Schütz et al. 2002; Baker et al. 2005b). However, by manipulating
temperature and seed moisture content, it is possible to accelerate after-ripening
(Probert 2000). The effect of temperature and seed moisture content on after-ripening
has not been fully investigated on seeds of Australian species, but general principles can
be derived from studies on other species (Baskin and Baskin 1979; Esashi et al. 1993;
Foley 1994; Sharif-Zadeh and Murdoch 2001; Steadman et al. 2003). Increasing the
temperature at which seeds are after-ripened generally increases the rate of dormancy
loss (Cohn and Hughes 1981; Foley 1994; Allen et al. 1995; Murdoch and Ellis 2000),
however, at high temperatures, seed aging may occur (Walters 1998; Steadman et al.
2003). In general, seeds stored at low moisture contents do not after-ripen and seeds
stored at high moisture contents lose viability. For example, Baskin and Baskin (1979)
found that Draba verna seeds failed to after-ripen when stored at 25oC for six months at
0-20% relative humidity (RH), decayed when stored between 70-100% RH, but after-
ripened at 40 and 60% RH.
In a number of native plant species, the use of germination promoting compounds such
as gibberellic acid (GA3) and smoke products (smoke water and karrikinolide) can
increase the germination of seeds that respond to after-ripening. For example, GA3 has
been shown to partially overcome the after-ripening requirement for Schoenia filifolia
subsp. subulifolia, an Australian arid zone daisy (Plummer et al. 2001). Smoke water
(SW) and karrikinolide (KAR1 - the butenolide, 3-methyl-2H-furo[2,3-c]pyran-2-one)
stimulate germination of many Australian plant species (Dixon et al. 1995; Flematti et
al. 2004; Merritt et al. 2006; Dixon et al. 2008; chapter 2) and seeds may become more
responsive to the smoke cue with after-ripening. For example, Anigozanthos manglesii
seeds become more smoke responsive with increasing shelf storage time (Tieu et al.
2001b) and germination of Actinotus leucocephalus seeds gradually increases with the
combination of increasing laboratory storage time and SW treatment (Baker et al.
2005b).

54
In the Australian arid zone, a solar salt facility operated by Shark Bay Salt and located
in the Shark Bay World Heritage Area, Western Australia, contains numerous borrow
pits (soil donor sites for road and bund construction), which are devoid of vegetation
and need to be rehabilitated. An understanding of seed biology is essential to enable
rehabilitation procedures. This study focuses on six species common in the vegetation
of the Shark Bay World Heritage Area; Acanthocarpus preissii (Dasypogonaceae),
Anthocercis littorea (Solanaceae), Dioscorea hastifolia (Dioscoreaceae), Eremophila
oldfieldii (Myoporaceae), Thryptomene baeckeacea (Myrtaceae) and Zygophyllum
fruticulosum (Zygophyllaceae). The species are representative of key life form types;
herb (Acanthocarpus preissii), shrub (Anthocercis littorea, Eremophila oldfieldii,),
climber (Dioscorea hastifolia), spreading to prostrate shrub (Thryptomene baeckeacea),
scrambling, succulent shrub (Zygophyllum fruticulosum) (Western Australian
Herbarium 1998–). Previous research has either identified low or slow germination and
postulated dormancy (Turner et al. 2006b; chapter 2). As seeds imbibe water and have
a developed embryo, they are likely to exhibit physiological dormancy. Physiologically
dormant seeds may after-ripen (Baskin and Baskin 2004c) and previous research on
genera related to the selected study species indicates that after-ripening may be a useful
strategy for overcoming dormancy. For example, dormancy of Thryptomene calycina
was partially alleviated by laboratory storage (20oC for 90 days) as fresh seeds failed to
germinate, whereas 50% of stored seeds germinated (Beardsell et al. 1993). Similarly,
seeds within 10 month old fruits of Eremophila maculata failed to germinate, whereas
germination occurred from seeds in 22 and 34 month old fruit (Richmond and
Ghisalberti 1994), implying an after-ripening effect.
The aim of this chapter was to investigate whether seeds of the study species after-ripen,
and if so, the effects of storage environment on the rate of after-ripening, to determine
the potential to overcome seed dormancy during ex-situ storage. Relatively high storage
temperatures were used, partly to accelerate dormancy loss and partly because seeds are
naturally exposed to high temperatures over the summer period at Shark Bay. Hence,
this study examined the effect of (a) seed moisture content, (b) storage temperature and
(c) GA3 and KAR1 on germination of seeds after storage.

55
3.3 Materials and Methods
3.3.1 Seed moisture content manipulation and storage
This experiment was implemented in two parts; the first part commencing in 2006 and
the second in 2007. Due to limited seed numbers, not all species/seed batches were
subject to all treatment combinations. Distribution maps of the recorded locations of the
species are displayed in Appendix 4.
For the first part of the experiment, indehiscent fruits (hereafter seeds) of Thryptomene
baeckeacea and seeds of Zygophyllum fruticulosum were collected on the Edel
Peninsula in Shark Bay, Western Australia (S 26o 07‘ 53‘‘ E 113
o 22‘ 58‘‘) in
November 2005. Seeds were cleaned and stored at ambient laboratory conditions (ca.
23oC). In May 2006, seeds of both species were sealed in porous nylon mesh bags (0.75
µm). Seed moisture content was adjusted by holding seeds in boxes over saturated salts
at one of three RHs (13%, 50% or 75%) for three weeks at ambient laboratory
conditions (ca. 23oC). Each RH was achieved by placing saturated salt solutions (LiCl
(13%), Ca(NO3)2 (50%) or NaCl (75%) (Winston and Bates 1960) in sealed
polyethylene boxes (27 x 19.5 x 10 cm). After moisture content adjustment, mesh bags
containing seeds were hermetically sealed in laminated aluminium foil bags to maintain
their moisture content and transferred to 45oC for storage. Separate aluminium foil bags
were used for each of six storage periods (1, 3, 6, 9, 12 and 18 months). Each foil bag
contained seeds of both Thryptomene baeckeacea and Zygophyllum fruticulosum for
germination testing, as well as extra seeds of Thryptomene baeckeacea on which seed
moisture content was assessed at the end of each storage period to ensure that it had not
changed during storage. Foil bags were not opened until the end of each storage period.
Germination was tested prior to storage and after each of the six storage periods using
four replicates of 25 seeds (Zygophyllum fruticulosum) or 100 seeds (Thryptomene
baeckeacea).
For the second part of the experiment, a greater suite of species (Acanthocarpus preissii,
Anthocercis littorea, Dioscorea hastifolia, Eremophila oldfieldii, Thryptomene
baeckeacea and Zygophyllum fruticulosum) were collected on the Edel Peninsula in
Shark Bay, Western Australia (S 26o 07‘ 53‘‘ E 113
o 22‘ 58‘‘) in October 2006, cleaned
and sealed in nylon mesh bags and in January 2007 equilibrated at each of two RHs
(13% and 50%) for three weeks, as described above. Seeds in mesh bags were then

56
hermetically sealed in aluminium foil bags and stored at 30oC or 45
oC. Separate
aluminium foil bags were used for each of the two equilibrium RHs, two storage
temperatures and five storage periods (1, 3, 6, 9, and 12 months). Each foil bag
contained seeds of all species (for germination testing) as well as seeds of a test species
(Anigozanthos manglesii D.Don), on which seed moisture content was assessed at the
end of the storage period to ensure that moisture content had not changed during
storage. Foil bags were not opened until the end of the storage period. Germination
testing intervals were based on seed availability. Germination of all species was tested
prior to storage and after 1, 3, 6 and 9 months (Anthocercis littorea and Dioscorea
hastifolia), 3, 6 and 12 months (Acanthocarpus preissii and Zygophyllum fruticulosum),
1, 3, 6 and 12 months (Eremophila oldfieldii) or 6 and 12 months (Thryptomene
baeckeacea) of storage. Four replicates of 20 seeds were used in all treatments, except
for Eremophila oldfieldii (four replicates of 20 indehiscent fruits) and Thryptomene
baeckeacea (four replicates of 90 seeds). A high number of seeds were used per
replicate for Thryptomene baeckeacea due to low seed viability, which was 16% as
determined using a cut test. For this test, imbibed seeds were cut in half and seeds with
firm, white embryos were considered viable whilst empty seeds, or those with shrivelled
or brown/black embryos, were non-viable (Baskin and Baskin 1998). Fruits of
Eremophila oldfieldii also have low seed fill, so prior to experimentation, Eremophila
oldfieldii fruits were x-rayed using a Faxitron Specimen Radiography System (MX-20
Cabinet X-ray Unit) to ensure that each fruit contained at least one seed. Indehiscent
fruits of Eremophila oldfieldii are hereafter referred to as seeds. Seed viability of the
remaining species was >90% (Turner et al. 2006b; chapter 2).
After equilibration and prior to storage, seed moisture content of all species was
measured gravimetrically (g H2O g-1
DW) by drying three replicates of > 0.1 g seeds (or
at least 10 seeds) at 103oC for 17 h (ISTA 2007). After each storage period, seeds were
removed from the foil bags, and moisture content was assessed gravimetrically on seeds
of a test species (either Thryptomene baeckeacea (2006) or Anigozanthos manglesii
(2007)) to ensure that foil bags remained hermetically sealed throughout the storage
period. Seed moisture content was only tested for Thryptomene baeckeacea in 2006 and
Anigozanthos manglesii in 2007, as there were insufficient seeds to test each of the
other species individually.

57
3.3.2 Seed germination testing prior to and after storage
To assess germination following each period of after-ripening, seeds were soaked for 24
h in either water (H20), 2.89 mM gibberellic acid (GA3) or 0.67 μM karrikinolide
(KAR1 - the butenolide 3-methyl-2H-furo[2,3-c]pyran-2-one, synthesised as described
by Flematti et al. (2005)). Seeds were then surface sterilised in 2% (w/v) calcium
hypochlorite (Ca(OCl)2) for 30 min, then rinsed three times with sterilised deionised
water and placed on 0.7% (w/v) water agar in 90 mm plastic Petri dishes. Seeds were
incubated under a 12/12 h alternating temperature regime of 26/13oC (determined as
optimal for several spp. in this region (chapter 2)). Germination was defined as the
emergence of the radicle, and was assessed regularly until no further germination was
observed. Germination rate was determined using time to 50% of the final germination
(t50), which was interpolated from cumulative germination curves of each replicate.
When germination of a particular treatment was low, viability of ungerminated seeds
was assessed using tetrazolium chloride (Moore 1972), whereby seeds were cut in half
and placed cut side down on germination test paper irrigated with 1% tetrazolium
chloride buffered to pH 7 with a phosphate buffer (KH2PO4 and Na2HPO4).
3.3.3 Statistical analysis
Germination percentages were arcsine transformed prior to analysis. Data was analysed
by analysis of variance (ANOVA) (P=0.05) using Genstat 8.1 (Copyright 2005, Lawes
Agricultural Trust). If significant differences were detected by ANOVA, Fishers LSD
was used to determine treatment differences. Untransformed data appears in all figures
and tables. Viability adjusted germination (VAG) is presented for Thryptomene
baeckeacea and was determined using the following formula (Sweedman and Merritt
2006): VAG = (germination (%) / viability (%)) x 100.
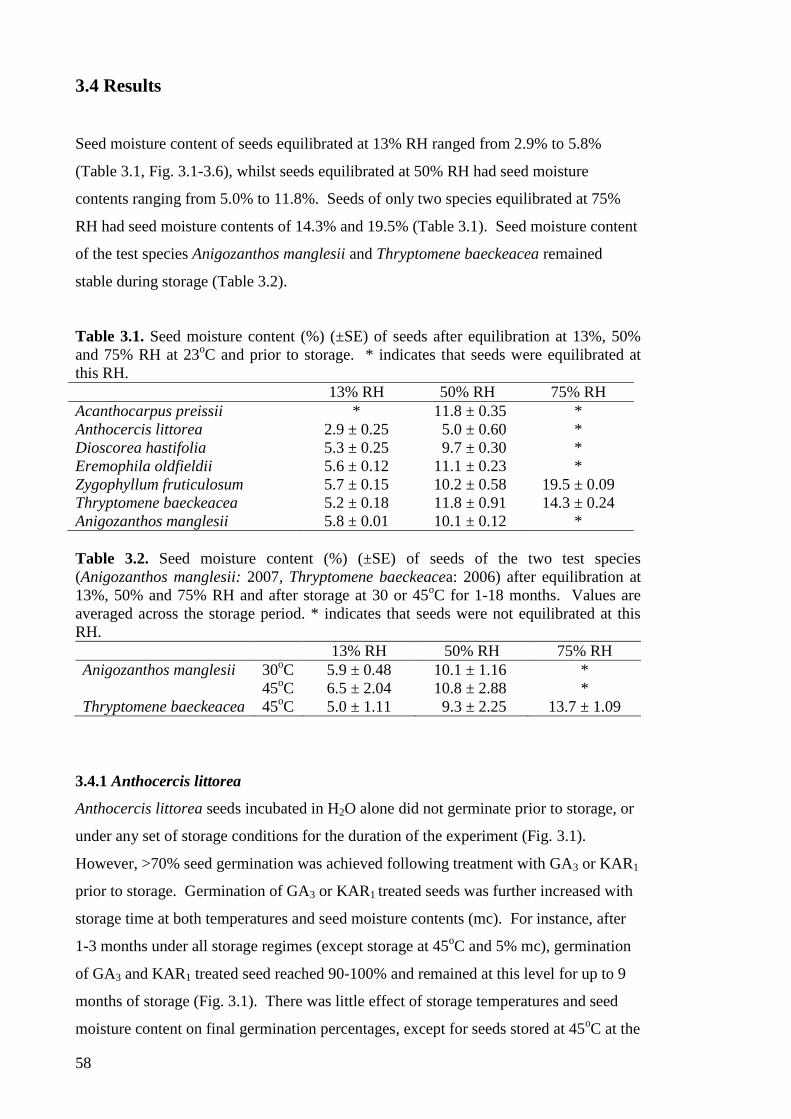
58
3.4 Results
Seed moisture content of seeds equilibrated at 13% RH ranged from 2.9% to 5.8%
(Table 3.1, Fig. 3.1-3.6), whilst seeds equilibrated at 50% RH had seed moisture
contents ranging from 5.0% to 11.8%. Seeds of only two species equilibrated at 75%
RH had seed moisture contents of 14.3% and 19.5% (Table 3.1). Seed moisture content
of the test species Anigozanthos manglesii and Thryptomene baeckeacea remained
stable during storage (Table 3.2).
Table 3.1. Seed moisture content (%) (±SE) of seeds after equilibration at 13%, 50%
and 75% RH at 23oC and prior to storage. * indicates that seeds were equilibrated at
this RH.
13% RH 50% RH 75% RH
Acanthocarpus preissii * 11.8 ± 0.35 *
Anthocercis littorea 2.9 ± 0.25 5.0 ± 0.60 *
Dioscorea hastifolia 5.3 ± 0.25 9.7 ± 0.30 *
Eremophila oldfieldii 5.6 ± 0.12 11.1 ± 0.23 *
Zygophyllum fruticulosum 5.7 ± 0.15 10.2 ± 0.58 19.5 ± 0.09
Thryptomene baeckeacea 5.2 ± 0.18 11.8 ± 0.91 14.3 ± 0.24
Anigozanthos manglesii 5.8 ± 0.01 10.1 ± 0.12 *
Table 3.2. Seed moisture content (%) (±SE) of seeds of the two test species
(Anigozanthos manglesii: 2007, Thryptomene baeckeacea: 2006) after equilibration at
13%, 50% and 75% RH and after storage at 30 or 45oC for 1-18 months. Values are
averaged across the storage period. * indicates that seeds were not equilibrated at this
RH.
13% RH 50% RH 75% RH
Anigozanthos manglesii 30oC 5.9 ± 0.48 10.1 ± 1.16 *
45oC 6.5 ± 2.04 10.8 ± 2.88 *
Thryptomene baeckeacea 45oC 5.0 ± 1.11 9.3 ± 2.25 13.7 ± 1.09
3.4.1 Anthocercis littorea
Anthocercis littorea seeds incubated in H2O alone did not germinate prior to storage, or
under any set of storage conditions for the duration of the experiment (Fig. 3.1).
However, >70% seed germination was achieved following treatment with GA3 or KAR1
prior to storage. Germination of GA3 or KAR1 treated seeds was further increased with
storage time at both temperatures and seed moisture contents (mc). For instance, after
1-3 months under all storage regimes (except storage at 45oC and 5% mc), germination
of GA3 and KAR1 treated seed reached 90-100% and remained at this level for up to 9
months of storage (Fig. 3.1). There was little effect of storage temperatures and seed
moisture content on final germination percentages, except for seeds stored at 45oC at the

59
higher seed moisture content (5% mc), which had lost viability after 6 months of storage
(Fig. 3.1). Also, there was little difference between germination of GA3 and KAR1
treated seeds within each combination of temperature and seed moisture content, except
for seeds stored at 30oC and 5% mc where germination of GA3 treated seeds was
significantly higher (P<0.05) than KAR1 treated seeds over the first 9 months of storage.
Germination rate increased with storage as indicated by a decrease in the time to 50%
germination, t50, from 18 d to 8-15 d after 1-3 months of storage (Fig. 3.1). The rate
then remained similar from 3-9 months of storage. There was some effect of storage
conditions on t50. After 3 months, seeds stored at 45oC at the higher moisture content
had significantly slower (P<0.05) germination, compared with the other storage
conditions. After 6 and 9 months, germination rate of seeds stored at 45oC and 2.9% mc
> seeds stored at 30oC and 5.0% mc, > seeds stored at 30
oC and 2.9% mc. Seeds treated
with GA3 and KAR1 generally had similar germination rates.
30oC, 2.9% mc
Germ
ination (
%)
0
20
40
60
80
10030
oC, 5.0% mc 45
oC, 5.0% mc45
oC, 2.9% mc
0 3 6 9
t 50 (
d)
0
5
10
15
20
Duration of storage (months)
0 3 6 9 0 3 6 9 0 3 6 9
H2O
GA3
KAR1
Figure 3.1. Percent germination (mean ± se) and time to 50% germination (t50) (days)
(mean ± se) of Anthocercis littorea seeds firstly stored at 30oC or 45
oC and 2.9%
moisture content (mc) or 5.0% mc for up to 9 months, then treated with water (H2O),
2.89 mM gibberellic acid (GA3) or 0.67 µM karrikinolide (KAR1) and incubated in Petri
dishes at 26/13oC.

60
3.4.2 Dioscorea hastifolia
Prior to storage, seeds of Dioscorea hastifolia exhibited 59-66% germination, with
germination increasing over 1-3 months of storage to a maximum of 97%. In general,
storage at higher temperature or higher seed moisture content increased germination
(Fig. 3.2), although, at the highest combination of temperature and seed moisture
content, viability (as assessed by tetrazolium chloride) was lost after 3 months. For
example, when incubated in H2O, seeds stored for 1 month at 45oC had higher
germination (P<0.05) than seeds stored at 30oC; and for seeds stored for 1-6 months at
30oC, the higher seed moisture content (9.7% mc) resulted in significantly greater
(P<0.05) germination than the lower seed moisture content (5.3% mc). Differences
between storage temperature and seed moisture content became less pronounced as
storage time increased. Differences between H2O, GA3 and KAR1 treated seeds were
minor, although the germination-promoting effect of GA3 was more prominent after a
short period (1 month) of storage at 30oC.
The germination rate of Dioscorea hastifolia seeds was affected by storage time and
conditions (Fig. 3.2). Storage time increased germination rate, with t50 of seeds prior to
storage ≥ 34 days, whereas t50 of stored seeds (45oC and 5.3% mc for 9 months) was 6-7
days. Germination rate increased with storage temperature, as after 1 month, t50 of
seeds after-ripened at 45oC (and treated with H2O and GA3) was significantly higher
(P<0.05) than those after-ripened at 30oC. Seeds stored at 30
oC with a higher moisture
content (9.7%) had a greater germination rate than seeds with a lower moisture content
(5.3%). GA3 and KAR1 had a minimal effect on germination rate in most instances (Fig.
3.2).

61
30oC, 5.3% mc
Ge
rmin
atio
n (
%)
0
20
40
60
80
100
30oC, 9.7% mc 45
oC, 5.3% mc 45
oC, 9.7% mc
0 3 6 9
t 50
(d
)
0
10
20
30
40
Duration of storage (months)
0 3 6 9 0 3 6 9 0 3 6 9
H2O
GA3
KAR1
Figure 3.2. Percent germination (mean ± se) and time to 50% germination (t50) (days)
(mean ± se) of Dioscorea hastifolia seeds firstly stored at 30oC or 45
oC and 5.3%
moisture content (mc) or 9.7% mc for up to 9 months, then treated with water (H2O),
2.89 mM gibberellic acid (GA3) or 0.67 µM karrikinolide (KAR1) and incubated in Petri
dishes at 26/13oC.
3.4.3 Zygophyllum fruticulosum
Germination of Zygophyllum fruticulosum seeds was only 16-28% prior to storage (Fig.
3.3). Similar to Anthocercis littorea and Dioscorea hastifolia, various combinations of
storage treatments and germination promoting compounds were able to increase
germination. Following 12 months storage at 30oC and 5.7% mc, germination of seeds
treated with GA3 and KAR1 was significantly higher (P<0.05) than germination of seeds
prior to storage. Also, seeds stored at 30oC and 10.2% mc had significantly higher
(P<0.05) germination after 3 months (KAR1 treated seeds only), 6 months (GA3 and
KAR1) and 12 months storage (KAR1 only), compared with germination of seeds prior
to storage. Although germination was not optimised after 12 months storage
(germination ≤55%), a tetrazolium test confirmed that seeds stored at 30oC and both
moisture contents had maintained constant levels of viability during storage.
Unlike storage at 30oC, storage at 45
oC increased germination of seeds treated with H2O
(Fig. 3.3). Seeds stored at 45oC and 5.7% mc for 3 and 9 months had significantly
higher (P<0.05) germination than seed prior to storage. In addition, treatment of seeds
with germination promoting compounds also resulted in increased germination after
most storage times (except 6 (KAR1 treated seeds) and 18 months (GA3 and KAR1
treated seeds)). Although germination of all seeds stored at 45oC and 5.7% mc was low
(5-11%) after 18 months, seeds maintained viability (as confirmed by a tetrazolium

62
test). Seeds stored at 45oC and 10.2% mc for 1-3 months exhibited significantly higher
(P<0.05) germination than seeds prior to storage (when treated with H2O, GA3 and
KAR1), however, after 6 months, seeds failed to germinate as they had lost viability, as
had seeds pre-equilibrated at 75% RH and stored at 45oC for 1 month (data not shown).
The germination rate of Zygophyllum fruticulosum seeds increased marginally following
storage at 30oC (t50 was 14-22 d prior to storage and 5-20 d after storage), but
differences were disparate amongst treatments and storage periods (data not shown). In
contrast, germination rate did not increase following storage at 45oC.
30oC, 5.7% mc
0 6 12 18
Ge
rmin
ation (
%)
0
20
40
60
80
10030
oC, 10.2% mc
0 6 12 18
45oC, 5.7% mc
Duration in storage (months)
0 6 12 18
45oC, 10.2% mc
0 6 12 18
H2O
GA1
KAR1
Figure 3.3. Percent germination (mean ± se) of Zygophyllum fruticulosum seeds firstly
stored 30oC or 45
oC and 5.7% moisture content (mc) or 10.2% mc for up to 18 months,
then treated with water (H2O), 2.89 mM gibberellic acid (GA3) or 0.67 µM karrikinolide
(KAR1) and incubated in Petri dishes at 26/13oC.
3.4.4 Acanthocarpus preissii
Seeds of Acanthocarpus preissii exhibited low germination (≤22%) prior to storage
(Fig. 3.4). While there was a significant effect of storage time on germination (P<0.05),
few treatments increased germination above that of seeds prior to storage. The only
conditions that increased germination, relative to seeds prior to storage, were seeds
stored for 3 months (and treated with KAR1) and 6 months (treated with H2O and
KAR1). Overall, there were few differences in germination between the germination
promoters GA3 and KAR1 and H2O treated seeds. Prior to storage, t50 ranged from 25-
47 d (data not shown) and after storage t50 ranged from 18-30 d. Storage time had a
significant effect on t50 (P<0.05) with 3-12 months storage decreasing t50 (for seeds
treated with H2O) compared with prior to storage. GA3 and KAR1 also affected t50
(P<0.05) (data not shown).

63
30oC, 11.8% mc
Duration of storage (months)
0 3 6 9 12
Ge
rmin
atio
n (
%)
0
20
40
60
80
100H2O
GA3
KAR1
Figure 3.4. Percent germination (mean ± se) of Acanthocarpus preissii seeds firstly
stored at 30oC and 11.8% moisture content (mc) for up to 12 months, then treated with
water (H2O), 2.89 mM gibberellic acid (GA3) or 0.67 µM karrikinolide (KAR1) and
incubated in Petri dishes at 26/13oC.
3.4.5 Thryptomene baeckeacea
Germination of Thryptomene baeckeacea seeds was <26% prior to storage (Fig. 3.5)
and there was little effect of storage or germination promoting compounds on
germination. For seeds stored at 30oC and 11.8% mc, it was only after 6 months that
germination was greater than prior to storage (for seeds treated with H2O). Seeds stored
at 45oC and 5.2% mc for 12 (GA3 treated) or 18 months (GA3 and KAR1 treated) had
higher germination than seeds prior to storage, as did seeds stored at 45oC and 11.8%
mc for 1 month (KAR1 treated). Seeds stored at 45oC failed to germinate after 3 months
(14.3% mc) or 9 months (11.8% mc). Storage had a limited effect on germination rate
(data not shown), as t50 ranged from 50-70 d for seed prior to storage, to 30-69 d
following storage.
45oC, 5.2% mc
0 6 12 18
45oC, 11.8% mc
Duration of storage (months)
0 6 12 18
45oC, 14.3% mc
0 6 12 18
H2O
GA3
KAR1
30oC, 11.8% mc
0 6 12 18
Ge
rmin
atio
n (
%)
0
10
20
30
40
50
Figure 3.5. Percent germination (mean ± se) of Thryptomene baeckeacea seeds firstly
stored at 30oC and 11.8% moisture content (mc) or 45
oC and 5.2% mc, 11.8% mc or
14.3% mc for up to 18 months, then treated with water (H2O), 2.89 mM gibberellic acid
(GA3) or 0.67 µM karrikinolide (KAR1) and incubated in Petri dishes at 26/13oC.

64
3.4.6 Eremophila oldfieldii
Prior to storage, germination of Eremophila oldfieldii seeds was <15%, with seeds not
responding to storage treatments (Fig. 3.6). There was no difference between
germination of seeds prior to storage and those stored from 3-12 months at 30oC and
5.6% mc. For seeds stored at 30oC and 11.1% mc, treatment with KAR1 after 3 months
storage was the only treatment able to increase germination, relative to seeds prior to
storage. Germination rate also did not differ following storage, with t50 of seeds prior to
storage ranging from 23-45 d and t50 of seed after storage ranging from 15-52d (data not
shown).
30oC, 5.6% mc
0 3 6 9 12
Germ
ination (
%)
0
10
20
30
40
5030
oC, 11.1% mc
Duration of storage (months)
0 3 6 9 12
H2O
GA3
KAR1
Figure 3.6. Percent germination (mean ± se) of Eremophila oldfieldii seeds firstly
stored at 30oC and 5.6% moisture content (mc) or 11.1% mc for up to 12 months, then
treated with water (H2O), 2.89 mM gibberellic acid (GA3) or 0.67 µM karrikinolide
(KAR1) and incubated in Petri dishes at 26/13oC.

65
3.5 Discussion
Dormancy loss of Anthocercis littorea, Dioscorea hastifolia and Zygophyllum
fruticulosum via storage (after-ripening) was promoted after one or more months at
various combinations of temperature and seed moisture content. In contrast, storage
under a range of conditions had limited effect on germination of Acanthocarpus preissii,
Eremophila oldfieldii and Thryptomene baeckeacea, implying that these seeds may not
respond to after-ripening. Whilst after-ripening is known to increase germination of
many Australian plant species (Bell 1999; Plummer et al. 2001; Schütz et al. 2002), it
has rarely been quantified in terms of both temperature and seed moisture content. This
study indicates that manipulation of the seed storage environment can affect the rate of
after-ripening and can improve germination performance of diverse arid zone plant
species.
It is well established that increasing the storage temperature increases the rate of after-
ripening/dormancy loss of many species (Baskin and Baskin 1976; Cohn and Hughes
1981; Esashi et al. 1993; Sharif-Zadeh and Murdoch 2001; Steadman et al. 2003). In
this study, such an effect was most pronounced in Dioscorea hastifolia with seeds
exhibiting a higher germination percentage and a lower t50 after one month of after-
ripening at 45oC (at either seed moisture contents), compared with 30
oC. In addition,
whilst germination of Zygophyllum fruticulosum seeds did not increase after storage at
30oC, storage at 45
oC resulted in increased germination after 3 and 9 months of storage
(for seeds incubated in water). Although there was little effect of storage temperature
on germination percentages of Anthocercis littorea following after-ripening, it is
possible that the use of a wider range of storage temperatures may have demonstrated a
difference in the rate of dormancy loss of these seeds, as they clearly showed an after-
ripening response. For example, in a study on Avena fatua seeds, germination was
similar after 6 months of after-ripening at both 30oC and 40
oC, but germination of seeds
after-ripened at 20oC was much lower (Foley 1994).
Seed moisture content during storage also affected the germination characteristics of
Anthocercis littorea, Dioscorea hastifolia and Zygophyllum fruticulosum. For instance,
at 30oC, Dioscorea hastifolia seeds stored at a higher moisture content (9.7% mc) had a
higher germination percentage and rate compared with seeds stored at a lower moisture
content (5.3% mc) after most storage times. This result is analogous to other studies

66
reporting that increasing the seed moisture content / storage RH can increase the rate of
after-ripening (within certain limits) (Baskin and Baskin 1979; Steadman et al. 2003).
In this study, at the higher storage temperature of 45oC, effect of seed moisture content
on germination was negligible for the first month of storage. However, after 3-6
months, viability loss of seeds pre-equilibrated at 50% or 75% RH became an issue.
For instance, after three months (Dioscorea hastifolia) or six months (Anthocercis
littorea and Zygophyllum fruticulosum) of storage at 45oC, seeds pre-equilibrated at
50% RH had lost viability. In addition, seeds of Zygophyllum fruticulosum pre-
equilibrated at 75% RH and stored at 45oC for 1 month lost viability. Other studies
have also found a loss in viability of seeds after-ripened at high (≥70%) RH (Baskin and
Baskin 1979; Sharif-Zadeh and Murdoch 2001) and seed longevity is reduced by both
high seed moisture (high RH) and high temperature (Walters 1998). Hence, after-
ripening appears to occur most effectively within an optimum range of RH: too low and
seeds do not after-ripen; too high, and viability declines (Baskin and Baskin 1979). The
optimum range of seed moisture contents appears to correlate with seed moisture
contents within region 2 of water binding for two species, Lolium rigidum and
Xanthium pennsylvanicum (Esashi et al. 1993; Steadman et al. 2003) and near the
boundary between region 1 and 2 for another species, Oryza sayiva (Leopold et al.
1988).
Germination rate of Dioscorea hastifolia was slow (t50 >30 d) prior to storage, however,
whilst storage time increased the germination rate of Dioscorea hastifolia and
Anthocercis littorea, it had a minimal effect on the germination rate of Zygophyllum
fruticulosum. Other studies have also noted slow germination of Dioscorea species.
Albrecht and McCarthy (2006) observed that germination of Dioscorea villosa did not
commence until after 8 weeks of incubation and Terui and Okagami (1993) observed
slow germination of Dioscorea tenuipes and Dioscorea balcanica. Both studies noted
an increase in germination rate and maximum percentage as seeds lost dormancy.
Germination rate commonly increases in concert with dormancy loss and this has also
been shown in an Australian study which observed an increase in germination rate
(decrease in time to 50% germination) as dormancy was lost following soil storage,
storage at 22-23oC or storage at diurnal fluctuation of 25/60
oC (McIvor and Howden
2000).

67
Germination promoting compounds generally had little effect on the germination of
Dioscorea hastifolia and Zygophyllum fruticulosum. In contrast, seeds of Anthocercis
littorea did not germinate unless exposed to KAR1 or GA3. A response to smoke
products is consistent with reports that Anthocercis is a post-fire ephemeral – a group of
plants that emerge after a fire, and are generally short lived, as illustrated by seed set
and senescence prior to the next fire event (Pate et al. 1985; Bell et al. 1993). Hence,
seeds of these species are stored in the soil awaiting the next fire event, and our results
suggest a possible obligate requirement for smoke to cue germination.
It is likely that Anthocercis littorea, Dioscorea hastifolia and Zygophyllum fruticulosum
exhibit physiological dormancy given that seeds imbibe water and have fully developed
linear (Anthocercis littorea and Zygophyllum fruticulosum) or capitate embryos
(Dioscorea hastifolia) (chapter 2). This corresponds with Baskin and Baskin (1998)
who concluded that three other Zygophyllum species exhibit this type of dormancy and
other members of the Dioscoreaceae are non-dormant or physiologically dormant.
Dormancy in other Dioscorea species has been overcome by cold stratification, rather
than after-ripening, as they occur in the northern hemisphere and mature in October
(autumn), and germinate in April (spring) (Albrecht and McCarthy 2006). Dioscorea
hastifolia matures in late spring and if dormancy is lost over summer, seeds may be
ready to germinate in autumn or winter, which is the period of reliable rainfall.
After-ripening had a limited effect on the germination of Acanthocarpus preissii,
Eremophila oldfieldii and Thryptomene baeckeacea. This finding contrasts with
Beardsell et al. (1993) who found germination of Thryptomene calycina increased after
dry storage. The highest germination occurred from fruits that had been soil aged for 2
years. In addition, a study on Eremophila maculata found that germination only
occurred from older seeds (22 and 34 months old), but not from younger seeds (10
month old) (Richmond and Ghisalberti 1994). Physiological dormancy of
Acanthocarpus preissii seeds has been overcome by warm stratification, which occurs
during autumn when there is intermittent rainfall and warm temperatures (Turner et al.
2006b) and such a strategy could be useful for Eremophila oldfieldii and Thryptomene
baeckeacea. It is also possible that dormancy is not overcome during the first summer
in these two species, and dormancy may be released by a combination of after-ripening
and stratification (Baskin and Baskin 2004b; Hoyle et al. 2008). In addition, there are
different levels of physiological dormancy, and while Acanthocarpus preissii exhibits

68
non-deep physiological dormancy (Turner et al. 2006b), the other two species may
exhibit intermediate or deep physiological dormancy.
In conclusion, germination of three arid zone species has been increased by after-
ripening. By using a combination of temperatures, and equilibrating seeds at various
relative humidities, this study has increased our understanding of dormancy loss in these
species through the interactions between temperature, seed moisture content and storage
time, and the effects of GA3 and KAR1. For improving germinability, this study
recommends that seeds be stored at warmer temperatures such as 45oC, however
moisture content of seeds must be taken into consideration. Storage at 45oC of seeds
equilibrated at 50% or 75% is not recommended for all species. In addition,
Anthocercis littorea should be treated with KAR1 or GA3. These findings have direct
implications for restoration in that they will enable dormancy to be overcome between
the time of seed collection (late spring) and the time when seed is needed for restoration
activities such as nursery seedling production and seed broadcasting (summer and
autumn). The findings may also be applicable to other dormant species from the arid
zone that experience similar environmental conditions.

69
C H A P T E R 4
Case study: Seed germination of Solanum spp. from
arid Australia
4.1 Abstract
Effective methods for propagation of native Solanum species are required for mine
rehabilitation and the native food industry in Australia. This study investigated seed
germination of eight native Solanum species with respect to incubation temperature and
the efficacy of germination promoting compounds gibberellic acid (GA3), the
butenolide isolated from smoke (karrikinolide, KAR1) and smoke water (SW). Seeds of
all species were tested under a temperature regime of 26/13°C or 33/18°C. In these
conditions, seeds of only two species, S. cunninghamii Benth. and S. phlomoides Benth.
germinated to high levels without treatment. Of the remaining six species, GA3 alone
promoted germination in S. chippendalei Symon, S. diversiflorum F.Muell. and S.
sturtianum F.Muell., whilst GA3, KAR1 and SW were effective at promoting
germination of S. centrale J.M.Black, S. dioicum W.Fitzg. and S. orbiculatum Dunal ex
Poir. to varying degrees. Additional incubation temperatures (10, 15, 20, 25 and 30°C)
were examined for S. centrale and S. orbiculatum. For both species, broadly similar
patterns were noted in the response of seeds to GA3, KAR1 and SW across all
temperatures. However, for S. centrale seeds, germination percentages were higher at
26/13°C than at any of the constant temperatures, and there was a trend of increasing
germination with increasing constant temperature for S. orbiculatum seeds. Analysis of
seed embryo type and imbibition characteristics and consideration of the subsequent
germination results indicates that dormant Solanum seeds possess physiological
dormancy.

70
4.2 Introduction
Solanum species occur across many ecosystems and in all continents. The genus
includes economically important food crops such as potato (Solanum tuberosum) and
eggplant (Solanum melongena). There are 47 native species of Solanum in Western
Australia and 11 naturalised species (Paczkowsa and Chapman 2000). Many of the
native species, commonly know as bush tomatoes, were used as a food source by
indigenous Australians and a number of species are in commercial production or
evaluation as bush tucker. Edible Solanum species, including S. centrale (Latz 1995;
Stefaniski 1998; Ahmed and Johnson 2000) and S. chippendalei (Courtenay pers.
comm.), are important food sources with fruits possessing high carbohydrate and
vitamin C content.
While the fruit of S. centrale and S. chippendalei can be collected from the wild,
commercial production of S. centrale is underway (Ahmed and Johnson 2000) and is
planned for S. chippendalei (Courtenay pers. comm.). However, information about
propagation is required. Also, propagation of Solanum species is required for minesite
restoration in Australia, particularly as a result of a resurgence in mining activity in the
arid zone where the genus most commonly occurs. Species required in restoration
include S. orbiculatum and S. diversiflorum as both species are common and widespread
components of the pre-mined vegetation. However, little is known about the seed
germination biology of arid zone vegetation in Australia, particularly with respect to
methods applicable to large scale propagation and restoration. Furthermore, poor seed
germination and limited horticultural information available on Solanum species are
hampering propagation and commercial production.
The two studies published on S. centrale seeds indicate that gibberellic acid (GA3) and
smoke may be useful germination promoting agents for Solanum spp. (Stefaniski 1998;
Ahmed et al. 2005). Stefaniski (1998) found gibberellic acid increased germination
from 7% to 20% while Ahmed et al. (2005) showed that a combination of seed-coat
nicking and aerosol smoke improved germination. In particular, fire related cues
warrant further investigation as disturbance by fire has been observed to encourage the
spread of Solanum species in natural ecosystems (Latz 1995). Smoke products are well
known to promote germination of a large number of Australian species (Dixon et al.
1995; Roche et al. 1997b) and the newly discovered active chemical in smoke, the

71
butenolide (3-methyl-2H-furo[2,3-c]pyran-2-one) (Flematti et al. 2004), now known as
karrikinolide (KAR1) (Dixon et al. 2008), has proved highly effective at promoting
germination of a broad range of Australian species, including arid zone species (Merritt
et al. 2006). Gibberellins are similarly known to be efficacious across a broad range of
Australian species (Bell et al. 1995; Plummer and Bell 1995) and are thought to act via
mechanisms that include promoting the growth potential of the embryo (Kucera et al.
2005), weakening endospermic cells (Groot and Karssen 1987; Groot et al. 1988;
Debeaujon and Koornneef 2000), and replacing after-ripening requirements (Baskin and
Baskin 2004b).
Optimal germination temperatures for seed germination usually correspond to the time
where water is non-limiting in the environment (Bell et al. 1993; Bell 1999; Bell et al.
1999). The distribution of the Solanum species in this study covers a range of
environmental conditions from wet summers and dry winters (Pilbara, Great Sandy
Desert and Dampierland regions), to an arid region with aseasonal rainfall (MacDonnell
Ranges in central Australia) and finally to areas that receive sporadic winter rain and
occasional summer cyclonic systems (Geraldton Sandplains and Murchison regions).
As these regions receive summer rainfall, it is likely that incubation temperatures
corresponding to the season of reliable rainfall may be higher than typically used in
nursery propagation of Australian species in southern Australia (15-20oC) (Bell 1999).
For example, Jurado and Westoby (1992) found that germination of a Solanum species
from arid Australia was higher at 28oC compared with 12
oC and 20
oC.
Therefore, the aim of this study was to develop an understanding of germination and
dormancy characteristics for an indicative range of eight Solanum species with
restoration and commercial value from the arid and semi-arid zone of Australia.
Specifically, for each species we determined (a) the seed and embryo morphology, (b)
whether seeds were permeable and able to imbibe water (via imbibition studies) and (c)
the effects and interactions of incubation temperature, gibberellic acid (GA3),
karrikinolide (KAR1) and smoke water (SW) on seed germination.

72
4.3 Materials and methods
4.3.1 Seed collection
Table 1.1 shows the collection date, location and region for the eight Solanum species
used in this study. The method of seed cleaning and storage conditions varied between
species. Following collection of fruits of S. cunninghamii, S. dioicum, S. phlomoides
and S. sturtianum, seeds were extracted from fruits and air dried and stored at -18oC
after collection. Seeds were retrieved from storage in June 2006 and used in
experiments immediately.
For S. orbiculatum, seeds were removed from freshly collected fruits using pectinase
(1%) to dissolve the fleshy fruit. Seeds were then air dried and stored at ambient
laboratory conditions (c. 22oC, 50% RH) for three months prior to use in experiments in
2005. S. orbiculatum seeds used for additional experiments at constant temperatures of
10, 15, 20, 25 and 30oC were collected in November 2005, cleaned as described above
and stored at ambient laboratory conditions for four months prior to the experiment in
2006.
Fruits of S. chippendalei and S. diversiflorum were air dried then cracked open to
remove the seeds. Seeds were stored at ambient laboratory conditions (c. 22oC, 50%
RH) after collection for three months (S. chippendalei) and six weeks (S. diversiflorum)
prior to use in experiments in 2005 and 2007 respectively.
Seeds of Solanum centrale were provided by Alice Springs Desert Park. Experiments
were undertaken in April 2007. Mature, collected fruits were hydrated in water to
soften and the seeds then separated from the pulp.
Specimens of each species were lodged at the Kings Park and Botanic Garden
Herbarium. Voucher numbers are as follows; S. centrale (LCOM4), S. chippendalei
(LCOM2), S. cunninghamii (LSWE1488), S. dioicum (LSWE1429), S. diversiflorum
(LCOM3), S. orbiculatum (LCOM1), S. phlomoides (LSWE1348), S. sturtianum
(LSWE6365).

73
Table 4.1. Collection date, location and Interim Biogeographic Regionalisation for
Australia (IBRA region) of eight Solanum species Species Collection
date
Location IBRA region
S. centrale Feb 2007 Napperby Station, north of Alice Springs
(S 23o 38‘ 51‘‘ E 133
o 51‘ 50‘‘)
Burt Plain
S. chippendalei Aug 2005 The Great Sandy Desert near Punju Njamal Great Sandy
Desert
S. cunninghamii 1993 Between Millstream and Pannawonica Pilbara
S. dioicum 1993 5.3 km on Shay Gap Road, near Marble Bar Pilbara
S. diversiflorum Feb 2007 Telfer mine
(S 21o 43‘ 26‘‘ E 122
o 12‘ 33‘‘).
Great Sandy
Desert
S. orbiculatum Nov 2004 Shark Bay Salt Lease
(S 26o 07‘ 53.7‘‘ E 113
o 22‘ 58.5‘‘)
Geraldton
Sandplains
S. phlomoides 1993 15 km south of Meekatharra Murchison
S. sturtianum 2004 Lake Carey
(S 28o 50‘ 04‘‘ E 122
o 11‘ 10‘‘)
Murchison
4.3.2 Seed and embryo characteristics, viability testing and imbibition studies
Given the consistency of seed coat colour, the colour of the seed coat of each species
was recorded from a simple observation. Seed diameter was determined for three
replicates of 10 seeds. Seed weight was determined by weighing three replicates of 100
seeds and multiplied by 10 to estimate 1000 seed weight. A cut test was used to
estimate the viability of the seeds prior to germination experiments. Three replicates of
20 imbibed seeds were cut in half and inspected for healthy embryonic tissue. Firm,
white embryos were considered viable and shrivelled or black embryos were considered
non-viable. Results of the cut test were confirmed by using tetrazolium chloride (Moore
1972) whereby seeds were cut in half and placed cut side down on germination test
paper irrigated with 1% tetrazolium chloride buffered to pH 7 with a phosphate buffer
(KH2PO4 and Na2HPO4). The embryos of dissected seeds were examined and classified
according to Martin (1946) and described as fully developed or underdeveloped (Baskin
and Baskin 2004c).
For each species three replicates of ≥0.03 g of seeds were weighed, placed on moist
germination test paper in Petri dishes for five minutes, patted dry with paper towel to
absorb water on the seed surface, then re-weighed. Seeds were returned to the moist
germination test paper and each replicate was weighed again after 2, 4, 6, 24, 48, 72 and
96 h. Seeds were kept at ambient laboratory conditions (c. 22oC, 50% RH) for the
duration of the experiment. Percent water uptake was determined gravimetrically.

74
4.3.3 Germination
Seeds of all species were soaked for 24 h in solutions of 2.89 mM gibberellic acid
(GA3) (Sigma Aldrich, Castle Hill, Australia, 90% GA3), smoke water (SW) (1:10 v/v),
0.67 μM karrikinolide (KAR1) (the butenolide, 3-methyl-2H-furo[2,3-c]pyran-2-one) or
deionised water (control). SW was prepared with straw using the process described by
Dixon et al. (1995). KAR1 was synthesised in pure form as described in Flematti et al.
(2005). After soaking, seeds were surface sterilised in 2% (w/v) calcium hypochlorite
(Ca(OCl)2) for 30 mins, then rinsed three times with sterilised deionised water.
Afterwards, four replicates of 25 seeds were placed in plastic Petri dishes (90mm) on
water agar (0.7% w/v) and incubated at a 12/12 h alternating temperature regime of
33/18oC or 26/13
oC. These two temperatures approximate summer and winter
temperatures in the arid environment of Western Australia where these plants
commonly occur. In addition, three replicates of 10 seeds of all species were nicked by
removing the portions of seed coat and endosperm covering the radicle tip. Nicked
seeds were then incubated only at 33/18oC as described above.
In a second germination experiment, additional incubation temperatures of 10, 15, 20,
25 and 30oC were examined for S. orbiculatum and S. centrale seeds, but could not be
performed on the other species due to limited seed numbers. For all experiments, Petri
dishes were sealed with plastic (food grade cling film), then wrapped in aluminium foil
to exclude light. Foil was removed each time germination was recorded in the
laboratory under ambient light conditions. Germination of intact seeds was defined as
the emergence of the radicle and germination of nicked seeds was defined as the
elongation of the radicle tip, the production of root hairs and subsequent development
into a normal seedling. Germination was assessed five days a week for two weeks, then
weekly until germination had ceased. Final percentage germination data are presented
for the first experiment, and both final percentage germination and time to 50% of the
final germination data are presented for the second experiment.
4.3.4 Statistical analysis
Germination percentages were arcsine transformed prior to analysis. Data analysis was
performed on individual species to determine temperature and treatment differences
however, data from germination of nicked seeds were not included in this analysis.
Germination data were analysed by analysis of variance (ANOVA) (P=0.05) using
Genstat 8.1 (Copyright 2005, Lawes Agricultural Trust, Rothamstead Experimental

75
Station, UK). If significant differences were detected by ANOVA, Fishers LSD was
used to determine treatment differences. Due to missing values, the control treatment
was not included in the analysis of time to 50% germination of S. centrale.

76
4.4 Results
4.4.1 Seed and embryo characteristics, viability testing and imbibition studies
Four species had dark (black/dark brown) seed coats including the larger massed species
S. chippendalei, S. diversiflorum and S. sturtianum and the remaining four had light
(white/cream) seed coats (Table 4.2). Seed diameter ranged from 2.1-4.7 mm. Seed
viability was generally high with the three lower massed species exhibiting 100%
viability. S. chippendalei had the lowest viability at 73% (Table 4.2). The seeds of all
eight species were endospermic and contained curved linear embryos. The curved
embryo was longer than the seed and was fully developed. Seeds of all species readily
imbibed water (Fig. 4.1). Increase in seed mass due to water uptake over 48 h ranged
from 17% (S. dioicum) to 46% (S. chippendalei).
Table 4.2. Seed coat colour, seed diameter, seed weight and viability (Mean ± SE) of
eight Solanum species.
Species Seed coat
colour
Seed diameter
(mm)
Weight of
1000 seeds (g)
Viability (%)
S. centrale Light 2.8 ± 0.04 2.4 ± 0.02 88 ± 5%
S. chippendalei Dark 4.7 ± 0.06 7.8 ± 0.10 73 ± 3%
S. cunninghamii Light 2.1 ± 0.02 1.1 ± 0.01 100 ± 0%
S. dioicum Dark 2.1 ± 0.04 1.4 ± 0.02 100 ± 0%
S. diversiflorum Dark 4.0 ± 0.01 8.1 ± 0.01 96 ± 3%
S. orbiculatum Light 2.9 ± 0.04 2.2 ± 0.03 95 ± 3%
S. phlomoides Light 2.3 ± 0.04 1.4 ± 0.02 100 ± 0%
S. sturtianum Dark 3.0 ± 0.03 4.0 ± 0.00 78 ± 2%

77
b) S. chippendalei
c) S. cunninghamii
Incre
ase
in
ma
ss (
%)
0
10
20
30
40
50d) S. dioicum
f) S. orbiculatum
g) S. phlomoides
Time (h)
0 12 24 36 48
Incre
ase
in
ma
ss (
%)
0
10
20
30
40
50h) S. sturtianum
Time (h)
0 12 24 36 48
e) S. diversiflorum
Incre
ase
in
ma
ss (
%)
0
10
20
30
40
50
a) S.centrale
Incre
ase
in
ma
ss (
%)
0
10
20
30
40
50
Figure 4.1. Imbibition (% water uptake) of eight Solanum species over 48 h at room
temperature (ca. 22oC) a) Solanum centrale, b) S. chippendalei, c) S. cunninghamii, d)
S. dioicum, e) S. diversiflorum, f) S. orbiculatum, g) S. phlomoides, and h) S.
sturtianum. Bars indicate standard error.

78
4.4.2 Germination
Whilst untreated (control) seeds of S. cunninghamii and S. phlomoides had less than
20% germination when incubated at 26/13oC, germination was 97% and 62%
respectively when incubated at 33/18oC (Fig. 4.2c, g). In contrast, untreated seeds of S.
centrale, S. dioicum and S. orbiculatum had only 1–27% germination at both 26/13oC
and 33/18oC (Fig. 4.2a, d, f). Seeds of S. diversiflorum did not germinate at 33/18
oC,
but demonstrated 2% germination when incubated at 26/13oC (Fig. 4.2e). Untreated
seeds of S. chippendalei and S. sturtianum failed to germinate at either temperature (Fig.
4.2b,h).
Treatment of seeds of all species with GA3 significantly increased (P<0.05)
germination, compared with the controls, at either one or both temperature regimes (Fig.
4.2). GA3 promoted germination of S. cunninghamii at 26/13oC, but when incubated at
33/18oC germination of both control and GA3 treated seeds was >95% (Fig. 4.2c). GA3
significantly increased (P<0.05) germination of S. phlomoides at 26/13oC, but
suppressed germination at 33/18oC (Fig. 4.2g). For the other six species, GA3
significantly increased (P<0.05) germination at both 26/13oC and 33/18
oC (Fig. 4.2a, b,
d, e, f, h). For most species germination of GA3 treated seeds was similar at both
temperatures, although S. orbiculatum seeds germinated to a higher percentage at
33/18oC than at 26/13
oC (P<0.05) (Fig. 4.2f) and S. centrale germinated to a higher
percentage at 26/13oC than at 33/18
oC (P<0.05) (Fig. 4.2a).
Unlike GA3, SW promoted germination in some, but not all species. SW significantly
increased (P<0.05) germination of S. centrale, S. dioicum and S. orbiculatum relative to
the control at both temperature regimes (Fig. 4.2a, d, f). For seeds of S. cunninghamii,
SW increased germination at 26/13oC but suppressed it at 33/18
oC (Fig. 4.2c).

79
b) S. chippendalei
c) S. cunninghamii
Germ
ination (
%)
0
20
40
60
80
100d) S. dioicum
f) S. orbiculatum
26/13oC
33/18oC
g) S. phlomoides
Control GA SW KAR Nicked
Germ
ination (
%)
0
20
40
60
80
100h) S. sturtianum
Control GA SW KAR Nicked
e) S. diversiflorum
Germ
ination (
%)
0
20
40
60
80
100
a) S. centrale
Germ
ination (
%)
0
20
40
60
80
100
Figure 4.2. Mean (± SE) germination (radicle emergence) of a) Solanum centrale, b) S.
chippendalei, c) S. cunninghamii, d) S. dioicum, e) S. diversiflorum, f) S. orbiculatum,
g) S. phlomoides, and h) S. sturtianum. Seeds were soaked for 24 h in water (Control),
in gibberellic acid (GA), in smoke water (SW), karrikinolide (KAR) or nicked, and
incubated at 12/12h alternating temperature regime of 26/13oC or 33/18
oC (Nicked
treatment only incubated at 33/18oC).

80
For S. phlomoides seeds, SW did not affect germination at 26/13oC, but suppressed
germination at 33/18oC (Fig. 4.2g). For the remaining three species S. chippendalei, S.
diversiflorum and S. sturtianum, germination of SW treated seeds was negligible (Fig.
4.2b, e, h).
Karrikinolide elicited higher germination than control seeds for five species at one or
both incubation temperatures (P<0.05). KAR1 increased germination of S. dioicum and
S. orbiculatum to at least the same level as GA3 and SW at both incubation temperatures
(Fig. 4.2d, f). For S. centrale seeds, germination of KAR1 treated seeds exceeded that
of control and SW treated seeds at both incubation temperatures (Fig. 4.2a).
Germination of S. cunninghamii and S. phlomoides was promoted by KAR1 at 26/13oC
but not at 33/18oC (Fig. 4.2c, g). For the remaining three species (S. chippendalei, S.
diversiflorum and S. sturtianum) germination in the presence of KAR1 was <5% (Fig.
4.2b, e, h). Coincidently, these three species (which also did not respond to SW) all had
dark seed coats, and had larger seeds (1000 seeds ≥4.0 g) compared with the other five
species (1000 seeds ≤2.4 g) (Table 2).
Nicking seeds did not elicit germination of S. chippendalei, S. diversiflorum or S.
sturtianum (Fig. 4.2b, e, h). Nicking seeds of S. centrale, S. dioicum and S. orbiculatum
increased germination relative to the control, and to similar levels as seeds treated with
GA3, SW or KAR1 (Fig. 4.2a, d, f). Nicked seeds of S. cunninghamii germinated to the
same percent as control seeds but those of S. phlomoides germinated to only half the
percentage of control seeds (Fig. 4.2c, g).
Additional experiments were undertaken on S. centrale and S. orbiculatum to examine
the effects of incubation temperature in greater detail. As in the first experiment,
control germination of S. centrale seeds was very low (<2%) across all incubation
temperatures. Germination of seeds treated with GA3 was high (81-99%) between 10-
25oC, but lower at 30
oC (65%) (Fig. 4.3a). Similarly, germination of seeds treated with
SW and KAR1 was slightly higher at 10, 15 and 20oC (7-35%), compared with at 25 and
30oC (<5%) (P<0.05). Germination of seeds treated with KAR1 was lower at the
constant incubation temperatures compared with the alternating temperatures of 26/13
and 33/18oC (63-84%) (P<0.05). Although germination of GA3, SW and KAR1 treated
seeds of S. centrale incubated at 10oC was significantly higher (P<0.05) than at 30
oC,
time to 50% germination was much longer (Fig. 4.3c). At 10oC, time to 50%

81
germination was around 22-24 days, compared with 2-6 days at 30oC. Time to 50%
germination did not differ from 15 to 30oC.
For S. orbiculatum, germination percentage of control seeds increased as the
temperature increased (Fig. 4.3b). All treatments significantly increased germination
(P<0.05) relative to the control at each temperature. Germination of GA3 and SW
treated seeds was higher at 20, 25 and 30oC compared with 10 and 15
oC (P<0.05),
whereas KAR1 treated seeds had high germination (90-98%) across all temperatures.
These treatments also increased the rate of germination (P<0.05) (i.e. decreased the
time to 50% germination) compared with the control at all temperatures (Fig. 4.3d). In
addition, the time to 50% germination decreased as the incubation temperature
increased, with the fastest germination observed at 20, 25 and 30oC (Fig. 4.3d).
Temperature (oC)
10 15 20 25 30
Ge
rmin
atio
n (
%)
0
20
40
60
80
100
Control
GA
SW
KAR
Temperature (oC)
10 15 20 25 30
Tim
e (
da
ys)
0
5
10
15
20
25
30c) d)
a) b)
Figure 4.3. Mean (± SE) germination of a) Solanum centrale and b) S. orbiculatum and
time to 50% of the final germination of c) S. centrale and d) S. orbiculatum seeds
treated with water (control), gibberellic acid (GA), smoke water (SW) and karrikinolide
(KAR) and incubated at constant temperatures of 10, 15, 20, 25 and 30oC.

82
4.5 Discussion
Germination was increased in all Solanum species at one or both incubation
temperatures using germination-promoting compounds and these results provide some
direction for more efficient methods for rehabilitation and commercial production. The
degree to which each compound was effective varied somewhat between species,
probably due to differing germination and dormancy characteristics and different seed
ages and storage histories. Some species germinated without treatment, whereas
germination in others was stimulated by SW, KAR1 or GA3. Firstly, germination of
untreated seeds of two species (S. cunninghamii and S. phlomoides) was moderate to
high at the incubation temperature 33/18oC. It is possible that these two species are
either non-dormant or they may have after ripened between collection and storage (the
time and conditions between collection and storage are unknown), hence dormancy may
have been partly or fully overcome. Secondly, species that exhibited little or no
germination of untreated seeds (S. centrale, S. chippendalei, S. dioicum, S.
diversiflorum, S. orbiculatum and S. sturtianum) could be considered dormant (i.e. do
not germinate within a period of time (30 days) when provided with normal physical
environmental factors (Baskin and Baskin 2004c)). However, species where control
germination was low, but germination of SW or KAR1 treated seeds was high (S.
centrale, S. dioicum and S. orbiculatum), may not be dormant, if smoke products are
considered as agents that promote germination independently of dormancy status as
suggested by some studies (Baker et al. 2005b; Merritt et al. 2007; Rokich and Dixon
2007). For the three species where germination of control, SW and KAR1 treated seeds
of S. chippendalei, S. diversiflorum and S. sturtianum was low or zero, but germination
was promoted by GA3, the presence of dormancy is likely, although this can not be
concluded absolutely as germination was tested over limited temperature conditions and
seed age varied.
If seeds are dormant, it is useful to know what type of dormancy they exhibit.
Imbibition studies indicated that seeds of all species readily take up water thus do not
exhibit physical or combinational dormancy. Observing seed morphology of all species
showed that the embryos were differentiated and fully developed indicating that the
seeds do not exhibit morphological or morphophysiological dormancy. As four classes
of dormancy have been ruled out, dormant species must therefore exhibit physiological
dormancy.

83
Germination promotion by smoke in the Australian flora is well established (Dixon et
al. 1995; Roche et al. 1997b) and the active compound in smoke, a butenolide, now
know as karrikinolide, has been recently discovered to promote germination of a range
of smoke responsive species from a wide variety of ecosystems including arid regions
(Flematti et al. 2004; Merritt et al. 2006; Stevens et al. 2007). The results of the present
study contrast with two other studies on S. centrale; one finding neither SW or aerosol
smoke effective at promoting germination (Stefaniski 1998) and the other finding
aerosol smoke only increased germination after seeds were nicked (Ahmed et al. 2005).
A difference in smoke responsiveness could be due to collection of S. centrale at
different locations and in different years. For example, Stevens et al. (2007) found a
difference in butenolide (KAR1) response of Brassica tournefortii depending on
collection year and location. In the present study both SW and KAR1 increased
germination of over half of the species (including S. centrale). Notably, germination of
KAR1 treated seeds of four species (S. centrale, S. cunninghamii, S. orbiculatum and S.
phlomoides) was higher than that of SW treated seeds at one or both incubation
temperatures. Increased germination in the presence of KAR1, as compared to SW, was
also found in a study on Australian Asteraceae (Merritt et al. 2006) and this was
explained by the presence of possible toxic compounds in SW. Similar evidence for
toxicity issues with SW have been noted by Flematti et al. (2004) who found that
undiluted SW reduced germination of Conostylis aculeata and Stylidium affine
compared with a 1 in 10 dilution.
For the three species where SW and KAR1 failed to elicit germination (S. chippendalei,
S. diversiflorum and S. sturtianum – which had dark seed coats and the largest seeds),
the seeds are either not smoke-responsive, or dormancy must be overcome before the
seeds become smoke-responsive. Seeds of two of these species were fresh when
experiments commenced, and the other had been stored for two years at -18oC,
suggesting these seeds may not have been sensitive to the smoke cue. In some studies,
freshly collected seeds have been found to be insensitive to smoke. For example, seeds
of some species are more responsive to smoke after dormancy has been released by dry
after-ripening (Tieu et al. 2001a) warm stratification (Merritt et al. 2007) or soil burial
(Tieu et al. 2001b; Baker et al. 2005b). Although germination of these three Solanum
species was not stimulated by SW or KAR1, it was stimulated by GA3. This observation
indicates that seeds of the study species exhibit physiological dormancy, as GA has
been observed to promote germination of other physiologically dormant seeds (Baskin

84
and Baskin 1998; Baskin and Baskin 2004c). However, nicking (scarification) is also
known to promote germination of seeds with non-deep physiological dormancy, as the
embryos within these seeds lack the growth potential to emerge through their covering
structures (Groot and Karssen 1987; Baskin and Baskin 1998; Baskin and Baskin
2004c). In this study, nicking did not promote germination of S. chippendalei, S.
diversiflorum and S. sturtianum suggesting that germination control is not simply via
mechanical restraint to embryo growth imposed by the seed coat. It is therefore
possible that the seeds of these three species exhibit intermediate physiological
dormancy as in these types of seeds scarification does not overcome dormancy, but GA
promotes germination (Baskin and Baskin 2004c).
Dormancy of S. centrale was recently classified by Ahmed et al. (2005). Like our
study, these authors found that germination of S. centrale seeds was promoted by
nicking. They inferred from this result that the seeds had a water impermeable seed
coat and that the species exhibited seed coat imposed dormancy. However, imbibition
was not tested to determine whether or not the seeds imbibed water prior to nicking. As
our study found all eight Solanum species readily imbibed, S. centrale seeds have a
water permeable seed coat and do not possess physical dormancy. Two recent studies
(Baskin and Baskin 2004c; Baskin et al. 2006a) have emphasised that mechanical
scarification promotes germination of both physically and physiologically dormant
seeds, and that some studies have incorrectly identified physical dormancy based on
increased germination of scarified seeds, highlighting the importance of imbibition
testing for identification of dormancy states.
Although there were some subtle differences between germination at 26/13oC and
33/18oC, for most species broadly similar responses at these two temperatures were
evident. In addition, KAR1 treated seeds of S. orbiculatum germinated to a high
percentage over the temperature range of 10oC to 30
oC. This apparent broad
temperature range for germination suggests that some Solanum species may be able to
germinate throughout the year, responding to moisture cues rather than temperature cues
(within their normal seasonal range), and enabling germination at any time during the
year (Ahmed et al. 2005). In a study on germination of central Australian plants, Jurado
and Westoby (1992) found that 30% of species tested did not show a preference for
germination temperature, although S. quadriloculatum had higher germination at 28oC
compared with 20oC and 12
oC. The range over which the Solanum species germinated

85
in this study was generally higher than that of species from the southwest of Australia
which have optimal germination between 13oC and 20
oC (Bell 1999). In addition, time
to 50% germination of S. orbiculatum decreased as the temperature increased. These
results will be important to those propagating Solanum species for restoration and
commercial production, particularly if propagation is to occur in areas outside the
normal range of the species.
In conclusion, this study has observed that SW, KAR1 and/or GA3 can promote
germination of eight Solanum species, the degree to which differs between species.
Seeds of some species may be dormant, and given that Solanum seeds have fully
developed embryos and seeds readily take up water, it is likely that dormancy is
physiological. This study also offers some insight into preferred germination
temperatures. The information about germination will be useful for propagation of
Solanum species for horticulture or restoration.

86

87
C H A P T E R 5
Assessment of vegetation in the borrow pits and
adjacent undisturbed areas
5.1 Abstract
A vegetation survey was undertaken at Shark Bay Salt (SBS) to quantify differences in
plant and species abundance, species richness, cover and frequency between
undisturbed areas on the lease, and in disturbed areas termed ‗borrow pits‘. The
decommissioned borrow pits are undergoing rehabilitation mainly through natural
migration of seed, and to a limited extent through brushing and topsoil replacement.
This study found differences between the vegetation in the undisturbed areas and the
borrow pits. Although the borrow pit that had been subject to topsoil replacement had
similar plant density and species richness compared with the undisturbed areas, it
showed a very different community structure to that of the undisturbed area. Borrow
pits without topsoil had low plant density and species richness. The reasons for limited
natural migration and the vegetation community in the topsoil replacement pit are
discussed. In addition, completion criteria and management strategies are suggested.

88
5.2 Introduction
Mining activities are generally associated with the removal of vegetation and soil
(topsoil and subsoil) (Gardner 2001). While vegetation can be returned in some
ecosystems through natural migration from vegetation that remains onsite (Hobbs and
Cramer 2007), vegetation reinstatement in most of the highly disturbed older landscapes
of the southern hemisphere requires human intervention through the return of seeds in
topsoil, the application of mulch containing canopy-stored seeds, broadcast seeding or
planting of seedlings (Rokich et al. 2002). This level of intervention is necessary due to
the loss of propagules available to rebuild the system including resprouting materials
from the soil (Grant and Loneragan 1999) and seeds from the topsoil seed bank. Seeds
are also unlikely to return via animal or bird dispersal as surrounding habitat may have
been lost. Given the difficulties associated with returning functional plant ecosystems
similar to the pre-mined natural ecosystem, vegetation in post-mined areas may differ
both in composition and structure to vegetation in pre-mined areas.
The first step in revegetating a disturbed area is to develop benchmark standards of
targets by defining the attributes of a normal, healthy ecosystem. This is achieved by
assessing the pre-disturbed vegetation community (reference sites) (Davy 2002) at least
in terms of composition and structure (e.g. dominant species, abundance and cover) and
if time permits, function (e.g. fecundity, growth, nutrient cycling, invertebrate activity).
The next step is to assess the post disturbed site to ascertain which species colonise via
natural migration. In addition, assessing sites that have been subject to rehabilitation
works (such as topsoil replacement) will determine if these works are successful in
replacing the pre-mined vegetation community. Knowledge of the vegetation
communities both in the pre- and post-mined areas will identify species that are absent
from the post-mined areas and that will therefore require re-introduction to those areas.
The aim of this study was to source reference sites and describe differences in plant and
species abundance, species richness, cover and frequency between the borrow pits and
the reference sites (the adjacent undisturbed areas) at Shark Bay Salt (SBS). In
particular, variation between disturbed areas that have been subjected to topsoil
replacement and disturbed areas subjected to natural migration were investigated. This
study identified the key perennial species that are absent from the borrow pits, and then
formed the basis of further research.

89
5.3 Methods
5.3.1 Site selection
Seven sites at SBS were chosen for the vegetation survey. The sites included four
borrow pits (named L, P, Q and R) and undisturbed vegetation adjacent to three of these
borrow pits (P, Q and R) (Appendix 1, 2). Borrow pits P (Fig. 5.1) and Q (Fig. 5.2, 5.3)
were decommissioned prior to 1993 and pit R (Fig. 5.4) was decommissioned in 1990.
The pits have been subject to some rehabilitation activities. Brushing (vegetation
prunings) from the town was spread in pit P in 1994 and seeding was undertaken in July
1996. The southern side of pit Q was cross ripped in 1996. Pit R was seeded in 1996
but the success rate was low. Topsoil was spread in pit L in 2003 (Fig. 5.5-5.10). This
topsoil came from borrow pit H, which has subsequently been turned into a pond. For
comparison, photos of the undisturbed vegetation are on pages 3, 169 and 170.
5.3.2 Site assessment
In September 2005, five quadrats of 10 x 10 m were laid out in each undisturbed site
and pit L. Due to the low numbers and patchy distribution of plants in borrow pits P, Q
and R, all of the dominant plants in each pit were recorded. Hence, there was no
replication within pits P, Q and R. Only the perennial species were recorded in all sites
as annual flora was not deemed to be important in this study, as the focus of
rehabilitation is on perennial species. At each quadrat, a GPS reading was taken (Table
5.1), the number of plants of the dominant perennial species present was recorded, and
voucher specimens were collected. Cover of perennial species was recorded using an
index of 0-5 (0 = <19% cover, 1 = 20-39% 2 = 40-59% 3 = 60-79% 4 = 80-99% 5 =
100% cover). Plant abundance (plants m-2
), species abundance (species 100 m-2
),
species richness (total number of species per site) and species frequency (the number of
quadrats per site in which the species occurs) were determined from the observations.
Plant and species abundance was determined by dividing the number of plants and
species, respectively, by the total area of each pit (1.5, 2.4 and 0.9 ha for pits P, Q and R
respectively) to achieve data per m2 for plant abundance, and per 100 m
2 for species
abundance. Plant and species abundance were analysed using ANOVA and means were
separated using Fishers LSD.
Species composition was determined by using diagrams developed using Primer
(Plymouth Marine Laboratory) that show how similar or dissimilar the vegetation is at

90
each site. Specifically, a cladogram was used to show the similarity by branching and
an ordination was used to show the sites as clusters which can be tightly or loosely
grouped, and may or may not overlap. The diagrams were based on presence/absence
data (whether individual species were present or absent from the site) and abundance
data (the number of species m-2
).
In addition, a summary of all borrow pit activities from 1993 to 2004 compiled from
SBS‘s Annual and Triennial Environmental reports (Shark Bay Salt Joint Venture 1993;
1994; 1995; 1996; 1997; 1998; 2001; 2002; 2003; 2004) is shown in Appendix 5.
Table 5.1. GPS co-ordinates at each survey location in the borrow pits (pits P, Q and R)
and the undisturbed sites (sites L, P, Q, R). For the borrow pits, the entire pit was
surveyed, so one GPS co-ordinate is listed. The total area of pits P, Q and R is 1.5, 2.4
and 0.9 ha respectively. For the undisturbed sites, five replicate surveys were
undertaken, and a GPS co-ordinate is listed for each (except site R, rep 5).
Site Rep GPS co-ordinates
Pit P 26o 09‘ 29.1‖ 113
o 23‘ 56.0‖
Pit Q 26o 10‘ 15.0‖ 113
o 23‘ 52.3‖
Pit R 26o 10‘ 37.0‖ 113
o 23‘ 58.4‖
Pit L 1 26o 08‘ 51.5‖ 113
o 23‘ 44‖
2 26o 08‘ 52.5‖ 113
o 23‘ 44.5‖
3 26o 08‘ 53‖ 113
o 23‘ 45‖
4 26o 08‘ 54‖ 113
o 23‘ 44.5‖
5 26o 08‘ 56‖ 113
o 23‘ 44.5‖
Site P 1 26o 09‘ 29‖ 113
o 23‘ 47‖
2 26o 09‘ 30.5‖ 113
o 23‘ 47‖
3 26o 09‘ 32‖ 113
o 23‘ 46.5‖
4 26o 09‘ 35‖ 113
o 23‘ 46‖
5 26o 09‘ 37‖ 113
o 23‘ 45‖
Site Q 1 26o 10‘ 18‖ 113
o 23‘ 43.5‖
2 26o 10‘ 19‖ 113
o 23‘ 42.5‖
3 26o 10‘ 21‖ 113
o 23‘ 42‖
4 26o 10‘ 23‖ 113
o 23‘ 42‖
5 26o 10‘ 24‖ 113
o 23‘ 42.5‖
Site R 1 26‘ 10‘ 45‘‘ 113‘ 23‘ 45‘‘‘
2 26‘ 10‘ 45‘‘ 113‘ 23‘ 55‘‘‘
3 26‘ 10‘ 44‘‘ 113‘ 23‘ 56‘‘‘
4 26‘ 10‘ 45‘‘ 113‘ 23‘ 54‘‘‘
5 -

91
Figure 5.1. Photograph of pit P taken in April 2005.
Figure 5.2. Aerial photo of pit Q taken in September 2004.

92
Figure 5.3. Photograph of pit Q taken in April 2005.
Figure 5.4. Photograph taken April 2005 of the grader ripping the soil in pit R.

93
Figure 5.5. Photograph taken in September 2004 of pit L, in which topsoil was
spread.
Figure 5.6. Photograph of pit L taken in June 2005.

94
Figure 5.7. Photograph of pit L taken in November 2005.
Figure 5.8. Photograph of pit L taken in May 2007.

95
Figure 5.10. Photograph (looking south) of pit L taken in October 2007.
Figure 5.9. Photograph (looking south) of pit L taken in October 2006.

96
5.4 Results
5.4.1 Vegetation characteristics
The survey recorded 50 perennial species from 42 genera and 28 families (Appendix 6).
The most dominant families were Chenopodiaceae (9 species), and Solanaceae (4
species). Three species that were new to the area (i.e. were not included in the
vegetation survey by Mattiske (1996)) were identified; Croton sp., Spyridium sp. and
Gastrolobium sp.. Furthermore, there were changes to priority status and name changes
since the last survey. Species are declared rare (R) or priority (P1-4) if they are
endangered or poorly known (Western Australian Herbarium 1998–). Pityrodia
cuneata is no longer a priority species, Plectrachne bromoides (R) has changed to
Triodia bromoides (P4), Brachyscome latisquamea has changed to Pembertonia
latisquamea and Loxocarya aspera has been changed to Desmocladus asper. In
addition to the native vegetation, three main weeds were identified.
Mesembryanthemum crystallinum (ice weed) and Brassica tournefortii (wild turnip)
were present in pits L and R, respectively, and Eucalyptus platypus, resulting from
seeds contained in the brushing material, was present in pit P. This Eucalyptus species,
whilst native to Western Australia, is not native to the area.
5.4.2 Plant abundance, species abundance and species richness in borrow pits and
undisturbed sites
Plant abundance ranged from 0.01 plants m-2
to 1.39 plants m-2
(Fig. 5.11a). The
undisturbed sites and pit L (topsoil replacement site) had higher plant abundance
compared with borrow pits P, Q and R. Also, pit L had significantly higher plant
abundance than the three undisturbed sites (P<0.05).
Species abundance ranged from 0.09 species 100 m-2
to 10.80 species 100 m-2
(Fig.
5.11b). Similar to plant abundance, pits P, Q and R had much lower species abundance
compared with the undisturbed sites and pit L. Undisturbed sites P, Q, R and pit L had
similar species abundance, although species abundance at site R was significantly
higher than at pit L (P<0.05) (Fig. 5.11b).
Species richness showed a different trend to plant and species abundance. Richness was
similar across all sites, ranging from 14 to 21 species (Fig. 5.11c). Pit Q had the highest
total number of species with pits L and P exhibiting the lowest.

97
Figure 5.11. a) Plant abundance (plants m-2
), b) species abundance (species 100 m-2
)
and c) species richness (total number of species in each site) in undisturbed sites R, Q
and P and borrow pits L, P, Q and R. Standard error is only indicated for the three
undisturbed sites, and borrow pit L, as there was no replication in pits P, Q and R.
5.4.3 Cover, abundance and frequency of select species
In general, Triodia plurinervata, Melaleuca cardiophylla, Thryptomene baeckeacea and
Acacia ligulata contributed to most of the cover in the undisturbed sites. However,
although the three undisturbed sites were in the same vegetation association determined
by Mattiske (1996), there were slight differences between sites. In particular, at site P,
Thryptomene baeckeacea and Melaleuca cardiophylla contributed over 20% (average
cover value >1) of the cover in each quadrat (Table 5.2), were the most abundant (Table
5.3) and occurred in all quadrats (Table 5.4). In sites Q and R, Triodia plurinervata
contributed to most of the cover (Table 5.2) followed by Melaleuca cardiophylla and
Acacia ligulata. In site Q, the most abundant and frequent species was Melaleuca
cardiophylla followed by Pembertonia latisquamea, while in site R, the most abundant
and frequent species were Atriplex bunburyana, Enchylaena tomentosa and
Pembertonia latisquamea followed by Pimelea microcephala subsp. microcephala.
Hence, from this data and observations, we can describe site P as mainly a Thryptomene
baeckeacea / Melaleuca cardiophylla heath, site Q is a low shrubland of Melaleuca
cardiophylla and Triodia plurinervata with Acacia ligulata and site R is similar to site
Q, but more diverse containing Atriplex bunburyana, Acacia tetragonophylla,
Enchylaena tomentosa, Pimelea microcephala subsp. microcephala and Scaevola
spinescens. In contrast, pit L had a very different species composition to the three
undisturbed sites, with Alyogyne cuneiformis and Stylobasium spathulatum dominating
the cover (Table 5.2) and occurring in high abundance (Table 5.3). Species with the
highest abundance in the remaining borrow pits (P, Q and R) were Acacia ligulata,
a) Plant abundance
0.0
0.5
1.0
1.5
2.0
P Q R L P Q R
Undisturbed
site
Borrow pit
Pla
nts
per
m2
b) Species abundance
0
2
4
6
8
10
12
P Q R L P Q R
Undisturbed
site
Borrow pit
Spe
cie
s p
er
100
m2
c) Species richness
0
5
10
15
20
25
P Q R L P Q R
Undisturbed
site
Borrow pit
To
tal num
ber
of specie
s

98
Alyogyne cuneiformis and Pimelea microcephala subsp. microcephala respectively
(Table 5.3).
The species that recruited from respread topsoil in pit L and through natural migration
in pits P, Q and R were investigated in further detail. Appendix 7 shows a list of nine
dominant species (i.e. were present in every quadrat in pit L) that recruited from topsoil.
Only three of these corresponded to the abundant species in the undisturbed sites (Table
5.3) – Acacia ligulata, Atriplex bunburyana and Enchylaena tomentosa. In addition,
several species that colonised borrow pits through natural migration (Appendix 7)
corresponded to abundant species in the undisturbed vegetation, including Acacia
ligulata, Atriplex bunburyana and Enchylaena tomentosa. However, many dominant
species in the undisturbed sites were under-represented in the borrow pits such as
Melaleuca cardiophylla, Thryptomene baeckeacea and Triodia plurinervata. Several
other species were only found in the undisturbed sites, for example Diplopeltis sp.
(Appendix 7), and did not colonise by natural migration. Many of these species,
including Diplopeltis sp. have limited distribution in the undisturbed sites (Appendix 8).
Table 5.2. Cover value of species at Shark Bay Salt in the undisturbed sites (US) P, Q R
and borrow pit (BP) L. Cover values range from 0-5 (0 = <19% cover, 1 = 20-39% 2 =
40-59% 3 = 60-79% 4 = 80-99% 5 = 100%). Values indicate cover value averaged
across 5 quadrats and average across all sites. Only species that contribute to over 20%
cover in two or more quadrats per site are shown. Species are arranged in order of
highest to lowest average cover value.
US BP
Species P Q R L av
Triodia plurinervata 0 4 2.2 0 1.6
Melaleuca cardiophylla 1.2 0.8 0.6 0 0.7
Thryptomene baeckeacea 1.6 0 0 0 0.4
Acacia ligulata 0.2 0.6 0.4 0 0.3
Stylobasium spathulatum 0 0 0 0.6 0.2
Alyogyne cuneiformis 0 0 0 0.4 0.1
Atriplex bunburyana 0 0 0.4 0 0.1
Acacia tetragonophylla 0 0 0.4 0 0.1
Enchylaena tomentosa 0 0 0.4 0 0.1
Pimelea microcephala subsp. microcephala 0 0 0.4 0 0.1
Scaevola spinescens 0 0 0.4 0 0.1

99
Table 5.3. Abundance of species at Shark Bay Salt in the undisturbed sites (US) P, Q R
and borrow pits (BP) L, P, Q, R. Values indicate number of plants per 100m2
. List of
species with the highest abundance (>5 plants per plot i.e. >0.05 plants per square
metre). Values for undisturbed sites (US) and borrow pit L are averaged across the five
quadrats. Values for borrow pits (BP) P, Q and R are not averaged as there was only
one replicate. Species are arranged in order of highest to lowest average abundance.
Table 5.4. Frequency of species at SBS the undisturbed sites (US) P, Q R and borrow
pit (BP) L. Values indicate how many quadrats the species is present in. Species shown
occur in all 5 quadrats at one or more sites. Species are arranged in order of highest to
lowest frequency.
US BP
Species P Q R L av
Melaleuca cardiophylla 5 5 4 0 4
Atriplex bunburyana 2 0 5 5 3
Enchylaena tomentosa 2 0 5 4 3
Pembertonia latisquamea 1 4 5 0 3
Pimelea microcephala subsp. microcephala 0 2 5 0 2
Alyogyne cuneiformis 0 0 0 5 1
Stylobasium spathulatum 0 0 0 5 1
Thryptomene baeckeacea 5 0 0 0 1
Species US BP av
P Q R L P Q R
Stylobasium spathulatum 53.60 0.13 0.10 17.94
Alyogyne cuneiformis 45.00 0.06 0.53 0.01 11.40
Thryptomene baeckeacea 41.40 0.03 13.81
Melaleuca cardiophylla 21.00 7.20 4.20 0.00 0.01 8.10
Atriplex bunburyana 0.40 10.80 8.80 0.01 0.10 0.10 3.37
Enchylaena tomentosa 1.00 9.40 5.20 0.21 0.02 3.17
Pembertonia latisquamea 0.40 4.40 8.40 0.01 0.04 2.65
Acacia ligulata 2.20 1.20 1.20 7.80 0.63 0.12 0.07 1.89
Lawrencia sp 0.00 8.40 8.40
Halgania littoralis 7.80 0.20 4.00
Pimelea microcephala
subsp. microcephala
0.80 6.20 0.17 0.36 1.88
Nicotiana occidentalis
subsp. hesperius
5.20 5.20

100
5.4.4 Species presence/absence and abundance
The presence/absence cladogram created in Primer illustrated that the four borrow pits
were not similar to the undisturbed vegetation sites, given the similarity index of 0 (Fig.
5.12). In addition, borrow pits P, Q and R, although similar to each other, were not
closely related to pit L (similarity index was 20). Quadrats within site L were related to
each other with an index of about 50.
Although the undisturbed sites were slightly related to each other, with a similarity
index of about 10 (Fig. 5.12), the three sites were also distinct from each other. The
undisturbed sites R and Q were the most closely related to each other with a similarity
index of about 30. These two sites were not very similar to site P, as there was a
similarity index of about 10. The quadrats within undisturbed sites R, Q and P were
quite closely related to each other being 60, 40 and 35 respectively. Correspondingly,
the presence/absence ordination illustrated distinctly grouped vegetation in the three
undisturbed vegetation sites and there were no overlaps (Fig. 5.13). In addition, the
relationship of the quadrats to each other within each site can also be seen in the
ordination, where R is the most tightly clustered, followed by Q then P (Fig. 5.13).
Figure 5.12. Presence/absence cladogram of the undisturbed sites P, Q R and borrow pit
L, P, Q, R. The entire of borrow pits P, Q and R were surveyed, and they are indicated
on the cladogram as numbers 16, 21 and 26 respectively. Five replicates at the
undisturbed sites were surveyed, and they are indicated by numbers 1-5 (site R), 6-10
(site Q) and 11-15 (site P). Five replicates in borrow pit L were surveyed, as indicated
by numbers 31-35.
Pit
P
Pit
Q
Pit
R
Borrow pits
Undisturbed sites
Pit L Site P Site R Site Q

101
Figure 5.13. Presence/absence ordination of the undisturbed sites P, Q R and borrow pit
L, P, Q, R. The entire of borrow pits P, Q and R were surveyed, and they are indicated
on the cladogram as numbers 16, 21 and 26 respectively. Five replicates at the
undisturbed sites were surveyed, and they are indicated by numbers 1-5 (site R), 6-10
(site Q) and 11-15 (site P) and are circled. Five replicates in borrow pit L were
surveyed, as indicated by numbers 31-35 and are circled.
The abundance cladogram (Fig. 5.14) showed similar trends as the presence/absence
cladogram (Fig. 5.12) in that the borrow pits were not similar to the undisturbed sites.
In addition, borrow pits P, Q and R were not similar to pit L (Fig. 5.14). Pits P, Q and R
were not as similar to each other in the abundance cladogram compared with the
presence/absence cladogram (Fig. 5.12, 5.14). Quadrats within pit L were related to
each other with a similarity index of about 40 (Fig. 5.14).
Unlike the presence/absence cladogram, the abundance cladogram showed that the
undisturbed site P was not related to sites Q and R as indicated by index of 0 (Fig. 5.14).
Sites Q and R were related with an index of >20 (Fig. 5.14). The abundance ordination
showed very distinct groupings between all sites and the stress value was 0.13 (Fig.
5.15).
Pit L (topsoil)
Site R
Site Q
Site P
Pit P
Pit Q Pit R

102
Figure 5.14. Abundance cladogram of the undisturbed sites P, Q R and borrow pit L, P,
Q, R. The entire of borrow pits P, Q and R were surveyed, and they are indicated on the
cladogram as numbers 16, 21 and 26 respectively. Five replicates at the undisturbed
sites were surveyed, and they are indicated by numbers 1-5 (site R), 6-10 (site Q) and
11-15 (site P). Five replicates in borrow pit L were surveyed, as indicated by numbers
31-35.
Figure 5.15. Abundance ordination of the undisturbed sites P, Q R and borrow pit L, P,
Q, R. The entire of borrow pits P, Q and R were surveyed, and they are indicated on the
cladogram as numbers 16, 21 and 26 respectively. Five replicates at the undisturbed
sites were surveyed, and they are indicated by numbers 1-5 (site R), 6-10 (site Q) and
11-15 (site P) and are circled. Five replicates in borrow pit L were surveyed, as
indicated by numbers 31-35 and are circled.
Site P
Site Q Site R
Pit L
Pit R Pit Q
Pit P
Pit
P
Pit
Q
Pit
R
Pit L Site P Site R Site Q
Borrow pits Undisturbed sites

103
5.5 Discussion
This study demonstrated that the borrow pits were generally not a reflection of the
adjacent undisturbed sites in terms of plant abundance and species composition. While
species such as Melaleuca cardiophylla and Thryptomene baeckeacea were dominant in
terms of cover, abundance and frequency in the undisturbed sites, this was not the case
in the borrow pits where topsoil was spread, given that Alyogyne cuneiformis and
Stylobasium spathulatum were more abundant.
5.5.1 Comparison of assessment techniques
Three methods - cover, abundance and frequency - were used to determine dominant
species in the undisturbed vegetation adjacent to borrow pits P, Q and R and in borrow
pit L. Generally, most species did not rank equally in all methods (exception being
Melaleuca cardiophylla). Therefore, using only one method would not have provided a
reliable assessment of dominant species. For example, Acacia ligulata was one of the
dominant species in pit Q, as it provided much of the cover. However, this cover was
provided by just a few large plants. In addition, it was useful to use both species
abundance and species richness data to achieve a reliable assessment. For instance,
whilst the undisturbed sites and borrow pits contained similar species richness (between
14 and 21 species), there was a large discrepancy in species abundance (0.08-10.80
species 100 m-2
). By monitoring a range of parameters, the dataset can provide a more
balanced approach species selection for rehabilitation purposes. The information can
also identify species that are lacking from borrow pits subjected to restoration. For
example, even though Atriplex bunburyana and Acacia ligulata were common to both
the undisturbed sites and the borrow pits, Melaleuca cardiophylla, Thryptomene
baeckeacea and Triodia plurinervata were absent from the borrow pits, and steps may
need to be taken to re-introduce these dominant species.
5.5.2 Differences between undisturbed vegetation sites
Although Mattiske (1996) classified the undisturbed vegetation sites (that are acting as
reference points in this study) as belonging to the same vegetation association, the
cladograms and ordinations showed differences between the sites. It is not surprising
that both cladograms rated undisturbed vegetation sites R and Q as more closely related
to each other than to site P as the vegetation was quite different. Site R was generally a
low shrubland / heath of Atriplex bunburyana, Melaleuca cardiophylla and Triodia

104
plurinervata with occasional emergent Acacia sp. Site Q was also a low shrubland of
Melaleuca cardiophylla and Triodia plurinervata but contained more Acacia ligulata.
Site P however was a Thryptomene baeckeacea and Melaleuca cardiophylla heath.
5.5.3 Borrow pit with topsoil replacement
Whilst there were differences between the undisturbed sites, they were more similar to
each other than they were to any of the borrow pits. Although topsoil was replaced on
borrow pit L in 2003, the vegetation was dissimilar to the undisturbed sites and this can
be attributed to a number of reasons. Firstly, given that some species store their seeds in
the canopy, their seeds would not be transferred to rehabilitation sites via topsoil
replacement (Rokich et al. 2002). These may include resprouter species and those that
employ vegetative reproduction would not be present (Grant and Loneragan 1999).
Further, it is likely that topsoil was stockpiled, which results in lower species richness
compared with direct returned topsoil (Ward et al. 1996). Loss of species may be due to
seed decomposition during stockpiling, as Rokich et al. (2000) noted lower seed
viability of several species stored in stockpiled topsoil compared with un-stockpiled
seed. Also, the topsoil came from pit H, which is not adjacent to the undisturbed
vegetation areas investigated. The original species composition at this site may have
been different to the undisturbed vegetation sites adjacent to pits P, Q and R that were
investigated in this study, even though they are in the same vegetation association as
defined by Mattiske (1996). Moreover, pit L contained high numbers of plants of
species such as Stylobasium spathulatum and Alyogyne cuneformis that were not
recorded in the undisturbed sites during the survey. It is possible that these species were
not necessarily absent from the undisturbed vegetation, but they may have been present
in low numbers (and hence not detected by the survey) and are colonisers or disturbance
‗opportunist‘ species. Koch and Ward (1994) found that colonising species (also
termed r-type species) were dominant in new jarrah forest rehabilitation sites. High
numbers of colonising species is not a concern as they generally have a short life span
and over time it is expected that their numbers will decline to frequencies matching
nearby vegetation. The difference between pit L and the undisturbed vegetation may
also be due to the absence of species otherwise dominant in the undisturbed vegetation
such as Melaleuca cardiophylla, Thryptomene baeckeacea and Triodia plurinervata.
According to the relay floristics model, these dominant species will establish in time.
The model implies that following disturbance, groups of species will replace each other
over time (Egler 1954). However, the relay floristics model may not apply in this

105
ecosystem. For example, Koch and Ward (1994) state that the initial floristic model is
more appropriate for the jarrah forest in the southwest of Australia. This model implies
that species present in the final community will establish immediately post-mining
(Koch and Ward 1994) (and not migrate from nearby undisturbed vegetation).
Analogously, Holmes and Richardson (1999) have similar findings for the South
African fynbos. They state that after disturbance (in this case, fire) all species return
immediately, and species composition changes over time as short-lived species die,
leaving longer-lived species in the final community. Similarly, Rokich (1999) states
that plant abundance and species richness are highest in the first year of rehabilitation in
Banksia woodland. In addition, a key priority in Banksia woodland rehabilitation is to
achieve the highest possible levels of recruitment of all species in the first year of
rehabilitation given the onset of soil settling in the second year that impedes further
seedling recruitment, seedling establishment and plant survival beyond that year.
Southwest Australia and South Africa are very similar in terms of climate, they both
have a rich flora, their landscapes are nutrient deficient and they have not been glaciated
since the Permian (Hopper and Gioia 2004). Both of these regions have been recently
termed OCBILs (old climatically-buffered infertile landscapes), and have been
described as containing few r-type species that are dominant in the contrasting
YODFELs (young, often-disturbed fertile landscapes) (Hopper 2007). These reasons
may explain why it is more appropriate to use the initial floristics model in both of these
ecosystems, while the relay floristics model may be more appropriate in YODFELs. In
addition, given the location of Shark Bay on the edge of southwest Australia, it may
also be considered an OCBIL and therefore it would be appropriate to consider the intial
floristics model in this area. Therefore, the absence of dominant species such as
Melaleuca cardiophylla, Thryptomene baeckeacea and Triodia plurinervata is of
particular concern and steps will need to be taken to introduce these important species.
5.5.4 Borrow pits with limited treatments
Borrow pits that have not been subject to rehabilitation treatments (apart from brushing)
have very low plant abundance. Ward et al. (1990) also found that revegetation is slow
in untreated post-mined areas. The differences in plant abundance between the
untreated borrow pits and the topsoil replacement pit highlights the importance of using
topsoil in restoration, which has also been shown to be highly beneficial in biodiverse
ecosystems (Rokich et al. 2000). Colonisation from surrounding vegetation has only
happened to a limited extent in this ecosystem. Limited colonisation may be due to

106
minimal seed dispersability. According to Hopper (2007), seed dispersability is
minimal in the previously described OCBILs. Minimal seed dispersal has also been
shown to restrict revegetation of some areas of former farm land in southwest Australia
(Standish et al. 2007) and seed dispersal distances are short in the South African fynbos
(Holmes and Richardson 1999) . This contrasts with other habitats where vacant land is
quickly colonised by species from surrounding areas (Hobbs and Cramer 2007). These
other habitats are the previously described YODFELs, where many species have wide
and rapid dispersal enabling fast colonisation as they are adapted to large scale
disturbance such as glaciation and volcanic activity (Hopper 2007).
5.5.5 Conclusion
This study has highlighted that vegetation in disturbed areas is highly different to
undisturbed areas and that there is a need for rehabilitation research and technologies to
address the species shortfall. In addition, this study has identified dominant species that
will need to be researched in order to maximise their establishment in rehabilitation
sites.

107
C H A P T E R 6
Topsoil replacement and broadcast seeding in borrow pits
6.1 Abstract
The borrow pits at Shark Bay Salt are generally devoid of vegetation. This chapter
investigated the potential to re-introduce flora through ‗borrowed‘ topsoil and seed
broadcasting. Seedling recruitment from ‗borrowed‘ topsoil was generally similar in the
donor site (natural vegetation) and the borrow pits. Seed broadcasting investigations
focused on the effect of year and season of sowing, and the effects of soil ripping,
raking and seed priming for improving emergence success. Seedling emergence
following seed broadcasting was greater in the wetter year and following sowing in
autumn compared with winter. Soil ripping and raking were generally beneficial to
seedling emergence, particularly in combination with higher rainfall, whereas seed
priming had little effect on seedling emergence.

108
6.2 Introduction
The absence of vegetation in the borrow pits at Shark Bay Salt (SBS) after >10 years of
clearing (chapter 5) highlights a need to actively rehabilitate decommissioned pits with
endemic flora. Typically, vegetation may be replaced in post-disturbed sites via three
sources; respread topsoil, broadcast seeds and nursery produced seedlings (hereafter
referred to as greenstock). It is well documented that respread topsoil can provide an
important source of seeds (Koch et al. 1996; Rokich et al. 2000). However, species that
do not store seeds in the topsoil seed bank, but instead store their seeds in the canopy
(serotinous taxa) (Rokich and Dixon 2007), and those that do not emerge from respread
topsoil, may be returned to site via broadcast seeding and greenstock planting.
Broadcast seeding and greenstock planting also play an essential role when topsoil is in
limited supply or is absent. In addition, broadcast seeding of species that do emerge
from topsoil is a common practice as it may increase plant density, cover and species
richness (Koch and Ward 1994). Planting greenstock propagated from seeds, or
vegetatively via cuttings or tissue culture, may be necessary when low seed numbers,
viability or germinability prevents establishment from broadcasting or the topsoil seed
bank (Gardner 2001). Two sources of vegetation replacement will be investigated in
this chapter – topsoil and seed broadcasting, while the third, greenstock, will be studied
in the following chapter.
Given the limited topsoil availability at SBS, an alternative means of sourcing the
topsoil resource is necessary. This may include ‗borrowing‘ topsoil, whereby topsoil is
sourced from natural vegetation adjacent to a post-disturbed site requiring rehabilitation.
To allow regeneration of these donor sites, some of the ‗borrowed‘ topsoil should be
returned. To optimise regeneration in both donor and post-disturbed sites, topsoil may
be mixed, divided in half, with 50% of the topsoil used in the post-disturbed sites, and
the remaining 50% returned to the donor site. While this is, at this time, an untested
technique, it has the potential to provide benefits to rehabilitation activities.
To optimise seedling emergence, growth and survival, seed broadcasting practices need
to be tailored to site and climatic conditions. Seed sowing times have an effect on
seedling emergence (Ward et al. 1996; Turner et al. 2006c), and therefore need to be
investigated in order to find the optimal sowing time. For instance, Turner et al.
(2006c) found that emergence of Banksia woodland seeds from southwest Western

109
Australia broadcast in May (autumn) was greater than seeds broadcast in July (winter).
Also, microsites provide niches for seedling emergence (Winkel et al. 1991;
Elmarsdottir et al. 2003; Doust et al. 2006) and these sites may be provided by furrows
in ripped soil. In addition, incorporating seeds into the soil via raking has been shown
to increase seedling emergence (Turner et al. 2006c), possibly due to reducing the loss
of seeds through predation or erosion (Rokich and Dixon 2007). Finally, the time taken
for seedlings to emerge can be slow and erratic (Heydecker et al. 1973; Karssen et al.
1989). Seed priming can be used to increase the rate and synchronicity of germination
(Heydecker et al. 1973; Heydecker and Coolbear 1977). Hydropriming involves
soaking seeds in water so the seeds imbibe and begin metabolism, then drying seeds just
prior to radical emergence. Priming can also be undertaken with germination stimulants
such as GA3. For instance, priming with water increased field emergence (rate and
percentage) of Atriplex amnicola, and emergence was further increased by priming with
GA3 (Stevens et al. 2006).
The aim of this chapter was to investigate the success of ‗borrowed‘ topsoil and
broadcast seed for rehabilitating the borrow pits at SBS. As topsoil is currently
unavailable for use in decommissioned borrow pits, topsoil was ‗borrowed‘ from
adjacent natural vegetation to enable understanding of the potential to rehabilitate both
the donor and recipient sites. To optimise seedling recruitment through seed
broadcasting, the effects of year, season, soil ripping, raking of seeds into the soil, and
seed priming were investigated.

110
6.3 Methods
6.3.1 Site description
Three borrow pits at SBS were chosen for the ‗borrowed‘ topsoil replacement (pits G, K
and P) and seed broadcasting experiments (pits P, Q and R) (Appendix 1, 2). There
were three criteria for pit selection: greater than 1ha in area, not recently subjected to
rehabilitation works, and accessible for machinery. Different pits were used for the
topsoil and seed broadcasting experiments due to space and machinery access
requirements. Pit K consisted of a large active area, whilst a small inactive area was
employed for the topsoil replacement trial. Pit P also had two distinct areas; topsoil
replacement was implemented in the smaller area which was higher in the topography,
and just north-east of a larger area in which the seed broadcasting was undertaken.
Prior to seed broadcasting, a 40 x 25 m area was marked out in pits Q, R and the larger
area of pit P. A grader ripped half of the area (20 x 25 m) with four 50-70 mm wide
tynes in April 2005. A 0.8 m high fence was then erected around the whole area to
discourage herbivory by kangaroos and rabbits.
Daily rainfall for the study period (January 2005 – March 2008) was monitored at the
site by SBS. Soil temperatures were monitored hourly from January to December 2007
using a Tinytag Plus 2 data logger. Loggers were placed in the larger area of pit P and
the natural vegetation adjacent to the pit. The probes of the loggers were placed level
with the surface of the soil.
6.3.2 Part A. Topsoil replacement
On 10 May (autumn) 2006, vegetation was cleared from three 5 x 1 m plots in vegetated
areas adjacent to three borrow pits (G, K and P). Topsoil was stripped from each plot to
a depth of 15 cm, placed on a tarpaulin and mixed. Half of the topsoil was replaced
onto the 5 x 1 m plot in the vegetated area (i.e. to a depth of 7.5 cm). The other half was
spread on a 5 x 1 m plot in the adjacent borrow pit. Seedling emergence was assessed
in all plots in October (spring) 2006, May (autumn) 2007, October (spring) 2007 and
April (autumn) 2008.
6.3.3 Part B. Seed broadcasting experiments conducted in 2005
Four species that were dominant in the natural vegetation and that covered a range of
life forms were chosen for the trial: Acacia tetragonophylla, Atriplex bunburyana,

111
Rhagodia baccata and Solanum orbiculatum. Seeds were cleaned by removing them
from pods (Acacia tetragonophylla), fleshy fruited berries (Rhagodia baccata, Solanum
orbiculatum), and utricles (Atriplex bunburyana). Optimised seed pre-treatments to
break dormancy or stimulate germination were determined from a previous experiment
(chapter 2). Acacia tetragonophylla seeds were soaked in hot (c. 90oC) water for 2 min.
Rhagodia baccata seeds were soaked in gibberellic acid (GA3) for 24 h. Solanum
orbiculatum seeds were soaked in karrikinolide (KAR1) for 24 h. Seeds that were
soaked were then dried at ambient laboratory conditions (ca. 22oC, 50% RH) for ease of
handling.
The effects of season, soil ripping and raking seeds into soil were tested individually
and in combination. The treatments were as follows: (a) neither ripping nor raking
(control), (b) ripping only, (c) raking only and (d) ripping + raking. For each treatment,
one hundred seeds of all species were broadcast in a 2 x 2 m plot (except Rhagodia
baccata where 150 seeds were used) on 29 April (autumn) and 28 June (winter) 2005.
Three replicate 2 x 2 m plots were implemented for each treatment in each of the three
pits (P, Q and R). Final seedling emergence data was collected on 13 November
(spring) 2005.
6.3.4 Part C. Seed broadcasting experiments conducted in 2006
Eight species that were dominant in the natural vegetation and that covered a range of
life forms were chosen for the trial: Acacia tetragonophylla, Atriplex bunburyana,
Aphanopetalum clematideum, Dioscorea hastifolia, Melaleuca cardiophylla, Senna
glutinosa, Solanum orbiculatum and Zygophyllum fruticulosum. Seeds of all species
were collected in 2005 from SBS (except for Dioscorea hastifolia and Zygophyllum
fruticulosum that were collected in 2004). Seeds were cleaned by removing them from
pods (Acacia tetragonophylla, Senna glutinosa), fleshy fruited berries (Solanum
orbiculatum), capsules (Dioscorea hastifolia, Melaleuca cardiophylla and Zygophyllum
fruticulosum), nuts (Aphanopetalum clematideum) and utricles (Atriplex bunburyana).
Prior to sowing, seed dormancy of certain species was broken or seeds were treated with
a stimulant to facilitate germination. Acacia tetragonophylla, Atriplex bunburyana and
Solanum orbiculatum were pre-treated as per Part B. Seeds of Dioscorea hastifolia and
Zygophyllum fruticulosum had after-ripened after >1 year at ambient laboratory
conditions (c. 23oC, 50% RH) (Appendix 9, 10), while Senna glutinosa was soaked in
hot (c. 90oC) water for 2 min. Seeds of the remaining species did not require treatment.

112
The treatments investigated in this trial were soil ripping, raking seeds into topsoil, and
seed priming. The trial was implemented at each of the three pits (P, Q and R) used in
the 2005 trial. The treatments were as follows: (a) neither ripping nor raking (control),
(b) ripping only, (c) raking only and (d) ripping + raking.
These treatments were repeated with primed seeds of Atriplex bunburyana and Solanum
orbiculatum only. Priming treatments were not undertaken on the remaining six species
due to limited seed numbers. Atriplex bunburyana seeds were soaked in GA3 for 24 h
and Solanum orbiculatum seeds were soaked in water for 48 h then KAR1 for 24 h.
Seeds were then dried at ambient laboratory conditions (ca. 22oC, 50% RH) (Appendix
11, 12).
For each treatment, one hundred seeds of each of the species were broadcast in a 2 x 2
m plot on 10 May (autumn) 2006. Each treatment was replicated in three 2 x 2 m plots
in each of the three pits. Final seedling emergence data was collected on 29 October
(spring) 2006.
6.3.5 Statistics
Each experiment was analysed using a general analysis of variance (ANOVA) to
determine whether there were significant effects of treatment, species or pit. For the
data displayed in each graph, two-way ANOVAs with Fisher‘s unprotected LSD (using
a significance level of 0.05) were used to separate the means. Percentage data were
arcsine transformed prior to analysis but untransformed data is presented in figures.
6.4 Results
6.4.1 Rainfall
Rainfall over the study period was highly variable in terms of amount and periodicity.
In 2005, rainfall was distributed over 10 months, and was highest May (autumn) and
June (winter), followed by April (autumn) and August (winter) (Fig. 6.1a). The total
yearly rainfall was 300 mm (Fig. 6.1a). Rainfall in 2006 and 2007 was very low, and
both years experienced <100 mm (Fig. 6.1b, c) with rain distributed over 10 months in
each year. The yearly average from 2005-2007 (146 mm) was lower than that from
1983-2007 (211 mm) (data not shown). Rainfall in 2008 was only recorded until the

113
end of April (autumn), however a large rainfall event in March (autumn) from ex-
tropical cyclone Pancho resulted in 308 mm of rain in four days, exceeding the total
rainfall for 2006 and 2007 combined (Fig. 6.2). Maximum soil temperatures in the
summer months were frequently >60oC (Fig. 6.3). Minimum soil temperatures were
<25oC for the duration of the year (Fig. 6.3).
a) 2005
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan
Daily
ra
infa
ll (m
m)
0
10
20
30
40
501 2 3
b) 2006
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan
Daily
ra
infa
ll (m
m)
0
10
20
30
40
50
c) 2007
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan
Daily
ra
infa
ll (m
m)
0
10
20
30
40
50
4 5 6
7 8
Figure 6.1. Rainfall (mm) per day in Useless Loop in a) 2005, b) 2006 and c) 2007. Red
lines and numbers indicate timing of experimental trials; 1: seed broadcasting April
2005, 2: seed broadcasting June 2005 and greenstock planting (chapter 7), 3:
broadcasting and greenstock assessment, 4: seed broadcasting 2006 and topsoil
replacement, 5: greenstock planting 2006 (chapter 7), 6: topsoil, broadcasting and

114
greenstock assessment, 7: topsoil and greenstock assessment, 8: topsoil and greenstock
assessment.
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan
Daily
rain
fall
(mm
)
0
50
100
150
200
250
Figure 6.2. Rainfall (mm) per day in Useless Loop from 1 January – 1 April 2008 (note
change of scale from Fig. 6.1). Red line indicates greenstock and topsoil assessment
date.
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Te
mp
era
ture
(oC
)
0
10
20
30
40
50
60
70
80Max
Av
Min
Figure 6.3. Maximum, minimum and average daily soil temperatures in the natural
vegetation adjacent to pit P from 1 January to 31 December 2007.

115
6.4.2 Part A. Topsoil replacement
Seedling recruitment from topsoil in October (spring) 2006 ranged from 1 to 53
seedlings m-2
(Fig. 6.4) with Pit K recording the greatest seedling numbers, and the only
site with differences between a borrow pit and the adjacent natural vegetation. Species
richness ranged from 3.7 to 9.7 species 5 m-2
with no difference in species numbers
between natural vegetation and borrow pits, however, species numbers were higher in
pits G and K compared with pit P. Very few seedlings were present in May (autumn)
2007 (data not shown) implying high seedling mortality over the summer months. In
October (spring) 2007 seedling numbers ranged from 0 to 41.2 seedlings m-2
as a result
of recruitment over winter 2007, with no difference between natural vegetation and
borrow pits, however numbers were higher in pit K compared to pit P only. Species
richness was generally similar to October 2006. There was a large amount of seedling
recruitment (up to 86 seedlings m-2
) in April (autumn) 2008 (Fig. 6.4), so that by this
time there was generally no difference between natural vegetation and borrow pit sites.
October (Spring) 2006
Seedlin
gs (
m-2
)
0
20
40
60
80
100
120
140October (Spring) 2007 April (Autumn) 2008
Nat Veg
Borrow Pit
G K P
Specie
s r
ichness (
5m
-2)
0
5
10
15
Pit
G K P G K P
a
ab
a
b
a a
ab abab
b
a a
ab
bc
c c
a a
c
cbc
c
ab
a
c
abc
bcabc
ab a
Figure 6.4. Number of seedlings (m
-2) and species richness (species 5 m
-2) from
‗borrowed‘ topsoil replacement at the natural vegetation and borrow pits at sites G, K
and P in October 2006, October 2007 and April 2008. Bars indicate standard error.
Letters indicate significant (P<0.05) differences between the means within each graph.

116
6.4.3 Part B. Seed broadcasting experiments conducted in 2005
The timing of seed broadcasting had the largest effect on seedling emergence. The
earlier broadcasting in April (autumn) 2005 resulted in seedling emergence of 0-27%
(Fig. 6.5a) compared with the later broadcasting in June (winter) 2005 that resulted in
0-2% seedling emergence (Fig. 6.5b).
When sown in autumn, raking or ripping alone increased the seedling emergence of
Acacia tetragonophylla by at least five-fold compared with the control, while a
combination of raking and ripping further increased seedling emergence by an
additional two-fold (Fig. 6.5a). Raking and/or ripping increased the seedling emergence
of Solanum orbiculatum by at least 50-fold; ripping and the combination of ripping +
raking increased the seedling emergence of Atriplex bunburyana by 10-fold; while there
was no effect of the treatments on the seedling emergence of Rhagodia baccata (Fig.
6.5a). In addition, the combination of ripping and raking increased total seedling
emergence (all species combined) compared with the control at each of the three pits by
>10-fold (Fig. 6.6). Given the low emergence across all treatments, when sown in
winter, treatment effects were less pronounced with the combination of ripping and
raking increasing the seedling emergence of only Acacia tetragonophylla, while raking
only increased the seedling emergence of Atriplex bunburyana (Fig. 6.5b).
There was a difference (P<0.05) in seedling emergence between the species.
Emergence of Acacia tetragonophylla was higher than that of Atriplex bunburyana and
Solanum orbiculatum, which in turn were higher than Rhagodia baccata (Fig. 6.5).
There was no difference between seedling emergence at each of the three pits with all
pits averaging approximately 6% emergence.

117
a) April
A. tetra
gonophylla
A. bunburyana
R. baccata
S. orbiculatum
Em
erg
ence
(%
)
0
10
20
30
40
100
b) June
A. tetra
gonophylla
A. bunburyana
R. baccata
S. orbiculatum
Control
Rake only
Rip only
Rip + rake
a
bb
c
aab
cbc
a
bcb
c
aba ab
b
a b a ab
Figure 6.5. Emergence (%) of Acacia tetragonophylla, Atriplex bunburyana, Rhagodia
baccata and Solanum orbiculatum sown in a) April 2005 and b) June 2005 and
subjected to one of four treatments; control, rake only, rip only, rip + rake. Values are
averages across the three borrow pits. Bars indicate standard error. Letters indicate
significant (P<0.05) differences between the means of each species at each month.
P Q R
Em
erg
ence (
%)
0
10
20
30
100 Control
Rake only
Rip only
Rip + rake
ab
abccd
d
a
abccd
cd
a
bcdabc
cd
Figure 6.6. Emergence (%) at pit P, Q and R of seeds broadcast in April 2005 across
one of four treatments; control, rake only, rip only, rip + rake. Values are averages
across the four species. Bars indicate standard error. Letters indicate significant
(P<0.05) differences between the means.

118
6.4.4 Part C. Seed broadcasting experiments conducted in 2006
Seedling emergence was very low with all treatments exhibiting less than 2.5%
emergence (Fig. 6.7). There was no effect of ripping or raking on the emergence of any
of the species (Fig. 6.7). There was a difference in seedling emergence between the
species as Acacia tetragonophylla and Zygophyllum fruticulosum had the highest
seedling emergence, while Dioscorea hastifolia and Melaleuca cardiophylla failed to
emerge (Fig. 6.7). The combination of ripping and raking increased seedling emergence
at pit R, whereas ripping and raking alone did not increase seedling emergence at any of
the pits (Fig. 6.8). Seedling emergence was higher at pits Q and R compared with pit P
(Fig. 6.8).
A. tetra
gonophylla
A. clematid
eum
D. hastifo
lia
M. cardiophylla
S. glutin
osa
Z. frutic
ulosum
Em
erg
ence (
%)
0
1
2
3
4
5
100 Control
Rake only
Rip only
Rip + rake
Figure 6.7. Emergence of seeds of six species sown in May 2006. Seeds were sown in
plots with the following treatments; control, rake only, rip only and rip + rake.
Emergence was scored in October 2006. Values are averages across all three pits. Bars
indicate standard error.
P Q R
Em
erg
ence (
%)
0
1
2
3
4
5
100Control
Rake only
Rip only
Rip + rake
abaab
bc
ababab
c
Figure 6.8. Emergence (%) at pit P, Q and R of seeds broadcast in May 2006 across one
of four treatments; control, rake only, rip only, rip + rake. Values are averages across
the six species. Bars indicate standard error. Letters indicate significant (P<0.05)
differences between the means.

119
Seed priming did not have an effect on the emergence of Atriplex bunburyana and
Solanum orbiculatum seeds (Fig. 6.9). Ripping and raking also did not have an effect
on the emergence of unprimed and primed seeds (Fig. 6.9). Furthermore, there was no
difference in emergence between the two species. There was a difference in emergence
between the three pits, with higher emergence at pit R compared with pits P and Q (Fig.
6.10).
unprimed primed unprimed primed
Em
erg
en
ce
(%
)
0
1
2
3
4
5
100
A. bunburyana S. orbiculatum
Control
Rake only
Rip only
Rip + rake
Figure 6.9. Emergence % of unprimed and primed seeds of Atriplex bunburyana and
Solanum orbiculatum sown in May 2006 and subject to one of four treatments; control,
rake only, rip only, rip + rake. Bars indicate standard error.
P Q R
Em
erg
en
ce
(%
)
0
1
2
3
4
5
100Unprimed
Primed
aab a
a
b
ab
Figure 6.10. Emergence of unprimed and primed seeds in pits P, Q and R. Values are
averaged across the two species. Bars indicate standard error. Letters indicate
significant (P<0.05) differences between the means.

120
6.5 Discussion
This is the first study into the use of topsoil replacement and seed broadcasting as
restoration tools in the Shark Bay World Heritage Area. It highlights the difficulties in
returning plants to a post-disturbed site in an arid environment where rainfall periods
and amount are difficult to predict, compounded by the high temperatures experienced
at the site. Seedling emergence from seed broadcasting was low, as was seedling
survival from replaced topsoil. While seedling emergence was strongly and negatively
affected by low rainfall years, treatments to improve seedling emergence, including
early seed sowing time, soil ripping and raking seeds into soil, are now recognised.
6.5.1 Topsoil replacement
Although there was some recruitment from topsoil in October (spring) 2006 (up to 53
seedlings m-2
), there were very few seedlings remaining in April (autumn) 2007. This
indicates that either a great proportion of annual flora emerged in October (spring)
2006, or that perennial flora emerged but did not survive over summer. The recruitment
from topsoil in October (spring) 2007, indicated that seeds persisted in the seed bank.
The seedlings present in April (autumn) 2008 were very small, hence they were unlikely
to be seedlings that emerged in 2007 and survived over summer, rather they may have
emerged after a high rainfall event (308 mm in early 2008) two weeks prior to scoring.
This pattern of inconsistent seedling emergence highlights the complex relationship
between dormancy status of seeds in the soil seed bank and the amount of rainfall that is
required for germination and survival. These results indicate the presence of a
persistent seed bank, where not all seeds of all species germinate during the first
occurrence of favourable conditions, but seeds within a species, or seeds of different
species are able to spread their germination across a number of years due to varying
levels of dormancy or varying requirements for germination. A persistent seed bank
(which is a ‗bet-hedging‘ strategy) is more common in unpredictable environments
(Philippi 1993; Fenner and Thompson 2005).
The difficulties with using topsoil in the arid zone may be due to an uneven distribution
of seed in soil. Organic matter and seeds were observed to collect under the clumps of
shrubs. Patchy distribution of seeds in the seed bank in semi-arid regions has been
previously described (Page et al. 2006). As seeds collect under shrubs, they may also
be removed from the bare areas between shrubs. For instance, greater proportion of

121
seeds under shrubs compared with areas between shrubs has been shown in four North
American deserts (Guo et al. 1998). Hence, in this study, recruitment may have been
limited due to wind erosion which may have removed soil and seeds. While this study
indicated the potential for seedling recruitment from both the donor and recipient sites,
an understanding of seedling recruitment and ultimately plant survival within an
undisturbed reference site is crucial to understand the success of employing ‗borrowed‘
topsoil.
6.5.2 Seed broadcasting experiments conducted in 2005 and 2006
The low seedling emergence from seed broadcasting in plots that were not subject to
treatment is consistent with other Australian studies which report 1.2% emergence for
species in Banksia woodland (Turner et al. 2006c), and 1.8-3.3% for rainforest species
(Doust et al. 2006). In the present study, amending the sowing time, undertaking soil
ripping and raking seeds into the soil increased emergence to 9.8-14.0% (in 2005).
Analogously, earlier optimal sowing time and raking seeds into the soil resulted in 11%
emergence in Banksia woodland in Western Australia (Turner et al. 2006c) and optimal
soil micro-sites resulted in up to 25-34% emergence at three sites in a Queensland
rainforest (Doust et al. 2006).
The higher seedling emergence achieved from seed broadcasting of three common
species (Acacia tetragonophylla, Atriplex bunburyana and Solanum orbiculatum) in
April (autumn) 2005 (≤27%) compared with May (autumn) 2006 (<4%) may be due to
differences in rainfall between the two years (300 mm in 2005 and 58 mm in 2006)
(Fig. 6.1a, b). Seasonal differences in recruitment has been noted in another area in
Australia with low rainfall (Knight et al. 1998). In addition, there was a difference in
seedling emergence between seed broadcasting in April (autumn) and June (winter)
2005 reflecting the distribution of rain throughout the 2005 winter. There was a large
amount of rainfall in May and June 2005, and very little in July and August 2005 (Fig.
6.1a). This rainfall distribution for 2005 resulted in 293 mm recorded between sowing
in April and scoring in October, and only 49 mm received between sowing in June and
scoring in October. Two other Australian studies have demonstrated differences in
seedling emergence depending on sowing time (Knight et al. 1998; Turner et al. 2006c).
In southwest Australia, seeds sown in May (autumn) exhibited greater seedling
emergence compared with seeds sown in July (winter) (Turner et al. 2006c). Also, at
one of the locations in a South Australian study, sowing in April (mid autumn) resulted

122
in greater emergence than May (late autumn), which in turn resulted in greater
emergence than sowing in June, July and August (winter) (Knight et al. 1998).
Raking seeds into the soil and/or soil ripping increased seedling emergence of three out
of the four species broadcast in April (autumn) 2005 with the combination of raking and
ripping increasing seedling emergence at all three pits. Firstly, it has been postulated
that raking may reduce seed losses from predation or erosion, and when seeds are
incorporated into the soil they experience lower soil temperatures compared with the
soil surface (Rokich and Dixon 2007). Other studies have also found that incorporating
seeds into the soil increases seedling emergence, such as Turner et al. (2006c) who
found that raking increased emergence of eight out of nine species and Doust et al.
(2006) who found that seeds of rainforest species that were buried had higher
establishment compared with seeds sown on the soil surface. Ripping may create
microsites that are favourable for seed germination and seedling emergence. Studies
have shown the importance of these microsites in seedling establishment (Winkel et al.
1991; Elmarsdottir et al. 2003; Petersen et al. 2004; Doust et al. 2006). For instance,
seeds sown in furrows (rip-lines) had higher establishment compared with those sown
on undisturbed soil (Doust et al. 2006). Also, seeds sown in cracks in the soil surface
had greater emergence compared with those sown on bare soil (Winkel et al. 1991).
These microsites may provide shelter for seedlings (Elmarsdottir et al. 2003). Shelter
may decrease wind exposure, which may decrease wind erosion and soil and seedling
desiccation compared with the smooth soil surface of the control plots. Wind erosion
has been shown to displace, on average, 67% of broadcast seeds from where they were
placed in post-mined restoration sites within Banksia woodland (Ord 2007). In addition
to providing shelter, water may collect in rip-lines, hence soil moisture would be higher
in rip-lines compared to flat control plots. In this study, seeds broadcast in ripped plots
were not only broadcast in rip-lines, they were also broadcast on mounds between rip-
lines. These mounds may not be as conducive for seedling establishment, as they are
more exposed to the wind, and may have lower soil moisture compared with the
riplines. Hence, it is not surprising that incorporating the seeds into the soil by raking
further improved emergence in some of the ripped plots.
Priming did not have an effect on in-situ seedling emergence in this study. Emergence
of primed and unprimed seed was low, probably due to the low rainfall in 2006 as stated
above. Therefore, it is difficult to determine whether priming would have had an effect

123
in a higher rainfall year where overall emergence might be higher. Field emergence of
primed seeds can be variable. Hardegree and Van Vactor (2000) postulated that
priming results were affected by sowing date and soil type. Also, there can be
differences between priming of seeds of different species across different genera and
across different species within the same genus (Hardegree and Van Vactor 2000;
Stevens et al. 2006). For instance, priming enhanced germination of Atriplex amnicola,
but not Atriplex nummularia and Atriplex undulata (Stevens et al. 2006). In addition,
priming increased total emergence of one species and emergence rate of three out of
four species, at one or more of the sowing dates (Hardegree and Van Vactor 2000).
While three species in 2005 had similar ex-situ germination to each other (>80%) and
the fourth (Atriplex bunburyana) had only slightly less (>70%) (Chapter 2), the
differences in the in-situ seedling emergence between the species might be attributed to
soil water availability and seed size. Soil water availability can affect germination of
species differently. For instance, water stress (-1 MPa) decreased germination of
Callitris verrucosa from 100% (at 0 MPa) to 82%, whereas water stress reduced
germination of Callitris preissii from 100% to 43% (Adams 1999). In addition, Doust
et al. (2006) postulated that seed size affects seedling establishment, finding that when
seeds were buried, larger seeds had greater emergence compared with smaller and
intermediate-sized seeds. Seed size seemed to correlate with emergence in this study.
In 2005, emergence of Acacia tetragonophylla > Solanum orbiculatum and Atriplex
bunburyana > Rhagodia baccata and the weight of 100 seeds of Acacia tetragonophylla
(1.6 g) > Atriplex bunburyana (0.5 g) > Solanum orbiculatum (0.2 g) > Rhagodia
baccata (0.06 g).
A final factor that may limit seedling recruitment from topsoil and seed broadcasting
could be the absence of plant cover. Although this factor was not investigated in the
present study, it could be useful to consider in future studies. Vegetation structure in
the Shark Bay region contains recumbent shrubs under which small shrubs and climbers
grow (Brooker 2000). These ‗shrub clumps‘ may contain up to eight species (with
means ranging from 1.7 to 1.9 species depending on the vegetation type) (Brooker
2000). The growth of shrubs under other shrubs is termed facilitation (or nurse-protégé
interactions), and has been shown to be common in other arid environments (Pugnaire
and Lazaro 2000; Flores and Jurado 2003). Facilitation may provide shelter from
extreme temperatures and protection from herbivory (Flores and Jurado 2003). In

124
addition, the soil under shrubs may have more organic matter and soil nutrients and
better water retention than bare soil (Pugnaire et al. 1996). One study has found that
some species only recruit under old shrubs, even though they occur in the soil seed bank
under young shrubs, indicating that shrub age also has an effect on facilitation (Pugnaire
and Lazaro 2000). Hence, the lack of plant cover may have limited recruitment as some
species may benefit facilitation for germination and survival.
6.5.3 Conclusion
This study highlights the difficulties in returning plants to post-disturbed areas in an arid
region. ‗Borrowed‘ topsoil replacement did not seem to be a successful practice,
probably due to the low rainfall years experienced in the study, and hence warrants
further investigation. The study requires incorporation of a reference site in the natural
vegetation to benchmark the level of success or failure. Seedling emergence from seed
broadcasting was very low, and was dependant on year, season, soil ripping, and raking,
with much variation in species performance. Rainfall appeared to have the largest effect
on seedling emergence, and there were minimal effects soil ripping and raking when
broadcasting was followed by low rainfall.

125
C H A P T E R 7
Soil properties and greenstock establishment for
restoration of borrow pits
7.1 Abstract
Planting seedlings (greenstock) can be an effective method for revegetation or
restoration of post-mined areas. Treatments to optimise greenstock establishment (in
terms of condition, height and survival) of six species, in addition to soil properties
including soil nutrients, compaction (impedance), moisture and texture, was
investigated for post-mined sites in the arid zone at Shark Bay Salt. Soil nutrient status
differed between some sites, in particular the high level of electrical conductivity in pit
Q compared with the adjacent natural vegetation. Soil compaction was higher in
unripped areas of the borrow pits compared with the undisturbed vegetation, but this
compaction was somewhat alleviated by ripping. The greenstock experiments were
implemented in two parts: in 2005 four species and four treatments were investigated,
and in 2006, two focus species and 16 treatments were undertaken, with the survival of
an additional three species monitored. The treatments (singly and in combination)
included ripping, slow release fertiliser application, nutrient loading, pruning,
application of moisture retaining gel (wetta gel) and air pruning of roots (using modified
containers). There was little effect of treatment on condition, height and survival of
greenstock planted in 2005. However, ripping vastly improved the condition and
survival of Atriplex bunburyana planted in 2006, and fertiliser increased the height of
plants in the ripped area. Survival differed between species and between years with
survival of Acacia tetragonophylla planted in 2006 less than 20% after three months,
compared with 2005 where survival was 47-90% after five months. Atriplex
bunburyana seems to establish on post-mined areas effectively, with little need for
treatments apart from soil ripping. Further investigation is needed to optimise
establishment of the other species.

126
7.2 Introduction
In addition to topsoil and seed broadcasting (chapter 6), planting greenstock may assist
rehabilitation endeavours via increasing plant numbers in disturbed areas, particularly
key dominant taxa, where broadcast seeding is unreliable or topsoil seed bank
replacement is limited or unavailable. There has been no research undertaken on
greenstock establishment and survival at Shark Bay Salt (SBS). Techniques to optimise
greenstock survival that have been developed for restoration and forestry in the
southwest of Western Australia and around the world should be considered to determine
their success in arid areas of Western Australia.
Prior to planting, consideration of the soil environment is essential to enable plant
growth and ecosystem restoration following a major disturbance (Rokich 1999). Three
soil factors that may limit plant growth in disturbed areas are soil compaction (Ashby
1997), water availability (Lamont et al. 1993), and soil nutrient status (Paschke et al.
2000). Treatments such as soil ripping, and the application of moisture retaining gel
(wetta gel) and fertiliser may be needed to overcome these limitations and optimise
plant growth and survival. Additional treatments may be used to improve the growth
and survival of seedlings including shoot pruning, root pruning and nutrient loading.
During disturbance, borrow pits may have had up to several metres of soil removed and
it is likely that the remaining soil profile will have been compacted by heavy machinery.
The soil is likely to have little or no organic matter, limited nutrients and low water
infiltration capacity. Australian studies have noted that soil compaction (otherwise
known as impedance) appears to be associated to poor root development (Burrows
1982; Enright and Lamont 1992; Rokich et al. 2001) and high seedling mortality
(Enright and Lamont 1992). Compaction in rehabilitation sites is commonly alleviated
by ripping (Ward et al. 1996; Rokich et al. 2001; Szota et al. 2007) whereby
soils are ripped with a bulldozer with 2 m long tynes spaced 1.5 m apart (Koch et al.
1996) or with an excavator with a single 0.8 m long tyne (Rokich et al. 2001). Ripping
has been shown to increase plant growth (Ashby 1997) and improve root architecture
(Rokich et al. 2001).
An additional limitation to seedling survival is water availability. Given that the
rehabilitation site at SBS is in the arid zone, and rainfall can be low and variable, water

127
may be limiting. Also, seedlings grown in post-mined areas with high soil impedance
have shallow root systems, indicating that seedlings are dependant on shallow soil water
and are vulnerable to drought (Enright and Lamont 1992). Moisture retaining gel such
as ‗wetta gel‘ (also called polymer gel or hydrogel), a product that retains water, making
water available to the roots for longer, may assist plants when water is limited. For
instance, wetta gel increased survival from 49 to 82 days and increased growth of Pinus
halepensis seedlings under drought conditions in a greenhouse (Hüttermann et al.
1999). However, not all experiments have produced such positive results, particularly
those in field conditions. For example, wetta gel incorporated into the soil in a semi-
arid region of the US did not improve plant establishment from direct seeding (Paschke
et al. 2000). In addition, in a quarry restoration site in the Mediterranean, wetta gel did
not affect shoot growth of three native shrubs (Clemente et al. 2004).
As soil depth increases, organic matter (Williamson and Neilsen 2003) and organic
carbon (Schwenke et al. 2000) decreases. Organic matter and soil nutrients may be
limited in the borrow pits due to the large depth of soil removal. One particular study
stated that logging activities in Tasmanian forests can remove the surface leaf litter and
part or all of the topsoil exposing the subsoil (Williamson and Neilsen 2003). The study
conducted a glasshouse trial and found that seedling growth of three Eucalyptus species
planted in pots containing subsoils (10-30 cm depth) was lower than that in pots
containing topsoil (0-10 cm depth) and that growth of Eucalyptus globulus seedlings
planted in pots containing subsoil was increased with nitrogen + phosphorus fertiliser
(Williamson and Neilsen 2003). Hence, fertiliser application can be used to counteract
low nutrient availability and result in earlier commencement of shoot growth (Close et
al. 2005b). Nitrogen fertiliser has also been shown to increase the height of Pinus
radiata trees in a New Zealand plantation one year after planting, and nitrogen +
phosphorus fertiliser increased the height of trees after four years, but fertiliser
application did not affect survival (Simcock et al. 2006). However, Clemente et al.
(2004) found that fertiliser response was species specific, with slow release fertiliser
applied at planting increasing shoot growth after 1.5 years of only one of the three study
species planted in a quarry restoration site in the Mediterranean. Despite mixed
findings in other studies, the effect of fertiliser on growth and survival of seedlings in
borrow pits at SBS is worthy of investigation.

128
As well as fertiliser additions at planting, fertiliser regimes in the nursery can also affect
seedling performance after planting. Fertiliser is usually applied to prevent nutrient
deficiencies, but can also be applied in luxury levels (i.e. levels above that considered
sufficient) (Timmer 1997). Feeding seedlings with luxury levels of liquid fertiliser prior
to planting is a technique termed nutrient loading and can increase the nutrient
concentration within the plant without increasing its mass. Once planted, seedlings
exhibit greater growth. For example, black spruce seedlings that have been subjected to
nutrient-loading have been shown to contain ≥33% more N, P and K in their tissue, but
had a similar biomass compared with conventionally fertilised seedlings. After
planting, the seedlings exhibited more growth (44% taller and 37% more biomass)
compared with conventionally fertilised seedlings (Timmer 1997).
High root:shoot ratio at planting is beneficial for increasing growth (Close et al. 2005b).
Pruning shoots increases the root:shoot ratio and may decrease transpiration and
increase survival. Although manipulating the root:shoot ratio may seem advantageous,
field results can be variable. For example, Liquidambar styraciflua seedlings shoot-
pruned to 50% of their height had greater height growth compared with non-pruned
seedlings, three years after planting into sites in the southeastern United States
(McNabb and Vanderschaaf 2005). However, shoot pruning did not affect seedling
survival, and tree volume, height and ground-line diameter were lower than seedlings
that had not been pruned. Similarly, Zaczek et al. (1997) found that Quercus ruba
seedlings that were pruned when planted had a faster growth rate during the first year
after planting compared with unpruned seedlings in a plantation in Pennsylvania, USA
in spite of being initially shorter, the heights of pruned seedlings were similar to those
of unpruned seedlings after the second year. There was also no effect of pruning on
survival. The effect of shoot pruning seems to be commonly ascertained within a
forestry context, particularly in temperate areas, and the effect of shoot pruning on
plants in arid regions is yet to be tested.
Root morphology is important as it affects plant survival (Rokich et al. 2001), plant
stability and root growth (Burdett et al. 1983). Techniques of seedling production can
affect root morphology (Burdett et al. 1983). For instance, seedlings grown in
containers can have malformed roots, as lateral roots grow vertically, circle the
container, or grow to the bottom of the container and reach a barrier then grow upwards
(Zahreddine et al. 2004). In addition to physically pruning roots prior to planting to

129
remove the malformed roots (Zahreddine et al. 2004), modifying the container design
can help improve root morphology. Modifications may include the use of vertical
ridges, vertical slits (air pruned pots), or container interiors treated with a chemical
(Mullan and White 2001; Zahreddine et al. 2004). Vertical ridges prevent roots from
circling the container, which is beneficial, but lateral roots then grow vertically down
the sides of the container, and may continue growing vertically (instead of laterally)
when planted. The vertical growth of lateral roots may be prevented with air pruned
pots, where lateral roots grow through the vertical slits, and the ends desiccate and die
when they are exposed to the air. Similarly, chemicals such as cupric carbonate are
used to coat the container interiors and can inhibit lateral root growth. Growth of lateral
roots will then resume after seedlings are planted. Results of root pruning are variable,
for instance, three to four years after planting in British Columbia, heights of Pinus
contorta seedlings grown in chemically-treated containers were greater than those
grown in untreated containers (Burdett et al. 1983). In addition, the use of containers
with vertical slits did not affect seedling dry weight of Pinus nigra seedlings five
months after sowing, but chemically-treated containers increased shoot dry weights.
However, four months after transplanting Pinus nigra seedlings in Ohio, the use of
vertical slits and chemically treated containers did not affect plant dry weight
(Zahreddine et al. 2004). Also, physical root pruning did not have an effect on seedling
growth or survival of Liquidambar styraciflua seedlings in a plantation in the southeast
of the United States (McNabb and Vanderschaaf 2005).
Greenstock planting is one method that could be used to revegetate borrow pits at SBS.
Survival of greenstock may be low, and could be limited by changed soil properties
caused by disturbance. To optimise greenstock survival, treatments to enhance
establishment must be investigated. The aim of this chapter was to examine soil
properties and greenstock establishment and survival. Soil properties investigated
included soil compaction, water content and nutrient content. Management practices
investigated to improve seedling establishment and survival on site included (a) soil
ripping prior to planting, (b) nutrient-loading in the nursery, (c) shoot pruning prior to
planting, (d) application of slow-release fertiliser at planting, (e) application of wetta gel
at planting, and (f) the use of air pruned pots to raise seedlings.

130
7.3 Methods
7.3.1 Site description and preparation
Three borrow pits at SBS were chosen (pits P, Q and R) on the basis that they were
greater than 1ha (to ensure sufficient area for the experiments), had not recently been
subjected to rehabilitation works, and provided easy access for machinery (Appendix 1,
2). The three borrow pits were subjected to soil ripping and were fenced in April 2005,
and then re-ripped in May 2006 (as described in chapter 6). The experiment was
implemented in three parts. For part A, soil properties were measured. Parts B and C
describe establishment and survival of greenstock planted in 2005 (part B) and 2006
(part C).
7.3.2 Part A. Soil properties
Soil properties were measured at each of nine sites; in the ripped and unripped areas of
pits P, Q and R, and in the undisturbed vegetation adjacent to each of the pits.
Soil moisture and impedance
Gravimetric soil moisture was determined from five replicates of 400-500 g of soil at
each of the nine sites in May (autumn) 2007. Each replicate of soil was transferred into
in a plastic container and weighed immediately after collection and then again after
drying at 80oC for 1 week. Gravimetric soil moisture was determined using the
following formula:
Gravimetric soil moisture (θG) = ((mass of wet soil – mass of dry soil) / mass of dry soil) x 100
Volumetric soil moisture was measured in May (autumn) 2007, October (spring) 2007
and April (autumn) 2008 using a MP406 Moisture probe with a MPM 160 Moisture
Probe Meter. Ten readings were taken at each of the nine sites. Volumetric soil
moisture was converted to gravimetric soil moisture using equations fitted to calibration
curves for each soil collection location. The calibration curves were developed by
obtaining seven samples of each soil, adding 0-60 mL of water to each sample, testing
the volumetric and gravimetric soil moisture of each sample, then plotting the values
and fitting a curve. The equations for each soil are shown in Appendix 13.
Soil impedance was measured in May (autumn) 2007, October (spring) 2007 and April
(autumn) 2008 using a Rimik CP 20 Cone Penetrometer. The penetrometer measured

131
the force needed to penetrate the soil. The instrument was inserted into the soil and
readings were taken every 20 mm up to a maximum of 600 mm, or until it could not be
inserted into the soil any further. Ten replicate insertions were performed at each of the
nine sites.
Soil nutrient and texture analysis
Soil was collected in May 2007 from each of the nine sites at a depth of 0-5 cm and 5-
10 cm. Three replicates made up of five bulked samples were collected for each depth
and at each site. The soil was analysed by CSBP Limited (Cumming Smith British
Petroleum) for Ammonium, Iron, Nitrogen, organic Carbon, Phosphorus, Potassium,
Sulphur, electrical conductivity and pH (determined using CaCl2 or H2O).
Soil texture analysis was performed by CSBP on soil from a depth of 0-5 cm only from
each of the nine sites. One replicate of soil was used, and was made up from soil taken
from the three bulk replicates collected for nutrient analysis.
7.3.3 Part B. Greenstock experiments conducted in 2005
The four species chosen for seed broadcasting in 2005 (chapter 6) were also used for
greenstock production in 2005; Acacia tetragonophylla, Atriplex bunburyana, Rhagodia
baccata and Solanum orbiculatum. Seed pre-treatments are the same as described in
chapter 6. After pre-treatment, seeds were sown in 123 mm forestry tubes in late
February (summer) 2005 at the Kings Park Nursery. The seedlings were transported to
SBS in April (autumn) 2005.
The effect of planting seedlings in ripped soil and the addition of fertiliser at planting
was investigated individually (for two species) and in combination (for two species).
Soil was ripped in April 2005 (as described in chapter 6), and for the fertiliser
treatments, one teaspoon (4.4 g) of Osmocote® Native Gardens fertiliser (a slow release
fertiliser used for Australian plants) was applied after planting into a slit in the soil
approximately 10 cm from each seedling. Seedlings of two species (Acacia
tetragonophylla and Atriplex bunburyana) were subjected to four treatments a) control,
b) fertiliser at planting, c) planting in ripped soil and d) planting in ripped soil with
fertiliser. Seedlings of Rhagodia baccata were subjected to two treatments, a) control,
b) planting in ripped soil. Seedlings of Solanum orbiculatum were subjected to two

132
treatments, a) planting in ripped soil, b) planting in ripped soil with fertiliser. Seedlings
were planted in one pit (Pit R) in June (winter) 2005.
The experiment was implemented in a split-plot design with ripping as the whole-plot
and fertiliser as the sub-plot. For each treatment there were three replicates and each
replicate consisted of four rows. Ten seedlings of each species were planted per
replicate. Seedlings of all species were planted alternately within each replicate.
For all seedlings, survival, condition and height was assessed in November (spring)
2005 and May (autumn) 2006. Condition was ranked from 1 to 5 (1 = dead, 2 = 1-25%
living tissue, 3 = 26-50% living tissue, 4 = 51-75% living tissue, 5 = 76-100% living
tissue, with living tissue deemed as stems exhibiting evidence of being green and alive)
(Fig. 7.1). All living plants (condition rank 2-5) were considered as having survived.
7.3.4 Part C. Greenstock experiments conducted in 2006
To further investigate the effect of greenstock treatments, two focus species were
chosen for 2006, given the wider set of treatments and treatment combinations, and the
limited fenced area in which to conduct the experiment. The two focus species were
Acacia tetragonophylla and Atriplex bunburyana and they were chosen due to their
dominance in the vegetation, their ease of propagation, and the ability to collect a
sufficient amount of seeds. Seedlings were grown at The Permaculture Nursery in
Geraldton (seeds were sown in January 2006) and were transported to SBS in June
2006. Six treatments were investigated; ripping (rip), slow release fertiliser application
(fert), nutrient loading (NL), pruning (prune), wetta gel application (WG) and air
pruning of pots (AP). Given the number of available seedlings, some (but not all) of
these treatments were applied in combination. Six single treatments were applied in the
unripped area and ripped areas and four treatment combinations were applied to the
ripped area only for a total of 16 treatments (Table 7.1).
Table 7.1. Treatments in the unripped and ripped areas of the borrow pits.
Unripped
(single treatments)
Ripped
(single treatments)
Ripped
(treatment combinations)
control control fert + NL
fert fert fert + prune
NL NL NL + prune
prune prune fert + NL + prune
AP AP
WG WG

133
For the fertiliser treatment, slow release Osmocote® Native Gardens fertiliser was
applied as described previously. Nutrient loading was undertaken between 18 May and
9 June 2006 where seedlings were subjected to twice weekly applications of nitrogen
fertiliser (4.4% urea and 2.2% blood) at a rate of 1:200 fertiliser:water for a total of
seven applications. For the pruning treatment, seedlings were pruned to around half
their initial size using secateurs just prior to planting. One teaspoon of Wetta gel
(Rainsaver water storing crystals, Hortex Australia Pty Ltd.) was applied in a similar
way to the slow release fertiliser. Air pruned pots were constructed by cutting two
parallel slits down each side of the pot (Fig. 7.2).
In July (winter) 2006, seedlings were planted using mechanised tree planters (potti
putkis) in the three pits (P, Q and R). Three replicates of 20 seedlings were planted in
each treatment. There was one row per treatment, and Acacia tetragonophylla and
Atriplex bunburyana were planted alternately in the row. Each replicate was
implemented in a block. Seedlings were unable to be planted in the un-ripped area at pit
P because the soil was impenetrable.
Condition and height was assessed on 29 October (spring) 2006, 1 May (autumn) 2007,
30 October (spring) 2007 and 16 April (autumn) 2008. Condition was ranked as
described previously (Fig. 7.1). All living plants (condition rank 2-5) were considered
as having survived.
Figure 7.1. Condition rating of Atriplex bunburyana seedlings from 1 to 5 (1 = dead, 2
= 1-25% living tissue, 3 = 26-50% living tissue, 4 = 51-75% living tissue, 5 = 75-100%
living tissue).
Figure 7.2. Root system of Atriplex bunburyana seedlings grown in conventional (left)
and air-pruned pots (right).
1 2
3 4 5

134
Opportunistically, seedlings of Solanum orbiculatum that were raised from seed by The
Permaculture Nursery in Geraldton and seedlings of Thryptomene baeckeacea and
Triodia plurinervata that were propagated from cuttings at the Kings Park nursery were
planted to assess their potential as planted greenstock material. Given the plant
availability, three replicates of 20 seedlings of each species were planted at pits Q and R
only in July (winter) 2006 and were not subjected to any treatments. Survival was
assessed on 29 October (spring) 2006, 1 May (autumn) 2007, 30 October (spring) 2007
and 16 April (autumn) 2008.
7.3.5 Statistics
For part A, a two way ANOVA was used to detect site differences in soil moisture and
maximum penetration depth with Fisher‘s unprotected LSD (using a significance level
of 0.05) used to separate the means. For part B, height, condition and survival were
analysed for each species individually using a one way ANOVA to determine if there
was an effect of treatment. If there was an effect, Fisher‘s unprotected LSD (using a
significance level of 0.05) was used to separate the means of the treatments. For part C,
condition, height and survival of Atriplex bunburyana and survival of Acacia
tetragonophylla was analysed using a general analysis of variance (ANOVA) to
determine whether there were significant effects of treatment or pit across the
assessment dates. Height of Acacia tetragonophylla was not analysed because heights
of dead plants were not recorded, and as there was low survival, there were
consequently many missing values. Greenstock planted in the ripped and unripped
areas were analysed separately due to the unbalanced experimental design (i.e. in the
un-ripped areas, only single treatments were used and not treatment combinations, also
greenstock was not planted in the unripped area of pit P). To determine differences
between pits across the assessment dates, a two way ANOVA with Fisher‘s unprotected
LSD (using a significance level of 0.05) was used to separate the means. The
effectiveness of the treatments was analysed at the final scoring time (autumn 2008).
To determine differences between treatments, a one way ANOVA was performed on
ripped and unripped areas separately and Fisher‘s unprotected LSD (using a
significance level of 0.05) was used to separate the means. For parts B and C,
percentage (survival) data was arcsine transformed prior to analysis but untransformed
data is presented in the figures.

135
7.4 Results
7.4.1 Part A. Soil properties
Soil moisture and impedance
Soil moisture trends were similar for both volumetric and gravimetric methods but soil
moisture percentages were generally higher using the volumetric method compared to
the gravimetric method (Fig. 7.3, 7.4a). There were differences in soil moisture
between pits, with gravimetric soil moisture higher in Pit Q compared with the adjacent
natural vegetation, and the other two pits in May 2007 (Fig. 7.3). Soil moisture
determined volumetrically in May 2007 was also highest in pit Q, both in the ripped and
unripped areas, with the other sites exhibiting similar soil moisture (Fig. 7.4). These
trends were similar across each measurement time of May 2007, October 2007 and
April 2008 (Fig. 7.4a, b, c).
Pit
P Q R
So
il m
ois
ture
(%
)
0
2
4
6
8
10 Rip
No rip
Nat Veg
a
c
a
dd
c
abbc c
Figure 7.3. Gravimetric soil moisture in the ripped and unripped areas of borrow pits P,
Q and R and the surrounding natural vegetation in May 2007. Bars indicate standard
error. Letters indicate significant (P<0.05) differences between the means.
a) May 2007
P Q R
So
il m
ois
ture
(%
)
0
5
10
15
20b) October 2007
Pit
P Q R
c) April 2008
P Q R
Rip
No Rip
Nat Veg
bcb
a
d
d
bcbc bc c bc
aabc
d
e
ababc
c bca
bab
b
d
ab abbc
c
Figure 7.4. Soil moisture determined volumetrically then converted to gravimetric soil
moisture in a) May 2007, b) October 2007 and c) April 2008 in the ripped (Rip) and
unripped (No Rip) areas of borrow pits P, Q and R and the surrounding natural
vegetation (Nat Veg). Bars indicate standard error. Letters indicate significant
(P<0.05) differences between the means within each graph.

136
Fig
ure
7.5
. S
oil
im
ped
ance
(K
Pa)
in M
ay 2
007,
Oct
ober
2007
and
Apri
l 20
08 i
n t
he
ripped
(R
ip)
and u
nri
pped
(N
o R
ip)
area
s at
Pit
s P
, Q
and R
,
and n
atura
l v
eget
atio
n (
Nat
Veg
) ad
jace
nt
to t
he
pit
s.
Aver
ages
wer
e only
cal
cula
ted w
hen
ther
e w
ere
at l
east
thre
e re
adin
gs
at t
hat
dep
th.
Bar
s
indic
ate
stan
dar
d e
rro
r
April 2008
Pit P Pit Q Pit R
Octo
ber
2007
Depth
(m
m)
0100
200
300
400
500
600
May 2
007
0
1000
2000
3000
4000
Soil impedance (KPa)
0
1000
2000
3000
4000
0100
200
300
400
500
600
0
1000
2000
3000
4000
0100
200
300
400
500
600
Rip
N
o R
ip
Nat
Veg

137
Soil impedance in the top 120 mm of the soil profile in the ripped areas of pits Q and R
was similar to the adjacent natural vegetation, whereas the unripped areas had higher
soil impedance. However, at pit P, the ripped and unripped areas showed similar soil
impedance, and they were higher than the adjacent vegetation (Fig. 7.5). Beyond 120
mm, there was generally no penetration in the unripped areas, and if there was, soil
impedance was higher than the ripped sites (Fig. 7.5). The highest maximum
penetration depth was recorded in the natural vegetation, followed by the ripped areas,
with the unripped areas having the lowest depth of penetration (Fig. 7.6). There were
few differences between the pits, except that in May 2007, the maximum penetration
depth was lower in the ripped and unripped areas in pit P compared with the ripped and
unripped areas in Pits Q and R (Fig. 7.6a).
a) May 2007
P Q R
De
pth
(m
m)
0
200
400
600b) October 2007
Pit
P Q R
c) April 2008
P Q R
Rip
No Rip
Nat Veg
b
a
d
c
b
e
c
b
cd
b
a
d
c
a
d
bc
a
d b
a
c
b
a
c
b
a
c
Figure 7.6. Maximum penetration depth (mm) in a) May 2007, b) October 2007 and c)
April 2008 in the ripped and unripped at Pits P, Q and R, and natural vegetation
adjacent to the pits. Bars indicate standard error. Letters indicate significant (P<0.05)
differences between the means within each graph.

138
g) Conductivity
dS
/m
0
5
10
15
b) Iron
mg
/kg
0
50
100
150Rip
No Rip
Nat Veg
f) Sulphur
mg
/kg
0
200
400
600
800
1000
1200e) Potassium
mg
/kg
0
200
400
600
800
1000
1200
d) Phosphorus
mg
/kg
0
10
20
30
40
a) Ammonium
mg
/kg
0
1
2
3
4
c) Nitrogen
mg
/kg
0
5
10
15
20
h) Organic Carbon
% O
rgan
ic c
arb
on
0.0
0.2
0.4
0.6
0.8
1.0
j) pH (CaCl2)
pH
0
2
4
6
8
10
12
5-100-5 5-10 0-5 0-5 5-10P Q R
i) pH (H2O)
pH
0
2
4
6
8
10
12
5-100-5 5-10 0-5 0-5 5-10P Q R
Figure 7.7. Soil nutrient analysis at 0-5 cm and 5-10 cm in the ripped (Rip) and
unripped (No Rip) areas in pits P, Q and R, and the natural vegetation (Nat Veg)
adjacent to those areas. Bars indicate standard error.

139
Soil nutrient and texture analysis
High electrical conductivity values and high amounts of potassium and sulphur were
found in the ripped and unripped areas of pits P and Q compared with the adjacent
natural vegetation and pit R (Fig. 7.7e, f, g). There was also a lower percentage of
organic carbon in the ripped and unripped areas of pits P and R compared with the
adjacent natural vegetation (Fig. 7.7h).
Soil texture was similar at pits P and Q, with each pit containing a large percentage of
coarse sand (Fig. 7.8). The soil at pit R had a lower amount of coarse sand, but more
fine sand compared with the other two pits. There seemed to be little differences
between the three sites (ripped, unripped and natural vegetation).
Perc
ent
of
tota
l sam
ple
0
20
40
60
80
100
Coarse sand
Fine sand
Silt
Clay
rip no rip nat veg rip no rip nat veg rip no rip nat veg
P Q R Figure 7.8. Soil texture in the ripped (rip) and unripped (no rip) areas in pits P, Q and
R, and the natural vegetation (nat veg) adjacent to those areas. Soil texture is indicated
by percentage of coarse sand (200-2000 µm), fine sand (20 - 200 µm), silt (>2 - <20
µm) and clay (<2 µm).

140
7.4.2 Part B. Greenstock experiments conducted in 2005
Five months after planting, the treatments (control, fertiliser only, rip only or fertiliser +
rip) did not have any effect on condition of three of the species, but condition of
Atriplex bunburyana was higher when planted in ripped areas with fertiliser, compared
with the other treatments (Fig. 7.9a). The treatments did not have any effect on height
or survival of the species (Fig. 7.9a, 7.10). Survival ranged from 47-90% with Solanum
orbiculatum (90%) exhibiting the highest survival after 5 months, followed by Atriplex
bunburyana (63-80%), Rhagodia baccata (67-77%) and Acacia tetragonophylla (47-
71%) (Fig. 7.10a). Eleven months after planting, survival decreased, ranging from 3-
50% (Fig. 7.10b) and there was no significant differences between treatments for each
species.
b) Height
A. tetra
gonophylla
A. bunburyana
R. baccata
S. orbiculatum
Heig
ht (c
m)
0
1
2
3
4
5
a) Condition
A. tetra
gonophylla
A. bunburyana
R. baccata
S. orbiculatum
Conditio
n
0
1
2
3
4
5Control
Fert
Rip
Rip + Fert
aa a
b
Figure 7.9. a) Condition rank (1-5) and b) height of seedlings of Acacia
tetragonophylla, Atriplex bunburyana, Rhagodia baccata and Solanum orbiculatum 5
months after planting and subjected to one of four treatments; control, fertiliser only
(Fert), rip only (Rip), rip + fertiliser (Rip + Fert) (Rhagodia baccata was only planted in
the control and rip treatments, and Solanum orbiculatum was only planted in the control
and fertiliser treatments). Condition was ranked from 1 to 5 (1 = dead, 2 = 1-25% living
tissue, 3 = 26-50% living tissue, 4 = 51-75% living tissue, 5 = 75-100% living tissue).
Bars indicate standard error. Letters indicate significant (P<0.05) differences between
the means within each species. An absence of letters indicates that there were no
differences between treatments within each species.

141
a) 5 months
A. tetra
gonophylla
A. bunburyana
R. baccata
S. orbiculatum
Surv
ival (%
)
0
20
40
60
80
100b) 11 months
A. tetra
gonophylla
A. bunburyana
R. baccata
S. orbiculatum
Control
Fert
Rip
Rip + Fert
Figure 7.10. Survival of seedlings of Acacia tetragonophylla, Atriplex bunburyana,
Rhagodia baccata and Solanum orbiculatum a) 5 months and b) 11 months after
planting and subjected to one of four treatments; control, fertiliser only, rip only, rip +
fertiliser (Rhagodia baccata was only planted in the control and rip treatments, and
Solanum orbiculatum was only planted in the control and fertiliser treatments). Bars
indicate standard error. Letters indicate significant (P<0.05) differences between the
means within each species. An absence of letters indicates that there were no
differences between treatments within each species.

142
7.4.3 Part C. Greenstock 2006
Seedlings of two species (Atriplex bunburyana and Acacia tetragonophylla were
planted in ripped and unripped areas of three pits, and subjected to five treatments
(fertiliser, nutrient loading, pruning, wetta gel and air pruned pots). Condition of
Atriplex bunburyana seedlings was lower in the unripped areas (<2) compared with the
ripped areas (>2) (Fig. 7.11, 7.12, 7.13). Season (assessment date), treatment and pit
significantly (P<0.05) affected the condition of Atriplex bunburyana planted in both the
ripped and unripped areas. Condition fluctuated across the seasons in the study period
in the ripped areas and was generally lower in autumn compared with spring (Fig.
7.11b, c). However, this was not observed at each pit, for example, condition in pit Q
was only lower in autumn 2008, and not autumn 2007, and condition in pit R was only
lower in autumn 2007 and not autumn 2008 (Fig. 7.12b). There were differences
between treatments in both the unripped and ripped areas. There was no difference in
condition between the control and any of the treatments, however condition was higher
in the fertiliser treatment compared with two treatments in the unripped areas (prune,
AP) and two treatments in the ripped areas (NL+prune, NL+fert+prune) (Fig 7.13).
There were differences in condition between the pits. In the unripped areas, condition
was significantly (P<0.05) higher in pit R compared with pit Q in all seasons except
autumn 2007 (Fig. 7.12a). In the ripped areas, differences between pits depended on the
season, with condition being higher in pits Q and R compared with pit P in spring 2006,
and higher in pit Q compared with pits P and R in autumn 2007 (Fig. 7.12b).
There was a significant (P<0.05) effect of season, treatment and pit on the height of
Atriplex bunburyana seedlings in both the unripped and ripped areas. Height increased
over the study period (Fig. 7.14, 7.15). Seedlings in the fertiliser treatment were
significantly (P<0.05) taller than those in the control, in both the unripped and ripped
areas (Fig. 7.16). Seedlings that were pruned were significantly (P<0.05) shorter than
the controls (Fig. 7.16) (due to the fact that they were pruned). There were few
differences between the heights of seedlings across the three pits, except that seedlings
were taller in pit Q compared with pit R in autumn 2007 (unripped and ripped areas)
and autumn 2008 (ripped area only) (Fig. 7.15).
Survival of Atriplex bunburyana seedlings decreased over the study period (Fig. 7.17,
7.18). Most of the deaths occurred between planting (in winter) and spring 2006 (for all
pits), and further deaths occurred prior to autumn 2007 (at pit R). Survival remained

143
relatively stable after autumn 2007, except for pit Q in autumn 2008 where survival
dropped to <30% (Fig. 7.18). Ripping also had an effect on survival, as survival was
lower in the unripped areas compared with the ripped areas. For instance, in autumn
2007 in pits Q and R, survival in the unripped areas was <40% whereas in the ripped
areas it was >60% (Fig. 7.18). There was no effect of treatment on survival in the
unripped areas, whilst in the ripped areas, all treatments had the same or lower survival
compared with the control (Fig. 7.19) with survival in three treatments (NL+fert,
fert+prune and NL+fert+prune) significantly (P<0.05) lower than the control. There
were some differences in survival between the three pits. In spring 2006, survival was
highest in pit R, followed by pit Q, then pit P, however, these differences became less
pronounced over time (Fig. 7.18), the exception being pit Q in autumn 2008 as
previously mentioned.
Unlike Atriplex bunburyana, Acacia tetragonophylla seedlings had very low survival in
all treatments at all sites. Generally, most plants died between the time of planting in
winter 2006 and monitoring in spring 2006 (Fig. 7.20, 7.21). There were no differences
between treatments in autumn 2008 within the ripped areas, whilst in the unripped
areas, survival of the wetta gel treatment was higher than the other treatments (Fig.
7.22). In the ripped areas, there were differences between pits. In most (but not all)
seasons, survival was highest in pit R, followed by pit P, then pit Q (Fig. 7.21). There
was little effect of treatments on height and condition (data not shown).
There were differences in survival of Atriplex bunburyana and Acacia tetragonophylla
depending on the planting year. Survival of Atriplex bunburyana seedlings planted in
ripped areas in 2005 and 2006 was generally high (>60%) in the first spring after
planting (Fig. 7.10a, 7.17). However survival of Atriplex bunburyana seedlings planted
in 2006 was higher (>60%) than those planted in 2005 (<60%) (Fig. 7.10b, 7.17).
Conversely, survival of Acacia tetragonophylla seedlings planted in 2005 was higher
(>40%) than seedlings planted in 2006 (<20%) (Fig. 7.10a, 7.20). After the first
summer, survival of Acacia tetragonophylla seedlings planted in 2005 was similar to
survival of seedlings planted in 2006 (<20%) (Fig. 7.10b, 7.20).
Condition of Solanum orbiculatum, Thryptomene baeckeacea and Triodia plurinervata
was generally low (<3), and declined from spring 2007 to autumn 2008 (Fig. 7.23).
Survival of the three species was 28% - 58% in the first spring, then decreased to 3% -
18% after summer (Fig. 7.24).

144
b) Rip, single treatments
Control
NL
Fert
Prune
AP
WG
c) Rip, multiple treatments
Fert + NL
NL + Prune
Fert + Prune
Fert + NL + Prune
Spring2006
Autumn2007
Autumn2008
Spring2007
Spring2006
Autumn2007
Autumn2008
Spring2007
a) No ripC
onditio
n
0
1
2
3
4
5Control
NL
Fert
Prune
AP
WG
Spring2006
Autumn2007
Autumn2008
Spring2007
Figure 7.11. Condition of Atriplex bunburyana seedlings planted in pits P, Q and R in
a) unripped and b) ripped areas and subjected to 6 treatments; control, nutrient loading
(NL), fertiliser (Fert), pruning (Prune), air pruned pots (AP), wetta gel (WG), and in c)
ripped areas with various combinations of these treatments. Values are averages across
the three pits.
a) No rip
Spring 06 Autumn 07 Spring 07 Autumn 08
Conditio
n
0
1
2
3
4
5b) Rip
Spring 06 Autumn 07 Spring 07 Autumn 08
P
Q
R
a
d
abb
a
cd
a
c
cd
ef ef
bc
ef
b
ef
f efde
a
ef
Figure 7.12. Condition of Atriplex bunburyana seedlings planted in a) unripped areas of
Q and R and b) ripped areas of pits P, Q and R. Values are averages across the
treatments within the ripped and unripped areas. Bars indicate standard error. Letters
indicate significant (P<0.05) differences between the means within each graph.
a) No rip
Conditio
n
0
1
2
3
4
5b) Rip
Control NL Fert Prune Air
prune
Wettagel
Control NL Fert Prune Air
prune
Wettagel
NL +
Fert
NL +
Prune
Fert +
Prune
NL +Fert +Prune
abc
abc
c
abcabc
abc
c
a ab
bc
abb b
a a
b
Figure 7.13. Condition of Atriplex bunburyana seedlings in autumn 2008 planted in a)
unripped and b) ripped areas. Values are averaged across the three pits. Bars indicate
standard error. Letters indicate significant (P<0.05) differences between the means
within each graph.

145
b) Rip, single treatments
Control
NL
Fert
Prune
AP
WG
a) No rip
Heig
ht (c
m)
0
10
20
30
40
Control
NL
Fert
Prune
AP
WG
c) Rip, multiple treatments
Fert + NL
NL + Prune
Fert + Prune
Fert + NL + Prune
Spring2006
Autumn2007
Autumn2008
Spring2007
Spring2006
Autumn2007
Autumn2008
Spring2007
Spring2006
Autumn2007
Autumn2008
Spring2007
Figure 7.14. Height of Atriplex bunburyana seedlings planted in pits P, Q and R in a)
unripped and b) ripped areas and subjected to 6 treatments; control, nutrient loading
(NL), fertiliser (Fert), pruning (Prune), air pruned pots (AP), wetta gel (WG), and in c)
ripped areas with various combinations of these treatments. Values are averages across
the three pits.
a) No rip
Spring 06 Autumn 07 Spring 07 Autumn 08
Heig
ht (c
m)
0
10
20
30
40b) Rip
Spring 06 Autumn 07 Spring 07 Autumn 08
P
Q
R
aba
d
a
bcdab
cd
bc
abcabcd
a
bcde
ef
ab
deef
bcdee
f
cde
Figure 7.15. Height of Atriplex bunburyana seedlings planted in a) unripped areas of Q
and R and b) ripped areas of pits P, Q and R. Values are averages across the treatments
within the ripped and unripped areas. Bars indicate standard error. Letters indicate
significant (P<0.05) differences between the means within each graph.
b) Ripa) No rip
Heig
ht (
cm
)
0
10
20
30
40
cd cd
e
a
de
a
bc
ab
cde de
Control NL Fert Prune Air
prune
Wettagel
Control NL Fert Prune Air
prune
Wettagel
NL +
Fert
NL +
Prune
Fert +
Prune
NL +Fert +Prune
bc
c
b
a
bccd
Figure 7.16. Height of Atriplex bunburyana seedlings in autumn 2008 planted in a)
unripped and b) ripped areas. Values are averaged across the three pits. Bars indicate
standard error. Letters indicate significant (P<0.05) differences between the means
within each graph.

146
a) No ripS
urv
ival
0
20
40
60
80
100 Control
NL
Fert
Prune
AP
Wetta gel
Spring
2006
Autumn
2007
Autumn
2008
Spring
2007
Winter
2006
b) Rip, single treatments
Control
NL
Fert
Prune
AP
WG
c) Rip, multiple treatments
Fert + NL
NL + Prune
Fert + Prune
Fert + NL + Prune
Spring
2006
Autumn
2007
Autumn
2008
Spring
2007
Winter
2006
Spring
2006
Autumn
2007
Autumn
2008
Spring
2007
Winter
2006 Figure 7.17. Survival of Atriplex bunburyana seedlings planted in pits P, Q and R in a)
unripped and b) ripped areas and subjected to 6 treatments; control, nutrient loading
(NL), fertiliser (Fert), pruning (Prune), air pruned pots (AP), wetta gel (WG), and in c)
ripped areas with various combinations of these treatments. Survival was monitored in
November (Spring) 06, May (Autumn) 07, November (Spring) 07 and April (Autumn)
08.
a) No rip
Spring 06 Autumn 07 Spring 07 Autumn 08
Surv
ival (%
)
0
20
40
60
80
100b) Rip
Spring 06 Autumn 07 Spring 07 Autumn 08
P
Q
R
a
c
a
b
a
b
a
b
bcd
e
f
bcd
ede
bc
cde cde
b
a
bcde
Figure 7.18. Survival of Atriplex bunburyana seedlings planted in a) unripped areas of
Q and R and b) ripped areas of pits P, Q and R. Values are averages across the
treatments within the ripped and unripped areas. Bars indicate standard error. Letters
indicate significant (P<0.05) differences between the means within each graph.
a) No rip
Surv
ival (%
)
0
20
40
60
80
100b) Rip
c
abc
c
abc
c
aab
a
abc
bc
Control NL Fert Prune Air
prune
Wettagel
Control NL Fert Prune Air
prune
Wettagel
NL +
Fert
NL +
Prune
Fert +
Prune
NL +Fert +Prune
Figure 7.19. Survival of Atriplex bunburyana seedlings in autumn 2008 planted in a)
unripped and b) ripped areas. Values are averaged across the three pits. Bars indicate
standard error. Letters indicate significant (P<0.05) differences between the means
within each graph.

147
a) No rip
Surv
ival (%
)
0
20
40
60
80
100 Control
NL
Fert
Prune
AP
WG
Spring
2006
Autumn
2007
Autumn
2008
Spring
2007
Winter
2006
b) Rip, single treatmentsControl
NL
Fert
Prune
AP
WG
c) Rip, multiple treatmentsFert + NL
NL + Prune
Fert + Prune
Fert + NL + Prune
Spring
2006
Autumn
2007
Autumn
2008
Spring
2007
Winter
2006
Spring
2006
Autumn
2007
Autumn
2008
Spring
2007
Winter
2006 Figure 7.20. Survival of Acacia tetragonophylla seedlings planted in pits P, Q and R in
a) unripped and b) ripped areas and subjected to 6 treatments; control, nutrient loading
(NL), fertiliser (Fert), pruning (Prune), air pruned pots (AP), wetta gel (WG), and in c)
ripped areas with various combinations of these treatments. Values are averages across
the three pits.
a) No rip
Spring 06 Autumn 07 Spring 07 Autumn 08
Surv
ival (%
)
0
10
20
30
40b) Rip
Spring 06 Autumn 07 Spring 07 Autumn 08
P
Q
R
a ab a ab a ab aab
bcb
d
b
a
bc
b
a
cd
b
a
cd
Figure 7.21. Survival of Acacia tetragonophylla seedlings planted in a) unripped areas
of Q and R and b) ripped areas of pits P, Q and R. Values are averages across the
treatments within the ripped and unripped areas. Bars indicate standard error. Letters
indicate significant (P<0.05) differences between the means within each graph.
a) No rip
Surv
ival (%
)
0
10
20
30
40b) Rip
Control NL Fert Prune Air
prune
Wettagel
Control NL Fert Prune Air
prune
Wettagel
NL +
Fert
NL +
Prune
Fert +
Prune
NL +Fert +Prune
a a a a ab
Figure 7.22. Survival of Acacia tetragonophylla seedlings in autumn 2008 planted in a)
unripped and b) ripped areas. Values are averaged across the three pits. Bars indicate
standard error. Letters indicate significant (P<0.05) differences between the means
within each graph.

148
a) Pit Q
Spring 07 Autumn 07
Conditio
n
0
1
2
3
4
5b) Pit R
Spring 07 Autumn 07
0
1
2
3
4
5S. orbiculatum
T. pleurinervata
T. baeckeacea
b bab
aa a
bc bc
c
ab aba
Figure 7.23. Condition of Solanum orbiculatum, Thryptomene baeckeacea and Triodia
plurinervata planted at a) pit Q and b) pit R. Bars indicate standard error.
a) Pit Q
Spring 06 Autumn 07
Surv
ival
0
20
40
60
80
100b) Pit R
Spring 06 Autumn 07
0
20
40
60
80
100 S. orbiculatum
T. pleurinervata
T. baeckeacea
b b
ab
a ab
a
bcc
c
ab ab
a
Figure 7.24. Survival of Solanum orbiculatum, Thryptomene baeckeacea and Triodia
plurinervata planted at a) pit Q and b) pit R. Bars indicate standard error.

149
7.5 Discussion
This study has highlighted the importance of soil ripping to alleviate soil impedance and
maximise greenstock survival. There was little effect of various greenstock treatments
on the condition, growth and survival of two focus and five additional species, apart
from plant pruning, which produced consistently lower survival than any other
treatment. There were differences between the two focus species, as Atriplex
bunburyana exhibited higher survival than Acacia tetragonophylla.
7.5.1 Greenstock 2006
Soil ripping effects on greenstock and soil impedance
Condition and survival of greenstock was higher in ripped than unripped areas. Soil
ripping had a reduced effect on Acacia tetragonophylla compared with Atriplex
bunburyana, as overall survival was very low for Acacia tetragonophylla. Soil ripping
is common practice in other post-mined areas in Western Australia (Ward et al. 1996;
Gardner 2001; Rokich et al. 2001; Szota et al. 2007) and is clearly effective in the post-
disturbed area in this study. Other studies have shown that ripping increases survival of
Pinus echinata (Gwaze et al. 2007), Pinus radiata (Simcock et al. 2006) and Acacia
hemiteles (Yates et al. 2000) seedlings. However, in a degraded woodland site in
southwestern Australia, ripping did not increase survival of Atriplex semibaccata
seedlings two years after planting (Yates et al. 2000), possibly as the seedlings were
planted into a woodland site, likely to have been compacted by animal grazing, unlike in
mined areas in which compaction is commonly caused by heavy machinery.
Soil ripping may have resulted in higher survival in this study as it alleviates
compaction (hence reduces bulk density), increases saturated hydraulic conductivity
(therefore increasing the water infiltration capacity) (Luce 1997) and reduces soil
impedance (allowing plant roots to penetrate deeper into the soil) (Rokich et al. 2001).
Bulk density and hydraulic conductivity were not measured in this study, but soil
impedance differed between the ripped and unripped areas in the three pits, and the
adjacent natural vegetation. Ripping reduced the impedance compared to the unripped
areas in two pits and increased the maximum penetration depth in all pits. These
findings are similar to those of Rokich et al. (2001) who reported lower soil impedance
and greater penetration depth in ripped areas compared with unripped areas in a post-
mined site in Banksia woodland. In the present study, the maximum depth of

150
penetration was greater in the natural vegetation compared with the pits, unlike Rokich
et al. (2001), where there was no difference in maximum penetration depth between the
undisturbed and ripped soils. Hence, in this study, higher survival in ripped areas
compared with unripped areas can be partly attributed to the alleviation of soil
impedance and the increase in the maximum penetration depth which would allow
improved root growth and development.
While soil ripping increased survival, there seemed to be little difference between the
heights of plants in ripped soil compared with unripped areas. This finding contrasts
with other studies, such as a post-mined area in Midwest America where seedlings were
taller when planted in ripped plots compared to unripped plots (Ashby 1997). Also, two
years after planting into ripped areas in a degraded woodland in southwestern Australia,
Acacia hemiteles, Atriplex semibaccata, Eucalyptus salmonophloia and Maireana
brevifolia seedlings were taller than those planted into unripped areas (Yates et al.
2000).
Soil moisture and nutrients
Soil moisture and nutritional status also differed between the sites, and appeared to have
an effect on plant survival and condition. Soil moisture differed between the three pits
and was greater in two of the borrow pits compared with the adjacent natural vegetation.
This study had similar findings to research conducted in Banksia woodland in southwest
Australia where the authors reported that soil moisture was higher in reconstructed soils
in post-mined areas compared with adjacent undisturbed vegetation (Rokich et al.
2001). However, that study found the soil moisture was lower in ripped areas compared
with unripped areas, whereas this study found similar soil moisture in ripped and
unripped areas. In the present study, there were differences in soil moisture between the
three pits. These differences may not be due to soil texture, as the texture analysis
showed similar percentages of sand, silt and clay in the pits and the adjacent natural
vegetation. However, there was a difference in the topography between the pits, with
pit Q having the lowest elevation (hence closest to the water table), then pit P, followed
by pit R. Differences in soil moisture were reflected in plant condition. Atriplex
bunburyana plants were in better condition in pit Q compared with pits P and R in
autumn 2007. Pit Q had higher soil moisture than the other pits, which may have kept
the plants in better condition over the summer. However, survival of seedlings in pit Q
decreased between spring 2007 and autumn 2008, and this is likely to be due to

151
inundation during the heavy rainfall event in March 2008 (308 mm of rain in four days
– Fig. 6.2).
Soil nutrient analysis showed that the concentration of many nutrients was similar in the
natural vegetation and the pits. There were some notable differences, for example,
higher organic carbon percentages were found in the natural vegetation compared with
pits P and R, similar to Rokich (1999). Organic carbon content is highly important, it
can be used as a key measure of soil formation, and therefore used to assess restoration
success (Koch and Hobbs 2007). Clearly, the difference between organic carbon
content of the disturbed and undisturbed soils indicates that restoration is still ongoing.
In addition, electrical conductivity was substantially higher in pits P and Q compared
with the natural vegetation. The soil in pit P (2-3 dS m-1
) is considered to be slightly
saline, and the soil in pit Q (10 dS m-1
) is very saline (DAFWA 2006). Growth of many
plants is limited at high salt concentrations, however, saline soils can be beneficial for
other plants (Barrett-Lennard et al. 2003) such as saltbushes (Atriplex spp.) as they have
adapted to saline soils. For example, growth of Atriplex amnicola increases by 10%
when grown at 5 dS m-1
(Barrett-Lennard et al. 2003). Similarly, in another study,
growth of Atriplex bunburyana was not affected by soil salinity up to 5 dS m-1
(Jefferson 2001). In this study, survival of Acacia tetragonophylla was lowest in pit Q
(0% survival after 1 year) which may be due to the high level of salinity in that pit.
Atriplex bunburyana did not seem to be adversely affected by the electrical conductivity
in pit Q.
Greenstock treatments
Fertiliser application had a positive effect on growth. Seedlings that were provided with
fertiliser were taller than their control counterparts two years after planting. In another
study, fertiliser (phosphate rock) applied three months after planting increased the
height of Pinus radiata seedlings in a New Zealand plantation after one and four years
(Simcock et al. 2006). Also, slow release fertiliser applied at planting increased the
height of Ceratonia siliqua seedlings in a Mediterranean quarry, 1.5 years after planting
(Clemente et al. 2004). However, in this study, fertiliser was not beneficial to condition
or survival. This is consistent with studies that did not find an effect of fertiliser on
survival of P. radiata (Simcock et al. 2006) and eight species planted in Banksia
woodland in southwestern Australia (Turner et al. 2005b).

152
In contrast to fertiliser, the other greenstock treatments (air pruning, wetta gel and
nutrient loading) were not beneficial to condition, growth or survival. There was no
effect of the use of air pruned pots on seedling condition, height and survival.
Similarly, Pinus nigra seedlings grown in containers with vertical slits (air pruned pots)
had comparable growth to those grown unmodified pots, four months after planting
(Zahreddine et al. 2004). In this study root morphology of seedlings grown in
conventional and air pruned pots was not assessed. It is possible that seedlings grown
in conventional pots had well formed root systems, and air pruned pots did not further
improve root morphology.
The finding that water-holding polymer (wetta gel) did not affect survival corresponds
to that of Paschke et al. (2000) who found no effect of wetta gel on survival of seedlings
of eight species. Paschke et al. (2000) suggested that the use of fertiliser may be more
important than improvements in the availability of soil moisture. In addition, water-
holding polymer gel did not affect growth of three species in a Mediterranean post-
mined site (Clemente et al. 2004). Further investigation is needed into the reasons why
the addition of wetta gel was not beneficial. One potential reason is that the low rainfall
during this study may not have been adequate to hydrate the gel, and hence store water
for the roots to access.
Nutrient loading was not beneficial to growth, condition or survival in this study.
However, nutrient loading has been shown to increase the biomass of Picea mariana
seedlings after planting in intact forest substrates in pots in a glasshouse (Timmer 1997).
Prior to planting, the nutrient loaded seedlings were similar in size, but had higher tissue
nutrient content compared with conventionally fertilised seedlings. In the present
experiment, the nutrient loading treatment ceased at least six weeks prior to planting.
Perhaps the delay between nutrient loading and planting had an effect on the outcome of
the experiment. Equally, other factors (namely water) may have been limiting growth
more than the nutrient status of the plant.
It was expected that pruning seedlings at planting would be a favourable treatment,
as studies have indicated that a low shoot:root ratio should be beneficial for growth
(Close et al. 2005b). However, pruning actually decreased survival and condition of
Atriplex bunburyana in this study. This finding contrasts with a study on Liquidambar
styraciflua in the southeastern United States that observed greater height growth and no

153
difference in survival between seedlings pruned to 50% of their original height and
unpruned seedlings. But that study did observe some detrimental effects of pruning,
with pruning resulting in a decrease in diameter growth. The reasons for the deleterious
effects of pruning in this study are unknown. Perhaps in this study the pruning
treatment was too severe, and removed too much leaf material and lowered the plants‘
ability to photosynthesise.
Most of the seedling mortality in the 2006 experiment occurred within the first 4 months
after planting. Seedling mortality shortly after planting is termed seedling transplant
shock (Close et al. 2005b). Transplant shock can be caused by stress while
acclimatising to environmental differences, such as light, between the nursery and the
field. However, in this study, seedlings were acclimatised at Shark Bay Salt prior to
planting. Another cause of transplant shock is competition with the surrounding
vegetation (Close et al. 2005b), however, in this case, seedlings were planted into bare
ground, and there was very little weed growth or emergence through natural migration
during the study. Drought stress is a widespread reason for transplant shock (Close et
al. 2005b), and probably the most likely explanation for seedling mortality in this study.
Only 58 mm of rain fell in 2006 (chapter 6), 12 mm of which fell between planting in
winter and monitoring in spring 2006. This amount of winter/spring rain is
substantially lower than the 25 year average rainfall for August, September and October
(40 mm) (SBS, unpublished data).
Species differences
Differences between the species were highlighted by higher survival of Atriplex
bunburyana (62% planted in the ripped area with no treatment) compared with Acacia
tetragonophylla (8%) after two years. Higher success of Atriplex bunburyana in post-
mine rehabilitation compared with other species has been noted in another Western
Australian study (Jefferson 2004). Another Australian study has noted survival
differences between Atriplex and Acacia species. Two years after planting, survival of
Acacia hemiteles was <50% compared with Atriplex semibaccata, which was between
approximately 60-80% (Yates et al. 2000). In this study, differences in survival could
be caused by differences in drought tolerance or tolerance to soil characteristics in the
post-disturbed areas. Atriplex species (saltbush) are considered halophytes (salt tolerant
plants) and hence have adaptations that may enable them to survive better in saline
environments. Saltbush species are able to withstand salinity by excreting salt from the

154
plant via bladder like cells on the leaf epidermis (Lambers et al. 1998). In addition, a
factor limiting the growth of Acacia tetragonophylla could be the possible lack of
rhizobia in the borrow pits, and the presence of rhizobia could be assessed by uprooting
plants and inspecting the roots. Rhizobia are bacteria that form a symbiotic relationship
with some species of plants (including legumes) to enable nitrogen fixation (Lambers et
al. 1998). The lack of rhizobia in soil can limit the growth of legumes, hence
restoration areas that have been subject to soil removal may benefit from rhizobia
inoculation (Rodríguez-Echeverría and Pérez-Fernández 2005). The findings in this
study highlight differences in soil properties (such as electrical conductivity) between
the pits, and that soil properties may affect species differently. Therefore, it is possible
that not all species will be able to be replaced by greenstock in all borrow pits. Species
survival in different soil types will need to be investigated before broad-scale planting
and seeding occurs.
7.5.2 Differences between greenstock planted in 2005 and 2006
Survival differences between the two years may be due to seasonal characteristics and
greenstock condition. There was more winter rainfall in 2005 compared with 2006,
which may have enabled high initial survival in 2005. However, greenstock planted in
2006 were in better condition compared with those planted in 2005. Seeds for
greenstock production in 2006 were sown earlier, and raised in a warmer climate
(Geraldton, compared with Perth), which may have enabled better survival of Atriplex
bunburyana over the first summer.
Ripping and fertiliser had little effect on greenstock planted in 2005, whereas ripping
had a major positive effect on survival of greenstock planted in 2006. This may also be
due to the differences in greenstock condition between the two years. Greenstock
planted in 2005 was small, and had a shallow root system, so may not have been
adversely affected by the underlying compaction. In addition, as rainfall was higher in
2005, roots may have been less likely to extend through the compact soil to access soil
moisture, compared with in 2006, where rainfall was low and roots may have had a
greater need to grow through the compacted soil to access moisture. It is possible that,
given the higher rainfall in 2005 and limited response to treatments, greenstock
treatments may be less beneficial in good years, compared with 2006 (and 2007) where
rainfall was low and some treatment effects were visible.

155
7.5.3 Conclusion
The findings of this study show that there are clear differences between soil properties
in disturbed and undisturbed areas in the Shark Bay World Heritage Area. One of these
properties was soil impedance, which was partly alleviated by soil ripping. In addition,
this study demonstrated the difficulty of returning vegetation to post-disturbed sites with
greenstock. While planting Atriplex bunburyana greenstock is a viable means of re-
vegetating post-disturbed areas at SBS, planting and sustaining the other species tested
may prove problematic. Ripping was the treatment with the largest effect on condition
and survival, and it seems to be a crucial practice in these post-disturbed areas. Apart
from a slight increase in height from fertiliser application, most of the greenstock
treatments had little effect on greenstock success, while pruning was detrimental.
However, given that the major part of the study was only conducted over two years,
both of which had very low rainfall, greater success could possibly be achieved in
higher rainfall years.

156

157
C H A P T E R 8
General Discussion
8.1 Introduction
Ecological restoration in southwest Western Australia is fraught with difficulties as a
result of the old landscape, highly evolved flora, high level of speciation and endemism,
short seed dispersal distances and limited vegetation succession (Cramer et al. 2007;
Hopper 2007). Many of these issues have most likely contributed to a high level of
novel dormancy mechanisms in seed of many Western Australian species to ensure
species regeneration, persistence and survival. Not surprisingly, the practice of
ecological restoration in Western Australia therefore requires a significant level of
empirical research.
Many ecological restoration studies undertaken in Western Australia have focused on
the southwest of the state, at post-mined sites operated by major resource industries
including Alcoa World Alumina (Koch et al. 1996; Ward et al. 1996; Roche et al.
1997b; Koch 2007; Koch and Hobbs 2007) and Rocla Quarry Products (Rokich et al.
2000; Rokich et al. 2001; Rokich et al. 2002; Turner et al. 2006c), and in remnant
vegetation (Yates and Hobbs 1997; Yates et al. 2000) or abandoned fields (Cramer et al.
2007; Standish et al. 2007; Cramer et al. 2008) within the agricultural landscape.
However, all these sites support vegetation associations within a more reliable climatic
zone compared with the site studied in this thesis, SBS, which is in a transition zone
between the South West and Eremaean Botanical Provinces. Whilst restoration
techniques (including seed pre-treatments) developed in southwest Western Australia
and other parts of the world may be applicable to the arid zone, it was necessary to trial
the techniques to determine their efficacy.

158
This thesis presents a body of information relating to the floristic associations of the
study site; seed dormancy and accompanying seed pre-treatments, the topsoil seed bank;
seed broadcasting; and greenstock planting – all key aspects for technical development
of restoration (‗technical reclamation‘ of Prach and Hobbs (2008)). The main findings
suggest that:
There is a complex spatial arrangement of floristic associations within the SBS
lease, with little evidence of natural colonisation or succession and where natural
colonisation has occurred there is a significant disparity in floristic composition
between reference sites and borrow pits.
The 18 dominant species, which were the focus of this study, exhibit a variety of
seed dormancy types including no dormancy (44% of species), physical dormancy
(11%) and physiological dormancy (44%).
The dominant species from SBS and Solanum spp. from across arid Australia
respond differently to temperature and stimulant cues, implying a variety of
strategies for germination timing and illustrating the difficulty of generalising across
species in a genus.
Dormancy of three of the physiologically dormant species can be alleviated by after-
ripening, and temperature and seed moisture content affect the efficacy of after-
ripening.
Whilst seed broadcasting has limited success, particularly in low rainfall years,
seedling emergence has the potential to be enhanced by soil ripping (to alleviate
high soil impedance in borrow pits) and raking (to ensure incorporation of seed into
the soil).
Greenstock planting has greater success than seed broadcasting, and can be
enhanced with soil ripping and fertiliser, while plant optimisation treatments (air-
pruned pots, wetta gel and nutrient loading) have no effects on plant survival.
This synthesis will provide a summary of the floristic differences between the disturbed
and undisturbed areas at SBS, an overview of seed germination requirements and
dormancy alleviation, a discussion into why restoration success may be improved by a
better understanding of seedling recruitment processes, and an outline of some of the
findings from restoration practices.

159
8.2 Floristics
This study identified a complex spatial arrangement of floristic associations and types
within the SBS lease, as evidenced by the plant diversity between the three undisturbed
natural vegetation sites adjacent to the borrow pits that were the focus of this study, and
between the natural vegetation sites and their correspondingly adjacent borrow pits
(chapter 5). The dominant species in the natural vegetation (as represented by cover
and abundance) included Acacia ligulata, Atriplex bunburyana, Melaleuca
cardiophylla, Thryptomene baeckeacea, and Triodia plurinervata. Three of these
species were not present in the borrow pits subject to natural migration nor were they
evident in the pit subjected to topsoil replacement, highlighting the need for human
intervention. Limitations for natural migration may include the lack of dispersal
mechanism (the case for Melaleuca cardiophylla and Thryptomene baeckeacea) and a
reliance on vegetative propagation (Triodia plurinervata). Absence from topsoil is not
surprising for Melaleuca cardiophylla, which supports canopy-stored seed rather than
soil-stored seed and for Thryptomene baeckeacea which has complex seed dormancy
(chapter 3). Other Western Australian studies have also found limited recruitment from
natural migration (Standish et al. 2007), and a need to supplement the topsoil seed bank
by seed broadcasting to achieve required species richness (Koch 2007), reinforcing the
need for active management practices at this site.
8.3 Investigation into seed germination requirements and dormancy
alleviation
From the research contained in this thesis, we have classified dormancy of dominant
species, determined the effect of incubation temperature and germination stimulants,
and overcome dormancy of three species using after-ripening. This information will be
invaluable to restoration practitioners at SBS in order to understand germination timing,
and hence timing of seed broadcasting practices. In addition, this information will
enable practitioners to treat seeds appropriately prior to seed broadcasting or prior to
sowing for seedling production. Germination of several species has not been optimised
and is therefore in need of further research.

160
8.3.1 Dormancy types in this ecosystem
Seeds of the 18 focus species from SBS were found to be either, non-dormant (44% of
species), physically dormant (11%) or physiologically dormant (44%) (chapters 2, 3 and
4). Based on these findings, a classification model illustrating dormancy alleviation and
germination stimulation requirements has been developed for 38 of the species that were
identified in the floristic assessment (chapter 5) including the 18 focus species. The 20
species not subject to dormancy research in this study (chapter 5) have been placed in
the model based on evidence from the literature (Baker et al. 2005a; Turner et al.
2005a; Turner et al. 2006a; Turner et al. 2006b), or unpublished results (Merritt and
Turner unpublished) that detail dormancy classification and/or germination stimulation.
From consolidation of all this available information for the 38 species encountered at
SBS, it can be inferred that 26% of species are non-dormant (ND), 24% exhibit physical
dormancy (PY) and 53% exhibit physiological dormancy (PD), providing a solid basis
for future germination research. Such proportions are similar to those that have been
reported across hot deserts (Baskin and Baskin 1998), with approximately 10% ND,
30% PY and 60% PD. Clearly, physiological dormancy is the most important
dormancy type in the SBS area and other arid ecosystems. However, dormancy type
may not be proportionally distributed within each plant life form. For instance, seeds of
desert trees commonly exhibit PY (Baskin and Baskin 1998) which seemed to be true at
SBS also, with the common trees (Acacia spp. and Alyogyne sp. – Fig. 8.1) exhibiting
PY.
8.3.2 Species for which germination mechanisms were resolved
Effect of temperature and germination stimulants
Seeds of most species that we concluded were non-dormant had higher germination
when incubated at 26/13oC compared with 33/18
oC, whereas seeds we determined to
have physical and physiological dormancy had similar germination at both temperatures
(although, for the physiologically dormant seeds, the temperatures over which seeds
will germinate will broaden as dormancy is alleviated) (chapter 2). This illustrates that
non-dormant seeds may be cueing their germination to times of the year that are cooler,
more likely to receive rainfall, and therefore more favourable for seedling
establishment. This temperature preference was not observed for two species with non-
dormant seeds, Melaleuca cardiophylla and Salsola tragus. Melaleuca cardiophylla
had only slightly higher germination at 26/13oC compared with 33/18
oC, and incubation
temperature had no effect on the germination of S. tragus. These two species store part

161
or all of their seeds in the canopy, the release of which controls the timing of
germination. Hence, dormancy, temperature optimum and canopy storage of seed all
appear to be controlling the timing of germination, perhaps to avoid germination over
summer when conditions may not be appropriate for establishment. This implies that an
appropriate time for seed broadcasting would be just prior to the winter rain and this
seems to be an appropriate strategy for most species. For those species that are able to
germinate at higher temperatures timing is less critical for broadcasting. For instance,
Solanum orbiculatum exhibited higher germination when incubated at 33/18oC
compared with 26/13oC, as did two additional Solanum sp. from other arid Australian
environments and an additional four (three of which were PD) was unaffected by
incubation temperature (chapter 4). While some of these species occur in ecosystems
that receive predominantly summer rainfall, and are hence likely to germinate at that
time, Solanum orbiculatum occurs at SBS, so may have the potential to germinate
during sporadic summer rainfall.
In a large number of species germination stimulants enabled increased germination.
Gibberellic acid (GA3) increased germination of ten species at one or both incubation
temperatures; Anthocercis littorea, Aphanopetalum clematideum, Austrostipa
elegantissima, Diplolaena grandiflora, Pembertonia latisquamea, Ptilotus exaltatus,
Rhagodia baccata, Solanum orbiculatum, and Thryptomene baeckeacea (chapter 2) and
all eight of the Solanum species tested (chapter 4). Ten species (Anthocercis littorea,
Aphanopetalum clematideum, Diplolaena grandiflora, Pembertonia latisquamea,
Ptilotus exaltatus, Solanum centrale, S. cunninghamii, S dioicum S. orbiculatum and S.
phlomoides) had higher germination when treated with KAR1 and/or SW at one or more
of the incubation temperatures. Clearly both agents act over a wide range of families
and on PD seeds, and in some cases would be essential pre-treatments for seeds destined
for seed broadcasting and greenstock production as without the stimulants, germination
is limited.
After-ripening
After-ripening was successful in overcoming PD of three species (Anthocercis littorea,
Dioscorea hastifolia, Zygophyllum fruticulosum; chapter 3). Understanding the
mechanism of after-ripening may assist in further development of optimal storage
conditions for other untested PD species at SBS. Optimising after-ripening is necessary

162
to maximise germination at the time when seeds are required for rehabilitation
activities.
Both temperature and RH affected after-ripening in this study. After-ripening has been
shown previously to occur within an optimum range of RH. It is clear from this study
and others that if the RH is too low, seeds will not after-ripen at all, or will do so only
slowly, whereas if it is too high, seed viability will decline. The ability of water to
influence metabolic/biological processes in seeds is closely linked to RH with at least
three regions of water activity clearly defined (Walters 1998). Other authors have
reported that this optimum range of RH appears to correlate with region two of water
binding for Lolium rigidum and Xanthium pennsylvanicum (Esashi et al. 1993;
Steadman et al. 2003). Seeds of another species, Avena fatua, also after-ripened most
efficiently at seed moisture contents within region two of water binding, but dormancy
loss was not exclusively constrained to this region (Foley 1994) and optimal after-
ripening of Oryza sativa occurred near the boundary between regions one and two of
water binding (Leopold et al. 1988). In our study, optimal after-ripening at 45oC
generally occurred when seeds were equilibrated at 13% RH (near the boundary
between regions one and two) and at 30oC, after-ripening occurred in seeds equilibrated
at 13% RH and 50% RH (region two). At different regions of water binding, water has
different properties. In region two, water is weakly associated with membranes and
begins to behave like a solvent (Leopold et al. 1988; Steadman et al. 2003). The types
of reactions in region two may be non-enzymatic and oxidative reactions known to
proceed within this range of seed moisture contents, in particular Amadori and Maillard
reactions, have been proposed to play a role in dormancy alleviation (Esashi et al. 1993;
Foley 1994). Regardless of the mechanism it is clear that manipulating the storage
environment of PD seeds can accelerate dormancy loss and provide useful options for
handling seeds between collection and restoration use.
8.3.3 Species with unknown germination mechanisms
For some species the approach/treatment applied did not optimise germination.
Germination of several species with either PY or PD, was not fully resolved and
therefore further research is needed to develop pre-treatments for seed destined for
rehabilitation activities. Physical dormancy is exhibited in 16 families (Baskin et al.
2000; Baskin et al. 2006b), and in this study it was identified in seeds of two families
(Fabaceae and Surianaceae). While PY was recorded for Acacia tetragonophylla

163
(>80% germination) and Stylobasium spathulatum (<40% germination) (chapter 2), the
hot water treatment used to overcome physical dormancy was not completely effective
for S. spathulatum, with <50% of seeds imbibing after exposure to hot (>90oC) water
(unpublished results) for various time periods. This experiment concurs with the
results of Baskin et al. (2006b), where hot water treatment overcame physical dormancy
of only 70-87% of Stylobasium seeds and germination percentages of seeds treated with
hot water were <50%. Other mechanisms for overcoming PY include mechanical
scarification, acid scarification, and exposure to high temperatures (Quinlivan 1961;
Quinlivan 1966; Baskin et al. 1998). Treatments for overcoming PY have been mainly
been developed on seeds in the majority of families with PY (13 out of 16) where the
water impermeable palisade layer (that causes PY) is located in the seed coat. However,
Stylobasium spathulatum (Surianaceae) is in one of only three families in which the
water impermeable layer is located in the fruit coat, compounding the difficulties in
developing a treatment to overcome dormancy. Further research is required to
successfully overcome physical dormancy of entire seed lots, and to determine whether
low germination is due to a combination of physical and physiological dormancy, or
other factors.
Three out of eight Solanum species (S. chippendalei, S. diversiflorum and S. sturtianum)
germinated to 17-64% when treated with GA3, and had minimal (≤2%) germination
with the other treatments. It is clear that these species have physiological dormancy, as
they imbibe water and have fully developed embryos. To overcome this dormancy,
after-ripening, in combination with a smoke cue could be investigated in a manner
similar to chapter 3. However, one key finding is that the differences in germination for
the eight Solanum species highlight the need for testing germination of individual
species, and not making generalisations within a genus. This finding will be important
when undertaking further research on untested species at SBS and may be important
when working with other arid zone ecosystems.
Whilst after-ripening promoted germination of three species with PD, for the other three
investigated in detail it was not effective. One of these three unresponsive species,
Acanthocarpus preissii, has been shown previously to respond to warm stratification
(Turner et al. 2006b). Stratification may occur during summer or autumn, when rainfall
moistens the soil but temperatures may be too high for germination. Stratification (6 or
12 weeks at 33/18oC) was also trialled on the other two PD species that did not respond

164
to after-ripening, Eremophila oldfieldii and Thryptomene baeckeacea, but germination
was low and the results were inconclusive due to several months of shelf storage prior
to the experiment and extremely low seed fill, compounding difficulties in the
experiment (data not presented). It is possible that the after-ripening or stratification
conditions were not optimal for these species. However, given that these stratification
conditions alleviate dormancy of Acanthocarpus preissii (Turner et al. 2006b), and 4-6
weeks stratification overcomes dormancy in other species with a stratification
requirement (Merritt et al. 2007), it is likely that dormancy of Eremophila oldfieldii and
Thryptomene baeckeacea is deep and complex. Species with such complex dormancy
may require several seasons of soil burial, and may need a combination of both
stratification and after-ripening to alleviate dormancy, followed by a stimulant to
promote germination (Merritt et al. 2007), therefore, long-term seed-burial or laboratory
experiments that closely mimic natural conditions may elucidate the process of
dormancy loss in these species. Interestingly, seeds of both species are contained within
a woody, indehiscent fruit (drupe for Eremophila oldfieldii and nut for Thryptomene
baeckeacea). Seeds of other genera contained within a woody fruit have also been
found to be difficult to germinate, such as Leucopogon, Persoonia and Styphelia
(Norman and Koch 2008). When excised from the fruit, seeds of both Eremophila
oldfieldii and Thryptomene baeckeacea germinate (data not presented), however, this is
not a practical treatment for large-scale restoration. As a result, these two genera and
the other eight that are listed in the model as having unknown mechanisms for
germination (Fig. 8.1) are in need of further investigation.

165
Fig
ure
8.1
. M
odel
of
do
rman
cy c
lass
ific
atio
n a
nd a
llev
iati
on,
and g
erm
inat
ion s
tim
ula
nt
requir
emen
ts f
or
38 d
om
inan
t ta
xa
at S
BS
iden
tifi
ed i
n
chap
ter
2.
The
model
is
bas
ed o
n t
axa
studie
d i
n c
hap
ters
2,
3 a
nd 4
(li
sted
in b
old
), a
nd t
axa
list
ed i
n t
he
veg
etati
on s
urv
ey i
n c
hap
ter
5 b
ut
not
studie
d (
unbold
ed).
T
he
num
ber
of
spec
ies
per
gen
us
is l
iste
d i
n b
rack
ets,
if
gre
ater
than
one.
D
orm
ancy
of
taxa
not
studie
d w
as a
scer
tain
ed f
rom
the
lite
ratu
re (
Bak
er e
t al.
2005a;
Turn
er e
t al.
20
05a;
Turn
er e
t al.
2006a;
Turn
er e
t al.
2006b).
S
pec
ies
wer
e fo
und t
o b
e non
-dorm
ant
or
do
rman
t
(physi
olo
gic
al (
PD
) o
r p
hysi
cal
(PY
)).
1T
he
nu
mber
and
per
centa
ge
of
spec
ies
wit
h e
ach
dorm
ancy
type
is l
iste
d i
n b
rack
ets
afte
r ea
ch d
orm
ancy
type.
2W
ithin
the
taxa
that
exhib
it P
Y,
those
whic
h m
ay e
xhib
it c
om
bin
atio
nal
dorm
ancy
(P
Y +
PD
) is
indic
ated
by *
. 3
Aft
er-r
ipen
ing i
ndic
ates
a
per
iod o
f w
arm
, dry
sto
rage.
4W
arm
str
atif
icat
ion i
ndic
ates
a p
erio
d o
f w
arm
, m
ois
t st
ora
ge.
5U
nknow
n m
echan
ism
indic
ates
that
ger
min
atio
n
requir
emen
ts a
re u
nknow
n.
See
ds
may
nee
d a
fter
-rip
enin
g,
war
m s
trat
ific
atio
n,
or
a co
mbin
atio
n o
f th
e tw
o.
6O
ver
com
ing t
he
wat
er i
mper
mea
ble
seed
or
fruit
co
at
is
esse
nti
al
to
over
com
e dorm
ancy
. T
reat
men
ts
may
in
clude
hot
wat
er,
man
ual
/aci
d
scar
ific
atio
n
or
hig
h
tem
per
ature
s.
7S
tim
ula
nts
in
clude
smoke
wat
er a
nd k
arri
kin
oli
de
and t
hey
may
be
repla
ced b
y g
ibber
elli
c ac
id.
8G
eosp
oro
us
refe
rs t
o s
eeds
that
are
rel
ease
d i
nto
and s
tore
d i
n t
he
soil
see
d b
ank a
nd s
eroti
nous
refe
rs t
o s
eeds
that
are
sto
red o
n t
he
par
ent
pla
nt.
Seed
s
(38
sp
ec
ies)
Do
rman
t
(28
sp
ec
ies)
(74
%)1
No
n-d
orm
an
t
(10
sp
ec
ies)
(26
%)1
CH
EN
OP
OD
IAC
EA
E
Sals
ola
MY
RT
AC
EA
E
Mela
leu
ca
AS
TE
RA
CE
AE
Pem
bert
on
ia
CH
EN
OP
OD
IAC
EA
E
Atr
iple
x(2
)
Rh
ag
od
ia(2
)
Sals
ola
CU
NO
NIA
CE
AE
Ap
han
op
eta
lum
PO
AC
EA
E
Au
str
os
tip
a
Ph
ysio
logic
al d
orm
ancy
(19
sp
ecie
s)
(53%
)1P
hys
ica
l do
rmancy
2
(9 s
pe
cie
s)
(24
%)1
Ove
rcom
e w
ate
r
imperm
ea
ble
seed
/fru
it c
oa
t6R
esponds t
o
aft
er-
rip
enin
g3
Re
sponds t
o
wa
rm s
tra
tifica
tion
4
DA
SY
PO
GO
NA
CE
AE
Acanth
oca
rpu
s
DIO
SC
OR
EA
CE
AE
Dio
sc
ore
a
ZY
GO
PH
YL
LA
CE
AE
Zyg
op
hyll
um
PO
AC
EA
E
Trio
dia
(2)
Re
quires s
tim
ula
nt7
SO
LA
NA
CE
AE
An
tho
cerc
is
Re
quires s
tim
ula
nt7
SO
LA
NA
CE
AE
So
lan
um
Sero
tinous
8
Un
know
n
mechanis
m5
AM
AR
AN
TH
AC
EA
E
Pti
lotu
s
EU
PH
OR
BIA
CE
AE
Cro
ton
GO
OD
EN
IAC
EA
E
Scaevola
(3)
MY
OP
OR
AC
EA
E
Ere
mo
ph
ila
(2)
MY
RT
AC
EA
E
Th
ryp
tom
en
e
PH
OR
MIA
CE
AE
Dia
nella
RE
ST
ION
AC
EA
E
De
sm
ocla
dus
RU
TA
CE
AE
Dip
lola
en
a
TH
YM
ELA
CE
AE
Pim
ele
a
ZY
GO
PH
YL
LA
CE
AE
Nit
rari
a
Geo
spo
rous
8
Geo
spo
rous
8
CA
ES
ALP
INIA
CE
AE
Senna
MA
LV
AC
EA
E
Aly
ogyne*
MIM
OS
AC
EA
E
Ac
acia
(2)
PA
PIL
ION
AC
EA
E
Gastr
olo
biu
m
RH
AM
NA
CE
AE
Spyrid
ium
SA
PIN
DA
CE
AE
Dip
lopeltis
* (2
)
SU
RIA
NA
CE
AE
Sty
lob
asiu
m
Seed
s
(38
sp
ec
ies)
Do
rman
t
(28
sp
ec
ies)
(74
%)1
No
n-d
orm
an
t
(10
sp
ec
ies)
(26
%)1
CH
EN
OP
OD
IAC
EA
E
Sals
ola
MY
RT
AC
EA
E
Mela
leu
ca
AS
TE
RA
CE
AE
Pem
bert
on
ia
CH
EN
OP
OD
IAC
EA
E
Atr
iple
x(2
)
Rh
ag
od
ia(2
)
Sals
ola
CU
NO
NIA
CE
AE
Ap
han
op
eta
lum
PO
AC
EA
E
Au
str
os
tip
a
Ph
ysio
logic
al d
orm
ancy
(19
sp
ecie
s)
(53%
)1P
hys
ica
l do
rmancy
2
(9 s
pe
cie
s)
(24
%)1
Ove
rcom
e w
ate
r
imperm
ea
ble
seed
/fru
it c
oa
t6R
esponds t
o
aft
er-
rip
enin
g3
Re
sponds t
o
wa
rm s
tra
tifica
tion
4
DA
SY
PO
GO
NA
CE
AE
Acanth
oca
rpu
s
DIO
SC
OR
EA
CE
AE
Dio
sc
ore
a
ZY
GO
PH
YL
LA
CE
AE
Zyg
op
hyll
um
PO
AC
EA
E
Trio
dia
(2)
Re
quires s
tim
ula
nt7
SO
LA
NA
CE
AE
An
tho
cerc
is
Re
quires s
tim
ula
nt7
SO
LA
NA
CE
AE
So
lan
um
Sero
tinous
8
Un
know
n
mechanis
m5
AM
AR
AN
TH
AC
EA
E
Pti
lotu
s
EU
PH
OR
BIA
CE
AE
Cro
ton
GO
OD
EN
IAC
EA
E
Scaevola
(3)
MY
OP
OR
AC
EA
E
Ere
mo
ph
ila
(2)
MY
RT
AC
EA
E
Th
ryp
tom
en
e
PH
OR
MIA
CE
AE
Dia
nella
RE
ST
ION
AC
EA
E
De
sm
ocla
dus
RU
TA
CE
AE
Dip
lola
en
a
TH
YM
ELA
CE
AE
Pim
ele
a
ZY
GO
PH
YL
LA
CE
AE
Nit
rari
a
Geo
spo
rous
8
Geo
spo
rous
8
CA
ES
ALP
INIA
CE
AE
Senna
MA
LV
AC
EA
E
Aly
ogyne*
MIM
OS
AC
EA
E
Ac
acia
(2)
PA
PIL
ION
AC
EA
E
Gastr
olo
biu
m
RH
AM
NA
CE
AE
Spyrid
ium
SA
PIN
DA
CE
AE
Dip
lopeltis
* (2
)
SU
RIA
NA
CE
AE
Sty
lob
asiu
m

166
8.4 Restoration ecology of disturbed land at Shark Bay Salt
8.4.1 Understanding seedling recruitment processes may improve restoration
success
While moderate establishment success was achieved by planting Atriplex
tetragonophylla seedlings, this study found limited success from seed broadcasting and
planting seedlings of other species. This limited success may have been due to a lack of
understanding of seedling recruitment dynamics in the Australian arid zone. Greater
understanding of the patterns and processes driving spatial organisation of vegetation in
arid areas may assist restoration efforts (Ludwig and Tongway 1995) particularly with
respect to seedling recruitment and plant survival. Understanding the relationship
between the soil seed bank environment and seed germination may improve the success
of direct seeding in a rehabilitation context (Allen and Meyer 1998), particularly
regarding succession, temporal variation (driven by rainfall) and spatial variation
(driven by existing vegetation patterning). For instance, understanding vegetation
succession may influence the timing of seed broadcasting. In addition, if seeds require
average or above average rainfall for recruitment, which seems likely from results in
this study, then low seedling recruitment (from both topsoil and broadcast seeds) in
years with below average rainfall will undoubtedly occur. Specific micro-sites (such as
‗nurse plants‘ or ‗resource islands) may be necessary for seedling recruitment and plant
survival, and if this is the case, restoration techniques will need to provide for this. For
instance, the creation of resource islands with greenstock can be beneficial for
revegetating borrow pits in arid America (Bainbridge 2007). Nurse plants and resource
islands are areas that require investigation in an Australian context.
Succession in the arid-zone
An understanding of the type of vegetation succession that occurs in the ecosystem will
have an effect on management practices. There are two models for recruitment and
succession after disturbance. These models are the initial floristic model and the relay
floristic model (Egler 1954). In the initial floristic model, seeds of all species in the
seed bank recruit immediately following disturbance, and over time some species
gradually disappear, whereas in the relay floristic model a dominant group of species
will be replaced by a new group of species, and this process is repeated over time (Egler
1954). If an ecosystem follows the initial floristic model, when implementing
rehabilitation, it would be prudent to broadcast seeds of all species in the first year. For

167
instance, the jarrah forest in Western Australia is believed to follow the initial floristic
model (Koch and Ward 1994), so restoration practitioners in that ecosystem attempt to
establish the complete suite of species immediately following disturbance (Norman et
al. 2006a). In an area of jarrah forest rehabilitation, species richness remained constant
from two years after rehabilitation to the end of the study (14 years) matching the model
proposed in Cramer et al. (2007) for southwest Western Australia. Other ecosystems
also appear to follow the initial floristic model, for instance in the South African fynbos,
where maximum species richness is observed after disturbance due to fire, and species
gradually disappear (Holmes and Richardson 1999). However, if an ecosystem follows
the relay floristic model, it would be wise to sow seeds of pioneer species first, followed
by those that are secondary successional species or species that require facilitation. It is
unknown whether the ecosystem in this study follows the initial or relay floristic model,
and research to determine this would benefit rehabilitation planning.
The effect of rainfall on seedling recruitment
This study suggests that rainfall is the major limiting factor for seedling recruitment.
Within arid environments that experience uncertain climatic conditions, an
understanding of seedling recruitment patterns and response to episodic rainfall events
is an important restoration consideration. It is assumed that there is little recruitment in
years that have rainfall substantially below the average, as demonstrated by this study,
and that mass seedling recruitment occurs in years with at least average, but most likely
well above average rainfall. However, given the relatively short time-frame of this
study, the effect of rainfall was not able to be fully investigated. One arid-zone study in
Peru and Chile has indicated that two tree species need a minimum threshold of
approximately 85 mm of rainfall for establishment, which is higher than the average
rainfall of 50 mm recorded for the study site (López et al. 2008). This study contrasts
with another study in a semi-arid rangeland in Argentina that found low precipitation
did not affect the water availability in the top layer of the soil, and did not affect
emergence, survival or biomass. Low precipitation did, however, affect the depth of
water infiltration (Cipriotti et al. 2008). Clearly, there are different effects of rainfall on
recruitment in different ecosystems, therefore quantifying the effects of variable annual
rainfall on recruitment is needed in the arid-zone of Western Australia. Understanding
seasonal recruitment would assist in determining appropriate management practices.
For instance, if broadcasting occurs in a low rainfall year, restoration practitioners need
to know if seedlings will emerge and die, or if seeds will not germinate and either

168
persist in the soil until the following year, lose viability or be subject to removal via
predation or erosion. Management practices would need to take these outcomes into
account, and if seedlings die, or seeds lose viability or are removed, broadcasting will
need to occur again in the following year. However, if seeds do not germinate, and
persist in the soil until a season when they are able to emerge and survive, follow-up
broadcasting may not be needed.
Implications of vegetation patterning on seedling recruitment
Whilst seedling recruitment can have temporal variation due to differences in annual
rainfall, it may also have spatial variation. Semi-arid landscapes often consist of
patches (trees, shrubs and clumps of small shrubs) (Ludwig and Tongway 1995). This
suggests that seedlings recruit in these patches (under ‗nurse plants‘ or within ‗resource
islands‘) rather than uniformly across the landscape. From observations and a study on
a nearby ecosystem (the Peron Peninsula in Shark Bay), Brooker (2000) reported that
the landscape in that part of the Western Australian arid zone is indeed dominated by
patches, particularly clumps of shrubs. These shrub clumps may act as ‗fertile islands‘
or ‗resource islands‘ (Figs 8.2-8.5), thereby supporting the patch theory. Patches may
be dominant in the landscape by trapping seed and/or increasing recruitment success. In
the presence of vegetation, wind blown seeds and organic matter have the potential to
become trapped by ground covers, shrubs and litter sitting on the soil surface under
vegetation. Other studies have shown that soil seed banks in some arid areas are
heterogeneous (Page et al. 2006), sometimes with greater numbers of seeds under
shrubs compared with between shrubs (Guo et al. 1998). Low emergence following
broadcast seeding in this study was possibly a result of seed loss through wind erosion
at SBS, made more pronounced in the absence of extant vegetation in the borrow pits
and the prevalence of strong winds at the site that could move seed from bare surfaces.
It is also possible that species within the arid region have evolved to recruit in the more
favourable conditions within patches. Resources in arid environments can be limited,
and these resources (such as nutrients) may be lost from bare areas and concentrated in
vegetation patches (Ludwig and Tongway 1995). As a result, understanding and
restoring vegetation patches in rehabilitation sites may play a crucial role in
revegetation success (Ludwig et al. 1994; Ludwig and Tongway 1995).

169
Figure 8.2. Natural gaps and species clumps (May 2006)
Figure 8.3. Aerial photo of shrub clumps and inter-patch gaps (Peron Peninsula near
Denham) (September 2004)

170
Figure 8.4 A shrub clump surrounded by annual flora (September 2005)
Figure 8.5. Shrub clump consisting of Acacia tetragonophylla, Pembertonia
latisquamea, Zygophyllum fruticulosum and Solanum orbiculatum (September 2005)

171
8.4.2 Restoration practices
Topsoil
The study identified that borrow pit restoration requires careful management to ensure
reinstatement of matched floristic types. The importance of this restoration step was
illustrated by the borrow pit that had been subjected to topsoil replacement. In spite of
the high plant density, and a limited number of dominant species, a different floristic
type to the nearby undisturbed vegetation was present (chapter 5). Whilst it is possible
that this topsoil may have been sourced from a dissimilar floristic type, this seems
unlikely, as SBS records indicate it was sourced from within the same vegetation
association as indicated by Mattiske (1996). Alternatively, the composition of species
that regenerated from topsoil (e.g. Acacia ligulata, Alyogyne cuneiformis, Atriplex
bunburyana and Stylobasium spathulatum) may be a reflection of the species that are
typically responsive to topsoil handling procedures in this environment. Species
dominance of this type is often attributed to seed dormancy status. Of the species
typically found to be regenerating from the topsoil, and therefore dominant components
of the topsoil (Acacia ligulata, Alyogyne cuneiformis, Atriplex bunburyana and
Stylobasium spathulatum), three exhibit physical dormancy (PY) (Fig. 8.1). This
indicates that either seeds of PY species are better able to survive the topsoil storage
process (otherwise known as topsoil stockpiling), compared with non-PY species, or
they benefit from soil disturbance. Koch et al. (1996) found that seeds of PY species
survive topsoil handling process better than other species in their study of post-mined
restoration in southwest Western Australia. Similarly, Rokich et al. (2000) reported that
PY (or hard-seeded) species survived topsoil stockpiling better than non-PY species.
However, another explanation is that physically dormant species benefit from
disturbance, as it promotes seed germination. Rokich (1999) demonstrated the presence
of three PY species: Acacia pulchella, Hovea pungens and Jacksonia densiflora, in
topsoil replaced sites whereas prior to topsoil stripping, all three species were absent
from undisturbed Banksia woodland. In addition, another two PY species:
Gompholobium tomentosa and Gastrolobium capitatum, were recorded in extremely
low numbers in undisturbed Banksia woodland compared with topsoil replaced sites.
Given the limited topsoil availability for use in the remaining borrow pits, the technique
of ‗borrowing‘ topsoil was investigated and demonstrated that species can recruit and
establish to some extent (by year 2) in both topsoil donor and recipient sites (chapter 6).
This is a new technique that, prior to this study, had not been tested. However, an

172
important aspect of this activity is to ensure that the topsoil donor site has the potential
to return to pre-disturbed plant abundance, richness and cover levels, together with
minimal weed invasion. Therefore, seedling recruitment and plant survival in topsoil
donor sites needs to be assessed and compared to adjacent undisturbed sites (reference
sites) over the longer term, possibly after five and 10 years. Given the duration of this
study, there was not an option to assess the return of topsoil donor sites to pre-
disturbance conditions, and therefore the potential of borrowing topsoil. Until further
studies indicate any potential in borrowing topsoil, vegetation will need to be re-
established in borrow pits at SBS through the use of broadcast seeding and greenstock
planting. However, once this potential is understood, there is still the likelihood that not
all species will return via topsoil replacement or indeed from natural migration. All
missing species will need to be replaced via seed broadcasting and/or greenstock
planting once seed dormancy issues, if present, are overcome.
Seed broadcasting and greenstock
Once seed dormancy was resolved, and pre-treatments to stimulate germination were
developed, seeds could be broadcast directly into the borrow pits, or sown in pots to
produce greenstock. Seedling recruitment and survival from broadcasting was affected
by environmental conditions, particularly water. For instance, combined seedling
emergence of the three species common to both years was 11-fold higher in the wetter
year (2005) compared to the drier year (2006) (chapter 6). In addition, soil ripping and
raking further increased emergence in the wetter year, and this was perhaps due to the
creation of microsites which may have resulted in enhanced seed:soil contact and
therefore increased access to soil moisture. Soil raking has been shown to increase
seedling emergence in Banksia woodland restoration (Turner et al. 2006c), and soil
ripping is beneficial for seedling establishment in a Queensland rainforest (Doust et al.
2006).
Soil ripping vastly increased survival of Atriplex bunburyana seedlings, and to a lesser
extent Acacia tetragonophylla (chapter 7). This was likely due to the decrease in soil
impedance after ripping. Most other treatments had no effect on seedling growth and
survival, except fertiliser which increased the height of Atriplex bunburyana seedlings,
and pruning, which was detrimental to survival. Apart from the probability that water
was the most over-riding and critical resource in the environment, we are unsure as to
why these treatments were ineffective. In the future, further investigation could be

173
undertaken into the treatments that did not improve survival to determine if they in fact
did what we expected them to do. For instance, foliar nutrient content of nutrient
loaded seedlings could be tested, to determine whether the treatment did indeed enhance
the internal nutrient concentration of seedlings. Root morphology of seedlings grown in
normal and air-pruned pots could be investigated before and after planting to determine
if the air-pruned pots improved root morphology. Wetta gel could be excavated to
determine whether it hydrated and whether the roots were able to access it.
When considering the source of plants for restoration operations, one needs to consider
the benefits and costs of employing seed broadcasting and/or greenstock plants
(Appendix 14). Whilst seed broadcasting is generally considered to be a quicker and
more efficient means of returning plants to restoration sites (Knight et al. 1998), it is
also considered to be a non-reliable source of seedlings (Glenn et al. 2001; Banerjee et
al. 2006). On the other hand, employing greenstock is more costly, but generally
ensures improved seedling quality control and reliability (Barrett-Lennard et al. 1991),
and uses substantially less seed than broadcasting (Greening Australia 2003). Not
surprisingly, it has been reported that that direct seeding in arid environments is rarely
successful, whereas planting seedlings can be more effective (Bainbridge 2007).
Employing greenstock may also assist in more rapid soil stabilisation in areas prone to
erosion (such as pit slopes) where broadcast seeding may be futile due to seed losses as
a result of high wind and/or water erosion. In addition, producing greenstock from
cuttings is necessary when seed viability and germination is low (Gardner 2001).
However, greenstock planting is an expensive option and requires significantly greater
labour input. Therefore, weighing up the costs versus the benefits (e.g. improved
survival) is an issue that restoration ecologists must face, and nowhere is this issue more
apparent than in arid-landscapes where seed supplies are limited.
Determining which source of plants provided the greater benefit to SBS restoration sites
is difficult given the yearly variation in climate and the different species employed from
year to year. However, when comparing a year in which the same species were
employed in both broadcast seeding and greenstock planting operations there is the
possibility of drawing some conclusions. A cost-benefit analysis of seed broadcasting
and greenstock is provided in Appendix 14. What is clear from this analysis is that
future opportunities for effective restoration may rely on a combination of greenstock
planting and broadcast seeding – the former for key dominant species such as trees and

174
large shrubs that are in low enough density for planting to be effective. Broadcast
seeding would then involve species that are common in the landscape with abundance
levels that preclude use of greenstock.
8.5 Conclusion
This study has initiated restoration research for SBS, and provided a body of benchmark
information to initiate a rehabilitation plan (Appendix 15). Foremost, the study has
highlighted the challenges re-establishing viable plant communities in the arid zone,
where rainfall is extremely variable and is possibly the singly determinant critical for
determining restoration success. Given that the study has improved our understanding of
the seed biology in the Australian arid zone, it has provided a means of returning plants
to post-disturbed areas utilising seed broadcasting and greenstock planting. The study
has also provided a window of opportunity to further examine the potential of a novel
technique of borrowing topsoil, when topsoil is in scarce supply. Management
recommendations and directions for further research have been developed from the
findings in this study (Appendix 15).

175
References
Adams R (1999) Germination of Callitris seeds in relation to temperature, water stress,
priming, and hydration-dehydration cycles. Journal of Arid Environments 43,
437-448.
Ahmed AK, Johnson KA (2000) Horticultural development of Australian native edible
plants. Australian Journal of Botany 48, 417-426.
Ahmed AK, Johnson KA, Burchett MD, Kenny BJ (2005) The effects of heat, smoke,
leaching, scarification, temperature and NaCl salinity on the germination of
Solanum centrale (the Australian bush tomato). Seed Science and Technology
34, 33-45.
Albrecht MA, McCarthy BC (2006) Seed germination and dormancy in the medicinal
woodland herbs Collinsonia canadensis L. (Lamiaceae) and Dioscorea villosa
L. (Dioscoreaceae). Flora (Jena) 201, 24-31.
Allen PS, Meyer SE (1998) Ecological aspects of seed dormancy loss. Seed Science
Research 8, 183-191.
Allen PS, Meyer SE, Beckstead J (1995) Patterns of seed after-ripening in Bromus
tectorum L. Journal of Experimental Botany 46, 1737-1744.
Anderson DC, Ostler WK (2002) Revegetation of degraded lands at U.S. Department of
Energy and U.S. Department of Defense installations: strategies and successes.
Arid Land Research and Management 16, 197-212.
Ashby WC (1997) Soil ripping and herbicides enhance tree and shrub restoration on
stripmines. Restoration Ecology 5, 169-177.
Bainbridge DA (2007) 'A Guide for Desert and Dryland Restoration: New Hope for
Arid Lands.' (Island Press: Washington)
Baker KS, Steadman KJ, Plummer JA, Dixon KW (2005a) Seed dormancy and
germination responses of nine Australian fire ephemerals. Plant and Soil 277,
345-358.
Baker KS, Steadman KJ, Plummer JA, Merritt DJ, Dixon KW (2005b) The changing
window of conditions that promotes germination of two fire ephemerals,
Actinotus leucocephalus (Apiaceae) and Tersonia cyathiflora
(Gyrostemonaceae). Annals of Botany 96, 1225-1236.
Baker KS, Steadman KJ, Plummer JA, Merritt DJ, Dixon KW (2005c) Dormancy
release in Australian fire ephemeral seeds during burial increases germination
response to smoke water or heat. Seed Science Research 15, 339-348.

176
Banerjee MJ, Gerhart VJ, Glenn EP (2006) Native plant regeneration on abandoned
desert farmland: Effects of irrigation, soil preparation, and amendments on
seedling establishment. Restoration Ecology 14, 339-348.
Barrett-Lennard EG, Frost F, Vlahos S, Richards N (1991) Revegetating salt-affected
land with shrubs. WA Journal of Agriculture 32, 124-129.
Barrett-Lennard EG, Malcolm CV, Bathgate A (2003) 'Saltland Pastures in Australia -
A Practical Guide.' (Land Water & Wool Sustainable Grazing on Saline Lands
Sub-program)
Baskin CC, Baskin JM (1998) 'Seeds: Ecology, Biogeography and Evolution of
Dormancy and Germination.' (Academic Press: San Diego, California)
Baskin CC, Baskin JM (2003a) Overview and recommendations for future research
priorities on native seed dormancy and germination of Australian plants.
Australasian Plant Conservation 11, 2-9.
Baskin CC, Baskin JM (2004a) Determining dormancy-breaking and germination
requirements from the fewest seeds. In 'Ex Situ Plant Conservation - Supporting
Species Survival in the Wild'. (Eds EO Guerrant Jr, K Havens and M Maunder)
pp. 162-179. (Island Press: Washington, DC)
Baskin CC, Baskin JM (2004b) Germinating seeds of wildflowers, an ecological
perspective. HortTechnology 14, 467-473.
Baskin CC, Baskin JM (2007) A revision of Martin's seed classification system, with
particular reference to his dwarf-seed type. Seed Science Research 17, 11-20.
Baskin CC, Thompson K, Baskin JM (2006a) Mistakes in germination ecology and how
to avoid them. Seed Science Research 16, 165-168.
Baskin JM, Baskin CC (1976) High temperature requirement for afterripening in seeds
of winter annuals. New Phytologist 77, 619-624.
Baskin JM, Baskin CC (1979) Effect of relative humidity on afterripening and viability
in seeds of the winter annual Draba verna. Botanical Gazette 140, 284-287.
Baskin JM, Baskin CC (2003b) Classification, biogeography and phylogenetic
relationships of seed dormancy. In 'Seed Conservation - Turning Science into
Practice'. (Eds R Smith, J Dickie, S Linnington, HW Pritchard and R Probert)
pp. 517-544. (The Royal Botanic Gardens, Kew)
Baskin JM, Baskin CC (2004c) A classification system for seed dormancy. Seed Science
Research 14, 1-16.

177
Baskin JM, Baskin CC, Dixon KW (2006b) Physical dormancy in the endemic
Australian genus Stylobasium, a first report for the family Surianaceae
(Fabales). Seed Science Research 16, 229-232.
Baskin JM, Baskin CC, Li X (2000) Taxonomy, anatomy and evolution of physical
dormancy in seeds. Plant Species Biology 15, 139-152.
Baskin JM, Nan X, Baskin CC (1998) A comparative study of seed dormancy and
germination in an annual and a perennial species of Senna (Fabaceae). Seed
Science Research 8, 501-512.
Beardsell DV, Knox RB, Williams EG (1993) Germination of seeds from the fruits of
Thryptomene calycina (Myrtaceae). Australian Journal of Botany 41, 263-273.
Bell DT (1994) Interaction of fire, temperature and light in the germination response of
16 species from the Eucalyptus marginata forest of south-western Western
Australia. Australian Journal of Botany 42, 501-509.
Bell DT (1999) The process of germination in Australian species. Australian Journal of
Botany 47, 475-517.
Bell DT, Bellairs SM (1992) Effects of temperature on the germination of selected
Australian native species used in the rehabilitation of bauxite mining
disturbances in Western Australia. Seed Science and Technology 20, 47-55.
Bell DT, King LA, Plummer JA (1999) Ecophysiological effects of light quality and
nitrate on seed germination in species from Western Australia. Australian
Journal of Ecology 24, 2-10.
Bell DT, Plummer JA, Taylor SK (1993) Seed germination ecology in southwestern
Western Australia. Botanical Review 59, 24-73.
Bell DT, Rokich DP, McChesney CJ, Plummer JA (1995) Effects of temperature, light
and gibberellic acid on the germination of seeds of 43 species native to Western
Australia. Journal of Vegetation Science 6, 797-806.
Bellairs SM, Ashwath N (2007) Seed biology of tropical Australian plants. In 'Seeds:
Biology, Development and Ecology'. (Eds S Adkins, S Ashmore and S Navie)
pp. 416-427. (CABI Publishing: Wallingford, Oxfordshire)
Bellairs SM, Bell DT (1990) Temperature effects on the seed germination of ten
kwongan species from Eneabba Western Australia. Australian Journal of Botany
38, 451-458.
Bewley JD (1997) Seed germination and dormancy. Plant Cell 9, 1055-1066.
Bewley JD, Black M (1994) 'Seeds: physiology of development and germination.'
(Plenum Press: New York)

178
Borger CP, Walsh M, Scott JK, Powles SB (2007) Tumbleweeds in the Western
Australian cropping system: seed dispersal characteristics of Salsola australis.
Weed Research 47, 406-414.
Bradshaw A (1997) Restoration of mined lands - using natural processes. Ecological
Engineering 8, 255-269.
Brooker B (2000) The range and habitat characteristics of the thick-billed grasswren
(Amytornis textilis) in the Shark Bay region, Western Australia. Wildlife
Research 27, 245-256.
Burbidge AA, George AS (1978) The flora and fauna of Dirk Hartog Island Western
Australia. Journal of the Royal Society of Western Australia 60, 71-90.
Burdett AN, Simpson DG, Thompson CF (1983) Root development and plantation
establishment success. Plant and Soil 71, 103-110.
Burke A (2003) Practical measures in arid land restoration after mining: A review for
the southern Namib. South African Journal of Science 99, 413-417.
Burrows FJ (1982) Root growth of clover on compacted colliery spoil. Environmental
Geochemistry and Health 4, 156-158.
CALM (2005) 'Issues Paper: Shark Bay terrestrial reserves and proposed terrestrial
reserve additions management plan.' Department of Conservation and Land
Management, Perth, Western Australia.
Cipriotti PA, Flombaum P, Sala OE, Aguiar MR (2008) Does drought control
emergence and survival of grass seedlings in semi-arid rangelands?: An example
with a Patagonian species. Journal of Arid Environments 72, 162-174.
Clemente AS, Werner C, Maguas C, Cabral MS, Martins-Loucao MA, Correia O (2004)
Restoration of a limestone quarry: Effect of soil amendments on the
establishment of native Mediterranean sclerophyllous shrubs. Restoration
Ecology 12, 20-28.
Close DC, Bail I, Hunter S, Beadle CL (2005a) Effects of exponential nutrient-loading
on morphological and nitrogen characteristics and on after-planting performance
of Eucalyptus globulus seedlings. Forest Ecology & Management 205, 397-403.
Close DC, Beadle CL, Brown PH (2005b) The physiological basis of containerized tree
seedling 'transplant shock': a review. Australian Forestry 68, 112-120.
Cochrane A, Kelly A, Brown K, Cunneen S (2002) Relationships between seed
germination requirements and ecophysiological characteristics aid the recovery
of threatened native plant species in Western Australia. Ecological Management
and Restoration 3, 47-55.

179
Cohn MA, Hughes JA (1981) Seed dormancy in Red Rice (Oryza sativa) I. Effect of
temperature on dry-afterripening. Weed Science 29, 402-404.
Cole I, Lunt ID, Koen T (2005) Effects of sowing treatment and landscape position on
establishment of the perennial tussock grass Themeda triandra (Poaceae) in
degraded eucalyptus woodlands in southeastern Australia. Restoration Ecology
13, 552-561.
Cramer VA, Hobbs RJ, Standish RJ (2008) What's new about old fields? Land
abandonment and ecosystem assembly. Trends in Ecology & Evolution 23, 104-
112.
Cramer VA, Standish RJ, Hobbs RJ (2007) Prospects for the recovery of native
vegetation in Western Australian old fields. In 'Old Fields: Dynamics and
Restoration of Abandoned Farmland'. (Eds VA Cramer and RJ Hobbs). (Island
Press: Washington, DC)
DAFWA (2006) The Department of Agriculture and Food Western Australia
(http://www.agric.wa.gov.au) accessed 6 June 2008.
Davy AJ (2002) Establishment and manipulation of plant populations and communities
in terrestrial systems. In 'Handbook of Ecological Restoration'. (Eds MR Perrow
and AJ Davy). (Cambridge University Press: Cambridge)
Daws MI, Davies J, Pritchard HW, Brown NA, van Staden J (2007) Butenolide from
plant-derived smoke enhances germination and seedling growth of arable weed
species. Plant Growth Regulation 51, 73-82.
Debeaujon I, Koornneef M (2000) Gibberellin requirement for Arabidopsis seed
germination is determined both by testa characteristics and embryonic abscisic
acid. Plant Physiology 22, 415-424.
DEP (2001) 'Shark Bay World Heritage Property: environmental values, cultural uses,
and potential petroleum industry impacts.' Department of Environmental
Protection, Perth, Western Australia.
Dixon KW, Merritt DJ, Flematti GR, Ghisalberti EL (2008) Karrikinolide - a
phytoreactive compound derived from smoke with applications in horticulture,
ecological restoration and agriculture. Acta Horticulturae in press.
Dixon KW, Roche S, Pate JS (1995) The promotive effect of smoke derived from burnt
native vegetation on seed germination of Western Australian plants. Oecologia
101, 185-192.
DoIR (Department of Industry and Resources) (2006) Mineral and petroleum resources
(available from

180
http://www.mpr.wa.gov.au/documents/investment/CompletionCriteria-
RevisedFeb06.pdf) accessed 19 March 2008.
Doust SJ, Erskine PD, Lamb D (2006) Direct seeding to restore rainforest species:
Microsite effects on the early establishment and growth of rainforest tree
seedlings on degraded land in the wet tropics of Australia. Forest Ecology &
Management 234, 333-343.
Egler FE (1954) Vegetation science concepts I. Initial floristic composition, a factor in
old-field vegetation development. Vegetatio 4, 412-417.
Elmarsdottir A, Aradottir AL, Trlica MJ (2003) Microsite availability and establishment
of native species on degraded and reclaimed sites. Journal of Applied Ecology
40, 815-823.
Enright NJ, Lamont BB (1992) Survival, Growth and Water Relations of Banksia
Seedlings on a Sand Mine Rehabilitation Site and Adjacent Scrub-Heath Sites.
Journal of Applied Ecology 29, 663-671.
Esashi Y, Ogasawara M, Gorecki R, Leopold AC (1993) Possible mechanisms of
afterripening in Xanthium seeds. Physiologia Plantarum 87, 359-364.
Fenner M, Thompson K (2005) 'The Ecology of Seeds.' (Cambridge University Press:
Cambridge)
Fennimore SA, Foley ME (1998) Genetic and physiological evidence for the role of
gibberellic acid in the germination of dormant Avena fatua seeds. Journal of
Experimental Botany 49, 89-94.
Finch-Savage WE, Leubner-Metzger G (2006) Seed dormancy and the control of
germination. New Phytologist 171, 501-523.
Flematti GR, Ghisalberti EL, Dixon KW, Trengove RD (2004) A compound from
smoke that promotes seed germination. Science 305, 977.
Flematti GR, Ghisalberti EL, Dixon KW, Trengove RD (2005) Synthesis of the seed
germination stimulant 3-methyl-2H-furo[2,3-c]pyran-2-one. Tetrahedron Letters
46, 5719-5721.
Flores J, Jurado E (2003) Are nurse-protégé interactions more common among plants
from arid environments? Journal of Vegetation Science 14, 911-916.
Foley ME (1994) Temperature and water status of seed affect afterripening in wild oat
(Avena fatua). Weed Science 42, 200-204.
Gardner J (2001) Rehabilitating mines to meet land use objectives: bauxite mining in
the jarrah forest of Western Australia. Unasylva 52, 3-8.

181
Gentilli J (1979) 'Western Landscapes.' (The University of Western Australia Press:
Nedlands, Western Australia)
Glenn EP, Waugh WJ, Moore D, McKeon C, Nelson SG (2001) Revegetation of an
abandoned uranium millsite on the Colorado Plateau, Arizona. Journal of
Environmental Quality 30, 1154-1162.
Goodman RC, Jacobs DF, Karrfalt RP (2006) Assessing viability of northern red oak
acorns with X-rays: application for seed managers. Native Plants Journal 7,
279-283.
Grant CD, Bell DT, Koch JM, Loneragan WA (1996) Implications of seedling
emergence to site restoration following bauxite mining in Western Australia.
Restoration Ecology 4, 146-154.
Grant CD, Loneragan WA (1999) The effects of burning on the understorey
composition of 11-13 year-old rehabilitated bauxite mines in Western Australia.
Plant Ecology 145, 291-305.
Grant CD, Ward SC, Morley SC (2007) Return of Ecosystem Function to Restored
Bauxite Mines in Western Australia. Restoration Ecology 15, S94-S103.
Grappin P, Bouinot D, Sotta B, Miginiac E, Jullien M (2000) Control of seed dormancy
in Nicotiana plumbaginifolia: Post-imbibition abscisic acid synthesis imposes
dormancy maintenance. Planta 210, 279-285.
Greening Australia (2003) 'Revegetation techniques: A guide for establishing native
vegetation in Victoria.' (Greening Australia: Victoria)
Groot SPC, Karssen CM (1987) Gibberellins regulate seed germination in tomato by
endosperm weakening: a study with gibberellin-deficient mutants. Planta 171,
525-531.
Groot SPC, Kieliszewska-Rokicka B, Vermeer E, Karseen CM (1988) Gibberellin-
induced hydrolysis of endosperm cell walls in gibberellin-deficient tomato seeds
prior to radicle protrusion. Planta 174, 500-504.
Guo Q, Rundel PW, Goodall DW (1998) Horizontal and vertical distribution of desert
seed banks: patterns, causes, and implications. Journal of Arid Environments 38,
465-478.
Gwaze D, Johanson M, Hauser C (2007) Long-term soil and shortleaf pine responses to
site preparation ripping. New Forests 34, 143-152.
Hardegree SP (1996) Optimization of seed priming treatments to increase low-
temperature germination rate. Journal of Range Management 49, 87-92.

182
Hardegree SP, Van Vactor SS (2000) Germination and emergence of primed grass seeds
under field and simulated-field temperature regimes. Annals of Botany 85, 379-
390.
Herbert MA (1990) Fertiliser/site interaction on the growth and foliar nutrient levels of
Eucalyptus grandis Forest Ecology and Management 30, 247-257.
Heydecker W, Coolbear P (1977) Seed treatments for improved performance survey and
attempted prognosis. Seed Science and Technology 5, 353-426.
Heydecker W, Higgins J, Gulliver RL (1973) Accelerated germination by osmotic seed
treatment. Nature 246, 42-44.
Hilhorst H, Bentsink L, Koornneef M (2006) Dormancy and Germination. In 'Handbook
of seed science and technology'. (Ed. AS Basra) pp. xviii + 795. (Food Products
Press: Binghamton)
Hilhorst HWM, Karssen CM (1992) Seed dormancy and germination: the role of
abscisic acid and gibberellins and the importance of hormone mutants. Plant
Growth Regulation 11, 225-238.
Hobbs RJ, Cramer VA (2007) Old field dynamics: Regional and local differences and
lessons for ecology and restoration. In 'Old Fields; Dynamics and Restoration of
Abandoned Farmland'. (Eds VA Cramer and RJ Hobbs). (Island Press:
Washington, DC)
Hocking RM (1990) Carnarvon Basin. In 'Geology and Mineral Resources of Western
Australia' pp. 457-495. (Western Australia Geological Survey: Memoir 3)
Hocking RM, Moors HT, van de Graaff WJE (1987) 'Geology of the Carnarvon Basin,
Western Australia.' Western Australia Geological Survey, Bulletin 133.
Holmes PM, Richardson DM (1999) Protocols for restoration based on recruitment
dynamics, community structure, and ecosystem function: Perspectives from
South African fynbos. Restoration Ecology 7, 215-230.
Hopper SD (2007) Landscape age, evolution and conservation of biodiversity in
Mediterranean regions. In 'Proceedings of the MEDECOS XI 2007 Conference,
2-5 September'. (Eds DP Rokich, G Wardell-Johnson, CJ Yates, JC Stevens,
KW Dixon, R McLellan and G Moss) pp. 123-124. (Kings Park and Botanic
Garden) Perth, Australia.
Hopper SD, Gioia P (2004) The Southwest Australian Floristic Region: evolution and
conservation of a global hot spot of biodiversity. Annual Review of Ecology,
Evolution, and Systematics 35, 623-650.

183
Hoyle GL, Daws MI, Steadman KJ, Adkins SW (2008) Mimicking a Semi-arid Tropical
Environment Achieves Dormancy Alleviation for Seeds of Australian Native
Goodeniaceae and Asteraceae. Annals of Botany 101, 701-708.
Hüttermann A, Zommorodi M, Reise K (1999) Addition of hydrogels to soil for
prolonging the survival of Pinus halepensis seedlings subjected to drought. Soil
& Tillage Research 50, 295-304.
ISTA (2007) 'International rules for seed testing, edition 2007.' (The International Seed
Testing Association: Bassersdorf)
Jefferson LV (2001) The biology and ecology of Maireana and Enchylaena: Intra- and
inter- specific competition in plant communities in the eastern goldfields of
Western Australia. PhD thesis, Curtin University of Technology.
Jefferson LV (2004) Implications of plant density on the resulting community structure
of mine site land. Restoration Ecology 12, 429-438.
Jurado E, Westoby M (1992) Germination biology of selected central Australian plants.
Australian Journal of Ecology 17, 341-348.
Karssen CM, Haigh A, van der Toorn P, Weges R (1989) 'Physiological Mechanisms
involved in Seed Priming.' (Plenum Press: New York)
Kermode AR (2005) Role of abscisic acid in seed dormancy. Journal of Plant Growth
Regulation 24, 319-344.
Knight AJ, Beale PE, Dalton GS (1998) Direct seeding of native trees and shrubs in low
rainfall areas and on non-wetting sands in South Australia. Agroforestry Systems
39, 225-239.
Koch JM (2007) Restoring a jarrah forest understorey vegetation after bauxite mining in
Western Australia. Restoration Ecology 15, S26-S39.
Koch JM, Hobbs RJ (2007) Synthesis: is Alcoa successfully restoring a jarrah forest
ecosystem after bauxite mining in Western Australia? Restoration Ecology 15,
S137-S144.
Koch JM, Ward SC (1994) Establishment of understory vegetation for rehabilitation of
bauxite-mined areas in the jarrah forest of Western Australia. Journal of
Environmental Management 41, 1-15.
Koch JM, Ward SC, Grant CD (1997) Soil seed bank research for mining rehabilitation:
a case study of bauxite in a species-rich ecosystem in south-west Western
Australia. In 'Second Australian Native Seed Biology for Restoration
Workshop'. (Eds SM Bellairs and JM Osborne) pp. 53-62. (Australian Centre
for Mining Rehabilitation Research).

184
Koch JM, Ward SC, Grant CD, Ainsworth GL (1996) Effects of bauxite mine
restoration operations on topsoil seed reserves in the jarrah forest of Western
Australia. Restoration Ecology 4, 368-376.
Koornneef M, Bentsink L, Hilhorst H (2002) Seed dormancy and germination. Current
Opinion in Plant Biology 5, 33-36.
Kucera B, Cohn MA, Leubner-Metzger G (2005) Plant hormone interactions during
seed dormancy release and germination. Seed Science Research 15, 281-307.
Kulkarni MG, Sparg SG, Van Staden J (2007) Germination and post-germination
response of Acacia seeds to smoke-water and butenolide, a smoke-derived
compound. Journal of Arid Environments 69, 177-187.
Lambers H, Chapin FS, Pons TL (1998) 'Plant Physiological Ecology.' (Springer: New
York)
Lamont BB, Witkowski ETF, Enright NJ (1993) Post-fire litter microsites: safe for
seeds, unsafe for seedlings. Ecology 74, 501-512.
Latz P (1995) 'Bushfires and Bushtucker: Aboriginal Plant Use in Central Australia.'
(IAD Press: Alice Springs, Northern Territory)
Leopold AC, Glenister R, Cohn MA (1988) Relationship between water content and
afterripening in red rice. Physiologia Plantarum 74, 659-662.
López BC, Holmgren M, Sabaté S, Gracia CA (2008) Estimating annual rainfall
threshold for establishment of tree species in water-limited ecosystems using
tree-ring data. Journal of Arid Environments 72, 602-611.
Luce CH (1997) Effectiveness of road ripping in restoring infiltration capacity of forest
roads. Restoration Ecology 5, 265-270.
Ludwig JA, Tongway DJ (1995) Spatial organisation of landscapes and its function in
semi-arid woodlands, Australia. Landscape Ecology 10, 51-63.
Ludwig JA, Tongway DJ, Marsden SG (1994) A flow-filter model for stimulating the
conservation of limited resources in spatially heterogeneous semi-arid
landscapes. Pacific Conservation Biology 1, 209-213.
Martin AC (1946) The comparative internal morphology of seeds. American Midland
Naturalist 36, 513-660.
Mattiske (1996) 'Flora and Vegetation - Useless Loop, Shark Bay.'
McIvor JG, Howden SM (2000) Dormancy and germination characteristics of
herbaceous species in the seasonally dry tropics of northern Australia. Austral
Ecology 25, 213-222.

185
McNabb K, Vanderschaaf C (2005) Growth of graded sweetgum 3 years after root and
shoot pruning. New Forests 29, 313-320.
Merritt DJ, Kristiansen M, Flematti GR, Turner SR, Ghisalberti EL (2006) Effects of a
butenolide present in smoke on light-mediated germination of Australian
Asteraceae. Seed Science Research 16, 29-35.
Merritt DJ, Turner SR, Clarke S, Dixon KW (2007) Seed dormancy and germination
stimulation syndromes for Australian temperate species. Australian Journal of
Botany 55, 336-344.
Merritt DJ, Turner SR, Commander LE, Dixon KW (2004) Integrated seed science for
delivering a better restoration outcome. In 'Proceedings of the Fifth Australian
Workshop on Native Seed Biology'. (Eds SW Adkins, PJ Ainsley, SM Bellairs,
DJ Coates and LC Bell) pp. 69-76. (Australian Centre for Minerals Extension
and Research) Brisbane, Australia.
Milton SJ (2001) Rethinking ecological rehabilitation in arid and winter rainfall regions
of southern Africa. South African Journal of Science 97, 47-48.
Montalvo AM, McMillan PA, Allen EB (2002) The relative importance of seeding
method, soil ripping, and soil variables on seeding success. Restoration Ecology
10, 52-67.
Moore RP (1972) Tetrazolium staining for assessing seed quality. In 'Seed ecology :
proceedings of the Nineteenth Easter School in Agricultural Science'. (Ed. W
Heydecker) pp. 347-366. (London, Butterworths) University of Nottingham.
Mott JJ (1972) Germination studies on some annual species from an arid region of
Western Australia. Journal of Ecology 60, 293-304.
Mott JJ, Groves RH (1981) Germination strategies. In 'The Biology of Australian
Plants'. (Eds JS Pate and AJ McComb) pp. 307-341. (University of Western
Australia Press: Nedlands, Western Australia)
Mullan GD, White PJ (2001) 'Seedling Quality: Making informed choices.' Bushcare
and the Department of Conservation and Land Management, Western Australia.
Murdoch AJ, Ellis RH (2000) Dormancy, viability and longevity. In 'Seeds: the ecology
of regeneration in plant communities'. (Ed. M Fenner) pp. 183-214. (CABI
Publishing: Wallingford, Oxfordshire)
Nikolaeva MG (2001) Ecological and physiological aspects of seed dormancy and
germination (review of investigations for the last century). Botanicheskii
Zhurnal 86, 1-14.

186
Noble JC, Whalley RD (1978) The biology and autecology of Nitraria L. in Australia.
II. Seed germination, seedling establishment and response to salinity. Australian
Journal of Ecology 3, 165-177.
Norman MA, Koch JM (2008) The effect of in situ seed burial on dormancy break in
three woody-fruited species (Ericaceae and Proteaceae) endemic to Western
Australia. Australian Journal of Botany 56, 493-500.
Norman MA, Koch JM, Grant CD, Morald TK, Ward SC (2006a) Vegetation
succession after bauxite mining in Western Australia. Restoration Ecology 14,
278-288.
Norman MA, Plummer JA, Koch JM, Mullins GR (2006b) Optimising smoke
treatments for jarrah (Eucalyptus marginata) forest rehabilitation. Australian
Journal of Botany 54, 571-581.
Ooi MKJ (2007) Dormancy classification and potential dormancy-breaking cues for
shrub species from fire-prone south-eastern Australia. In 'Seeds: Biology,
Development and Ecology'. (Eds S Adkins, S Ashmore and S Navie) pp. 205-
216. (CABI Publishing: Wallingford, Oxfordshire)
Ooi MKJ, Auld TD, Whelan RJ (2007) Distinguishing between persistence and
dormancy in soil seed banks of three shrub species from fire-prone southeastern
Australia. Journal of Vegetation Science 18, 405-412.
Ord R (2007) Assessing the effects of wind erosion and invertebrate activity on
broadcast seed removal in Banksia woodland restoration. Honours thesis, The
University of Western Australia.
Paczkowsa G, Chapman AR (2000) 'The Western Australian flora: a descriptive
catalogue.' (Wildflower Society of Western Australia: Perth)
Page MJ, Baxter GS, Lisle AT (2006) Evaluating the adequacy of sampling germinable
soil seed banks in semi-arid systems. Journal of Arid Environments 64, 323-341.
Paschke MW, DeLeo C, Redente EF (2000) Revegetation of roadcut slopes in Mesa
Verde National Park, U.S.A. Restoration Ecology 8, 276-282.
Pate JS, Casson NE, Rullo J, Kuo J (1985) Biology of fire ephemerals of the sandplains
of the kwongan of southwestern Australia. Australian Journal of Plant
Physiology 12, 641-656.
Paynter BH, Dixon KW (1990) Seed Viability and Embryo Decline in Geleznowia-
Verrucosa Turcz. Rutaceae. Scientia Horticulturae 45, 149-158.

187
Petersen SL, Roundy BA, Bryant RM (2004) Revegetation methods for high-elevation
roadsides at Bryce Canyon National Park, Utah. Restoration Ecology 12, 248-
257.
Philippi T (1993) Bet-hedging germination of desert annuals: beyond the first year.
American Naturalist 142, 474-487.
Plummer JA, Bell DT (1995) The effect of temperature, light and gibberellic acid (GA3)
on the germination of Australian everlasting daisies (Asteraceae, Tribe Inuleae).
Australian Journal of Botany 43, 93-102.
Plummer JA, Rogers AD, Turner DW, Bell DT (2001) Light, nitrogenous compounds,
smoke and GA3 break dormancy and enhance germination in the Australian
everlasting daisy, Schoenia filifolia subsp. subulifolia. Seed Science and
Technology 29, 321-330.
Prach K, Hobbs RJ (2008) Spontaneous Succession versus Technical Reclamation in the
Restoration of Disturbed Sites. Restoration Ecology 16, 363-366.
Probert R (2000) The role of temperature in the regulation of seed dormancy and
germination. In 'Seeds: the ecology of regeneration in plant communities'. (Ed.
M Fenner) pp. 261-292. (CABI Publishing: Wallingford, Oxfordshire)
Pugnaire FI, Haase P, Puigdefabregas J (1996) Facilitation between higher plant species
in a semiarid environment. Ecology 77, 1420-1426.
Pugnaire FI, Lazaro R (2000) Seed bank and understorey species composition in a semi-
arid environment: The effect of shrub age and rainfall. Annals of Botany 86,
807-813.
Quinlivan BJ (1961) The effect of constant and fluctuating temperatures in the
permeability of the hard seeds of some legume species. Australian Journal of
Agricultural Research 19, 1009-1022.
Quinlivan BJ (1966) The relationship between temperature fluctuations and the
softening of hard seeds of some legume species. Australian Journal of
Agricultural Research 17, 625-631.
Richmond GS, Ghisalberti EL (1994) Seed dormancy and germination mechanisms in
Eremophila (Myoporaceae). Australian Journal of Botany 42, 705-715.
Robley AJ, Short J, Bradley S (2001) Dietary overlap between the burrowing bettong
(Bettongia lesueur) and the European rabbit (Oryctolagus cuniculus) in semi-
arid coastal Western Australia. Wildlife Research 28, 341-349.

188
Roche S, Dixon KW, Pate JS (1997a) Seed ageing and smoke: Partner cues in the
amelioration of seed dormancy in selected Australian native species. Australian
Journal of Botany 45, 783-815.
Roche S, Dixon KW, Pate JS (1998) For everything a season: Smoke-induced seed
germination and seedling recruitment in a Western Australian Banksia
woodland. Australian Journal of Ecology 23, 111-120.
Roche S, Koch JM, Dixon KW (1997b) Smoke enhanced seed germination for mine
rehabilitation in the southwest of Western Australia. Restoration Ecology 5, 191-
203.
Rodríguez-Echeverría S, Pérez-Fernández MA (2005) Potential use of Iberian shrubby
legumes and rhizobia inoculation in revegetation projects under acidic soil
conditions. Applied Soil Ecology 29, 203-208.
Rokich DP (1999) Banksia Woodland Restoration. PhD thesis, The University of
Western Australia.
Rokich DP, Bell DT (1995) Light quality and intensity effects on the germination of
species from the Jarrah (Eucalyptus marginata) forest of Western Australia.
Australian Journal of Botany 43, 169-179.
Rokich DP, Dixon KW (2007) Recent advances in restoration ecology, with a focus on
the Banksia woodland and the smoke germination tool. Australian Journal of
Botany 55, 375-389.
Rokich DP, Dixon KW, Sivasithamparam K, Meney KA (2000) Topsoil handling and
storage effects on woodland restoration in Western Australia. Restoration
Ecology 8, 196-208.
Rokich DP, Dixon KW, Sivasithamparam K, Meney KA (2002) Smoke, mulch, and
seed broadcasting effects on woodland restoration in Western Australia.
Restoration Ecology 10, 185-194.
Rokich DP, Meney KA, Dixon KW, Sivasithamparam K (2001) The impact of soil
disturbance on root development in woodland communities in Western
Australia. Australian Journal of Botany 49, 169-183.
Schütz W, Milberg P, Lamont BB (2002) Seed dormancy, after-ripening and light
requirements of four annual Asteraceae in south-western Australia. Annals of
Botany 90, 707-714.
Schwenke GD, Mulligan DR, Bell LC (2000) Soil stripping and replacement for the
rehabilitation of bauxite-mined land at Weipa. I. Initial changes to soil organic
matter and related parameters. Australian Journal of Soil Research 38, 345-369.

189
Sharif-Zadeh F, Murdoch AJ (2001) The effects of temperature and moisture on after-
ripening of Cenchrus ciliaris seeds. Journal of Arid Environments 49, 823-831.
Shark Bay Salt Joint Venture (1993) 'Environmental Report, December 1993.'
Shark Bay Salt Joint Venture (1994) 'Environmental Report, December 1994.'
Shark Bay Salt Joint Venture (1995) 'Environmental Report, 1995.'
Shark Bay Salt Joint Venture (1996) 'Environmental Report, December 1996.'
Shark Bay Salt Joint Venture (1997) 'Environmental Report, December 1997.'
Shark Bay Salt Joint Venture (1998) 'Triennial Environmental Report, January 1996 -
December 1998.'
Shark Bay Salt Joint Venture (2001) 'Triennial Environmental Report, 1999 - 2001.'
Shark Bay Salt Joint Venture (2002) 'Annual Environmental Report, 2002.'
Shark Bay Salt Joint Venture (2003) 'Annual Environmental Report, December 2003.'
Shark Bay Salt Joint Venture (2004) 'Triennial Environmental Report, January 2002 -
December 2004.'
Shark Bay Solar Salt Industry Agreement Act (1983).
Short J, Turner B (2000) Reintroduction of the burrowing bettong Bettongia lesueur
(Marsupialia: Potoroidae) to mainland Australia. Biological Conservation 96,
185-196.
Simcock RC, Parfitt RL, Skinner MF, Dando J, Graham JD (2006) The effects of soil
compaction and fertilizer application on the establishment and growth of Pinus
radiata. Canadian Journal of Forest Research 36, 1077-1086.
Snyman HA (2003) Revegetation of bare patches in a semi-arid rangeland of South
Africa: an evaluation of various techniques. Journal of Arid Environments 55,
417-432.
Standish RJ, Cramer VA, Wild SL, Hobbs RJ (2007) Seed dispersal and recruitment
limitation are barriers to native recolonization of old-fields in Western Australia.
Journal of Applied Ecology 44, 435-445.
Steadman KJ, Crawford AD, Gallagher RS (2003) Dormancy release in Lolium rigidum
seeds is a function of thermal after-ripening time and seed water content.
Functional Plant Biology 30, 345-352.
Stefaniski A (1998) Manipulating seed dormancy in bush tomato (Solanum centrale).
Honours thesis, The University of Sydney.
Stevens JC, Barrett-Lennard EG, Dixon KW (2006) Enhancing the germination of three
fodder shrubs (Atriplex amnicola, A. nummularia, A. undulata;

190
Chenopodiaceae): implications for the optimisation of field establishment.
Australian Journal of Agricultural Research 57, 1279-1289.
Stevens JC, Merritt DJ, Flematti GR, Ghisalberti EL, Dixon KW (2007) Seed
germination of agricultural weeds is promoted by the butenolide 3-methyl-2H-
furo[2,3-c]pyran-2-one under laboratory and field conditions. Plant and Soil
298, 113-124.
Sweedman L, Merritt DJ (2006) 'Australian Seeds - A Guide to their Collection,
Identification and Biology.' (CSIRO Publishing: Collingwood, Victoria)
Szota C, Veneklaas EJ, Koch JM, Lambers H (2007) Root architecture of Jarrah
(Eucalyptus marginata) trees in relation to post-mining deep ripping in Western
Australia. Restoration Ecology 15, S65-S73.
Tacey WH, Glossop BL (1980) Assessment of Topsoil Handling Techniques for
Rehabilitation of Sites Mined for Bauxite within the Jarrah Forest of Western
Australia. Journal of Applied Ecology 17, 195-202.
Terui K, Okagami N (1993) Temperature effects on seed germination of East Asian and
Tertiary relict species of Dioscorea (Dioscoreaceae). American Journal of
Botany 80, 493-499.
Tieu A, Dixon KW, Meney KA, Sivasithamparam K (2001a) The interaction of heat
and smoke in the release of seed dormancy in seven species from southwestern
Western Australia. Annals of Botany 88, 259-265.
Tieu A, Dixon KW, Meney KA, Sivasithamparam K (2001b) Interaction of soil burial
and smoke on germination patterns in seeds of selected Australian native plants.
Seed Science Research 11, 69-76.
Timmer VR (1997) Exponential nutrient loading: a new fertilization technique to
improve seedling performance on competitive sites. New Forests 13, 279-299.
Turner SR, Merritt DJ, Baskin CC, Dixon KW, Baskin JM (2005a) Physical dormancy
in seeds of six genera of Australian Rhamnaceae. Seed Science Research 15, 51-
58.
Turner SR, Merritt DJ, Baskin JM, Baskin CC, Dixon KW (2006a) Combinational
dormancy in seeds of the Western Australian endemic species Diplopeltis
huegelii (Sapindaceae). Australian Journal of Botany 54, 565-570.
Turner SR, Merritt DJ, McLean B, Rokich DP, Pearce B (2005b) 'Bold Park Research
Final Report.' Botanic Gardens and Parks Authority, Perth, Western Australia.

191
Turner SR, Merritt DJ, Ridley E, Commander LE, Baskin JM, Baskin CC, Dixon KW
(2006b) Ecophysiology of seed dormancy in the Australian endemic species
Acanthocarpus preissii (Dasypogonaceae). Annals of Botany 98, 1137-1144.
Turner SR, Pearce B, Rokich DP, Dunn RR, Merritt DJ, Majer J, Dixon KW (2006c)
Influence of polymer seed coatings, soil raking and time of sowing on seedling
performance in post mining restoration. Restoration Ecology 14, 267-277.
Vleeshouwers LM, Bouwmeester HJ, Karssen CM (1995) Redefining seed dormancy:
An attempt to integrate physiology and ecology. Journal of Ecology 83, 1031-
1037.
Walters C (1998) Understanding the mechanisms and kinetics of seed aging. Seed
Science Research 8, 223-244.
Ward SC, Koch JM, Ainsworth GL (1996) The effect of timing of rehabilitation
procedures on the establishment of a jarrah forest after bauxite mining.
Restoration Ecology 4, 19-24.
Ward SC, Koch JM, Nichols OG (1990) Bauxite mine rehabilitation in the Darling
Range, Western Australia. Proceedings of the Ecological Society of Australia
16, 557-565.
Western Australian Herbarium (1998–) FloraBase — The Western Australian Flora.
Department of Conservation and Land Management
(http://florabase.calm.wa.gov.au/ ) accessed 1 March 2005.
Williams RR, Holliday KC, Bennell MR (1989) Cultivation of the pink mulla mulla
Ptilotus exaltatus Nees 1. Seed germination and dormancy. Scientia
Horticulturae 40, 267-274.
Williamson JR, Neilsen WA (2003) The effect of soil compaction, profile disturbance
and fertilizer application on the growth of eucalypt seedlings in two glasshouse
studies. Soil & Tillage Research 71, 95-107.
Wills R, Dogramaci S (2000) 'Carnarvon Basin, Hydrogeology and Hydrochemistry of
the Confined Groundwater Systems, WA ' Western Australia Water and Rivers
Commission, Hydrogeology Report 170, (unpublished).
Windsor DM, Clements A (2001) A germination and establishment field trial of
Themeda australis (kangaroo grass) for mine site restoration in the Central
Tablelands of New South Wales. Restoration Ecology 9, 104-110.
Winkel VK, Roundy BA, Cox JR (1991) Influence of Seedbed Microsite Characteristics
on Grass Seedling Emergence. Journal of Range Management 44, 210-214.

192
Winston P, Bates D (1960) Saturated solutions for the control of humidity in biological
research. Ecology 41, 232-237.
Woodhouse J, Johnson MS (1991) Effect of Superabsorbent Polymers on Survival and
Growth of Crop Seedlings. Agricultural Water Management 20, 63-70.
Yates CJ, Hobbs RJ (1997) Woodland restoration in the Western Australian wheatbelt:
A conceptual framework using a state and transition model. Restoration Ecology
5, 28-35.
Yates CJ, Hobbs RJ, Atkins L (2000) Establishment of perennial shrub and tree species
in degraded Eucalyptus salmonophloia (salmon gum) remnant woodlands:
Effects of restoration treatments. Restoration Ecology 8, 135-143.
Zaczek JJ, Steiner KC, Bowersox TW (1997) Northern red oak planting stock: 6-year
results. New Forests 13, 175-189.
Zahreddine HG, Struve DK, Quigley M (2004) Growing Pinus nigra seedlings in
Spinout(TM)-treated containers reduces root malformation and increases growth
after transplanting. Journal of Environmental Horticulture 22, 176-182.

193
Appendices
Appendix 1. Shark Bay Salt pond system transected by roads and bunds and
surrounded by borrow pits (shown in green, and labelled with letters).

194
Appendix 2. Aerial photograph showing the Shark Bay Salt pond system in the middle
of the peninsula, transected by roads and bunds and surrounded by borrow pits. The
borrow pits used in this study are indicated.

195
Appendix 3. Species present in each of the vegetation associations found in the Shark
Bay Salt Joint Venture lease at Useless Loop, August 1996, from Mattiske (1996).
Associations 1-12 are shrublands, associations 13-14 are herblands, association 15 is the
halophytic complex and associations 16-17 are the coastal complexes
Shrublands
1. Closed to Open Tall Shrubland of Melaleuca cardiophylla, Acacia bivenosa and
Alyogyne huegelii thickets in deep sand on midslopes of sand dunes on the western
side of the Useless Inlet.
2. Closed to Open Tall Shrubland of Acacia ligulata with bare ground in flat between
sand dunes on red sandy soil.
3. Open Shrubland of Acacia bivenosa, Acacia ligulata on limestone plate above
birridas.
4. Shrubland of Thryptomene baeckeacea with scattered taller emergent shrubs
dominated by Acacia ligulata on slopes of cream sand dunes.
5. Closed to Low Shrubland of Melaleuca huegelii subsp. pristicensis thickets fringing
inlets and birridas.
6. Open Low Shrubland of Atriplex species, Salsola kali and mixed species of shrubs
with a few emergent Pittosporum phylliraeoides subsp. phylliraeoides on relatively
flat area between sand dunes on Useless Inlet.
7. Closed to Open Low Shrubland of Thryptomene baeckeacea, Salsola kali, Rhagodia
preissii subsp. obovata, Atriplex bunburyana, and Acacia tetragonophylla with
occasional emergent Acacia ligulata, Acacia rostellifera and/or Acacia
sclerosperma on mid to upper slopes of sand dunes of Useless Inlet.
8. Low Shrubland of Thryptomene baeckeacea with Plectrachne bromoides, Melaleuca
cardiophylla and emergent Acacia ligulata on the slopes of cream to red sand dunes.
9. Low Closed to Open Shrubland with occasional emergent Acacia ligulata over
Triodia plurinervata and/or the Declared Rare species, Plectrachne bromoides on
red sand dunes, occasionally with limestone pebbles large than 20cm, on the lower
to upper slopes above birridas.
10. Shrubland of Diplolaena grandiflora and other shrubs and annuals including
Priority 2 species, Rhodanthe oppositifolia subsp. ornata on shale limestone
outcrops with eroded, sand accumulated depressions.
11. Low Closed Shrubland of Melaleuca sp. Shark Bay and Triodia plurinervata with
mixed shrubs, on midslopes to upper slopes of sand dunes on Freycinet Reach with
exposed limestone rocks outcropping.
12. Closed to Open Low Heath dominated by Melaleuca cardiophylla with scattered
emergent taller shrubs of Acacia species with large areas of mixed Asteraceae
species, in creamy-yellow sand on upper slopes of dunes.
Herblands
13. Low Herbland with Muellerolimon salicorneaceum, Frankenia setosa and Olearia
axillaris subsp. obovata on flats beside the tidal inlet in clay or shale soil with
subangular pebbles larger than 20cm.
14. Low Herbland of Wilsonia humilis on shale flats surrounding birridas.
Halophytic Complex
15. Halophytic Complex dominated by Halosarcia species on clay soil.
Coastal Complexes 16. Coastal Complex of Spinifex longifolius, Olearia spp. Acanthocarpus preissii and
low shrubs, including Nitraria billardieri, on calcareous sand.
17. Mangrove Coastal Complex of Avicennia maritima flats surrounding the inlets and
some birridas in clay.

196
Appendix 3. cont.
Vegetation Association
Species 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Abutilon geranioides + +
Abutilon sp. Hamelin
(A.M. Ashby 2196) + +
Acacia bivenosa + + + +
Acacia idiomorpha var.
(Maslin 3692) + + +
Acacia ligulata + + + + + + + +
Acacia rostellifera + + +
Acacia sclerosperma +
Acacia tetragonophylla + + + + + +
Acanthocarpus preissii + +
Acanthocarpus robustus + + + + + + +
Actinobole drummondiana +
Alyogyne cuneiformis + + + + + + +
Alyogyne huegelii subsp.
huegelii +
Alyogyne pinoniana subsp.
pinoniana +
Anthobolus foveolatus +
Anthocercis littorea +
Anthocercis sp. Shark Bay
(Aplin 3335) + +
Aphanopetalum
clematideum + + + +
Atriplex bunburyana + + + + + + + + + + + + + + +
Atriplex ?cinerea +
Atriplex ?nummularia +
Avena barbata + + + + +
Avicennia marina +
Beyeria cinerea + + + + + +
Bossiaea walkeri +
Brachyscome ciliocarpa + + + + + +
Brachyscome latisquamea + + + + + + +
Bromus arenarius + + +
Calandrinia polyandra + + + + + + + + + + +
Calotis multicaulis + +
Carpobrotus aff. rossii + + + + + + +
Cassytha pomiformis + + +
Cassytha racemosa +
* Centaurea melitensis +
Centrolepis humillima + +
Cephalipterum
drummondii + + + +
Chenopodium
gaudichaudianum + + +
Commersonia sp. + + +
Commicarpus australis +

197
Appendix 3. cont.
Vegetation Association
Species 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Crassula colorata var.
colorata + + + + + + + +
Cuscuta epithymum + + +
Cymbopogon ambiguus +
Dampiera incana var.
fuscescens +
Dianella revoluta + + + + +
Dichopogon tyleri +
Dioscorea hastifolia + +
Diplolaena grandiflora + + + + + + +
Diplopeltis intermedia var.
intermedia +
Dodonaea bursariifolia +
Duboisia hopwoodii +
Dysphania plantaginella + + + + +
* Ehrharta brevifolia +
Enchylaena tomentosa + + + + + + +
Eragrostis barrelieri + + + +
Eremophila clarkei +
Eremophila glabra subsp.
albicans + + + +
Eremophila oldfieldii + +
* Erodium cicutarium + + + + + +
Erodium cygnorum subsp.
cygnorum + + + + + +
Euphorbia australis + + + + + + + +
Euphorbia boophthona + + + + + + + +
Exocarpos aphyllus + + + + + +
Frankenia pauciflora + + + + + + + +
Frankenia setosa + + + + + +
Glycine canescens +
Goodenia beardiana + + + + + +
Grevillea brachystachya +
Gunniopsis septifraga +
Halgania littoralis + + + + +
Haloragis trigonocarpa +
Halosarcia bidens subsp.
indica + + + +
Halosarcia halocnemoides + + + +
Halosarcia pruinosa + + + +
Halosarcia pterygosperma
subsp. denticulata + +
Halosarcia sp. +
Heterodendrum
oleaefolium + + +
* Hypochaeris glabra + + +
Indigofera boviperda + +

198
Appendix 3. cont.
Vegetation Association
Species 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Jasminum calcareum + + +
Jasminum didymum +
Jasminum sp. +
Lawrencia densiflora + +
Lawrencia viridigrisea + +
Lepidium phlebopetalum + + + + +
Leptosema brachycarpum + +
Lomandra maritima +
Lotus australis +
Loxocarya aspera + +
Maireana stipitata + + + + +
Maireana tomentosa +
Maireana trichoptera + +
Marsdenia australis +
Marsdenia graniticola +
Melaleuca cardiophylla + + + + + + + + + +
Melaleuca huegelii subsp.
pristicensis + + + +
Melaleuca sp. Shark Bay +
Mirbelia ramulosa + + +
Mirbelia sp. + +
Muellerolimon
salicorniaceum + + +
Myoporum insulare + + +
Nicotiana occidentalis
subsp. hesperis + + + + + + + +
Nitraria billardierei + + +
Olearia axillaris subsp.
obovata + + + +
Olearia dampieri subsp.
dampieri + +
Olearia occidentissima + +
Olearia revoluta +
Opercularia vaginata + + +
Ophioglossum lusitanicum +
Paractaenum novae-
hollandiae subsp. novae-
hollandiae + + + + +
Parietaria debilis + + + + +
Phyllanthus calycinus +
Phyllanthus fuernrohrii +
Pimelea gilgiana + +
Pimelea microcephala
subsp. microcephala + + + + + + + + +
Pittosporum
phylliraeoides var.
phylliraeoides + + + + + + + +
Pityrodia cuneata

199
Appendix 3. cont.
Vegetation Association
Species 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Plectrachne bromoides + + +
Podolepis canescens +
Podotheca angustifolia + + +
Podotheca gnaphalioides + + + + +
* Polycarpon tetraphyllum +
Poranthera microphylla + + +
Ptilotus divaricatus var.
divaricatus + + + + +
Ptilotus exaltatus + + + +
Ptilotus gaudichaudii
subsp. parviflorus + +
Ptilotus obovatus + +
Ptilotus villosiflorus + +
* Raphanus raphanistrum + + + +
Rhagodia baccata + +
Rhagodia latifolia subsp.
latifolia + +
Rhagodia preissii subsp.
obovata + + + + + + + + +
Rhodanthe humboldtianum + +
Rhodanthe oppositifolia
subsp. ornata +
Rhodanthe polycephala +
* Rostraria pumila
Salsola kali + + + + + + + + +
Samolus repens var.
paucifolius + +
Santalum acuminatum + + + +
Santalum spicatum + +
Sarcocornia blackiana +
Sarcostemma viminale
subsp. australe +
Scaevola anchusifolia + +
Scaevola crassifolia + + + + +
Scaevola spinescens + + + + +
Scaevola tomentosa + + + + + + +
Schoenia ayersii + + + +
Senecio lautus subsp.
dissectifolius +
Senna glutinosa subsp.
chatelainiana + + + + + +
Sida calyxhymenia + + + + + +
* Silene gallica +
Solanum hesperium +
Solanum orbiculatum + + + + + +
* Sonchus oleraceus + + + + + + +
Spinifex longifolius + + + + +
Sporobolus virginicus + + + +

200
Appendix 3. cont.
Vegetation Association
Species 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Stenanthemum
complicatum +
Stenanthemum
divaricatum +
Stenopetalum pedicellare + + + +
Stipa elegantissima + +
Stipa nitida + + + +
Stylobasium spathulatum + +
Swainsona longicarinata + + + + + +
Sysymbrium erysimoides + + + +
Templetonia retusa + +
Tetragonia implexicoma + + + +
Threlkeldia diffusa + + + + + + +
Thryptomene baeckeacea + + + + + + + + +
Thysanotus patersonii + + + + +
Thysanotus speckii + + +
Trichanthodium
scarlettianum + + + + + + +
Triglochin aff.
calcitrapum +
Triglochin centrocarpa + + + +
Triglochin mucronata +
Triglochin trichophora + + +
Triglochin trichosperma +
Triodia plurinervata + +
* Urospermum picroides + + + + +
Waitzia podolepis + + + +
Westringia dampieri +
Wilsonia humilis + +
Wurmbea monantha + + + + + + +
Zygophyllum apiculatum + + + +
Zygophyllum aurantiacum +
Zygophyllum fruticulosum + + + + + + + + +
Zygophyllum simile +

201
Appendix 4. Distribution maps of the 19 species in this study (Western Australian
Herbarium 1998–)

202
Appendix 4. cont.

203
Appendix 5. Borrow pit status from 1993 to 2004 compiled from Shark Bay
Salt‘s Annual and Triennial Environmental reports. Pit 1993 1994
A Active. Current salvage yard.
B Inactive. Inactive, old asbestos dump.
C Inactive. Inactive, blown out sand dune area. No vegetation.
D Inactive. Inactive, sand dune blow out, no topsoil. Nitraria billardierei
seed was planted in 1992, germination occurred but no seedlings
survived.
E Inactive. Inactive sand dune blow out. Small pockets of vegetation are
present.
F Nth Inactive. Prolific salt bush on quarry floor
G Wst Under rehabilitation. Deep ripped and sealed. Germination but low survival. Fenced in
June.
G Sth Deep ripped, contoured,
mulched, fertilised and seeded
by June. Germination high but
survival low.
Topsoil has been spread
G Nth
H
H Nth Active.
H Sth Active. Active. As this pit is developed, topsoil and vegetation will be
stockpiled for future rehabilitation
I
J Wst Inactive. The main pit was closed and blended in April
J Est Inactive.
K Active. Active
L Under rehabilitation. Inactive, closed.
M Inactive, closed 1990. Old sand stone blowout, arrested by
mulching and spreading topsoil. Vegetation has returned.
M Nth Under rehabilitation.
M Sth Under rehabilitation.
N Inactive.
O Opened up in late 1993. Topsoil was stockpiled and replaced
upon completion. Ripped and sprayed with terrolas by January
1994.
O Sth topsoiled and terrolassed.
P Under rehabilitation. Cliff faces were battered in August, blending the pit into the
existing topography. The area is currently being mulched.
P-1
Q Under rehabilitation.
Q1
R Under rehabilitation. Inactive, closed 1990. Vegetation is slowly returning and could
benefit from seed.
S Part active, part under
rehabilitation.
Progressive rehabilitation, active
T Opened in May 1993 to supply the town with topsoil. A large
stockpile was pushed up and the area was closed, mulched,
seeded and stabilised immediately. Vegetation is returning to this
area.
U Closed in February. Fertilised
and seeded by May 1993.
Showing signs of recovery.
U Nth
V Opened and closed within a 3
month time span.
Small area to the north-east was reworked for levy reinforcing.
Relatively stable with small pockets of vegetation returning.
W Similar to pit V using the same rehabilitation methods.
X Created in August for levy extensions. Topsoil was pushed over it
on completion of the levy.
Y
Z

204
Appendix 5. cont. Pit 1995
A Active – current salvage yard.
B Inactive, no change.
C Inactive, blown out sand dune area.
D Inactive, limited vegetation.
E Inactive, sand dune area.
F Nth Inactive, no change.
G Wst Seed trial commenced in June 1995. Vegetation is prolific.
G Sth Inactive, cover increasing. Germination from brushing of non-indigenous genus not expected to
survive.
G Nth Inactive, vegetation cover improving. Area still unstable.
H contoured and topsoiled
H Nth Inactive. Topsoil and vegetation from the R.O. plant has been stockpiled along the eastern edge of the
pit.
H Sth Active, no change.
I Active
J Wst Active, no change. No topsoil present.
J Est Inactive, relatively stable. Vegetation under stress due to poor rainfall.
K Active.
L 90% inactive, 10% active. Little change.
M 80% inactive, 20% active. Stable, vegetation maturing.
M Nth
M Sth
N Active.
O Inactive. Vegetation under stress due to low rainfall. Terrolas is starting to break up. Seed
germination trials have been unsuccessful.
O Sth
P Inactive. Brushing has been completed.
P-1
Q Inactive, there is little vegetation.
Q1
R Inactive. Stable with vegetation slowly increasing.
S 50% inactive, 50% active. Little change to vegetation in rehabilitated area.
T 90% inactive, 10% active. Topsoil pit. Vegetation in the area increasing.
U Inactive – Vegetation on the banks sparse. Little vegetation on the floor, the non indigenous plants
(Eucalyptus sp.) from past brushing are not expected to survive. Brushing has improved cover.
U Nth
V Inactive, vegetation improving.
W 70% inactive. Frankenia colonising well.
X Inactive. Juvenile vegetation evident.
Y Active. Topsoil stored.
Z Active. No topsoil stored
Z1 Inactive
Z2 Inactive. Steep gradient with cliff faces, poorly revegetated. Area to the South/East has been battered
and cross ripped. Vegetation is returning gradually.
Z3 Active.
Z4 Active.
Z5 Active.
Z6 Active.

205
Appendix 5. cont. Pit 1996
A Active.
B Inactive, no change. 5% vegetation cover.
C Inactive. 1% vegetation. Soil from recent crystalliser works used to help encourage vegetation.
D Inactive. 2% vegetation. Soil from recent crystalliser works used to help encourage vegetation.
E Inactive. 20% vegetation. Soil from recent crystalliser works used to help encourage vegetation. An
embankment was creased to help reduce wind channelling.
F Nth Delisted – now part of the crystallizer development.
G Sth Seed trial area. 45% vegetation
G Nth Inactive. 50% vegetation. Dune slopes still barren.
G Inactive. 30% vegetation.
H Nth
H Sth Inactive. 5% vegetation.
H Active.
I Inactive. 70% vegetation. In February the pit was battered, topsoil pushed over the area and the track
was cross ripped. Hand seeding took place in July.
J Wst Active, no change.
J Est Inactive. 25% vegetation.
K Active.
L Inactive (10% active). 35% vegetation.
M Nth
M Sth Inactive, unstable. 5% vegetation. Batters and tracks were cross ripped. Hand seeded in July.
N Inactive, stable. 50% vegetation.
O Active.
O Sth Inactive. 50% vegetation.
P
P-1 Inactive. 50% vegetation. Seeded in July.
Q
Q1 Inactive. 25% vegetation. Cross ripping in January on the southern side. Evidence of vegetation
present in the cross rips.
R Active.
S Inactive. 25% vegetation. Contours were moulded into the existing landscape in February. Seeding
was conducted with winter rains. Success rate was low due to the nature of the terrain.
T 25% inactive. 2% vegetation. The steep batter was moonscaped to slow erosion. In July the troughs
were hand seeded.
U 90% inactive. 60% vegetation.
U Nth Inactive. 60% vegetation. Cross ripped in February and seeded in July.
V
W Inactive. 50% vegetation.
X 70% inactive.
Y Inactive. 5% vegetation. Sand cliff faces were contoured to the existing topography. The access was
ripped and windrows were created. Hand seeded in July.
Z Active.
Z1 Active.
Z2 Inactive. 20% vegetation. Tidied up in February and seeded in July. Germination was disappointing.
Z3 Inactive. Moonscaped in February and hand seeded in July. Low but encouraging germination rates.
Z4 Active.
Z5 Active.
Z6 Active.
Z7 Active.

206
Appendix 5. cont. Pit 1997
A Active.
B Brushing covered 60%
C Inactive
D inactive
E
F Nth
G Sth Seed trial area, prolific vegetation within the exclusion fence, area outside is vegetating slowly
G Nth Area stabilising, encouraging vegetation diversity
G Increased stability, plant diversity encouraging
H Nth
H Sth
H Inactive.
I Active.
J Wst
J Est Active.
K relatively stable, colonisation occurring with good diversity
L Active
M Nth 90% inactive, 10% active.
M Sth
N Inactive - evidence of direct seeding encouraging
O Inactive, stable. Vegetation colonising
O Sth Active.
P Topdressed with topsoil immediately after disturbance.
P-1
Q Inactive, vegetation throughout pit area increasing. Infestation of E. platypus and Schinus sp. evident
from brushing will be culled
Q1 Pit construction started in 1997
R Inactive - vegetation diversity and plant coverage encouraging. E. platypus require culling
S Active
T Inactive - stable with vegetation cover returning
U 25% inactive, 75% active.
U Nth 90% inactive, 10% active.
V Inactive - Vegetation increasing. E. platypus seedlings will be culled.
W
X Inactive - vegetation improving, colonisation of Frankenia, Atriplex, Halosarcia and native grasses
evident despite sheep grazing.
Y 70% inactive, 30% active
Z Inactive. 5% vegetation.
Z1 Active.
Z2
Z3 Inactive.
Z4 Inactive.
Z5 Active.
Z6 Active.
Z7 Active.

207
Appendix 5. cont. 1998 1999 2000 2001
Pit active under rehab.
A 100%
B 100%
C 100%
D 100%
E 100%
F Nth 100%
G Sth 100%
G Nth 100% Topsoiled
G 100%
H Nth 100% 50% used for used for
foreshore expansion.
H Sth 100% Active.
H
I 100%
J Wst 100% Active.
J Est 100%
K Active.
L 10% 90%
M Nth 100%
M Sth 100%
N Active.
O 100
O Sth Vegetation returned to
sustainable level
P 100
P-1 100% Fenced with
polyethylene fencing
Fencing removed. Vegetation
at 85%
Q 100%
Q1 100%
R 100%
S 75% 25% Active.
T 10% 90% 30% reopened to
stockpile soil for
revegetation
U 100%
U Nth Topsoiled (previously pit
V?).
V 100%
W 30% 70%
X 100%
Y 100% Active. Now called
F south?
Z 100% Active.
Z1 100%
Z2 100%
Z3 100%
Z4 100%
Z5 100%
Z6 100%
Z7

208
Appendix 5. cont. Previously 2002 2003 2004
Delisted Under Under Under
Pit Areas (Ha) rehab. Active Total rehab. Active Total rehab. Active Total
A 2.9 0 0 0
B 1.1 1.1 1.1 1.1 1.1 1.1
C 2.9 2.9 2.9 2.9 2.9 2.9
D 2.4 2.4 2.4 2.4 2.4 2.4
E 5.3 5.3 5.3 5.3 5.3 5.3
F Nth 2 2 2 2 2 2
G Sth 2.9 2.9 2.9 2.9 2.9 2.9
G Nth 4.1 4.1 4.1 4.1 4.1 4.1
G
H Nth
H Sth
H 13.2 9.6 9.6 4.6 4.6 Delisted:
16.6
New: 12 0
I 1 0 0 0
J Wst 0.6 0.6 0.6 0.6 0.6 0.6
J Est 0.6 0.6 0.6 0.6 0.6 0.6
K 5 5 5 5 5 5
L 2.8 0.7 3.5 3.5 3.5 3.5 3.5
M Nth 0.6 0.6 0.6 0.6 0.6 0.6
M Sth 1.6 0 0 0
N 6.3 6.3 6.3 6.3 6.3 6.3
O
O Sth 3.5 0 0 0
P 1.5 1.5 1.5 1.5 1.5 1.5
P-1 12 8 20 12 8 20 12 8 20
Q 2.4 2.4 2.4 2.4 2.4 2.4
Q1 0.5 0.5 0.5 0.5 0.5 0.5
R 0.9 0.9 0.9 0.9 0.9 0.9
S 1 3.3 4.3 1 3.3 4.3 1 3.3 4.3
T 1.2 0.8 2 2 2 2 2
U 0.5 0.5 0.5 0.5 0.5 0.5
U Nth 5 5 5 5 5 5
V 0.9 0.9 0.9 0 0
W 1.1 1 1 1 1 1 1
X 0.7 0 0 0
Y 0.9 0.9 0.9 0.9 0.9 0.9
Z 1 1 1 1 1 1
Z1 0.3 0.3 0.3 0.3 0.3 0.3
Z2 1.1 1.1 1.1 1.1 1.1 1.1
Z3 0.8 0.8 0.8 0.8 0.8 0.8
Z4 1.2 1.2 1.2 1.2 1.2 1.2
Z5 1.9 1.9 1.9 1.9 1.9 1.9
Z6 1.1 1.1 1.1 1.1 1.1 1.1
Z7 0.7 0 0 0

209
Appendix 6. Species present in each of the undisturbed sites (US) P, Q, R and the
borrow pits (BP) P, Q, R, L.
US BP
Species P Q R P Q R L
Acacia ligulata + + + + + + +
Acacia tetragonophylla + + + +
Acanthocarpus preissii +
Alyogyne cuneiformis + + + +
Anthocercis sp. Shark Bay (Aplin 3335) +
Aphanopetalum clematideum + + +
Atriplex bunburyana + + + + + +
Atriplex cinerea? +
Beyeria sp. +
Carpobrotus sp. + +
Croton sp. +
Dianella revoluta + + + +
Dioscorea hastifolia + +
Diplolaena grandiflora
Diplopeltis intermedia var. intermedia
Diplopeltis sp. +
Enchylaena tomentosa + + + + +
Eremophila glabra (subsp. Albicans) +
Eremophila sp. +
Exocarpos sp. +
Frankenia pauciflora + + +
Gastrolobium sp. + +
Halgania littoralis + +
Halosarcia halocnemoides +
Indigofera sp. +
Lawrencia sp. +
Loxocarya sp. +
Maireana sp. + +
Melaleuca cardiophylla + + + +
Nicotiana occidentalis subsp. hesperius +
Olearia axillaris subsp. obovata + +
Opercularia sp. + +
Pembertonia latisquamea + + + + +
Pimelea microcephala subsp. microcephala + + + +
Rhagodia latifolia subsp. latifolia + + + + + +
Rhagodia preissii subsp. obovata +
Salsola kali + + + +
Scaevola spinescens +
Scaevola tomentosa + + + + +
Scaevola sp. +
Senna glutinosa subsp. chatelainiana +
Solanum hesperium
Solanum orbiculatum + + +
Spyridium sp. + +
Stylobasium spathulatum + + +
Threlkeldia diffusa + + + + +
Thryptomene baeckeacea + + +
Triodia plurinervata + + + +
Triodia longipleura? +
Zygophyllum fruticulosum + + + + +

210
Appendix 7. Dominant species that recruit from topsoil (present in every quadrat in pit
L), recruit from natural migration (>5 plants in pit P, Q or R), or are only found in the
undisturbed vegetation.
Topsoil Natural migration Undisturbed vegetation
Acacia ligulata Acacia ligulata Beyeria sp.
Alyogyne cuneiformis Acacia tetragonophylla Diplopeltis sp.
Atriplex bunburyana Alyogyne cuneiformis Eremophila glabra small (subsp.
Enchylaena tomentosa Anthocercis sp. Shark Bay Albicans)
Frankenia pauciflora Atriplex bunburyana Eremophila sp.
Lawrencia sp. Carpobrotus sp. Gastrolobium sp.
Mariana sp. Enchylaena tomentosa Halgania littoralis
Nicotiana occidentalis Frankenia pauciflora Indigofera sp.
subsp. hesperius Halosarcia halocnemoides Loxocarya sp.
Stylobasium spathulatum Olearia axillaris subsp. Scaevola sp.
obovata Scaevola spinescens
Pimelea microcephala subsp.
microcephala
Senna glutinosa subsp.
chatelainiana
Rhagodia latifolia subsp. Spyridium sp.
latifolia Triodia longipleura?
Scaevola tomentosa
Stylobasium spathulatum
Threlkeldia diffusa
Zygophyllum fruticulosum
Appendix 8. Species of widespread distribution (occur in at least 5 sites) and limited
distribution (occur only in 1 site).
Widespread distribution Limited distribution
Acacia ligulata Acanthocarpus preissii
Atriplex bunburyana Anthocercis sp. Shark Bay
Rhagodia latifolia subsp. latifolia Beyeria sp.
Pembertonia latisquamea Croton sp
Enchylaena tomentosa Diplopeltis sp.
Scaevola tomentosa Eremophila glabra small (subsp. Albicans)
Threlkeldia diffusa Eremophila 'big white flower'
Zygophyllum fruticulosum Exocarpos
Halosarcia halocnemoides
Indigofera sp.
Lawrencia sp.
Loxocarya sp.
Nicotiana occidentalis subsp. hesperius
Rhagodia preissii subsp. obovata
Scaevola sp
Scaevola spinescens
Senna glutinosa subsp. chatelainiana
Triodia longipleura?

211
Appendix 9. Germination of Dioscorea hastifolia treated with water (control),
Gibberellic acid (GA), smoke water (SW) or karrikinolide (KAR) after 49 days
incubation at 12/12h 26/13oC (white bars) or 12/12h 33/18
oC (grey bars) after a) 4
months or b) 16 months storage at ambient laboratory temperatures.
b)
Control GA SW KAR
0
20
40
60
80
10026/13
33/18
a)
Control GA SW KAR
Germ
ina
tion
(%
)
0
20
40
60
80
100
26/13
33/18
Appendix 10. Germination of Zygophyllum fruticulosum treated with water (control),
gibberellic acid (GA), smoke water (SW) or karrikinolide (KAR) after 49 days
incubation at 12/12h 26/13oC (white bars) or 12/12h 33/18
oC (grey bars) after a) 3
months or b) 15 months storage at ambient laboratory temperatures.
b)
Control GA SW KAR
0
20
40
60
80
10026/13
33/18
a)
Control GA SW KAR
Ge
rmin
atio
n (
%)
0
20
40
60
80
10026/13
33/18

212
Appendix 11. Time to 50% germination of Atriplex bunburyana seeds soaked for 0 ,
12, 18 or 24 hours in water (control), GA or KAR, dried, then incubated on water agar
at 20oC.
Priming time (h)
0 12 18 24
Tim
e t
o 5
0%
ge
rmin
atio
n (
days)
0
1
2
3control
GA
KAR
Appendix 12. Cumulative germination of Solanum orbiculatum not primed and treated
with karrikinolide (KAR) prior to incubation (not primed), primed in water for 1 day
and treated with KAR prior to incubation (water 1), primed in KAR for 1 day (KAR 1)
and primed in water for 2 days then butenolide for 1 day (water 2 KAR 1). Seeds were
incubated on water agar at 20oC.
Time (days)
0 2 4 6 8 10 12 14
Cu
mu
lative
ge
rmin
atio
n (
%)
0
20
40
60
80
100
not primed
water 1
KAR 1
water 2 KAR 1

213
Appendix 13. Equations developed from volumetric soil moisture calibration.
Volumetric and gravimetric soil moisture was determined on seven samples of each soil
(no rip, rip and natural vegetation areas at pits P, Q and R). The values were plotted
against each other, with volumetric (in milli volts) on the x-axis and gravimetric on the
y-axis. A trendline was determined, and the resulting equation was used to convert
volumetric soil moisture readings obtained in the field to gravimetric soil moisture.
Pit Site Equation R2 value
P no rip y = 0.0289x – 0.2569 0.9375
P rip y = 0.0339x – 1.4972 0.9379
P natural vegetation y = 2E-07x3 – 0.0002x
2 + 0.1266x – 4.5688 0.9762
Q no rip y = 1E-07x3 – 0.0001x
2 +0.0461x – 2.7454 0.7952
Q ripa
y = 1E-07x3 – 0.0001x
2 +0.0461x – 2.7454 0.7952
Q natural vegetation y = 5E-08x3 – 9E-05x
2 + 0.075x – 1.7212 0.9885
R no rip y = 1E-07x3 – 0.0002x
2 + 0.1129 – 6.7878 0.9865
R rip y = 10.491Ln(x) – 44.675 0.9587
R natural vegetation y = 2E-07x3 – 0.0002x
2 + 0.1217x – 5.2904 0.9705
aThe equation developed for pit Q, no rip, was used for pit Q, rip, due to the similarity
in readings, and insufficient data points in the relevant part of the curve for pit Q, rip.

214
Appendix 14. Cost-benefit analysis
Results tend to suggest that the plant source may be somewhat species specific, and this
can be illustrated by the two focus species tested at SBS in 2006, Acacia
tetragonophylla and Atriplex bunburyana. For example, while Acacia tetragonophylla
had higher emergence (4-fold under ripping to 9-fold under no ripping) than Atriplex
bunburyana following seed broadcasting, Atriplex bunburyana greenstock had higher
establishment (5-fold under ripping to >43 fold under no ripping) than Acacia
tetragonophylla. However, following seed broadcasting, the 4-fold higher level in
seedling emergence for Acacia tetragonophylla in the (preferred) ripped profile was
offset by a 4-fold higher price in seed, which translated into no difference in the price
between species for a seedling unit (ca. 23-24 cents) (Table 1). Following greenstock
planting under the same conditions, the 5-fold lower establishment of Acacia
tetragonophylla seedlings (at 18.7%) compared to Atriplex bunburyana, translated into
a seedling unit price of $6 while the Atriplex bunburyana was only $1.21 given the 91%
establishment (Table 2).
Obviously, the broadcasting of seeds is a significantly cheaper option, but if seed
supplies are limited, and when considering the low emergence (0.55%) of Atriplex
bunburyana in the (preferred) rip profile following seed broadcasting (Table 1) -
possibly as a result of seeds being lost to wind erosion - it may be wise to consider
spending $1.21 for a seedling unit and planting greenstock of Atriplex bunburyana.
This will ensure the conservation of seeds, given that there is a 91% chance that a seed,
that has been sown to produce a greenstock unit, will develop into an established
seedling. Conversely, for every 200 seeds of Atriplex sown as broadcast seed, only one
will develop into a seedling.
For Acacia tetragonophylla, the significantly higher priced seedling unit following
greenstock planting ($6.07), together with the higher seedling emergence following
broadcasting seeding operations, when compared to Atriplex bunburyana, may warrant
the broadcasting of Acacia tetragonophylla seeds instead.
The major limitation of this study is that it was only performed over two consecutive
years, whilst the aforementioned conclusions are based on broadcast seeding and
greenstock planting in only one year, 2006 (which was considered a dry year). Due to

215
rainfall variables at the site, and given the dry 2006 year, a longer term study would
assist the development of more solid conclusions.
Table 1. Cost of seedlings from broadcast seeding in 2006. Price per gram was
ascertained from the price list of a local seed merchant (Kimseed). Emergence
percentages are based on the treatment averaged across all three pits. Emergence was
recorded in spring 2006.
price per
gram
($)
100
seed wt
(g)
price per
100 seeds
($)
emergence
(%)
price per
seedling
($)
a b a x b (c) d c / d
Acacia tetragonophylla rake only 0.36 1.42 0.51 0.22 2.34
Atriplex bunburyana rake only 0.25 0.50 0.125 0.11 1.14
Acacia tetragonophylla rip+rake 0.36 1.42 0.51 2.11 0.24
Atriplex bunburyana rip+rake 0.25 0.50 0.125 0.55 0.23
Table 2. Cost of seedlings from greenstock planting in 2006. Establishment (recorded
in spring 2006) percentages are based on the control treatment, averaged across all pits.
Establishment of Acacia tetragonophylla seedlings in the no rip area was actually 0%,
but changed to 1% for the purposes of comparison.
price per
gram
($)
100
seed wt
(g)
price per
200 seeds
($)
price per
seedling
($)
price per 100
seedlings
($)
establishment
(%)
price per
seedling
($)
a b
a x b x 2
(c) d
c + (d x 100)
(e) f e / f
Acacia
tetragonophylla
no
rip 0.36 1.43 1.03 1.10 111.02 1.0 111.03
Atriplex
bunburyana
no
rip 0.25 0.50 0.25 1.10 110.25 43.3 2.55
Acacia
tetragonophylla
rip
0.36 1.43 1.03 1.10 111.02 18.3 6.07
Atriplex
bunburyana
rip
0.25 0.50 0.25 1.10 110.25 91.1 1.21

216
Appendix 15. Management recommendations
The topics covered in this study have been integrated into a working rehabilitation plan
for SBS (Fig. 1). The rehabilitation plan is an example of an adaptive management
approach to rehabilitation whereby the recipients of the rehabilitation plan acknowledge
that they are in a continual phase of learning, improvement and reassessment of their
rehabilitation operations. It is suggested that SBS approach their rehabilitation
operations as a series of experiments and allow rehabilitation management decisions to
be guided by research-based ecological rehabilitation. By adopting an adaptive
management approach, SBS has the potential to develop benchmark standards –
completion criteria – to guide their rehabilitation efforts and to ultimately assist the de-
listing process of completed rehabilitation borrow-pits. I feel that it is particularly
important for SBS to establish some completion criteria in order to have a goal for their
rehabilitation, and also as a means of delisting borrow pits they consider to be
rehabilitated. Currently there are no completion criteria for borrow pits at SBS. At
present, borrow pits are delisted when they are ‗considered to be sufficiently
rehabilitated‘(Shark Bay Salt Joint Venture 2004). However, there is no means of
quantifying when the borrow pits have been rehabilitated to an acceptable standard. In
order to compile completion criteria, it is first necessary to set out a rehabilitation
objective. For example, the rehabilitation objective of a mine in the jarrah forest of
Western Australia is to ‗return a self-sustaining jarrah forest ecosystem that fulfils all of
the pre-mining land uses‘ (Gardner 2001). This objective has lead to the development
of completion criteria including the establishment of an overstorey and an understorey.
These criteria are quantified, for example, minimum and maximum eucalypt stems per
hectare are specified, the minimum density of legumes, and the minimum species
richness (DoIR (Department of Industry and Resources) 2006). Hence, completion
criteria of borrow pits at SBS could include a minimum density of dominant species
(such as Acacia ligulata, Melaleuca cardiophylla, Thryptomene baeckeacea and Triodia
plurinervata), and minimum species richness. In addition to restoring flora (and fauna)
diversity, it is also important to restore vegetation structure and ecosystem functions
(Koch and Hobbs 2007). If a diversity of species is returned to the area, then the
resulting vegetation structure should be similar to the pre-mined structure. Ecosystem
functions include litter decomposition and nutrient cycling (Grant et al. 2007). An
alternative method of measuring restoration success is landscape function analysis,
which is commonly used in arid Australia (Koch and Hobbs 2007). So, in summary,

217
SBS should have a goal for borrow pit rehabilitation, and a method of assessing whether
they have been successful in attaining this goal.
Figure 1. Rehabilitation plan for borrow pits at SBS. Coloured boxes indicate areas
covered by this thesis. White boxes indicate areas that need to be implemented by SBS.
Develop
completion
criteria
Produce
greenstock
Compare
revegetation
methods
(broadcasting
vs
greenstock)
Plant
greenstock
(when soil is
moist)
Compare
with
completion
criteria
Monitor
(annually)
Classify as
successfully
rehabilitated
and delist
Collect
cuttings of
species with
unresolved
seed
dormancyDevelop seed mix based on
required density in rehabilitation
and average emergence
Determine which species do not
return via broadcast seeding
therefore must be planted, and
determine planting density based
on required density and average
survival
Weed
management
Spread
topsoil
(if available)
Compare
annual
variation
Assess soil
properties
Monitor
(5 yearly)
Determine
which
species can
grow in
which pit
Collect seed
(Sept – Dec)
Floristic
benchmarks
for natural
vegetation
Pre-treat
seed
Broadcast
seed (prior
to winter
rain)
Revegetated
borrow pit
Rip soil
(between
summer and
winter rain)
Erosion
control on pit
slopes
Resolve
seed
germination
and
dormancy
issues
StartChapters 2, 3, 4
Chapter 5
Chapter 6
Chapter 7
Chapter 8
Chapters 6, 7
Optimise
seed
broadcasting
technology

218
Whilst the rehabilitation process is only in its infancy, this study has provided a body of
benchmark information in an attempt to assist the initial phase of rehabilitation at SBS.
The recommendations for rehabilitation and further research that follow are based on
this body of benchmark information.
Part I. Seed biology
Finding/s: Seeds of many species (Aphanopetalum clematideum, Atriplex bunburyana,
Austrostipa elegantissima, Melaleuca cardiophylla, Nitraria billardierei,
Pembertonia latisquamea, Rhagodia baccata) had improved germination at the
lower incubation temperature, indicating that they may germinate more readily in
winter.
Recommendation/s: Seeds should be broadcast prior to winter rain, not summer rain.
When sowing seeds in the summer months for greenstock production, cool
incubation conditions are recommended.
Finding/s: Seeds of some species (Acacia tetragonophylla, Anthocercis littorea, Ptilotus
exaltatus, Solanum orbiculatum, Stylobasium spathulatum) required a pre-
treatment to break dormancy or promote germination.
Recommendation/s: Seeds will need to be pre-treated with hot water to break physical
dormancy; or smoke water, karrikinolide or gibberellic acid to promote
germination prior to broadcast seeding or greenstock production.
Finding/s: Solanum species displayed a range of germination needs: they were either
non-dormant, required smoke water or karrikinolide for germination, or were
physiologically dormant. They were not physically dormant.
Recommendation/s: Solanum orbiculatum seeds will need to be pre-treated with smoke
water or karrikinolide to promote germination prior to broadcast seeding or
greenstock production. Given the responsiveness of the seeds to a wide range of
temperatures, they may be able to germinate with summer rain, and therefore can
be broadcast to capitalise on summer rains. When sowing seeds for greenstock
production, cool incubation conditions are not required.

219
Finding/s: Dormancy of 50% of the physiological dormant species investigated
(Anthocercis littorea, Dioscorea hastifolia, Zygophyllum fruticulosum) was
overcome by after-ripening.
Recommendation/s: Seeds will require after-ripening at 45oC and 10% RH for 3 months.
After-ripening at 30oC and 50% RH for 3 months is also acceptable for two of the
species.
Finding/s: Dormancy of the remaining 50% of the physiological dormant species
investigated (Acanthocarpus preissii, Eremophila oldfieldii, Thryptomene
baeckeacea) was not overcome by after-ripening.
Recommendation/s: Further research is required to optimise germination of two of these
species, given that one species is responsive to warm stratification.
Finding/s: Karrikinolide or gibberellic acid was required to promote germination of
Anthocercis littorea.
Recommendation/s: Seeds will require pre-treatment with Karrikinolide or gibberellic
acid.
Part II. Rehabilitation
Finding/s: Borrow pits had lower plant density and unrelated species composition
compared to their undisturbed counterparts.
Recommendation/s: All other borrow pits at SBS (undergoing rehabilitation and those
previously delisted) should be surveyed to quantify plant and species abundance
and species richness, and to determine if they are in fact similar to the undisturbed
vegetation, or whether remedial action will need to be undertaken in those areas
also. Revegetation requires human intervention through seed broadcasting or
greenstock planting, and if topsoil is employed, habitat matching to reinstate the
appropriate plant density and species richness.
Finding/s: A non-local eucalypt species, which established from seeds contained in re-
spread canopy mulch, was identified in Pit P (Fig. 2).
Recommendation: Non-local plants should be removed and mulch, which has been
derived from vegetation that supports non-local species, should be avoided in
rehabilitation efforts.

220
Figure 2. Non-local Eucalyptus in pit P (April 2008)
Finding/s: Topsoil borrowing in a low rainfall year resulted in similar seedling
recruitment in recipient and donor sites, however, few seedlings survived the first
summer in both sites.
Recommendation/s: Topsoil borrowing requires significant further investigation in an
average rainfall year, with the investigation needing to incorporate an undisturbed
reference site to ensure that the topsoil donor site has the potential to return to
pre-disturbance plant abundance, richness and cover levels.
Finding/s: Seed broadcasting was more effective when implemented in autumn (≤ 27%
emergence) compared with winter (<4% emergence).
Recommendation/s: Seeds should be broadcast prior to the onset of winter rain.
Finding/s: Seedling emergence following seed broadcasting was 14-fold higher in
ripped and raked soil.
Recommendation/s: Soil ripping, followed by raking seeds into the soil (with a scarifier)
to ensure seed burial and stabilisation, should become standard rehabilitation
practice.

221
Finding/s: High seed losses in pits P and Q, following seed broadcasting, occurred as a
result of water erosion (Fig. 3).
Recommendation/s: Research is required to determine options for decreasing water
erosion. Some options to investigate include decreasing the slope angle of pit
walls, ripping along the contour of pit walls, jute-matting the soil surface and
greenstock planting.
Figure 3. Erosion on the slopes of pit Q (June 2005)
Finding/s: Rehabilitation areas that were subject to soil ripping were characterised by
improved soil penetrability, and seedling establishment from greenstock (Fig. 4).
Recommendation/s: Soil ripping in borrow pits should become standard rehabilitation
practice.
Finding/s: Differences in plant survival between the borrow-pits and species was highly
evident and may be related to the soil nutrient and salinity status.
Recommendation/s: Borrow-pit soils should be analysed prior to planting and/or
seeding to determine their potential to support specific species. For example,
highly saline pits (such as Q) may need to be rehabilitated with species from

222
analogous sites (e.g. birridas) rather than adjacent vegetation. Therefore, ex-situ
trials to determine soil effects on dominant species should be implemented.
Finding/s: The use of air-pruned pots, nutrient loading and plant pruning did not benefit
plant survival.
Recommendation/s: The use of air-pruned pots and pruning seedlings are not
recommended. Nutrient loading should be more rigorously investigated to
determine its potential.
Finding/s: Fertiliser improved growth, but not survival of Atriplex bunburyana
seedlings.
Recommendation/s: Further investigation is required into the effectiveness of fertiliser
in higher rainfall years.
Finding/s: Whilst seed broadcasting was more cost-effective then greenstock planting,
establishment of seedlings derived from a seed unit from broadcast seeding was
lower than those derived from greenstock planting, with level of success
dependent on species and rainfall.
Recommendation/s: Further research into the two plant sources is highly recommended
and should involve a cost-benefit analysis, a greater suite of species, seasonal
variation, and ―resource islands‖. In the interim, both broadcast seeding and
greenstock planting should be undertaken to ‗bet hedge‘ in the event of low
rainfall.

223
a)
b)
Figure 4. Survival in the a) ripped and b) unripped areas of pit Q.