seasonal parasitism and biological characteristics of habrobracon hebetor (hymenoptera: braconidae)...
TRANSCRIPT

This article was downloaded by: [Kungliga Tekniska Hogskola]On: 10 October 2014, At: 10:13Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Biocontrol Science and TechnologyPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/cbst20
Seasonal parasitism and biologicalcharacteristics of Habrobraconhebetor (Hymenoptera: Braconidae)– a potential larval ectoparasitoid ofHelicoverpa armigera (Lepidoptera:Noctuidae) in a chickpea ecosystemHem Saxena a , Duraimurugan Ponnusamy a & Mir Asif Iquebal ba Division of Crop Protection , Indian Institute of Pulses Research ,Kanpur , Indiab Division of Biometrics and Statistical Modelling , IndianAgricultural Statistics Research Institute , New Delhi , IndiaAccepted author version posted online: 16 Jan 2012.Publishedonline: 13 Mar 2012.
To cite this article: Hem Saxena , Duraimurugan Ponnusamy & Mir Asif Iquebal (2012) Seasonalparasitism and biological characteristics of Habrobracon hebetor (Hymenoptera: Braconidae) – apotential larval ectoparasitoid of Helicoverpa armigera (Lepidoptera: Noctuidae) in a chickpeaecosystem, Biocontrol Science and Technology, 22:3, 305-318, DOI: 10.1080/09583157.2012.656579
To link to this article: http://dx.doi.org/10.1080/09583157.2012.656579
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoeveror howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.

This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

RESEARCH ARTICLE
Seasonal parasitism and biological characteristics of Habrobraconhebetor (Hymenoptera: Braconidae) � a potential larval ectoparasitoid
of Helicoverpa armigera (Lepidoptera: Noctuidae) in a chickpeaecosystem
Hem Saxenaa*, Duraimurugan Ponnusamya and Mir Asif Iquebalb
aDivision of Crop Protection, Indian Institute of Pulses Research, Kanpur, India; bDivision ofBiometrics and Statistical Modelling, Indian Agricultural Statistics Research Institute, New
Delhi, India
(Received 28 September 2011; final version received 6 January 2012)
Seasonal parasitism of Habrobracon hebetor (Say) on Helicoverpa armigera(Hubner) in chickpea was studied for three consecutive years. Parasitism byH. hebetor on larvae of H. armigera reached 12.3%. The parasitoid maintainedreproductive activity on H. armigera from February to April coinciding with podformation and maturation stages of the crop. In laboratory assays, we investigatedthe suitability of larval instars of H. armigera to the parasitoid H. hebetor. Thisparasitoid attacked third to sixth instars, though fourth and fifth instar larvaewere found most suitable with 100% parasitism and development to adults.Parasitoid developmental time was longest in fifth instar (9.1 days) compared toother instars (8.1�8.9 days). Fifth instar larvae resulted in highest numbers ofcocoons and adult emergence. In addition, suitability of seven lepidopteranspecies to H. hebetor was investigated. Corcyra cephalonica, Galleria mellonellaand H. armigera were the most suitable hosts with 100% parasitism anddevelopment to adults. It was followed by Maruca vitrata and Autographanigrisigna with 60�76.7% and 40�70% parasitism and parasitoid developmentalsuccess, respectively. Though there was 23.3% parasitism, there was no parasitoiddevelopment in Spodoptera litura. No parasitism was recorded in Spilarctiaobliqua. Development of H. hebetor was most rapid in C. cephalonica (8.7 days),and longest in G. mellonella (9.3 days). Parasitoids that developed on these hostsresulted in highest numbers of cocoons and adult emergence. The parasitoid couldbe exploited for the biological control of H. armigera in a chickpea ecosystem.
Keywords: Habrobracon hebetor; Helicoverpa armigera; chickpea; host instar;host species; biological control
Introduction
Chickpea (Cicer arietinum L.) is the third most important grain legume in the world
after dry beans (Phaseolus spp.) and field pea (Pisum sativum L.). It is an important
source of protein, minerals, fibre and vitamins in the diets of millions of people in
Asia and Africa. Chickpea is cultivated on about 11 m ha adding 8.8 m tonnes to the
global food basket (Rao, Birthal, Bhagavatula, and Bantilan 2010). There are many
factors which influence the production of chickpea among which are insect pests,
*Corresponding author. Email: [email protected]
Biocontrol Science and Technology,
Vol. 22, No. 3, March 2012, 305�318
ISSN 0958-3157 print/ISSN 1360-0478 online
# 2012 Taylor & Francis
http://dx.doi.org/10.1080/09583157.2012.656579
http://www.tandfonline.com
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

particularly the gram pod borer Helicoverpa armigera (Hubner) (Lepidoptera:
Noctuidae) (Rao et al. 2010).
Among the natural enemies of H. armigera recorded in chickpea ecosystem,
Campoletis chlorideae Uchida (Hymenoptera: Ichneumonidae) is an important larval
endoparasitoid that causes up to 50% parasitisation under natural conditions (Nema
2010). However, activity of the parasitoid occurs only during November to February,
coinciding with the vegetative stage of the crop. In North India, during March to
April, H. armigera damage coincides with the fruiting stage of chickpea causing
heavy losses (Sachan and Katti 1994). In a survey conducted to explore a potential
parasitoid coinciding with the fruiting stage of chickpea, we recorded a gregarious
larval ectoparasitoid Habrobracon (Bracon) hebetor (Say) (Hymenoptera: Braconi-
dae) on H. armigera in chickpea fields in Kanpur district, Uttar Pradesh (Saxena and
Duraimurugan 2009). Therefore, the present study was aimed at understanding the
seasonal parasitism of H. hebetor on H. armigera in chickpea ecosystems. We also
examined the suitability of H. armigera and six other lepidopteran species as hosts to
H. hebetor. Such information is essential for mass culture and utilisation of
H. hebetor as a major biological control agent of H. armigera.
Materials and methods
Test insects
Laboratory culture of H. armigera was established from larvae collected in chickpea
fields at the New Research Farm, Indian Institute of Pulses Research (IIPR), Kanpur
during November and December 2008. The larvae were maintained on a chickpea-
based semi-synthetic diet (Armes, Bond, and Cooter 1992) and reared in groups until
the early third instar. Henceforth, they were reared individually in plastic containers
(6 cm height�4 cm diameter). Air circulation in the container was ensured through
a hole (diameter 1 cm) in the lid, covered with nylon mesh. The container and diet
were replaced on alternate days and larvae were continuously reared until they
pupated. Four days after pupation, pupae were washed in running tap water, surface
sterilised by immersing in 0.5% sodium hypochlorite solution, followed by rinsing in
water and then placed on filter paper to dry. The pupae were transferred to a
40�40�40 cm emergence cage. Newly emerged adults were sexed and five pairs of
adults were transferred to plastic buckets (7 L capacity) maintaining the sex ratio of
1:1 for mating and oviposition. Adults were fed with 10% sugar solution enriched
with ABDEC vitamin solution. The plastic bucket was covered with sterile white
muslin cloth, which served as an oviposition substrate. Daily, the cages were checked
and the muslin cloth on which the eggs were laid was removed and replaced by a
fresh, sterile one. The sugar solution in the glass vials was also refilled daily. The
cages were kept in a culture room maintained at 90% RH, 278C with a photoperiod
of 14:10 (L:D). Newly hatched larvae from the eggs laid on muslin cloth were
transferred to plastic trays (30�15�5 cm) containing a 3 mm layer of semi-
synthetic diet. The tray was kept in an inverted position in dark after closing the lid
tightly. The larvae were allowed to grow until the early third instar stage followed by
rearing individually in plastic containers, as described earlier.
Field collected larvae of the tobacco caterpillar, Spodoptera litura (Fabricius)
[Lepidoptera: Noctuidae]; the Bihar hairy caterpillar, Spilarctia obliqua (Walker)
306 H. Saxena et al.
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

[Lepidoptera: Arctiidae] and legume pod borer, Maruca vitrata (Geyer) [Lepidop-
tera: Pyralidae] (collected from mungbean during October 2008), the semilooper,
Autographa nigrisigna (Walker) [Lepidoptera: Noctuidae] (collected from chickpea
during December 2008) were continuously reared on their natural hosts. Thelaboratory hosts rice moth, Corcyra cephalonica (Stainton) [Lepidoptera: Pyralidae]
and greater wax moth, Galleria mellonella (Linnaeus) [Lepidoptera: Galleridae] were
obtained from the Department of Entomology, Narendra Dev University of
Agriculture and Technology, Faizabad were reared on respective natural hosts
broken maize and wax comb.
Parasitoid
A laboratory colony of H. hebetor was established from parasitised H. armigera
larvae collected from New Research Farm (IIPR, Kanpur) during March 2009.
H. hebetor was reared in the laboratory at 25928C, 6595% RH, and a 14:10 (L:D)
photoperiod using larvae of H. armigera as hosts. H. armigera larvae wereindividually placed in a plastic container (30�15 cm) and a pair of adult parasitoids
was released. A cotton swab soaked in 50% honey solution was kept in the container
for H. hebetor adults. The mouth of the container was covered with black muslin
cloth and fastened with rubber band. After 48 h, the parasitoid was removed and the
parasitised larvae were held until adult parasitoids emerged. These were then used
for the experiments.
Seasonal parasitism of H. hebetor on H. armigera in chickpea
Field studies were carried out on chickpea at New Research Farm (26827?N, 80814?E;
152.4 m), IIPR, Kanpur during post-rainy season over a period of three years (2007�2010). The chickpea seeds were sown in sandy loam soil with pH 8.16, EC 0.21 dS/m,organic carbon 0.24%, available P 11.88 kg/ha and available K 126 kg/ha. In all the
three years, the kabuli chickpea variety ‘JKG 1’ was sown during last week of
November with a spacing of 30�10 cm and plot size of 0.20 ha. A fertiliser of 20 kg
N and 50 kg P2O was applied. Recommended crop management practices were
adopted except that no insecticide was sprayed. To record the natural parasitism and
seasonal activity of parasitoids, the larval population of H. armigera was monitored
at weekly intervals beginning 15�21 days after the date of sowing and continued until
crop maturity. On each sampling date, observations were taken from 100 randomlyselected plants at four places (25 plants/place) selected diagonally across the field.
The plants were checked for the presence of H. armigera larvae and when collected,
were kept individually in a plastic container. The field-collected larvae were brought
back to the laboratory and reared on a chickpea based semi-synthetic diet (Armes,
Bond, and Cooter 1992) under laboratory conditions [25928C, 6595% RH, and a
14:10 (L:D) photoperiod] until the parasitoids emerged, or until H. armigera adult
emergence (Figures 1 and 2). Parasitoids were identified by Prof. M. Hayat, Emeritus
Scientist, Department of Zoology, Aligarh Muslim University, Aligarh, UttarPradesh, India.
A correlation coefficient was estimated to determine the relationship between
weather parameters and per cent parasitism (Schliserman 2001). For the analysis, the
mean maximum temperature, minimum temperature, relative humidity, wind speed,
Biocontrol Science and Technology 307
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

Figure 1. Field collected healthy and parasitised larvae of Helicoverpa armigera in chickpea ecosystem.
30
8H
.S
ax
ena
eta
l.
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

Figure 2. Different developmental stages of Habrobracon hebetor on Helicoverpa armigera.
Bio
con
trol
Scien
cea
nd
Tech
no
log
y3
09
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

sunshine hours and rainfall during the sampling week and the corresponding per
cent parasitism were used.
Host instar suitability for the parasitoid
A no-choice test was conducted to determine the suitability of different larval instars
of H. armigera to H. hebetor under laboratory conditions. For studying the host
instar acceptance (paralysation, parasitisation and parasitioid developmental
success) by the parasitoid, newly-moulted larvae of each instar (first to sixth instar
larva) of H. armigera were placed individually in a plastic container (10�5 cm)
along with the diet. Larval instars were determined by checking the shed headcapsules. A single, four-day old, mated H. hebetor female, without prior access to
hosts, was released into each container along with diet. Forty-eight hours after their
introduction, the parasitoids were removed from the plastic container and the per
cent of the host instar larvae paralysed was assessed. For assessing parasitism, host
instar larvae were examined, and the presence of parasitoid eggs or larvae
determined. Parasitised host larvae were monitored daily and proportion of
parasitised hosts that successfully yielded live wasps (parasitoid developmental
success) was determined. For studying the host instar suitability for the parasitoid,
newly moulted third, fourth, fifth and sixth instars of H. armigera (host instars that
did not yield a live parasitoid in the acceptance test were discarded) were individually
kept in a plastic container together with artificial diet. Individual four-day old mated
female parasitoids, without prior access to hosts, were released into each plastic
container as described earlier. After 48 h, the parasitoid was removed and parasitised
larvae were allowed to develop. The parasitised larvae were checked at 12 h intervals
and the developmental time (egg to adult emergence), number of cocoons formed,
number of adult parasitoids emerged, sex ratio and longevity of adult parasitoids
were recorded. The emerged parasitoids were transferred to test tubes (5�1.5 cm)
containing 50% honey solution and adult longevity was calculated by observing daily
mortality. In all the experiments, 30 larvae for each host-instar (three replicates each
of 10 larvae) were used in completely randomised design.
Suitability of parasitoid on different hosts
Seven lepidopteran species (rice moth, Corcyra cephalonica; greater wax moth,
Galleria mellonella; tobacco caterpillar, Spodoptera litura; semilooper, Autographa
nigrisigna; legume pod borer, Maruca vitrata and Bihar hairy caterpillar, Spilarctia
obliqua; and gram pod borer, Helicoverpa armigera) were tested for acceptance and
suitability by the parasitoid in a no-choice test using plastic container (10�5 cm)
with a single host larva under laboratory conditions. A laboratory colony of
H. hebetor established from parasitised H. armigera larvae and cultured on
H. armigera as described earlier was used for the experiment. Newly moulted fifth
instar larvae of each host were individually placed in a plastic container along with
their natural hosts and a 4-day old mated female parasitoid was released as described
earlier. After 48 h, the female adult parasitoid was removed and the parasitised larva
was held in the container and allowed to develop. Observations on hosts paralysed,
parasitism and parasitoid developmental success were similar to the method
310 H. Saxena et al.
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

described earlier for studying the influence of different larval instars of H. armigera
on the parasitoid.
Statistical analysis
The association between weather parameters and per cent parasitism were computed
using Pearson’s correlation coefficient using SPSS software. An analysis of variance
(ANOVA) was conducted on all data related to suitability of the parasitoid on host
instars of H. armigera and on different host species using SAS statistical softwareversion 9.2. Following ANOVA, differences between data-sets were determined using
Tukey-Kramer post hoc tests and the accepted level of significance was P50.05 in
all instances. Data are presented as means9standard error (SE), where means within
the same column and followed by different letters are significantly different
(P50.05; Tukey’s test).
Results
Seasonal parasitism of Habrobracon hebetor on Helicoverpa armigera in chickpea
The data on seasonal parasitism of H. armigera by H. hebetor in chickpea during the
post-rainy season of three consecutive years (2007�2010) are presented in Figure 3.
The occurrence and percentage parasitism varied between years. In all three years,
the parasitoid remained almost nil up to the third week of February and itsparasitism was observed only in the last week of February to third week of April,
coinciding with pod formation and maturation stage of the crop. Mean parasitism by
H. hebetor on larvae of H. armigera during all three years ranged from 0.9 (2010) to
12.3% (2009). Higher parasitism of 2.0�12.3% occurred between the last week of
February to second week of April. Adult emergence per parasitised larva of
H. armigera during these three years varied from 7.0 to 17.3 and the progeny sex
ratio varied across the standard meteorological weeks and years (Figure 4). Parasitic
activity did not show any significant correlation with abiotic factors.
Host instar suitability
Habrobracon hebetor paralysed all the developmental stages of H. armigera presented
under no-choice tests. However, significant differences were observed in theparasitism and parasitoid developmental success (egg to adult) by H. hebetor in
different larval instars of H. armigera (Tukey’s test, P50.05). In both cases, fourth
and fifth instar larvae were most suitable, representing 100% parasitism and
parasitoid development. Sixth instar larvae were the next most suitable for
parasitism (96.7%) and parasitoid development (95.0%) followed by third instars
(68.3% parasitism and 56.7% parasitoid development). There was no parasitism and
parasitoid development in first and second instars, though the parasitoid paralysed
these instars (Figure 5).Different instars of H. armigera greatly affected the biology and development of
the parasitoid (Table 1). When parasitism took place on fifth instars, parasitoid
developmental time (egg to adult emergence) was significantly longer (9.1 days) than
on larvae from other instars (8.1�8.9 days). Similarly, fifth instar larvae provided
Biocontrol Science and Technology 311
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

Figure 3. Percentage of Helicoverpa armigera larvae parasitised by Habrobracon hebetor and
its association with weather parameters.
312 H. Saxena et al.
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014
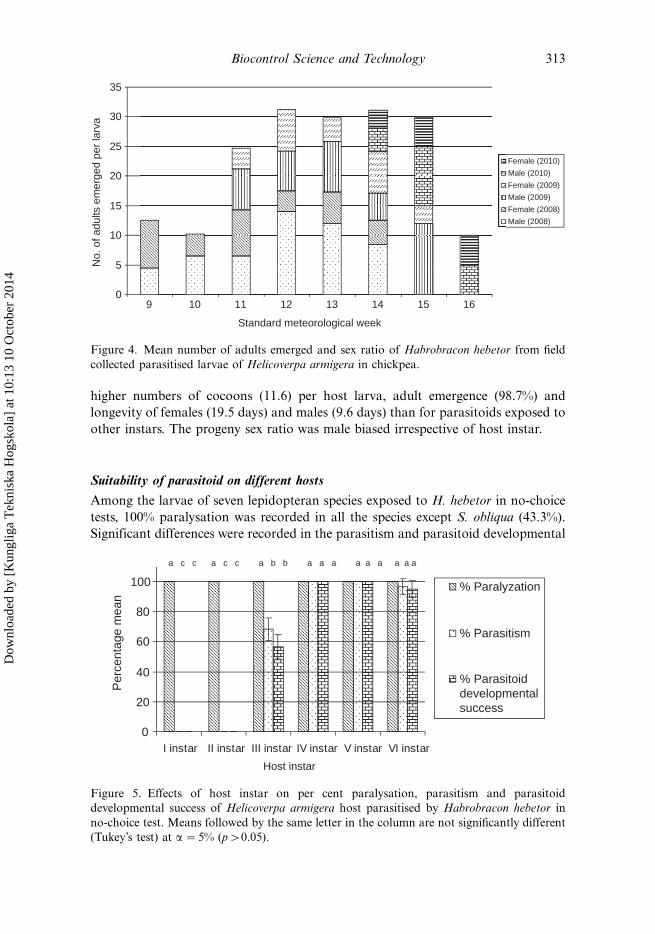
higher numbers of cocoons (11.6) per host larva, adult emergence (98.7%) and
longevity of females (19.5 days) and males (9.6 days) than for parasitoids exposed to
other instars. The progeny sex ratio was male biased irrespective of host instar.
Suitability of parasitoid on different hosts
Among the larvae of seven lepidopteran species exposed to H. hebetor in no-choice
tests, 100% paralysation was recorded in all the species except S. obliqua (43.3%).
Significant differences were recorded in the parasitism and parasitoid developmental
0
5
10
15
20
25
30
35
9 10 11 12 13 14 15 16
Standard meteorological week
Female (2010)
Male (2010)
Female (2009)
Male (2009)
Female (2008)
Male (2008)
No.
of a
dults
em
erge
d pe
r la
rva
Figure 4. Mean number of adults emerged and sex ratio of Habrobracon hebetor from field
collected parasitised larvae of Helicoverpa armigera in chickpea.
0
20
40
60
80
100
I instar II instar III instar IV instar V instar VI instar
Per
cent
age
mea
n
% Paralyzation
% Parasitism
% Parasitoiddevelopmentalsuccess
Host instar
a c c a c c a b b a a a a a a a a a
Figure 5. Effects of host instar on per cent paralysation, parasitism and parasitoid
developmental success of Helicoverpa armigera host parasitised by Habrobracon hebetor in
no-choice test. Means followed by the same letter in the column are not significantly different
(Tukey’s test) at a�5% (p�0.05).
Biocontrol Science and Technology 313
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

success in different host species (Tukey’s test, P50.05). Among the seven host
species, C. cephalonica, G. mellonella and H. armigera were the most suitable hosts,
with 100% parasitism and parasitoid development. It was followed by M. vitrata and
A. nigrisigna with 60�76.7% and 40�70% parasitism and parasitoid development,
respectively. There was no parasitoid developmental success in S. litura, though there
was 23.3% parasitism. No parasitism was recorded in S. obliqua (Figure 6).
Different host species significantly affected the survival and development of the
parasitoid (Table 2). The developmental time of H. hebetor on G. mellonella wassignificantly longer than other hosts (9.3 days), and similar to H. armigera (9.0 days)
and M. vitrata (9.0 days). Most rapid parasitoid development was observed in
C. cephalonica (8.7 days), which was on par with A. nigrisigna (8.8 days). The
parasitoids that developed on G. mellonella and C. cephalonica gave the highest
number of cocoons per host larva (16.7 and 14.4, respectively). It was followed by
H. armigera and M. vitrata which recorded a higher number of cocoons per host larva
(11.1 and 7.9, respectively) as compared to A. nigrisigna (4.2). Emergence of adults
was highest on C. cephalonica (97.6%), H. armigera (97.3%) and G. mellonella (95.4%).Similarly, adult longevity of female and male parasitoid was highest on C. cephalonica
(21.3 and 13.7 days) and G. mellonella (20.8 and 12.7 days), followed by H. armigera
(19.3 and 9.3 days). In contrast, parasitoids developing on A. nigrisigna and
M. vitrata recorded lowest adult emergence and adult longevity. The development
of parasitoids in all host species showed a male-biased sex ratio in their progeny.
Discussion
Although a large number of parasitoids have been reported attacking various stages
of H. armigera, only a few parasitoids contribute to regulation of H. armigera
populations in the chickpea ecosystem due to presence of glandular and non-glandular trichomes on the surface of the calyxes and pods and also due to the acid
exudates (Romeies, Shanmower, and Gupta 1997). In North India, Campoletis
chlorideae is the most efficient parasitoid of H. armigera in chickpea, and it
preferentially attacks early instars (second and third instar larvae) which feed mostly
Table 1. Survival and development of Habrobracon hebetor parasitising host instars of
Helicoverpa armigera.
Mean9SE
Adult longevity (days)
Host instar
Egg to adult
emergence
(days)
No. of
cocoon per
larva
Percentage of
adult
emergence
Sex ratio
(M:F) Female Male
Third instar 8.190.25b 3.390.65c 67.8911.01b 0.8390.20a 16.991.16b 8.590.39b
Fourth
instar
8.390.14b 8.390.82b 96.792.95a 0.7190.14ab 19.490.38a 9.390.36a
Fifth instar 9.190.20a 11.691.22a 98.790.93a 0.7390.09ab 19.590.48a 9.690.41a
Sixth instar 8.990.43a 8.190.72b 94.693.86a 0.5590.12b 18.790.34a 8.790.20b
Note: Means followed by the same superscript letter in the column are not significantly different (Tukey’stest) at a�5% (p�0.05).
314 H. Saxena et al.
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

on leaves (Dhillon and Sharma 2009). However, its occurrence and parasitism during
pod formation in March and April are very low (Yadava, Lal, Ahmad, and Sachan
1991). The braconid parasitoid, H. hebetor is known almost exclusively as a
parasitoid of pyralid moth larvae infesting stored grain (Eliopoulos and Stathas
2008). Previously, field parasitism of lepidopteran insects by H. hebetor was reported
in tea (Subbiah 1995), cotton (Gerling 1971) and coconut (Nasser and Abdurahiman
2001). However, no previous information is available on the seasonal occurrence of
the parasitoid on H. armigera in the chickpea ecosystem.
Seasonal occurrence and field parasitism of H. hebetor over three years indicated
parasitoid activity coinciding with the pod formation and pod maturation (February
to April) and mean percentage parasitism ranged from 0.9 to 12.3. Saxena (2007)
discussed the influence of climate change and development of large-seeded varieties
in chickpea on the feeding behaviour of H. armigera. She suggested that late instar
larvae prefer feeding on grain while completely within the pod, as compared to the
normal feeding habit of larvae as they feed with only their head inside the pod. This
behaviour may favour parasitism and population growth of H. hebetor on
H. armigera in chickpea. Interesting aspects of this parasitoid bio-ecology, that is,
occurrence in February to March and preference for late instar larvae of H. armigera,
indicate its non-competitive nature with C. chlorideae, which prefers early instar
larvae and occurs during November to February.
Many parasitoids exhibit a marked preference for a specific instar or larval stage
(Mackauer 1990; Mattiacci and Dicke 1995; McGregor 1996). We observed that late
instar larvae of H. armigera (fourth to sixth instars) were most suitable for successful
parasitism and development of H. hebetor. Parasitoid developmental rate is
0
10
20
30
40
50
60
70
80
90
100
Helicoverpaarmigera
Marucavitrata
Autographanigrisigna
Spodopteralitura
Spilarctiaobliqua
Corcyracephalonica
Galleriamellonella
Host species
Per
cent
age
mea
n
% Paralyzation % Parasitism % Parasitoid developmental success
a a a a b c a c d b c d a a a a a aa ab b
Figure 6. Effects of lepidopteran host species on per cent paralysation, parasitism and
parasitoid developmental success by Habrobracon hebetor in no-choice test. Means followed
by the same letter in the column are not significantly different (Tukey’s test) at a�5%
(p�0.05).
Biocontrol Science and Technology 315
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

frequently greater with older host larvae, presumably because more abundant
nutrients are available to support development of parasitoids in larger hosts (Vinson
and Iwantsch 1980; Lawrence 1990; Pennacchio, Vinson, and Tremblay 1993;
Harvey, Kadash, and Strand 2000). Higher numbers of cocoons, and greater adult
emergence and longer adult longevity of H. hebetor was recorded in the later instar
larvae (fourth to sixth instars) as compared to third instar larvae. This agrees with
Beckage and Templeton (1985), who found that maximum host larval size attracts
female oviposition and supports development of their progeny. The parasitoid,
Campoletis sonorensis (Hymenoptera: Ichneumonidae) develops faster in younger
H. virescens larvae and, specifically, total developmental time from oviposition to
adult emergence was significantly shorter when first or second instar hosts were
parasitised compared to later instars (Hu and Vinson 2000). Similarly, our results
showed that the developmental time of parasitoids was shorter on third and fourth
instar larvae as compared to fifth and sixth instars (Table 1).
Most parasitoids have the ability to determine host quality during oviposition
and will often accept or reject hosts on this basis (Charnov and Skinner 1985; Strand
and Pech 1995). The current study shows strong influence of different lepidopteran
host species on acceptance and survival of H. hebetor. Of the seven hosts presented to
H. hebetor, H. armigera, C. cephalonica and G. mellonella were the most suitable hosts
with 100% paralysation, parasitism and parasitoid development. M. vitrata, and A.
nigrisigna were least suitable with lower per cent parasitism (60.0�76.7%) and
parasitoid development (40.0�70.0%). Spodoptera litura and Spilarctia obliqua were
not suitable for the development of H. hebetor. This is similar to the findings of
Edwards, Weaver, and Marris (2001), who reported that failure to develop within a
given host may be due to the inability of the parasitoid to overcome their host
immune system. The parasitoid did not parasitise S. obliqua due to the presence of
hairs in the body, though it paralysed some of the larvae through the ventral body
Table 2. Survival and development of Habrobracon hebetor parasitising different lepidopteran
host species.
Mean9SE
Adult longevity (days)
Host species
Egg to adult
emergence
(days)
No. of
cocoon per
larva
Percentage of
adult
emergence
Sex ratio
(M:F) Female Male
Helicoverpa
armigera
9.090.25a 11.191.43b 97.392.33a 0.8690.18a 19.391.12b 9.390.73b
Maruca vitrata 9.090.23a 7.990.58b 83.1913.86ab 0.7990.27a 14.490.64c 8.390.60bc
Autographa
nigrisigna
8.890.52b 4.291.13c 77.2919.63b 0.7590.17a 11.890.16d 7.990.70c
Corcyra
cephalonica
8.790.31b 14.492.35a 97.693.83a 0.6890.05a 21.390.83a 13.791.26a
Galleria
mellonella
9.390.37a 16.793.08a 95.494.96a 0.6990.10a 20.890.48a 12.790.49a
Note: Means followed by the same superscript letter in the column are not significantly different (Tukey’stest) at a�5% (p�0.05).
316 H. Saxena et al.
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

surface which is devoid of hairs. The offspring of hymenopteran parasitoids depend
on the host for their nutritional needs. The nutritional quality may be different in
different hosts (Harvey et al. 2000; Harvey and Strand 2002). The developmental
time and survival to adult eclosion for H. hebetor reared on different host species
may give some indication of the suitability of hosts for the parasitoid. In the present
study, the developmental time from egg to adult emergence was longer on
G. mellonella as compared to other hosts.
Our results suggest that late, rather than early instars of any one of the hosts
H. armigera, C. cephalonica and G. mellonella are preferable for mass rearing of
H. hebetor under laboratory conditions. Field releases of H. hebetor should coincide
with the occurrence of field populations of late instar larvae of H. armigera in March
and April to maximise parasitism of this pest infesting chickpea in India.
Acknowledgements
We thank Prof. M. Hayat, Emeritus Scientist, Department of Zoology, Aligarh MuslimUniversity, Aligarh, Uttar Pradesh, India for his kind identification of the parasitoid. Authorsthank Dr C. Chattopadhyay, Head (Crop Protection) and Dr N. Nadarajan, Director, IndianInstitute of Pulses Research, Kanpur for their expert advice and for providing facilities. Wethank Dr Y.S. Rathore, Principal Scientist and Dr Aditya Pratap, Senior Scientist for Englishlanguage editing. We thank the two referees for their valuable comments for improvement ofthis study.
References
Armes, N.J., Bond, G.S., and Cooter, R.J. (1992), The Laboratory Culture and Development ofHelicoverpa armigera, Chatham: Natural Resources Institute, pp. 1�22.
Beckage, N.E., and Templeton, T.J. (1985), ‘Temporal Synchronization of Emergence ofHyposoter exiguae and H. fugitivus (Hymenoptera: Ichneumonidae) with ApolysisPreceding Larval Molting in Manduca sexta (Lepidoptera: Sphingidae)’, Annals of theEntomological Society of America, 78, 775�782.
Charnov, E.L., and Skinner, S.W. (1985), ‘Complementary Approaches to the Understandingof Parasitoid Oviposition Decision’, Environmental Entomology, 14, 383�391.
Dhillon, M.K., and Sharma, H.C. (2009), ‘Temperature Influences the Performance andEffectiveness of Field and Laboratory Strains of the Ichneumonid Parasitoid, Campoletischlorideae’, BioControl, 54, 743�750.
Edwards, J.P., Weaver, R.J., and Marris, G.C. (2001), ‘Endocrine Changes in LepidopteranLarvae: Potential Challenges to Parasitoid Development and Survival’, in EndocrineInteractions of Insect Parasites and Pathogens, eds. J.P. Edwards and R.J. Weaver, Oxford:BIOS Science Publications, pp. 1�32.
Eliopoulos, P.A., and Stathas, G.J. (2008), ‘Life Tables of Habrobracon hebetor (Hymenoptera:Braconidae) Parasitizing Anagasta kuehniella and Plodia interpunctella (Lepidoptera:Pyralidae): Effect of Host Density’, Journal of Economic Entomology, 101, 982�988.
Gerling, D. (1971), ‘Occurrence, Abundance, and Efficiency of Some Local ParasitoidsAttacking Spodoptera littoralis (Lepidoptera: Noctuidae) in Selected Cotton Fields inIsrael’, Annals of the Entomological Society of America, 64, 492�499.
Harvey, J.A., Kadash, K., and Strand, M.R. (2000), ‘Differences in Larval Feeding BehaviorCorrelate with Altered Developmental Strategies in two Parasitic Wasps: Implications forthe Size-fitness Hypothesis’, Oikos, 88, 621�629.
Harvey, J.A., and Strand, M.R. (2002), ‘The Developmental Strategies of EndoparasitoidWasps Vary with Host Feeding Ecology’, Ecology, 83, 2439�2451.
Hu, J.S., and Vinson, S.B. (2000), ‘Interaction between the Larval Endoparasitoid Campoletissonorensis (Hymenoptera: Ichneumonidae) and its Host the Tobacco Budworm (Lepidop-tera: Noctuidae)’, Annals of the Entomological Society of America, 93, 220�224.
Biocontrol Science and Technology 317
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014

Lawrence, P.O. (1990), ‘The Biochemical and Physiological Effects of Insects Host on theDevelopment and Ecology of their Insect Parasites: an Overview’, Archives of InsectBiochemistry and Physiology, 13, 217�228.
Mackauer, M. (1990), ‘Host Discrimination by Parasitoids’, in Semiochemical: Their Role inPest Control, eds. M. Mackauer, L.E. Ehler and J. Roland, Andover: Intercept, pp. 41�62.
Mattiacci, L., and Dicke, M. (1995), ‘The Parasitoid Cotesia glomerata (Hymenoptera:Braconidae) Discriminates between First and Fifth Larval Instars of its Host Pierisbrassicae on the Basis of Contact Cues from Frass, Silk and Herbivore-damaged LeafTissue’, Journal of Insect Behavior, 8, 485�497.
McGregor, R. (1996), ‘Phenotypic Selection by Parasitoids in the Timing of Life History in aLeafmining Moth’, Evolution, 50, 1579�1584.
Nasser, M., and Abdurahiman, U.C. (2001), ‘Biological Control of the Coconut CaterpillarOpisina arenosella (Lepidoptera: Xylorictidae): Achievements and Prospects’, in BiocontrolPotential and its Exploitation in Sustainable Agriculture (Volume II), eds. R.K. Upadhayay,K.G. Mukerji and B.P. Chamda, New York: Plenum publishers, pp. 285�302.
Nema, K.K. (2010), ‘Chickpea Insect Pest Management in Madhya Pradesh with SpecialReference to Helicoverpa armigera’, in Souvenir, All India Coordinated Research Project onChickpea, eds. D.R. Saxena, S.C. Gupta, A.N. Tikle, K.K. Nema, M. Yasin, R.P. Singh andA. Saxena, Sehore, India: R.A.K. College of Agriculture, pp. 35�38.
Pennacchio, F., Vinson, S.B., and Tremblay, E. (1993), ‘Growth and Development ofCardiochilies nigriceps Viereck (Hymenoptera: Braconidae) Larvae and Their Synchroniza-tion with Some Changes of the Hemolymph Composition of Their Host, Heliothis virescens(F.) (Lepidoptera: Noctuidae)’, Archives of Insect Biochemistry and Physiology, 24, 65�77.
Rao, P.P., Birthal, P.S., Bhagavatula, S., and Bantilan, M.C.S. (2010), ‘Chickpea and PigeonpeaEconomies in Asia: Facts, Trends and Outlook’, Andhra Pradesh, India: International CropsResearch Institute for the Semi-Arid Tropics, pp. 9�25.
Romeies, J., Shanmower, T.G., and Gupta, M. (1997), ‘Failure of Trichogramma Mass Releasesin Pigeonpea and Chickpea’, International Chickpea and Pigeonpea Newsletter, 4, 27�28.
Sachan, J.N., and Katti, G. (1994), ‘Integrated Pest Management’, in Proceedings of NationalSymposium on Pulses Research, pp. 23�26.
Saxena, H. (2007), ‘Behavioral Change in Helicoverpa armigera � an Emerging Challenge’, inProceedings of National Symposium on Legumes for Ecological Sustainablity: EmergingChallenges and Opportunities, pp.134�135.
Saxena, H., and Duraimurugan, P. (2009), ‘Record of Bracon hebetor Say (Hymenoptera:Braconidae) on Helicoverpa armigera (Hubner) in Chickpea’, Journal of Biological Control,23, 191�192.
Schliserman, P. (2001), ‘Seasonal abundance of hymenopterous parasitoids of Anastrephafraterculus and Ceratitis capitata (Diptera: Tephritidae) in areas of secondary forest in theSierra de San Javier, Tucuman’, MSc Thesis, Regional Research Center, Science andTechnology Transfer, National University of La Rioja, Argentina, 120 pp.
Strand, M.R., and Pech, L.L. (1995), ‘Immunological Basis for Compatibility in Parasitoid-Host Relationships’, Annual Review of Entomology, 40, 31�56.
Subbiah, K. (1995), ‘The Parasitism by Bracon hebetor (Say) (Hymenoptera: Braconidae) onthe Larvae of Cydia leucostoma in Field Conditions’, Journal of Economic Entomology, 7,299�302.
Vinson, S.B., and Iwantsch, G.F. (1980), ‘Host Suitability for Insect Parasitoids’, AnnualReview of Entomology, 25, 397�419.
Yadava, C.P., Lal, S.S., Ahmad, R., and Sachan, J.N. (1991), ‘Influence of Abiotic Factor onRelative Abundance of Pod Borers of Chickpea Cicer arietinum’, Indian Journal ofAgricultural Science, 61, 512�515.
318 H. Saxena et al.
Dow
nloa
ded
by [
Kun
glig
a T
ekni
ska
Hog
skol
a] a
t 10:
13 1
0 O
ctob
er 2
014