rnainterference$(rnai)$$...
TRANSCRIPT

RNA interference (RNAi) and its applica3ons
Carlos Camilleri

Content of the presenta3on
• Introduc1on • Argonaute proteins • piRNAs: biogenesis and gene silencing • miRNA and siRNA
– Differences – Biogenesis – Gene Silencing – Transla1onal ac1va1on
• Applica1ons of RNA interference – Poten1al therapeu1c applica1on in gene1c diseases – Poten1al an1viral response: HIV
• Current drawbacks of using RNAi • Conclusion

Introduc3on
• RNA interference is a process of gene silencing
• Three different types of RNA involved: – Micro interference RNA (miRNA) – Small interference RNA (siRNA) – Piwi-‐interac1ng RNA (piRNA)
• RNA-‐induced silencing complex (RISC)
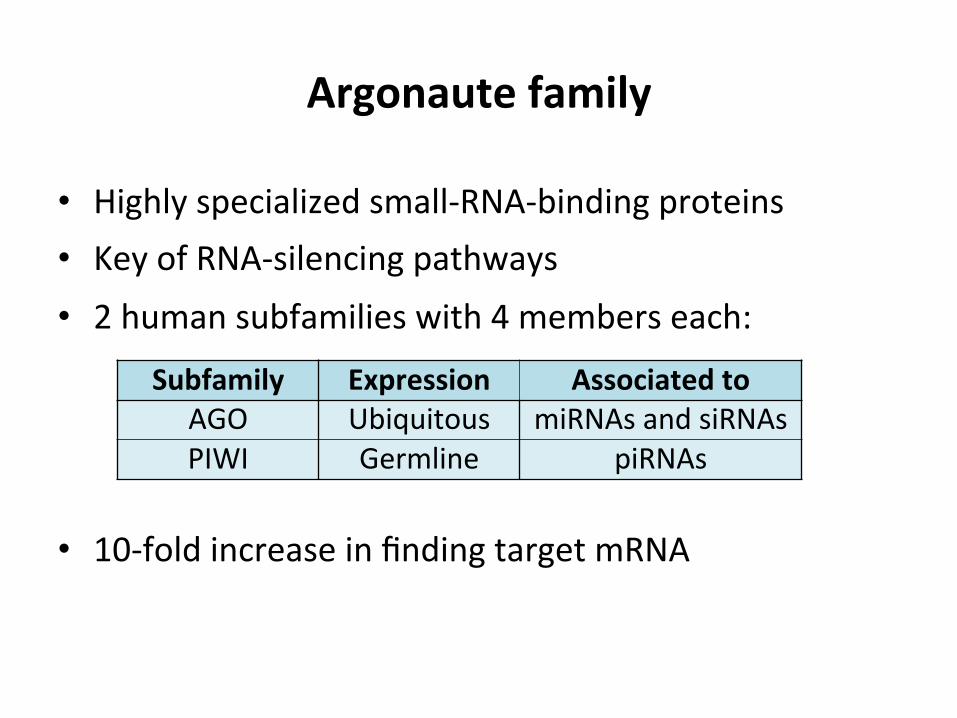
Argonaute family
• Highly specialized small-‐RNA-‐binding proteins • Key of RNA-‐silencing pathways • 2 human subfamilies with 4 members each:
• 10-‐fold increase in finding target mRNA
Subfamily Expression Associated to AGO Ubiquitous miRNAs and siRNAs PIWI Germline piRNAs

piRNAs biogenesis and gene silencing
• Derive from transposons à share homology
• Transcribed from piRNA clusters: – Long single-‐stranded transcript – Sense or an1sense transcrip1on
• Biogenesis of piRNAs follows 2 different pathways: – Primary processing pathway – Amplifica1on pathway (ping-‐pong cycle)
• Mature piRNAs are 25-‐33 nt in length
• Mechanism of transposon silencing in mammals

piRNAs biogenesis and gene silencing
anchors the 5′ end of the small RNA by providing a binding pocket in which the 5′ terminal base engages in stacking interactions with a conserved tyrosine. In addition, several hydrogen bonds coordinate correct 5′ end binding8. The N domain is required for small RNA loading and assists in unwinding the small RNA duplex10.
The PIWI domain is structurally similar to RNase H, and it has indeed been shown that Argonaute proteins can function as endonucleases and cleave target RNA that is fully complementary to the bound small RNA8. The PIWI domain contains a catalytic triad composed of DDX (where X is D or H). Recent structural work on an Argonaute protein from yeast revealed that a fourth residue is also essential turning the catalytic centre into a tetrad of DEDX11. Generally, only a subset of Argonaute proteins possesses cleavage activity. In mam-mals, for example, only AGO2 of the AGO subfamily is catalytically active and functions as an endonuclease12,13. Besides the PIWI domain, an unstructured loop in the N domain is also important for cleavage14.
Recently, crystal structures of human AGO2 have been reported15,16. Overall, their structure is strikingly similar to known bacterial and archaeal Argonaute pro-teins, underlining the high conservation of the basic principles of small RNA pathways.
Mechanisms of Argonaute loadingThe AGO subfamily and the PIWI subfamily use differ-ent mechanisms for small RNA loading, and they will be discussed separately with an emphasis on AGO clade proteins.
Loading AGO clade members. Dicer processing gener-ates a short double-stranded RNA of about 20–24 nt in length. However, only one strand associates with the AGO protein and becomes the guide strand. How is the correct strand selected, and how is it loaded into AGO proteins? One important determinant for strand selection lies in the small RNA duplex itself: both in the miRNA pathway and the siRNA pathway, the strand with the less stably paired 5′ end is preferentially loaded into AGO proteins. These thermodynamic differ-ences between the small RNA ends are known as the asymmetry rule17,18.
In various organisms, Dicer proteins directly inter-act with AGO proteins and strand selection, and load-ing is achieved within such complexes19–21. In addition, Dicer proteins partner with various different dsRNA-binding proteins, which have been most extensively studied in D. melanogaster. In flies, two Dicer proteins exist: Dcr1 processes precursor miRNAs (pre-miRNAs) and loads Ago1 for miRNA-guided gene silencing, whereas Dcr2 cleaves perfectly paired long dsRNA and loads siRNAs into Ago2. Dcr1 interacts with loquacious (Loqs), which is essential for the miRNA pathway22. Dcr2 requires R2d2, which is important for the siRNA pathway. R2d2 functions as a sensor for the thermodynamic asymmetry within an siRNA duplex and positions Dcr2 for correct strand selection23,24. In mammals, the dsRNA-binding domain proteins TRBP
Nature Reviews | Genetics
Primaryprocessing
PrimarypiRNAs
PIWI protein
Sense transcript
Trimming
Secondaryprocessing(ping-pong cycle)
Mature sensepiRNA
Mature antisense piRNA
Antisense piRNASense piRNA
Antisense transcript
Trimming
piRNA cluster
Sense or antisensetranscript
Zuc
Box 2 | piRNA biogenesis and function
Mobile genetic elements such as transposons are a constant threat for the genome. PIWI-interacting RNAs (piRNAs) protect germline cells from transposons in organisms as diverse as flies, fish or mammals. piRNAs are 25 to 33 nt in length, depending on the PIWI clade protein that they bind to. piRNAs derive from distinct transposons that are referred to as piRNA clusters, but the piRNAs from each locus are characterized by a complex mixture of sequences spanning large portions of the transposon5. piRNA clusters are transcribed in the sense or antisense direction, and the long single-stranded RNA serves as the basis for piRNA production.
The biogenesis of piRNAs is independent of Dicer165 and requires other nucleases. Two biogenesis pathways are important for piRNA production. First, a primary processing pathway generates primary piRNAs, and these are then amplified by an amplification cycle referred to as the ping-pong loop5. In the primary biogenesis pathway, the long transposon transcript is initially cleaved by the nuclease zucchini (Zuc; see the figure)166–168, which probably generates the 5′ ends of primary piRNAs. The other steps of primary piRNA maturation are not understood5.
In the ping-pong cycle (see the lower right panel of the figure), mature sense primary piRNAs guide PIWI clade proteins to complementary sequences on antisense transcripts from the same piRNA cluster. PIWI proteins use their slicer activity to cleave the target antisense transcript to generate a new 5′ end. This 5′ end is bound by another PIWI protein. In subsequent steps, the 3′ end is trimmed to the length of the mature piRNA, leading to a mature antisense secondary piRNA, which can now target sense transcripts transcribed from the piRNA cluster. In Drosophila melanogaster, the two Piwi proteins Aubergine and Ago3 cooperate in secondary piRNA production to generate sense and antisense piRNAs. However, antisense piRNAs dominate, and a protein called Qin, which contains E3 ligase and Tudor domains, seems to modulate such a heterotypic ping-pong cycle169. In the mouse germ line, the PIWI proteins MILI and MIWI collaborate in piRNA generation. After trimming, piRNAs receive a methyl group at the 3′ end by the methyltransferase HEN1. Primary piRNAs carry such modifications as well5. In Caenorhabditis elegans, an additional piRNA production pathway exists. Short and capped RNA polymerase II transcripts are decapped, processed and directly loaded into PIWI proteins170.
piRNAs guide PIWI proteins to complementary RNAs derived from transposable elements. Similarly to in RNA interference, PIWI proteins cleave the transposon RNA, leading to silencing. In flies, mutations in piwi, aub and Ago3 (which encode the Piwi proteins in &�|OGNCPQICUVGT) are required for transposon silencing in the germ line5. Similar observations were made when the mouse PIWI proteins MILI and MIWI were genetically inactivated171. Here, long interspersed nuclear elements (LINE) and long terminal repeat (LTR) retrotransposons accumulated172.
REVIEWS
NATURE REVIEWS | GENETICS VOLUME 14 | JULY 2013 | 449
© 2013 Macmillan Publishers Limited. All rights reserved

miRNA and siRNA
• O[en referred to as the ‘true’ interference RNA • Similar biogenesis but s1ll key differences
miRNA siRNA
Mature structure dsRNA of 19-‐25nt with a 2nt 3'overhang
dsRNA of 21-‐23nt with a 2nt 3'overhang
TargeBng Mul1ple targets, par1ally complementary
One highly specific target with fully complementariety
Mechanisms of gene regula3on
1. Transla1onal repression 2. mRNA degrada1on
3. Endonucleotydic cleavage 1. Endonucleotydic cleavage

Biogenesis of miRNA and siRNA
• Both form RNA duplexes (dsRNA) a[er transcrip1on
• miRNA requires an extra processing in the nucleus
• Both are exported to the cytoplasm by Expor1n-‐5
• In the cytoplasm, their processing is the same

www.moleculartherapy.org/mtna
siRNA Versus miRNA TherapeuticsLam et al.
3
backbone of the mRNA between bases 10 and 11 relative to the 5!end of the guide strand.30 The mRNA fragments gener-ated are subsequently degraded by different exonucleases.31 By contrast, the target recognition of miRNA is more complex, as different binding sites and different degree of complemen-tarity between the miRNA and the target RNA exist. This is a consequence of imperfect base pairing; miRNA only needs
to be partially complementary to its target mRNA. The com-plementary pairing between mRNA and the mature miRNA typically occurs at the 3! untranslated region (UTR) of the for-mer and the seed region (nucleotides 2–7 from the 5! end) of the latter (Figure 2).32,33 Other miRNA binding sites, such as the centered sites, 3! supplementary sites and bulged sites, are considered to be atypical.32,34,35 Since miRNA-mRNA
Figure 1 Gene silencing mechanisms of siRNA and miRNA. siRNA: dsRNA (either transcribed or artificially introduced) is processed by Dicer into siRNA which is loaded into the RISC. AGO2, which is a component of RISC, cleaves the passenger strand of siRNA. The guide strand then guides the active RISC to the target mRNA. The full complementary binding between the guide strand of siRNA and the target mRNA leads to the cleavage of mRNA. miRNA: Transcription of miRNA gene is carried out by RNA polymerase II in the nucleus to give pri-miRNA, which is then cleaved by Drosha to form pre-miRNA. The pre-miRNA is transported by Exportin 5 to the cytoplasm where it is processed by Dicer into miRNA. The miRNA is loaded into the RISC where the passenger strand is discarded, and the miRISC is guided by the remaining guide strand to the target mRNA through partially complementary binding. The target mRNA is inhibited via translational repression, degradation or cleavage.
miRNA gene
Pol II
Transcription
Pri-miRNA Pre-miRNA
Exportin 5Nucleus
Cytoplasm
miRNADICER
RISC
AGO
Passenger strand iscleaved
Complementary binding of theguide strand to target mRNA
Incomplete complementarybinding of the guide strand to
target mRNA
Passenger strand isdiscarded
3!UTR
Translational repression,mRNA degradation,mRNA cleavage
mRNA cleavage
Activated RISC
mRNA mRNA
miRISC
RISC
AGO
dsRNA
siRNA
Drosha

miRNA and siRNA gene silencing
Endonucleotydic cleavage of target mRNA
• Performed by the AGO2
• Needs perfect complementarity
• Cleavage followed by mRNA degrada1on ####P14(%CR
0CVWTG�4GXKGYU�^�)GPGVKEU
�
�
&GCFGP[NCVKQP
&GECRRKPI
6TCPUNCVKQPCNGNQPICVKQP
6TCPUNCVKQP�KPKVKCVKQP
####P14(%CR
#)1�)9���
2#$2��5
��5
��5
��5
� 764� 764
C
� 6TCPUETKRV�FGEC[
#)1�
6TCPUETKRV�ENGCXCIGD
Seed regionA region consisting of nucleotides 2–8 counted from the 5 end of miRNAs that participates in the interaction between a small RNA and target transcript.
Moreover, miRNA duplexes often contain sufficient bulges to prevent slicing of miR* strands even by com-petent enzymes. Therefore, it has been proposed that miR* strands dissociate in a cleavage-independent man-ner by unwinding — a process that is facilitated by the presence of mismatches in the loaded duplexes113,118,132. Biochemical evidence supports unwinding as a passive, ATP-independent process, with degradation of the miR* strand on its release. It is unclear how plant Argonautes remove the miR* or passenger strand during RISC mat-uration. MiR* and passenger strands could be cleaved through the slicer activity of AGOs (similar to fly AGO2) or unwound passively (like fly AGO1)12–14,132.
The impact of sorting on target regulationThe ultimate result of accurate strand selection and sort-ing is that an active RISC is formed, which is imbued with the ability to regulate a target gene or process. Argonaute family members differ in their biochemical properties, subcellular localization and expression pat-terns, and matching the right small RNA with the correct partner is key to proper biological function.
Although AGO proteins evolved as ribonucleases, animal miRNAs affect their targets without the need for this activity. miRNAs generally interact with their tar-gets through limited base-pairing interactions that are insufficient to place the scissile phosphate of the target in the enzyme active site where cleavage can occur. The prevalence of cleavage-independent repression modes is also reflected in the diversity of the Argonaute family. In mammals, three of the four AGO proteins have lost catalytic potential, and AGO1, the D. melanogaster AGO protein into which most miRNAs are sorted, is a poor enzyme compared with its siRNA-binding cousin113.
miRNA-directed target cleavage has only been reported in a few cases144,145. However, this is assumed to be the principal regulatory mode for endo-siRNAs and for piRNA-mediated repression of transposons. Here again, the choice of a particular AGO partner is crucial. Piwi family members all retain catalytic compe-tence and D. melanogaster AGO2, the main partner for endogenous and viral siRNAs, is tuned for highly active slicing (BOX 4).
AGO1-associated plant miRNAs usually share exten-sive sequence complementary with their mRNA targets and these interactions often result in target cleavage146. However, recent studies have indicated that cleavage-independent translational repression is widespread in plants, even for highly complementary target sites147. Nevertheless, miRNA-mediated cleavage is of key importance for some processes like the biogenesis of ta-siRNAs, for which the initial slicing event is key to RdRP recruitment and dsRNA synthesis88.
Notably, small RNAs that direct cleavage, for exam-ple, plant miRNAs, piRNAs and fly endo-siRNAs, often have a 2 -O-methyl modification on their 3 termini. Although the purpose of this modification was initially mysterious, it is now clear that this functions as a pro-tective group to prevent small RNA destruction64,142,143. In flies and mammals, small RNAs that have extensive complementarity to their targets can be recognized by terminal uridyl transferases, which mark small RNAs for degradation64. The uridylation event is blocked by the 2 -O-methyl modification, preserving these small RNAs, which have evolved to function through cleav-age64. The balance between protection and targeted destruction has been proposed as a quality control on small RNA sorting and as an evolutionary mechanism to drive animal miRNAs toward a cleavage-independent repression mode64.
hc-siRNAs are thought to function by different mechanisms148,149. They must be sorted into a particu-lar Argonaute, AGO4, which they guide to target DNA loci by base pairing with nascent non-coding transcripts synthesized by RNAPV. Effector proteins, such as the chromatin-remodelling factor DRD1, the de novo meth-yltransferase DRM2 and other factors, are then recruited, resulting in DNA methylation at cytosine residues150,151. As this regulation functions by repressing RNA synthe-sis, it was termed transcriptional gene silencing to distin-guish it from post-transcriptional gene-silencing modes. Some piRNAs in flies and mammals must associate with particular Piwi-family proteins — that is, PIWI and
Box 4 | Mechanisms of target regulation in Drosophila melanogaster
Individual Argonaute (AGO) proteins differ in their expression patterns, subcellular localization and enzymatic properties. Thus, distinct AGOs can function through many different effector modes that may involve slicing of target transcripts, cleavage-inde-pendent regulation and chromatin modification (reviewed in REFS 15–17). Another layer of complexity is added by the degree of sequence complementarity between the AGO-bound small RNA and target transcripts, which determines the mechanism of regulation. a | In flies, AGO1-associated microRNAs (miRNAs) typically target mRNAs in their 3 UTRs to reduce protein synthesis. Owing to limited sequence complementarity between the small RNA (seed region) and the mRNA, such interactions usually do not result in direct cleavage of the targeted transcript. Instead, AGO1 and its partner protein GW182 are likely to disrupt crucial interactions between the polyA tail and the cap of the transcript, leading to a reduction in translational initiation and an induction of mRNA decay173. In mammals, it was recently shown that reduced protein output is predominantly owing to destabilization of the target transcript174. b | Small RNAs bound to Drosophila melanogaster AGO2 do not exhibit a bias towards binding their targets in the 3 UTR. AGO2 primed with a small RNA sharing extensive complementarity with its target typically directs endonucleolytic cleavage of the mRNAs through AGO2 slicer activity. The 2-O-methyl modification of AGO2-bound small RNAs prevents their degradation when targeting perfectly complementary transcripts64,142,143. However, other modes are possible: AGO2 can also regulate targets with limited sequence complementarity through a block in translation initiation (not shown)173. PABP, poly(A)-binding protein
REVIEWS
28 | JANUARY 2011 | VOLUME 12 www.nature.com/reviews/genetics
© 2011 Macmillan Publishers Limited. All rights reserved
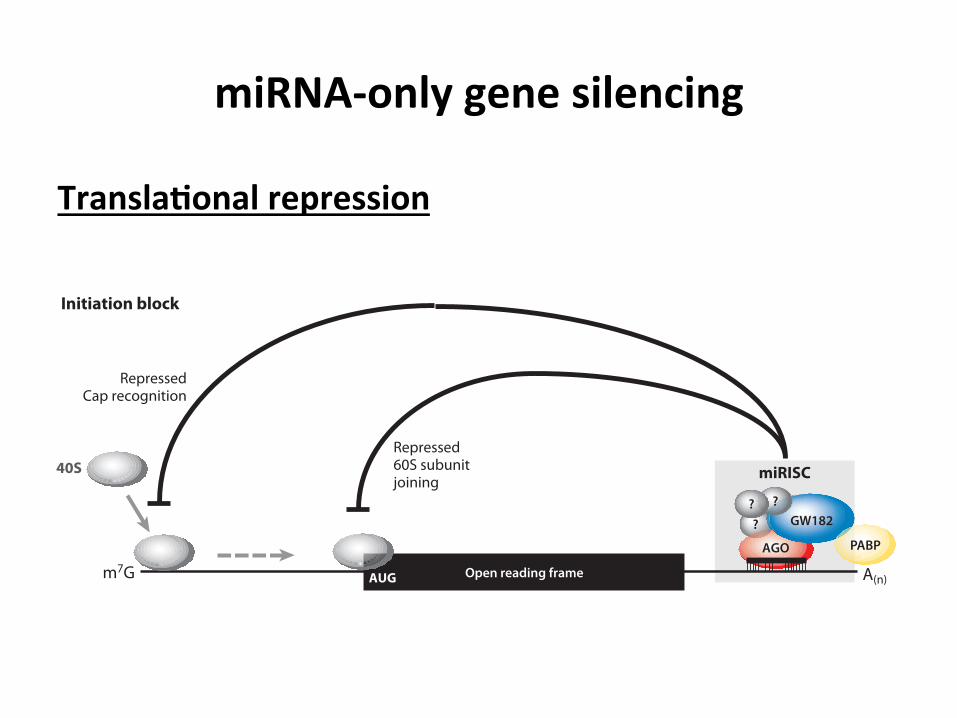
miRNA-‐only gene silencing
Transla3onal repression
ANRV413-BI79-13 ARI 27 April 2010 19:36
A(n)
AGO
GW182?? ?
PABP
miRISC
eIF4E
eIF4E
Elongationblock Proteolysis
Nascentpolypeptide
RepressedCap recognition
Repressed60S subunitjoining
Ribosomedrop-o!
40S
40S
60S
m7G
A(n)m7G
a Initiation block
b Postinitiation block
AGO
GW182?? ?
PABP
miRISC
Open reading frameAUG
Figure 2Schematic diagram of miRNA-mediated translational repression. (a) Initiation block: The miRISC inhibits translation initiation byinterfering with eIF4F-cap recognition and 40S small ribosomal subunit recruitment or by antagonizing 60S subunit joining andpreventing 80S ribosomal complex formation. The reported interaction of the GW182 protein with the poly(A)-binding protein(PABP) (106, 156) might interfere with the closed-loop formation mediated by the eIF4G-PABP interaction and thus contribute to therepression of translation initiation. (b) Postinitiation block: The miRISC might inhibit translation at postinitiation steps by inhibitingribosome elongation, inducing ribosome drop-off, or facilitating proteolysis of nascent polypeptides. There is no mechanistic insight toany of these proposed “postinitiation” models. The 40S and 60S ribosomal subunits are represented by small and large gray spheres,respectively. Ovals with question marks represent potential additional uncharacterized miRISC proteins that might facilitatetranslational inhibition. Abbreviations: AGO, Argonaute; A(n), poly(A) tail; m7G, the 5!-terminal cap.
358 Fabian · Sonenberg · Filipowicz
Ann
u. R
ev. B
ioch
em. 2
010.
79:3
51-3
79. D
ownl
oade
d fr
om w
ww
.ann
ualre
view
s.org
Acc
ess p
rovi
ded
by U
nive
rsita
t Aut
onom
a de
Bar
celo
na o
n 12
/12/
15. F
or p
erso
nal u
se o
nly.

miRNA-‐only gene silencing
Transla3onal repression
ANRV413-BI79-13 ARI 27 April 2010 19:36
A(n)
AGO
GW182?? ?
PABP
miRISC
eIF4E
eIF4E
Elongationblock Proteolysis
Nascentpolypeptide
RepressedCap recognition
Repressed60S subunitjoining
Ribosomedrop-o!
40S
40S
60S
m7G
A(n)m7G
a Initiation block
b Postinitiation block
AGO
GW182?? ?
PABP
miRISC
Open reading frameAUG
Figure 2Schematic diagram of miRNA-mediated translational repression. (a) Initiation block: The miRISC inhibits translation initiation byinterfering with eIF4F-cap recognition and 40S small ribosomal subunit recruitment or by antagonizing 60S subunit joining andpreventing 80S ribosomal complex formation. The reported interaction of the GW182 protein with the poly(A)-binding protein(PABP) (106, 156) might interfere with the closed-loop formation mediated by the eIF4G-PABP interaction and thus contribute to therepression of translation initiation. (b) Postinitiation block: The miRISC might inhibit translation at postinitiation steps by inhibitingribosome elongation, inducing ribosome drop-off, or facilitating proteolysis of nascent polypeptides. There is no mechanistic insight toany of these proposed “postinitiation” models. The 40S and 60S ribosomal subunits are represented by small and large gray spheres,respectively. Ovals with question marks represent potential additional uncharacterized miRISC proteins that might facilitatetranslational inhibition. Abbreviations: AGO, Argonaute; A(n), poly(A) tail; m7G, the 5!-terminal cap.
358 Fabian · Sonenberg · Filipowicz
Ann
u. R
ev. B
ioch
em. 2
010.
79:3
51-3
79. D
ownl
oade
d fr
om w
ww
.ann
ualre
view
s.org
Acc
ess p
rovi
ded
by U
nive
rsita
t Aut
onom
a de
Bar
celo
na o
n 12
/12/
15. F
or p
erso
nal u
se o
nly.
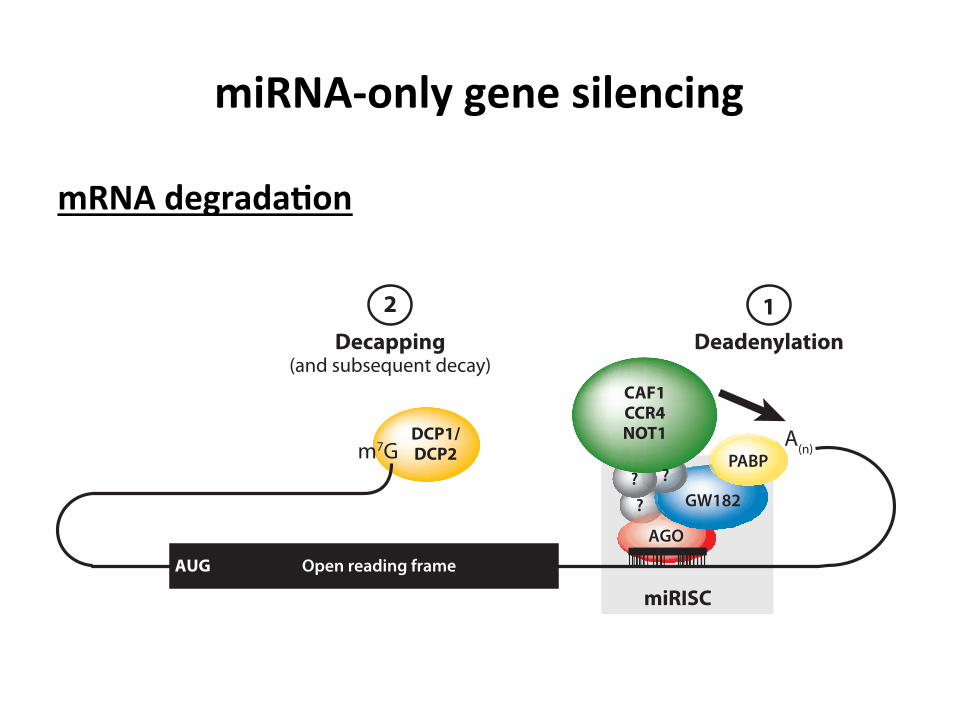
miRNA-‐only gene silencing
mRNA degrada3on
ANRV413-BI79-13 ARI 27 April 2010 19:36
CAF1CCR4NOT1DCP1/
DCP2
DeadenylationDecapping(and subsequent decay)
12
AGO
GW182?? ?
miRISC
A(n)m7G
AUG Open reading frame
PABP
Figure 3Schematic diagram of miRNA-mediated mRNA decay. The miRISC interacts with the CCR4-NOT1deadenylase complex to facilitate deadenylation of the poly(A) tail [denoted by A(n)]. Deadenylation requiresthe direct interaction of the GW182 protein with the poly(A)-binding protein (PABP) (see previous figures).Following deadenylation, the 5!-terminal cap (m7G) is removed by the decapping DCP1-DCP2 complex.The open reading frame is denoted by a black rectangle. Abbreviations: AGO, Argonaute; CAF1,CCR4-associated factor; CCR4, carbon catabolite repression 4 protein; NOT1, negative on TATA-less.
The conclusions from the cell culture stud-ies are strongly supported by in vitro exper-iments using cell-free systems that faithfullyrecapitulate the action of miRNAs in cells.In cell-free extracts from mouse Krebs II as-cites cells (referred to as Krebs extracts) (97),Drosophila embryos (96), and HEK293 cells(95), inhibition of mRNA translation or dead-enylation was dependent on the ability of the“seed sequence” of the miRNA to base-pairto the target sequence in the mRNA. Addi-tion of oligonucleotides, which are comple-mentary to miRNAs (antimiRs), to the ex-tract prevented miRNA function. The threesystems mentioned above made use of an en-dogenous miRNA targeting for in vitro syn-thesized mRNAs. All these aforementionedstudies concluded that the miRNA-mediatedtranslation inhibition occurs at the initiationstep and is due to the interference with thecap recognition process. This is further sup-ported by the findings that miRNAs failed toinhibit IRES-dependent translation or trans-lation from ApppG-capped mRNAs (95–97).More detailed analyses revealed that miRNAsinhibited ribosome initiation complex forma-tion; miR-2 inhibited both 40S ribosomalsubunit recruitment and 80S initiation com-
plex formation in fly embryo extract (96),and 80S initiation complex formation was im-paired in mouse Krebs extract (97). A study byZdanowicz et al. (98) points to the 5!-cap structure itself as being a direct targetof miRNA-mediated translational repression.miR-2-targeted mRNAs bearing modificationsto the triphosphate bridge of the 5! cap demon-strated increased translational repression inboth Drosophila embryonic extracts and S2cells (98). Strong evidence for the notion thatthe cap recognition machinery is indeed thetarget for miRNA-mediated translational re-pression was the demonstration that addinga purified eIF4F complex to the Krebs ex-tract alleviated translational repression of let-7-targeted mRNAs (97). In contrast to theseresults, Wang et al. (99) showed that in arabbit reticulocyte lysate the CXCR4 artificialmiRNA impairs translation by inhibiting thejoining of the 60S subunit, even though the 5!
cap was required for the inhibition. It is pos-sible that this inconsistence with former re-sults stems from nuclease-treated rabbit retic-ulocyte lysate not displaying cap-poly(A) tailtranslational synergy (100, 101), which may, inturn, alter the outcome of miRNA-mediatedrepression.
www.annualreviews.org • Regulation of mRNAs by microRNAs 359
Ann
u. R
ev. B
ioch
em. 2
010.
79:3
51-3
79. D
ownl
oade
d fr
om w
ww
.ann
ualre
view
s.org
Acc
ess p
rovi
ded
by U
nive
rsita
t Aut
onom
a de
Bar
celo
na o
n 12
/12/
15. F
or p
erso
nal u
se o
nly.

Transla3onal ac3va3on by miRNA
• Mechanism s1ll under study • Only found in quiescent cells arrested in G0/G1 • Involves recruitment of FXR1 (fragile X-‐related protein 1)

Applica3ons of RNA interference
• Poten1al therapeu1cs for gene1c diseases – Dominant gene1c disorders caused by a mutant allele in the presence of a second, normal allele
– Currently no cures: difficult to treat the origin, in some cases the muta1on is a SNP
Demonstrated in vitro that a SNP is enough for RNAi to select the mutant allele over the normal one
S1ll under development

Applica3ons of RNA interference
• Poten1al an1viral responses: HIV – Successful targe1ng of HIV-‐encoded RNAs – Successful downregula1on of cellular cofactors needed for the HIV replica1on
– Inhibi1on of HIV replica1on achieved in several human cell lines
Big challenge going from in vitro to in vivo – High viral muta1on rate à escaping mutants – Challenging delivery of the RNAi to the infected cells
S1ll under development

Current major drawbacks of using iRNA
• Off-‐target effects – Gene silencing of unwanted genes
• Delivery methods – Large size of iRNAs (around 14 kDa) – Nega1ve charge of RNA – Generally unstable in vivo – Problems targe1ng specific cells or 1ssues

Conclusion Clinical trials with RNAi are currently underway, but major obstacles, such as off-‐target effects, toxicity and unsafe delivery methods, have to be overcome before RNAi can
be considered a conven1onal drug

Bibliography
1. Lingel, A. & Izaurralde, E. RNAi : Finding the elusive endonuclease. RNA 10, 1675–1679 (2004).
2. Freeley, M. & Long, A. Advances in siRNA delivery to T-‐cells: poten1al clinical applica1ons for inflammatory disease, cancer and infec1on. Biochem. J. 455, 133–47 (2013).
3. Czech, B. & Hannon, G. J. Small RNA sor1ng: matchmaking for Argonautes. Nat. Rev. Genet. 12, 19–31 (2011).
4. Ender, C. & Meister, G. Argonaute proteins at a glance. J. Cell Sci. 123, 1819–23 (2010).
5. Fabian, M. R., Sonenberg, N. & Filipowicz, W. Regula1on of mRNA transla1on and stability by microRNAs. Annu. Rev. Biochem. 79, 351–79 (2010).
6. Wilson, R. C. & Doudna, J. a. Molecular mechanisms of RNA interference. Annu. Rev. Biophys. 42, 217–39 (2013).
7. Lam, J. K. W., Chow, M. Y. T., Zhang, Y. & Leung, S. W. S. siRNA Versus miRNA as Therapeu1cs for Gene Silencing. Mol. Ther. Nucleic Acids 4, e252 (2015).
8. Meister, G. Argonaute proteins: func1onal insights and emerging roles. Nat. Rev. Genet. 14, 447–59 (2013).
9. Chak, L.-‐L. & Okamura, K. Argonaute-‐dependent small RNAs derived from single-‐stranded, non-‐structured precursors. Front. Genet. 5, 172 (2014).