role of iron and iron chelation therapy in oxygen free radical mediated tissue injury in an...
TRANSCRIPT

Pergamon Comp. hnmun. Microbiol. infect. Dis. Vol. 20, No. 4, pp. 299-307, 1997
Published by Elsevier Science Ltd. All rights reserved Printed in Great Britain
PI I : S0147-9571(97)00001-5 0147-9571/97 $17.00 + 0.00
R O L E O F I R O N A N D I R O N C H E L A T I O N T H E R A P Y I N
O X Y G E N F R E E R A D I C A L M E D I A T E D T I S S U E
I N J U R Y I N A N A S C E N D I N G M O U S E
M O D E L O F C H R O N I C P Y E L O N E P H R I T I S
R. G U P T A , I* S. G U P T A , 2 K. J O S H I 3 and N . K . G A N G U L Y 2
~Government Medical College, Department of Microbiology, Chandigarh, India, -'PGIMER, Department of Experimental Medicine, Chandigarh, India and 3Department of Pathology,
Chandigarh, India
(Received for publication 12 January 1997)
Abstract--The contribution of iron towards the free radical generation leading to renal tissue damage was assessed using a non-obstructive ascending mouse model for chronic pyelonephritis. The parameters studied include luminol dependent chemiluminescence (LDCL), histopathology and some biochemical investigations. We found that iron enhanced the renal tissue damage and led to renal scarring, an end point in chronic renal inflammation, irrespective of the bacterial strain studied. In addition a role of iron chelation therapy as a treatment for chronic renal inflammation is also suggested. Published by Elsevier Science Ltd
Key words: pyelonephritis, ascending route, reactive oxygen species, iron chelation, iron overload, lipid peroxidation, LDCL, enzyme.
I N T R O D U C T I O N
Urinary tract infections range in severity from acute symptomatic pyelonephritis to chronic pyelonephritis and have the potential to lead to renal failure and death [1]. Despite the high prevalence of the infection, and years of study by many investigators, factors playing consequential roles in pathogenesis of chronic pyelonephritis have not yet been identified. A positive correlation, between the extent of free radical mediated renal tissue damage during the acute pyelonephritis and chronic renal scarring has been shown[2]. Further, several transition metal salts have been shown to play very important roles in the generation of the free radicals. In terms of the possibility of free radical generation in vivo, the most important transition metal is iron[3]. Iron facilitates the generation of free radicals either via Fenton reaction or via Haber Weiss reaction. Both the reactions account for part of the damage that is caused to living cells by excessive generation of reactive oxygen species (ROS). If no catalytic iron ions are available then ROS at physiological concentration may have limiting damaging effects [4].
This study was therefore designed to assess the contribution of iron in the pathogenesis of chronic pyelonephritis in an ascending non obstructive mouse model.
*To whom all correspondence should be addressed at: 144, NAC, Shivalik Enclave, Manimajra, CHAND1GARH, 160 101, INDIA.
299

300 R. Gupta et al.
MATERIAL AND METHODS
Bacterial strains
Two strains of Escherichia coli were used for the study: (i) strain 31-B, a nalidixic acid resistant derivative of strain TN 675, this strain produces type-I pili and is uropathogenic; and (ii) strain BH-5 is mutant defective in the ability to produce type-I pili. Both these strains were kindly supplied by Takeda Chemical Industries Ltd, Japan.
Animal model
Female BALB/c mice (Central Animal House, Punjab University, Chandigarh, India) weighing 20-30 g were used for the study. They were fed on standard antibiotic free synthetic feed (JDB Agencies Private Ltd., India). Mice free bacteria as confirmed by urine culture were selected for the study.
Chronic pyelonephritis and renal scarring was established in mice via ascending route[4]. The animals were infected intrauretherally using a soft intermediate non-radio-opaque polyethylene catheter (Clay Adams, U.S.A.). The inoculum (0.1 ml, l0 s cfu/ml) was injected slowly into the bladder to avoid leakage. To promote persistent chronic inflammation and renal scarring the mice were reinfected twice in the same way 2 weeks and 16 weeks after the first infection. Groups of mice (n = 6) infected with either strain of E. coli, were killed by employing cervical dislocation 4 weeks, 8 weeks, 12 weeks, 16 weeks and 20 weeks after the first infection. The kidneys were removed aseptically and were subjected to histopathological and biochemical investigations. Blood cultures were done to exclude bacteremia. In addition the peritoneal macrophages were also isolated [5].
Experimental groups
The animals were divided into 9 groups depending upon the infecting organisms and their iron availability status. (i) Control (C)-only normal saline is injected. (ii) Jectofer Treated (jf)-iron overloading was induced injectable iron (i.e. iron sorbitol solution, Jectofer). (iii) Desferal Treated (D0- i ron depleted conditions were induced using an iron chelator (i.e. Desferal). (iv) 31B Infected (31B)-animals were infected with type-1 piliated E. coli strain 31B. (v) 31B Infected and Jectofer Treated (31B + )-animals were infected with E. coli 31B along with iron overloading (vi) 31B Infected and Desferal Treated (31B-)- i ron depleted animals were infected with E. coli 31B (vii) BH5 Infected (BH5)-animals were infected with non-piliated E. coli strain BH5. (viii) BH5 Infected and Jectofer Treated (BH5+)-animals were infected with E. coli BH5 along with iron overloading (ix) BH5 Infected and Desferal Treated (BH5-) - i ron depleted animals were infected with E. coli BH5.
Doses
Iron sorbitol solution (Jectofer, Astra, India, 150 mg/ml) was diluted with phosohate buffered saline (pH 7.0) to make final concentration of 5 mg/10 ml.). 0.2 ml of the solution was injected intramuscularly per day. The dose was started 5 days before every infection and continued upto, 3 days after every infection (total 9 days) in all the animals treated with jectofer.

Role of iron and iron chelation therapy in oxygen 301
Iron depleted conditions were induced in animals using desferal (Sigma). Animals, l0 mg/Kg body weight, were administered intraperitonealy per day following the same schedule as that for desferal.
Histopathological examination
The kidney tissue preserved in formalin was dehydrated in an ethanol gradient of 70 100%. The tissue was embedded in paraffin, sectioned and stained with haematoxylin and eosin. Grading of the severity of pathological lesion was also done [6].
Biochemical estimations
Lactate dehydrogenase was estimated by the method of Vassault [7]. The LDH activity was measured at 30°C from the amount ofpyruvate consumed by continuously monitoring the decrease in absorbency owing to oxidation of N A D H at 339 nm.
Glucose-6-PO4 dehydrogenase was estimated by the method of Dror et al. [8]. The rate of increase of absorbency at 339 nm, which will measure the rate of appearance of NADPH, taken indirectly as a measure of G-6-P-DH activity.
Method of Goldberg and Spooner [9] was followed to quantitate glutathione reductase activity. Catalytic activity of the enzyme was measured by following the decrease in absorbency owing to the oxidation of NADPH at 339 nm.
Thiobarbituric acid reactive lipid peroxides were measured [10], which is used as an index for the quantitation of MDA. Protein was estimated by the method of Lowry et al. [11].
Chemiluminescence response
Chemiluminescenge response of phagocytic cells isolated from peritoneal cavity was measured [12] using latex as non specific stimulant. Counts were noted after every 10 secs until it reached a peak. Results were expressed as cpm/106 macrophages.
RESULTS
Figure 1 shows the effect of iron overload and iron chelation on the histopathology of mouse kidney during the course of chronic pyelonephritis. It was found that desferal significantly decreases the severity of pathological damage induced by both the strains of E. coli, at all the stages of infection. Whereas jectofer (i.e iron overload) significantly enhanced the same. Desferal itself does not bring about any histopathological changes of the kidney tissue as observed in the control animals receiving desferal only, however excess of iron damages the tissue even in the absence of infection (Fig. 2a). Further E. coli strain 31-B was not able to induce renal scarring in the group of mice not receiving jectofer, whereas, scarring was observed with both the E. coli studied in the group of mouse receiving jectofer, even at 12 weeks post infection interval (Fig. 2b).
Figure 3a shows a significant increase (p < 0.001) in the levels of MDA formation in both the jectofer treated group and the jectofer treated + infected groups with either of the strain used, in comparison to their respective controls. Desferal treatment in the absence of any infection does not significantly alter the MDA levels, however both the infected groups treated with desferal showed decreased lipid peroxidation in comparison to the corresponding-infected group without desferal treatment.
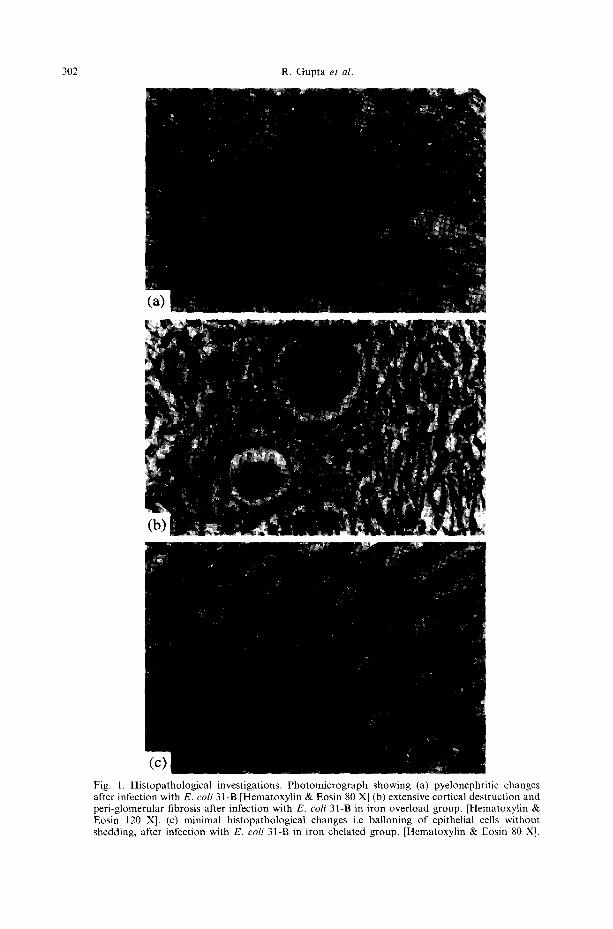
302 R. Gupta et al.
Fig. 1. Histopathological investigations. Photomicrograph showing (a) pyelonephritic changes after infection with E. coli 31-B [Hematoxylin & Eosin 80 X] (b) extensive cortical destruction and peri-glomerular fibrosis after infection with E. coli 31-B in iron overload group. [Hematoxylin & Eosin 120 X]. (c) minimal histopathological changes i.e balloning of epithelial cells without shedding, after infection with E. coli 31-B in iron chelated group. [Hematoxylin & Eosin 80 X].

Role of iron and iron chelation therapy in oxygen
(a) C q T i m e (weeks)
~ 4 [ ~ 8 ~ 1 2 ~ 1 6 ~ 2 0 Jf .... .......q
Df
31B
31B+
31B-
BH5
b
B H 5 + ~']/,')//,'///,'],'/,'/~////)g//H///H///M///M//~//////~
~ H H ~ . ' H H / ) H H J A BH5- ' \ \ \ \ \ \ \ \ \ \ l
i i
0 2 4 6 10 12 Histopathological score
14
1 0 0 l
Time (weeks)
~ 1 2
80- [ ~ 16 ~ 2 0
60-,
i ~ 4o
2
_ _ j - _ _
31B 31B+
r 1 I J
t i
I l
I I
[ [
T ~ 31B- BH5 BH5+
Groups
(b)
B~IS-
Fig. 2. Histopathological investigations (a) Severity score of histopathological lesions in different groups (b) percentage of scarred kidneys in different group of animals at different time intervals.
303
F i g u r e 3 b s h o w s t h a t i r o n o v e r l o a d n o t o n l y s i g n i f i c a n t l y e n h a n c e s t he L D C L r e s p o n s e
b u t a l so d e c r e a s e s t h e t i m e in w h i c h p e a k c h e m i l u m i n e s c e n c e ( C L ) w a s o b s e r v e d . T h e
r e v e r s e is t r u e fo r t h e d e s f e r a l t r e a t e d g r o u p s .

304 R. Gupta et al.
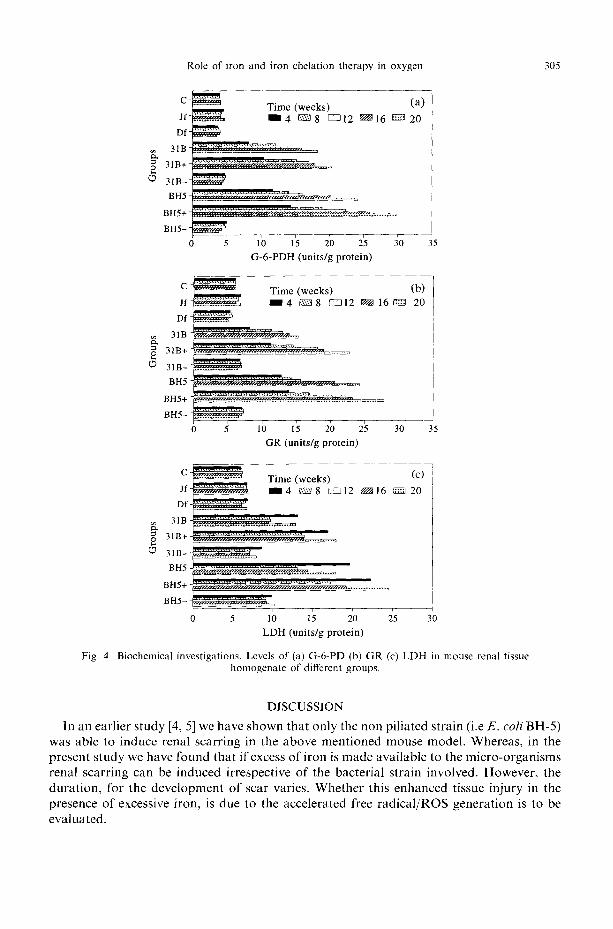
Neither desferal nor jectofer treatment in the absence of any infection significantly alters the levels of various enzymes in the renal tissue homogenate. However, both the infected groups treated with jectofer showed enhanced levels of all the enzyme studied, at all the stages of infection. Whereas desferal does the reverse (Fig. 4).
L
C ~ - -- (a)
i t D f
~ Time (weeks)
31B ~ 4
~ ~ 8 I
3 1 B + ~ ~ , , ~ [[ I 12
....................................................... ~ ~¢~ 16
31 B- @~ili~ 20
BH5
BH5+
BH5-
i
10 20 3; 4'0 Amount of MDA (n moles/g protein)
5O
c (b)
J# /
Df ' ~ Time (weeks)
31B ~ ~ 1 4
~ ~ 8 31B+ [Z[] 12 i
BH5 . . . . . . . ........ 7 i
BH5+ ~:~
BH5- ! • . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
0 5 10 15 20 25 30 35 CPM x 104/106 macrophages
Fig. 3. Quantitation of reactive oxygen species. (a) Amount of MDA in mouse renal tissue homogenate of different groups; (b) chemiluminescence response of peritoneal macrophages of mice
from different groups.
Role of iron and iron chelation therapy in oxygen 305
c ~
D f ~ 31B ~ ; ;~ -2 ; ' }~ - : : : ' ; 2 ;2 .~
3 1 B - ~ '
BH5 - ; : : : : : ' : : ' : 7 " ; : ; : : 2 7 : : . 7 2 . : : : : : : . . . .
BH5+ ~ ; . ; . ; . ; . ' ' ;2 ' ; ' - '~ ' ' ' ' '~ - -~- -~ '~ ' - -~ ....................
B H 5 - ~ ~
o 5 1o 15 2'0 2; ~o G-6-PDH (units/g protein)
Time (weeks) (a) ram4 c~s~8 [55312 9 1 6 ~ 2 0
I
J 35
e Time (we ks) (b) Jf. ............. ~ I 4 ~s~8 ~ 1 2 ~ 1 6 ~ 20
31B ]
31B+ ~-----;2;';5 "2-'2 ~ : ~ .. . . . . . . ]
3 1 B - ~
BH5 ~ : ' : : ' : Y : : ' : 2 : ' : : ' 2 " : 2 : 2 ' 2 : : : : : : :
BH5+ . . . . . . . ~ ~ - -
B H 5 - F n - - - 0 5 r 10 15 20 25 3'0 35
GR (units/g protein)
c ~ . : ; : : : : : : : (c) F ................ Time (weeks) J f ~ ' " 4 ~ 8 [25312 NN16 ~ 2 0
D f ~ i ' BIB ....................... 2222222': . . . . . . . . .
o~ 31 B+ ~Y.-'.-"..-'"...£'.LZ;;-;;;';;'Z"2.';7"~.~
0 5 10 15 20 25 30
LDH (units/g protein)
Fig. 4. Biochemical investigations. Levels of (a) G-6-PD (b) GR (c) LDH in mouse renal tissue homogenate of different groups.
DISCUSSION
In an earlier study [4, 5] we have shown that only the non piliated strain (i.e E . c o l i BH-5) was able to induce renal scarring in the above ment ioned mouse model. Whereas, in the present study we have found that if excess of i ron is made available to the micro-organisms renal scarring can be induced irrespective of the bacterial strain involved. However, the dura t ion , for the development of scar varies. Whether this enhanced tissue injury in the presence of excessive iron, is due to the accelerated free radica l /ROS generat ion is to be evaluated.

306 R. Gupta et al.
Major ROS formed by incomplete reduction of molecular oxygen include superoxide anion (O2_), hydrogen peroxide (H202), hydroxyl radical (OH) and singlet oxygen (O:) [13]. Hydrogen peroxide and 0 2 may be directly damaging, or more often, they interact to form a highly reactive species that can attack every molecule in living cells [14]. This species is the hydroxyl radical, (OH). Wolf[15] has shown the precise reaction sequence that comprises the hydroxyl radical attack, and its damaging effect on the living cell, Hydroxyl radicals attack membrane lipids by hydrogen atom abstraction, leaving behind a carbon radical (lipid) that swiftly combines with oxygen, ultimately forming a peroxy radical (lipid-O;). Further this peroxy radical can attack adjacent lipid in the cell wall membrane abstracting hydrogen atom to form lipid peroxide (lipid-O2H) and this propagates the chain reaction. Additionally, lipid peroxides can fragment to give a wide range of products including more radicals, hydrocarbon gases and aldehydes that are highly cytotoxic even in minute amounts. Hydroxyl radicals also attack other molecules such as proteins and DNA bases, leading to peroxy radical formation and this initiates similar chain reactions [16].
We have found that iron alone (i.e in the absence of any infection) enhances the production of ROS shown by increased lipid peroxidation and LDCL. However values of the enzyme parameters studied remained unaltered. This data is supported by the findings of Halliwell and Gutteridge [16]. These workers had suggested that iron accelerates generation of 02 and H202 by non enzymatic oxidation of several molecules including glutathione. Further, the group of animals receiving jectofer, when infected with either of the 2 strains studied showed increased levels of all the parameters studied, hence increased ROS generation leading to more tissue damage. This data is in accordance with our previous study[17] whereby we have shown that biochemical parameters i.e. glucose-6-phosphate dehydrogenase, glutathione reductase, lactate dehydrogenase and lipid peroxidation can be used as indicators of free radical generation and have a positive correlate with the extent of renal tissue damage. This data is also supported by the findings of Halliwell et al. [18] who have shown that iron enhances the free radical generation either via Fenton reaction or via iron catalysed Haber Weiss reaction. In addition excess of iron has been reported to be toxic for phagocytes as well as being due to the formation of excessive ROS. These species alter phagocytosis through peroxidation of neutrophil membrane lipids leading to impaired phagocytic function [19].
In another set of experiments, where a group of mice depleted of iron was infected with E. coli 31-B and BH-5, we found that damage caused by the infecting bacterial strains is neutralized to some extent by limiting the iron availability to the them. This data corroborates the findings of many of the previous studies [20-22] which showed that both Haber Weiss and Fenton reactions can be inhibited using an iron chelator. Therefore the present study shows that renal tissue damage is the direct consequence of free radicals generation during the infective process and iron is directly involved in oxygen free radicals mediated tissue injury.
REFERENCES 1. Contran R. S. and Pennington J. E. Urinary tract infection, pyelonephritis and re-flux nephropathy. In The
Kidney (Eds Bremmer B. M. and Rector F. C.), pp. 1571-1632. (1981). 2. Meylan P. R., Markert M., Bille J. and Glauser M. P. (1989) Relationship between neutrophil mediated
oxidative injury during acute experimental pyelonephritis and chronic renal scarring, lt~fectious Immunology 57, 2196-2202.

Role of iron and iron chelation therapy in oxygen 307
3. Halliwell B. and Gutteridge J. M. C. Free Radicals in Biology and Medicine. 2nd Ed. Clarendon Press, Oxford. (1989).
4. Gupta R., Ganguly N. K., Ahuja V., Joshi K. and Sharma S. (1995) An ascending non obstructive model for chronic pyelonephritis in BALB/c mice. Journal of Medical Microbiology 43, 33-36.
5. Bjornson A. B. and Michael J. G, (1971) Contribution of humoral and cellular factors to the resistance to experimental infection by pseudomonas aeruginosa in mice. Interaction between immunoglobulins, heat labile serum factors and phagocytic cells in the killing of bacteria. Infectious Immunology 4, 462-467.
6. Garg U. C., Ganguly N. K., Sharma S., Chakravarti R. N. and Bhatnagar R. K. (1987) Quantitative histopathological methods for evaluation of experimental ascending pyelonephritis. Medical Science Research 15, 365-368.
7. Vassault A. (1983) Lactate Dehydrogenase. In Methods of Enzymatic Analysis Vol Ill (Eds Bergmeyer H. U. and Grs B. l.), pp. 118-126. Verlag Chemic Grab H. Weinheim.
8. Dror Y., Sasson H. F., Watson J. J. and Johnson B. C. (1970) G-6-PD assay in liver and blood. Clinica Chimica Acta 28, 291-298.
9. Goldberg D. M. and Spooner R. J. (1983) Glutathione reductase. In Methods qf Enzymatic Analysis Vol Ill (Eds Bergmeyer H. U. and Grs B. l,), pp. 258-265. Verlag Chemic Gmb H, Weinheim.
I0. Ohkawa H., Ohishi N. and Yagi K. (1979) Assay of lipid peroxides in animal tissues by TBA reaction. Annals of Biochemistry 95, 351 358.
11. Lowry O. H., Rosenbrough N. J., Farr A. 1. and Randall R. J. (1951) Protein measurement with the folin phenol reagent. Journal o f Biological Chemistry 193, 265275.
12. Cheung K., Archibald A. C. and Robinson M. F. (1984) Chemiluminescence produced by PMNS stimulated by immune complex. Australian Journal of Experimental Biology and Medical Science 62, 403-419.
13. Halliwell B. and Gutteridge J. M. C. (1987) Oxygen free radicals and iron in relation to medicine. Some problems and concepts. Archives of Biochemistry and Biophysics 246, 501-514.
14. Yagi K. (1986) Oxygen radicals and tissue injury. Trends in Biochemical Science II, 18 19. 15. Wolf S. P., Garner A. and Dean R. T. (1986) Free radicals, lipids and protein degradation. Trends in
Biochemical Science ! 1, 27-31. 16. Halliwell B. and Gutteridge J. M. C. (1986) lron and free radicals: two aspects of antioxidant protection.
Trends" in Biochemical Science 11, 372-375. 17. Gupta R. and Ganguly N. K. (1996) Contribution of Type-1 pili in the pathogenesis of experimental murine
in relation to biochemical parameters. Medical Microbiology Letters 5, 105-112. 18. Halliwell B., Gutteridge J. M. C. and Cross C. E. (1992) Free radicals, anti oxidants and human disease:
where are we now. Journal of Laboratory and Clinical Medicine i19, 598-620. 19. Waterlot Y., Contineux B., Harriga-Muller C., Laurent E. D. M., Vanherwegham J. L. and Fandu P. (1985)
Impaired phagocytic activity of neutrophils in patients receiving hemedialysis: the critical role of iron overload. British Medical Journal 291, 501-504.
20. Ortize de M. P. R. (1990) Free radical modification of prosthetic heine groups. Pharmacological Therapeutics 48, 95-120.
21. Kyle M. E., Nakai D. and Sakaida I. (1989) Endocytosis of superoxide dismutase is required in order for the enzyme to protect hepatocytes from the cytotoxicity, of hydrogen peroxide. Journal o]' Biological Chemistry 263, 3784~3789.
22. Halliwell B. (1987) Oxidants and human diseases: some of the new concepts. FASEB Journal 1, 358-364.