rubella virus induces apoptosis in culture cells · 2017-02-21 · consequence of viral...
TRANSCRIPT

Rubella Virus Induces Apoptosis in Culture Cells
Konstantin V. Pugachev and Teryl K. Frey1
Department of Biology, Georgia State University, Atlanta, Georgia 30303
Received April 17, 1998; returned to author for revision May 26, 1998; accepted August 25, 1998
The replication of rubella virus (RUB) in Vero cells, an adherent cell line, results in apoptotic death of infected cells asdetected by chromatin fragmentation assays. In infected cultures, virtually all of the cells that had become detached (ahallmark feature of RUB-induced cytopathology) were apoptotic; they were predominantly dead as shown by propidium iodideand trypan blue exclusion tests. In contrast, the majority of the cells in the infected monolayers that remained adherent werealive and contained intact chromatin. Thus simple counting of detached cells in the medium is a convenient way of measuringthe extent of RUB-induced apoptosis. RUB-induced cytopathology was inhibited by z-VAD-fmk, an inhibitor of caspases thatare involved in the execution stages of apoptosis, confirming the induction of apoptosis by RUB. The lack of apoptoticadherent cells (maximally 1% at any time point through 6 days postinfection) indicates that the induction of apoptosis isasynchronous since cells become uniformly virus antigen-positive by day 2 postinfection. To elucidate whether thisasynchronicity and the ability of RUB to persistently infect Vero cells were due to a suppression of apoptosis, we examinedwhether RUB can suppress chemically induced apoptosis. Staurosporine (ST) was found to be an efficient inducer ofapoptosis in Vero cells. ST treatment of RUB-infected and RUB persistently infected cells resulted in a much higher proportionof detached cells, higher even than in Vero cells treated with ST alone. This indicates that RUB does not suppress ST-inducedapoptosis and, rather, that ST and RUB acted cumulatively in inducing apoptosis, possibly indicating that they use differentinduction pathways. © 1998 Academic Press
INTRODUCTION
Apoptosis, or programmed cell death, is a commonconsequence of viral replication, as has been demon-strated for a growing number of RNA and DNA viruses incell culture and in vivo, which can contribute to thesymptomology of viral diseases (Razvi and Welsh, 1995;Teodoro and Branton, 1997). The purpose of this studywas to determine whether rubella virus (RUB) also be-longs to the category of viruses that kill infected culturecells via induction of apoptosis. Although RUB infectionis generally asymptomatic or benign in nature (typified bya rash of short duration), it can result in complicationssuch as transient and chronic polyarthralgia and arthritis(especially in adult women), thrombocytopenic purpura,and postinfectious encephalopathy (medical aspects ofrubella reviewed by Wolinsky, 1996). The most seriousconsequence of RUB infection is known as congenitalrubella syndrome, a complex of severe birth defects(deafness, blindness, heart and brain damage, or mis-carriage) caused by virus replication in the fetus afterinfection during the first trimester of pregnancy.
RUB is the only member of the Rubivirus genus of theTogaviridae family that also includes the Alphavirus ge-nus. RUB is an enveloped single-stranded, plus-senseRNA virus (the molecular biology of RUB is reviewed in
Frey, 1994). In the virion, the genomic RNA of ;10,000nucleotides in length, which is capped and polyadenyl-ated, is enclosed within an icosahedral capsid com-posed of the capsid protein C. The capsid is surroundedby an envelope with two embedded viral glycoproteins,E1 and E2. The whole replication cycle of RUB, includingviral RNA replication, protein synthesis, and assembly ofnew virions, occurs in the cytoplasm. Three viral RNAspecies are found in infected cells: the genomic RNA,which is also the mRNA for the nonstructural proteinsnecessary for viral RNA synthesis; the complementarygenome-length, minus-sense RNA, which is a templatefor synthesis of plus-strands; and a subgenomic RNA,which corresponds to the 39-terminal third of the genomeand serves as the mRNA for the structural proteins.
RUB replicates slowly and to relatively low titers in anumber of primary cell cultures and continuous cell linesof vertebrates, generally producing little or no cytopathiceffect (CPE) (reviewed by Frey, 1994). RUB grows tohigher titers (;107 PFU/ml), induces CPE, and formsplaques within 5–6 days in Vero cells (African greenmonkey kidney cells) without shutting down host cellmacromolecular synthesis. Viral growth kinetics in Verocells are characterized by a lag phase of ;12 h followedby a gradual raise in virus titers through 2–3 days postin-fection (p.i.) (Frey, 1994; Pugachev et al., 1997). DespiteCPE and cell death, persistent infection in Vero cells isreadily established.
In contrast to RUB, replication of Sindbis virus (SIN),1 To whom reprint requests should be addressed. Fax: (404) 651-
2509. E-mail: [email protected].
VIROLOGY 250, 359–370 (1998)ARTICLE NO. VY989395
0042-6822/98 $25.00Copyright © 1998 by Academic PressAll rights of reproduction in any form reserved.
359

the prototype member of the alphaviruses, is rapid (themaximum level of virus output is achieved by 6 h p.i.) andis accompanied by arrest of host macromolecular syn-thesis and profound CPE in practically all vertebrate cellcultures (however, not in mosquito cell cultures, in whichpersistent infection is readily established) (reviewed byStrauss and Strauss, 1994). Apoptosis triggered by SINhas been detected in various vertebrate (but not mos-quito) cell cultures and brain neurons of infected mice(Karpf and Brown, 1998; Levine et al., 1993; Lewis et al.,1996). Neuronal apoptosis and hormonal stress re-sponses have been proposed to be the key contributorsto development of fatal encephalitis caused by SIN insusceptible mice (Griffin et al., 1994; Levine et al., 1996;Lewis et al., 1996; Trgovcich et al., 1997).
Mechanisms of virus-induced apoptosis, as well asapoptosis induced by other agents, are not completelyunderstood because of the complexity of the underlyingbiochemical cascades and because not all of the partic-ipating host factors have been identified (Teodoro andBranton, 1997). SIN-induced apoptosis has been re-ported to be inhibited by Bcl-2 (Levine et al., 1993, 1996)or delayed by Ras (Joe et al., 1996) and to require NF-kBactivation (Lin et al., 1995). In addition, caspases wereimplicated in SIN-induced programmed cell death (Navaet al., 1998). Caspases are cysteine proteases involved inthe execution stage of apoptosis. Recently, identifiedcleavages by caspases of a number of cellular proteins,such as Bcl-2, protein kinases, the retinoblastoma pro-tein, cytoskeletal proteins, DFF protein, and caspaseprecursors (Cheng et al., 1997, and references therein),are believed to trigger DNA fragmentation as well asother biochemical and morphological changes duringapoptosis. In addition, a caspase-dependent DNase(CAD) was recently discovered that is located in thecytoplasm in a complex with its inhibitor ICAD.Caspase-3 cleaves ICAD, which results in activation ofCAD and its migration to the nucleus to degrade cellDNA (Enari et al., 1998; Sakahira et al., 1998).
In this study, we demonstrate that death of RUB-in-fected Vero cells also occurs due to apoptosis and thatthis apoptosis is caspase dependent. We were alsointerested in whether the ability of RUB to initiate persis-tent infection involved interference with apoptosis. How-ever, we found no evidence of suppression of apoptosisin either acutely or persistently infected cells.
RESULTS
RUB-induced cytopathology in Vero cells
Two strains of RUB were used in this study: w-Therien,which induces moderate CPE and forms opaque plaquesin Vero cells, and its clear-plaque laboratory variant,f-Therien, which is more cytopathic (Pugachev et al.,1997). In this study, infection with these viruses was doneusing a relatively high multiplicity of infection (m.o.i.) (3
PFU/cell), which typically results in initial infection of50–60% of the cells (Hemphill et al., 1988). Morphologicalchanges in the infected monolayers begin to appear at;2 days p.i., when some of the cells become roundedand then detach and float in the medium. These de-tached cells (floaters) appear shrunken in comparison toVero cells in suspension obtained by trypsinization ofmock-infected or RUB-infected monolayers. By this time,virually all of the monolayer cells are RUB antigen-posi-tive as determined by an indirect immunofluorescence.The cell rounding and detachment progresses such thatby day 5 p.i., ;40% and ;10% of the cells infected withw-Therien and f-Therien, respectively, remain in themonolayer (Pugachev at al., 1997), whereas there is prac-tically no observable cell rounding and detachment inmock-infected cultures during $10 days of incubation.
Cytopathology during the establishment of persistentinfection is as follows. During the first five weekly pas-sages of an infected culture, the cells remain subconflu-ent, and pronounced cytopathology is observed (round-ed cells in the monolayer and large numbers of detachedcells in the medium; during this period, the rate of cellkilling is obviously equilibrated by cell multiplication).After the fifth passage, the condition of the culture beginsto improve in that the monolayer reaches confluency andfewer cells are detached. By approximately the 12thpassage, cytopathology in the culture is minimal.
Chromatin fragmentation in RUB-infected Vero cells
To examine whether RUB-infected Vero cells die dueto induction of apoptosis, total cellular DNA from 105
monolayer and detached cells was extracted using anSDS-extraction protocol and analyzed in agarose gels.As can be seen in Fig. 1A, no significant DNA fragmen-tation was detected in mock-infected cells or in w-Ther-ien RUB-infected monolayers through 6 days p.i.,whereas chromatin in the detached cells was almostcompletely fragmented (Fig. 1B). A similar result wasobtained with the f-Therien strain (data not shown). Thisobservation of a DNA ladder in the floaters indicates thatRUB induces apoptosis. On the other hand, the absenceof a pronounced DNA laddering in the monolayer sug-gests that the majority of attached cells are not in the latestages of apoptosis when DNA fragmentation is induced.When an alternative, Triton X-100 extraction protocol wasused that yields only low-molecular-weight (extranucle-ar) DNA present in apoptotic cells but not in normalhealthy cells, a characteristic DNA ladder was detectedin both w-Therien and f-Therien-infected monolayer cellsas early as 2 days p.i. (and through day 4 p.i.) (Fig. 1C).No chromatin fragmentation was detected at 1 day p.i. orin mock-infected cells. The higher sensitivity in the laterexperiment was because extranuclear DNA from ;5 3106 could be loaded onto each of the lanes without
360 PUGACHEV AND FREY

overloading the gel, which is 50 times more than in theprevious experiment.
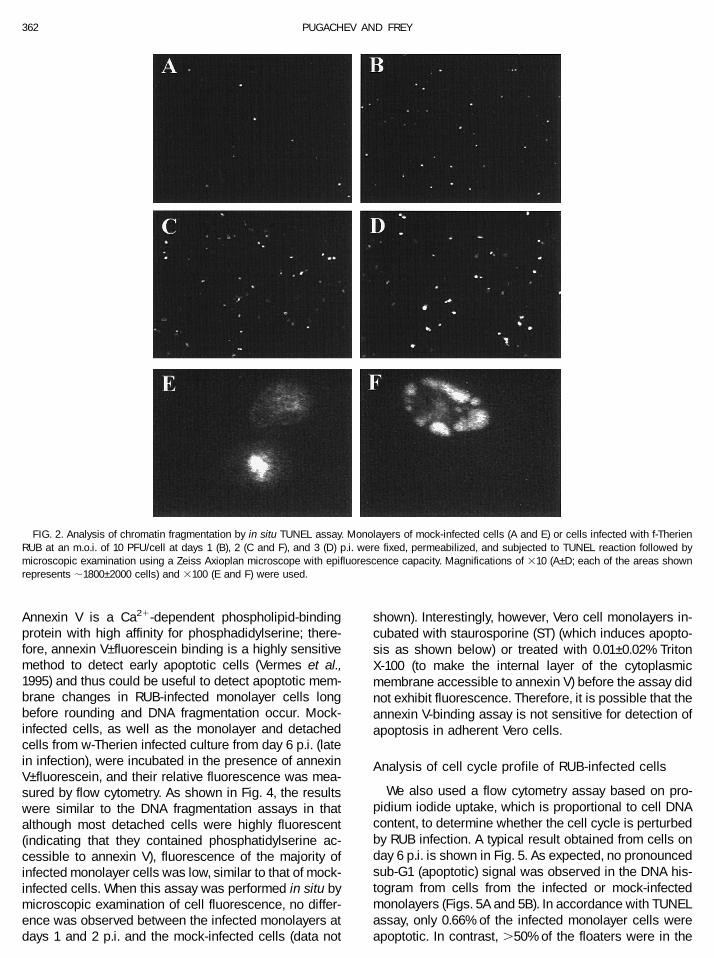
An in situ terminal deoxynucleotidyl transferase-medi-ated dUTP nick end labeling (TUNEL) assay was used todetermine how many monolayer cells are undergoingDNA fragmentation at any particular time. A few apopto-tic cells were observed in the mock-infected monolayer(Fig. 2A), whereas their number increased roughly fivetimes at 1 day p.i. to ;1% of total cells (Fig. 2B) andremained similar at later times p.i. (Figs. 2C and 2D).Interestingly, at day 1 p.i. and in the mock-infected cul-ture, the chromatin of apoptotic cells was mostly con-densed (Fig. 2E), whereas starting at say 2 p.i., cells withsegmented and condensed chromatin were observed(Fig. 2F). Costaining with an anti-E1 monoclonal antibodyrevealed that ;70% of the cells in the infected culturewere antigen-positive at 1 day p.i. (some apoptotic cellsdid not costain with the antibody), whereas virtually allcells were antigen-positive at days 2 and 3 p.i. (data notshown). Thus, the segmented condensation pattern wasunique to RUB-infected cells.
The fact that the majority of infected monolayer cellscontain intact chromatin was confirmed by flow cytom-etry. Monolayer and detached cells from RUB-infectedcultures were harvested and subjected to the TUNELassay at different times (up to 6 days) p.i., and theFITC-fluorescence (which is proportional to the extent ofDNA fragmentation in these cells) was measured. Atypical result obtained with w-Therien-infected cells isshown in Fig. 3. Fluorescence in the majority of cells inthe monolayer on day 4 p.i. (94.59%) was low (Fig. 3B)
and similar to that in mock-infected cells from day 1 p.i.(Figs. 3A and 3D), whereas most of the detached cells(86.96%) exhibited fluorescence that was ;100 timesstronger than the infected monolayer or mock-infectedcontrol (Figs. 3C and 3D). Using this assay, the propor-tion of apoptotic cells in the infected monolayer (0.32%)was somewhat lower than that in the in situ assay (;1%).This could be due to the relatively wide range in theintensity of fluorescence in apoptotic cells, which couldbe scored differently by microscopy compared with flowcytometry, or because the sample preparation for flowcytometry involves multiple centrifugations, which couldlead to selective loss of some apoptotic monolayer cells,which have an increased tendency to adhere to plasticcentrifuge tubes compared with monolayer cells.
We also examined DNA fragmentation patterns withthese methods in cultures persistently infected with thew-Therien strain. During the establishment of the persis-tent infection (passages 3 and 8 were examined), DNApatterns were similar to those observed during acuteinfection: DNA from monolayer cells was predominantlyintact, whereas DNA from detached cells was degraded(data not shown).
Annexin V-binding assay to detect early stages ofapoptosis
In early stages of apoptosis, phosphatidylserine,which is located in the inner layer of the plasma mem-brane of healthy cells, is translocated to the cell surface(outer layer of the membrane) (Vermes et al., 1995).
FIG. 1. Apoptotic DNA-laddering in RUB-infected cells. Vero cells were mock-infected (mo) or infected with the w-Therien (w-RUB) or f-Therien strain(f-RUB) of RUB at an m.o.i. of 3 PFU/cell. Total DNA from 105 monolayer (A) or detached cells (B) was extracted using a total DNA extraction protocolat indicated days p.i. and resolved by agarose gel electrophoresis. (C) Low-molecular-weight (extranuclear) DNA from ;5 3 106 monolayer cells wasextracted using an alternative Triton X-100 extraction protocol at indicated days p.i. The molecular weight markers (mw) are phage l DNA digestedwith HindIII and phage fX174 DNA digested with HaeIII loaded in the same lane (A and B) or in adjacent lanes (C).
361RUBELLA VIRUS-INDUCED APOPTOSIS
Annexin V is a Ca21-dependent phospholipid-bindingprotein with high affinity for phosphadidylserine; there-fore, annexin V–fluorescein binding is a highly sensitivemethod to detect early apoptotic cells (Vermes et al.,1995) and thus could be useful to detect apoptotic mem-brane changes in RUB-infected monolayer cells longbefore rounding and DNA fragmentation occur. Mock-infected cells, as well as the monolayer and detachedcells from w-Therien infected culture from day 6 p.i. (latein infection), were incubated in the presence of annexinV–fluorescein, and their relative fluorescence was mea-sured by flow cytometry. As shown in Fig. 4, the resultswere similar to the DNA fragmentation assays in thatalthough most detached cells were highly fluorescent(indicating that they contained phosphatidylserine ac-cessible to annexin V), fluorescence of the majority ofinfected monolayer cells was low, similar to that of mock-infected cells. When this assay was performed in situ bymicroscopic examination of cell fluorescence, no differ-ence was observed between the infected monolayers atdays 1 and 2 p.i. and the mock-infected cells (data not
shown). Interestingly, however, Vero cell monolayers in-cubated with staurosporine (ST) (which induces apopto-sis as shown below) or treated with 0.01–0.02% TritonX-100 (to make the internal layer of the cytoplasmicmembrane accessible to annexin V) before the assay didnot exhibit fluorescence. Therefore, it is possible that theannexin V-binding assay is not sensitive for detection ofapoptosis in adherent Vero cells.
Analysis of cell cycle profile of RUB-infected cells
We also used a flow cytometry assay based on pro-pidium iodide uptake, which is proportional to cell DNAcontent, to determine whether the cell cycle is perturbedby RUB infection. A typical result obtained from cells onday 6 p.i. is shown in Fig. 5. As expected, no pronouncedsub-G1 (apoptotic) signal was observed in the DNA his-togram from cells from the infected or mock-infectedmonolayers (Figs. 5A and 5B). In accordance with TUNELassay, only 0.66% of the infected monolayer cells wereapoptotic. In contrast, .50% of the floaters were in the
FIG. 2. Analysis of chromatin fragmentation by in situ TUNEL assay. Monolayers of mock-infected cells (A and E) or cells infected with f-TherienRUB at an m.o.i. of 10 PFU/cell at days 1 (B), 2 (C and F), and 3 (D) p.i. were fixed, permeabilized, and subjected to TUNEL reaction followed bymicroscopic examination using a Zeiss Axioplan microscope with epifluorescence capacity. Magnifications of 310 (A–D; each of the areas shownrepresents ;1800–2000 cells) and 3100 (E and F) were used.
362 PUGACHEV AND FREY

apoptotic area of the histogram (Fig. 5C). Interestingly,relative to the mock-infected cells, a slight shift in theproportion of cells in the S and G2/M phases of the cellcycle was noticed in the infected monolayer. Specifically,75.52%, 14.43%, and 8.93% of the cells in the infectedmonolayer were in G0/G1, S, and G2/M phases, respec-tively, whereas percentages of mock-infected cells inthese phases were 92.04%, 2.89%, and 4.86%, respec-tively. This was most probably due to the mock-infectedmonolayer, but not the RUB-infected monolayer, becom-ing overconfluent by the time of the analysis (see below).
Analysis of floaters
To determine whether the detached cells in the me-dium were alive or dead, we first used a propidiumiodide (PI)-permeability assay in which PI fluorescence ismeasured by flow cytometery (live cells are not perme-
able). As can be seen in the histogram in Fig. 6, at 6 daysp.i. with w-Therien, the majority of cells in the monolayer(79.88%) exhibited a low intensity of fluorescence andthus were alive. Interestingly, a slight shift upward in theposition of the live cell peak in comparison with themock-infected cells was observed. The reason for thisshift is unknown but may be due to changes in thepermeability of the plasma membrane in infected cells.In contrast, the majority of detached cells (94.95%) werehighly fluorescent, and thus the floaters were predomi-nantly dead. These results were confirmed by a standardtrypan blue exclusion test: ;90% of mock-infected cellsand ;80% of the cells in the RUB-infected monolayerexcluded trypan blue, whereas 98% of the RUB-infecteddetached cells were stained with trypan blue and thuswere dead. In Fig. 6, the detached cells were harvestedafter a 24-h period (the medium was changed at 5 days
FIG. 3. Analysis of chromatin fragmentation by TUNEL assay followed by flow cytometry. (A) Fluorescence of mock-infected cells harvested at 1day p.i. (B and C) Fluorescence of w-Therien RUB-infected monolayer and detached cells, respectively, harvested at 4 days p.i. (D) Histograms in A–Ccombined. After TUNEL reactions, the relative FITC fluorescence of 10,000 cells were assayed using a flow cytometer; cell counts are plotted as afunction of intensity of fluorescence (FL1-H). The percentages of cells exhibiting low (M1) and high (M2) relative fluorescence are given.
363RUBELLA VIRUS-INDUCED APOPTOSIS

p.i. and floaters were harvested on day 6). Detachedcells produced during as short an interval as 1 h werealso examined with a similar result that they were pre-domonantly dead (data not shown), indicating that celldeath occurs before or concomitantly with detachment.
We next examined whether cells remain capable ofproduction of virus after detachment. For this purpose,a monolayer of f-Therien-infected cells at day 3 p.i. anddetached cells harvested from this culture during a 1-hinterval were washed with PBS and incubated ingrowth medium at 35°C for 2 h. Cells were counted,titers of virus in the media before and after incubationwere determined, and rates of virus production werethen calculated. Adherent monolayer cells produced;220 times more PFU (2.27 PFU/cell/h) than detachedcells (0.01 PFU/cell/h). In addition, a relative ratio ofvirus trapped inside these cells was determined bymultiple freeze–thaw cycles of cell suspensions fol-lowed by determination of virus titers in the clarifiedmedia. Roughly 30 times more virus was releasedusing this procedure from the monolayer cells (1.85PFU/cell) compared with detached cells (0.064 PFU/cell). Thus, virus production by detached cells is sig-nificantly reduced compared with the infected mono-layer cells, and these cells are not a repository ofsignificant amounts of trapped virus.
Inhibition of RUB-induced apoptosis by z-VAD
Caspases play a crucial role in the execution stage ofapoptosis, and apoptosis can be inhibited by pseudo-substrates of these proteases. A chemically modifiedoligopeptide called z-VAD-fmk [z-Val-Ala-Asp(OMe)-CH2F]has been shown to efficiently and specifically inhibitapoptosis, including SIN-induced apoptosis (Nava etal., 1998; Zhivotovsky et al., 1995). To learn whetherz-VAD can inhibit the cytopathology observed duringRUB infection, f-Therien RUB-infected and mock-in-fected Vero cells were incubated in the presence andabsence of z-VAD (the f-Therien strain was chosen forthis experiment because of its greater cytopathogenic-ity). The mock-infected cells appeared normal through-out the experiment, both in the presence and in theabsence of z-VAD. RUB-specific CPE was observablein the f-Therien-infected culture without z-VAD on day2 after infection and became profound on day 3.5. Incontrast, the infected culture incubated with z-VADshowed little cytopathology on day 2, and by day 3.5,the numbers of deformed and rounded cells in themonolayer and detached cells in the medium werenoticeably lower than in the culture incubated withoutz-VAD. On day 3.5, the numbers of attached and de-tached cells in these cultures were determined (Table1). In the untreated f-Therien-infected culture, 43.8% of
FIG. 4. Result of annexin V-binding assay. Trypsinized mock-infected (shadowed) and w-Therien-infected monolayer cells at day 6 p.i. (thick line)and detached cells from the infected culture (thin line) were incubated in the presence of annexin V-fluorescein. Relative FITC fluorescence of 10,000cells was measured by flow cytometry and plotted versus cell counts ( y axis).
364 PUGACHEV AND FREY

the total cells were detached, whereas in the infectedculture incubated with z-VAD, floaters constituted only7.5% of all cells. z-VAD did not affect virus replicationbecause RUB titers in the media from the two infectedcultures were identical. These results confirm thatVero cells infected with RUB die due to apoptosis andthat caspases are involved in the process. Interest-ingly, the total number of the RUB-infected cells in themonolayer and in the medium combined (incubatedboth with and without z-VAD) was ;2.5 times less thanthe number of cells in the mock-infected culture, indi-cating that RUB reduces the rate of Vero cell multipli-cation. z-VAD and/or DMSO (z-VAD solvent) also some-what slowed the growth of mock-infected Vero cells(1.65 times fewer cells were present in the treatedcompared with the untreated, mock-infected culture).
ST-induced apoptosis in RUB-infected cells
To determine whether the slow and asynchronousinduction of apoptosis in Vero cells after RUB infection orthe ability of RUB to initiate a persistent infection wasdue to a virus-induced suppression of apoptosis, weattempted to find a proapoptotic agent that would mimicRUB-induced apoptosis in Vero cells but induce it moreefficiently. ST was found to be the most suitable for thispurpose. The addition of ST to Vero cells resulted in;30–80% of the cells becoming detached within 15 h andfloating in the medium, depending on the freshness ofthe reagent (ST is unstable and its activity diminishedwith multiple freeze–thaw cycles). Interestingly, the effec-tiveness of ST was also dependent on the age of theculture and/or the batch of the reagent (experimentsdescribed here were done on Vero cells that had been
FIG. 5. Cell cycle analysis. Shown are DNA content histograms of mock-infected (A) and w-Therien-infected monolayer cells (B) analyzed at 6 daysp.i. and floaters (C) harvested during a 24-h interval (day 4–5) from the w-Therien-infected culture. Fixed and permeabilized cells were stained withpropidium iodide (PI), followed by measurement of relative PI-fluorescences (FL2-H), which is proportional to DNA content, using a flow cytometer.Signals from 5000 singlets were plotted in each histogram. Proportions of cells in the different phases of the cell cycle are given. The subG1 (M1)area is composed of apoptotic cells.
365RUBELLA VIRUS-INDUCED APOPTOSIS

propagated in our laboratory for ;1.5 years, whereasnew Vero cells obtained from the American Type CultureCollection did not produce as many floaters whentreated with a different batch of ST). In contrast to ST,etoposide and mitomycin C, as well as incubation of thecells for #4 days in serum-free medium, did not inducepronounced cytopathology, whereas 1-methyl-3-nitro-1-nitrosoguanidine appeared overly toxic in that the wholemonolayer detached without breaking into individualcells within a 14-h incubation period. Although the cyto-pathology induced by ST resembled RUB-induced CPE,
ST caused significant blebbing of cells, making countingof cells (especially floaters) somewhat difficult (to mini-mize the contribution of blebs to cell counts, we did notcount particles that were arbitrarily too small). As foundin RUB-infected cells, the chromatin of detached cellscollected after incubation with ST was fragmented aswas shown by their strong fluorescence in TUNEL assay(Fig. 7A) and by the presence of a ladder in extractedDNA (data not shown). Although chromatin fragmenta-tion in the ST-treated cells that remained in the mono-layer was almost undetectable in terms of DNA laddering
FIG. 6. Cell viability. Mock-infected cells (shadowed) and w-Therien-infected monolayer cells (thick line) from day 6 p.i., and detached cells fromthe w-Therien-infected culture collected during day 6 p.i. (thin line) were stained with propidium iodide (PI) without prior fixation and permeabilization.Relative PI fluorescences (FL3-H) of 10,000 cells were measured by flow cytometry and plotted versus cell counts ( y axis). Percentages of live cellsthat exclude PI and thus exhibit low fluorescence (M1 area of the histogram) and dead cells exhibiting high fluorescence (M2 area) are given. Mockindicates mock-infected cells; ML, monolayer cells; FL, floaters.
TABLE 1
Inhibition of Cytopathic Effect of RUB by z-VAD
CultureCells in monolayer a
(3105)Floatersa
(3105)Total cells
(3105)Floaters
(%)RUB titer b
(PFU/ml)
RUB 4.18 3.26 7.44 43.8% 1.05 3 107
RUB 1 VAD 7.46 0.61 8.07 7.5% 1.03 3 107
Mock 1 VAD 12.67 0 12.67 0% —Mock 21.0 0 21.0 0% —
Note. Vero cells in 35-mm plates were infected (RUB) or mock-infected (mock) with f-Therien RUB and incubated in the presence (1VAD) or absenceof z-VAD. Cells in the monolayers and in the media were counted at 3.5 days postinfection.
a Absolute numbers of cells; mean values of two counts are given.b Mean values of titers obtained in two independent experiments are given.
366 PUGACHEV AND FREY

(data not shown), there was a pronounced shift toward astronger fluorescence of these cells in TUNEL assays incomparison with the monolayer cells in RUB-infectedcultures (compare Figs. 7A and 3). In contrast to RUBinfection, the majority (;70%) of the detached cells gen-erated by ST treatment excluded trypan blue and PI andthus were viable (data not shown). ST-induced apoptosiswas also found to be caspase dependent because co-incubation of cells with z-VAD and ST reduced theamount of detached cells by ;10-fold.
ST was added to w-Therien RUB-infected cells at day2 p.i. and to the persistently infected culture at the sev-enth passage. Interestingly, ST treatment resulted in amore profound CPE (90% and 96% of the total cells in theinfected and persistently infected cultures, respectively,were floating in the medium) compared with the parallelinfected and persistently infected cultures (10% and 11%floaters, respectively), as well as uninfected cells incu-bated in the presence of the reagent (37% floaters) (Fig.7B). RUB therefore does not cause an inhibitory effect onST-induced apoptosis, and the two interact cumulativelyto induce cell detachment.
DISCUSSION
Apoptosis is a natural, genetically controlled processof cell self-destruction that is vital in the development ofan organism and in its life-long maintenance free fromaberrant cells that can be generated by various factorssuch as radiation, oncogenesis, and viral infections. Al-though the replication of many viruses has been shown
to lead to apoptosis and some viruses are believed toactively induce apoptosis (e.g., blebbing of apoptoticcells is speculated to facilitate dissemination of nonen-veloped, nonlytic viruses), other viruses have evolvedmechanisms to suppress apoptosis via interference withapoptotic cascades by certain virus-specified proteins toprolong the life of the host cell (Razvi and Welsh, 1995;Shen and Shenk, 1995; Teodoro and Branton, 1997).
In this study, we demonstrated that cytopathology andcell death in Vero cells, a line of culture cells in whichthese are induced by RUB, are due to the induction ofapoptosis. The chromatin of detached cells (floaters) thataccumulated in the medium of infected cultures wasfragmented in patterns specific for apoptosis as mea-sured by two assays; these cells were also predomi-nantly dead. In contrast to detached cells, cells withchromatin fragmentation were at minimal levels in themonolayers (;1% of total cells by in situ TUNEL assay).Thus the majority of cells in the monolayers containintact chromatin. Our experiments to detect cells under-going early stages of apoptosis by using an annexinV-binding assay that detects membrane changes gave asimilar profile. However, the annexin V-binding assayappeared to be insensitive on attached Vero cells (an-nexin V-binding assay has been successfully usedmostly on suspension cultures). Because our assaysthus detected only the terminal stages of apoptosis, wedo not know the time course of RUB-induced apoptosis.Presumably, the process is initiated in adherent cells andculminates with cell rounding, detachment from the
FIG. 7. Effects of staurosporine (ST). (A) Induction of apoptosis in Vero cells by ST as detected by TUNEL assay. Cells were incubated for 15 h inthe presence or absence of 10 mM ST. Mock-treated cells (shadowed), ST-treated monolayer cells (thick line), and detached cells from the treatedculture (thin line) were subjected to TUNEL reaction and analyzed by flow cytometry. Signals from 10,000 cells in each sample were measured andplotted. (B) Effects of ST on RUB-infected cells. The diagram shows total numbers of monolayer cells (m) and floaters (f) in the following parallelcultures (in 35-mm plates): mock-infected cells (mock), RUB-infected cells at 63 h p.i. (RUB), and persistently infected cells incubated for 63 h afterthe seventh passage (Pers) in the absence or presence during the last 15 h of incubation of ST (1ST).
367RUBELLA VIRUS-INDUCED APOPTOSIS

monolayer, and cell death. The detached cells wereneither significant repositories of virus nor sources ofvirus shedding.
We found that RUB-induced cytopathology could bedramatically reduced by the addition of z-VAD, a specificpseudosubstrate inhibitor of caspases (Zhivotovsky etal., 1995), without affecting virus replication, indicatingthat RUB-induced apoptosis is caspase dependent. Arecent report demonstrated that SIN-induced apoptosisis also caspase dependent (Nava et al., 1998). Our find-ing thus was not surprising and instead served as evi-dence supporting our conclusion that RUB can causeapoptosis because to our knowledge, no caspase-inde-pendent apoptotic pathway has been described.
Because at any time after 1 day p.i., only 1% of themonolayer is demonstrably apoptotic and floaters areproduced continuously from 2 days p.i. onward while70% and 100% of the adherent cells are RUB antigenpositive at 1 and 2 days p.i., respectively, apoptosis isinduced asynchronously. Other viruses (albeit large DNAviruses) have been shown to be capable of suppressingapoptosis, and such capability on the part of RUB couldexplain the asynchronicity of apoptosis and the ability ofRUB to initiate persistent infection in Vero cells. Wetherefore tested the ability of RUB to inhibit apoptosisinduced by a different agent. For this purpose, ST, apotent protein kinase and topoisomerase II inhibitor(Lassota et al., 1996; Seynaeve et al., 1994; Wang et al.,1996), was found to induce profound apoptosis in Verocells. Although generally resembling RUB-induced cyto-pathology, ST treatment produced significant blebbing ofcells, induced detectable fragmentation in adherentcells, and generated detached cells that were predomi-nantly alive. The addition of ST to RUB-infected cells andto a persistently infected culture resulted in a moreprofound CPE that was even greater than the cytopathol-ogy induced by ST treatment of mock-infected Vero cells.Therefore, RUB did not inhibit ST-induced apoptosis, andthe two agents appeared to work cumulatively to pro-duce more profound apoptotic effect than induced byeither alone. Considering this and the differences be-tween ST- and RUB-induced apoptosis, it is possible,however, that RUB and ST use different upstream apop-totic pathways; in such a case, a possible suppressiveeffect of RUB on apoptosis could be masked.
Similar results in terms of detectable apoptosis beingmostly confined to detached cells were obtained withtwo strains of RUB, w-Therien and f-Therien, which differin plaque morphology and cell killing; thus these strainsdo not kill cells via different mechanisms and simplydiffer in ability to induce apoptosis. The results of thisstudy show that apoptosis can be quantified by simplycounting floaters, and this simple assay will be of greatuse in our ongoing studies to more closely define cellkilling determinants with these two strains. Using thesestrains, we mapped the genetic determinants of cell
killing to the nonstructural proteins (Pugachev et al.,1997), and thus the inducer of apoptosis may be one ofthe nonstructural proteins (these two strains differ in theamount of nonstructural proteins produced) or some vi-rus function mediated by the nonstructural proteins (Pu-gachev and Frey, 1998; Pugachev et al., 1997). Becausevirus macromolecular synthesis is greater in Vero cellsthan in other cell lines, the lack of apoptosis in theseother lines may be due to the lack of sufficient accumu-lation of the nonstructural proteins. In terms of persistentinfection, the eventual reduction in cytopathology ob-served after ;12 weeks after initial infection may in partbe explained by changes in the virus itself, specifically,by selection of virus population that does not efficientlyinduce apoptosis (although cells resistant to apoptosismay also be selected). In this regard, we demonstratedpreviously that by 68–76 days postinitiation, RUB-specificRNAs detected in persistently infected cells are mostlydefective interfering RNAs, whereas the full-lengthgenomic RNA (the mRNA for the nonstructural proteins)is no longer detectable, and the total amount of RUB-specific RNA has declined (Frey and Hemphill, 1988).
Finally, the only medical manifestation of RUB infectionin which RUB cytopathology has been implicated inpathogenesis is congenital infection (reviewed by Frey,1994). Although necrotic lesions that could be caused byvirus-induced apoptosis are apparent, particularly in thebrain, the hallmark of congenital infection is underdevel-opment. In infected fetal tissues, only one of 104 or 105
cells is infected, and it has been proposed that virusinfection early in development adversely affects undiffer-entiated precursor cells, leading to underdevelopment.RUB has been shown to have an inhibitory effect ondivision in primary fetal cell cultures, and we also ob-served an inhibitory effect on cell division because thereis a 3-fold reduction in the total cell numbers in RUB-infected cultures compared with parallel mock-infectedcultures. The results of this study show that RUB caninduce apoptosis, and it is possible that during congen-ital infection, RUB destroys precursor cells by inductionof apoptosis.
MATERIALS AND METHODS
Cells, viruses, and reagents
Vero cells obtained from the American Type CultureCollection (ATCC CCL 81) were grown in DMEM (GIBCOBRL) containing 5% FBS and gentamicin (10 mg/ml) at35°C under 5% CO2. The w-Therien and f-Therien strainsof RUB were propagated as described previously (Hemp-hill et al., 1988; Pugachev et al., 1997; Wang et al., 1994).ST (Biomol Research Laboratories, Plymouth Meeting,PA) was dissolved in DMSO to a concentration of 5 mM(5003 stock solution). z-VAD-fmk [z-Val-Ala-Asp(OMe)-CH2F; Enzyme Systems Products, Dublin, CA] was dis-solved in DMSO to a concentration of 20 mM (5003
368 PUGACHEV AND FREY

stock solution). Etoposide (Sigma) and 1-methyl-3-nitro-1-nitrosoguanidine (Aldrich) solutions were made inDMSO (both 10 mg/ml), and mitomycin C (Sigma) wasdissolved at a concentration of 2 mg/ml in water. Theseconcentrated stock solutions were diluted directly intomedium immediately before the addition to cells.
Electrophoretic DNA fragmentation assays
Subconfluent Vero cell monolayers were mock-infected or infected with w-Therien or f-Therien RUB at anm.o.i. of 3 PFU/cell. At indicated times p.i., media werecollected, and cells in the monolayers were trypsinized.Detached cells in the media (floaters) and cells from thetrypsinized suspensions were collected by low-speedcentrifugation (400g, 5 min, 12°C) and resuspended at106–107 cells/ml in DMEM for immediate use or inDMEM containing 5% FBS and 10% DMSO for storage at270°C until future use. Cell concentrations in thesesuspensions were determined by counting using a he-macytometer. Total cellular DNA was extracted by theSDS-extraction procedure essentially as described byJeurissen et al. (1992). Briefly, 105 cells were lysed inSDS-extraction buffer [20 mM Tris–HCl, pH 7.5, 2 mMEDTA, 0.2% SDS, 0.6 mg/ml proteinase K (Boehringer-Mannheim)], and the lysates were incubated for 1 h at37°C. The samples were then extracted twice with phe-nol and once with chloroform, followed by ethanol pre-cipitation. The DNA was resuspended in TE buffer (10mM Tris–HCl, pH 8.0, 1 mM EDTA) containing 0.5 mg/mlRNase I and incubated at room temperature for 10 minbefore electrophoresis in 1% agarose gels containingethidium bromide.
Alternatively, extranuclear/low-molecular-weight DNAwas extracted by modification of a procedure describedby Duke and Cohen (1994). Monolayers of RUB-infectedand mock-infected cells grown in 60-mm plates weredirectly lysed in TE buffer containing 0.2% Triton X-100.Intact chromatin/nuclei were removed by centrifugationof the lysates for 10 min at 13,000g, and fragmented DNAwas recovered from the supernatants by trichloroaceticacid (TCA) precipitation and dissolved in TE buffer. Be-fore agarose gel electrophoresis, samples were phenol–chloroform extracted, ethanol precipitated, and redis-solved in TE containing RNase I.
TUNEL assays
TUNEL assays were done using a Fluorescein in situCell Death Detection Kit (Boehringer-Mannheim) accord-ing to the manufacturer’s procedure. Briefly, cells har-vested as described above or monolayers grown ineight-well chamber slides (Nunc) were fixed in fresh 4%formalin dissolved in PBS, permeabilized in 0.1% TritonX-100 in 0.1% sodium citrate, and labeled in a TUNELreaction mixture. Between each of these steps, monolay-ers on the slides were rinsed twice with PBS, and cells
in suspensions were washed twice with PBS containing1% BSA. Fluorescein (FITC) emission of cells was exam-ined microscopically (on slides) using a Zeiss Axioplanmicroscope with epifluorescence capacity or by FACSanalysis using a Becton Dickinson FACSCalibur flowcytometer (equipped with 488-nm, 16-mW argon laser)under CELLQuest software (Becton Dickinson).
Annexin V-fluorescein binding assays
An Annexin-V-FLUOS Staining kit (Boehringer-Mann-heim) was used for these analyses. For flow sytometry,106 detached cells or trypsinized monolayer cells werewashed with PBS and resuspended in 200 ml of a Ca21-containing staining solution (provided in the kit) supple-mented with 4 ml of annexin V-fluorescein reagent. Flu-orescein emission of cells in the samples was examinedusing a Becton Dickinson FACSCalibur flow cytometer.For analyses in situ, cell monolayers grown in eight-wellchamber slides (Nunc) were washed with PBS and incu-bated for 10–15 min with 100 ml of the staining solutioncontaining annexin V-fluorescein, followed by micro-scopic examination using a Zeiss Axioplan microscopewith epifluorescence capacity.
DNA content analysis
Sample preparation was essentially as described byOrmerod (1994). Cells suspended in PBS were mixedwith five volumes of cold 70% ethanol and incubated onice for 1 h. The fixed cells were harvested by low-speedcentrifugation, resuspended in PBS, and incubated at4°C overnight to allow low-molecular-weight DNA dif-fuse from cells. RNase I and PI were added to 100 and 50mg/ml concentrations, respectively. Measurements ofDNA fluorescence in individual cells (cell aggregateswere excluded by appropriate gating) were done using aBecton Dickinson FACSCalibur flow cytometer accordingto the manufacturer’s procedures.
Cell viability tests
Viability tests using flow cytometry were done accord-ing to a Becton Dickinson protocol. Suspensions of 0.5 3106 cells in 0.5 ml of PBS were mixed with 10 ml of a PIsolution in PBS (5 mg/ml) and directly examined using aFACSCalibur flow cytometer. Alternatively, a standardtrypan blue exclusion test (Strober, 1994) was used.
ACKNOWLEDGMENTS
This study was supported by U.S. Public Health Service Grant AI-21389 from NIAID. A Becton Dickinson FACSCalibur flow cytometerwas purchased with a grant from the Georgia Research Alliance.
REFERENCES
Cheng, E. H.-Y., Kirsch, D. G., Clem, R. J., Ravi, R., Kastan, M. B., Bedi, A.,Ueno, K., and Hardwick, J. M. (1997). Conversion of Bcl-2 to Bax-likedeath effector by caspases. Science 278, 1966–1968.
369RUBELLA VIRUS-INDUCED APOPTOSIS

Duke, R. C., and Cohen, J. J. (1994). Morphological and biochemicalassays of apoptosis. In “Current Protocols in Immunology” (J. E.Coligan, A. M. Kruisbeek, et al., Eds.), pp. 3.17.1–3.17.16. Wiley-Greene,New York.
Enari, M., Sakahira, H., Yokoyama, H., Okawa, K., Iwamamatsu, A., andNagata, S. (1998). A caspase-activated DNase that degrades DNAduring apoptosis, and its inhibitor ICAD. Nature 391, 43–50.
Frey, T. K. (1994). Molecular biology of rubella virus. Adv. Virus Res. 44,69–160.
Frey, T. K., and Hemphill, M. L. (1988). Generation of defective-interfer-ing particles by rubella virus in Vero cells. Virology 164, 22–29.
Griffin, D. E., Levine, B., Tyor, W. R., Tucker, P. C., and Hardwick, J. M.(1994). Age-dependent susceptibility to fatal encephalitis: alphavirusinfection of neurons. Arch. Virol. 9, 31–39.
Hemphill, M. L., Forng, R.-Y., Abernathy, E. S., and Frey, T. K. (1988). Timecourse of virus-specific macromolecular synthesis during rubellavirus infection in Vero cells. Virology 162, 65–75.
Jeurissen, S. H. M., Wagenaar, F., Pol, J. M. A., van der Eb, A. J., andNoteborn, M. H. M. (1992). Chicken anemia virus causes apoptosis ofthymocytes after in vivo infection and cell lines after in vitro infection.J. Virol. 66, 7383–7388.
Joe, A. K., Ferrari, G., Jiang, H. H., Liang, X. H., and Levine, B. (1996).Dominant inhibitory Ras delays Sindbis virus-induced apoptosis inneuronal cells. J. Virol. 70, 7744–7751.
Karpf, A. R., and Brown, D. T. (1998). Comparison of Sindbis virus-induced pathology in mosquito and vertebrate cell cultures. Virology240, 193–201.
Lassota, P., Singh, G., and Kramer, R. (1996). Mechanism of topoisom-erase II inhibition by staurosporine and other protein kinase inhibi-tors J. Biol. Chem. 271, 26418–26423.
Levine, B., Goldman, J. E., Jiang, H. H., Griffin, D. E., and Hardwick, J. M.(1996). Bcl-2 protects mice against fatal alphavirus encephalitis.Proc. Natl. Acad. Sci. USA 93, 4810–4815.
Levine, B., Huang, Q., Isaacs, J. T., Reed, J. C., Griffin, D. E., andHardwick, J. M. (1993). Conversion of lytic to persistent alphavirusinfection by the bcl-2 cellular oncogene. Nature (Lond) 361, 739–742.
Lewis, J., Wesselingh, S. L., Griffin, D. E., and Hardwick, J. M. (1996).Alphavirus-induced apoptosis in mouse brains correlates with neu-rovirulence. J. Virol. 70, 1828–1835.
Lin, K.-I., Lee, S.-H., Narayanan, R., Baraban, J. M., Hardwick, J. H., andRatan, R. R. (1995). Thiol agents and bcl-2 identify an alphavirus-induced apoptotic pathway that requires activation of the transcrip-tion factor NFkB. J. Cell Biol. 131, 1149–1161.
Nava, V. E., Rosen, A., Veliuona, M. A., Clem, R. J., Levine, B., andHardwick, J. M. (1998). Sindbis virus induces apoptosis through acaspase-dependent, CrmA-sensitive pathway. J. Virol. 72, 452–459.
Ormerod, M. G. (1994). Analysis of DNA: general methods. In “Flow
Cytometry: A Practical Approach” (M. G. Ormerod, Ed.), pp. 119–135.Oxford University Press, New York.
Pugachev, K. V., Abernathy, E. S., and Frey, T. K. (1997). Improvement ofthe specific infectivity of the rubella virus (RUB) infectious clone:determinants of cytopathogenicity induced by RUB map to the non-structural proteins. J. Virol. 71, 562–568.
Pugachev, K. V., and Frey. T. K. (1998). Effects of defined mutations inthe 59 nontranslated region of rubella virus genomic RNA on virusviability and macromolecule synthesis. J. Virol. 72, 641–650.
Razvi, E. S., and Welsh, R. M. (1995). Apoptosis in viral infections. Adv.Virus Res. 45, 1–60.
Sakahira, H., Enari, M., and Nagata, S. (1998). Cleavage of CAD inhibi-tod in CAD activation and DNA degradation during apoptosis. Nature391, 96–99.
Seynaeve, C. M., Kazanietz, M. G., Blumberg, P. M., Sausville, E. A., andWorland, P. J. (1994). Differential inhibition of protein kinase Cisozymes by UCN-01, a staurosporine analogue. Mol. Pharmacol. 45,1207–1214.
Shen, Y., and Shenk, T. (1995). Viruses and apoptosis. Curr. Opin. Genet.Dev. 5, 105–111.
Strauss, J. H., and Strauss, E. G. (1994). The alphaviruses: gene expres-sion, replication, and evolution. Microbiol. Rev. 58, 491–562.
Strober, W. (1994). Trypan blue exclusion test of cell viability. In “CurrentProtocols in Immunology” (J. E. Coligan, A. M. Kruisbeek, et al., Eds.),pp. A.3.3–A.3.4. Wiley-Greene, New York.
Teodoro, J. G., and Branton, P. E. (1997). Regulation of apoptosis by viralgene products. J. Virol. 71, 1739–1746.
Trgovcich, J., Ryman, K., Extrom, R., Eldridge, J. C., Aronson, J. F., andJohnston, R. E. (1997). Sindbis virus infection of neonatal mice resultsin a severe stress response. Virology 227, 234–238.
Vermes, I., Haanen, C., Steffens-Nakken, H., and Reutelingsper-ger, C. (1995). A novel assay for apoptosis: flow cytometric detection ofphosphatidylserine expression on early apoptotic cells using fluores-cein labelled annexin V. J. Immunol. Methods 184, 39–51.
Wang, C. Y., Dominguez, G., and Frey, T. K. (1994). Construction ofrubella virus genome-length cDNA clones and synthesis of infec-tious RNA transcripts. J. Virol. 68, 3550–3557.
Wang, X., Zelenski, N. G., Yang, J., Sakai, J., Brown, M. S, andGoldstein, J. L. (1996). Cleavage of sterol regulatory elementbinding proteins (SREBPs) by CPP32 during apoptosis. EMBO J.15, 1012–1020.
Wolinsky, J. S. (1996). Rubella. In “Virology” (B. N. Fields, D. M. Knipe, etal., Eds.), 3rd Ed., pp. 899–929. Lippincott-Raven Publishers, Phila-delphia.
Zhivotovsky, B., Gahm, A., Ankarcrona, M., Nicotera, P., and Orrenius, S.(1995). Multiple proteases are involved in thymocyte apoptosis. Exp.Cell Res. 221, 404–412.
370 PUGACHEV AND FREY