scleractinian corals produce calcite, and grow more slowly, in artificial cretaceous seawater
TRANSCRIPT

Geology
doi: 10.1130/G22600.1 2006;34;525-528Geology
Justin B. Ries, Steven M. Stanley and Lawrence A. Hardie Cretaceous seawaterScleractinian corals produce calcite, and grow more slowly, in artificial
Email alerting servicescite this article
to receive free e-mail alerts when new articleswww.gsapubs.org/cgi/alertsclick
Subscribe to subscribe to Geologywww.gsapubs.org/subscriptions/click
Permission request to contact GSAhttp://www.geosociety.org/pubs/copyrt.htm#gsaclick
viewpoint. Opinions presented in this publication do not reflect official positions of the Society.positions by scientists worldwide, regardless of their race, citizenship, gender, religion, or politicalarticle's full citation. GSA provides this and other forums for the presentation of diverse opinions and articles on their own or their organization's Web site providing the posting includes a reference to thescience. This file may not be posted to any Web site, but authors may post the abstracts only of their unlimited copies of items in GSA's journals for noncommercial use in classrooms to further education andto use a single figure, a single table, and/or a brief paragraph of text in subsequent works and to make
GSA,employment. Individual scientists are hereby granted permission, without fees or further requests to Copyright not claimed on content prepared wholly by U.S. government employees within scope of their
Notes
The Geological Society of America, Inc.
on May 5, 2012geology.gsapubs.orgDownloaded from
q 2006 Geological Society of America. For permission to copy, contact Copyright Permissions, GSA, or [email protected]; July 2006; v. 34; no. 7; p. 525–528; doi: 10.1130/G22600.1; 4 figures. 525
Scleractinian corals produce calcite, and grow more slowly, inartificial Cretaceous seawaterJustin B. Ries Morton K. Blaustein Department of Earth and Planetary Sciences, Johns Hopkins University, Baltimore,
Maryland 21218, USASteven M. Stanley Department of Geology and Geophysics, University of Hawaii, 1680 East-West Road, Honolulu, Hawaii
96822, USALawrence A. Hardie Morton K. Blaustein Department of Earth and Planetary Sciences, Johns Hopkins University, Baltimore,
Maryland 21218, USA
ABSTRACTThe mineralogies of most biotic and abiotic carbonates have alternated in synchroneity
between the calcite (hexagonal) and aragonite (orthorhombic) polymorphs of CaCO3
throughout the Phanerozoic Eon. These intervals of calcite and aragonite production, orcalcite seas and aragonite seas, are thought to be caused primarily by secular variationin the molar magnesium/calcium ratio of seawater (mMg/Ca . 2 5 aragonite 1 high-Mgcalcite; mMg/Ca , 2 5 low-Mg calcite), a ratio that has oscillated between 1.0 and 5.2throughout the Phanerozoic. In laboratory experiments, we show that three species ofscleractinian corals, which produce aragonite in modern seawater and which have flour-ished as important reef builders primarily during aragonite seas of the past, began pro-ducing calcite in artificial seawater with an ambient mMg/Ca ratio below that of modernseawater (5.2). The corals produced progressively higher percentages of calcite and cal-cified at lower rates with further reduction of the ambient mMg/Ca ratio. In artificialseawater of imputed Late Cretaceous composition (mMg/Ca 5 1.0), which favors the pre-cipitation of the calcite polymorph, scleractinian corals produced skeletons containing.30% low-Mg calcite (skeletal mMg/Ca , 0.04). These results indicate that the skeletalmineral used by scleractinian corals is partially determined by seawater chemistry. Fur-thermore, slow calcification rates, resulting from the production of largely aragonitic skel-etons in chemically unfavorable seawater (mMg/Ca , 2), probably contributed to thescleractinians’ diminished reef-building role in the calcite seas of Late Cretaceous andearly Cenozoic time.
Keywords: scleractinian corals, calcification, calcite seas, aragonite seas, biomineralization,seawater Mg/Ca, reef building.
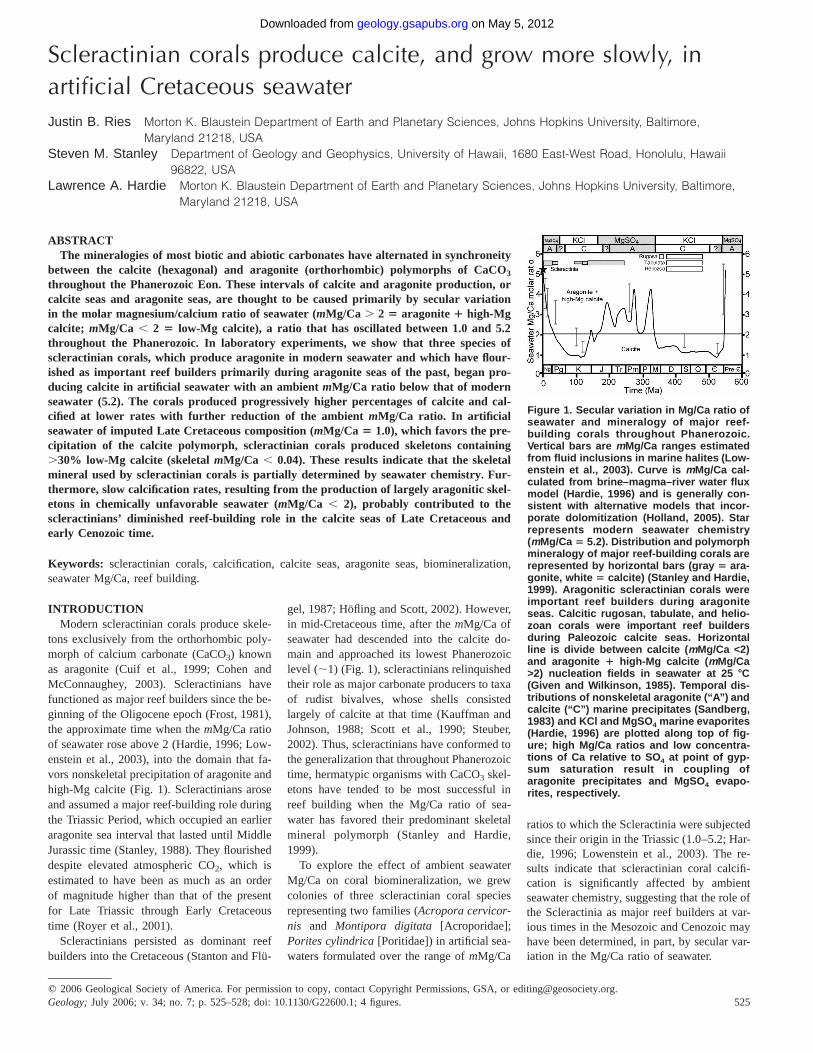
Figure 1. Secular variation in Mg/Ca ratio ofseawater and mineralogy of major reef-building corals throughout Phanerozoic.Vertical bars are mMg/Ca ranges estimatedfrom fluid inclusions in marine halites (Low-enstein et al., 2003). Curve is mMg/Ca cal-culated from brine–magma–river water fluxmodel (Hardie, 1996) and is generally con-sistent with alternative models that incor-porate dolomitization (Holland, 2005). Starrepresents modern seawater chemistry(mMg/Ca 5 5.2). Distribution and polymorphmineralogy of major reef-building corals arerepresented by horizontal bars (gray 5 ara-gonite, white 5 calcite) (Stanley and Hardie,1999). Aragonitic scleractinian corals wereimportant reef builders during aragoniteseas. Calcitic rugosan, tabulate, and helio-zoan corals were important reef buildersduring Paleozoic calcite seas. Horizontalline is divide between calcite (mMg/Ca <2)and aragonite 1 high-Mg calcite (mMg/Ca>2) nucleation fields in seawater at 25 8C(Given and Wilkinson, 1985). Temporal dis-tributions of nonskeletal aragonite (‘‘A’’) andcalcite (‘‘C’’) marine precipitates (Sandberg,1983) and KCl and MgSO4 marine evaporites(Hardie, 1996) are plotted along top of fig-ure; high Mg/Ca ratios and low concentra-tions of Ca relative to SO4 at point of gyp-sum saturation result in coupling ofaragonite precipitates and MgSO4 evapo-rites, respectively.
INTRODUCTIONModern scleractinian corals produce skele-
tons exclusively from the orthorhombic poly-morph of calcium carbonate (CaCO3) knownas aragonite (Cuif et al., 1999; Cohen andMcConnaughey, 2003). Scleractinians havefunctioned as major reef builders since the be-ginning of the Oligocene epoch (Frost, 1981),the approximate time when the mMg/Ca ratioof seawater rose above 2 (Hardie, 1996; Low-enstein et al., 2003), into the domain that fa-vors nonskeletal precipitation of aragonite andhigh-Mg calcite (Fig. 1). Scleractinians aroseand assumed a major reef-building role duringthe Triassic Period, which occupied an earlieraragonite sea interval that lasted until MiddleJurassic time (Stanley, 1988). They flourisheddespite elevated atmospheric CO2, which isestimated to have been as much as an orderof magnitude higher than that of the presentfor Late Triassic through Early Cretaceoustime (Royer et al., 2001).
Scleractinians persisted as dominant reefbuilders into the Cretaceous (Stanton and Flu-
gel, 1987; Hofling and Scott, 2002). However,in mid-Cretaceous time, after the mMg/Ca ofseawater had descended into the calcite do-main and approached its lowest Phanerozoiclevel (;1) (Fig. 1), scleractinians relinquishedtheir role as major carbonate producers to taxaof rudist bivalves, whose shells consistedlargely of calcite at that time (Kauffman andJohnson, 1988; Scott et al., 1990; Steuber,2002). Thus, scleractinians have conformed tothe generalization that throughout Phanerozoictime, hermatypic organisms with CaCO3 skel-etons have tended to be most successful inreef building when the Mg/Ca ratio of sea-water has favored their predominant skeletalmineral polymorph (Stanley and Hardie,1999).
To explore the effect of ambient seawaterMg/Ca on coral biomineralization, we grewcolonies of three scleractinian coral speciesrepresenting two families (Acropora cervicor-nis and Montipora digitata [Acroporidae];Porites cylindrica [Poritidae]) in artificial sea-waters formulated over the range of mMg/Ca
ratios to which the Scleractinia were subjectedsince their origin in the Triassic (1.0–5.2; Har-die, 1996; Lowenstein et al., 2003). The re-sults indicate that scleractinian coral calcifi-cation is significantly affected by ambientseawater chemistry, suggesting that the role ofthe Scleractinia as major reef builders at var-ious times in the Mesozoic and Cenozoic mayhave been determined, in part, by secular var-iation in the Mg/Ca ratio of seawater.
on May 5, 2012geology.gsapubs.orgDownloaded from

526 GEOLOGY, July 2006
Figure 2. A: X-ray diffraction (XRD) patternfor Porites cylindrica coral that producedexclusively aragonite when grown in artifi-cial seawater favoring nucleation of arago-nite polymorph (mMg/Ca 5 5.2). Primaryaragonite peak occurs at 2u 5 26.28 [d(111);3.40 A]. B: XRD pattern for pure calcite, re-vealing primary calcite peak at 2u 5 29.48–29.58 [d(104); 3.03–3.04 A]. C: XRD patternfor P. cylindrica coral that produced 33% 63% calcite and 67% 6 3% aragonite whengrown in artificial seawater favoring nucle-ation of calcite polymorph (mMg/Ca 5 1.0).Proportion of calcite to aragonite was cal-culated as ratio of areas beneath primarycalcite and aragonite peaks.
Figure 3. Backscattered electron (BSE) images (A, D: JEOL 8600 Superprobe), Sr maps (B,E: wavelength-dispersive spectrometry), and Mg maps (C, F: energy-dispersive spectrom-etry) showing, respectively, skeletal microstructure, aragonite, and calcite in portions oftwo specimens of Porites cylindrica that were grown in artificial seawater favoring abioticprecipitation of calcite over aragonite (mMg/Ca 5 1.0). Light areas (mSr/Ca 5 0.011–0.016)of Sr maps (B, E) and dark areas (mMg/Ca 5 0.003–0.007) of Mg maps (C, F) correspond toaragonite, while dark areas (mSr/Ca 5 0.001–0.005) of Sr maps (B, E) and light areas (mMg/Ca 5 0.019–0.033) of Mg maps (C, F) correspond to calcite. Calcite is located on outermostportion of coral skeleton, directly beneath coral ectoderm and, therefore, in contact withcoral’s mechanism of calcification. Scale bars are 100 mm.
METHODSWe grew 70 comparably sized individuals
(50 living 1 20 dead controls) of each of the3 species for 60 days in 5 10 gal aquaria con-taining artificial seawaters that were identical-ly formulated (at modern values) except fortheir mMg/Ca ratios, which were fixed at 1.0,1.5, 2.5, 3.5, and 5.2 (Ries, 2004). Coral spec-imens were acclimated to the artificial sea-water treatments over a 30 day period in stag-es, to minimize any chemical shock fromexposure to the modified seawater. Water wasmaintained at 25 6 1 8C using a 50 W heater,circulated with two motor-driven pumps, andcontinuously filtered at a rate of 600 L/h. Cor-al lights that emit wavelengths commensuratewith midday and dawn or twilight natural lightprovided 8 h/day of irradiation. Corals werefed 1.8 mL of Red Sea CoralGro plankton sus-pension daily. Red Sea Vitamin (0.75 mL),Coral Trace Element (1.5 mL), and iodine(0.75 mL) supplements were added to theaquaria weekly.
RESULTS AND DISCUSSIONScleractinian Corals Produce Calcite inArtificial Cretaceous Seawater
X-ray diffraction analysis of new skeletalgrowth (Fig. 2) revealed that corals grown inartificial seawaters formulated with lower thannormal Mg/Ca ratios (mMg/Ca , 5.2) beganincorporating the hexagonal calcite polymorphof CaCO3 into their skeleton. Dead corals (n5 60) placed alongside the live corals as con-trols to monitor the formation of abiotic cal-cite cements within the coral skeletons failedto incorporate any calcite. The proportion ofskeletal calcite in the living corals (Fig. 3) in-creased with reduction of the ambient Mg/Caratio (Fig. 4A). All three species of corals ac-creted about one-third of their skeletal mate-rial as low-Mg calcite when grown in the low-est Mg/Ca artificial calcite seawater (mMg/Ca5 1.0). This suggests that scleractinian corals
accreted at least part of their skeleton as low-Mg calcite in the calcite seas of Late Creta-ceous and early Cenozoic time (Floris, 1972;Fig. 1).
Mg Fractionation in CoralsThe Mg/Ca ratio of the calcitic portions of
the corals’ skeletons increased proportionallywith the Mg/Ca ratio of the ambient seawater(Fig. 4B). Low-Mg calcite was produced withambient mMg/Ca , 2 and high-Mg calcite(mMg/Ca . 0.04) was produced with ambientmMg/Ca . 2. This relationship is consistentwith that observed for Mg incorporation incalcareous animals (echinoids, crabs, shrimp,serpulid worms) (Ries, 2004), coralline algae(Stanley et al., 2002), and some species ofcoccolithophores (Stanley et al., 2005). TheMg/Ca ratios of the aragonite accreted by thecorals also increased with the ambient Mg/Ca
on May 5, 2012geology.gsapubs.orgDownloaded from

GEOLOGY, July 2006 527
Figure 4. Mineralogy, geochemistry, and cal-cification rates of Acropora cervicornis(white symbols), Montipora digitata (graysymbols), and Porites cylindrica (black sym-bols) grown in artificial seawater treatments.A: Molar % calcite in coral skeletons (n 5 15for each species). B: mMg/Ca ratio of calcit-ic (squares) and aragonitic (diamonds) por-tions of coral skeleton, determined by mi-croprobe (energy-dispersive spectrometry).Solid black lines represent Mg-fractionationcurves for coral calcite (y 5 0.0213x1.46, R2
5 0.95, n 5 45) and coral aragonite (y 50.0020x, R2 5 0.75, n 5 35), calculated usingleast squares regression. C: Calcificationrate of corals (n 5 50 for each species) de-termined by buoyant weighing at beginningand end of experiment. Ca concentration ofseawater decreases as Mg/Ca increases. Er-ror bars indicate instrument error and spec-imen variation.
ratio (Fig. 4B). Therefore, paleotemperaturereconstructions based on the Mg/Ca ratio offossil coral skeletons require correction for theeffect of secular variation in seawater Mg/Ca(Mitsuguchi et al., 1996).
Calcification RatesEach of the three species of scleractinian
corals calcified most rapidly in the aragoniteseawater treatments (Fig. 4C). A significant
decrease in the rate of calcification occurredwhen seawater crossed into the calcite nucle-ation field at mMg/Ca , 2, most likely due tothe difficulty of precipitating aragonite—thecorals’ predominant skeletal mineral—in thismedium.
In pioneering experiments on the influenceof Mg and Ca concentrations on coral biomin-eralization, Swart (1980) grew specimens ofAcropora aquamosa, Pocillopora damicornis,Acropora cuneata, and Porites lutea for 10days in seawater with Mg concentrations 100and 200 mg/L higher than modern values andCa concentrations 100, 200, and 400 mg/Lhigher than modern values. Ambient Mg con-centrations significantly higher than that ofmodern seawater prevented new skeletalgrowth, while Ca concentrations twice thoseof modern seawater resulted in reduced ratesof calcification. The effect of elevated Ca isconsistent with our findings that coral calcifi-cation rates decreased significantly in artificialseawaters with mMg/Ca ratios that were low(1.0, 1.5) relative to modern values. The shortduration of these experiments and the employ-ment of higher concentrations of Mg and Cathan have ever characterized Phanerozoic sea-water preclude interpretation of the results inthe context of actual past marine conditions.
Other experiments on specimens of Acro-pora sp., Stylophora pistillata, A. cervicornis,and A. formosa showed that calcification ratesincrease as Ca concentrations are elevatedfrom ;20% to 80% of normal marine values(Chalker, 1976; Gattuso et al., 1998). At Caconcentrations .80%, calcification rates lev-eled out and, in some cases, declined. How-ever, these experiments did not adjust Mg con-comitantly with Ca, and therefore failed tocontrol for the effects of changes in the Mg/Ca ratio of the artificial seawater. When theobserved variation in calcification rate is cor-related with the implicit variation in Mg/Caratio of the artificial seawaters, the results con-form to those observed in our study. In addi-tion, the short durations (2–2.5 h) of the ex-periments may not have allowed sufficienttime for the corals to fully equilibrate to themodified ambient conditions.
Modes of Coral Calcification and CaCO3
Polymorph ControlIt has been proposed that organic matter
within centers of calcification of scleractiniancorals (as much as 1% by volume) is a vestigeof organic matrices and templates that specifynucleation of aragonite (Cuif et al., 1999). Thefact that corals transferred to artificial calciteseawater still precipitated two-thirds of theirskeletal material as aragonite supports the hy-pothesis that corals exert significant controlover their skeletal mineralogy. However, ourdiscovery that corals precipitate approximate-ly one-third of their skeleton as low-Mg cal-
cite in the artificial calcite seawater indicatesthat their biomineralogical control can be par-tially overridden by ambient seawater chem-istry. The tendency of corals to incorporateMg (this study) and other trace elements (Co-hen and McConnaughey, 2003) into their skel-etons in proportions reflecting their abundancein seawater is additional evidence of their lim-ited biomineralogical control. Furthermore,the acicular morphology and spherulitic or-ganization of aragonitic crystals in the scler-odermites of coral skeletons closely resemblethe aragonite of marine cements, suggestingthat coral aragonite could be deposited in away similar to abiotic precipitation (Cohenand McConnaughey, 2003).
Seawater reaches the coral’s region of cal-cification, beneath the calicoblastic ectoderm,by moving through and between cells and bydiffusion through the porous skeleton (Cohenand McConnaughey, 2003). An ATPase pumpelevates Ca in the region of calcification onlyslightly above the level of ambient seawater(Al-Horani et al., 2003), thereby suggestingthat corals precipitate their skeletons fromonly slightly modified seawater. The high pre-cipitation rate of the coral skeleton, nearly 100times that of inorganic CaCO3 precipitation(Cohen and McConnaughey, 2003), has in-stead been attributed to the pumping of pro-tons from the calcification medium (Al-Horaniet al., 2003). It is therefore not surprising thatthe Mg/Ca ratio of seawater has a significanteffect on the chemistry of a coral’s calcifica-tion medium and, therefore, on its skeletalmineralogy.
While a secondary, abiogenic mode of cal-cification in scleractinian corals is known tooccur beneath the ectodermal calcificationspace, filling pore spaces no longer occupiedby living tissue (Enmar et al., 2000), the cal-cite observed in our experimental corals ac-tually appears to be primary skeletal material.The biogenic nature of this coral calcite is ev-ident in that: (1) the calcite occurs on the out-ermost portion of the skeleton (Fig. 3), im-mediately beneath the corals’ ectodermaltissue and, thus, in direct contact with the cor-als’ mechanism of calcification; (2) the fine-scale integration of the calcite crystals withthe skeletal aragonite of the experimental cor-als (Fig. 3) generally preserves the morphol-ogy and integrity of the corals’ skeletons; and(3) dead corals placed alongside the live coralsto monitor the formation of abiogenic calcitecements within the coral skeletons failed toincorporate any calcite.
Implications for the Geologic History ofScleractinian Reef Construction
Our results suggest that scleractinian corals’diminished reef-building role from mid-Cretaceous through Eocene time (Frost, 1981;Stanton and Flugel, 1987; Hofling and Scott,
on May 5, 2012geology.gsapubs.orgDownloaded from

528 GEOLOGY, July 2006
2002) was caused, at least in part, by a dropin oceanic Mg/Ca over this interval (Fig. 1)to the degree that seawater no longer favoredthe precipitation of the Scleractinia’s predom-inantly aragonitic mineralogy (Stanley andHardie, 1999). Depressed calcification ratesover this interval may have opened the reef-building environments to the rudist bivalves(Stanley and Hardie, 1999), which were pri-marily calcitic after the Cenomanian (Steuber,2002) and became important constituents ofcarbonate reef facies during the mid-Cretaceous (Kauffman and Johnson, 1988;Scott et al., 1990; Moussavian, 1992; Gili etal., 1995).
The Scleractinia failed to resume their roleas major reef builders until well after the ex-tinction of the rudists. They began to formmassive widespread reefs again early in theOligocene (Frost, 1981), shortly after ambientmMg/Ca ratios had risen above two (Fig. 1)into the aragonite stability field (Stanley andHardie, 1999). A progressive decline in theconcentration of atmospheric CO2 over theOligocene Epoch (Pagani et al., 2005), whichwould have increased the CaCO3 saturationstate of seawater, may have also contributedto the Scleractinia’s resurgence over this in-terval. The highest calcification rates for thethree scleractinian species in this experimentoccurred both in modern seawater (mMg/Ca5 5.2) and in seawater with mMg/Ca 5 3.5(Fig. 4C). We hypothesize that the corals’ el-evated calcification rates in seawater withmMg/Ca 5 3.5, a level approached in Oligo-cene time, resulted from the combined effectof a favorable ambient Mg/Ca ratio (mMg/Ca. 2) and a high concentration of Ca relativeto that of modern seawater (Stanley et al.,2005).
CONCLUSIONSWe grew colonies of three species of scler-
actinian corals representing two families infive artificial seawaters that differed only intheir mMg/Ca ratios, which ranged from 5.2(that of the modern ocean) to 1.0 (a Late Cre-taceous level) (Hardie, 1996; Lowenstein etal., 2003). For all three species, stepwise re-duction of the ambient Mg/Ca ratio resultedin a progressive decrease in the rate of skeletalcalcification and an increase in the proportionof skeletal calcite. We conclude that sclerac-tinians exert only partial control over theirskeletal mineralogy, but we suggest that thepredominance of the aragonite polymorph insome, if not all, of their species played a rolein their mid-Cretaceous decline and their Ol-igocene resurgence as major reef builders.These experimental results support the broaderfossil and geochemical evidence that secularvariation in seawater Mg/Ca has influencedthe reef-building and sediment-producing
roles of many hypercalcifying marine taxathroughout the Phanerozoic (Stanley and Har-die, 1999; Ries, 2004, 2006; Stanley et al.,2002, 2005).
ACKNOWLEDGMENTSThis manuscript was improved by the reviews of
Peter Swart, Anne Cohen, and Thomas Steuber. Weacknowledge the National Science Foundation andthe Petroleum Research Fund for funding this study,and thank the Moody Gardens Aquarium for sup-plying coral specimens.
REFERENCES CITEDAl-Horani, F.A., Al-Moghrabi, S.M., and De Beer,
D., 2003, The mechanism of calcification andits relation to photosynthesis and respiration inthe scleractinian coral Galaxea fascicularis:Marine Biology, v. 142, p. 419–426.
Chalker, B.E., 1976, Calcium-transport during ske-letogenesis in hermatypic corals: Compara-tive Biochemistry and Physiology, v. 54,p. 455–459.
Cohen, A.L., and McConnaughey, T.A., 2003, Ageochemical perspective on coral mineraliza-tion, in Dove, P.M., et al., eds., Biominerali-zation: Reviews in Mineralogy and Geochem-istry Volume 54, p. 151–187.
Cuif, J.P., Dauphin, Y., Doucet, J., Gautret, P., andZibrowius, H., 1999, Biochemical markers ofzooxanthellae symbiosis in soluble matrices ofskeleton in scleractinian coral skeletons: Geo-chimica et Cosmochimica Acta, v. 67,p. 75–83.
Enmar, R., Lazar, B., Bar-Matthews, M., Sass, E.,Katz, A., and Stein, M., 2000, Diagenesis inlive corals from the Gulf of Aqaba. I. The ef-fect on paleo-oceanography tracers: Geo-chimica et Cosmochimica Acta, v. 64,p. 3123–3132.
Floris, S., 1972, Scleractinian corals from the UpperCretaceous and lower Tertiary of Nugssuaq,West Greenland: Museum de Mineralogie etde Geologie de L’Universite de Copenhague,Communications Paleontologiques no. 183,132 p.
Frost, S.H., 1981, Oligocene reef coral biofacies ofthe Vicentin, northeast Italy, in Toomey, D.F.,ed., European fossil reef models: Society ofEconomic Paleontologists and MineralogistsSpecial Publication 30, p. 483–539.
Gattuso, J.P., Frankignoulle, M., Bourge, I., Ro-maine, S., and Buddemeier, R.W., 1998, Effectof calcium carbonate saturation of seawater oncoral calcification: Global and PlanetaryChange, v. 18, p. 37–46.
Gili, E., Masse, J.-P., and Skelton, P.W., 1995, Rud-ists as gregarious sediment-dwellers, not reef-builders, on Cretaceous carbonate platforms:Palaeogeography, Palaeoclimatology, Palaeo-ecology, v. 118, p. 245–267.
Given, R.K., and Wilkinson, B.H., 1985, Kineticcontrol of morphology, composition, and min-eralogy of abiotic sedimentary carbonates:Journal of Sedimentary Petrology, v. 55,p. 109–119.
Hardie, L.A., 1996, Secular variation in seawaterchemistry: An explanation for the coupledsecular variation in the mineralogies of marinelimestones and potash evaporites over the past600 m.y: Geology, v. 24, p. 279–283.
Hofling, R., and Scott, R.W., 2002, Early and mid-Cretaceous build-ups, in Kiessling, W., et al.,eds., Phanerozoic reef patterns: SEPM (Soci-ety for Sedimentary Geology) Special Publi-cation 72, p. 521–548.
Holland, H.D., 2005, Sea level, sediments and thecomposition of seawater: American Journal ofScience, v. 305, p. 220–239.
Kauffman, E.G., and Johnson, C.C., 1988, The mor-phological and ecological evolution of middleand Upper Cretaceous reef-building rudistids:Palaios, v. 3, p. 194–216.
Lowenstein, T.K., Hardie, L.A., Timofeeff, M.N.,and Demicco, R.V., 2003, Secular variation inseawater chemistry and the origin of calciumchloride basinal brines: Geology, v. 31,p. 857–860.
Mitsuguchi, T., Matsumoto, E., Abe, O., Uchida, T.,and Isdale, P.J., 1996, Mg/Ca thermometry incoral skeletons: Science, v. 274, p. 961–963.
Moussavian, E., 1992, On Cretaceous bioconstruc-tions: Composition and evolutionary trends ofcrust-building associations: Facies, v. 26,p. 117–144.
Pagani, M., Zachos, J.C., Freeman, K.H., Tipple, B.,and Bohaty, S., 2005, Marked decline in at-mospheric carbon dioxide concentrationsduring the Paleogene: Science, v. 309,p. 600–603.
Ries, J.B., 2004, Effect of ambient Mg/Ca ratio onMg fractionation in calcareous marine inver-tebrates: A record of the oceanic Mg/Ca ratioover the Phanerozoic: Geology, v. 32,p. 981–984.
Ries, J.B., 2006, Aragonitic algae in calcite seas:Effect of seawater Mg/Ca on codiacean bio-mineralization: Journal of Sedimentary Re-search (in press).
Royer, D.L., Berner, R.A., and Beerling, D.J., 2001,Phanerozoic atmospheric CO2 change: Evalu-ating geochemical and paleobiological ap-proaches: Earth-Science Reviews, v. 54,p. 349–392.
Sandberg, P.A., 1983, An oscillating trend in Phan-erozoic nonskeletal carbonate mineralogy: Na-ture, v. 305, p. 19–22.
Scott, R.W., Fernandez-Mendiola, P.A., Gili, E., andSimo, A., 1990, Persistence of coral-rudistreefs into the Late Cretaceous: Palaios, v. 5,p. 98–110.
Stanley, G.D., 1988, The history of Mesozoic reefcommunities: A three-step process: Palaios,v. 3, p. 170–183.
Stanley, S.M., and Hardie, L.A., 1999, Hypercalci-fication: Paleontology links plate tectonics andgeochemistry to sedimentology: GSA Today,v. 9, p. 1–7.
Stanley, S.M., Ries, J.B., and Hardie, L.A., 2002,Low-magnesium calcite produced by corallinealgae in seawater of Late Cretaceous compo-sition: National Academy of Sciences Pro-ceedings, v. 99, p. 15323–15326.
Stanley, S.M., Ries, J.B., and Hardie, L.A., 2005,Seawater chemistry, coccolithophore popula-tion growth, and the origin of Cretaceouschalk: Geology, v. 33, p. 593–596.
Stanton, R.J., and Flugel, E., 1987, Paleoecology ofUpper Triassic reefs in the northern Calcare-ous Alps: Reef communities: Facies, v. 16,p. 157–186.
Steuber, T., 2002, Plate tectonic control on the evo-lution of Cretaceous platform-carbonate pro-duction: Geology, v. 30, p. 259–262.
Swart, P., 1980, The effect of seawater chemistry onthe growth of some scleractinian corals, inTardent, P., and Tardent, R., eds., Developmentand cellular biology of Coelenterates: Amster-dam, Elsevier, p. 203–208.
Manuscript received 16 January 2006Revised manuscript received 7 February 2006Manuscript accepted 9 February 2006
Printed in USA
on May 5, 2012geology.gsapubs.orgDownloaded from