rna editing
TRANSCRIPT

ARIJIT SHOME M-5490 (Veterinary Biochemistry)
Credit Seminaron
1
RNA editing in disease and differentiation : A Potential target of therapeutics

Let us refresh a bit about RNA family and their functional relationship:
DNA-(P)-RNA Pol II-----------Pre RNA (Primary Transcript)---------------Undergo processing to give rise to ------------mRNA, tRNA, rRNA
Processing include: 1. Capping—adds 7-methylguanylate to the 5’ end2. Polyadenylation----adds a poly(A) tail to the 3’ end3. Splicing-----remove introns and joins exons4. RNA editing----alters RNA sequence and transcritome diversity
Non-coding RNAs:Small non-coding RNAs (sncRNAs): three classes - short interfering
RNAs (siRNAs), microRNAs (miRNAs) and PIWI-interacting RNAs (piRNAs): important in mRNA degradation, translational repression, or both, therefore regulating gene expression.
Long non-coding RNAs (lncRNAs, >100 nucleotides): pivotal molecule for the transcriptional and post-transcriptional regulation of gene expression
sncRNAs and lncRNAs play critical roles in defining DNA methylation patterns, as well as chromatin remodeling thus having a substantial effect in epigenetics.
4
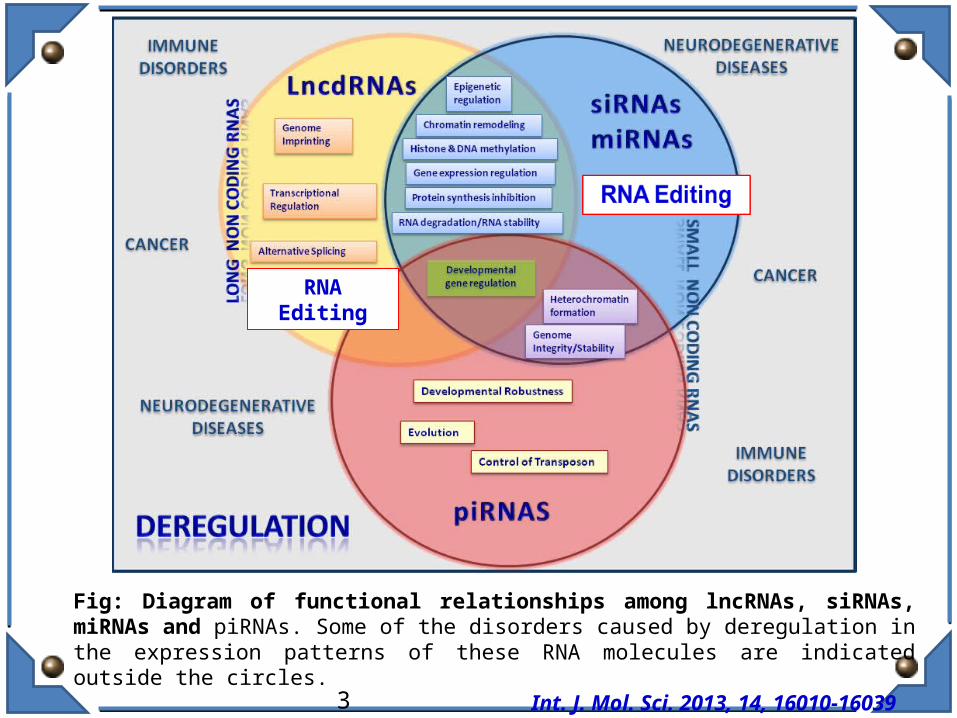
Fig: Diagram of functional relationships among lncRNAs, siRNAs, miRNAs and piRNAs. Some of the disorders caused by deregulation in the expression patterns of these RNA molecules are indicated outside the circles. Int. J. Mol. Sci. 2013, 14, 16010-16039
RNA Editing
3
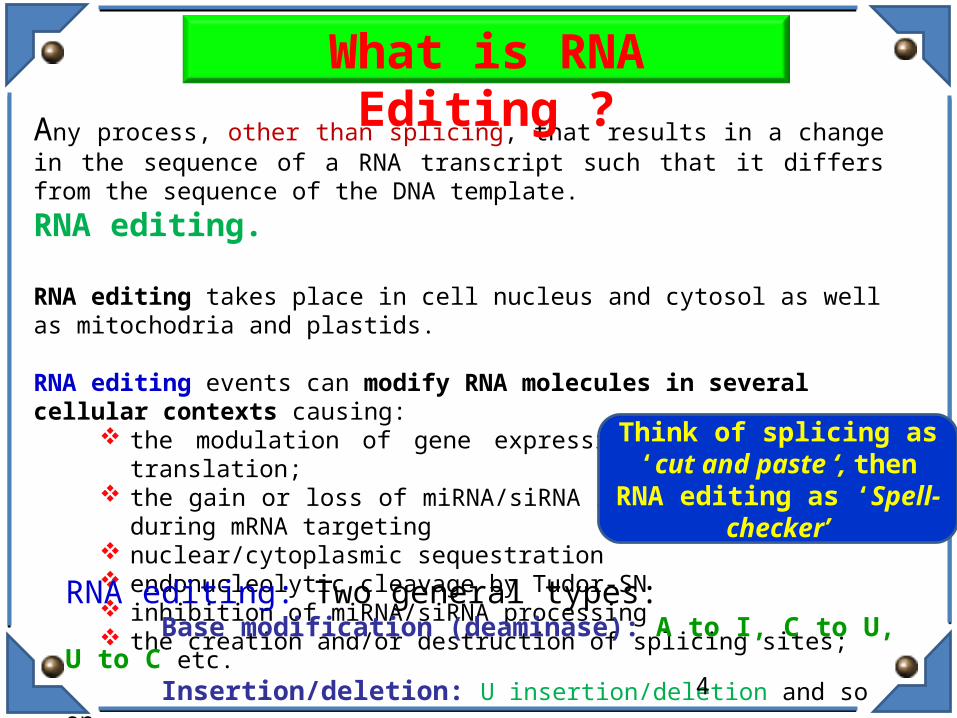
Any process, other than splicing, that results in a change in the sequence of a RNA transcript such that it differs from the sequence of the DNA template.
RNA editing.
RNA editing takes place in cell nucleus and cytosol as well as mitochodria and plastids.
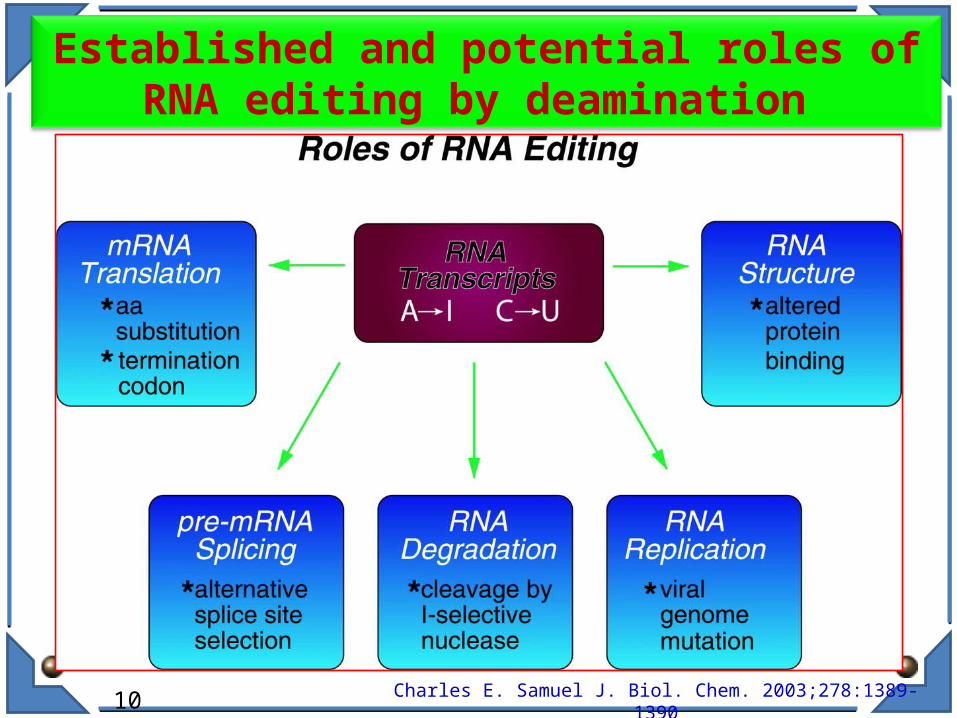
RNA editing events can modify RNA molecules in several cellular contexts causing: the modulation of gene expression pathways during translation; the gain or loss of miRNA/siRNA recognition elements during mRNA
targeting nuclear/cytoplasmic sequestration endonucleolytic cleavage by Tudor-SN inhibition of miRNA/siRNA processing the creation and/or destruction of splicing sites;
RNA editing: Two general types:Base modification (deaminase): A to I, C to U, U to C etc.Insertion/deletion: U insertion/deletion and so on
4
What is RNA Editing ?
Think of splicing as ‘cut and paste ‘, then RNA editing as ‘Spell-
checker’
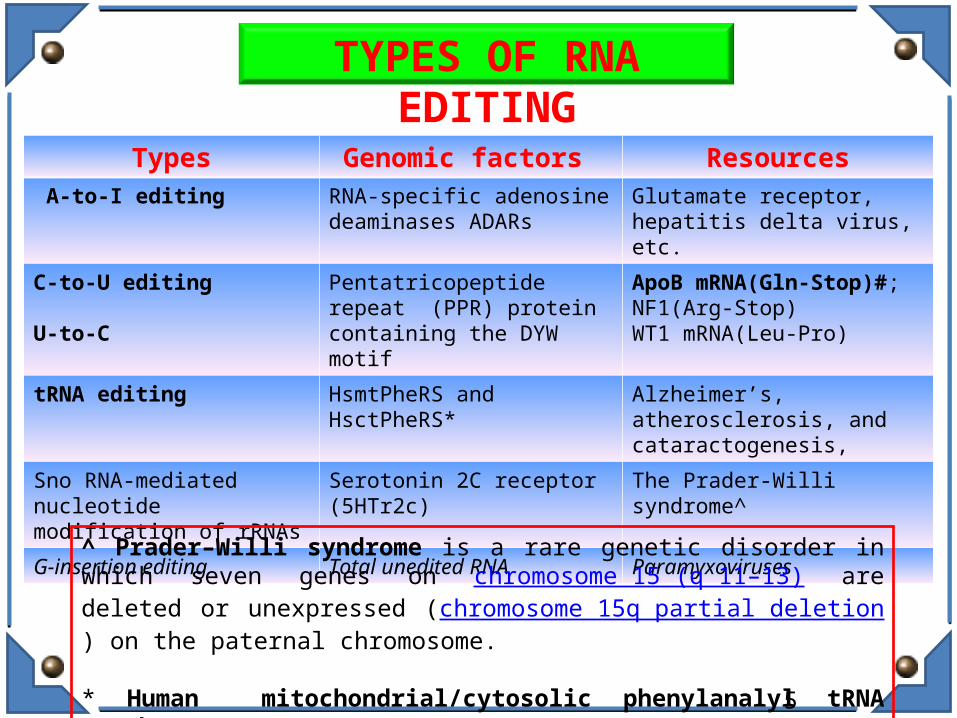
Types Genomic factors Resources A-to-I editing RNA-specific adenosine
deaminases ADARsGlutamate receptor, hepatitis delta virus, etc.
C-to-U editing
U-to-C
Pentatricopeptide repeat (PPR) protein containing the DYW motif
ApoB mRNA(Gln-Stop)#; NF1(Arg-Stop)WT1 mRNA(Leu-Pro)
tRNA editing HsmtPheRS and HsctPheRS* Alzheimer’s, atherosclerosis, and cataractogenesis,
Sno RNA-mediated nucleotidemodification of rRNAs
Serotonin 2C receptor (5HTr2c) The Prader-Willi syndrome^
G-insertion editing Total unedited RNA Paramyxoviruses
TYPES OF RNA EDITING
^ Prader–Willi syndrome is a rare genetic disorder in which seven genes on chromosome 15 (q 11–13) are deleted or unexpressed (chromosome 15q partial deletion) on the paternal chromosome.
* Human mitochondrial/cytosolic phenylanalyl tRNA synthetase
5
6
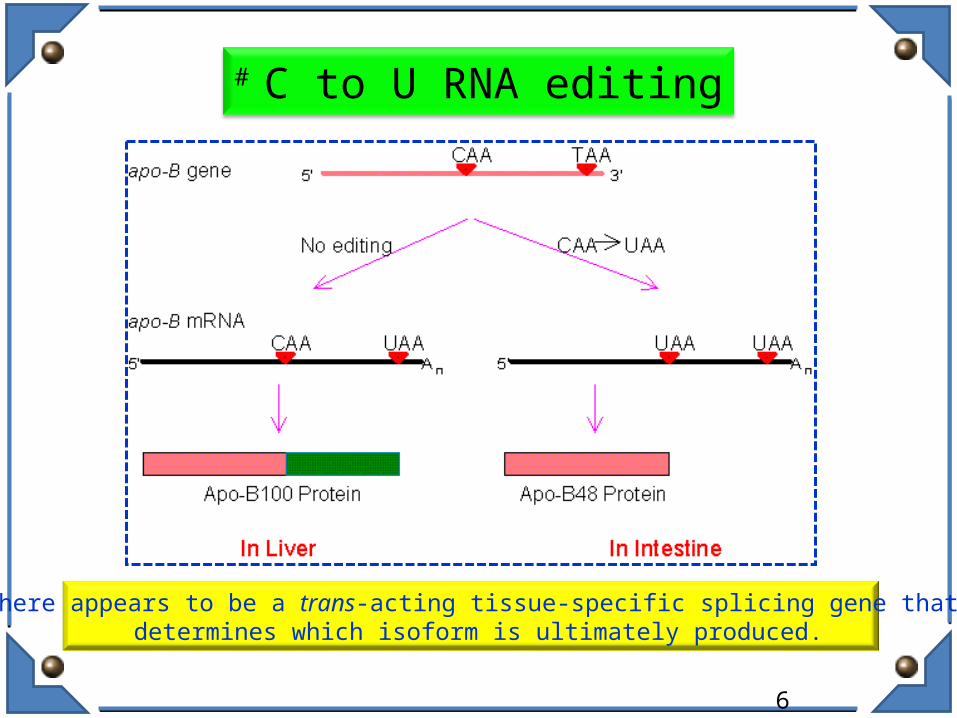
# C to U RNA editing
There appears to be a trans-acting tissue-specific splicing gene that determines which isoform is ultimately produced.
• Human Apolioprotein (ApoB100) essential for removal of LDL in Liver ; Tissue specific cytidyl deamination (C6666-U) introduces in-frame stop codon giving truncated ApoB48 by ApoB mRNA editing enzyme catalytic polypeptide 1 (ApoBEC1) in intestine.
• The RNA-binding protein RBM47 is required for normal Cytidine to Uridine RNA editing in mice and is sufficient for the C to U editing activity
of ApoBEC1.
• RBM47 is found in the epithelial cells of the intestine and interacts with APOBEC1
• BM47 is sufficient with APOBEC1 for C to U RNA editing.
• C to U editing of ApoB and other RNA is impaired in Rbm47-deficient mice.
Fossat et al. EMBO Reports (2014) 15(8): 903–910
7
C to U RNA editing
RECENTLY...
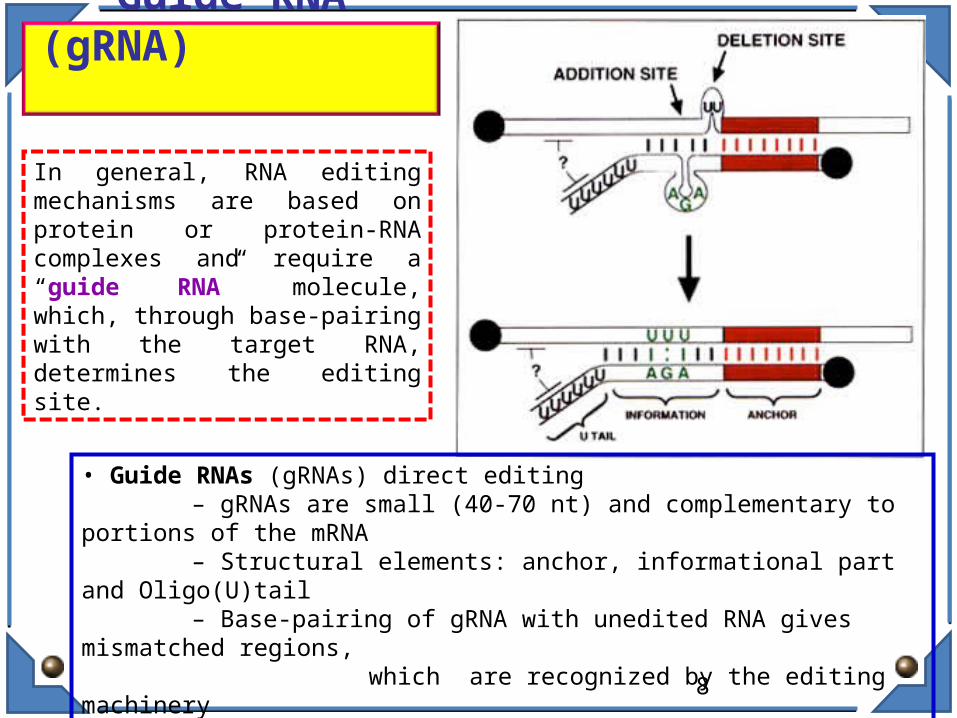
Guide RNA (gRNA)
• Guide RNAs (gRNAs) direct editing – gRNAs are small (40-70 nt) and complementary to portions of the mRNA – Structural elements: anchor, informational part and Oligo(U)tail – Base-pairing of gRNA with unedited RNA gives mismatched regions,
which are recognized by the editing machinery
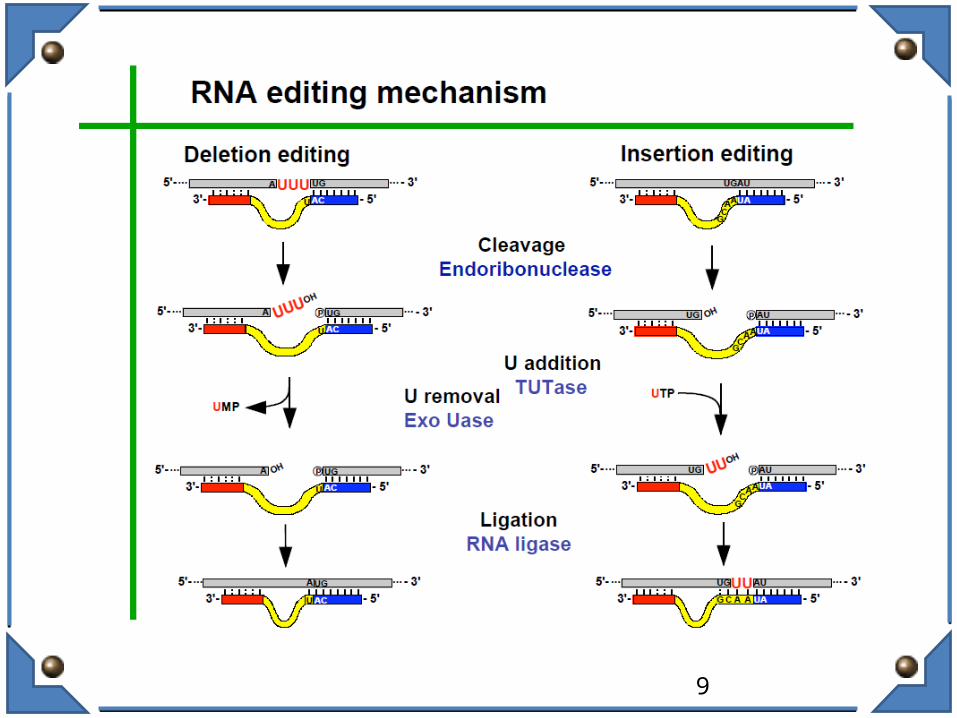
– Machinery includes an Endonuclease, a Terminal Uridylyl Transferase (TUTase),and a RNA ligase• Editing is directional, from 3’ to 5’
In general, RNA editing mechanisms are based on protein or protein-RNA complexes and require a “guide RNA” molecule, which, through base-pairing with the target RNA, determines the editing site.
8
9
Charles E. Samuel J. Biol. Chem. 2003;278:1389-139010
Established and potential roles of RNA editing by deamination
Annu Rev Biochem.(December 2010)
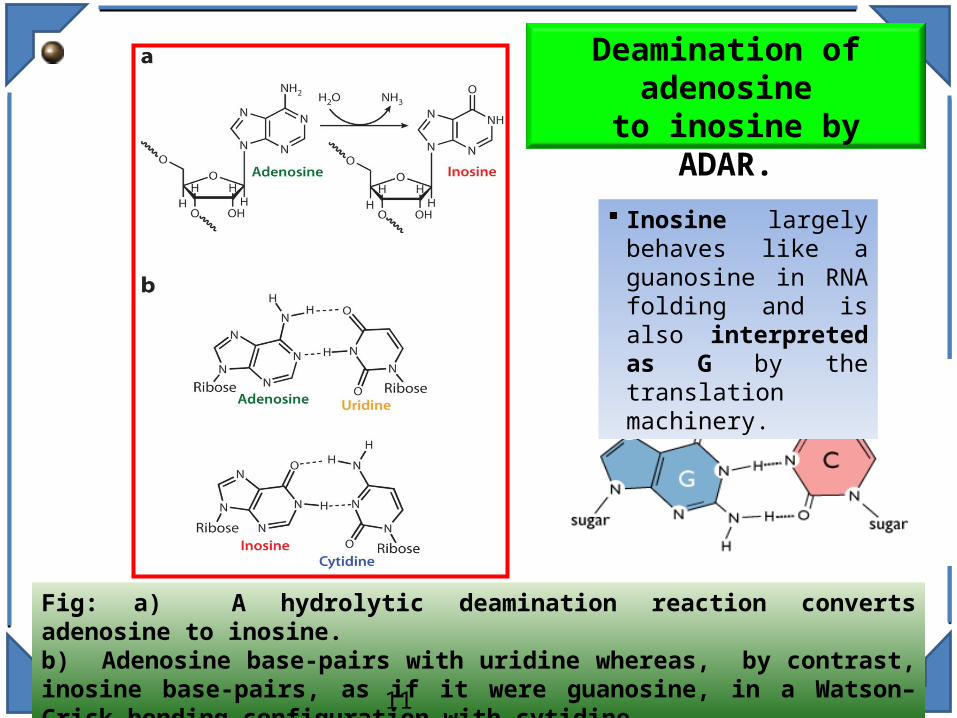
Deamination of adenosine
to inosine by ADAR.
Fig: a) A hydrolytic deamination reaction converts adenosine to inosine. b) Adenosine base-pairs with uridine whereas, by contrast, inosine base-pairs, as if it were guanosine, in a Watson–Crick-bonding configuration with cytidine.
Inosine largely behaves like a guanosine in RNA folding and is also interpreted as G by the translation machinery.
11
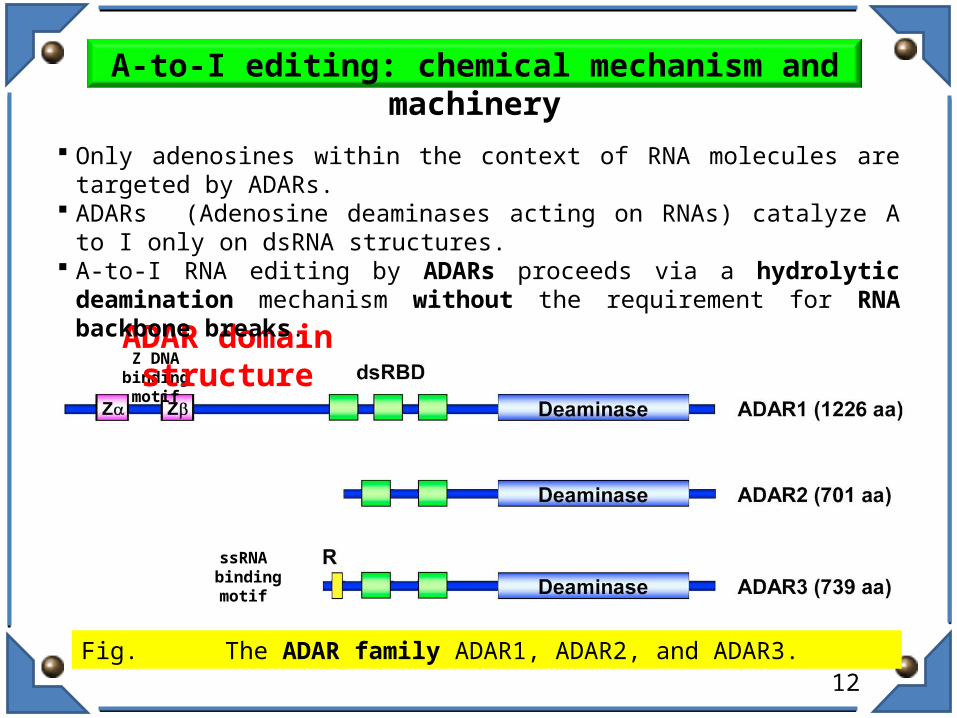
Fig. The ADAR family ADAR1, ADAR2, and ADAR3.
Z DNA binding motif
ssRNA binding motif
ADAR domain structure
Only adenosines within the context of RNA molecules are targeted by ADARs. ADARs (Adenosine deaminases acting on RNAs) catalyze A to I only on dsRNA
structures. A-to-I RNA editing by ADARs proceeds via a hydrolytic deamination mechanism
without the requirement for RNA backbone breaks.
12
A-to-I editing: chemical mechanism and machinery

ADAR1p150
ADAR1p110
Fig: The ADAR family. ADARs undergo both homo- and heterodimerization and are likely catalytically active as dimers.
Splice variants
13
The C terminal deaminase domain, coordinates a Zn2+ ion in its catalytic center and the
functional deaminase fold requires the incorporation of an
inositol hexaphosphate (IP6) molecule.
Zhou and Jin . ADAR-mediated RNA editing in non-coding RNA sequences. Sci China Life Sci, 2013, 56: 944–952,
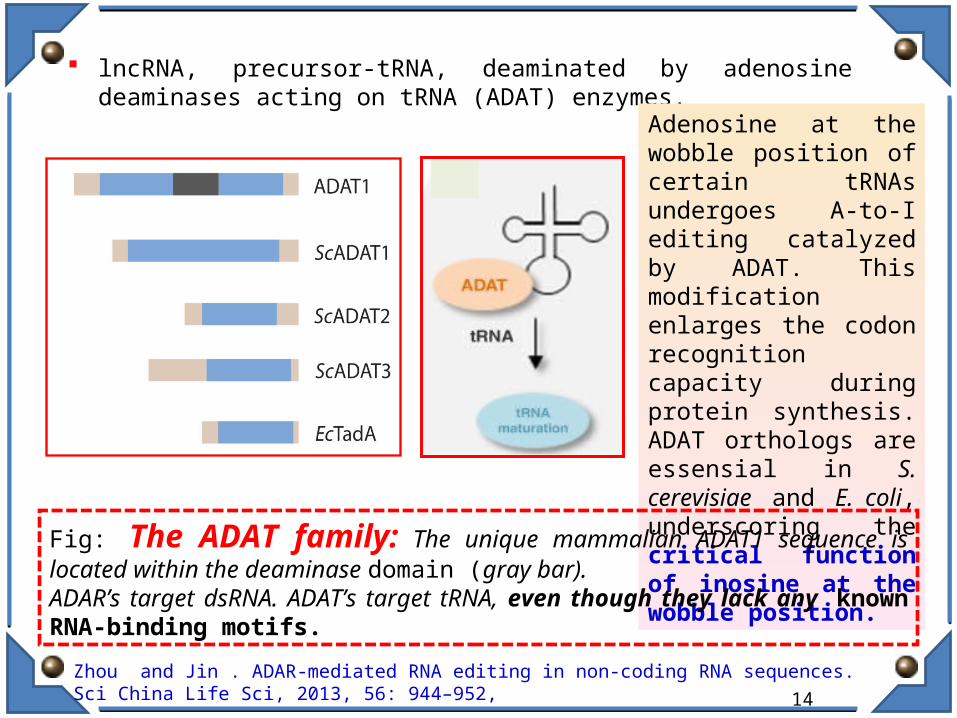
lncRNA, precursor-tRNA, deaminated by adenosine deaminases acting on tRNA (ADAT) enzymes.
Adenosine at the wobble position of certain tRNAs undergoes A-to-I editing catalyzed by ADAT. This modification enlarges the codon recognition capacity during protein synthesis. ADAT orthologs are essensial in S. cerevisiae and E. coli, underscoring the critical function of inosine at the wobble position.
14
Fig: The ADAT family: The unique mammalian ADAT1 sequence is located within the deaminase domain (gray bar). ADAR’s target dsRNA. ADAT’s target tRNA, even though they lack any known RNA-binding motifs.

Fig: Substrates of ADARsYang Y, Zhou X X, Jin Y F. ADAR-mediated RNA editing in non-coding RNA sequences. Sci China Life Sci, 2013, 56: 944–952,
A-to-I RNA editing sitesalso abundantly occur inIntronic regions as well
asin 3′-UTRs.
A-to-I RNA editing: two forms - Promiscuous and Specific
The promiscuous Occurs within longer duplexes of hundreds of nucleotides (e.g., Alu elements). Up to 60% of adenosines could be edited.
The specific Occurs in short and/or unstable duplex RNA regions. Up to10% of their adenosines selectively could undergo deamination. eg. A-to-I RNA editing in miRNA.
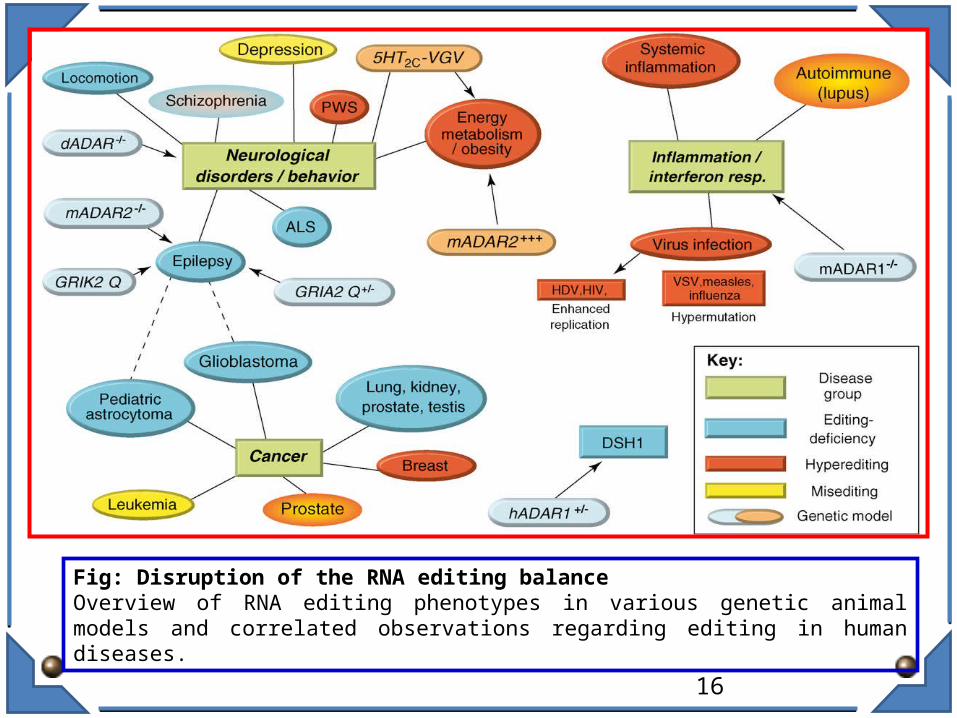
Fig: Disruption of the RNA editing balanceOverview of RNA editing phenotypes in various genetic animal models and correlated observations regarding editing in human diseases.
16

L-glutamate is the predominant excitatory neurotransmitter in vertebrate nervous systems, has been implicated in neuronal plasticity, memory and learning.
RNA editing affects protein function and higher order physiology. In mammals, particular attention is given to the GluA2, an ionotropic glutamate receptor subunit, and Kv1.1, a voltage-dependent K channel⁺ , because they are particularly well understood.
L-glutamate is the predominant excitatory neurotransmitter in vertebrate nervous systems, has been implicated in neuronal plasticity, memory and learning.
RNA editing affects protein function and higher order physiology. In mammals, particular attention is given to the GluA2, an ionotropic glutamate receptor subunit, and Kv1.1, a voltage-dependent K channel⁺ , because they are particularly well understood.
Channels open and close, or gate, in response to an extrinsic factor, such as concentration of a specific ligand (eg. ionotropic glutamate receptors) or the changes in the transmembrane voltage (eg. Voltage-dependent K channels).⁺
Channels open and close, or gate, in response to an extrinsic factor, such as concentration of a specific ligand (eg. ionotropic glutamate receptors) or the changes in the transmembrane voltage (eg. Voltage-dependent K channels).⁺
Four AMPA subunits: GluA1–4 Five Kinate receptor subunits:GluK1–5
Three basic groups of ionotropic glutamate receptors: α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPA) receptors, kinate receptors, and N-methyl-d-aspartate (NMDA) receptors.
Three basic groups of ionotropic glutamate receptors: α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPA) receptors, kinate receptors, and N-methyl-d-aspartate (NMDA) receptors.
17
ROLE IN NERVOUS SYSTEM
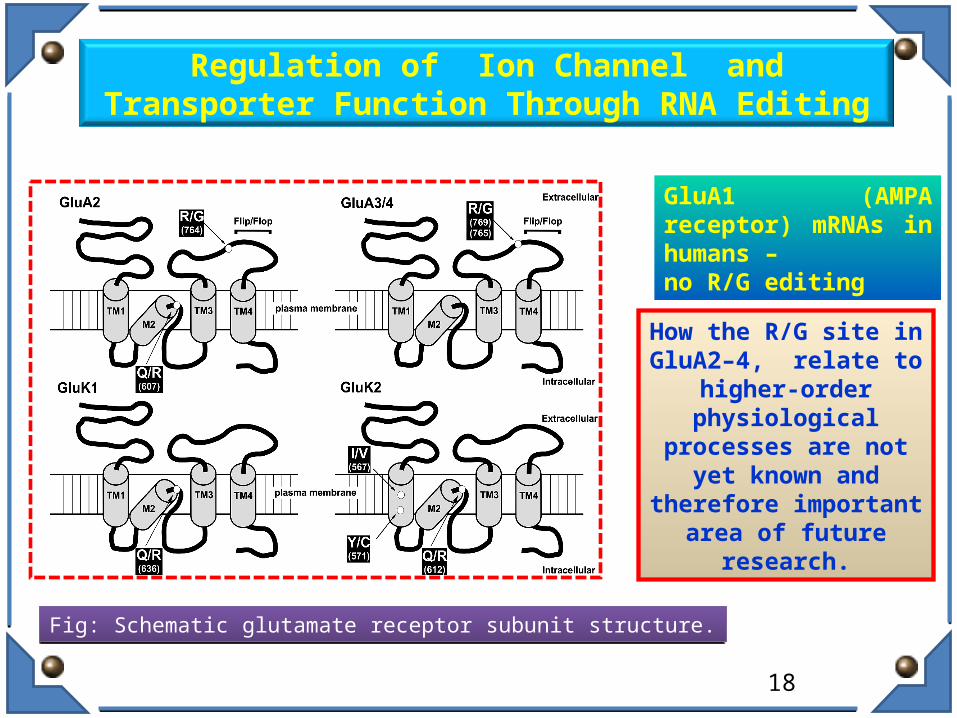
Fig: Schematic glutamate receptor subunit structure. Fig: Schematic glutamate receptor subunit structure.
Regulation of Ion Channel and Transporter Function Through RNA Editing
How the R/G site in GluA2–4, relate to higher-
order physiological processes are not yet known and therefore
important area of future research.
GluA1 (AMPA receptor) mRNAs in humans – no R/G editing
18
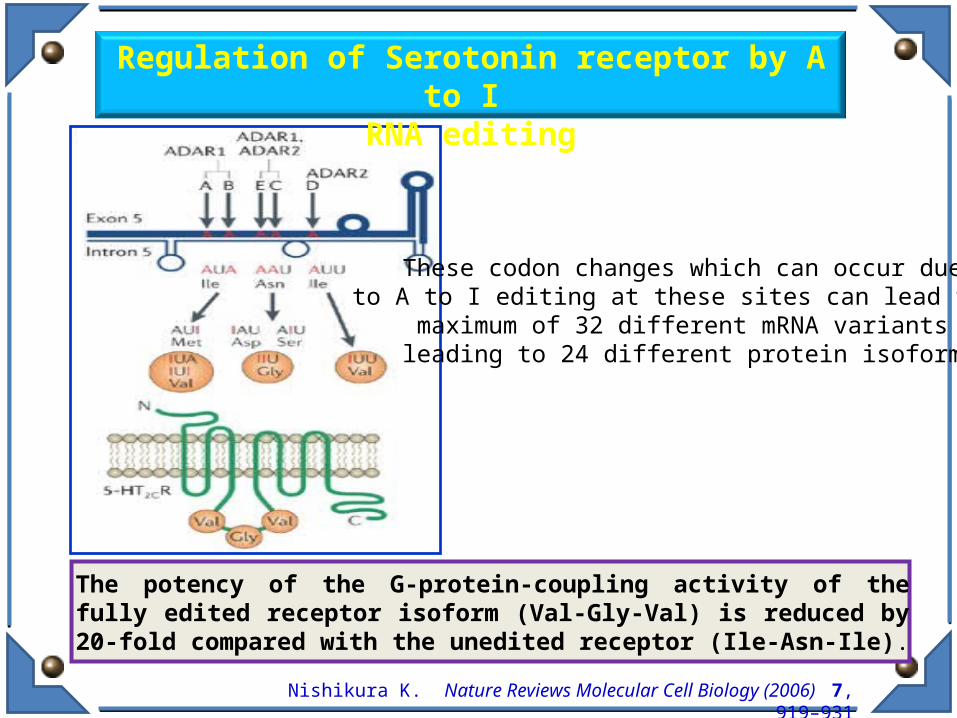
Nishikura K. Nature Reviews Molecular Cell Biology (2006) 7, 919–931
The potency of the G-protein-coupling activity of the fully edited receptor isoform (Val-Gly-Val) is reduced by 20-fold compared with the unedited receptor (Ile-Asn-Ile).
Regulation of Serotonin receptor by A to I
RNA editing
These codon changes which can occur due to A to I editing at these sites can lead to a maximum of 32 different mRNA variants leading to 24 different protein isoforms
Father plans to sell his own kidney to fund life-saving treatment for his three morbidly-obese children
Prader-Willi syndrome (PWS)•a constant desire to eat food,
•which seems driven by a permanent feeling
•of hunger and can easily lead to dangerous
•weight gain, restricted growth,
•leading to short stature
•reduced muscle tone
•learning difficulties
•lack of sexual development
•behavioural problems.
20
ZEE NEWS : 17th APRIL, 21:42
Obese: (From left) Yogita, five, Harsh, 18 months, and Anisha, three, weigh 34kg,15kg and 48kg respectively
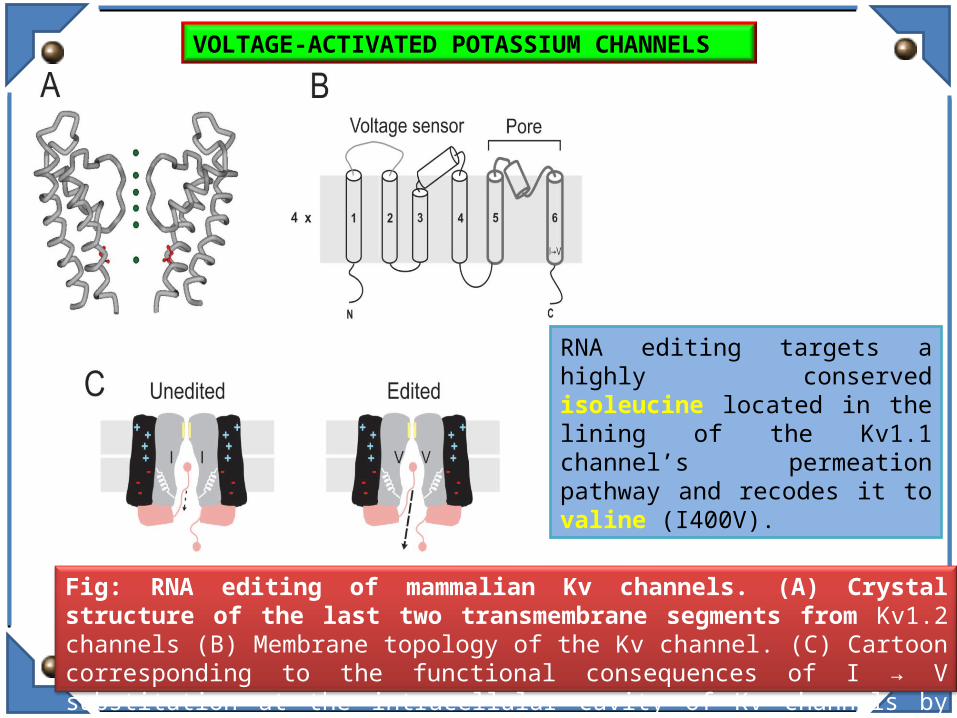
Fig: RNA editing of mammalian Kv channels. (A) Crystal structure of the last two transmembrane segments from Kv1.2 channels (B) Membrane topology of the Kv channel. (C) Cartoon corresponding to the functional consequences of I → V substitution at the intracellular cavity of Kv channels by ADAR2.
RNA editing targets a highly conserved isoleucine located in the lining of the Kv1.1 channel’s permeation pathway and recodes it to valine (I400V).
VOLTAGE-ACTIVATED POTASSIUM CHANNELS

Recently, three RNA editing sites were discovered in mRNAs encoding rodent Cav1.3 channels, clustered in four contiguous amino acids located in the cytoplasmic C terminus
Much work remains before we get a clear understanding of how RNA editing is used to regulate animal physiology in general, and excitability in
particular.
Holmgren and Rosenthal (2014) Curr. Issues Mol. Biol. 17: 23-36.
A recent study on the Drosophila developmental transcriptome has predicted more than 600 recoding events due to adenosine deamination, and more than 100 of these occur in ion channels, transporters or the machinery for synaptic vesicle release (Graveley et al.,2011).
22
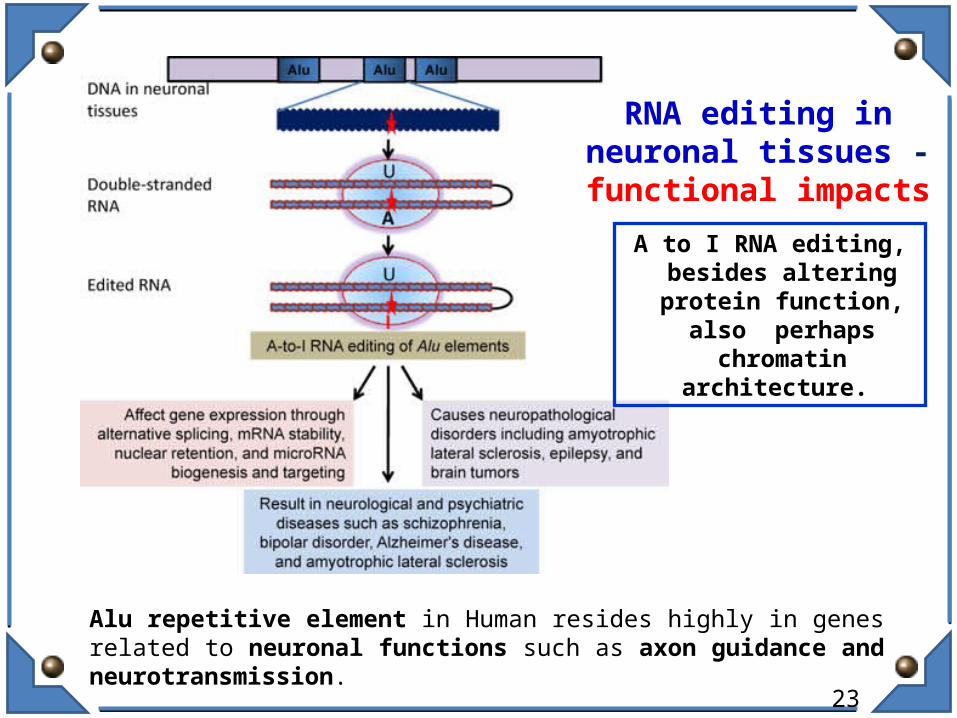
RNA editing in neuronal tissues - functional
impacts
A to I RNA editing, besides altering protein function, also perhaps chromatin
architecture.
Alu repetitive element in Human resides highly in genes related to neuronal functions such as axon guidance and neurotransmission.
23
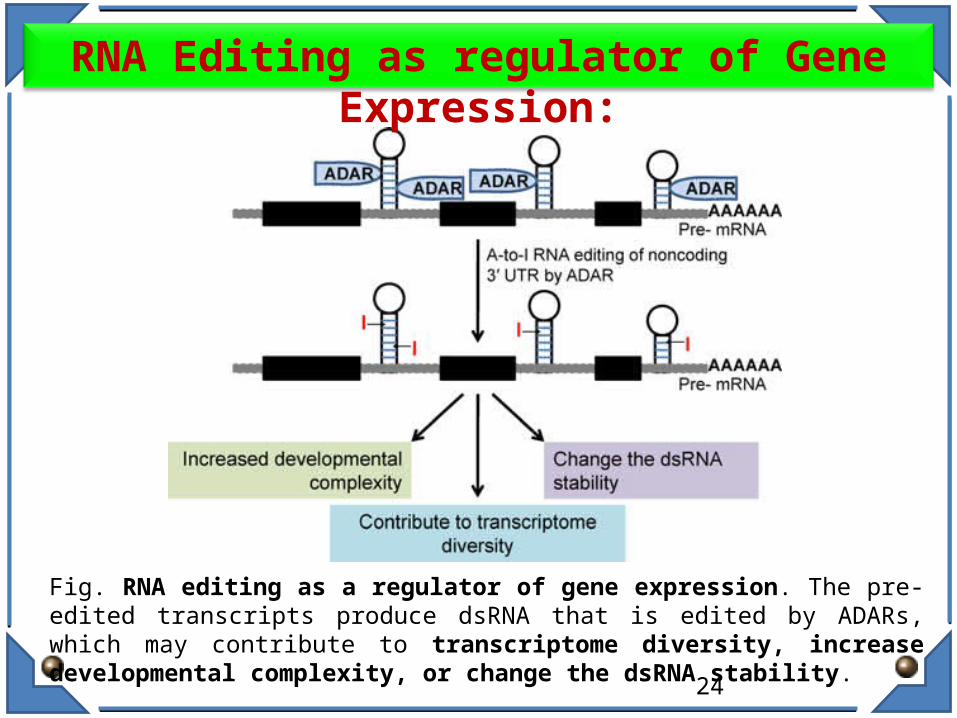
Fig. RNA editing as a regulator of gene expression. The pre-edited transcripts produce dsRNA that is edited by ADARs, which may contribute to transcriptome diversity, increase developmental complexity, or change the dsRNA stability.
24
RNA Editing as regulator of Gene Expression:

Either by inactivating a tumor supressor or activating gene that promote tumor development and progression.
In tumor supressor gene NF1(Neurofibromatosis type 1) mRNA editing at 3916 from C-U results in introduction of inframe stop codon. Similarly, WT1 (Wilms’ tumor gene) mRNA U-C conversion at 839 position takes place in colorectal cancer.
A-I editing is responsible for structure change and base pairing features of the RNA molecule and involved in cell differentiation
A-I editing is involved in human embyogenesis.
Other cancer related RNA editing targets: Antizyme Inhibitor1(AZIN1)– A-I editing is increased in hepatocellular carcinoma and Glioma associated oncogene 1 (GLI1).
Imbalance in expression of ADAR enzymes is highly correlated with cancer development and progression.
25
RNA Editing in tumorigenesis / Cancer:
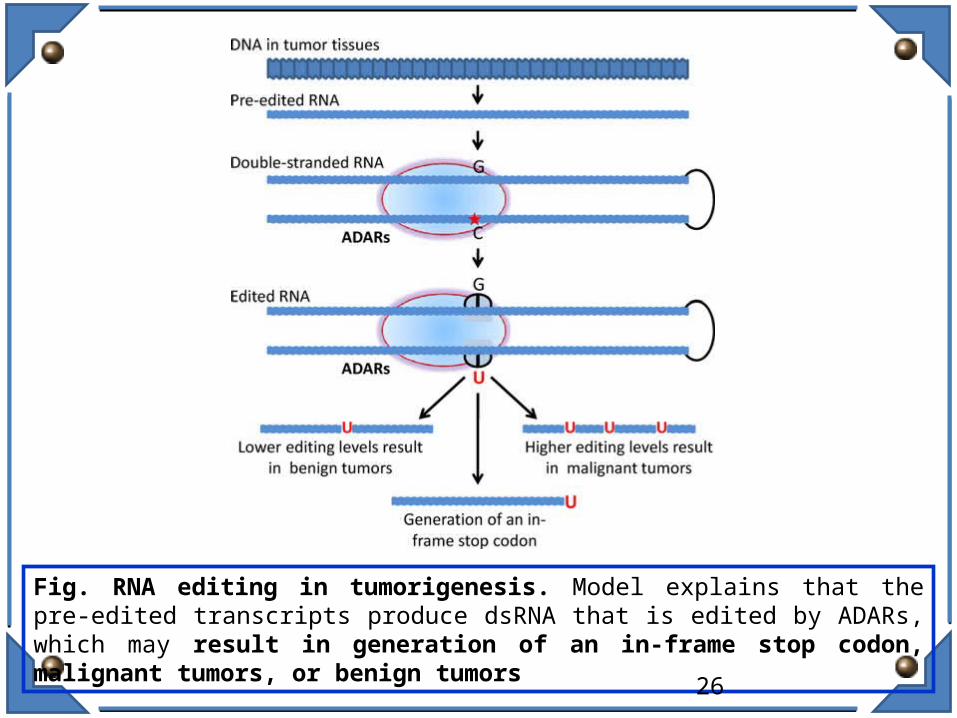
Fig. RNA editing in tumorigenesis. Model explains that the pre-edited transcripts produce dsRNA that is edited by ADARs, which may result in generation of an in-frame stop codon, malignant tumors, or benign tumors
26
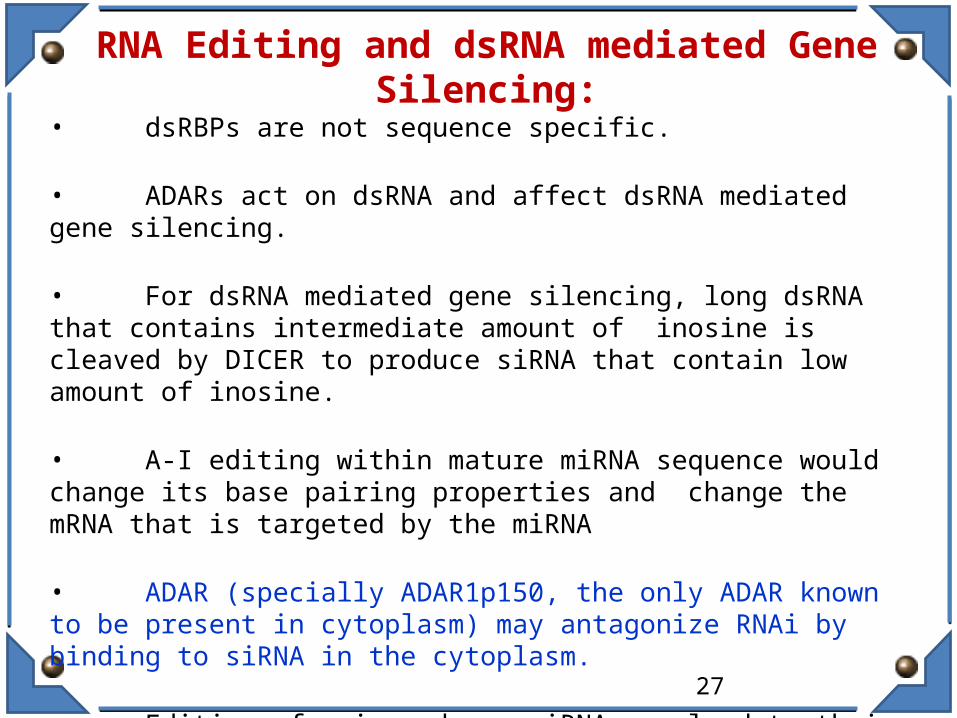
RNA Editing and dsRNA mediated Gene Silencing:
• dsRBPs are not sequence specific.
• ADARs act on dsRNA and affect dsRNA mediated gene silencing.
• For dsRNA mediated gene silencing, long dsRNA that contains intermediate amount of inosine is cleaved by DICER to produce siRNA that contain low amount of inosine.
• A-I editing within mature miRNA sequence would change its base pairing properties and change the mRNA that is targeted by the miRNA
• ADAR (specially ADAR1p150, the only ADAR known to be present in cytoplasm) may antagonize RNAi by binding to siRNA in the cytoplasm.
• Editing of pri- and pre-miRNA may lead to their degradation, possibly by Staphylococcal nuclease, Tudor-SN, a component of RSIC (RNA induced silencing complex). Cleavage of dsRNA is stimulated by presence of 3-4 contiguous IU base pairs.
27
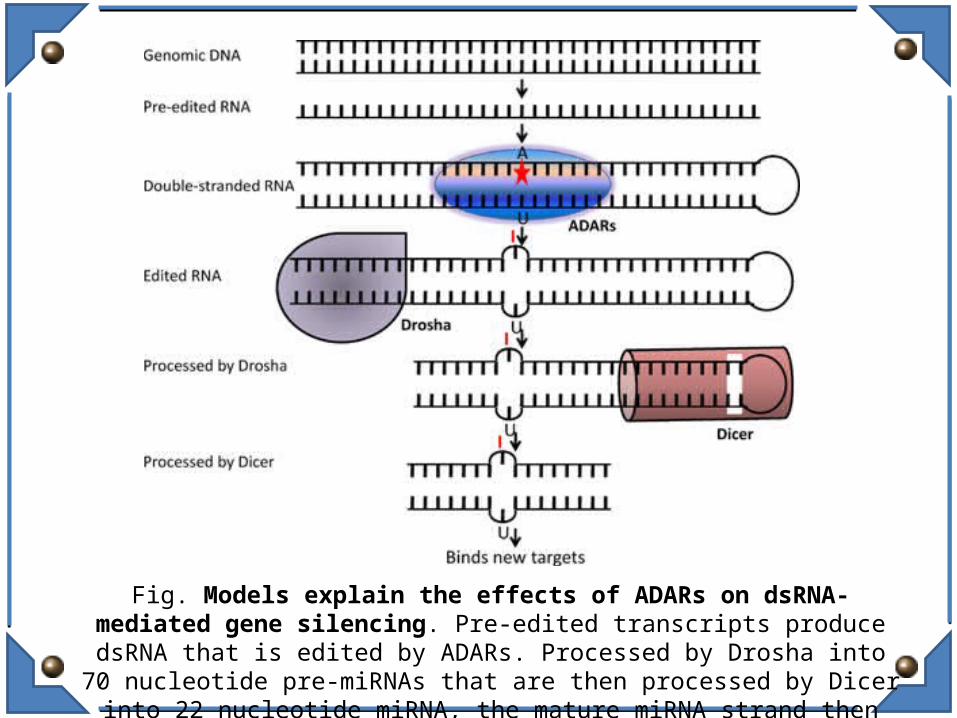
Fig. Models explain the effects of ADARs on dsRNA-mediated gene silencing. Pre-edited transcripts produce dsRNA that is edited by ADARs. Processed by
Drosha into 70 nucleotide pre-miRNAs that are then processed by Dicer into 22 nucleotide miRNA, the mature miRNA strand then pairs with the target mRNA
Nishikura Nature Reviews Molecular Cell Biology 7, 919–931 (2006)
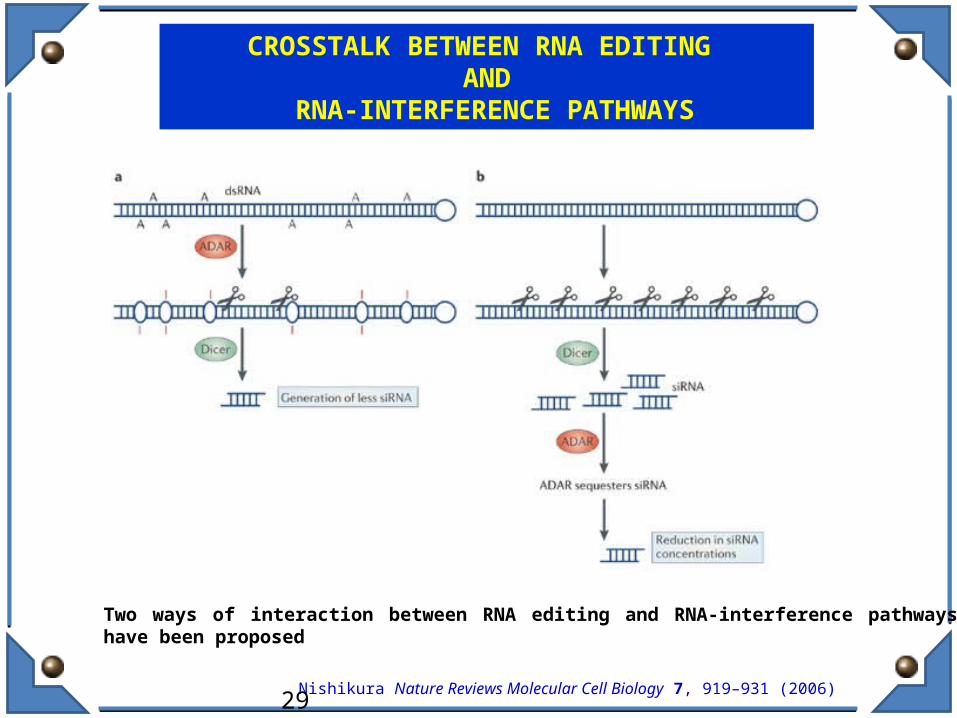
Two ways of interaction between RNA editing and RNA-interference pathways have been proposed
CROSSTALK BETWEEN RNA EDITING AND
RNA-INTERFERENCE PATHWAYS
29

The detection of editing sites in RNA molecules in particular cellular conditions is very difficult considering that RNA editing is a dynamic spatial–temporal process.
Recently, Sakurai et al. (2014) combined the ICE (inosine chemical erasing) method with HTS (ICE seq) for an unbiased genome-wide screening of novel A-to-I editing sites. ICE seq is able to detect editing sites in both repeat elements and short hairpins, rendering this a currently unique method for genome-wide identification of A-to-I editing events in both tissues and clinical specimens without genomic DNAs.
RNA Editing sites – The first centralized repository was DARNED (2010), whose last release contains more than 300,000 editing sites. Later, Ramaswami and Li (2014) built RADAR, a rigorously manually curated database of annotated A-to-I editing sites, amounting to about 1.4 million editing events.
(Nigita et al. (2015). Front. Bioeng. Biotechnol. 3:18)
30
Detection of Editing sites
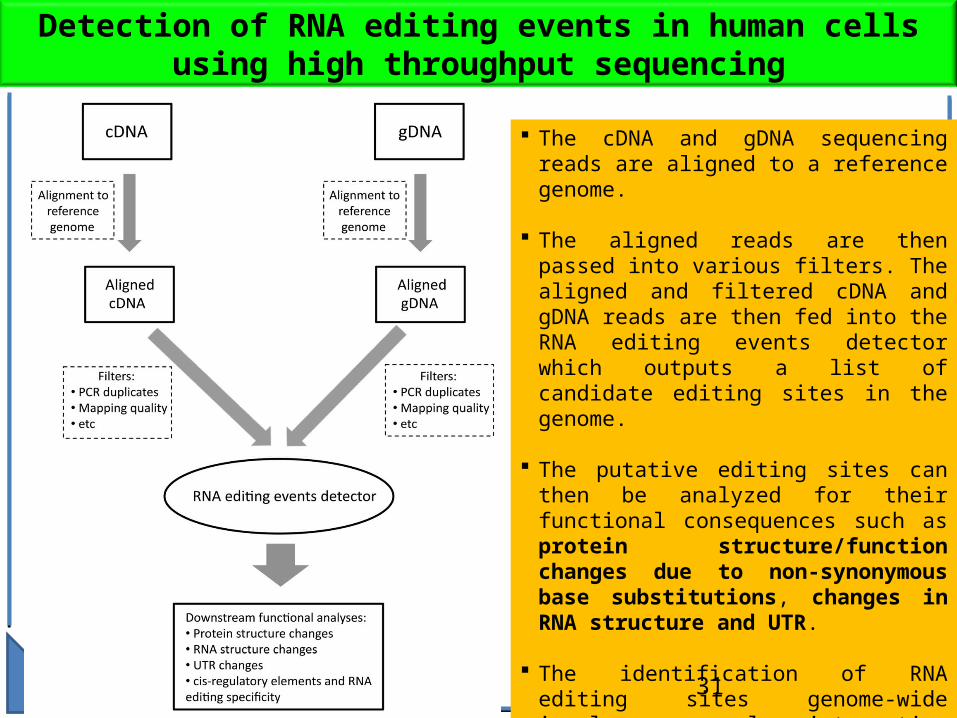
Detection of RNA editing events in human cells using high throughput sequencing
The cDNA and gDNA sequencing reads are aligned to a reference genome.
The aligned reads are then passed into various filters. The aligned and filtered cDNA and gDNA reads are then fed into the RNA editing events detector which outputs a list of candidate editing sites in the genome.
The putative editing sites can then be analyzed for their functional consequences such as protein structure/function changes due to non-synonymous base substitutions, changes in RNA structure and UTR.
The identification of RNA editing sites genome-wide involves a complex interaction of cis-regulatory sequences, base-paired RNA structures and trans-acting elements.
31

Green Glow Shows RNA Editing in Real Time
Glowing genes: White arrows show hot spots of ADAR activation
Researchers have devised a technique for seeing RNA editing process in real time—the corrected strand gives off a green glow—and even for the restoration of functionality report Nature Methods 9, 189–194 (2012)
Researchers have devised a technique for seeing RNA editing process in real time—the corrected strand gives off a green glow—and even for the restoration of functionality report Nature Methods 9, 189–194 (2012)
Reenan and colleagues produced fruit flies that included an altered version of the gene for the GFP. The protein product of their genetic construct would not glow—until it was first repaired by ADAR Nature Methods 9, 189–194 (2012)
Reenan and colleagues produced fruit flies that included an altered version of the gene for the GFP. The protein product of their genetic construct would not glow—until it was first repaired by ADAR Nature Methods 9, 189–194 (2012)
Because the technique actually repairs an error and restores a function, it suggests promise for gene therapy-based treatments—but at the RNA rather than DNA level. Such treatment would target transcription mistakes, which may contribute to diseases such as epilepsy, schizophrenia and suicidal depression.
Because the technique actually repairs an error and restores a function, it suggests promise for gene therapy-based treatments—but at the RNA rather than DNA level. Such treatment would target transcription mistakes, which may contribute to diseases such as epilepsy, schizophrenia and suicidal depression.
32

RNA Editing and its regulation in specific disease: Correct and regulated RE is important for functional nervous system and
avoiding neurological diseases.
Imbalance of RE may lead to dysfunction of some proteins involved in normal physiology such as neural and immune function.
In ALS, excitotoxic motor neuron death is mediated by glutamate induced Ca2+ influx through Ca2+ permeable AMPA receptor (GluR2 pre-mRNA codes for glutamine (Q) at 586 position of protein – ADAR2 takes up A-I editing converting to Arginine (R). Arginine at 586 position in edited GluR2 decreases calcium permeability because of positive charge of Arginine.) leading to Ca2+ overload and death.
33

Global Regulator of mRNA Editing Found
Using the nematode Caenorhabditis elegans as their model organism, Hundley et al., identified more than 400 new mRNA editing sites – the majority regulated by a single protein called ADR-1, which does not directly edit mRNA but rather
regulated how editing occurred by binding to the messenger RNAs subject to editing
Using the nematode Caenorhabditis elegans as their model organism, Hundley et al., identified more than 400 new mRNA editing sites – the majority regulated by a single protein called ADR-1, which does not directly edit mRNA but rather
regulated how editing occurred by binding to the messenger RNAs subject to editing
Protein controls editing, expanding the information content of DNA
Protein controls editing, expanding the information content of DNA
RNA editing within the mRNA allows a single gene to create multiple mRNAs that are subject to different modes of regulation. How exactly this process can
be modulated has not been clearly understood
34
RNA Editing and Drug target / Drugs:
Isoforms generated by RE that have preferential physiological effects and may provide additional drug target.
RNA editing as a therapeutic target for CNS disorders:Notable targets of RNA editing in the CNS include transcripts encoding subunits of the AMPA and kainate (KA) subtypes of glutamate receptor, the a3 subunit of the GABA receptor, the serotonin 2C (5 HT2C) receptor, and the Kv1.1 voltage-gated potassium channel. RNA editing can modulate the functional properties of the encoded protein products and variations in the editing of glutamate receptor subunit and 5 HT2C mRNAs have been observed in several CNS disorders.
Morabeto and Emeson (2009) Neuropsychopharmacology Reviews 34, 246
Therefore RE of these receptors and channels may change their protein functions leading to disease and could be therapeutic targets. Eg: Novel inhibitors of Tryp. brucei RE ligase IAre reportedly potential therapeutic drugs
RNA Editing and Generation of Antibody Diversity:
Activation induced cytidine deaminase initiates somatic hypermutation of Ab gene by deaminating deoxycytosine to deoxyuracil (C-U).
Chemical RNA Editing as a Possibility Novel Therapy for Genetic DisordersLuyen et al. (2012) International Journal of Advanced Computer Science, 2:6, 237-241,.
35

Major Strength in RNA Editing Research:
RE has been integrated into biological networks of gene expression, regulatory pathways and genome evolution.
Major weakness in RNA Editing Research:
1. How RE activity is globally regulated is unknown.2. Many editing site in human transcriptome remains to be discovered.3. Monitoring of global activity of RE in-vivo is rare.
Major future RNA Editing Research challenges:
1. Delineation of physiological significance of particular editing events identified in tumor, neuronal tissues and brain or other specialized organ.
2. Molecular mechanisms on how RE affect the expression or functions of many genes through the alteration of codon, splice pattern, stability or localization of protein coding transcripts, modulation of regulatory RNA biogenesis and function.
3. Monitoring of global activity of RE in-vivo as a useful early biomarker for patient with disease.
4. Therapeutic applications.
36

“It takes courage to tackle very
hard problems in science”
- Venky

38
ACKNOWLEDGEMENTS
I would never have been able to finish my seminar without the guidance ofmy teachers , help from friends, and support from my father .
I would like to express my deepest gratitude to my teacher, Dr. Manish Mahawar, for intoducing the topic and infusing the scientific acumen in me.
I would also like to thank my advisor Dr. Mohini Saini for providing me with the articles needed for the presentation. Without you madam this wouldn’t be possible.
I would like to thank Dr. Sumit Ranjan Mishra, who as a good friend, was always willing to help and give his best suggestions in improving my presentation.
Lastly but not the least I thank my dear father Dr. Bibek R. Shome who guided me step by step in the entire journey of my seminar and helped in understanding the subject in detail.